

Abstract
Initially used as antimalarial drugs, hydroxychloroquine (HCQ) and, to a lesser extent, chloroquine (CQ) are currently being used to treat several diseases. Due to its cost‐effectiveness, safety and efficacy, HCQ is especially used in rheumatic autoimmune disorders (RADs), such as systemic lupus erythematosus, primary Sjögren's syndrome and rheumatoid arthritis. Despite this widespread use in the clinic, HCQ molecular modes of action are still not completely understood. By influencing several cellular pathways through different mechanisms, CQ and HCQ inhibit multiple endolysosomal functions, including autophagy, as well as endosomal Toll‐like receptor activation and calcium signalling. These effects alter several aspects of the immune system with the synergistic consequence of reducing pro‐inflammatory cytokine production and release, one of the most marked symptoms of RADs. Here, we review the current knowledge on the molecular modes of action of these drugs and the circumstances under which they trigger side effects. This is of particular importance as the therapeutic use of HCQ is expanding beyond the treatment of malaria and RADs.
Keywords: calcium, chloroquine, cytokines, lysosome, toll‐like receptors
Subject Categories: Immunology, Chemical Biology
Hydroxychloroquine has been heavily discussed in the context of COVID19, but this anti‐malarial drug is primarily used in rheumatic autoimmune disorders (RADs). This comprehensive review recapitulates our knowledge on the mode of action of this drug in RADs, and on its potential applications and side effects.
Glossary
- Antigen‐presenting cells (APC)
Cells that process proteins derived from pathogens or from dying/dead cells, into peptides that get presented on their surface, thereby activating T cells and initiating an immune response.
- Autophagy
An intracellular process that delivers unwanted cytoplasmic material into lysosome for degradation.
- B cells
A type of lymphocytes (white blood cells) that plays a crucial role in the adaptive immune response by producing antigen‐specific antibodies.
- Calcium (Ca2+)
Is the most abundant mineral in the human body and is vital for a multitude of cellular and physiological function. It is also an important second messenger in numerous signal transduction pathways.
- Chloroquine (CQ)/hydroxychloroquine (HCQ)
Originally developed to fight malaria, these drugs are used to treat rheumatic autoimmune diseases and are currently tested in clinical trials as therapies for other conditions.
- Cytokines
Small secreted proteins that mediate communication and modulate interactions between cells, including immune cells.
- Endosomes
Intracellular organelles that mainly function as a sorting and recycling hub for endocytosed and biosynthetic components, on their route to lysosomes.
- Immune system
A network consisting of a variety of different cell types that defend the body against infections and other potentially harmful anomalies, and which, when misregulated, contributes or causes the development of an inflammatory disease.
- Lysosome
Intracellular organelles containing a large battery of digestive enzymes that degrade extracellular and cytoplasmic material delivered to their interior by endocytosis and autophagy, respectively.
- NADPH oxidase
A membrane‐bound multi‐subunit enzymatic complex at either the plasma or endosomal membrane, which participates in a variety of cellular functions, ranging from cellular signalling and gene expression to host defence mechanisms.
- Primary Sjögren's syndrome
An autoimmune disease that belongs to the group of rheumatic autoimmune diseases, which affect saliva‐producing glands leading to symptoms such as dry mouth and dry eyes.
- Retinopathy
Condition characterized by a damaged retina, which causes vision impairment, and is a documented adverse effect that can occur when taking HCQ and CQ.
- Rheumatic autoimmune diseases
A group of conditions characterized by a dysregulated immune system, which primarily affect the muscles, joints, connective tissue and bones.
- Systemic lupus erythematosus
An autoimmune disease that belongs to the group of rheumatic autoimmune diseases, which is the most common form of lupus and is associated with symptoms such as severe fatigue, joint pain and joint swelling.
- T cells
A type of lymphocytes (white blood cells) that is a key component of the adaptive immune system and that orchestrates other cell types in response to antigens.
- Toll‐like receptors (TLR)
Transmembrane proteins that recognize specific molecules at either the plasma membrane or endosomes, and subsequently initiate signalling pathways that are crucial for the innate immune response.
Introduction
Antimalarial drugs have a long history, starting around 400 years ago when quinine, a substance in the bark of the cinchona tree, was first used to fight Plasmodium falciparum infections (Woodward & Doering, 1945; Haładyj et al, 2018). CQ was the first potent and mass‐producible drug against malaria and was synthesized as an analogue of quinine (Shanks, 2016). Despite its remarkable antimalarial efficiency, CQ was deemed too toxic due to its side effects such as gastrointestinal and skin complications, retinopathy, cardiotoxicity or myopathy (Kalia & Dutz, 2007; Haładyj et al, 2018). The discovery of HCQ mitigated this issue, and HCQ is now regularly used in clinics under the brand name Plaquenil (Furst, 1996; Aviña‐Zubieta et al, 1998; Al‐Bari, 2014; Haładyj et al, 2018). Already during the Second World War, the positive effects of these two antimalarial drugs on RADs were observed. Soldiers taking CQ and HCQ as prophylaxis reported improvement of rashes and inflammatory arthritis. Today, CQ and particularly HCQ are commonly used to treat rheumatic and dermatological diseases, and are further being tested in clinical trials as potential drug candidates for COVID‐19, several types of cancer, diabetes type I and II, multiple sclerosis, recurrent miscarriages and myocardial infarction (Al‐Bari, 2014; clinicaltrials.gov).
RADs, such as systemic lupus erythematosus (SLE) (Ruiz‐Irastorza et al, 2010; Willis et al, 2012; Wu et al, 2017), rheumatoid arthritis (RA) (Khraishi & Singh, 1996) and primary Sjögren's syndrome (pSS) (Oxholm et al, 1998; Rihl et al, 2009; Kumar & Clark, 2012; Demarchi et al, 2017), are caused by a malfunctioning immune system that targets healthy tissues (Smith & Germolec, 1999) such as joints (Kumar & Clark, 2012). CQs and HCQs therapeutic role in RADs is linked to its anti‐inflammatory and immunomodulatory effects (Plantone & Koudriavtseva, 2018). These effects are achieved through the modulation of the autoimmune response by (i) impairing functions of the endolysosomal system through its lysosomotropic effects (Ziegler & Unanue, 1982; Kaufmann & Krise, 2007; Yoon et al, 2010), (ii) decreasing the levels of circulating pro‐inflammatory cytokines (Sperber et al, 1993; Van Den Borne et al, 1997), (iii) inhibiting T‐cell proliferation (Landewe et al, 1995; Costedoat‐Chalumeau et al, 2014), (iv) blocking Toll‐like receptors (TLRs) (Kyburz et al, 2006) and (v) autophagy inhibition (An et al, 2017c). However, numerous questions remain regarding both the mechanism of action of CQ and HCQ in RADs and the side effects caused by this compound.
In this review, we report on HCQ and CQ modes of action at the molecular and cellular levels in the context of RADs. Additionally, we discuss the relevance of these drugs in the treatment of cancer and infectious diseases. Finally, we summarize the side effects reported in patients taking HCQ for RADs and discuss how some of those can be explained by the current knowledge on CQ and HCQ.
CQ and HCQ: modes of action
So far, CQ and HCQ have been reported to inhibit four sets of cellular functions: (i) endolysosomal activities, including autophagy; (ii) cytokine signalling, including endosomal Toll‐like receptor (TLRs); (iii) NADPH oxidase (NOX) signalling; and (iv) calcium (Ca2+) mobilization from the endoplasmic reticulum (ER). They might further modulate other cellular and organismal processes, e.g. Golgi trafficking (Mauthe et al, 2018), but the underlying mechanisms remain to be identified.
Inhibition of lysosomal activity and autophagy
CQ and HCQ are weak bases that easily cross cell membranes and accumulate in acidic subcellular compartments such as lysosomes and endosomes, where they remain trapped in a protonated state (Ohkuma & Poole, 1978). This leads to a pH increase in lysosomes from 4 to 6, causing inhibition of acidic proteases and other enzymes within the endolysosomal compartments (Fig 1A) (Ohkuma & Poole, 1978; Poole & Ohkuma, 1981; Ziegler & Unanue, 1982; Haładyj et al, 2018). As a result, antigen processing and subsequent presentation by MHC‐II complex on the cell surface of both macrophages and lymphoid dendritic cells are impaired (Guidos et al, 1984; Chesnut & Grey, 1985; Fox, 1993), dampening the adaptive immune response (Fig 2) (Fox, 1993). CQ and HCQ also increase pH levels within the Golgi stacks. This causes functional alterations of this organelle that possibly contribute to the cellular effects of these two drugs, e.g. by impairing transforming growth factor beta (TGF‐β) activity (Perkett et al, 2006; Rivinoja et al, 2009; Mauthe et al, 2018).
Figure 1. Molecular mechanisms of CQ and HCQ .
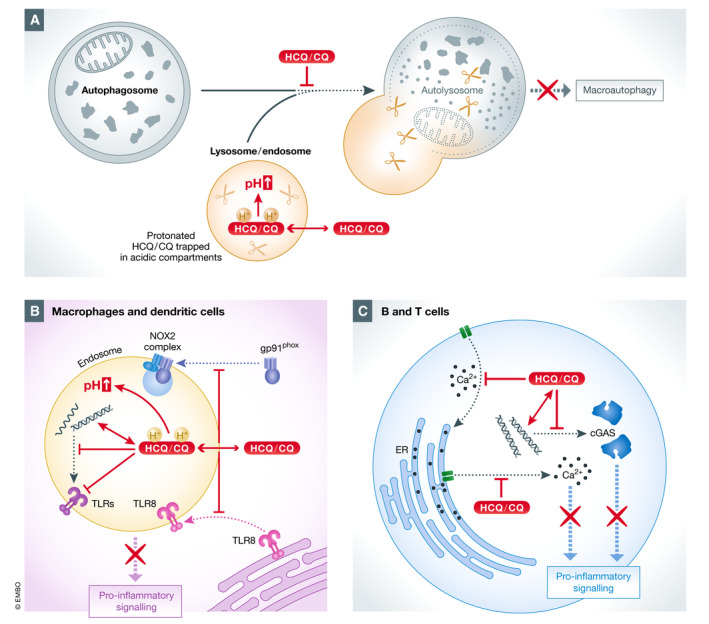
(A) CQ and HCQ are weak bases that accumulate inside acidic subcellular compartments, e.g. endosomes and lysosomes. They remain trapped in a protonated state, causing an increase of pH and thereby inhibiting the functions of these cellular compartments. Impairment of the autophagosome–lysosome fusion leads to autophagy inhibition. (B) CQ and HCQ alter endosomal TLR activation by increasing endosomal pH, by blocking the interaction between nucleic acids and endosomal TLRs (TRL3, TLR7 and TLR9) and by preventing translocation of TLR8 to endosomes. HCQ also blocks the correct assembly of the NOX2 complex by preventing the translocation of the NOX2 subunit gp91phox onto endosomes and consequently the formation of an active NOX2. (C) CQ and HCQ impair the release of Ca2+ from the ER, resulting in inhibition of Ca2+‐dependent signalling pathways. HCQ further inhibits the replenishing of intracellular Ca2+ stores from the extracellular space.
