

Abstract
Even with substantial advances in cardiovascular therapy, the morbidity and mortality rates of diabetic cardiomyopathy (DCM) continually increase. Hence, a feasible therapeutic approach is urgently needed. Objectives. This work is aimed at systemically reviewing literature and addressing cell targets in DCM through the possible cardioprotection of G. lucidum through its antioxidant effects by using the Open Targets Platform (OTP) website. Methods. The OTP website version of 19.11 was accessed in December 2019 to identify the studies in DCM involving G. lucidum. Results. Among the 157 cell targets associated with DCM, the mammalian target of rapamycin (mTOR) was shared by all evidence, drug, and text mining data with 0.08 score association. mTOR also had the highest score association 0.1 with autophagy in DCM. Among the 1731 studies of indexed PubMed articles on G. lucidum published between 1985 and 2019, 33 addressed the antioxidant effects of G. lucidum and its molecular signal pathways involving oxidative stress and therefore were included in the current work. Conclusion. mTOR is one of the targets by DCM and can be inhibited by the antioxidative properties of G. lucidum directly via scavenging radicals and indirectly via modulating mTOR signal pathways such as Wnt signaling pathway, Erk1/2 signaling, and NF-κB pathways.
1. Introduction
Cardiovascular complications are associated with diabetes and lead to high mortality [1, 2]. Diabetic cardiomyopathy (DCM) is one of the main causes of heart injury and death in patients with diabetes. A total of 1.6 million deaths worldwide are directly attributed to diabetes every year [3]. Independent of coronary artery disease, DCM has increased prevalence during the last two decades and is experienced by 55% of patients with diabetes [4]. With diabetes being a global epidemic, the number of patients with DCM has increased. For the last two decades, the number of people with diabetes worldwide has increased from 151 million in 2000 to 425 million in 2017 and is estimated to increase to 629 million by 2045 [5]. The risk of developing DCM is higher for patients with diabetes than that for those without diabetes [6] and increases 2 to 4 times for those with more than a 10-year span of diabetes [7, 8]. Once DCM has developed, reducing its morbidity and mortality is difficult even with pharmacological improvement in terms of regulating blood glucose and insulin sensitivity. Clinical and preclinical investigations have examined the complexity of the pathophysiological consequences of DCM.
Clinical studies in patients with DCM reported that the pathological remodeling of the heart, which is characterized by left ventricular concentric hypertrophy and perivascular and interstitial fibrosis commencing to diastolic dysfunction and extended contraction and relaxation [9, 10], shortens ventricular ejection and increases wall stiffness [11, 12]. The influence of the diabetic condition on heart and cardiomyocyte function has been experimentally evaluated.
DCM and cardiac dysfunction are initiated in diabetic-induced experimental animals from 2 to 12 weeks [13]. Streptozotocin-induced diabetes in mice leads to the morphological changes of heart tissues, interstitial collagen deposition, cardiac hypertrophy, fibrosis, and remarkable elevation of paracrine of angiotensin II level in myocardium and NADPH oxidase activities, which are considered the primary source of free radicals in the cardiomyocytes of diabetic heart [14]. Connective tissue growth factor mediates cardiac fibrosis in diabetes [13, 15]. In diabetic mice with cardiomyopathy, the expression of sarcoplasmic reticulum calcium ATPase and [Ca+2] ion transient is reduced [16]. Sarcoplasmic reticulum calcium ATPase is a primary cardiac isoform of calcium pump transporting calcium from cytoplasm to sarcoplasmic reticulum during diastolic relaxation [17].
Even with substantial advances in cardiovascular therapy, diabetic morbidity and mortality rate are continually increasing, and a feasible therapeutic approach for DCM is still lacking. Exploring the medication targets for DCM may further identify novel drugs and improve specific therapies for DCM. Therapeutic targets for DCM with natural resources are considered as one of the main reservoirs for drug discovery. Therefore, novel therapeutics for a range of targets must be developed to prevent DCM progression. This study identifies molecular target involvement and its association with DCM by using the Open Targets Platform (OTP) website established by Biogen, EMBL European Bioinformatics Institute, GlaxoSmithKline, and Wellcome Trust Sanger Institute. The OTP provides comprehensive and up-to-date data for drug molecular targets associated with relative diseases. Oxidative stress (OS) may be a key factor in the molecular and cellular mechanisms of diabetes-induced DCM [18]. Hence, targeting OS-related processes could be a promising therapeutic strategy for DCM.
Ganoderma (G.) lucidum, which is known in Chinese as “Lingzhi,” is a medicinal mushroom commonly used as a Chinese herbal medicine and the main ingredient in many conventional combinations or dietary supplements [19]. This name has been proposed by Petter Adolf Karsten from England in the late 19th century and has been applied in various places such as Asia, Africa, Oceania, and Europe [20]. Lingzhi has been widely cultivated in China and has a long history as a traditional Chinese medicine. Chinese G. lucidum exhibits high variability of basidioma morphology and more or less consistency in its microscopic characters, e.g., short clavate cutis elements, Bovista-type ligative hyphae, and strongly echinulated basidiospores [21]. G. lucidum also contains various bioactive compounds, such as flavonoids, ganoderic acid, phenolics, and polysaccharides [21], that can treat many chronic diseases including diabetes and its complications by counteracting OS. Preclinical studies reported the beneficial effects of G. lucidum against OS-induced diseases, its liver protection against CCl4-induced OS [22], skin protection against croton oil-induced lipid peroxidation in mice [23], and thymus and spleen protection against 5-fluorouracil-induced OS in mice [21]. This systematic review is aimed at discussing the potential cell targets and cardioprotective pathway of G. lucidum based on preclinical and clinical investigations.
2. Methods
The OTP website version 19.11 (OTP V 19.11) was used to prioritize and identify the targets associated with DCM. The OTP provides score and rank target-disease associations and integrates evidence from six resources, including genetics, genomics, transcriptomics, drugs, animal models, and scientific literature [24, 25]. Two main steps of searching were performed in December 2019. In the first step, the term “diabetic cardiomyopathy” was used, and all the targets associated with DCM were listed according to available evidence recorded through bioinformatic processing, including data evidence of drug, text mining, genetic association, somatic mutation, pathways and signals, RNA signal, and animal model. The resulting targets with the highest association with DCM from the first step were used to further search for evidence on G. lucidum cardioprotection.
This systematic review on the antioxidant activity of G. lucidum was described as follows. Abstracts published from 1985 to July 2019 were reported as guided by the Preferred Reporting Items for Systematic Reviews and Meta-Analyses [26] (Figure 1). The key terms used were G. lucidum and spore of G. lucidum. In this step, the studies were divided into seven 5-year periods to easily read and select related abstracts. The search was limited to studies published in English and Chinese languages. Inclusion criteria were as follows: studies must focus on (1) G. lucidum and its (2) antioxidant, antidiabetic, and cardioprotective activities. Exclusion criteria were as follows: studies focusing on (1) mushrooms other than G. lucidum and (2) not related to its antioxidant activities such as the botanical and genetic studies of G. lucidum.
Figure 1.
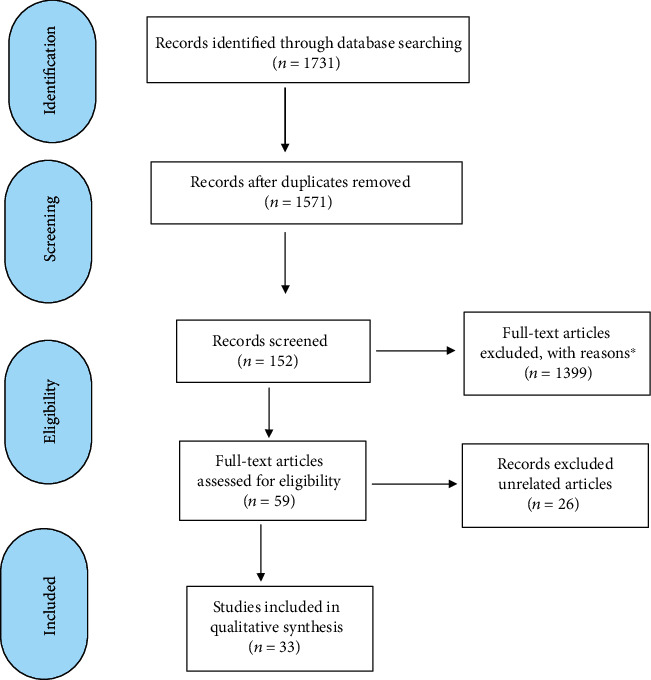
A PRISMA flow diagram summarising the study selection process. Antioxidant of G. Lucidum; PRISMA: Preferred Reporting Items for Systematic Reviews and Meta-Analyses. ∗After exclusion of other antioxidant activity studies of G. Lucidum.
3. Results
3.1. Targets Associated with DCM in Diabetes Integrated by OTP
3.1.1. DCM and Its Associated Targets
A total of 309 targets were associated with DCM based on evidence from drug and text mining data with overall association scores from 0.004 to 0.177 (Table 1, supplementary file (available here)). Among the selected drug data, only two targets, namely, carnitine palmitoyltransferase 1B (CPT1B) and 2 (CPT2) were associated with DCM with 0.1 score association. A total of 306 targets were identified from text mining. Only the mechanistic target of rapamycin kinase (mTOR) was common in both types of data. A total of 309 targets were expressed in 32 tissue organs including the heart and were involved in 19 pathway types (Table 2, supplementary file (available here)). Among these 309 targets, 155 were expressed in the heart tissues with overall association scores ranging from 0.007 to 0.177 (Table 1). Among the 19 pathways, 4 targets were included in autophagy (Table 2), namely, mTOR, beclin 1 (BECN1), parkin RBR E3 ubiquitin protein ligase (PRKN), and voltage-dependent anion channel 1 (VDAC1) with scores of 0.1, 0.06, 0.05, and 0.03, respectively.
Table 1.
Association sore of 155 targets associated with diabetic cardiomyopathy in heart tissue.
