

Abstract
Efficient methods for multigene transformation are important for developing novel crop varieties. Methods based on random integrations of multiple genes have been successfully used for metabolic engineering in plants. However, efficiency of co‐integration and co‐expression of the genes could present a bottleneck. Recombinase‐mediated integration into the engineered target sites is arguably a more efficient method of targeted integration that leads to the generation of stable transgenic lines at a high rate. This method has the potential to streamline multigene transformation for metabolic engineering and trait stacking in plants. Therefore, empirical testing of transgene(s) stability from the multigene site‐specific integration locus is needed. Here, the recombinase technology based on Cre‐lox recombination was evaluated for developing multigenic lines harboring constitutively‐expressed and inducible genes. Targeted integration of a five genes cassette in the rice genome generated a precise full‐length integration of the cassette at a high rate, and the resulting multigenic lines expressed each gene reliably as defined by their promoter activity. The stable constitutive or inducible expression was faithfully transmitted to the progeny, indicating inheritance‐stability of the multigene locus. Co‐localization of two distinctly inducible genes by heat or cold with the strongly constitutive genes did not appear to interfere with each other's expression pattern. In summary, high rate of co‐integration and co‐expression of the multigene cassette installed by the recombinase technology in rice shows that this approach is appropriate for multigene transformation and introduction of co‐segregating traits.
Significance Statement
Recombinase‐mediated site‐specific integration approach was found to be highly efficacious in multigene transformation of rice showing proper regulation of each gene driven by constitutive or inducible promoter. This approach holds promise for streamlining gene stacking in crops and expressing complex multigenic traits.
Keywords: Cre‐lox, gene stacking, multigene transformation, site‐specific recombination, trait engineering
1. INTRODUCTION
The demand for resilient, productive, and value‐added crops mandates breeding with multiple genes. Biotechnology innovates crop improvement through rapid introduction of genes; however, independently added genes are difficult to stack into cultivars. The population size needed to isolate a stacked F2 plant increases exponentially with the increasing number of unlinked genes, and screening of thousands of F2 plants becomes necessary for breeding 5–6 unlinked genes as compared to a handful of plants for a single gene. Therefore, breeding could be turned into a high efficiency process by simply linking the genes and stacking them into a locus (Petolino & Kumar, 2016; Que et al., 2010).
Multigene stacking could be obtained through co‐bombardment of plasmids that tend to recombine and co‐integrate into the plant genome; however, co‐expression of the introduced genes, in this method, generally occurs at a low rate (Chen et al., 1998; Gelvin, 1998; Schmidt, LaFayette, Artelt, & Parrott, 2008; Zhu et al., 2008). Agrobacterium‐mediated T‐DNA transfer is also an excellent method of multigene transformation as T‐DNA harboring many genes enters into the plant cell and integrates into the genome (Collier, Thomson, & Thilmony, 2018; Lin, Liu, Xu, & Li, 2003; Ruiz‐Lopez, Haslam, Usher, Napier, & Sayanova, 2015). However, integration of tandem repeats or truncated copies of T‐DNA, and disruption of critical genomic regions cannot be ruled out. These features of random transformation methods pose major challenges for multigene transformation, rendering a high number of the recovered events unsuitable for product development (Anand & Jones, 2018; Halpin, 2005).
The targeted integration approaches, on the other hand, are ideal for precise integrations into selected genomic sites. A number of targeted methods have been described that can be grouped into (a) recombinase‐mediated integration into pre‐characterized sites, and (b) integrations into double‐stranded breaks (DSB) induced by site‐specific DSB reagents such as CRISPR/Cas9 (Chen & Ow, 2017; Petolino & Kumar, 2016; Srivastava & Thomson, 2016; Weeks, Spalding, & Yang, 2016). Each method has its own advantages and disadvantages. While recombinase‐mediated methods are highly efficient in targeted integrations, for example, with Cre‐lox, FLP‐FRT, and other site‐specific recombination systems, they require placement of recombination target sites that can subsequently be targeted for gene stacking (Anand et al., 2019; Li et al., 2009; Ow, 2003; Srivastava & Gidoni, 2010). The CRISPR/Cas9 method, on the contrary, can target virtually any site in the genome, but the nonhomologous mode of DSB repair in plant cells overrides the integration of exogenous DNA into the DSB site (Puchta & Fauser, 2014; Voytas, 2013). As a result, the current DSB‐based approaches require extensive efforts and expanded post‐transformation analysis for isolating the rare individuals that contain targeted integrations (Svitashev et al., 2015; Yang, Luo, Mo, & Liu, 2019). Therefore, although, recombinase‐mediated approach can only be practiced on the “prepared” target lines, it provides a promising platform for developing gene stacking technologies for crops.
Cre‐lox site‐specific recombination is one of the most attractive systems for recombinase‐mediated genome engineering. Its high efficiency has been demonstrated in a number of plant species, most of which develop into healthy fertile plants (Gidoni, Srivastava, & Carmi, 2008; Srivastava & Thomson, 2016). The phenotypic effects of Cre recombinase in plants are either undetectable or reversible owing to its high specificity and undetectable off‐target activity (Coppoolse et al., 2003; Ream et al., 2005; Srivastava & Nicholson, 2006). Other site‐specific recombination systems that show high efficiency in plant cells include FLP‐FRT, phiC31‐att, and Bxb1‐att systems (Anand et al., 2019; Hou et al., 2014; Li et al., 2009; Nandy & Srivastava, 2011; Ow, 2011; Thomson & Ow, 2006). Recombinase‐mediated gene stacking has been demonstrated in soybean by inserting gene silencing and overexpression constructs through two rounds of transformation (Li et al., 2010). However, more research is needed to test the efficiency of gene stacking by recombinases, and verify its efficacy for co‐expression of the stacked genes. Further, since the stacked genes could consist of conditionally or developmentally regulated genes, it is important to test the recombinase‐mediated gene stacking approach with inducible genes.