Figure 2. Effects of HCQ on the immune system.
At the cellular level, HCQ inhibits antigen presentation, B‐ and T‐cell activation and NOX signalling. In addition, it rebalances Treg/Th17 cell ratio. These multifaceted effects on different immune cells result in a decreased production and release of pro‐inflammatory cytokines.
The ability to block lysosomal degradation also makes CQ and HCQ potent macroautophagy inhibitors (Fig 1A). Macroautophagy, hereafter called autophagy, is a conserved intracellular degradation pathway that is required to maintain cellular homeostasis by recycling damaged or unwanted cytoplasmic proteins, complexes and organelles (Eskelinen & Saftig, 2009). Autophagy plays a role in many physiological processes, and its misregulation is linked to pathologies such as cancer, neurodegeneration and inflammatory diseases (Mizushima et al, 2008; Levine et al, 2011; Dikic & Elazar, 2018; Levine & Kroemer, 2019). During autophagy, cytoplasmic cargoes are sequestered by double‐membrane vesicles called autophagosomes, which fuse with lysosomes to generate autolysosomes (Eskelinen & Saftig, 2009). Fusion with lysosomes and activity of the lysosomal enzymes are required to break down the autophagosomal cargoes and recycle the resulting metabolites. Impairment of both autophagosome–lysosome fusion and lysosomal degradative activity blocks autophagy (Klionsky et al, 2016). Although CQ and HCQ decrease the acidity of lysosomes (Seglen et al, 1979; Poole & Ohkuma, 1981; Mizushima et al, 2010), the primary inhibitory effect of these drugs on autophagy is blocking the fusion of autophagosomes and lysosomes, which is at least in part mediated by the dysregulation of the recruitment of specific SNARE proteins onto autophagosomes (Mauthe et al, 2018). This block results in an accumulation of autophagosomes in the cytoplasm (Mauthe et al, 2018), which can contribute to an enhanced autophagosome‐mediated signalling output (Martinez‐Lopez et al, 2013; Barrow‐McGee et al, 2016) and even compromise tumour cell viability (Button et al, 2017). Although HCQ and CQ have been extensively described as autophagy inhibitors, there is emerging evidence that these drugs induce a non‐canonical form of endocytosis (Florey et al, 2015; Jacquin et al, 2017).
Inhibition of cytokine signalling
Activation of TLRs, especially in macrophages, monocytes and T helper cells, but also in neutrophils and endothelial cells, induces the production and secretion of pro‐inflammatory cytokines, a hallmark of RADs (Beutler & Cerami, 1989; Feldmann & Maini, 2001; Kim & Moudgil, 2017). Hence, inhibition of endosomal TLRs by HCQ or CQ is a powerful therapy approach for these diseases (Lafyatis et al, 2006). TLR9, activated by DNA in immune cells, can thus be inhibited by HCQ and CQ (Yi et al, 1998; Ahmad‐Nejad et al, 2002). TLR7, activated by guanosine analogues, can also be inhibited by CQ, but to a lesser extent than TLR9 (Lee et al, 2003), indicating different inhibitory mechanisms. TLR3 is mainly activated by poly(I‐C), but also by debris originating from necrotic synovial fluid cells in RA patients, and both modes of activation are hampered by HCQ and CQ (Brentano et al, 2005; Jolly et al, 2014; Imaizumi et al, 2017). In general, inhibition of TLR3, TLR7 and TLR9 by HCQ and CQ has been attributed to their ability to impair endosomal acidification (Macfarlane & Manzel, 1998; Lafyatis et al, 2006; Schrezenmeier & Dörner, 2020), as activation of endosomal TLRs and subsequent downstream signalling only takes place within acidified compartments (Fig 1B) (Blasius & Beutler, 2010).
Beside endosomal acidification, Kuznik and colleagues discovered a second mechanism by which CQ impairs TLR signalling. They showed that CQ could inhibit endosomal TLR signalling after stimulation with nucleic acids at concentration too low to influence the endosomal pH. Under those conditions, CQ blocks endosomal TLR activation by directly interacting with TLR ligands, such as nucleic acids, which changes the nucleic acid secondary structure and prevents their binding to endosomal TLRs (Macfarlane & Manzel, 1998; Kužnik et al, 2011). This notion is further supported by the observation that HCQ specifically blocks activation of dendritic cells and macrophages by DNA but not by LPS, although LPS also stimulates these cells via a signalling cascade emanating from endosomes (Häcker et al, 1998).
A third mechanism that interferes with inflammatory cytokine production is the ability to disrupt GMP‐AMP synthase (cGAS) signalling (An et al, 2015, 2018). cGAS is a crucial component of the cGAS–stimulator of interferon gamma (IFN) genes (STING) signalling cascade that is required for the IFN type I response in immune cells (Sun et al, 2013), making it an important player in activation of pro‐inflammatory response in autoimmune diseases (Gao et al, 2015; Kato et al, 2018). cGAS is also upregulated in a portion of SLE patients (An et al, 2017a,b), and interestingly, HCQ and CQ can inhibit cGAS binding to its ligands, e.g. DNA, in vitro and in a T‐cell line (An et al, 2015). Importantly, inhibition of cGAS activation results in reduced IFNβ expression (An et al, 2015) (Fig 1C).
Inhibition of NADPH oxidase
NOX is a protein complex involved in numerous pro‐inflammatory signalling cascades, such as tumour necrosis factor alpha (TNFα)‐ and interleukin (IL)‐1β‐induced cascades. Activation of endosomal NOX, which leads to the generation of reactive oxygen species (ROS), requires the endocytic internalization and delivery to endosomes of cell surface ligand–receptor complexes (Müller‐Calleja et al, 2017). HCQ blocks the NOX‐mediated signalling cascades triggered by TNFα and IL‐1β in monocytes by blocking translocation of gp91phox, the catalytic subunit of NOX, from the cytosol onto endosomal membranes without changing the endosomal pH (Müller‐Calleja et al, 2017). This inhibition prevents the correct assembly and activation of NOX, hindering the downstream cellular events and the production of the pro‐inflammatory cytokines TNFα and IL‐8. HCQ also prevents the redistribution of TLR8 from the ER to endosomes, which is necessary to mediate the inflammatory response (Müller‐Calleja et al, 2017) (Fig 1B).
Inhibition of Ca2+ signalling
Ca2+ mobilization from both the ER and extracellular space into the cytoplasm and subsequent Ca2+‐dependent signalling is an important mechanism to activate cells of the immune system, such as T and B cells (Feske, 2007). High cytoplasmic levels of Ca2+ act as a second messenger for the activation of signalling pathways and transcription factors that regulate the expression and secretion of cytokines and other immune regulatory factors (Izquierdo et al, 2014). Ca2+ release from the ER can be impaired by HCQ (Goldman et al, 2000; Xu et al, 2015; Wu et al, 2017), leading to the inhibition of intracellular signals. In particular, T‐cell and B‐cell receptor‐mediated intracellular Ca2+ mobilization from both intracellular stores and the extracellular milieu is inhibited by HCQ in a dose‐dependent manner (Goldman et al, 2000). This impairment of Ca2+ mobilization is at least partially caused by the reduction of the Ca2+ stored intracellularly and the inability to replenish these intracellular stores with extracellular Ca2+ (Goldman et al, 2000). This further enhances its negative impact on the Ca2+‐dependent signalling pathways (Fig 1C) (Feske, 2007). The precise mechanism of HCQ‐induced reduction of internal Ca2+ mobilization remains unknown. However, it has been shown that HCQ does not reduce the availability of inositol 1,4,5‐trisphosphate, but rather the binding to its intracellular receptors that promotes Ca2+ release (Misra et al, 1997).
The impact of CQ and HCQ on the immune system in autoimmunity
Autoimmunity is characterized by an overreaction of the immune system (Smith & Germolec, 1999), which is linked to both innate and adaptive immunity (Mescher, 2016). The innate immune system is responsible for the initial recognition of pathogens, which is mostly carried out by antigen‐presenting cells (APCs), e.g. dendritic cells, and eventually triggers the activation of the adaptive immune system (Mescher, 2016). In particular, when APCs get directly activated through exposure to pathogen‐associated molecular patterns, they initiate both cell‐ and antibody‐mediated immune responses, which are mediated by the T and B cells, respectively (Christmas, 2010). The cell‐mediated response is executed by T cells that get activated by APCs through antigen presentation at their surface via MHC molecules. In contrast, B cells are activated through T helper (Th) cells and cytokines that are secreted by APCs (Mescher, 2016). Activated B cells produce and secrete additional pro‐inflammatory cytokines and antibodies to further stimulate the immune reaction (Mescher, 2016).
HCQ and CQ negatively regulate many aspects of these innate and adaptive immune responses by reducing inflammation, and ultimately the severity of autoimmune diseases (Fig 2).
Inhibition of pro‐inflammatory cytokine secretion
Through the inhibition of endosomal TLR signalling, HCQ and CQ treatment decreases the levels of pro‐inflammatory cytokines produced by peripheral mononuclear cells in the blood, including IFNγ (Van Den Borne et al, 1997), TNFα (Picot et al, 1991; Van Den Borne et al, 1997; Jang et al, 2006), IL‐1 (Picot et al, 1991; Sperber et al, 1993; Jang et al, 2006), IL‐6 (Sperber et al, 1993; Van Den Borne et al, 1997; Jang et al, 2006) and IL‐2 (Landewe et al, 1995). The reduction of TLR signalling‐mediated activation of immune cells by both drugs consequently decreases the aberrant immune response and diminishes inflammation symptoms observed in rheumatic patients (da Silva et al, 2013). In addition to directly inhibiting endosomal TLR signalling, CQ and HCQ can interfere with the intracellular signals that lead to both the release of phorbol ester‐induced arachidonic acid and the block of pro‐inflammatory cytokines secretion (e.g. TNFα and IL‐1) in mouse macrophages (Bondeson & Sundler, 1998). In particular, activation of phospholipase A2 by phorbol esters, but not by Ca2+, is inhibited by HCQ and CQ, which blocks the synthesis of arachidonic acid. Furthermore, these compounds negatively impact the generation of zymosan‐induced formation of inositol phosphates, a product of phospholipase C activity (Matsuzawa & Hostetler, 1980), suggesting that they have an inhibitory effect on this enzyme as well (Bondeson & Sundler, 1998). HCQ also inhibits Ca2+‐activated K+ channels in macrophages, and consequently K+ efflux, which could result in impaired inflammasome activation and pro‐inflammatory cytokine release (Eugenia Schroeder et al, 2017).