| Target name | Target symbol | Association score | |||
|---|---|---|---|---|---|
| Data types Known drug |
Data types Literature |
Overall | |||
| 1 | Tripartite motif containing 55 | TRIM55 | 0 | 0.177 | 0.177 |
| 2 | Peroxisome proliferator-activated receptor alpha | PPARA | 0 | 0.117 | 0.117 |
| 3 | Mechanistic target of rapamycin kinase | MTOR | 0.1 | 0.054 | 0.113 |
| 4 | Interleukin 6 | IL-6 | 0 | 0.113 | 0.113 |
| 5 | Carnitine palmitoyltransferase 1B | CPT1B | 0.1 | 0.000 | 0.100 |
| 6 | Carnitine palmitoyltransferase 2 | CPT2 | 0.1 | 0.000 | 0.100 |
| 7 | Tripartite motif containing 54 | TRIM54 | 0 | 0.081 | 0.081 |
| 8 | Nuclear factor, erythroid 2 like 2 | NFE2L2 | 0 | 0.072 | 0.072 |
| 9 | Hydroxysteroid 11-beta dehydrogenase 1 | HSD11B1 | 0 | 0.070 | 0.070 |
| 10 | Fibroblast growth factor 1 | FGF1 | 0 | 0.070 | 0.070 |
| 11 | Colony-stimulating factor 3 | CSF3 | 0 | 0.062 | 0.062 |
| 12 | Beclin 1 | BECN1 | 0 | 0.062 | 0.062 |
| 13 | Cytochrome P450 family 2 subfamily J member 2 | CYP2J2 | 0 | 0.061 | 0.061 |
| 14 | Angiotensin I-converting enzyme 2 | ACE2 | 0 | 0.060 | 0.060 |
| 15 | Aldehyde dehydrogenase 2 family member | ALDH2 | 0 | 0.059 | 0.059 |
| 16 | Glycogen synthase kinase 3 beta | GSK3B | 0 | 0.057 | 0.057 |
| 17 | Gelsolin | GSN | 0 | 0.055 | 0.055 |
| 18 | Toll-like receptor 2 | TLR2 | 0 | 0.054 | 0.054 |
| 19 | Parkin RBR E3 ubiquitin protein ligase | PRKN | 0 | 0.054 | 0.054 |
| 20 | Apelin | APLN | 0 | 0.053 | 0.053 |
| 21 | ST3 beta-galactoside alpha-2,3-sialyltransferase 4 | ST3GAL4 | 0 | 0.052 | 0.052 |
| 22 | Peroxisome proliferator activated receptor gamma | PPARG | 0 | 0.052 | 0.052 |
| 23 | Corin, serine peptidase | CORIN | 0 | 0.052 | 0.052 |
| 24 | Titin | TTN | 0 | 0.049 | 0.049 |
| 25 | Angiogenin | ANG | 0 | 0.049 | 0.049 |
| 26 | Protein kinase D1 | PRKD1 | 0 | 0.049 | 0.049 |
| 27 | PPARG coactivator 1 alpha | PPARGC1A | 0 | 0.048 | 0.048 |
| 28 | Vascular endothelial growth factor A | VEGFA | 0 | 0.048 | 0.048 |
| 29 | Insulin-like growth factor 1 | IGF1 | 0 | 0.047 | 0.047 |
| 30 | CD36 molecule | CD36 | 0 | 0.047 | 0.047 |
| 31 | Nitric oxide synthase 3 | NOS3 | 0 | 0.046 | 0.046 |
| 32 | Apolipoprotein A1 | APOA1 | 0 | 0.044 | 0.044 |
| 33 | Gap junction protein alpha 1 | GJA1 | 0 | 0.041 | 0.041 |
| 34 | Calsequestrin 2 | CASQ2 | 0 | 0.041 | 0.041 |
| 35 | Decorin | DCN | 0 | 0.040 | 0.040 |
| 36 | Urocortin | UCN | 0 | 0.040 | 0.040 |
| 37 | Cellular communication network factor 2 | CCN2 | 0 | 0.040 | 0.040 |
| 38 | Matrix metallopeptidase 2 | MMP2 | 0 | 0.040 | 0.040 |
| 39 | Periostin | POSTN | 0 | 0.039 | 0.039 |
| 40 | Fibroblast growth factor 2 | FGF2 | 0 | 0.039 | 0.039 |
| 41 | BCL6 transcription repressor | BCL6 | 0 | 0.039 | 0.039 |
| 42 | Tax1-binding protein 1 | TAX1BP1 | 0 | 0.038 | 0.038 |
| 43 | Solute carrier family 2 member 4 | SLC2A4 | 0 | 0.038 | 0.038 |
| 44 | Rho-associated coiled-coil containing protein kinase 2 | ROCK2 | 0 | 0.037 | 0.037 |
| 45 | NADPH oxidase 4 | NOX4 | 0 | 0.036 | 0.036 |
| 46 | Mitogen-activated protein kinase 9 | MAPK9 | 0 | 0.036 | 0.036 |
| 47 | Insulin-like growth factor 2 | IGF2 | 0 | 0.036 | 0.036 |
| 48 | Angiotensin II receptor type 2 | AGTR2 | 0 | 0.036 | 0.036 |
| 49 | Lipoprotein lipase | LPL | 0 | 0.036 | 0.036 |
| 50 | Insulin receptor | INSR | 0 | 0.035 | 0.035 |
| 51 | Angiopoietin 1 | ANGPT1 | 0 | 0.035 | 0.035 |
| 52 | Interleukin 33 | IL33 | 0 | 0.035 | 0.035 |
| 53 | Caveolin 3 | CAV3 | 0 | 0.034 | 0.034 |
| 54 | Angiotensin I-converting enzyme | ACE | 0 | 0.034 | 0.034 |
| 55 | Patatin-like phospholipase domain containing 2 | PNPLA2 | 0 | 0.034 | 0.034 |
| 56 | ATPase sarcoplasmic/endoplasmic reticulum Ca2+ transporting 2 | ATP2A2 | 0 | 0.033 | 0.033 |
| 57 | Glucokinase | GCK | 0 | 0.032 | 0.032 |
| 58 | Dimethylarginine dimethylaminohydrolase 2 | DDAH2 | 0 | 0.032 | 0.032 |
| 59 | Xenotropic and polytropic retrovirus receptor 1 | XPR1 | 0 | 0.032 | 0.032 |
| 60 | Vascular endothelial growth factor B | VEGFB | 0 | 0.032 | 0.032 |
| 61 | Phosphodiesterase 5A | PDE5A | 0 | 0.031 | 0.031 |
| 62 | MAPK-activated protein kinase 2 | MAPKAPK2 | 0 | 0.031 | 0.031 |
| 63 | Heat shock protein family E (Hsp10) member 1 | HSPE1 | 0 | 0.031 | 0.031 |
| 64 | Sirtuin 2 | SIRT2 | 0 | 0.031 | 0.031 |
| 65 | DIRAS family GTPase 3 | DIRAS3 | 0 | 0.030 | 0.030 |
| 66 | SMAD family member 3 | SMAD3 | 0 | 0.030 | 0.030 |
| 67 | Dual specificity phosphatase 5 | DUSP5 | 0 | 0.030 | 0.030 |
| 68 | Kruppel-like factor 4 | KLF4 | 0 | 0.030 | 0.030 |
| 69 | Ryanodine receptor 2 | RYR2 | 0 | 0.029 | 0.029 |
| 70 | Prohibitin | PHB | 0 | 0.029 | 0.029 |
| 71 | Estrogen related receptor gamma | ESRRG | 0 | 0.028 | 0.028 |
| 72 | Nebulin | NEB | 0 | 0.028 | 0.028 |
| 73 | Peroxiredoxin 3 | PRDX3 | 0 | 0.028 | 0.028 |
| 74 | Adrenoceptor beta 2 | ADRB2 | 0 | 0.028 | 0.028 |
| 75 | Solute carrier family 9 member A1 | SLC9A1 | 0 | 0.028 | 0.028 |
| 76 | Transglutaminase 2 | TGM2 | 0 | 0.027 | 0.027 |
| 77 | Poly(ADP-ribose) polymerase 1 | PARP1 | 0 | 0.027 | 0.027 |
| 78 | Insulin receptor substrate 1 | IRS1 | 0 | 0.027 | 0.027 |
| 79 | Voltage dependent anion channel 1 | VDAC1 | 0 | 0.026 | 0.026 |
| 80 | AKT serine/threonine kinase 1 | AKT1 | 0 | 0.025 | 0.025 |
| 81 | Myocyte enhancer factor 2A | MEF2A | 0 | 0.025 | 0.025 |
| 82 | Dual specificity phosphatase 1 | DUSP1 | 0 | 0.025 | 0.025 |
| 83 | Musculin | MSC | 0 | 0.025 | 0.025 |
| 84 | Diacylglycerol kinase zeta | DGKZ | 0 | 0.024 | 0.024 |
| 85 | Death associated protein kinase 2 | DAPK2 | 0 | 0.024 | 0.024 |
| 86 | Solute carrier family 25 member 4 | SLC25A4 | 0 | 0.023 | 0.023 |
| 87 | SMAD family member 7 | SMAD7 | 0 | 0.023 | 0.023 |
| 88 | Natriuretic peptide A | NPPA | 0 | 0.023 | 0.023 |
| 89 | Coiled-coil domain containing 47 | CCDC47 | 0 | 0.022 | 0.022 |
| 90 | Lipase E, hormone sensitive type | LIPE | 0 | 0.022 | 0.022 |
| 91 | Leptin | LEP | 0 | 0.022 | 0.022 |
| 92 | Arylsulfatase A | ARSA | 0 | 0.021 | 0.021 |
| 93 | Nitric oxide synthase 2 | NOS2 | 0 | 0.021 | 0.021 |
| 94 | Nuclear receptor subfamily 3 group C member 2 | NR3C2 | 0 | 0.021 | 0.021 |
| 95 | Sirtuin 3 | SIRT3 | 0 | 0.021 | 0.021 |
| 96 | Plasminogen | PLG | 0 | 0.020 | 0.020 |
| 97 | Spindlin 1 | SPIN1 | 0 | 0.020 | 0.020 |
| 98 | Serpin family E member 1 | SERPINE1 | 0 | 0.020 | 0.020 |
| 99 | Tachykinin receptor 1 | TACR1 | 0 | 0.020 | 0.020 |
| 100 | RNA binding fox-1 homolog 2 | RBFOX2 | 0 | 0.020 | 0.020 |
| 101 | Fatty acid binding protein 4 | FABP4 | 0 | 0.019 | 0.019 |
| 102 | Potassium voltage-gated channel subfamily H member 2 | KCNH2 | 0 | 0.019 | 0.019 |
| 103 | Cell adhesion molecule 1 | CADM1 | 0 | 0.019 | 0.019 |
| 104 | Prolylcarboxypeptidase | PRCP | 0 | 0.018 | 0.018 |
| 105 | Nucleotide-binding oligomerization domain containing 1 | NOD1 | 0 | 0.018 | 0.018 |
| 106 | Activating transcription factor 3 | ATF3 | 0 | 0.018 | 0.018 |
| 107 | Vasoactive intestinal peptide | VIP | 0 | 0.018 | 0.018 |
| 108 | Egl-9 family hypoxia inducible factor 3 | EGLN3 | 0 | 0.018 | 0.018 |
| 109 | Fibronectin 1 | FN1 | 0 | 0.018 | 0.018 |
| 110 | Endothelin 1 | EDN1 | 0 | 0.018 | 0.018 |
| 111 | C-C motif chemokine ligand 2 | CCL2 | 0 | 0.018 | 0.018 |
| 112 | Solute carrier family 5 member 1 | SLC5A1 | 0 | 0.018 | 0.018 |
| 113 | Fibrinogen-like 2 | FGL2 | 0 | 0.017 | 0.017 |
| 114 | Monoamine oxidase A | MAOA | 0 | 0.017 | 0.017 |
| 115 | Sphingosine-1-phosphate receptor 1 | S1PR1 | 0 | 0.017 | 0.017 |
| 116 | Signal transducer and activator of transcription 3 | STAT3 | 0 | 0.017 | 0.017 |
| 117 | Toll-like receptor 3 | TLR3 | 0 | 0.017 | 0.017 |
| 118 | Tripartite motif containing 63 | TRIM63 | 0 | 0.017 | 0.017 |
| 119 | TIMP metallopeptidase inhibitor 2 | TIMP2 | 0 | 0.017 | 0.017 |
| 120 | Nerve growth factor | NGF | 0 | 0.017 | 0.017 |
| 121 | Natriuretic peptide receptor 2 | NPR2 | 0 | 0.016 | 0.016 |
| 122 | Cyclin-dependent kinase inhibitor 1A | CDKN1A | 0 | 0.016 | 0.016 |
| 123 | Cathepsin D | CTSD | 0 | 0.016 | 0.016 |
| 124 | Thrombospondin 1 | THBS1 | 0 | 0.015 | 0.015 |
| 125 | Kinase insert domain receptor | KDR | 0 | 0.015 | 0.015 |
| 126 | Serine/threonine kinase 11 | STK11 | 0 | 0.015 | 0.015 |
| 127 | Enolase 3 | ENO3 | 0 | 0.015 | 0.015 |
| 128 | Gasdermin D | GSDMD | 0 | 0.015 | 0.015 |
| 129 | Cytochrome c, somatic | CYCS | 0 | 0.015 | 0.015 |
| 130 | Kallikrein B1 | KLKB1 | 0 | 0.015 | 0.015 |
| 131 | TIMP metallopeptidase inhibitor 4 | TIMP4 | 0 | 0.015 | 0.015 |
| 132 | Transforming growth factor beta 3 | TGFB3 | 0 | 0.015 | 0.015 |
| 133 | Zinc finger and BTB domain containing 16 | ZBTB16 | 0 | 0.015 | 0.015 |
| 134 | Collagen type I alpha 1 chain | COL1A1 | 0 | 0.015 | 0.015 |
| 135 | Endothelin receptor type A | EDNRA | 0 | 0.014 | 0.014 |
| 136 | Cellular communication network factor 1 | CCN1 | 0 | 0.014 | 0.014 |
| 137 | Secreted protein acidic and cysteine rich | SPARC | 0 | 0.014 | 0.014 |
| 138 | Glucagon like peptide 1 receptor | GLP1R | 0 | 0.014 | 0.014 |
| 139 | Cystatin C | CST3 | 0 | 0.014 | 0.014 |
| 140 | Intercellular adhesion molecule 1 | ICAM1 | 0 | 0.014 | 0.014 |
| 141 | Elastin | ELN | 0 | 0.014 | 0.014 |
| 142 | Tenascin C | TNC | 0 | 0.014 | 0.014 |
| 143 | PTEN-induced kinase 1 | PINK1 | 0 | 0.014 | 0.014 |
| 144 | Calpastatin | CAST | 0 | 0.014 | 0.014 |
| 145 | CCAAT enhancer binding protein beta | CEBPB | 0 | 0.012 | 0.012 |
| 146 | Acyl-coA thioesterase 1 | ACOT1 | 0 | 0.012 | 0.012 |
| 147 | G protein-coupled bile acid receptor 1 | GPBAR1 | 0 | 0.010 | 0.010 |
| 148 | Annexin A1 | ANXA1 | 0 | 0.010 | 0.010 |
| 149 | Apolipoprotein L2 | APOL2 | 0 | 0.008 | 0.008 |
| 150 | Natriuretic peptide B | NPPB | 0 | 0.008 | 0.008 |
| 151 | Leptin receptor | LEPR | 0 | 0.008 | 0.008 |
| 152 | Serum response factor | SRF | 0 | 0.008 | 0.008 |
| 153 | Heat shock protein family B (small) member 3 | HSPB3 | 0 | 0.007 | 0.007 |
| 154 | Angiotensin II receptor type 1 | AGTR1 | 0 | 0.007 | 0.007 |
| 155 | Protein phosphatase 5 catalytic subunit | PPP5C | 0 | 0.007 | 0.007 |
Table 2.