This study evaluated Cre‐lox mediated site‐specific integration of a set of five genes consisting of three constitutively overexpressed and two inducible genes in the crop model rice by analyzing transformation efficiency and expression stability over three generations. High transformation efficiency of the multigene construct harboring repeat sequences, strong expression of the constitutively expressed genes, and proper regulation of the inducible genes demonstrates that Cre‐lox mediated integration into the engineered target sites is a practical approach for gene stacking and trait engineering into the crop genomes.
2. RESULTS
2.1. Molecular strategy
This study used the Cre‐lox mediated site‐specific integration approach based on the use of mutant lox sites, lox75, and lox76, to stabilize the integration locus as described earlier (Albert, Dale, Lee, & Ow, 1995; Srivastava & Ow, 2002). Specifically, the rice target line, T5, is retransformed by the donor vector pNS64 to obtain the multigene site‐specific integration locus. T5 target locus consists of a lox76 placed between the maize ubiquitin‐1 promoter (ZmUbi1) and cre coding sequence (Figure 1a), and pNS64 contains a loxP and lox75‐flanked gene construct consisting of a promoter‐less selectable marker (NPT II) and four genes‐of‐interest (GFP, GUS, DREB1a, and pporRFP), each expressed by their dedicated promoters (Figure 1b). Upon entry into T5 cells, pNS64 undergoes Cre‐lox recombination leading to site‐specific integration of the gene construct (without vector backbone) into the target site (Figure 1c). The resulting site‐specific integration (SSI) is selectable due to the placement of NPT II downstream of strong promoter (ZmUbi1), and characterized by distinct left and right junctions.
FIGURE 1.

Molecular approach for site‐specific integration (SSI) of a multigene stacking. (a) T5 locus in rice cv. Taipei‐309 consisting of a single‐copy of T‐DNA encoding Cre activity and the target lox76 site (black triangle). (b) Donor vector, pNS64, in pBluescript SK backbone (not shown) containing promoterless NPT II gene and four expression units (GFP, GUS, AtDREB1A, and pporRFP) between loxP and lox75 (black triangles). The loxP x lox75 recombination circularize the gene construct, which subsequently integrates into T5 locus to generate the site‐specific integration (SSI) structure. The NPT II gene captures the maize ubiquitin‐1 promoter (ZmUbi1) at T5 locus to make the event selectable on geneticin™, and SSI locus expresses four genes, two constitutive (GFP and GUS) and two inducible (AtDREB1A and pporRFP) genes. (c) Structure of the predicted site‐specific integration (SSI) locus that expresses a stack of four genes (NPT II, GFP, GUS, AtDREB1A, and pporRFP). The primers sites and the expected PCR products are indicated below the structure, while EcoRI sites (E) and the fragment sizes are shown above the structure. 35S, Cauliflower Mosaic Virus 35S promoter; NPT II, neomycin phosphotransferase II; GFP, green fluorescent protein; GUS, β‐Glucuronidase; AtRD29a, Arabidopsis thaliana RD29a promoter; AtDREB1A, Arabidopsis thaliana dehydration responsive element 1A; HSP17.5E, soybean heat‐shock 17.5E gene promoter, and pporRFP, sea coral Porites porites red fluorescent protein. Each gene carries a nopaline synthase (nos 3′) transcription terminator (not shown)
2.2. Characterization of the transgenic lines
Using gene gun mediated transformation of T5 line (cv. Taipei‐309) with pNS64, 32 T0 plants were obtained (Table S1). Four of the T0 plants were removed from the analysis due to their weak stature at the young vegetative stage, and remaining 28 were analyzed by PCR. Twenty‐seven of these lines were found to contain the SSI junctions indicated by 0.5 kb and 1 kb left and the right junction bands, respectively (Figure S1a). Five of these lines contained biallelic integrations as indicated by target site PCR and genetic segregation among T1 progeny (Figure S1a; Table S1). Southern blot hybridization of EcoRI‐digested genomic DNA showed the presence of predicted 3.2 kb left junction and 2.1 kb right junction, upon hybridization with GFP and pporRFP probes, respectively (Figure S2a,b), and hybridization with GUS and AtDREB1a probes, in a subset of 22 lines, showed the predicted 2.5 kb GUS or 2.0 kb AtDREB1a in all except four lines (Figure S2c,d). Thus, full‐length SSI copy was found in 18 lines, and the remaining four contained truncation of the middle portion of the cassette (Table S1). Long PCR (4 kb) at the right junction verified this finding and identified another line (#4) with full‐length SSI (Figure S1b; Table S1). The lines lacking GUS and AtDREB1a genes in the Southern blot failed this PCR, while the remaining showed the predicted 4 kb amplicon, indicating a full‐length SSI structure (Figure S1b). In addition, Southern analysis revealed that 13 lines contained only the SSI copy, whereas the remaining contained additional 1–3 copies (Figure S2; Table S1). Two clonal lines showing identical hybridization patterns with four genes were also identified (Figure S2; Table S1). In summary, the presence of multigene SSI locus was validated in 19 of the 30 SSI lines (removing two clonal lines from the initial total of 32) developed from the bombardment of 30 callus plates. Eleven of these lines were selected, based on plant vigor and seed abundance, for gene expression analysis (Table S1).
2.3. Expression of the constitutive genes
Three constitutively‐expressed genes, NPT II, GFP, and GUS, in the order of arrangement in the SSI locus, are each controlled by a strong constitutive promoter (Figure 1). While NPT II serves as the selectable marker, GFP and GUS represent the genes‐of‐interest. Expression of these genes was determined at transcript and protein levels in the T0 lines and their progeny using T5 line as the negative control. Transcript analysis by RT‐qPCR, in the T0 plants, showed that most lines abundantly expressed the three genes within 2‐ to 5‐fold range (Figure 2a–c). However, SSI line #12 behaved atypically as it expressed NPT II and GFP transcripts within the range but showed markedly lower levels of GUS transcripts (Figure 2c). Although, gene expression at transcript levels in SSI lines has not been evaluated in previous reports, >10‐fold lower GUS transcript in line #12 was somewhat surprising. Transcript abundance analysis in the T1 progeny was performed in 10‐day old seedlings grown in the germination media. This analysis showed a greater variation in the transcript levels of the three genes (Figure 2d–f); however, all T1 plants abundantly expressed the three genes. Notably, T1 progeny of line #12 showed a similar pattern as found in the parental line, consisting of higher levels or NPT II and GFP transcripts but markedly lower levels of GUS transcripts (Figure 2f).