High levels of pro‐inflammatory cytokines are a central characteristic of the RA pathogenesis (McInnes & Schett, 2007; Blasius & Beutler, 2010; Pollard et al, 2013; Schinnerling et al, 2017; Muskardin & Niewold, 2018). In particular, stimulatory cytokines (i.e. IL‐1, IL‐6, IL‐12, IL‐15, IL‐17, IL‐23 and type I and II IFN for T cells, and B‐cell activating factor (BAFF) for B cells) activate T and B cells, which in turn produce pro‐inflammatory cytokines and autoantibodies, respectively. Pro‐inflammatory cytokines contribute to RA pathogenesis by promoting autoimmunity, maintaining chronic inflammatory synovitis and stimulating the destruction of joint tissues. They also play a role in the maturation and activation of osteoclasts, the cells responsible for breaking down bone tissue (McInnes & Schett, 2007).
Excessive production of BAFF, a cytokine essential for B‐cell physiology, alters the immune tolerance by contributing to the maturation and survival of self‐reactive B cells, the major source for autoantibodies contributing to joint inflammation (Mahdy et al, 2014). Reduction of the high BAFF levels in the serum from RA patients by HCQ (Mahdy et al, 2014) improves symptoms of RADs, both in animal models and in clinical trials (Sun et al, 2008).
Cytokines like BAFF, TNFα, IFNα and IFNγ are also major contributors to SLE severity, by promoting B‐cell survival and autoantibody production, and contributing to organ inflammation (Rönnblom & Elkon, 2010). Thus, the modulation of their levels represents a potential therapeutic avenue (Rönnblom & Elkon, 2010). This is supported by a cohort study showing that treatment of SLE patients with HCQ results in a decrease of type I IFN levels and concomitant reduction of disease severity (Willis et al, 2012). HCQ can also directly affect the production of autoantibodies by B cells through TLR9 inhibition. Particularly, HCQ interferes with the differentiation of memory B cells into antibody‐producing plasmablasts, a subset of B cells, by inhibiting TLR9 activation (Torigoe et al, 2018).
Although the pathogenesis of pSS is not fully understood yet, activation of exocrine gland epithelium cells is thought to lead to the release of pro‐inflammatory cytokines such as IFNα and IFNβ (both type I IFN), IL‐7 and BAFF, and chemokines (Retamozo et al, 2018). These factors stimulate further activation of APCs, but also of T and B cells, which promotes inflammation and autoimmunity (Retamozo et al, 2018). Only a few studies investigated HCQ administration in pSS patients. Nonetheless, pSS patients treated with HCQ have a significant lower BAFF levels in the serum, and an improvement in saliva production (Mumcu et al, 2013), indicating that this drug might be a promising therapy for pSS as well.
Inhibition of B‐ and T‐cell activation through Ca2+ signalling
Through T‐cell receptors (TCRs) on their surface, T cells recognize antigens that are presented by APCs and get activated (Goldman et al, 2000). This results in both their proliferation and the release of various cytokines, including IL‐6 and TNFα (Sperber et al, 1993). One important step in the signalling cascade downstream of TCRs is the increase of intracellular Ca2+ levels, which is released from internal Ca2+ storages such as the ER. As previously mentioned, HCQ can impair the release of Ca2+ from the ER, which consequently inhibits T‐cell activation (Goldman et al, 2000; Xu et al, 2015; Schmidt et al, 2017). HCQ also negatively influences the expression and activity of CD154 on T cells, which is needed for B‐cell activation (Wu et al, 2017; Dewitte et al, 2020). CD154 expression is controlled by the nuclear factor of activated T cells (NFAT), a transcription factor that relies on Ca2+ release from the ER (Wu et al, 2017). By impairing this event, HCQ inhibits NFAT nuclear translocation, resulting in decreased gene expression of CD154 (Wu et al, 2017). Altogether, these studies show that blocking Ca2+ release from the ER by HCQ leads to a multilevel inhibition of T‐ and B‐cell activation, thereby hindering the immune response (Fig 2).
Modulation of Th17 and Treg populations
Alterations in autophagic activity play an important role in the pathophysiology of T‐ and B‐cell‐mediated autoimmunity (Weindel et al, 2015; van Loosdregt et al, 2016; Alessandri et al, 2017; Mocholi et al, 2018; Zhang et al, 2019). In this context, autophagy is required to maintain cellular homeostasis in T cells (An et al, 2017c) and autophagy deficiency impairs MHC class II presentation and contributes to the generation of autoreactive T cells by thymic epithelial cells (Levine et al, 2011). Moreover, plasma cells require autophagy to sustain immunoglobulin production and B‐cell development (Wu & Adamopoulos, 2017). An imbalance within the T‐cell populations, more specifically an increase in the number of Th17 cells and a decrease in that of Treg cells, has been linked to pathogenesis of autoimmune diseases (Yang et al, 2011a; Jadidi‐Niaragh & Mirshafiey, 2012; Álvarez‐Rodríguez et al, 2019), including SLE (An et al, 2017c; Álvarez‐Rodríguez et al, 2019). This imbalance leads to an increased secretion of pro‐inflammatory cytokines such as IL‐17 and IL‐6, and a reduction of the levels of circulating factors like TGF‐β, which suppresses inflammation and autoimmunity (An et al, 2017c; Geng et al, 2020). This latter effect can be dampened with HCQ and CQ, as those drugs rebalance the Th17/Treg ratio (An et al, 2017c; Yang et al, 2018; Álvarez‐Rodríguez et al, 2019; Park et al, 2019; Geng et al, 2020). Mechanistically, this could be caused by an alteration of autophagy, as an induction of this process is observed in SLE patients (An et al, 2017c). Thus, An and colleagues thought to suppress hyperactivated autophagy by administrating HCQ to lupus MLR/pr mice, an animal model for SLE. In addition to lowering autophagic activity in this model, HCQ rebalanced Th17 and Treg cell numbers, which led to a decrease in pro‐inflammatory cytokine levels (Fig 2) and a concomitant augmentation of anti‐inflammatory cytokines, resulting in the suppression of the autoimmune response (An et al, 2017c). Moreover, CQ positively regulates Treg differentiation by stimulating transcriptional activity of Nurr1 and FOXP3, while simultaneously suppressing Th17 differentiation and gene expression (Álvarez‐Rodríguez et al, 2019; Park et al, 2019). More evidence that Th17 cells play a central role in RA and SLE pathogenesis comes from the detection of IL‐6, IL‐17 and IL‐22 in synovial fluids from patients suffering from those diseases (Lubberts et al, 2005; da Silva et al, 2013). High levels of these cytokines correlate with synovial inflammation, T‐cell activation and the osteoclast activity upregulation causing bone erosion (da Silva et al, 2013). Administration of HCQ reduces Th17 cell activation and consequently production of IL‐6, IL‐17 and IL‐22 (da Silva et al, 2013; Yang et al, 2018).
Impact of NOX inhibition on the immune system
NOX inhibition by HCQ impairs the production of pro‐inflammatory cytokines and the correct distribution of TLR8, thereby dampening the immune response (Müller‐Calleja et al, 2017). This inhibition also positively affects nitric oxide (NO) bioavailability (Gómez‐Guzmán et al, 2014). NO is involved in a multitude of physiologic functions, including the regulation of blood vessel tone and vasodilation, and is rapidly inactivated by ROS (Nagy et al, 2010). In SLE patients, NO bioavailability is severely lowered by high ROS levels, particularly O2−, resulting in endothelial dysfunction (Griendling & Alexander, 1997; Landmesser & Harrison, 2001; Gómez‐Guzmán et al, 2014). By blocking NOX, the major producer of O2− in the vascular wall, HCQ treatment reduces ROS levels and helps to prevent endothelial dysfunction in a mouse model for SLE (Gómez‐Guzmán et al, 2014). In agreement with this concept, NOX inhibition by HCQ reduces thrombus formation, which is a well‐known clinical manifestation in SLE, in a venous thrombus mouse model (Müller‐Calleja et al, 2017; Miranda et al, 2019) (Fig 2).
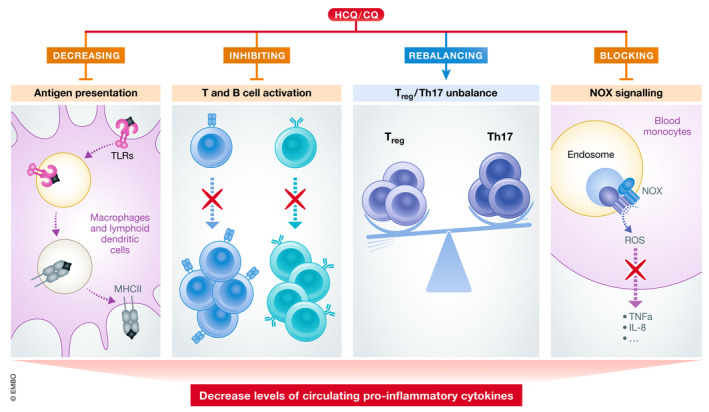
Thus, at the cellular level, HCQ and CQ inhibit antigen presentation, NOX signalling, B‐ and T‐cell activation, and rebalance Treg/Th17 cell ratio. These multifaceted effects on different immune cells synergistically result in a decreased production and release of pro‐inflammatory cytokines, a common hallmark of RADs (Fig 2).
Clinical impact of HCQ on RADs
HCQ is administered orally in tablet form as hydroxychloroquine sulphate (Pastick et al, 2020). It is absorbed in the gastrointestinal tract (Mclachlan et al, 1994) before being widely distributed throughout the body to muscles, liver, spleen, lungs, kidneys, pituitary and adrenal glands, and tissues that contain melanin (Haładyj et al, 2018). Daily dosage of HCQ ranges from 200 to 600 mg for RADs, from 200 to 400 mg for dermatological disorders (Ben‐Zvi et al, 2012), from 200 to 1,200 mg in cancers (Chude & Amaravadi, 2017) and from 200 to 800 mg for various infectious diseases. Its half‐life in the body ranges between 40 and 50 days (Mclachlan et al, 1994), and 30–40% of HCQ is protein‐bound (Furst, 1996), resulting in 60–70% unbound, pharmacologically active drug (Rang et al, 2016). The majority of HCQ is excreted through the kidneys, while the rest is metabolized by the liver or excreted through faeces (Furst et al, 1999; Haładyj et al, 2018). Contraindications for taking HCQ are a history of retinopathy or visual field changes, hypersensitivity to 4‐aminoquinoline compounds and long‐term therapies in children (https://www.fda.gov/). HCQ is, however, considered safe during pregnancy (Kaplan et al, 2016; Haładyj et al, 2018).
HCQ ameliorates classical RAD symptoms, such as skin problems and joint pain, predominantly by decreasing the inflammation reaction in patients (Fig 3). In SLE, HCQ is given to patients as either a single or a combinatorial therapy together with steroids and immunosuppressive drugs, to improve patients’ life expectancy by reducing lupus flares and accrual of organ damage (Ponticelli & Moroni, 2017). Case studies have revealed that HCQ treatment reduces SLE symptoms and improves long‐term survival of patients, while individuals not treated with HCQ have an increased risk of severe SLE exacerbations (James et al, 2007; Ruiz‐Irastorza et al, 2010; Willis et al, 2012).