Nineteen pathway types involved in the heart tissues of 155 targets expressed.
| No. | Pathway (No. of targets) |
|---|---|
| 1. | Signal transduction (63) |
| 2. | Immune system (47) |
| 3. | Metabolism of proteins (39) |
| 4. | Metabolism (31) |
| 5. | Gene expression (transcription) (25) |
| 6. | Hemostasis (23) |
| 7. | Disease (22) |
| 8. | Developmental biology (20) |
| 9. | Extracellular matrix organization (18) |
| 10. | Cellular responses to external stimuli (14) |
| 11. | Transport of small molecules (11) |
| 12. | Muscle contraction (11) |
| 13. | Vesicle-mediated transport (10) |
| 14. | Organelle biogenesis and maintenance (4) |
| 15. | Programmed cell death (4) |
| 16. | Autophagy (4) |
| 17. | Neuronal system (3) |
| 18. | Cell cycle (3) |
| 19. | Circadian clock (3) |
mTOR was further investigated, and its association with heart diseases ranged from 0.0004 to 0.8588, which is the overall association score for 49 subtypes of heart diseases. mTOR had 0.1 and 0.8 overall association scores with DCM and hypertrophic cardiomyopathy, respectively (Table 3).
Table 3.
mTOR score association with 49 heart diseases.
| No. | Heart disease | Association score | ||||
|---|---|---|---|---|---|---|
| Data types Genetic |
Data types Known drug |
Data types Literature |
Data types Animal model |
Overall | ||
| 1 | Heart disease | 0.00041 | 0.79550 | 0.14161 | 0.19028 | 0.8588 |
| 2 | Cardiomyopathy | 0.00000 | 0.77847 | 0.11930 | 0.19028 | 0.8393 |
| 3 | Hypertrophic cardiomyopathy | 0.00000 | 0.77222 | 0.10214 | 0.00000 | 0.7978 |
| 4 | Heart failure | 0.00000 | 0.25000 | 0.05235 | 0.00000 | 0.2631 |
| 5 | Dilated cardiomyopathy | 0.00000 | 0.00000 | 0.07568 | 0.19028 | 0.2092 |
| 6 | Congestive heart failure | 0.00000 | 0.20000 | 0.02636 | 0.00000 | 0.2066 |
| 7 | Diastolic heart failure | 0.00000 | 0.20000 | 0.00000 | 0.00000 | 0.2000 |
| 8 | Barth syndrome | 0.00000 | 0.00000 | 0.00000 | 0.19028 | 0.1903 |
| 9 | Coronary heart disease | 0.00000 | 0.00000 | 0.12009 | 0.00000 | 0.1201 |
| 10 | Diabetic cardiomyopathy | 0.00000 | 0.10000 | 0.05391 | 0.00000 | 0.1135 |
| 11 | Coronary artery disease | 0.00000 | 0.00000 | 0.10961 | 0.00000 | 0.1096 |
| 12 | Systemic scleroderma | 0.00000 | 0.00000 | 0.09914 | 0.00000 | 0.0991 |
| 13 | Cardiotoxicity | 0.00000 | 0.00000 | 0.09016 | 0.00000 | 0.0902 |
| 14 | Glycogen storage disease due to acid maltase deficiency | 0.00000 | 0.00000 | 0.08380 | 0.00000 | 0.0838 |
| 15 | Myocardial infarction | 0.00000 | 0.00000 | 0.06467 | 0.00000 | 0.0647 |
| 16 | Persistent truncus arteriosus | 0.00000 | 0.00000 | 0.06144 | 0.00000 | 0.0614 |
| 17 | Heart neoplasm | 0.00000 | 0.00000 | 0.06126 | 0.00000 | 0.0613 |
| 18 | Emery-Dreifuss muscular dystrophy | 0.00000 | 0.00000 | 0.05780 | 0.00000 | 0.0578 |
| 19 | Ischemia reperfusion injury | 0.00000 | 0.00000 | 0.05702 | 0.00000 | 0.0570 |
| 20 | Myocardial ischemia | 0.00000 | 0.00000 | 0.05658 | 0.00000 | 0.0566 |
| 21 | Carney complex | 0.00000 | 0.00000 | 0.05494 | 0.00000 | 0.0549 |
| 22 | Down syndrome | 0.00000 | 0.00000 | 0.05488 | 0.00000 | 0.0549 |
| 23 | Cardiac rhabdomyoma | 0.00000 | 0.00000 | 0.05475 | 0.00000 | 0.0547 |
| 24 | Autosomal dominant Emery-Dreifuss muscular dystrophy | 0.00000 | 0.00000 | 0.05280 | 0.00000 | 0.0528 |
| 25 | Polyarteritis nodosa | 0.00000 | 0.00000 | 0.04343 | 0.00000 | 0.0434 |
| 26 | Steinert myotonic dystrophy | 0.00000 | 0.00000 | 0.04273 | 0.00000 | 0.0427 |
| 27 | Acute myocardial infarction | 0.00000 | 0.00000 | 0.03798 | 0.00000 | 0.0380 |
| 28 | Cardiac arrhythmia | 0.00041 | 0.00000 | 0.03721 | 0.00000 | 0.0373 |
| 29 | Myocarditis | 0.00000 | 0.00000 | 0.03263 | 0.00000 | 0.0326 |
| 30 | Duchenne muscular dystrophy | 0.00000 | 0.00000 | 0.03253 | 0.00000 | 0.0325 |
| 31 | Gaucher disease | 0.00000 | 0.00000 | 0.03230 | 0.00000 | 0.0323 |
| 32 | Cardiac arrest | 0.00000 | 0.00000 | 0.02847 | 0.00000 | 0.0285 |
| 33 | Atrial fibrillation | 0.00000 | 0.00000 | 0.02720 | 0.00000 | 0.0272 |
| 34 | Aortic stenosis | 0.00000 | 0.00000 | 0.01910 | 0.00000 | 0.0191 |
| 35 | Acute coronary syndrome | 0.00000 | 0.00000 | 0.01900 | 0.00000 | 0.0190 |
| 36 | Sleep disorder | 0.00000 | 0.00000 | 0.01840 | 0.00000 | 0.0184 |
| 37 | Williams syndrome | 0.00000 | 0.00000 | 0.01640 | 0.00000 | 0.0164 |
| 38 | Supravalvular aortic stenosis | 0.00000 | 0.00000 | 0.01640 | 0.00000 | 0.0164 |
| 39 | Autoimmune myocarditis | 0.00000 | 0.00000 | 0.01560 | 0.00000 | 0.0156 |
| 40 | Friedreich ataxia | 0.00000 | 0.00000 | 0.01480 | 0.00000 | 0.0148 |
| 41 | Obstructive sleep apnea | 0.00000 | 0.00000 | 0.01480 | 0.00000 | 0.0148 |
| 42 | PHACE syndrome | 0.00000 | 0.00000 | 0.01440 | 0.00000 | 0.0144 |
| 43 | Glycogen storage disease due to LAMP-2 deficiency | 0.00000 | 0.00000 | 0.01440 | 0.00000 | 0.0144 |
| 44 | Idiopathic pulmonary arterial hypertension | 0.00000 | 0.00000 | 0.01400 | 0.00000 | 0.0140 |
| 45 | Fabry disease | 0.00000 | 0.00000 | 0.01340 | 0.00000 | 0.0134 |
| 46 | Becker muscular dystrophy | 0.00000 | 0.00000 | 0.00840 | 0.00000 | 0.0084 |
| 47 | Hemopericardium | 0.00000 | 0.00000 | 0.00720 | 0.00000 | 0.0072 |
| 48 | Aortic coarctation | 0.00000 | 0.00000 | 0.00680 | 0.00000 | 0.0068 |
| 49 | Atrial flutter | 0.00041 | 0.00000 | 0.00000 | 0.00000 | 0.0004 |
mTOR is a serine/threonine-protein kinase playing as a central regulator of cellular metabolism, growth, and survival in response to hormone growth factor [27], nutrients, energy, and stress signals [28, 29]. According to UniPort, mTOR can be found in different subcellular locations including the membranes of endoplasmic reticulum, Golgi apparatus, outer mitochondrion, microsome, and lysosome; lysosome, cytoplasm, nucleus, and PML nuclear body. The RNA and protein expression levels of mTOR are present in several organs including the heart, e.g., the medium RNA and high protein levels of mTOR are expressed in the left ventricle, atrium, and coronary artery but not in the heart muscles (Figure 2).
Figure 2.
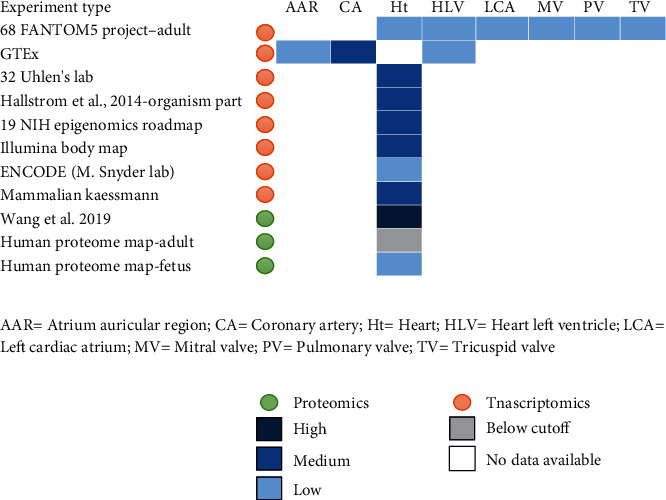
mRNA and protein baseline expression of mTOR in the heart.
3.1.2. Evidence on the Cardioprotection of G. lucidum
A total of 1731 articles were identified (Figure 3) and further divided into seven 5-year time periods. The first period ranged from 1985 to 1989, and the last period ranged from August 2018 to August 2019 (Figure 1). These articles were reviewed in the following three phases. First, 1571 articles remained after the duplicated ones were removed. Second, articles that did not satisfy the inclusion criteria based solely on their titles (remaining 1399 articles) and abstracts (remaining 59 articles) were excluded. Lastly, the remaining articles were scanned, and those that did not meet our inclusion criteria were excluded. After the initial screening of titles and abstracts, the 59 remaining articles were screened for the second time by two individual reviewers. Inclusion of full articles was agreed upon by two reviewers prior to data extraction. Finally, 33 studies were considered eligible for the review (Figure 2). In this section, the collected pieces of evidence were divided into two main parts, namely, the in vivo antioxidant of G. lucidum (14 studies, Table 4), in which the in vivo effect of antioxidant on the parameters related to OS was discussed, and the in vitro antioxidant of G. lucidum (19 studies, Table 5), in which the in vitro effect of antioxidant activities and possible molecular mechanisms was elaborated.