FIGURE 2.

Transcript abundance analysis of constitutively expressed genes by real time quantitative PCR (RT‐qPCR). Relative expression of NPT II, GFP, and GUS genes in the T0 plants (a–c) and the T1 progeny seedlings (d–e) of site‐specific integration (SSI) lines in comparison to the T5 negative control. The SSI line numbers are given on x‐axis. The values are the average of two biological replicates and two technical replicates of each. Standard errors are indicated as the error bars
Steady state mRNA levels can only partially predict protein abundance as posttranscriptional regulation and cell perturbation among other factors influence the correlation of transcript and protein abundance (Vogel & Marcotte, 2012). Therefore, gene expression analysis at the protein level is important for interpreting the functional effects. In this study, NPT II ELISA, GFP fluorescence, and GUS activity were measured to understand gene expression from the stacked locus. The NPT II activity in T0 plant was expected be strong owing to the selection during tissue culture; therefore, NPT II ELISA was done only in T1 progeny. We found a 4‐fold variation in GFP fluorescence and 2.8‐fold variation in GUS activity among SSI lines (Figure 3a,b), indicating a good correlation of these protein activities with their transcript levels. However, line #12, to our surprise, showed a comparable GUS activity (Figure 3b), even though it was found to express markedly lower levels of the GUS transcripts (Figure 2c). The reason behind this discrepancy is not clear but it could be an artifact of the GUS assay. If optimum amount of substrate is not used in the reaction, the enzyme rate may not be directly proportional to the enzyme concentration (Robinson, 2015). To further check GUS activity in line #12, histochemical staining of leaf cuttings was done according to Jefferson, Kavanagh, and Bevan (1987). A lower staining intensity in line #12 compared to other lines such as #10 (Figure 3c), suggested that #12 contains a lower GUS activity, and the transcript abundance in this line is a good indicator of its GUS levels. Among T1 progeny of the SSI lines, only <3‐fold variation for NPT II and GFP abundance, and ~5‐fold variation for GUS activity was observed (Figure 3d–f). Finally, a subset of four SSI lines was selected for T2 progeny analysis. T2 seedlings of these lines were found to abundantly express GFP fluorescence and GUS activities at more or less similar levels (Figure 3g,h), indicating that stacked genes at the SSI locus are stably transmitted to the progeny. The stability of the genes was also reflected by the gene dosage effect as the biallelic lines generally expressed GFP and GUS at higher levels compared to the monoallelic lines, although, a significant difference was only found for GFP (Figure 3i). In summary, stable expression of the three stacked genes regulated by strong promoters at the SSI locus was found in all lines tested in this study. Only one SSI line (#12) showed suboptimal expression of one of the three genes (GUS), while all other showed comparable expression that were transmitted to the subsequent generations.
FIGURE 3.
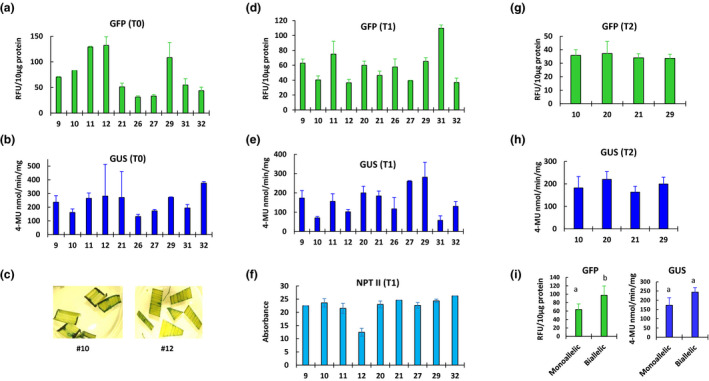
Protein abundance analysis of constitutively‐expressed genes in the site specific integration (SSI) lines. (a–b) Quantitative GFP fluorescence and GUS activity in T0 plants. (c) comparative histochemical GUS staining of leaf cuttings from SSI lines #10 and #12. (d–f) GFP fluorescence, GUS activity, and NPT II ELISA in T1 progeny seedlings. (g–h) quantitative GFP fluorescence and GUS activity in T2 progeny seedlings. (i) average GFP and GUS activities in monoallelic and biallelic T0 SSI lines (described in Table S1). The SSI line numbers are given on x‐axis. Statistical differences, shown by the alphabets, were determined by student t test at p = .05. Standard errors are indicated as the error bars
2.4. Expression of inducible genes
The SSI locus contained two inducible genes, a transcription factor, AtDREB1a, and a fluorescent protein, pporRFP, driven by RD29a and HSP17.5E promoters, respectively. These genes are expected to be “off” at room temperature but abundantly induced by specific treatments, that is, cold‐shock for RD29a and heat‐shock for HSP17.5E. The expression pattern of these genes was studied by RT‐qPCR in T0 plants and their progeny, at room temperature and upon treatment, using T5 as the negative control. Expression analysis of AtDREB1a in T0, T1, and T2 plants showed abundant transcripts upon cold‐shock treatment (Figure 4a–c). Similarly, pporRFP analysis in T0, T1, and T2 plants showed abundant transcripts in heat‐shock‐treated samples (Figure 4d–f). These experiments verified that the two inducible genes work properly and respond to the treatment. Notably, both genes expressed at a basal level at the room temperature, indicating proper regulation of the genes in SSI lines, and its faithful transmission to the progeny. The fold induction was highly variable between lines, but most lines showed strong induction upon treatment. Finally, specificity of the genes was determined by checking their expression upon nonspecific perturbation, that is, induction of RD29a:AtDREB1a by heat‐shock and HSP17.5E:pporRFP by cold‐shock treatment. As expected, heat‐shock treatment had little effect on AtDREB1a and cold‐shock treatment did not induce pporRFP (Figure S3a,b), showing that stacking these two genes closely together did not break their promoter‐specificity in the SSI lines.