Figure 3. Beneficial and side effects caused by HCQ in RAD patients.
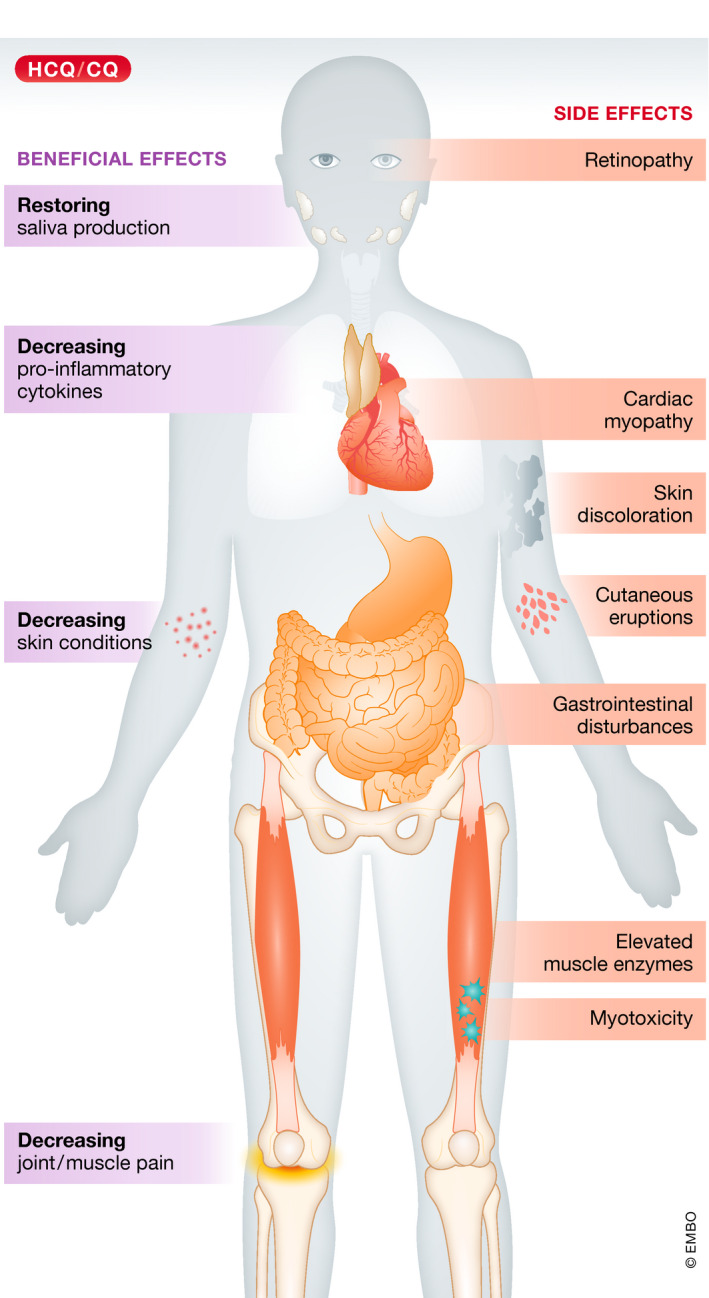
In RADs, HCQ treatment predominantly alleviates the symptoms (purple boxes) by inhibiting the production and release of pro‐inflammatory cytokines. As a consequence, HCQ diminishes skin conditions. There are also indications that HCQ both decreases cartilage degradation and consequently reduces joint and muscle pain, and helps to restore saliva production. Usage of HCQ can cause side effects (orange boxes); the most common are gastrointestinal disturbances, skin discoloration, cutaneous eruptions and elevated muscle enzymes, whereas retinopathy, cardiac myopathy and myotoxicity are rare, but severe.
Similarly, HCQ treatment produces significant clinical improvement and functional capacity in RA patients (Smolen et al, 2014; Haładyj et al, 2018). In RA, prevention of cartilage degradation, which causes joint destruction, is an important aspect of the therapeutic approach (Kumar & Clark, 2012). Cartilage degradation is mostly caused by pro‐inflammatory cytokines, such as IL‐1, IL‐17 and TNFα, and their production can be repressed by HCQ treatment (Picot et al, 1991; Sperber et al, 1993; Van Den Borne et al, 1997; Jang et al, 2006; McInnes & Schett, 2007; da Silva et al, 2013). In vitro experiments have also established that CQ inhibits proteoglycan turnover (Fulkerson et al, 1979; Ackerman et al, 1981; Schug & Kalbhen, 1995; Rainsford et al, 2015), and early autoradiographic studies following tritium‐labelled HCQ have revealed that this drug accumulates in the cartilage of mice (Cecchi & Porzio, 1964). These findings and its water‐soluble properties led to the proposition that HCQ accumulates in the cartilage by binding acidic proteoglycans and protecting them from degradation by proteolytic enzymes (Rainsford et al, 2015). Although an early study pointed out that CQ and HCQ can indeed inhibit cartilage breakdown, slowing down the disease progression and preventing further joint damage in RA patients (Julkunen et al, 1976), more recent investigations could not confirm a positive effects on joint damage (Sanders, 2000; Smolen et al, 2014; Haładyj et al, 2018).
The therapeutic benefits of HCQ administration on pSS classical symptoms, e.g. sicca symptoms, remain controversial; some studies documented beneficial effects (Tishler et al, 1999; Rihl et al, 2009; Yavuz et al, 2011; Mumcu et al, 2013), while others reported none (Gottenberg et al, 2014; Yoon et al, 2016; Wang et al, 2017). HCQ treatment, however, ameliorates extraglandular symptoms (Fox et al, 1996; Demarchi et al, 2017), and according to the Sjögren's Syndrome Foundation's clinical practice guidelines (https://www.sjogrens.org/), disease‐modifying anti‐rheumatic drugs are recommended to treat musculoskeletal pain, with HCQ being the therapeutic approach of choice (Carsons et al, 2015). HCQ also reduces immunological alterations of pSS, such as decreased levels of immunoglobulins, erythrocyte sedimentation rate, serology and IL‐6 production (Tishler et al, 1999; Yavuz et al, 2011; Mumcu et al, 2013). Furthermore, in a retrospective analysis, HCQ administration to pSS patients significantly improved saliva production (Rihl et al, 2009). This improvement was more pronounced in patients who were positive for autoantibodies against anti‐α‐fodrin, an intracellular filamentous cytoskeleton protein. While the cause for this difference remains unknown, a possible explanation is that HCQ could improve saliva production by decreasing elevated levels of cholinesterase, an enzyme that counteracts saliva production (Dawson et al, 2005).
HCQ and CQ in non‐rheumatologic diseases
Anti‐viral effects
The anti‐viral function of HCQ and CQ has mainly been linked to their ability to increase the pH of the endosomal system and the trans‐Golgi network (TGN) (Savarino et al, 2003). Thus, these drugs are able to inhibit cell entry of numerous viruses, as a low endosomal pH is required for the fusion of endocytosed virions with the limiting membrane of endosomes. In this context, CQ and HCQ decrease replication of viruses such as dengue virus (DENV2), chikungunya virus, hepatitis A and C virus, influenza A virus, Zika virus, severe acute respiratory syndrome coronavirus (SARS‐CoV) and Borna disease virus in cellular models (Bishop, 1998; Gonzalez‐Dunia et al, 1998; Keyaerts et al, 2004; Vincent et al, 2005; Blanchard et al, 2006; De Clercq, 2006; Eng et al, 2006; Di Trani et al, 2007; Sourisseau et al, 2007; Khan et al, 2010; Ashfaq et al, 2011; Boonyasuppayakorn et al, 2014; Farias et al, 2015; Delvecchio et al, 2016; Shiryaev et al, 2017). For some viral structural proteins, a maturation step involving post‐translational modification and/or processing in the TGN is crucial for their function and ultimately for the assembly of infectious viral particles, e.g. glycosylation of HIV gp120 (Tsai et al, 1990; Savarino et al, 2004) or cleavage of the DENV2 prM protein (Randolph et al, 1990). Glycosylation in the TGN is also required for the correct assembly of ACE2, the entry receptor for SARS‐CoV (Vincent et al, 2005). Thus, HCQ and CQ contribute to inhibit viral infections by neutralizing the pH of intracellular organelles, interfering with important processes required for viral life cycle.
Although HCQ and CQ have shown beneficial therapeutic effects in animal models for DENV2, hepatitis C virus, avian influenza A virus, Zika virus and SARS‐CoV infections, clinical trials have so far failed to conclusively prove their anti‐viral potential in humans (Rodrigo et al, 2020; Fragkou et al, 2020; McKee et al, 2020). This might be due to the fact that drug concentrations required to de‐acidify intracellular compartments cannot easily be reached in humans (Al‐Bari, 2017). Therefore, neither HCQ nor CQ is currently recommended as anti‐viral drugs (Rodrigo et al, 2020). During the SARS‐CoV‐2 pandemic in 2020, the need to find an effective medication has brought major attention to HCQ and CQ due to their ability to both inhibit viral infections and dampen the massive cytokine response that is observed in SARS‐CoV‐2‐infected patients (Badgujar et al, 2020; Ibáñez et al, 2020; Moore & June, 2020). The effectiveness of HCQ and CQ against SARS‐CoV‐2, however, has so far not been proven in humans, and the results at the time that this review was completed were still controversial (Boulware et al, 2020; Fragkou et al, 2020).
Anti‐cancer therapy
CQ and HCQ are being increasingly used in clinical trials to treat cancer (https://clinicaltrials.gov/). Because high doses are required to achieve anti‐tumoural effects in monotherapies, they are often used in combination with radiotherapy and/or other chemotherapeutical drugs (Plantone & Koudriavtseva, 2018). We briefly discuss here possible mechanisms of action for HCQ and CQ in cancer. For a more detailed discussion on this topic, more specific reviews are available (Manic et al, 2014; Pascolo, 2016; Levy et al, 2017; Shi et al, 2017; Verbaanderd et al, 2017).
Elevated autophagic activity is crucial for tumour cell survival and growth as it supplies the high demand of nutrients within a developed tumour (Amaravadi et al, 2016). This is especially relevant for autophagy‐dependent cancers that rely on this pathway when faced with metabolic stress. Consequently, HCQ or CQ treatment has been successful in regressing the growth of some of those cancers in preclinical studies (e.g. with RAS pathway mutations (Guo et al, 2011; Lock et al, 2011), such as specific pancreatic cancers (Mancias & Kimmelman, 2011; Yang et al, 2011b; Sousa et al, 2016), or BRAF‐driven tumours (Levy et al, 2014; Strohecker et al, 2013; Xie et al, 2015). The effectiveness of HCQ and CQ in cancer therapy is, however, controversial. In animal models, HCQ dosages are often 50 mg/kg/day or higher, which is too high to be administered in humans (Pascolo, 2016), and with lower dosages, autophagy is not sufficiently inhibited to achieve tumour regression (Pascolo, 2016). Moreover, some cancer cells (e.g. derived from breast tumours or melanomas or KRAS‐driven cancer cell lines) have shown CQ‐mediated cell growth inhibition that was independent of autophagy (Maycotte et al, 2012; Maes et al, 2014; Eng et al, 2016).