Figure 3.
Number of studies on G. lucidum during 1985-2019.
Table 4.
In vivo studies of G. lucidum.
| No. | Animal | Form | Dosage (mg/kg) | Antioxidant parameters | Biological activity | Pathway | References |
|---|---|---|---|---|---|---|---|
| 1 | CCl4-induced acute liver injury mice | GLPS | 100 - 150 | NOS CYP2E1 MDA, GSH | Suppressing free radical lipid peroxidation | Decreasing of the protein expression levels of NLRP3, ASC, and caspase-1 in acute liver injury. ASC (apoptosis-associated speck-like protein) NLRP3 (NOD-like receptor 3) Caspase-1 GAPDH (glyceraldehyde-3-phosphate hydrogenase |
[22] |
|
| |||||||
| 2 | Croton oil applied skin edema in rats | Ethanol extract of sporocarps | 500 and 1000 mg/kg | Antiperoxidative, anti-inflammatory, and antimutagenic activities | Direct anti-inflammatory and free radical scavenging properties of the extract | [23] | |
|
| |||||||
| 3 | Photoreceptor cell lesions induced by N-methyl-N-nitrosourea (MNU) in female SD arts | Ganoderma spore lipid (GSL) | 500, 1000, 2000, and 4000 mg/kg | Expressions of Bax, Bcl-xl, and caspase-3 | Improve A-wave amplitude (μv) decreased apoptosis levels | Regulate the expressions of Bax, Bcl-xl, and caspases-3, inhibiting MNU-induced rat, photoreceptor cell apoptosis, and protecting retinal function | [30] |
|
| |||||||
| 4 | A carotid-artery-ligation mouse model | Ganoderma triterpenoid (GT) | 300 mg/kg/day | Intimal hyperplasia structural changes VCAM-1, TNF-α, and IL-6 | Atheroprotective properties | Endothelin-1, von Willebrand factor, and monocyte chemoattractant protein-1 | [33] |
|
| |||||||
| 5 | Swimming-induced oxidative stress in skeletal muscle mice | GLPS | 50, 100, and 200 mg/kg | SOD, GPX, and CAT activities as well as by the MDA levels | Attenuates exercise-induced oxidative stress in skeletal muscle | Increasing antioxidant enzyme activities and decrease the MDA levels. Protective effects against exhaustive exercise-induced oxidative stress | [32] |
|
| |||||||
| 6 | Rat gastric cancer model | GLPS | 400-800 mg/kg for 20 weeks | SOD, CAT, and GSH-Px | Antioxidant | Induced the levels of serum IL-6 and TNF-α levels and increased the levels of serum IL-2, IL-4, and IL-10 in GLP-treated rats compared to gastric cancer model rats | [37] |
|
| |||||||
| 7 | BALB/c female mice | GLPS i.p. daily | 50 mg/kg, 100 mg/kg, and 200 mg/kg | SOD and GSH-Px | Antioxidant | Improved immunity in mice. Increased thymus and spleen index; improved SOD and GSH-Px contents in the mice body | [31] |
|
| |||||||
| 8 | T2DM rats | GLPS | 200, 400, and 800 mg × kg−1 for 16 weeks | NO, SOD, MDA, GSH-Px, and CAT MDA in cardiac tissue | Antioxidation in cardiac tissue of T2DM rats | Reduce MDA in cardiac tissue and improve the myocardial ultrastructure | [34] |
|
| |||||||
| 9 | Male BALB/c mice (age19-21 months) (aged mice) | Ethanolic extract of G. lucidum | 50 and 250 mg/kg, once daily for 15 days | GSH Mn-SOD, GPx, and GST | Antioxidant in heart tissues | Elevated the levels of GSH as well as activities of MnSOD, GPx, and GST and decreased significantly the levels of lipid peroxidation, AOPP, and ROS. Improve the age-related decline of antioxidant status which was partly ascribed to free radical scavenging activity | [38] |
|
| |||||||
| 10 | B16 mouse melanoma | Methanol extract containing total terpenoids (GLme) and a purified methanol extract containing mainly acidic terpenoids (GLpme) | A daily i.p. injection of 100 mg/kg body weight (b.w.) | Production of oxygen radical caspase-dependent apoptotic cell death-mediated production of reactive oxygen species | Anticancer | The mechanism of antitumor activity of GLme comprised inhibition of cell proliferation and induction of caspase-dependent apoptotic cell death mediated by upregulated p53 and inhibited Bcl-2 expression | [86] |
|
| |||||||
| 11 | With non-insulin-dependent diabetes mellitus (NIDDM) | Ganoderma lucidum spores | 250 mg/kg × d, for 10 | Xanthine oxidase (XOD), myeloperoxidase (MPO), and mitochondrial succinate dehydrogenase (SDH) in the testis | Reducing free radical-induceddamage to the testicular tissue | Protect the testis of diabetic rats by reducing free radical-induced damage to the testicular tissue and enhancing the activity of SDH | [35] |
|
| |||||||
| 12 | Epididymal cells of type 2 diabetes rats | Ganoderma lucidum spores (GLS) | 250 mg/kg × d, for 10 weeks | Contents of mitochondrial calcium & cytochrome C | Antipoptosis induced by DM | Protect epididymal cells and counteract their apoptosis in diabetic condition | [36] |
|
| |||||||
| 13 | Liver tissue of rats | Ganoderma lucidum peptide | 27.1 μg/mL | Malondialdehyde level | Antioxidant | Substantial antioxidant activity in the rat liver tissue homogenates and mitochondrial membrane peroxidation systems | [87] |
|
| |||||||
| 14 | Lupus mice | Ganoderma tsugae | 0.5 mg/kg/day | Decreased proteinuria, decreased serum levels of antidsDNA autoantibody | Prevention of autoantibody | Prevention of autoantibody formation | [88] |
Table 5.
In vitro studies of G. lucidum.
| No. | Form | Conc. | Chemical antioxidant tests | Biological text of in vitro | Exp. parameters | Biological activity | Pathway | References |
|---|---|---|---|---|---|---|---|---|
| 1 | GLP | 0.5-3.0 mg/mL | RS FR |
= | Scavenging of free radicals and reducing power | Antioxidant | NM | [89] |
| 2 | G. lucidum and Egyptian Chlorella vulgaris | CVE (63.5 μg/mL) was mixed with GLE (4.1 μg/mL) | RS FR AP Other tests |
Lipopolysaccharide-stimulated white blood cells | Nitric oxide, tumor necrosis factor- (TNF-) α | Antioxidant and anti-inflammatory | Downregulate NF-κB | [39] |
| 3 | Polysaccharides in G. lucidum | 2 mg/mL | RS FR AP Other tests |
NM | Radical scavenging reducing power | Antioxidant | NM | [90] |
| 4 | G. lucidum extract | 50 mg | RS FR AP Other tests |
NM | Reducing power | Antimicrobial and antioxidant | NM | [40] |
| 5 | Ganoderma lucidum G2 | 0.32 mg | RS FR AP Other tests |
DNA protection | Radical scavenging reducing power | Antimicrobial and antioxidant | NM | [41] |
| 6 | Protein extracts | 2–13 μg protein/mL | AP Other tests |
DNA protection | Radical scavenging reducing power | Antioxidant, antibacterial | NM | [42] |
| 7 | Polysaccharides extraction | = | FR AP Other tests |
MCF-7 breast cancer cell line and HeLa cells | Radical scavenging | Antioxidant Anticancer |
NM | [43] |
| 8 | G. lucidum and G. resinaceum | 0.1–1 & 0.64 ± 0.04 0–2.25 mg/mL |
FR AP Other tests |
In vitro cell line | Radical-scavenging chelating lipoxygenase assay | Antiproliferative & antioxidant | NM | [44] |
| 9 | Diff, organic solvent o G. lucidum | 1-200 μg/mL | FR AP Other tests |
NM | Radical scavenging, chelating lipid peroxidation | Antioxidant Anticholinesterase |
NM | [45] |
| 10 | Both aqueous and methanolic extracts | 0.2–30 mg/mL of extraction | FR AP Other tests |
NM | Radical scavenging, chelating lipid peroxidation | Antioxidant | NM | [46] |
| 11 | Low-molecular-weight β-1,3-glucan | 0–200 μg/mL | AP Other tests |
Mouse monocyte-macrophage cell line, RAW 264.7 | H2O2-induced apoptosis | Antioxidant | Attenuating intracellular reactive oxygen species (ROS) and inhibiting sphingomyelinase (SMase) activity | [51] |
| 12 | Polysaccharides | 0.16-10 mg/mL | FR AP Other tests |
NM | Radical scavenging, chelating reducing power | Antioxidant | NM | [47] |
| 13 | G. lucidumwater-soluble and water-insoluble | 80-1100 μg/ml | FR AP Other tests |
Human uroepithelial cell (HUC-PC) cells | Radical scavenging, chelating reducing power | Antioxidant | Oxidative DNA damage. Lingzhi-induced apoptosis in bladder chemoprevention | [48] |
| 14 | Ganoderma lucidum polysaccharides | 0.1-0.6 mg/ml | RS | CCl-induced injury hepatocytes DNA protection | MDA, SOD, CYP3A, caspase-3, andcaspase-8 | Suppressing inflammatory responses | Reduction of NF-κB activation inhibition of caspase-3, caspase-6, and caspase-9, indicating and suppression extrinsic-induced apoptosis | [52] |
| 15 | Ganoderic acid A | 10-80 lM/mL | NM | Pancreatic cells | Radical scavenging Antiproliferative |
Antioxidant Anticancer |
β-Catenin in Wnt signaling pathway | [54] |
| 16 | Aqueous extract of G. lucidum | 5-20 μL | NM | DNA protection | Radical scavenging | Antioxidant DNA repair |
Enhancing reactivity of apurinic/apyrimidinic endonucleases (APE1) a major enzyme of base excision repair (BER) | [91] |
| 17 | Methanolic extract of G. lucidum | 65 & 130 μg/mL | NM | Human gastric tumor cells | Increased the formation of autophagosomes | Induces autophagy | Increasing of the cellular levels of LC3-II and decreasing p62 (autophagy-related protein) | [92] |
| 18 | G. lucidum (GLPS) and G. sinense (GSPS) | 19–300 μg/mL | NM | RAW 264.7 mouse macrophage cells | Nitric oxide secretion of cytokines | Immunomodulatory | Promoting macrophage phagocytosis, increasing their release of nitric oxide and cytokines interleukin- (IL-) 1a, IL-6, IL-10, and tumor necrosis factor-α | [56] |
| 19 | Proteopolysaccharide from G. lucidum | 2 - 10 μg/mL | NM | RAW264.7, a mouse macrophage cell line | Nitrite production Expression levels of cytokines |
Activation the immune system by modulating cytokine production. | NM | [57] |
NM = not mention; RS = radical scavenging; FR = ferric reducing; AP = antilipid peroxidation.
3.2. In Vivo Antioxidant Activity and Protective Effect of G. lucidum
According to 10 in vivo experimental studies, G. lucidum has antioxidant activities and protects against OS through four main factors in different tissues, such as the heart, liver, thymus, spleen, eyes, and skeletal muscles, and by regulating chemical-level OS parameters in blood circulation (Table 4). G. lucidum exhibits its antioxidant effects by increasing the antioxidant enzymes and inhibiting the enzymes involved in OS. G. lucidum also increases the activities of superoxide dismutase (SOD), glutathione-S-transferase (GST), glutathione peroxidase (GPx), catalase (CAT), mitochondrial succinate dehydrogenase (SDH), and Mn-SOD and reduces glutathione (GSH) levels. By contrast, G. lucidum decreases the activities of nitric oxide synthase (NOS), cytochrome P450 2E1 (CYP2E1), xanthine oxidase (XOD), and myeloperoxidase (MPO). G. lucidum also significantly decreases lipid peroxidation levels, advanced oxidation protein products (AOPPs), and malondialdehyde (MDA) levels.