FIGURE 4.

Expression analysis of the inducible genes by real time quantitative PCR (RT‐qPCR) in the site‐specific integration (SSI) lines relative to T5 negative control. (a–c) AtDREB1a expression analysis at room temperature (white bars) or upon cold‐induction (20 hr in ice pack, magenta bars) in T0, T1, and T2 plants. (d–f) pporRFP expression at room temperature (white bars) or upon heat‐induction (42°C for 3 hr; red bars) in T0, T1, and T2 plants. The SSI line numbers are given on x‐axis. The values are the average of two biological replicates with standard error indicated as the error bars
Downstream targets of AtDREB1a transcription factor during cold stress are not clearly known in rice (Oh et al., 2005); therefore, it's downstream effects in SSI lines could not be analyzed. The functional effects of AtDREB1a expression would require phenotypic analysis in stress conditions. Therefore, protein abundance analysis was performed only on ppor RFP. Confocal imaging of root or leaf samples was done on the T1 seedlings of three SSI lines (#9, #10, #12) using GFP as the internal control. pporRFP is a complex dsRed type RFP that undergoes homo‐dimerization to become functional and emit fluorescence (Sacchetti, Subramaniam, Jovin, & Alberti, 2002). In a constitutive expression system, dimerization would occur continuously. However, in the inducible expression system, it is important to determine the lag time for pporRFP maturation. For this, SSI #9 seedlings were heat‐treated and their roots were imaged at three different time intervals. Optimum fluorescence was observed 72 hr after heat‐treatment (Figure S4). This is in agreement with the published report on temporal decoupling of dsRed RFP mRNA and protein expression. In tobacco, dsRed RFP mRNA was found to peak at 2–3 days after transformation, while fluorescence peak occurred 3–5 days later (Jansing & Buyel, 2019). Accordingly, seedlings of SSI lines #9, #10, and #12 were subjected to 3 hr of heat‐shock treatment at 42°C followed by fluorescence imaging 72 hr later in confocal microscope. As expected, each SSI line abundantly expressed GFP regardless of the treatment. Whereas, pporRFP showed an inducible pattern, characterized by undetectable red fluorescence at room temperature (RT) and enhanced fluorescence upon heat‐shock (HS) in the root or leaf tissues (Figure 5). In summary, the two co‐localized inducible genes that are regulated by specific stress treatments, expressed properly from the stacked SSI locus without apparent interference of the neighboring strong constitutive genes or the inducible genes.
FIGURE 5.

Confocal imaging of GFP (top) and pporRFP (bottom) in the roots and leaves of the 10 days old T1 seedlings of site‐specific integration lines #9, #10, and #12. All images were taken 72 hr postheat‐shock treatment at 20x magnification. T5: Target line (negative control); HS, heat‐shock; RT: room temperature. Scale bar in leaves: 100 µm and in roots: 50 µm
3. DISCUSSION
Multigene transformation has overcome the technical barriers of high load gene transfer but the challenges related to co‐integration and co‐expression remain to be fully addressed. Both Agrobacterium and gene gun are competent at multigene transfers; however, complexity of DNA integration into the plant cell leads to multiple random outcomes consisting of complex integrations into unique genomic sites (Saika, Nishizawa‐Yokoi, & Toki, 2014; Somers & Makarevitch, 2004). Co‐bombardment of gene vectors or multigenic T‐DNA have been used for metabolic engineering in plants (Naqvi et al., 2009; Ruiz‐Lopez et al., 2015), however, low rate of co‐expression from multigene assemblies poses a major bottleneck (Ghareeb, Laukamm, & Lipka, 2016; Schmidt et al., 2008). Recombinase‐mediated site‐specific integration overcomes the complexity of DNA integration and simplifies the structure, which, in turn, removes gene silencing triggers and creates a favorable environment for stable gene expression (Akbudak, More, Nandy, & Srivastava, 2010; Chawla, Ariza‐Nieto, Wilson, Moore, & Srivastava, 2006; Day et al., 2000; Li et al., 2010; Nanto, Sato, Katayama, & Ebinuma, 2009). In this study, we evaluated the recombinase approach for multigene transformation, specifically, asking how efficiently a multigene construct integrates into the targeted site and whether the integrated genes express faithfully in the plants. Using an available target line, we developed site‐specific integration of a five‐gene cassette through Cre‐lox recombination in rice. The molecular strategy of site‐specific integration has been described earlier (Srivastava & Ow, 2002). Using gene gun mediated particle bombardment, 30 transgenic events were obtained from 30 shots, 19 of which contained precise multigene integration characterized as full‐length integration of the cassette into the target site. This efficiency of co‐transformation is exceptionally higher than generally found in conventional methods. As observed in previous studies (Albert et al., 1995; Srivastava, Ariza‐Nieto, & Wilson, 2004), the majority of these lines (13 out of 19) contained only the SSI copy, and random integrations were low or undetectable. Most importantly, all tested lines expressed each of the five genes, affording stable expression at 100% rate for the multigene assembly in this study. The strong constitutive genes showed abundant expression levels and the inducible genes functioned according to their promoter specificity. Notably, the two inducible genes showed no apparent interference of the neighboring genes. Additionally, the multigene cassette was faithfully transmitted to the progeny as shown by gene expression analysis in T1 and T2 progeny. Interestingly, the repeated use of nos terminator in the construct did not seem to affect the structure stability. However, for product development, this feature should be avoided as repeat sequence in multigene assemblies could induce homology‐directed truncations. In conclusion, recombinase‐mediated site‐specific integration approach proved to be highly efficient in developing site‐specific integration lines harboring precise integration of the five‐gene cassette, and the efficacy of the approach was further demonstrated by proper expression of all genes and their inheritance by the progeny. The recombinase mediated multigene transformation approach can also be practiced with the “cassette exchange” strategy as described earlier (Li et al., 2010). In addition, the strategy can easily be modified to incorporate marker‐removal feature by employing another recombinase system (Nandy & Srivastava, 2012), that have been found to function in plant cells (Chen & Ow, 2017; Cody, Graham, Zhao, Swyers, & Birchler, 2020).