Various cancer cells express high levels of TLR9, e.g. breast and prostate cancer cells (Merrell et al, 2006; Verbaanderd et al, 2017), which is linked to cancer invasiveness in vitro and associated with poor prognosis (Väisänen et al, 2013; Verbaanderd et al, 2017). TLR9‐mediated NF‐κB signalling is required for cancer cell migration and proliferation in gastric cancer cell models, which is inhibited by CQ (Zhang et al, 2015). The exact molecular mechanism of TLR9 signalling inhibition in cancer cells remains unknown.
Another mechanism by which HCQ affects cancer growth is by modulating the immune system. Tumour‐associated macrophages (TAMs), which are phenotypically described as M2 macrophages, play a role in promoting tumour growth and immune escape, angiogenesis and metastasis (Mantovani et al, 2017; Li et al, 2018). In contrast, tumour killing macrophages (M1 macrophages) have an opposite effect and are activated by cytokines such as IFNγ, which are released from T cells (De Palma & Lewis, 2013; Ostuni et al, 2015). Interestingly, in a melanoma‐bearing mouse model, intraperitoneal injection of 75 mg/kg CQ effectively inhibited melanoma growth in a T‐cell‐dependent manner, and prolonged animal survival (Chen et al, 2018). Mechanistically, CQ can switch TAMs into M1 macrophages by raising lysosomal pH, and thereby mobilizing lysosomal Ca2+ through upregulation of the lysosomal Ca2+ channel MUCOLIPIN1. The release of lysosomal Ca2+ then activates the p38 and NF‐κB pathways, but also the transcription factor EB, resulting in an enhanced anti‐tumour T‐cell response (Chen et al, 2018). By stimulating the T‐cell‐mediated immune response and simultaneously decreasing immune inhibitory cells, including TAMs and Tregs, and cytokines such as TGF‐β and IL‐10, CQ treatment reduced breast cancer growth and prolonged mice survival in a breast xenograft model (Zhang et al, 2017). Another important aspect of anti‐cancer immunity is the activation of immune cells by sensing danger signals (e.g. HMGB1). Danger signals are subsequently recognized by receptors, such as TLR4 on dendritic cells (Apetoh et al, 2007). One function of TLR4 is to preserve engulfed tumour antigens from enhanced degradation, and thereby favour antigen presentation. The loss of antigen presentation capacity in TLR4‐deficient dendritic cells can be restored by CQ, possibly by raising lysosomal pH, which contributed to tumour size reduction in a tlr4 −/− thymoma mouse model (Apetoh et al, 2007). Along these lines, CQ reduced breast cancer growth in mice after irradiation by enhancing apoptotic and immunogenic tumour cell death (Ratikan et al, 2013). The enhanced immune response was attributed to a decreased degradation of tumour antigens in dendritic cells, resulting in an increased antigen presentation (Ratikan et al, 2013).
HCQ and CQ can also inhibit CXCL12/CXCR4 signalling, which is involved in chemotaxis and adhesion of tumour cells and of growth factors secretion that are key for cancer progression (Sun et al, 2010; Kim et al, 2012; Verbaanderd et al, 2017). Moreover, HCQ and CQ interfere with the activation of growth‐promoting pathways in cancer stem cells, thereby suppressing the regrowth of tumours (Li et al, 2008; Balic et al, 2014; Choi et al, 2014).
Multiple reports further describe the mechanisms by which CQ triggers cell death in tumour cells. CQ induces apoptosis of cancer cells by either stimulating the mitochondrial apoptotic pathway (Du Jiang et al, 2010) or activating the p53‐dependent transcription of pro‐apoptotic genes (Zhou et al, 2002; Loehberg et al, 2007, 2012; Maclean et al, 2008; Kim et al, 2010; Bieging et al, 2014). Additionally, several studies have suggested that CQ intercalates into DNA and disturbs chromatin topology (O'Brien et al, 1966; Sternglanz et al, 1969; Field et al, 1978; Yin et al, 2003), which could lead to an impairment in DNA repair mechanisms, and in turn cause DNA damage and enhance cell death (Michael & Williams, 1974; Liang et al, 2016; Weyerhäuser et al, 2018).
Besides directly targeting tumour cells, CQ also affects tumour angiogenesis by altering endothelial cell functionality. CQ administration leads to NOTCH1 accumulation in endothelial cell endosomes, stimulating the downstream signalling that leads to tumour vessel normalization, and resulting in reduced tumour invasion and metastasis (Maes et al, 2014). Therefore, CQ also improves the delivery and efficacy of other chemotherapeutics (Maes et al, 2014).
HCQ and CQ thus show potential in inhibiting tumour growth and modulating tumour immune response through various mechanisms. It is, however, important to reiterate that the doses used to achieve relevant effects in cancer therapies are often substantially higher than the doses used to treat RADs. Moreover, when treating cancer or viral infections, one has to keep in mind that HCQ and CQ also have immune suppressive functions that could negatively influence its beneficial effect for the patients.
Side effects of HCQ in RADs
Side effects of HCQ treatment are rare, but nonetheless exist, and can be very serious, especially during prolonged administration (Haładyj et al, 2018). In Table EV1, we provide a comprehensive overview of the known side effects caused by HCQ in RADs and their prevalence. Overall, the most common side effects in RAD patients taking HCQ or CQ are gastrointestinal disturbances, skin discolorations, cutaneous eruptions and elevated muscle enzymes. Although rare, retinopathy, neuromuscular and cardiac toxicities (Fig 3) are the most serious and life‐threatening side effects potentially triggered by HCQ (Plantone & Koudriavtseva, 2018).
Retinopathy
Prolonged administration of HCQ or CQ can cause retinopathy and loss of retinal function that, when ignored, can result in permanent vision loss (Jorge et al, 2018). The primary site of toxicity in the retina is the photoreceptor layer, with secondary degeneration occurring later in retinal pigment epithelium (RPE) cells (De Sisternes et al, 2015; Yusuf et al, 2017). Some studies offer a potential explanation for this severe side effect.
By inhibiting the lysosomal degradation capacity and possibly endocytosis in RPE cells, HCQ and CQ are preventing the degradation of old and spent outer segments of photoreceptors in the RPE, a process that is required to maintain its function and preserve vision (Kevany & Palczewski, 2010; Yusuf et al, 2017). Furthermore, HCQ entrapment in the RPE might lead to an accumulation of lipofuscin, which is associated with photoreceptor function impairment and consequent vision loss (Kevany & Palczewski, 2010; Yusuf et al, 2017). It has been speculated that, due to this entrapment, retinopathy still continues in some cases after cessation of HCQ treatment (Michaelides et al, 2011). Accumulation of CQ in the pigmented ocular tissue, which comprises RPE cells, the iris, the choroid and the ciliary body, and eventually in the retina, was also observed in rhesus monkeys when CQ was administered for 52 months (Rosenthal et al, 1978). This caused an initial damage to the photoreceptors and the ganglion cells, followed by a disruption of both the RPE and choroid, which ultimately led to visual impairments and retinopathy (Rosenthal et al, 1978).
High levels of HCQ inhibit the function of the organic anion transporting polypeptide 1A2 (OATP1A2), a plasma membrane importer expressed in many tissues, including RPE cells (Xu et al, 2016). In particular, OATP1A2 transports all‐trans‐retinol (atROL), a retinol precursor essential for the classic visual cycle (Chan et al, 2015), into RPE cells. By blocking this transporter, HCQ causes an extracellular accumulation of atROL and disrupts the classic visual cycle (Xu et al, 2016).
Cardiac side effects and myotoxicity
HCQ can cause acute and chronic cardiac adverse effects (Chatre et al, 2018). Acute adverse effects are linked to a very high dose of HCQ, which provokes a block of Na+ and Ca2+ channels. This inhibition can lead to membrane‐stabilization effects in cardiac muscle cells, which in turn causes conduction disturbances with atrioventricular block and QRS interval widening (White, 2007). Chronic adverse effects are connected to long‐term treatment with a high cumulative dose of HCQ (Chatre et al, 2018). As described above, HCQ treatment impairs the degradative activity of lysosomes, which leads to an accumulation of material such as glycogen and phospholipids in their interior (Chatre et al, 2018). In myocytes, this causes a vascularization of the cytoplasm and myofibrillar disorganization, which contributes to the development of cardiac myopathy and myocardial fibrosis (Yogasundaram et al, 2014). This phenomenon can also be seen in the Fabry and Danon lysosomal storage diseases, which have similar phenotypes (Roos et al, 2002; D'souza et al, 2014; Chatre et al, 2018). Moreover, HCQ‐mediated accumulation of autophagosomes in muscles and peripheral nerves can lead to myotoxicity or myotoxicity combined with peripheral nerve dysfunction (Shukla et al, 2019). Notably, HCQ and CQ also have proarrhythmic activity (Landmesser & Harrison, 2001; Khobragade et al, 2013; Chansky & Werth, 2017; Naksuk et al, 2020), which is of particular importance because of the potential use of this drug to treat COVID‐19 patients. These patients are burdened by arrhythmic events, and consequently, HCQ and CQ could worsen this pathological feature. It is still under investigation whether this proarrhythmic activity is caused by SARS‐CoV‐2 infection and whether HCQ and CQ are influencing it (Lazzerini et al, 2020).
Conclusions
HCQ is nowadays widely used for the treatment of RADs and has shown great success in improving the quality of life of many patients. Over the years, research on the molecular and cellular mode of action of HCQ (and CQ) revealed that this compound modulates molecular processes and cellular responses in multiple ways. At least four mechanisms of action that, directly or indirectly, influence the immune system by synergistically dampening pro‐inflammatory responses, have been described. Although lysosomal inhibition and autophagy impairment are the most studied, HCQ also influences other important immune regulatory pathways by inhibiting specific steps, such as activation of endosomal TLR‐, cGAS and NOX signalling and Ca2+ mobilization for the ER. The beneficial therapeutic effect of HCQ in RADs probably lies in its multifaceted properties, which also makes it a promising candidate in other medical fields, such as oncology (Onorati et al, 2018) and microbiology (Savarino et al, 2003; Cortegiani et al, 2020; Yao et al, 2020).