The first factor is the four toxic substances, including CCl4-induced oxidative stress (OS) in the liver, croton oil produced OS in the skin through inflammation, N-methyl-N-nitrosourea (MNU) causing retinal photoreceptor cell lesions in the eyes, and 5-fluorouracil-induced OS in the thymus and spleen of mice. Oral administration of G. lucidum polysaccharides (GLPs) represses free radical lipid peroxidation induced by CCl4 to reduce the enzyme activities of NOS and CYP2E1. Significant inhibition of NOS and CYP2E1 activities and MDA and IL-1β levels was noted in liver tissues, and depleted levels of interleukin- (IL-) 1β, IL-18, IL-6, and tumor necrosis factor-α were found in serum. In CCl4-induced liver damage, highly reactive trichloromethyl free radicals are generated by the cytochrome P450 isozymes (P450s) of the endoplasmic reticulum [22]. Topical administration of G. lucidum ethanol extract inhibits the croton oil-induced lipid peroxidation in the skin of mice [23]. Ganoderma spore lipid (GSL) shows a protective effect on MNU-induced retina injury by inhibiting the related apoptosis to modulate the expression levels of Bax, Bcl-xl, and caspase-3 [30]. GLPs also exhibit an antioxidant effect in 5-fluorouracil-induced OS and improve SOD, an intracellular compound that protects against oxidative processes initiated by superoxide anion and GPx contents in the spleen and thymus of mice [31].
The second factor creates conditions in biological systems that can induce OS, such as exercise-like exhaustive swimming, which is OS induced in skeletal muscles, and a carotid artery ligation, which disturbs the flow-induced OS level of manganese-dependent superoxide dismutase (Mn-SOD) in blood vessels. GLPs show protective effects against comprehensive swimming-induced OS by improving the activities of antioxidant enzymes (SOD, GPx, and CAT) and decreasing the MDA levels in the skeletal muscle of mice [32]. Oral ganoderma triterpenoids (GTs) protect against disturbed flow-induced OS through carotid artery ligation, which leads to chronic OS and inflammation that are features of early atherogenesis in mice, and by preventing neointimal thickening 2 weeks after ligation. Early atherogenesis includes neointimal hyperplasia and endothelial dysfunction due to flow turbulence in the ligated artery as induced by OS. GTs alleviate OS and restore the atheroresistent status of endothelium by inhibiting endothelin-1 induction, von Willebrand factor, and monocyte chemoattractant protein-1 after 3-day ligation as atherogenic factors [33]. Inflammatory cytokines, OS-induced endothelial dysfunction, and chronic OS contribute to endothelial impairment and induces atherogenesis.
The third factor in OS includes diseases such as type II diabetes mellitus (DM) and cancer. In type II DM, the beneficial effects of G. lucidum on abnormal heart and testis and epididymal cells of rats with streptozotocin-induced type II DM were evaluated. GLPs improve the myocardial ultrastructure by reducing MDA, activating antioxidant enzymes (GSH-Px, CAT, SOD, and NO) in cardiac tissues, and reducing lipid peroxidation in type II DM rats [34]. G. lucidum spores protect the testis of rats with type II DM by substantially increasing the mitochondrial SDH and decreasing the activities of XOD and MPO [35]. G. lucidum spores protect epididymal cells and counteract their apoptosis that damages the mitochondria and disequilibrium of calcium homeostasis by reducing the amount of mitochondrial cytoplasm cytochrome C in type II DM rats [36]. GLP administration enhances the immunity and antioxidant activities in N-methyl-N9-nitro-nitrosoguanidine-induced gastric cancer in Wistar rats. GLP remarkably reduces the levels of serum IL-6 and TNF-α and increases the levels of serum IL-2, IL-4, and IL-10. In addition, GLP improves the levels of SOD, CAT, and GSH-Px in serum and gastric tissues [37].
The fourth factor involved in OS is aging. G. lucidum administration ameliorates the age-related decline of antioxidant status in aged mice, substantially elevates the activities of GST, Mn-SOD, GPx, and CAT, and reduces GSH. By contrast, lipid peroxidation, AOPP, and reactive oxygen species (ROS) are reduced [38] (Table 4).
3.3. In Vitro Antioxidant of G. lucidum and Its Possible Pathway
Chemical antioxidant tests consistently revealed the free radical scavenging activity of G. lucidum. Twelve studies reported the scavenging activity of G. lucidum for different free radicals including 2,2-diphenylpicrylhydrazyl radical (DPPH·), 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid) radical (ABTS·+), hydroxyl radical (HO·), and hydrogen peroxide radicals (H2O2) [39–49] (Table 5). G. lucidum also inhibits lipid peroxidation [23, 49]. In some studies, G. lucidum protects against DNA damage [41, 42, 50]. The results of chemical antioxidant tests regarding the antioxidant properties of G. lucidum are also in agreement with the cell-based antioxidant assays. G. lucidum shows free radical scavenging activity for H2O2 in RAW264.7 cells incubated with G. lucidum lipopolysaccharide and protects against H2O2-induced cell death [48]. G. lucidum also hinders sphingomyelinase activity in incubated RAW264.7 cells with lipopolysaccharide [51]. In addition, G. lucidum prevents lipid peroxidation in two cell models, namely, WBCs incubated with lipopolysaccharide to induce OS [39] and hepatocytes incubated with CCl4 to induce OS [52]. In both cell models, G. lucidum showed protection by elevating the antioxidant enzyme activity (SOD, GPx, and GR) and improving the GSH level. Moreover, G. lucidum protects macrophages in human monocytic cells incubated with lipopolysaccharide to stimulate NO production [53].
Wnt, Erk1/2, and NF-κB are the possible signaling pathways of G. lucidum that support its antioxidant and protective effects. A pancreatic cell study suggested β-catenin in the Wnt signaling pathway as a target of ganoderic acid A, thus leading to cell protection and effective scavenging of ROS [54]. The Wnt signaling pathways transfer the signals from extracellular to intercellular and are stimulated by the Wnt protein binding to the cytoplasmic family receptor, which occurs in downstream cell signaling and controls the transcription of genes. In the canonical Wnt pathway, β-catenin accumulates in the cytoplasm and is further translocated into the nucleus, and this phenomenon is widely recognized as a regulation marker of fat and glucose metabolism and β-catenin/Wnt signaling involved in insulin secretion [54]. In 2006, Thyagarajan and his colleagues mentioned that G. lucidum modulates Erk1/2 signaling and transcription factors AP-1 and NF-κB and downregulates c-Fos, whose expression can be induced by OS as the result of the inhibited OS-induced invasive behavior of breast cancer cells. A high H2O2 concentration (5 mM) can stimulate Erk1/2 signaling in MCF-7 cells [55].
In addition to its antioxidant activities, G. lucidum also exhibits an anti-inflammatory property and modulates the immune system. It can reverse LPS-induced inflammation by downregulating inflammatory mediators such as NF-κB, thus substantially inhibiting NOS and reducing NO level [39]. G. lucidum also modulates the immune system byregulating cytokine production in RAW264.7 macrophages [56, 57]. Moreover, it increases the formation of autophagosomes and controls proteins (Vps34, beclin 1, LC3-I, LC3-II, and p62) that induce autophagy in a gastric adenocarcinoma cell line. G. lucidum increases the cellular levels of LC3-II and decreases the cellular levels of p62 (Table 5).
4. Discussion
Among the 155 targets associated with DCM, mTOR, CPT1B, and CPT2 have the highest association. mTOR acts as a core regulator of cellular metabolism, growth, and survival in response to hormone growth factors, nutrients, energy, and stress signals. An animal study confirmed that streptozotocin-induced diabetes increases mTOR levels in rats [58]. mTOR can be found in different cellular locations including membrane, cytoplasm, and nucleus and different cellular organs (mitochondria, Golgi, and endoplasmic reticulum) and therefore is involved directly or indirectly in regulating the phosphorylation of at least 800 proteins (OPT.V19.11). mTOR functions through two distinct signaling complexes of mTORC1 and mTORC2 [59]. When activated, mTORC1 upregulates protein synthesis by phosphorylating the key regulators of mRNA translation and ribosome synthesis. mTORC1 also regulates protein synthesis [29], lipid synthesis [60], and mitochondrial biogenesis and stimulates the pyrimidine biosynthesis pathway through acute and delayed regulations. In acute regulation, mTORC1 stimulates pyrimidine biosynthesis through the ribosomal protein S6 kinase B1-mediated phosphorylation of biosynthetic enzyme carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase; these enzymes catalyze the first three steps in de novo pyrimidine synthesis [61]. In delayed regulation, mTORC1 stimulates pyrimidine biosynthesis through the transcriptional enhancement of the pentose phosphate pathway, which produces 5-phosphoribosyl-1-pyrophosphate, an allosteric activator of pyrimidine biosynthesis enzyme at a later step in the synthesis. In addition, mTORC1 regulates ribosome synthesis by activating RNA polymerase III-dependent transcription through the phosphorylation and inhibition of MAF1 protein, a RNA polymerase III-repressor. When nutrients are available and mTOR kinase is active, MAF1 is hyperphosphorylated, and RNA polymerase III is engaged in the transcription [62]. Stress-induced MAF1 dephosphorylation resulted in nuclear localization, increased targeting of gene-bound RNA polymerase III, and decreased transcriptional readout [63, 64]. Moreover, mTORC1 is involved in the negative feedback regulation of autophagy on upstream growth factor signaling during microtubule regulation [64–66].
mTORC2 regulates other cellular processes such as survival and organization of cytoskeleton, actin cytoskeleton [67], osteoclastogenesis, and circadian clock function. In a pressure-overloaded male mouse heart, mTORC2 maintains a contractile function [68]. In brown adipose tissues, mTOR complex 2 has a role in β3-adrenoceptor-stimulated glucose uptake by stimulating the translocation of newly synthesized GLUT1 to the plasma membrane, thereby increasing the glucose uptake [69]. mTOR complex 2 regulates the proper turnover of insulin receptor substrate-1 [70].
G. lucidum exhibits cardiac protection via its antioxidant properties through OS modulation. This systemic review of 33 studies has documented its antioxidant activities. At the molecular and cellular levels, OS is a key in diabetes-induced DCM [18]. The antioxidant effects of G. lucidum are facilitated by increasing the antioxidant enzymes and inhibiting the enzymes involved in OS [33–35, 38]. G. lucidum consistently shows free radical scavenging activity against several free radicals including DPPH·, ABTS·+, HO·, and H2O2. As confirmed by the in vitro (chemical and cell-based) antioxidant tests, G. lucidum inhibits lipid peroxidation and protects against DNA damage.
G. lucidum modulates several signal pathways including Erk1/2, NF-κB, and Wnt. Its antioxidant activity protects against inflammation and directly modulates immunity through scavenging radicals and through the oxidative signal pathways, thereby protecting the cells. These effects of G. lucidum may contribute to its positive influence on DCM.