4. EXPERIMENTAL PROCEDURES
4.1. Vector construction and transformation
The multigene vector, pNS64 (Figure 1b), was developed through the standard restriction digestion and ligation method. The individual gene cassettes from pUC vectors were ligated one by one into pAA12 backbone that contains a promoterless neomycin phosphotransferase II (NPT II) gene between loxP and lox75 (Akbudak & Srivastava, 2017). Two gene cassettes, 35S:GFP and 35S:GUS were already available that contained CaMV 35S promoter driving GFP or GUS gene. To build RD29a:AtDREB1a cassette, Arabidopsis thaliana dehydration responsive element B1A (AtDREB1A) and Arabidopsis cold inducible RD29a promoter sequences were generated by PCR. For HSP17.5E:pporRFP cassette, the promoter fragment of Gmhsp17.5E was obtained by PCR on soybean DNA and pporRFP from pANIC6A that contains coral Porites porites RFP (Alieva et al., 2008; Mann et al., 2012). Primers used for cloning are given in Table S2. All gene constructs contain nopaline synthase transcription termination (nos 3′).
The rice line T5 (Taipei 309) that contains a Cre‐lox target site was used in the present study to develop site‐specific integration lines. The development of T5 line is described by Srivastava and Ow (2002). The pNS64 vector was delivered by gene gun (PDS 1000, Bio‐Rad Inc.) into the embryogenic callus derived from T5 mature seeds. The bombarded callus was selected on 100 mg/L geneticin™ to isolate the site‐specific integration lines. The tissue culture media for callus induction and plant regeneration were used according to Nishimura, Aichi, and Matsuoka (2006).
4.2. Molecular analysis
The polymerase chain reaction (PCR) was performed on genomic DNA using Emerald Amp MAX PCR Master Mix (Takara Bio) using the primers given in Table S2. Southern blot analysis was performed on genomic DNA digested with EcoR1 using 32P‐labeled DNA probes of GFP, RFP, GUS, and AtDREB1A. Gene expression analysis by reverse transcriptase (RT)‐quantitative (q) PCR was performed on total RNA isolated using Trizol reagent (Invitrogen, Inc.) according to manufacturer's protocol, and quantified on Nano‐drop 2000 (Thermo‐Fisher Inc.). Two microgram of total RNA, treated with RQ1‐RNAse free DNase (Promega Inc.), was used for the cDNA synthesis using PrimeScript RT reagent kit (Takara Bio), and used for qPCR using TB green Premix Ex Taq II (Takara Bio) on Bio‐Rad CFX 96 C1000. The product specificity was verified by the melt curve analysis and the Ct values were normalized against seven ubiquitin. The relative expression was calculated against T5 negative control and the untreated controls using delta‐delta Ct method (Livak & Schmittgen, 2001). Each line contained two to three biological replicates with two technical replications of each. For inducible gene expression analysis, excised leaf blades from the greenhouse plants wrapped in aluminum foil or seedlings germinated on MS/2 media in petri dishes, were placed on ice pack for 20 hr for cold‐shock treatment or 42°C incubator for 3 hr for heat‐shock treatment. The room temperature controls were handled in the same way except they were kept at the ambient room temperature.
4.3. Protein analysis
NPT II enzyme linked immunosorbent assay (ELISA) were conducted using a commercial kit (Agdia Inc.) according to the manufacturer's instructions. Fifty milligrams of fresh leaf from 1‐month old plants was ground in the protein extraction buffer provided in the kit, and centrifuged to collect the crude protein extract. The NPT II protein provided in the kit and the T5 crude protein extract were used as a positive and negative controls, respectively. ELISA plates were read at A650 in the Synergy Biotek Cytation 3. The ratio of the absorbance of samples to T5 negative control was used as the measure of NPT II expression.
GFP fluorescence was observed in the young tissue under the Leica 56D stereoscope fitted with the 440–460 nm excitation and 500–560 nm (band pass) emission filters (Night Sea, Lexington, MA). For the quantitative estimation, fresh tissue was ground in 10 mM of Tris–EDTA, pH 8.0, at 4°C and centrifuged at 15000 g for 20 min to collect the supernatant. A strongly expressing rice GFP line, C30‐1, described earlier (Pathak, Pruett, Guan, & Srivastava, 2019), was used as a reference. Ten microgram of protein was used for GFP fluorescence measurement in Versa‐fluorimeter (Bio‐Rad Inc) equipped with 490 ± 5 nm excitation filter and 510 ± 5 nm emission filter. All lines were measured against C30‐1 positive control and the T5 negative control. A unit of GFP was defined as relative fluorescence units per 10 µg of total protein (RFU/10 µg).
Histochemical GUS staining was done by submerging leaf cuttings in the GUS staining solution containing 1 mM of X‐Gluc (Gold Biotechnologies) according to Jefferson et al. (1987). For quantitative measurements, 10 µg of total protein extracted in 50 mM of phosphate buffer, pH 7, was used for GUS assay using 4‐methylumbelliferyl b‐D‐glucuronide (4‐MUG) as the substrate, and the kinetics of the reaction measured in spectrophotometer by appearance of a colored product 4‐methylumbelliferone (4‐MU) according to Jefferson et al. (1987). A unit of GUS activity was defined as nmol 4‐MU produced per minute from 1 mg of the protein (nmol min−1 mg−1). For each of the above assays, 2–3 biological replicates with the two technical replicates were included, and all protein estimations were done using Bradford reagent (VWR Inc.).