Generally, HCQ is considered a safe drug with low prevalence of side effects. These side effects nevertheless exist and can impact the life of a patient tremendously. Among them, the most severe, i.e. retinopathy and cardiomyopathy, is linked to the induced lysosomal activity inhibition. This suggests that the unwanted negative effects of HCQ could be due to its lysosomotropic properties. In this context, it has been reported that the effect of HCQ on endosomal and lysosomal pH at therapeutic concentrations is negligible (Kužnik et al, 2011) and that the pH changes observed in vitro might not reflect the in vivo reality. Therefore, a higher dose of HCQ (or a higher cumulative dose) could lead to a pH increase in the compartments of the endolysosomal system and thus cause more side effects (Latasiewicz et al, 2017; Jorge et al, 2018). The well‐documented list of side effects caused by HCQ during the treatment of RADs should be considered when using HCQ to treat other pathologies such as cancer (Onorati et al, 2018), neurodegenerative disorders (Hedya et al, 2019), metabolic diseases (Pasquier, 2016) and microbial infections (Savarino et al, 2003), especially since treatment of some pathologies requires high HCQ doses (Leung et al, 2015).
While the search for a unifying mechanism of action for HCQ is tempting, current knowledge shows that this small molecule has more than a single target. As a result, future research should aim at identifying potential additional cellular and organismal pathways specifically modulated by HCQ. The mechanisms by which HCQ causes side effects could also provide important information. Increasing our understanding of HCQ mode of action would improve patient outcome by promoting therapeutic benefits while reducing side effects.
Pending issues.
-
(i)
Investigate whether all HCQ modes of action described with in vitro experiments are relevant in patients, and whether one of these mechanisms is predominantly causing the observed side effects.
-
(ii)
Determine whether HCQ has other molecular effects than the ones described, which could help to better understand HCQ treatment outcomes in patients.
-
(iii)
Chemically improve HCQ to make it more effective and less toxic, and thereby render it more suitable for the treatment of other diseases (e.g. specific cancers).
-
(iv)
Understand how the anti‐inflammatory role of HCQ influences the anti‐viral and anti‐tumorigenic action of this drug in patients, and whether this could explain the observed discrepancies between the in vitro and in vivo results.
For more information
Conflict of interest
The authors declare that they have no conflict of interest.
Supporting information
Table EV1
Acknowledgements
M.M. is supported by ZonMW TOP (91217002) grant. F.R. is supported by ALW Open Programme (ALWOP.310), Marie Skłodowska‐Curie Cofund (713660) and Marie Skłodowska Curie ETN (765912) and ZonMW TOP (91217002) grants. We thank Zarina Rorije and Asaad Amasha for helping assembling information about the reported side effects of HCQ. We thank Muriel Mari, Ruben Gomez Sanchez, Adriana Tami and Pauline Verlhac for the critical reading of the manuscript. The authors apologize in advance to those authors whose contributions have been omitted due to lack of space and felt that their work was one of the highlights.
EMBO Mol Med (2020) 12: e12476
References
- Ackerman NR, Jubb SN, Marlowe SL (1981) Effects of various anti‐inflammatory and anti‐rheumatic agents on the synthesis, secretion, and activity of a cartilage proteoglycan‐degrading enzyme and other macrophage enzymes. Biochem Pharmacol 30: 2147–2155 [DOI] [PubMed] [Google Scholar]
- Ahmad‐Nejad P, Häcker H, Rutz M, Bauer S, Vabulas RM, Wagner H (2002) Bacterial CpG‐DNA and lipopolysaccharides activate toll‐like receptors at distinct cellular compartments. Eur J Immunol 32: 1958–1968 [DOI] [PubMed] [Google Scholar]
- Al‐Bari AA (2014) Chloroquine analogues in drug discovery: New directions of uses, mechanisms of actions and toxic manifestations from malaria to multifarious diseases. J Antimicrob Chemother 70: 1608–1621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al‐Bari MAA (2017) Targeting endosomal acidification by chloroquine analogs as a promising strategy for the treatment of emerging viral diseases. Pharmacol Res Perspect 5: e00293 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alessandri C, Ciccia F, Priori R, Astorri E, Guggino G, Alessandro R, Rizzo A, Conti F, Minniti A, Barbati C et al (2017) CD4 T lymphocyte autophagy is upregulated in the salivary glands of primary Sjögren's syndrome patients and correlates with focus score and disease activity. Arthritis Res Ther 19: 178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Álvarez‐Rodríguez L, Martínez‐Taboada V, Calvo‐Alén J, Beares I, Villa I, López‐Hoyos M (2019) Altered Th17/treg ratio in peripheral blood of systemic lupus erythematosus but not primary antiphospholipid syndrome. Front Immunol 10: 391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amaravadi R, Kimmelman AC, White E (2016) Recent insights into the function of autophagy in cancer. Genes Dev 30: 1913–1930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- An J, Woodward JJ, Sasaki T, Minie M, Elkon KB (2015) Cutting edge: antimalarial drugs inhibit IFN‐β production through blockade of cyclic GMP‐AMP synthase–DNA interaction. J Immunol 194: 4089–4093 [DOI] [PubMed] [Google Scholar]
- An J, Durcan L, Karr RM, Briggs TA, Rice GI, Teal TH, Woodward JJ, Elkon KB (2017a) Expression of cyclic GMP‐AMP synthase in patients with systemic lupus erythematosus. Arthritis Rheumatol 69: 800–807 [DOI] [PubMed] [Google Scholar]
- An J, Minie M, Sasaki T, Woodward JJ, Elkon KB (2017b) Antimalarial drugs as immune modulators: new mechanisms for old drugs. Annu Rev Med 68: 317–330 [DOI] [PubMed] [Google Scholar]
- An N, Chen Y, Wang C, Yang C, Wu ZH, Xue J, Ye L, Wang S, Liu HF, Pan Q (2017c) Chloroquine autophagic inhibition rebalances Th17/Treg‐mediated immunity and ameliorates systemic lupus erythematosus. Cell Physiol Biochem 44: 412–422 [DOI] [PubMed] [Google Scholar]
- An J, Woodward JJ, Lai W, Minie M, Sun X, Tanaka L, Snyder JM, Sasaki T, Elkon KB (2018) Inhibition of cyclic GMP‐AMP synthase using a novel antimalarial drug derivative in Trex1‐deficient mice. Arthritis Rheumatol 70: 1807–1819 [DOI] [PubMed] [Google Scholar]
- Apetoh L, Ghiringhelli F, Tesniere A, Obeid M, Ortiz C, Criollo A, Mignot G, Maiuri MC, Ullrich E, Saulnier P et al (2007) Toll‐like receptor 4‐dependent contribution of the immune system to anticancer chemotherapy and radiotherapy. Nat Med 13: 1050–1059 [DOI] [PubMed] [Google Scholar]
- Ashfaq UA, Javed T, Rehman S, Nawaz Z, Riazuddin S (2011) Lysosomotropic agents as HCV entry inhibitors. Virol J 8: 163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aviña‐Zubieta JA, Galindo‐Rodriguez G, Newman S, Suarez‐Almazor ME, Russell AS (1998) Long term effectiveness of antimalarial drugs in rheumatic diseases. Ann Rheum Dis 57: 582–587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Badgujar KC, Badgujar AB, Patil VP, Dhangar DV (2020) Hydroxychloroquine for COVID‐19: a review and a debate based on available clinical trials/case studies. J Drug Deliv Ther 10: 304–311 [Google Scholar]
- Balic A, Sørensen MD, Trabulo SM, Sainz B, Cioffi M, Vieira CR, Miranda‐Lorenzo I, Hidalgo M, Kleeff J, Erkan M et al (2014) Chloroquine targets pancreatic cancer stem cells via inhibition of CXCR4 and hedgehog signaling. Mol Cancer Ther 13: 1758–1771 [DOI] [PubMed] [Google Scholar]
- Barrow‐McGee R, Kishi N, Joffre C, Ménard L, Hervieu A, Bakhouche BA, Noval AJ, Mai A, Guzmán C, Robert‐Masson L et al (2016) Beta 1‐integrin‐c‐Met cooperation reveals an inside‐in survival signalling on autophagy‐related endomembranes. Nat Commun 7: 11942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben‐Zvi I, Kivity S, Langevitz P, Shoenfeld Y (2012) Hydroxychloroquine: from malaria to autoimmunity. Clin Rev Allergy Immunol 42: 145–153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beutler B, Cerami A (1989) The biology of cachectin/TNF ‐ A primary mediator of the host response. Annu Rev Immunol 7: 625–655 [DOI] [PubMed] [Google Scholar]
- Bieging KT, Mello SS, Attardi LD (2014) Unravelling mechanisms of p53‐mediated tumour suppression. Nat Rev Cancer 14: 359–370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bishop NE (1998) Examination of potential inhibitors of hepatitis A virus uncoating. Intervirology 41: 261–271 [DOI] [PubMed] [Google Scholar]
- Blanchard E, Belouzard S, Goueslain L, Wakita T, Dubuisson J, Wychowski C, Rouillé Y (2006) Hepatitis C virus entry depends on clathrin‐mediated endocytosis. J Virol 80: 6964–6972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blasius AL, Beutler B (2010) Intracellular toll‐like receptors. Immunity 32: 305–315 [DOI] [PubMed] [Google Scholar]
- Bondeson J, Sundler R (1998) Antimalarial drugs inhibit phospholipase A2 activation and induction of interleukin 1β and tumor necrosis factor α in macrophages: implications for their mode of action in rheumatoid arthritis. Gen Pharmacol 30: 357–366 [DOI] [PubMed] [Google Scholar]
- Boonyasuppayakorn S, Reichert ED, Manzano M, Nagarajan K, Padmanabhan R (2014) Amodiaquine, an antimalarial drug, inhibits dengue virus type 2 replication and infectivity. Antiviral Res 106: 125–134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boulware DR, Pullen MF, Bangdiwala AS, Pastick KA, Lofgren SM, Okafor EC, Skipper CP, Nascene AA, Nicol MR, Abassi M et al (2020) A randomized trial of hydroxychloroquine as postexposure prophylaxis for Covid‐19. N Engl J Med 10.1056/nejmoa2016638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brentano F, Schorr O, Gay RE, Gay S, Kyburz D (2005) RNA released from necrotic synovial fluid cells activates rheumatoid arthritis synovial fibroblasts via Toll‐like receptor 3. Arthritis Rheum 52: 2656–2665 [DOI] [PubMed] [Google Scholar]
- Button RW, Roberts SL, Willis TL, Oliver Hanemann C, Luo S (2017) Accumulation of autophagosomes confers cytotoxicity. J Biol Chem 292: 13599–13614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carsons SE, Parke A, Vivino F, Carteron N, Brasington R, Fox R, Kassan S, Scofield RH, Birnbaum J, Mandel S et al (2015) Sjögren's foundation clinical practice guidelines. Arthritis Care Res 10: 13 [Google Scholar]