DM is a state of persistent inflammation that upregulates mTOR at different levels of the myocardium, thereby influencing several signal pathways. The elevation of cellular cAMP levels disrupts phosphodiesterase-Rheb interaction, increases Rheb-mTOR interaction, and consequently leads to mTOR1 activation. Phosphodiesterase binds with Rheb and thereby inhibits the latter's ability to activate mTOR [71]. Heart myocardium responds to high blood glucose by adapting its energy metabolism and using only fatty acids as a substrate, thus increasing OS through the upregulation of NADPH-oxidases, NO synthases [72], and reversible oxidative modifications for myocardial titin elastic protein [73]. mTOR upregulation and oxidative modification alter titin-based stiffness and titin isoform composition, thereby impairing myocardium contractility. The PI3K-Akt-mTOR kinase axis regulates the composition of titin isoform [73]. OS decreases NO levels, leading to the impairment of the NO-soluble guanylate cyclase- (sGC-) cyclic guanosine monophosphate- (cGMP-) protein kinase G (PKG) pathway, an important regulator of cardiac contractility [72]. Chronic intrude accumulation to high free fatty acids downregulates PPAR-α and impairs mTOR-PPAR-α, thereby causing mitochondrial dysfunction in rodent cardiomyocytes and further deteriorating cardiac function through the inhibition of fatty acid oxidation and increase in intracellular fat accumulation. PPAR-α is involved in the upregulation of carnitine palmitoyltransferase I, which increases the uptake of long-chain fatty acid in the mitochondria and facilitates the beta-oxidation of fatty acids. mTOR-PPAR-α axis modification can lead to inflammation [74] and immune dysfunction [75]. mTOR upregulation leads to the impaired response to adrenergic stimulation in DCM mice and further reduces heart contractility [58]. mTOR inhibition improves contractility via the chronic administration of PDE inhibitor in animals and patients with diabetes [76] and restores the impaired response to adrenergic stimulation in DCM mice [58]. G. lucidum shows its effects via several signal pathways such as Wnt, Erk1/2, and NF-κB pathway and consequently reduces the upregulated mTOR and its effects. mTOR is the main target of G. lucidum, and this finding supports its antioxidant and cardioprotective effects. G. lucidum inhibits the Wnt pathway [54] and may decrease the activity of mTOR via the Wnt/GSk/mTOR signal pathway. A pathologically stressed heart reactivates the Wnt signal pathway, which is modulated during left ventricular remodeling [77]. In heart cells, the Wnt pathway plays a role in the release of intracellular Ca2+ whose accumulation activates several Ca+2-sensitive proteins, fat and glucose metabolism, and cell fate decisions, such as renewal, differentiation, and apoptosis. Wnt dysregulation has an important role in cardiac diseases such as hypertrophy and fibrosis [78]. The Wnt pathway is important in the response to heart injuries leading to adverse effects on the heart [79] and is integrated with bioenergetic status to control mTOR activity [80]. Wnt is activated in late-stage inflammation of heart tissue [81]. G. lucidum suppresses Erk1/2 signaling [55] and consequently reduces the mTOR level. Erk1/2 signaling inhibits the TSC1/2 complex, which is the downregulator of mTOR, and thus activates mTOR [82]. The antioxidant properties of G. lucidum abolish the activation of the Erk pathway by OS. NADPH oxidase 2 is involved in Erk activation [83], and the inhibition of Erk/mTOR by G. lucidum also prevents NF-κB. mTOR activates NF-κB by phosphorylating the NF-κB p65 subunit, increasing p65 nuclear translocation, and activating gene transcription. With its anti-inflammatory effect, G. lucidum inhibits NF-κB via decreasing inflammatory mediators and cytokines such as TNF or IL-1, and innate immune response effectors activate NF-κB via the IKK complex through IκB protein phosphorylation with subsequent ubiquitination and degradation [84]. Inhibiting mTOR and NF-κB may improve the contractility of the heart, abolish the angiotensin II-induced hypertrophic response of cardiomyocytes [83], and prevent heart failure. A prolonged NF-κB activation promotes heart failure by evoking signals that induce chronic inflammation through the enhancement of cytokines including tumor necrosis factor, IL-1, and IL-6, commencing to endoplasmic reticulum stress responses and cell death [85].
Our results concluded that the antioxidant properties of G. lucidum and the cardioprotection of its polysaccharides may have a direct effect. Its free radical scavenging ability reduces OS and upregulates mTOR via several pathways including Wnt, Erk1/2, and NF-κB/IKK/TOR, thereby improving myocardium contractility (Figure 4). The anti-inflammatory properties may enhance the cAMP/cGMP/mTOR/PPAR pathway and its related protein or/and pathway and mitochondrial function, thus improving myocardium hemostasis. Further study is needed to identify the specific target of GLP in heart tissues.
Figure 4.
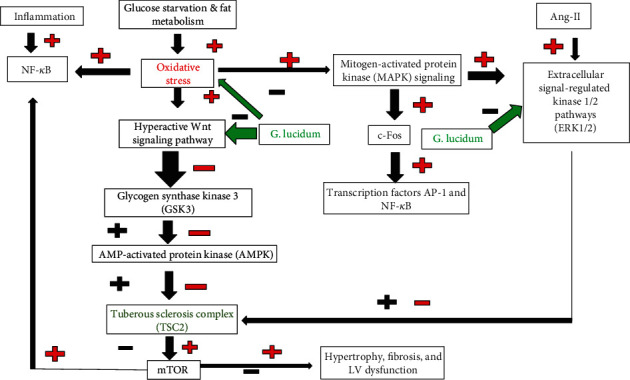
G. lucidum inhibits mTOR via several signal pathways (- inhibit; + stimulate). Red +/- effects of inflammation, glucose starvation and fat metabolism, and oxidative stress on different effectors of different pathways in cardiocytes; black +/- protective effects of G lucidum at different effectors of different pathways in cardiocytes.
Acknowledgments
This research was funded by the Jiamusi University, Basic Medicine College team, under grant number JDXKTD-2019002.
Abbreviations
- ABTS·+:
2,2′-azino-bis (3-ethylbenzthiazoline-6-sulphonic acid) radical
- AOPP:
Advanced oxidation protein products
- Bax:
BCL2 associated X, apoptosis regulator
- Bcl-xl:
B-cell lymphoma-extra large
- BECN1:
Beclin 1
- cAMP:
Cyclic adenosine monophosphate
- CAT:
Catalase
- CCl4:
Carbon tetrachloride
- c-Fos:
A protooncogene
- cGMP:
Cyclic guanosine monophosphate
- CPT1B and CPT2:
Carnitine palmitoyltransferase 1B and 2
- CYP2E1:
Cytochrome P450 2E1
- DCM:
Diabetic cardiomyopathy
- DM:
Diabetic mellitus
- DNA:
Deoxyribonucleic acid
- DPPH·:
2,2-diphenylpicrylhydrazyl radical
- EMBL:
European Molecular Biology Laboratory
- Erk1/2:
Extracellular signal-regulated kinase
- GLPs:
Ganoderma lucidum polysaccharides
- GPx:
Glutathione peroxidase
- GR:
Glutathione reductase
- GSH:
Reduced glutathione
- GSH-Px:
Glutathione peroxidase
- GST:
Glutathione-S-transferase
- GTs:
Ganoderma triterpenoids
- H2O2:
Hydrogen peroxide radicals
- HO·:
Hydroxyl radical
- IL-6:
Interleukin 6
- LC3:
Light chain 3
- LPS:
Lipopolysaccharide
- MAF1:
Protein negative regulator of RNA polymerase III
- MCF-7 cells:
Breast cancer cell line
- MDA:
Malondialdehyde level
- Mn-SOD:
Manganese-superoxide dismutase
- MNU:
N-methyl-N-nitrosourea
- MPO:
Myeloperoxidase
- mTOR:
Mammalian target of rapamycin
- mTORC:
mTOR complex
- NF-κB:
Nuclear factor-κB
- NO:
Nitrous oxide
- NOS:
Nitric oxide synthase
- OS:
Oxidative stress
- OTP:
Open Targets Platform
- PDE:
Phosphodiesterase
- PKG:
Protein kinase G
- PML:
Promyelocytic leukemia.
Conflicts of Interest
The authors declare no conflict of interest.
Authors' Contributions
Hongbin Qiu and Shuqiu Wang performed the conceptualization; Fahmi Shaher and Mahfoudh A.M. Abdulghani did the methodology; Hisham AL-ward, Salem Baldi, and Yu Hu participated in the software; Shaobo Zhou, Mahfoudh A.M. Abdulghani, and Weiqun Wang contributed to the validation; Salem Baldi, Fahmi Shaher, and Mahfoudh A.M. Abdulghani performed the formal analysis; Yu Zhang and Yao Wei participated in the investigation; Shuqiu Wang contributed to acquiring resources; Fahmi Shaher helped in the data curation; Fahmi Shaher and Mahfoudh A.M. Abdulghani wrote and prepared the original draft; Shaobo Zhou wrote, reviewed, and edited the manuscript; Mahfoudh A.M. Abdulghani and Fahmi Shaher performed the visualization; Hongbin Qiu supervised the study; Shuqiu Wang did the project administration; Shuqiu Wang helped in funding acquisition. Authorship must be limited to those who have contributed substantially to the work reported.
Supplementary Materials
Supplementary Table 1: association sore of 309 targets associated with diabetic cardiomyopathy in 30 recorded tissues. Supplementary Table 2: thirty-two tissue organs expressed 309 targets and types of pathways.