4.4. Confocal imaging
The pporRFP detection was performed using confocal imaging in the 7–10 days old seedlings. The seedlings were heat‐shocked as described in the molecular analysis section, and imaging was done at 24, 48 and 72 hr post heat‐shock treatment. The images were captured using a Leica TCS SP5 confocal microscope by the bandwidth adjustment for the fluorescence detection. For roots imaging, the samples were excited using 514 Argon and 594 HeNe laser channels and emission was collected at 542–582 mm for GFP, and at 610–710 nm for pporRFP. For leaf imaging, samples were excited at 514 Argon laser channel, and emission was collected at 590–610 nm for blocking the chlorophyll auto‐fluorescence. The leaf images were captured through sequential scan to prevent the bleed‐through between chlorophyll autofluorescence and fluorescent protein(s). A GFP positive line, C30‐1, described earlier (Pathak et al., 2019), and the parental T5 seedlings were used as controls. For all samples, first the gain, zoom, and offset were adjusted for T5 negative control, and then, all images were captured using the same parameters at 20x magnification.
4.5. Accession numbers
The NCBI accession numbers of the genes used in this study:
GFP: EF090408.1
NPT II: AF485783
GUS: AF485783
pporRFP: DQ206380.1
AtDREB1a: NM_118680.2.
CONFLICT OF INTEREST
Authors state no conflict of interest.
AUTHOR CONTRIBUTION
VS conceived the idea. BP conducted all experiments. VS and BP analyzed the data and drafted the manuscript. Both authors read and approved the manuscript.
Supporting information
Fig S1‐S4
Table S1
Table S2
ACKNOWLEDGMENT
We thank Soumen Nandy for pNS64 construction, Shan Zhao for rice transformations, Betty Martin for confocal imaging, and Jessica Kivett for the greenhouse support. This project was supported in part by USDA‐NIFA grant 2017‐38821‐26412 and NSF‐EPSCoR grant 1826836.
Pathak B, Srivastava V. Recombinase‐mediated integration of a multigene cassette in rice leads to stable expression and inheritance of the stacked locus. Plant Direct. 2020;4:1–10. 10.1002/pld3.236
This manuscript was previously deposited as a preprint at https://www.biorxiv.org/content/10.1101/2020.04.16.045435v1.
REFERENCES
- Akbudak, M. A. , More, A. B. , Nandy, S. , & Srivastava, V. (2010). Dosage‐dependent gene expression from direct repeat locus in rice developed by site‐specific gene integration. Molecular Biotechnology, 45, 15–23. 10.1007/s12033-009-9235-z [DOI] [PubMed] [Google Scholar]
- Akbudak, M. A. , & Srivastava, V. (2017). Effect of gene order in DNA constructs on gene expression upon integration into plant genome. 3 Biotech, 7, 94 10.1007/s13205-017-0729-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albert, H. , Dale, E. C. , Lee, E. , & Ow, D. W. (1995). Site‐specific integration of DNA into wild‐type and mutant lox sites placed in the plant genome. The Plant Journal, 7, 649–659. 10.1046/j.1365-313x.1995.7040649.x [DOI] [PubMed] [Google Scholar]
- Alieva, N. O. , Konzen, K. A. , Field, S. F. , Meleshkevitch, E. A. , Hunt, M. E. , Beltran‐Ramirez, V. , … Matz, M. V. (2008). Diversity and evolution of coral fluorescent proteins. PLoS One, 3(7), e2680 10.1371/journal.pone.0002680 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anand, A. , & Jones, T. J. (2018). Advancing Agrobacterium‐based crop transformation and genome modification technology for agricultural biotechnology. Current Topics in Microbiology and Immunology, 418, 489–508. 10.1007/82_2018_97 [DOI] [PubMed] [Google Scholar]
- Anand, A. , Wu, E. , Li, Z. , TeRonde, S. , Arling, M. , Lenderts, B. , … Chilcoat, N. D. (2019). High efficiency Agrobacterium‐mediated site‐specific gene integration in maize utilizing the FLP‐FRT recombination system. Plant Biotechnology Journal, 17, 1636–1645. 10.1111/pbi.13089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chawla, R. , Ariza‐Nieto, M. , Wilson, A. J. , Moore, S. K. , & Srivastava, V. (2006). Transgene expression produced by biolistic‐mediated, site‐specific gene integration is consistently inherited by the subsequent generations. Plant Biotechnology Journal, 4, 209–218. 10.1111/j.1467-7652.2005.00173.x [DOI] [PubMed] [Google Scholar]
- Chen, L. , Marmey, P. , Taylor, N. J. , Brizard, J. P. , Espinoza, C. , D'Cruz, P. , … Fauquet, C. M. (1998). Expression and inheritance of multiple transgenes in rice plants. Nature Biotechnology, 16, 1060–1064. 10.1038/3511 [DOI] [PubMed] [Google Scholar]
- Chen, W. , & Ow, D. W. (2017). Precise, flexible and affordable gene stacking for crop improvement. Bioengineered, 8, 451–456. 10.1080/21655979.2016.1276679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cody, J. P. , Graham, N. D. , Zhao, C. , Swyers, N. C. , & Birchler, J. A. (2020). Site‐specific recombinase genome engineering toolkit in maize. Plant Direct, 4, 1–9. 10.1002/pld3.209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collier, R. , Thomson, J. G. , & Thilmony, R. (2018). A versatile and robust Agrobacterium‐based gene stacking system generates high‐quality transgenic Arabidopsis plants. The Plant Journal, 95, 573–583. 10.1111/tpj.13992 [DOI] [PubMed] [Google Scholar]
- Coppoolse, E. R. , de Vroomen, M. J. , Roelofs, D. , Smit, J. , van Gennip, F. , Hersmus, B. J. , … van Haaren, M. J. (2003). Cre recombinase expression can result in phenotypic aberrations in plants. Plant Molecular Biology, 51, 263–279. 10.1023/A:1021174726070 [DOI] [PubMed] [Google Scholar]