- Cecchi E, Porzio F (1964) Affinité de l‐hydroxychloroquine pour les tissues articulaires. Rhumatologie 16: 399–400 [PubMed] [Google Scholar]
- Chan T, Zhu L, Madigan MC, Wang K, Shen W, Gillies MC, Zhou F (2015) Human organic anion transporting polypeptide 1A2 (OATP1A2) mediates cellular uptake of all‐trans‐retinol in human retinal pigmented epithelial cells. Br J Pharmacol 172: 2343–2353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chansky PB, Werth VP (2017) Accidental hydroxychloroquine overdose resulting in neurotoxic vestibulopathy. BMJ Case Rep 2017: bcr2016218786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatre C, Roubille F, Vernhet H, Jorgensen C, Pers YM (2018) Cardiac complications attributed to chloroquine and hydroxychloroquine: a systematic review of the literature. Drug Saf 41: 919–931 [DOI] [PubMed] [Google Scholar]
- Chen D, Xie J, Fiskesund R, Dong W, Liang X, Lv J, Jin X, Liu J, Mo S, Zhang T et al (2018) Chloroquine modulates antitumor immune response by resetting tumor‐associated macrophages toward M1 phenotype. Nat Commun 9: 873 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chesnut RW, Grey HM (1985) Antigen presenting cells and mechanisms of antigen presentation. Crit Rev Immunol 5: 263–316 [PubMed] [Google Scholar]
- Choi DS, Blanco E, Kim YS, Rodriguez AA, Zhao H, Huang THM, Chen CL, Jin G, Landis MD, Burey LA et al (2014) Chloroquine eliminates cancer stem cells through deregulation of Jak2 and DNMT1. Stem Cells 32: 2309–2323 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christmas P (2010) Toll‐like receptors: sensors that detect infection. Nat Educ 3: 85 [Google Scholar]
- Chude CI, Amaravadi RK (2017) Targeting autophagy in cancer: update on clinical trials and novel inhibitors. Int J Mol Sci 18: 1279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortegiani A, Ingoglia G, Ippolito M, Giarratano A, Einav S (2020) A systematic review on the efficacy and safety of chloroquine for the treatment of COVID‐19. J Crit Care 57: 279–283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costedoat‐Chalumeau N, Dunogué B, Morel N, Le Guern V, Guettrot‐Imbert G (2014) Hydroxychloroquine: a multifaceted treatment in lupus. Press Medicale 43: e167–e180 [DOI] [PubMed] [Google Scholar]
- Dawson LJ, Caulfield VL, Stanbury JB, Field AE, Christmas SE, Smith PM (2005) Hydroxychloroquine therapy in patients with primary Sjögren's syndrome may improve salivary gland hypofunction by inhibition of glandular cholinesterase. Rheumatology 44: 449–455 [DOI] [PubMed] [Google Scholar]
- De Clercq E (2006) Potential antivirals and antiviral strategies against SARS coronavirus infections. Expert Rev Anti Infect Ther 4: 291–302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Palma M, Lewis CE (2013) Macrophage regulation of tumor responses to anticancer therapies. Cancer Cell 23: 277–286 [DOI] [PubMed] [Google Scholar]
- De Sisternes L, Hu J, Rubin DL, Marmor MF (2015) Localization of damage in progressive hydroxychloroquine retinopathy on and off the drug: inner versus outer retina, parafovea versus peripheral fovea. Invest Ophthalmol Vis Sci 56: 3415–3426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delvecchio R, Higa LM, Pezzuto P, Valadão AL, Garcez PP, Monteiro FL, Loiola EC, Dias AA, Silva FJM, Aliota MT et al (2016) Chloroquine, an endocytosis blocking agent, inhibits zika virus infection in different cell models. Viruses 8: 322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demarchi J, Papasidero S, Medina MA, Klajn D, Chaparro del Moral R, Rillo O, Martiré V, Crespo G, Secco A, Catalan Pellet A et al (2017) Primary Sjögren's syndrome: extraglandular manifestations and hydroxychloroquine therapy. Clin Rheumatol 36: 2455–2460 [DOI] [PubMed] [Google Scholar]
- Dewitte A, Villeneuve J, Lepreux S, Bouchecareilh M, Gauthereau X, Rigothier C, Combe C, Ouattara A, Ripoche J (2020) CD154 induces interleukin‐6 secretion by kidney tubular epithelial cells under hypoxic conditions: inhibition by chloroquine. Mediators Inflamm 2020: 6357046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Trani L, Savarino A, Campitelli L, Norelli S, Puzelli S, D'Ostilio D, Vignolo E, Donatelli I, Cassone A (2007) Different pH requirements are associated with divergent inhibitory effects of chloroquine on human and avian influenza A viruses. Virol J 4: 39 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dikic I, Elazar Z (2018) Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol 19: 349–364 [DOI] [PubMed] [Google Scholar]
- D'souza RS, Levandowski C, Slavov D, Graw SL, Allen LA, Adler E, Mestroni L, Taylor MRG (2014) Danon disease clinical features, evaluation, and management. Circ Hear Fail 7: 843–849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du Jiang P, Zhao YL, Deng XQ, Mao YQ, Shi W, Tang QQ, Li ZG, Zheng YZ, Yang SY, Wei YQ (2010) Antitumor and antimetastatic activities of chloroquine diphosphate in a murine model of breast cancer. Biomed Pharmacother 64: 609–614 [DOI] [PubMed] [Google Scholar]
- Eng EO, Chew JSW, Jin PL, Chua RCS (2006) In vitro inhibition of human influenza A virus replication by chloroquine. Virol J 3: 39 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eng CH, Wang Z, Tkach D, Toral‐Barza L, Ugwonali S, Liu S, Fitzgerald SL, George E, Frias E, Cochran N et al (2016) Macroautophagy is dispensable for growth of KRAS mutant tumors and chloroquine efficacy. Proc Natl Acad Sci USA 113: 182–187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eskelinen EL, Saftig P (2009) Autophagy: a lysosomal degradation pathway with a central role in health and disease. Biochim Biophys Acta Mol Cell Res 1793: 664–673 [DOI] [PubMed] [Google Scholar]
- Eugenia Schroeder M, Russo S, Costa C, Hori J, Tiscornia I, Bollati‐Fogolín M, Zamboni DS, Ferreira G, Cairoli E, Hill M (2017) Pro‐inflammatory Ca++‐activated K+ channels are inhibited by hydroxychloroquine. Sci Rep 7: 1–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farias KJS, Machado PRL, Muniz JAPC, Imbeloni AA, Da Fonseca BAL (2015) Antiviral activity of chloroquine against dengue virus type 2 replication in aotus monkeys. Viral Immunol 28: 161–169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldmann M, Maini RN (2001) Anti‐TNFα therapy of rheumatoid arthritis: what have we learned? Annu Rev Immunol 19: 163–196 [DOI] [PubMed] [Google Scholar]
- Feske S (2007) Calcium signalling in lymphocyte activation and disease. Nat Rev Immunol 7: 690–702 [DOI] [PubMed] [Google Scholar]
- Field RC, Gibson BR, Holbrook DJ, McCall BM (1978) Inhibition of precursor incorporation into nucleic acids of mammalian tissues by antimalarial aminoquinolines. Br J Pharmacol 62: 159–164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Florey O, Gammoh N, Kim SE, Jiang X, Overholtzer M (2015) V‐ATPase and osmotic imbalances activate endolysosomal LC3 lipidation. Autophagy 11: 88–99 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox RI (1993) Mechanism of action of hydroxychloroquine as an antirheumatic drug. Semin Arthritis Rheum 23: 82–91 [DOI] [PubMed] [Google Scholar]
- Fox RI, Dixon R, Guarrasi V, Krubel S (1996) Treatment of primary Sjögren's syndrome with hydroxychloroquine: a retrospective, open‐label study. Lupus 5: 31–36 [PubMed] [Google Scholar]
- Fragkou PC, Belhadi D, Peiffer‐Smadja N, Moschopoulos CD, Lescure X, Janocha H, Karofylakis E, Yazdanpanah Y, Mentré F, Skevaki C et al (2020) Review of trials currently testing treatment and prevention of COVID‐19. Clin Microbiol Infect 10.1016/j.cmi.2020.05.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulkerson JP, Ladenbauer‐Bellis IM, Chrisman OD (1979) In vitro hexosamine depletion of intact articular cartilage by E‐prostaglandins: prevention by chloroquine. Arthritis Rheum 22: 1117–1121 [DOI] [PubMed] [Google Scholar]
- Furst DE (1996) Pharmacokinetics of hydroxychloroquine and chloroquine during treatment of rheumatic diseases. Lupus 5: S11–S15 [PubMed] [Google Scholar]
- Furst DE, Lindsley H, Baethge B, Botstein GR, Caldwell J, Dietz F, Ettlinger R, Golden HE, McLaughlin GE, Moreland LW, et al (1999) Dose‐loading with hydroxychloroquine improves the rate of response in early, active rheumatoid arthritis: a randomized, double‐blind six‐week trial with eighteen‐week extension. Arthritis Rheum 42: 357–365 [DOI] [PubMed] [Google Scholar]
- Gao D, Li T, Li XD, Chen X, Li QZ, Wight‐Carter M, Chen ZJ (2015) Activation of cyclic GMP‐AMP synthase by self‐DNA causes autoimmune diseases. Proc Natl Acad Sci USA 112: E5699–E5705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geng L, Tang X, Wang S, Sun Y, Wang D, Tsao BP, Feng X, Sun L (2020) Reduced let‐7f in Bone marrow‐derived mesenchymal stem cells triggers Treg/Th17 imbalance in patients with systemic lupus erythematosus. Front Immunol 11: 233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman FD, Gilman AL, Hollenback C, Kato RM, Premack BA, Rawlings DJ (2000) Hydroxychloroquine inhibits calcium signals in T cells: a new mechanism to explain its immunomodulatory properties. Blood 95: 3460–3466 [PubMed] [Google Scholar]
- Gómez‐Guzmán M, Jiménez R, Romero M, Sánchez M, Zarzuelo MJ, Gómez‐Morales M, O'Valle F, López‐Farré AJ, Algieri F, Gálvez J et al (2014) Chronic hydroxychloroquine improves endothelial dysfunction and protects kidney in a mouse model of systemic lupus erythematosus. Hypertension 64: 330–337 [DOI] [PubMed] [Google Scholar]
- Gonzalez‐Dunia D, Cubitt B, de la Torre JC (1998) Mechanism of borna disease virus entry into cells. J Virol 72: 783–788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottenberg JE, Ravaud P, Puéchal X, Le Guern V, Sibilia J, Goeb V, Larroche C, Dubost JJ, Rist S, Saraux A et al (2014) Effects of hydroxychloroquine on symptomatic improvement in primary Sjögren syndrome: the JOQUER randomized clinical trial. J Am Med Assoc 312: 249–258 [DOI] [PubMed] [Google Scholar]
- Griendling KK, Alexander RW (1997) Oxidative stress and cardiovascular disease. Circulation 96: 3264–3265 [PubMed] [Google Scholar]
- Guidos C, Wong M, Lee KC (1984) A comparison of the stimulatory activities of lymphoid dendritic cells and macrophages in T proliferative responses to various antigens. J Immunol 133: 1179–1184 [PubMed] [Google Scholar]