References
- 1.Raghavan S., Vassy J. L., Ho Y. L., et al. Diabetes mellitus-related all-cause and cardiovascular mortality in a national cohort of adults. Journal of the American Heart Association. 2019;8(4, article e011295) doi: 10.1161/JAHA.118.011295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Glovaci D., Fan W., Wong N. D. Epidemiology of diabetes mellitus and cardiovascular disease. Current Cardiology Reports. 2019;21(4):p. 21. doi: 10.1007/s11886-019-1107-y. [DOI] [PubMed] [Google Scholar]
- 3.Bertoluci M. C., Rocha V. Z. Cardiovascular risk assessment in patients with diabetes. Diabetology & Metabolic Syndrome. 2017;9(1):p. 25. doi: 10.1186/s13098-017-0225-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Williams L. J., Nye B. G., Wende A. R. Diabetes-related cardiac dysfunction. Endocrinology and Metabolism. 2017;32(2):171–179. doi: 10.3803/EnM.2017.32.2.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Karuranga S., da Rocha Fernandes J., Huang Y., Malanda B. IDF Diabetes Atlas. 8th. Brussels: IDF; 2017. [DOI] [PubMed] [Google Scholar]
- 6.Battiprolu P. K., Gillette T. G., Wang Z. V., Lavandero S., Hill J. A. Diabetic cardiomyopathy: mechanisms and therapeutic targets. Drug Discovery Today: Disease Mechanisms. 2010;7(2):e135–e143. doi: 10.1016/j.ddmec.2010.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Booth G. L., Kapral M. K., Fung K., Tu J. V. Relation between age and cardiovascular disease in men and women with diabetes compared with non-diabetic people: a population-based retrospective cohort study. The Lancet. 2006;368(9529):29–36. doi: 10.1016/S0140-6736(06)68967-8. [DOI] [PubMed] [Google Scholar]
- 8.Wannamethee S. G., Shaper A. G., Whincup P. H., Lennon L., Sattar N. Impact of diabetes on cardiovascular disease risk and all-cause mortality in older men: influence of age at onset, diabetes duration, and established and novel risk factors. Archives of Internal Medicine. 2011;171(5):404–410. doi: 10.1001/archinternmed.2011.2. [DOI] [PubMed] [Google Scholar]
- 9.Boudina S., Abel E. D. Diabetic cardiomyopathy revisited. Circulation. 2007;115(25):3213–3223. doi: 10.1161/CIRCULATIONAHA.106.679597. [DOI] [PubMed] [Google Scholar]
- 10.Mandavia C. H., Aroor A. R., DeMarco V. G., Sowers J. R. Molecular and metabolic mechanisms of cardiac dysfunction in diabetes. Life Sciences. 2013;92(11):601–608. doi: 10.1016/j.lfs.2012.10.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Poirier P., Bogaty P., Garneau C., Marois L., Dumesnil J. G. Diastolic dysfunction in normotensive men with well-controlled type 2 diabetes: importance of maneuvers in echocardiographic screening for preclinical diabetic cardiomyopathy. Diabetes Care. 2001;24(1):5–10. doi: 10.2337/diacare.24.1.5. [DOI] [PubMed] [Google Scholar]
- 12.Ernande L., Bergerot C., Rietzschel E. R., et al. Diastolic dysfunction in patients with type 2 diabetes mellitus: is it really the first marker of diabetic cardiomyopathy? Journal of the American Society of Echocardiography. 2011;24(11):1268–1275.e1. doi: 10.1016/j.echo.2011.07.017. [DOI] [PubMed] [Google Scholar]
- 13.Xi S., Zhou G., Zhang X., Zhang W., Cai L., Zhao C. Protective effect of total aralosides of Aralia elata (Miq) Seem (TASAES) against diabetic cardiomyopathy in rats during the early stage, and possible mechanisms. Experimental & Molecular Medicine. 2009;41(8):538–547. doi: 10.3858/emm.2009.41.8.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sun X., Chen R. C., Yang Z. H., et al. Taxifolin prevents diabetic cardiomyopathy in vivo and in vitro by inhibition of oxidative stress and cell apoptosis. Food and Chemical Toxicology. 2014;63:221–232. doi: 10.1016/j.fct.2013.11.013. [DOI] [PubMed] [Google Scholar]
- 15.Zhou G., Li C., Cai L. Advanced glycation end-products induce connective tissue growth factor-mediated renal fibrosis predominantly through transforming growth factor β-independent pathway. The American Journal of Pathology. 2004;165(6):2033–2043. doi: 10.1016/S0002-9440(10)63254-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pereira L., Matthes J., Schuster I., et al. Mechanisms of [Ca2+] i transient decrease in cardiomyopathy of db/db type 2 diabetic mice. Diabetes. 2006;55(3):608–615. doi: 10.2337/diabetes.55.03.06.db05-1284. [DOI] [PubMed] [Google Scholar]
- 17.Sulaiman M., Matta M. J., Sunderesan N. R., Gupta M. P., Periasamy M., Gupta M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. American Journal of Physiology-Heart and Circulatory Physiology. 2010;298(3):H833–H843. doi: 10.1152/ajpheart.00418.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schaffer S. W., Jong C. J., Mozaffari M. Role of oxidative stress in diabetes-mediated vascular dysfunction: unifying hypothesis of diabetes revisited. Vascular Pharmacology. 2012;57(5-6):139–149. doi: 10.1016/j.vph.2012.03.005. [DOI] [PubMed] [Google Scholar]
- 19.Wachtel-Galor S., Yuen J., Buswell J. A., Benzie I. F. Herbal Medicine: Biomolecular and Clinical Aspects. 2nd. CRC Press/Taylor & Francis; 2011. Ganoderma lucidum (Lingzhi or Reishi) [Google Scholar]
- 20.Kwon O., Lee C.-S., Park Y.-J. SNP and SCAR markers for specific discrimination of antler-shaped Ganoderma lucidum. Microorganisms. 2019;7(1):p. 12. doi: 10.3390/microorganisms7010012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wang X.-C., Xi R.-J., Li Y., Wang D.-M., Yao Y.-J. The species identity of the widely cultivated Ganoderma,‘G. lucidum’(Ling-zhi), in China. PLoS One. 2012;7(7) doi: 10.1371/journal.pone.0040857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen Y.-S., Chen Q. Z., Wang Z. J., Hua C. Anti-inflammatory and hepatoprotective effects of Ganoderma lucidum polysaccharides against carbon tetrachloride-induced liver injury in Kunming mice. Pharmacology. 2019;103(3-4):143–150. doi: 10.1159/000493896. [DOI] [PubMed] [Google Scholar]
- 23.Lakshmi B., Ajith T. A., Sheena N., Gunapalan N., Janardhanan K. K. Antiperoxidative, anti-inflammatory, and antimutagenic activities of ethanol extract of the mycelium of Ganoderma lucidum occurring in South India. Teratogenesis, Carcinogenesis, and Mutagenesis. 2003;23(S1):85–97. doi: 10.1002/tcm.10065. [DOI] [PubMed] [Google Scholar]
- 24.Koscielny G., An P., Carvalho-Silva D., et al. Open Targets: a platform for therapeutic target identification and validation. Nucleic Acids Research. 2017;45(D1):D985–D994. doi: 10.1093/nar/gkw1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Carvalho-Silva D., Pierleoni A., Pignatelli M., et al. Open Targets Platform: new developments and updates two years on. Nucleic Acids Research. 2019;47(D1):D1056–D1065. doi: 10.1093/nar/gky1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liberati A. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. Annals of internal medicine. 2009;151(4):p. W. doi: 10.7326/0003-4819-151-4-200908180-00136. [DOI] [PubMed] [Google Scholar]
- 27.Hsu P. P., Kang S. A., Rameseder J., et al. The mTOR-regulated phosphoproteome reveals a mechanism of mTORC1-mediated inhibition of growth factor signaling. Science. 2011;332(6035):1317–1322. doi: 10.1126/science.1199498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kim D.-H., Sarbassov D. D., Ali S. M., et al. mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery. Cell. 2002;110(2):163–175. doi: 10.1016/S0092-8674(02)00808-5. [DOI] [PubMed] [Google Scholar]
- 29.Brugarolas J., Lei K., Hurley R. L., et al. Regulation of mTOR function in response to hypoxia by REDD1 and the TSC1/TSC2 tumor suppressor complex. Genes & Development. 2004;18(23):2893–2904. doi: 10.1101/gad.1256804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gao Y., Deng X. G., Sun Q. N., Zhong Z. Q. Ganoderma spore lipid inhibits N-methyl-N-nitrosourea-induced retinal photoreceptor apoptosis in vivo. Experimental Eye Research. 2010;90(3):397–404. doi: 10.1016/j.exer.2009.11.017. [DOI] [PubMed] [Google Scholar]
- 31.Wang J., Wang Y., Liu X., Yuan Y., Yue T. Free radical scavenging and immunomodulatory activities of Ganoderma lucidum polysaccharides derivatives. Carbohydrate Polymers. 2013;91(1):33–38. doi: 10.1016/j.carbpol.2012.08.017. [DOI] [PubMed] [Google Scholar]
- 32.Zhonghui Z., Xiaowei Z., Fang F. Ganoderma lucidum polysaccharides supplementation attenuates exercise-induced oxidative stress in skeletal muscle of mice. Saudi journal of biological sciences. 2014;21(2):119–123. doi: 10.1016/j.sjbs.2013.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hsu P.-L., Lin Y. C., Ni H., Mo F. E. Ganoderma triterpenoids exert antiatherogenic effects in mice by alleviating disturbed flow-induced oxidative stress and inflammation. Oxidative Medicine and Cellular Longevity. 2018;2018:11. doi: 10.1155/2018/3491703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Xue H., Qiao J., Meng G., et al. Effect of Ganoderma lucidum polysaccharides on hemodynamic and antioxidation in T2DM rats. China journal of Chinese materia medica. 2010;35(3):339–343. doi: 10.4268/cjcmm20100318. [DOI] [PubMed] [Google Scholar]
- 35.Wang S., Qin W. B., Kang Y. M., et al. Intervention effect of ganoderma lucidum spores on the changes of XOD, MPO and SDH in the testis tissue of NIDDM rats. National Journal of Andrology. 2008;14(9):792–795. [PubMed] [Google Scholar]
- 36.Ma X., Zhou C. F., Wang S. Q., et al. Effects of ganoderma lucidum spores on mitochondrial calcium ion and cytochrome C in epididymal cells of type 2 diabetes rats. National Journal of Andrology. 2007;13(5):400–402. [PubMed] [Google Scholar]
- 37.Pan K., Jiang Q., Liu G., Miao X., Zhong D. Optimization extraction of Ganoderma lucidum polysaccharides and its immunity and antioxidant activities. International Journal of Biological Macromolecules. 2013;55:301–306. doi: 10.1016/j.ijbiomac.2013.01.022. [DOI] [PubMed] [Google Scholar]
- 38.Sudheesh N. P., Ajith T. A., Ramnath V., Janardhanan K. K. Therapeutic potential of _Ganoderma lucidum_ (Fr.) P. Karst. against the declined antioxidant status in the mitochondria of post-mitotic tissues of aged mice. Clinical Nutrition. 2010;29(3):406–412. doi: 10.1016/j.clnu.2009.12.003. [DOI] [PubMed] [Google Scholar]
- 39.Abu-Serie M. M., Habashy N. H., Attia W. E. In vitro evaluation of the synergistic antioxidant and anti-inflammatory activities of the combined extracts from Malaysian Ganoderma lucidum and Egyptian Chlorella vulgaris. BMC Complementary and Alternative Medicine. 2018;18(1):p. 154. doi: 10.1186/s12906-018-2218-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sharif S., Shahid M., Mushtaq M., Akram S., Rashid A. Wild mushrooms: a potential source of nutritional and antioxidant attributes with acceptable toxicity. Preventive nutrition and food science. 2017;22(2):124–130. doi: 10.3746/pnf.2017.22.2.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sarnthima R., Khammaung S., Sa-ard P. Culture broth of Ganoderma lucidum exhibited antioxidant, antibacterial and α-amylase inhibitory activities. Journal of Food Science and Technology. 2017;54(11):3724–3730. doi: 10.1007/s13197-017-2839-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sa-ard P., Sarnthima R., Khammuang S., Kanchanarach W. Antioxidant, antibacterial and DNA protective activities of protein extracts from Ganoderma lucidum. Journal of Food Science and Technology. 2015;52(5):2966–2973. doi: 10.1007/s13197-014-1343-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chen P., Yong Y., Gu Y., Wang Z., Zhang S., Lu L. Comparison of antioxidant and antiproliferation activities of polysaccharides from eight species of medicinal mushrooms. International journal of medicinal mushrooms. 2015;17(3):287–295. doi: 10.1615/IntJMedMushrooms.v17.i3.80. [DOI] [PubMed] [Google Scholar]
- 44.Saltarelli R., Ceccaroli P., Buffalini M., et al. Biochemical characterization and antioxidant and antiproliferative activities of different Ganoderma collections. Journal of Molecular Microbiology and Biotechnology. 2015;25(1):16–25. doi: 10.1159/000369212. [DOI] [PubMed] [Google Scholar]
- 45.Tel G., Ozturk M., Duru M. E., Turkoglu A. Antioxidant and anticholinesterase activities of five wild mushroom species with total bioactive contents. Pharmaceutical Biology. 2014;53(6):824–830. doi: 10.3109/13880209.2014.943245. [DOI] [PubMed] [Google Scholar]
- 46.Rani P., Lal M. R., Maheshwari U., Krishnan S. Antioxidant potential of lingzhi or reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes) cultivated on Artocarpus heterophyllus sawdust substrate in India. International Journal of Medicinal Mushrooms. 2015;17(12):1171–1177. doi: 10.1615/IntJMedMushrooms.v17.i12.70. [DOI] [PubMed] [Google Scholar]