- Day, C. D. , Lee, E. , Kobayashi, J. , Holappa, L. D. , Albert, H. , & Ow, D. W. (2000). Transgene integration into the same chromosome location can produce alleles that express at a predictable level, or alleles that are differentially silenced. Genes & Development, 14, 2869–2880. 10.1101/gad.849600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gelvin, S. B. (1998). Multigene plant transformation: More is better!. Nature Biotechnology, 16, 1009–1010. 10.1038/3452 [DOI] [PubMed] [Google Scholar]
- Ghareeb, H. , Laukamm, S. , & Lipka, V. (2016). COLORFUL‐Circuit: A platform for rapid multigene assembly, delivery, and expression in plants. Frontiers in Plant Science, 7, 246 10.3389/fpls.2016.00246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gidoni, D. , Srivastava, V. , & Carmi, N. (2008). Site‐specific excisional recombination strategies for elimination of undesirable transgenes from crop plants. In Vitro Cellular & Developmental Biology ‐ Plant, 44, 457–467. 10.1007/s11627-008-9140-3 [DOI] [Google Scholar]
- Halpin, C. (2005). Gene stacking in transgenic plants – The challenge for 21st century plant biotechnology. Plant Biotechnology Journal, 3, 141–155. 10.1111/j.1467-7652.2004.00113.x [DOI] [PubMed] [Google Scholar]
- Hou, L. , Yau, Y.‐Y. , Wei, J. , Han, Z. , Dong, Z. , & Ow, D. W. (2014). An open‐source system for in planta gene stacking by Bxb1 and Cre recombinases. Molecular Plant, 7, 1756–1765. 10.1093/mp/ssu107 [DOI] [PubMed] [Google Scholar]
- Jansing, J. , & Buyel, J. F. (2019). The correlation between DsRed mRNA levels and transient DsRed protein expression in plants depends on leaf age and the 5′ untranslated region. Biotechnology Journal, 14, e1800075 10.1002/biot.201800075 [DOI] [PubMed] [Google Scholar]
- Jefferson, R. A. , Kavanagh, T. A. , & Bevan, M. W. (1987). GUS fusions: beta‐glucuronidase as a sensitive and versatile gene fusion marker in higher plants. The EMBO Journal, 6, 3901–3907. PMCID: PMC553867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Z. , Xing, A. , Moon, B. P. , McCardell, R. P. , Mills, K. , & Falco, S. C. (2009). Site‐specific integration of transgenes in soybean via recombinase‐mediated DNA cassette exchange. Plant Physiology, 151, 1087–1095. 10.1104/pp.109.137612 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Z. , Moon, B. P. , Xing, A. , Liu, Z.‐B. , McCardell, R. P. , Damude, H. G. , & Falco, S. C. (2010). Stacking multiple transgenes at a selected genomic site via repeated recombinase‐mediated DNA cassette exchanges. Plant Physiology, 154, 622–631. 10.1104/pp.110.160093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, L. , Liu, Y.‐G. , Xu, X. , & Li, B. (2003). Efficient linking and transfer of multiple genes by a multigene assembly and transformation vector system. Proceedings of the National Academy of Sciences, 100, (10), 5962–5967. 10.1073/pnas.0931425100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livak, K. J. , & Schmittgen, T. D. (2001). Analysis of relative gene expression data using real‐time quantitative PCR and the 2− ΔΔCT method. Methods, 25, 402–408. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- Mann, D. G. J. , LaFayette, P. R. , Abercrombie, L. L. , King, Z. R. , Mazarei, M. , Halter, M. C. , … Neal Stewart Jr, C. (2012). Gateway‐compatible vectors for high‐throughput gene functional analysis in switchgrass (Panicum virgatum L.) and other monocot species. Plant Biotechnology Journal, 10, 226–236. 10.1111/j.1467-7652.2011.00658.x [DOI] [PubMed] [Google Scholar]
- Nandy, S. , & Srivastava, V. (2011). Site‐specific gene integration in rice genome mediated by the FLP‐FRT recombination system. Plant Biotechnology Journal, 9, 713–721. 10.1111/j.1467-7652.2010.00577.x [DOI] [PubMed] [Google Scholar]
- Nandy, S. , & Srivastava, V. (2012). Marker‐free site‐specific gene integration in rice based on the use of two recombination systems. Plant Biotechnology Journal, 10, 904–912. 10.1111/j.1467-7652.2012.00715.x [DOI] [PubMed] [Google Scholar]
- Nanto, K. , Sato, K. , Katayama, Y. , & Ebinuma, H. (2009). Expression of a transgene exchanged by the recombinase‐mediated cassette exchange (RMCE) method in plants. Plant Cell Reports, 28, 777–785. 10.1007/s00299-009-0683-5 [DOI] [PubMed] [Google Scholar]
- Naqvi, S. , Zhu, C. , Farre, G. , Ramessar, K. , Bassie, L. , Breitenbach, J. , … Christou, P. (2009). Transgenic multivitamin corn through biofortification of endosperm with three vitamins representing three distinct metabolic pathways. Proceedings of the National Academy of Sciences, 106, 7762–7767. 10.1073/pnas.0901412106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura, A. , Aichi, I. , & Matsuoka, M. (2006). A protocol for Agrobacterium‐mediated transformation in rice. Nature Protocols, 1, 2796 10.1038/nprot.2006.469 [DOI] [PubMed] [Google Scholar]
- Oh, S. J. , Song, S. I. , Kim, Y. S. , Jang, H. J. , Kim, S. Y. , Kim, M. , … Kim, J. K. (2005). Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiology, 138, 341–351. 10.1104/pp.104.059147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ow, D. W. (2003) Site‐specific gene stacking method In Vasil I. K. (Eds.), Plant biotechnol. 2002 and beyond. Dordrecht, the Netherlands: Springer; 10.1007/978-94-017-2679-5_41 [DOI] [Google Scholar]
- Ow, D. W. (2011). Recombinase‐mediated gene stacking as a transformation operating system. Journal of Integrative Plant Biology, 53, 512–519. 10.1111/j.1744-7909.2011.01061.x [DOI] [PubMed] [Google Scholar]
- Pathak, B. P. , Pruett, E. , Guan, H. , & Srivastava, V. (2019). Utility of I‐SceI and CCR5‐ZFN nucleases in excising selectable marker genes from transgenic plants. BMC Research Notes, 12, 272 10.1186/s13104-019-4304-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petolino, J. F. , & Kumar, S. (2016). Transgenic trait deployment using designed nucleases. Plant Biotechnology Journal, 14, 503–509. 10.1111/pbi.12457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puchta, H. , & Fauser, F. (2014). Synthetic nucleases for genome engineering in plants: Prospects for a bright future. The Plant Journal, 78, 727–741. 10.1111/tpj.12338 [DOI] [PubMed] [Google Scholar]
- Que, Q. , Chilton, M. D. , de Fontes, C. M. , He, C. , Nuccio, M. , Zhu, T. , … Shi, L. (2010). Trait stacking in transgenic crops: Challenges and opportunities. GM Crops, 1, 220–229. 10.4161/gmcr.1.4.13439 [DOI] [PubMed] [Google Scholar]
- Ream, T. S. , Strobel, J. , Roller, B. , Auger, D. L. , Kato, A. , Halbrook, C. , … Birchler, J. A. (2005). A test for ectopic exchange catalyzed by Cre recombinase in maize. Theoretical and Applied Genetics, 111, 378–385. 10.1007/s00122-005-2031-7 [DOI] [PubMed] [Google Scholar]
- Robinson, P. K. (2015). Enzymes: Principles and biotechnological applications. Essays in Biochemistry, 59, 1–41. 10.1042/bse0590001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz‐Lopez, N. , Haslam, R. P. , Usher, S. , Napier, J. A. , & Sayanova, O. (2015). An alternative pathway for the effective production of the omega‐3 long‐chain polyunsaturates EPA and ETA in transgenic oilseeds. Plant Biotechnology Journal, 13, 1264–1275. 10.1111/pbi.12328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacchetti, A. , Subramaniam, V. , Jovin, T. M. , & Alberti, S. (2002). Oligomerization of DsRed is required for the generation of a functional red fluorescent chromophore. FEBS Letters, 525, 13–19. 10.1016/S0014-5793(02)02874-0 [DOI] [PubMed] [Google Scholar]
- Saika, H. , Nishizawa‐Yokoi, A. , & Toki, S. (2014). The non‐homologous end‐joining pathway is involved in stable transformation in rice. Frontiers in Plant Science, 5, 560 10.3389/fpls.2014.00560 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt, M. A. , LaFayette, P. R. , Artelt, B. A. , & Parrott, W. A. (2008). A comparison of strategies for transformation with multiple genes via microprojectile‐mediated bombardment. In Vitro Cellular & Developmental Biology ‐ Plant, 44(3), 162–168. 10.1007/s11627-007-9099-5 [DOI] [Google Scholar]
- Somers, D. A. , & Makarevitch, I. (2004). Transgene integration in plants: Poking or patching holes in promiscuous genomes? Current Opinion in Biotechnology, 15, 126–131. 10.1016/j.copbio.2004.02.007 [DOI] [PubMed] [Google Scholar]
- Srivastava, V. , Ariza‐Nieto, M. , & Wilson, A. J. (2004). Cre‐mediated site‐specific gene integration for consistent transgene expression in rice. Plant Biotechnology Journal, 2, 169–179. 10.1111/j.1467-7652.2003.00061.x [DOI] [PubMed] [Google Scholar]
- Srivastava, V. , & Gidoni, D. (2010). Site‐specific gene integration technologies for crop improvement. . In Vitro Cellular & Developmental Biology ‐ Plant, 46, 219–232. 10.1007/s11627-009-9274-y [DOI] [Google Scholar]
- Srivastava, V. , & Nicholson, S. J. (2006). Cre/lox technologies for plant transformation. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources, 1(34), 1–12. 10.1079/pavsnnr20061034 [DOI] [Google Scholar]
- Srivastava, V. , & Ow, D. W. (2002). Biolistic mediated site‐specific integration in rice. Molecular Breeding, 8, 345–349. 10.1023/A:1015229015022 [DOI] [Google Scholar]
- Srivastava, V. , & Thomson, J. (2016). Gene stacking by recombinases. Plant Biotechnology Journal, 14, 471–482. 10.1111/pbi.12459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svitashev, S. , Young, J. K. , Schwartz, C. , Gao, H. , Falco, S. C. , & Cigan, A. M. (2015). Targeted mutagenesis, precise gene editing, and site‐specific gene insertion in maize using Cas9 and guide RNA. Plant Physiology, 169, 931–945. 10.1104/pp.15.00793 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomson, J. G. , & Ow, D. W. (2006). Site‐specific recombination systems for the genetic manipulation of eukaryotic genomes. Genesis, 44, 465–476. 10.1002/dvg.20237 [DOI] [PubMed] [Google Scholar]
- Vogel, C. , & Marcotte, E. (2012). Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nature Reviews Genetics, 13, 227–232. 10.1038/nrg3185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voytas, D. F. (2013). Plant genome engineering with sequence‐specific nucleases. Annual Review of Plant Biology, 64, 327–350. 10.1146/annurev-arplant-042811-105552 [DOI] [PubMed] [Google Scholar]
- Weeks, D. P. , Spalding, M. H. , & Yang, B. (2016). Use of designer nucleases for targeted gene and genome editing in plants. Plant Biotechnology Journal, 14, 483–495. 10.1111/pbi.12448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, X. , Luo, L. , Mo, B. , & Liu, L. (2019). Targeted genome engineering and its application in trait improvement of crop plants. Agricultural Science, 10, 1312–1342. 10.4236/as.2019.1010097 [DOI] [Google Scholar]
- Zhu, C. , Naqvi, S. , Breitenbach, J. , Sandmann, G. , Christou, P. , & Capell, T. (2008). Combinatorial genetic transformation generates a library of metabolic phenotypes for the carotenoid pathway in maize. Proceedings of the National Academy of Sciences of the United States of America, 105, 18232–18237. 10.1073/pnas.0809737105 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig S1‐S4
Table S1
Table S2