- 47.Liu W., Wang H., Pang X., Yao W., Gao X. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. International Journal of Biological Macromolecules. 2010;46(4):451–457. doi: 10.1016/j.ijbiomac.2010.02.006. [DOI] [PubMed] [Google Scholar]
- 48.Yuen J., Gohel M. The dual roles of Ganoderma antioxidants on urothelial cell DNA under carcinogenic attack. Journal of Ethnopharmacology. 2008;118(2):324–330. doi: 10.1016/j.jep.2008.05.003. [DOI] [PubMed] [Google Scholar]
- 49.Mau J.-L., Lin H.-C., Chen C.-C. Antioxidant properties of several medicinal mushrooms. Journal of Agricultural and Food Chemistry. 2002;50(21):6072–6077. doi: 10.1021/jf0201273. [DOI] [PubMed] [Google Scholar]
- 50.Kim K. C., Kim I. Ganoderma lucidum extract protects DNA from strand breakage caused by hydroxyl radical and UV irradiation. International Journal of Molecular Medicine. 1999;4(3):273–277. [PubMed] [Google Scholar]
- 51.Kao P.-F., Wang S.-H., Hung W.-T., Liao Y.-H., Lin C.-M., Yang W.-B. Structural Characterization and Antioxidative Activity of Low-Molecular- Weights Beta-1,3-Glucan from the Residue of Extracted Ganoderma lucidum Fruiting Bodies. Journal of Biomedicine and Biotechnology. 2012;2012:8. doi: 10.1155/2012/673764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Liu Y.-J., du J. L., Cao L. P., et al. Anti-inflammatory and hepatoprotective effects of Ganoderma lucidum polysaccharides on carbon tetrachloride-induced hepatocyte damage in common carp (Cyprinus carpio L.) International Immunopharmacology. 2015;25(1):112–120. doi: 10.1016/j.intimp.2015.01.023. [DOI] [PubMed] [Google Scholar]
- 53.Woo C. W. H., Man R. Y. K., Siow Y. L., et al. Ganoderma lucidum inhibits inducible nitric oxide synthase expression in macrophages. Molecular and Cellular Biochemistry. 2005;275(1-2):165–171. doi: 10.1007/s11010-005-1352-9. [DOI] [PubMed] [Google Scholar]
- 54.Gill B. S., Kumar S. Ganoderic acid A targeting β-catenin in Wnt signaling pathway: in silico and in vitro study. Interdisciplinary Sciences: Computational Life Sciences. 2018;10(2):233–243. doi: 10.1007/s12539-016-0182-7. [DOI] [PubMed] [Google Scholar]
- 55.Thyagarajan A., Jiang J., Hopf A., Adamec J., Sliva D. Inhibition of oxidative stress-induced invasiveness of cancer cells by Ganoderma lucidum is mediated through the suppression of interleukin-8 secretion. International Journal of Molecular Medicine. 2006;18(4):657–664. [PubMed] [Google Scholar]
- 56.Meng L.-Z., Xie J., Lv G. P., et al. A comparative study on immunomodulatory activity of polysaccharides from two official species of Ganoderma (Lingzhi) Nutrition and Cancer. 2014;66(7):1124–1131. doi: 10.1080/01635581.2014.948215. [DOI] [PubMed] [Google Scholar]
- 57.Ji Z., Tang Q., Zhang J., Yang Y., Jia W., Pan Y. Immunomodulation of RAW264. 7 macrophages by GLIS, a proteopolysaccharide from Ganoderma lucidum. Journal of Ethnopharmacology. 2007;112(3):445–450. doi: 10.1016/j.jep.2007.03.035. [DOI] [PubMed] [Google Scholar]
- 58.West T. M., Wang Q., Deng B., et al. Phosphodiesterase 5 associates with β2 adrenergic receptor to modulate cardiac function in type 2 diabetic hearts. Journal of the American Heart Association. 2019;8(15, article e012273) doi: 10.1161/JAHA.119.012273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jacinto E., Loewith R., Schmidt A., et al. Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive. Nature Cell Biology. 2004;6(11):1122–1128. doi: 10.1038/ncb1183. [DOI] [PubMed] [Google Scholar]
- 60.Porstmann T., Santos C. R., Griffiths B., et al. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metabolism. 2008;8(3):224–236. doi: 10.1016/j.cmet.2008.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Robitaille A. M., Christen S., Shimobayashi M., et al. Quantitative phosphoproteomics reveal mTORC1 activates de novo pyrimidine synthesis. Science. 2013;339(6125):1320–1323. doi: 10.1126/science.1228771. [DOI] [PubMed] [Google Scholar]
- 62.Michels A. A., Robitaille A. M., Buczynski-Ruchonnet D., et al. mTORC1 directly phosphorylates and regulates human MAF1. Molecular and Cellular Biology. 2010;30(15):3749–3757. doi: 10.1128/MCB.00319-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Orioli A., Praz V., Lhôte P., Hernandez N. Human MAF1 targets and represses active RNA polymerase III genes by preventing recruitment rather than inducing long-term transcriptional arrest. Genome Research. 2016;26(5):624–635. doi: 10.1101/gr.201400.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Holczer M., Hajdú B., Lőrincz T., Szarka A., Bánhegyi G., Kapuy O. A double negative feedback loop between mTORC1 and AMPK kinases guarantees precise autophagy induction upon cellular stress. International Journal of Molecular Sciences. 2019;20(22):p. 5543. doi: 10.3390/ijms20225543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Koren I., Reem E., Kimchi A. DAP1, a novel substrate of mTOR, negatively regulates autophagy. Current biology. 2010;20(12):1093–1098. doi: 10.1016/j.cub.2010.04.041. [DOI] [PubMed] [Google Scholar]
- 66.Zhao D.-Y., Yu D. D., Ren L., Bi G. R. Ligustilide protects PC12 cells from oxygen-glucose deprivation/reoxygenation-induced apoptosis via the LKB1-AMPK-mTOR signaling pathway. Neural Regeneration Research. 2020;15(3):473–481. doi: 10.4103/1673-5374.266059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sarbassov D. D., Ali S. M., Kim D.-H., et al. Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Current biology. 2004;14(14):1296–1302. doi: 10.1016/j.cub.2004.06.054. [DOI] [PubMed] [Google Scholar]
- 68.Shende P., Xu L., Morandi C., et al. Cardiac mTOR complex 2 preserves ventricular function in pressure-overload hypertrophy. Cardiovascular Research. 2015;109(1):103–114. doi: 10.1093/cvr/cvv252. [DOI] [PubMed] [Google Scholar]
- 69.Olsen J. M., Sato M., Dallner O. S., et al. Glucose uptake in brown fat cells is dependent on mTOR complex 2–promoted GLUT1 translocation. Journal of Cell Biology. 2014;207(3):365–374. doi: 10.1083/jcb.201403080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kim S. J., DeStefano M. A., Oh W. J., et al. mTOR complex 2 regulates proper turnover of insulin receptor substrate-1 via the ubiquitin ligase subunit Fbw8. Molecular Cell. 2012;48(6):875–887. doi: 10.1016/j.molcel.2012.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kim H. W., Ha S. H., Lee M. N., et al. Cyclic AMP controls mTOR through regulation of the dynamic interaction between Rheb and phosphodiesterase 4D. Molecular and Cellular Biology. 2010;30(22):5406–5420. doi: 10.1128/MCB.00217-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Mátyás C., Németh B. T., Oláh A., et al. Prevention of the development of heart failure with preserved ejection fraction by the phosphodiesterase-5A inhibitor vardenafil in rats with type 2 diabetes. European Journal of Heart Failure. 2017;19(3):326–336. doi: 10.1002/ejhf.711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Koser F., Loescher C., Linke W. A. Posttranslational modifications of titin from cardiac muscle: how, where, and what for? The FEBS Journal. 2019;286(12):2240–2260. doi: 10.1111/febs.14854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Sarnelli G., D’Alessandro A., Iuvone T., et al. Palmitoylethanolamide modulates inflammation-associated vascular endothelial growth factor (VEGF) signaling via the Akt/mTOR pathway in a selective peroxisome proliferator-activated receptor alpha (PPAR-α)-dependent manner. PLoS One. 2016;11(5, article e0156198) doi: 10.1371/journal.pone.0156198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Maile R., Stepp W. H., Eitas T., Cairns B. A. The Mammalian Target of Rapamycin (mTOR)/Peroxisome Proliferator-Activated Receptor γ (PPARγ) axis drives immune dysfunction and outcome after burn injury. 2018.
- 76.Das A., Durrant D., Salloum F. N., Xi L., Kukreja R. C. PDE5 inhibitors as therapeutics for heart disease, diabetes and cancer. Pharmacology & Therapeutics. 2015;147:12–21. doi: 10.1016/j.pharmthera.2014.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Bergmann M. W. WNT signaling in adult cardiac hypertrophy and remodeling: lessons learned from cardiac development. Circulation Research. 2010;107(10):1198–1208. doi: 10.1161/CIRCRESAHA.110.223768. [DOI] [PubMed] [Google Scholar]
- 78.Deb A. Cell–cell interaction in the heart via Wnt/β-catenin pathway after cardiac injury. Cardiovascular Research. 2014;102(2):214–223. doi: 10.1093/cvr/cvu054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Haybar H., Khodadi E., Shahrabi S. Wnt/β-catenin in ischemic myocardium: interactions and signaling pathways as a therapeutic target. Heart Failure Reviews. 2019;24(3):411–419. doi: 10.1007/s10741-018-9759-z. [DOI] [PubMed] [Google Scholar]
- 80.Inoki K., Ouyang H., Zhu T., et al. TSC2 integrates Wnt and energy signals via a coordinated phosphorylation by AMPK and GSK3 to regulate cell growth. Cell. 2006;126(5):955–968. doi: 10.1016/j.cell.2006.06.055. [DOI] [PubMed] [Google Scholar]
- 81.Aisagbonhi O., Rai M., Ryzhov S., Atria N., Feoktistov I., Hatzopoulos A. K. Experimental myocardial infarction triggers canonical Wnt signaling and endothelial-to-mesenchymal transition. Disease Models & Mechanisms. 2011;4(4):469–483. doi: 10.1242/dmm.006510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ma L., Chen Z., Erdjument-Bromage H., Tempst P., Pandolfi P. P. Phosphorylation and functional inactivation of TSC2 by Erk. Cell. 2005;121(2):179–193. doi: 10.1016/j.cell.2005.02.031. [DOI] [PubMed] [Google Scholar]
- 83.Li Y., Ha T., Gao X., et al. NF-κB activation is required for the development of cardiac hypertrophy in vivo. American Journal of Physiology-Heart and Circulatory Physiology. 2004;287(4):H1712–H1720. doi: 10.1152/ajpheart.00124.2004. [DOI] [PubMed] [Google Scholar]
- 84.Hayden M., Ghosh S. Signaling to NF-κB. Genes & development. 2004;18(18):2195–2224. doi: 10.1101/gad.1228704. [DOI] [PubMed] [Google Scholar]
- 85.Gordon J. W., Shaw J. A., Kirshenbaum L. A. Multiple facets of NF-κB in the heart: to be or not to NF-κB. Circulation Research. 2011;108(9):1122–1132. doi: 10.1161/CIRCRESAHA.110.226928. [DOI] [PubMed] [Google Scholar]
- 86.Harhaji Trajković L. M., Mijatović S. A., Maksimović-Ivanić D. D., et al. Anticancer properties of Ganoderma lucidum methanol extracts in vitro and in vivo. Nutrition and Cancer. 2009;61(5):696–707. doi: 10.1080/01635580902898743. [DOI] [PubMed] [Google Scholar]
- 87.Sun J., He H., Xie B. J. Novel antioxidant peptides from fermented mushroom Ganoderma lucidum. Journal of Agricultural and Food Chemistry. 2004;52(21):6646–6652. doi: 10.1021/jf0495136. [DOI] [PubMed] [Google Scholar]
- 88.Lai N.-S., Lin R.-H., Lai R.-S., Kun U.-C., Leu S.-C. Prevention of autoantibody formation and prolonged survival in New Zealand black/New Zealand white F1 mice with an ancient Chinese herb Ganoderma tsugae. Lupus. 2016;10(7):461–465. doi: 10.1191/096120301678416006. [DOI] [PubMed] [Google Scholar]
- 89.Zeng X., Li P., Chen X., et al. Effects of deproteinization methods on primary structure and antioxidant activity of Ganoderma lucidum polysaccharides. International Journal of Biological Macromolecules. 2019;126:867–876. doi: 10.1016/j.ijbiomac.2018.12.222. [DOI] [PubMed] [Google Scholar]
- 90.Tan X., Sun J., Xu Z., et al. Effect of heat stress on production and in-vitro antioxidant activity of polysaccharides in Ganoderma lucidum. Bioprocess and Biosystems Engineering. 2018;41(1):135–141. doi: 10.1007/s00449-017-1850-7. [DOI] [PubMed] [Google Scholar]
- 91.Kumari B., Das P., Kumari R. Accelerated processing of solitary and clustered abasic site DNA damage lesions by APE1 in the presence of aqueous extract of Ganoderma lucidum. Journal of Biosciences. 2016;41(2):265–275. doi: 10.1007/s12038-016-9614-0. [DOI] [PubMed] [Google Scholar]
- 92.Reis F., Lima R., Morales P., Ferreira I., Vasconcelos M. Methanolic extract of Ganoderma lucidum induces autophagy of AGS human gastric tumor cells. Molecules. 2015;20(10):17872–17882. doi: 10.3390/molecules201017872. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Table 1: association sore of 309 targets associated with diabetic cardiomyopathy in 30 recorded tissues. Supplementary Table 2: thirty-two tissue organs expressed 309 targets and types of pathways.