- Guo JY, Chen HY, Mathew R, Fan J, Strohecker AM, Karsli‐Uzunbas G, Kamphorst JJ, Chen G, Lemons JMS, Karantza V et al (2011) Activated Ras requires autophagy to maintain oxidative metabolism and tumorigenesis. Genes Dev 25: 460–470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Häcker H, Mischak H, Miethke T, Liptay S, Schmid R, Sparwasser T, Heeg K, Lipford GB, Wagner H (1998) CpG‐DNA‐specific activation of antigen‐presenting cells requires stress kinase activity and is preceded by non‐specific endocytosis and endosomal maturation. EMBO J 17: 6230–6240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haładyj E, Sikora M, Felis‐Giemza A, Olesinska M (2018) Antimalarials ‐ are they effective and safe in rheumatic diseases? Reumatologia 56: 164–173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedya SA, Safar MM, Bahgat AK (2019) Hydroxychloroquine antiparkinsonian potential: Nurr1 modulation versus autophagy inhibition. Behav Brain Res 365: 82–88 [DOI] [PubMed] [Google Scholar]
- Ibáñez S, Martínez O, Valenzuela F, Silva F, Valenzuela O (2020) Hydroxychloroquine and chloroquine in COVID‐19: should they be used as standard therapy? Perspect Rheumatol 39: 2461–2465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imaizumi T, Hayakari R, Matsumiya T, Yoshida H, Tsuruga K, Watanabe S, Kawaguchi S, Tanaka H (2017) Chloroquine attenuates TLR3/IFN‐β signaling in cultured normal human mesangial cells: a possible protective effect against renal damage in lupus nephritis. Mod Rheumatol 27: 1004–1009 [DOI] [PubMed] [Google Scholar]
- Izquierdo J‐H, Bonilla‐Abadía F, Cañas CA, Tobón GJ (2014) Calcium, channels, intracellular signaling and autoimmunity. Reumatol Clínica 10: 43–47 [DOI] [PubMed] [Google Scholar]
- Jacquin E, Leclerc‐Mercier S, Judon C, Blanchard E, Fraitag S, Florey O (2017) Pharmacological modulators of autophagy activate a parallel noncanonical pathway driving unconventional LC3 lipidation. Autophagy 13: 854–867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jadidi‐Niaragh F, Mirshafiey A (2012) The deviated balance between regulatory T cell and Th17 in autoimmunity. Immunopharmacol Immunotoxicol 34: 727–739 [DOI] [PubMed] [Google Scholar]
- James JA, Kim‐Howard XR, Bruner BF, Jonsson MK, McClain MT, Arbuckle MR, Walker C, Dennis GJ, Merrill JT, Harley JB (2007) Hydroxychloroquine sulfate treatment is associated with later onset of systemic lupus erythematosus. Lupus 16: 401–409 [DOI] [PubMed] [Google Scholar]
- Jang CH, Choi JH, Byun MS, Jue DM (2006) Chloroquine inhibits production of TNF‐α, IL‐1β and IL‐6 from lipopolysaccharide‐stimulated human monocytes/macrophages by different modes. Rheumatology 45: 703–710 [DOI] [PubMed] [Google Scholar]
- Jolly L, Stavrou A, Vanderstoken G, Meliopoulos VA, Habgood A, Tatler AL, Porte J, Knox A, Weinreb P, Violette S et al , et al (2014) Influenza promotes collagen deposition via αvβ6 integrin‐mediated transforming growth factor β activation. J Biol Chem 289: 35246–35263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorge A, Ung C, Young LH, Melles RB, Choi HK (2018) Hydroxychloroquine retinopathy — implications of research advances for rheumatology care. Nat Rev Rheumatol 14: 693–703 [DOI] [PubMed] [Google Scholar]
- Julkunen H, Rokkanen P, Laine H (1976) Chloroquine treatment and bone changes in rheumatoid arthritis. Scand J Rheumatol 5: 36–38 [PubMed] [Google Scholar]
- Kalia S, Dutz JP (2007) New concepts in antimalarial use and mode of action in dermatology. Dermatol Ther 20: 160–174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan YC, Ozsarfati J, Nickel C, Koren G (2016) Reproductive outcomes following hydroxychloroquine use for autoimmune diseases: a systematic review and meta‐analysis. Br J Clin Pharmacol 81: 835–848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato Y, Park J, Takamatsu H, Konaka H, Aoki W, Aburaya S, Ueda M, Nishide M, Koyama S, Hayama Y et al (2018) Apoptosis‐derived membrane vesicles drive the cGAS‐STING pathway and enhance type i IFN production in systemic lupus erythematosus. Ann Rheum Dis 77: 1507–1515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann AM, Krise JP (2007) Lysosomal sequestration of amine‐containing drugs: analysis and therapeutic implications. J Pharm Sci 96: 729–746 [DOI] [PubMed] [Google Scholar]
- Kevany BM, Palczewski K (2010) Phagocytosis of retinal rod and cone photoreceptors. Physiology 25: 8–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keyaerts E, Vijgen L, Maes P, Neyts J, Van Ranst M (2004) In vitro inhibition of severe acute respiratory syndrome coronavirus by chloroquine. Biochem Biophys Res Commun 323: 264–268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan M, Santhosh SR, Tiwari M, Lakshmana Rao PV, Parida M (2010) Assessment of in vitro prophylactic and therapeutic efficacy of chloroquine against Chikungunya virus in Vero cells. J Med Virol 82: 817–824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khobragade SB, Gupta P, Gurav P, Chaudhari G, Gatne MM, Shingatgeri VM (2013) Assessment of proarrhythmic activity of chloroquine in in vivo and ex vivo rabbit models. J Pharmacol Pharmacother 4: 116–124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khraishi MM, Singh G (1996) The role of anti‐malarials in rheumatoid arthritis ‐ The American experience. Lupus 5: S41–S44 [PubMed] [Google Scholar]
- Kim EL, Wüstenberg R, Rübsam A, Schmitz‐Salue C, Warnecke G, Bücker EM, Pettkus N, Speidel D, Rohde V, Schulz‐Schaeffer W et al (2010) Chloroquine activates the p53 pathway and induces apoptosis in human glioma cells. Neuro Oncol 12: 389–400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J, Yip MLR, Shen X, Li H, Hsin LYC, Labarge S, Heinrich EL, Lee W, Lu J, Vaidehi N (2012) Identification of anti‐malarial compounds as novel antagonists to chemokine receptor CXCR4 in pancreatic cancer cells. PLoS ONE 7: e31004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim EY, Moudgil KD (2017) Immunomodulation of autoimmune arthritis by pro‐inflammatory cytokines. Cytokine 98: 87–96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klionsky DJ, Abdelmohsen K, Abe A, Abedin MJ, Abeliovich H, Arozena AA, Adachi H, Adams CM, Adams PD, Adeli K et al (2016) Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 12: 1–222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar P, Clark ML (2012) Kumar and clark's clinical medicine, 9th edn London: Elsevier Health Science; [Google Scholar]
- Kužnik A, Benčina M, Švajger U, Jeras M, Rozman B, Jerala R (2011) Mechanism of endosomal TLR inhibition by antimalarial drugs and imidazoquinolines. J Immunol 186: 4794–4804 [DOI] [PubMed] [Google Scholar]
- Kyburz D, Brentano F, Gay S (2006) Mode of action of hydroxychloroquine in RA ‐ Evidence of an inhibitory effect on toll‐like receptor signaling. Nat Clin Pract Rheumatol 2: 458–459 [DOI] [PubMed] [Google Scholar]
- Lafyatis R, York M, Marshak‐Rothstein A (2006) Antimalarial agents: closing the gate on toll‐like receptors? Arthritis Rheum 54: 3068–3070 [DOI] [PubMed] [Google Scholar]
- Landewe RBM, Miltenburg AMM, Verdonk MJA, Verweij CL, Breedveld FC, Daha MR, Dijkmans BAC (1995) Chloroquine inhibits T cell proliferation by interfering with IL‐2 production and responsiveness. Clin Exp Immunol 102: 144–151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landmesser U, Harrison DG (2001) Oxidative stress and vascular damage in hypertension. Coron Artery Dis 12: 455–461 [DOI] [PubMed] [Google Scholar]
- Latasiewicz M, Gourier H, Yusuf IH, Luqmani R, Sharma SM, Downes SM (2017) Hydroxychloroquine retinopathy: an emerging problem. Eye 31: 972–976 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazzerini PE, Boutjdir M, Capecchi PL (2020) COVID‐19, arrhythmic risk and inflammation: mind the gap!. Circulation 142: 7–9 [DOI] [PubMed] [Google Scholar]
- Lee J, Chuang TH, Redecke V, She L, Pitha PM, Carson DA, Raz E, Cottam HB (2003) Molecular basis for the immunostimulatory activity of guanine nucleoside analogs: activation of toll‐like receptor 7. Proc Natl Acad Sci USA 100: 6646–6651 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung LSB, Neal JW, Wakelee HA, Sequist LV, Marmor MF (2015) Rapid onset of retinal toxicity from high‐dose hydroxychloroquine given for cancer therapy. Am J Ophthalmol 160: 799–805.e1 [DOI] [PubMed] [Google Scholar]
- Levine B, Mizushima N, Virgin HW (2011) Autophagy in immunity and inflammation. Nature 469: 323–335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levine B, Kroemer G (2019) Biological functions of autophagy genes: a disease perspective. Cell 176: 11–42 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy JMM, Thompson JC, Griesinger AM, Amani V, Donson AM, Birks DK, Morgan MJ, Mirsky DM, Handler MH, Foreman NK et al (2014) Autophagy inhibition improves chemosensitivity in BRAFV600E brain tumors. Cancer Discov 4: 773–780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy JMM, Towers CG, Thorburn A (2017) Targeting autophagy in cancer. Nat Rev Cancer 17: 528–542 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Lewis MT, Huang J, Gutierrez C, Osborne CK, Wu MF, Hilsenbeck SG, Pavlick A, Zhang X, Chamness GC et al (2008) Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J Natl Cancer Inst 100: 672–679 [DOI] [PubMed] [Google Scholar]
- Li Y, Cao F, Li M, Li P, Yu Y, Xiang L, Xu T, Lei J, Tai YY, Zhu J et al (2018) Hydroxychloroquine induced lung cancer suppression by enhancing chemo‐sensitization and promoting the transition of M2‐TAMs to M1‐like macrophages. J Exp Clin Cancer Res 37: 259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang DH, Choi DS, Ensor JE, Kaipparettu BA, Bass BL, Chang JC (2016) The autophagy inhibitor chloroquine targets cancer stem cells in triple negative breast cancer by inducing mitochondrial damage and impairing DNA break repair. Cancer Lett 376: 249–258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lock R, Roy S, Kenific CM, Su JS, Salas E, Ronen SM, Debnath J (2011) Autophagy facilitates glycolysis during Ras‐mediated oncogenic transformation. Mol Biol Cell 22: 165–178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loehberg CR, Thompson T, Kastan MB, Maclean KH, Edwards DG, Kittrell FS, Medina D, Conneely OM, O'Malley BW (2007) Ataxia telangiectasia‐mutated and p53 are potential mediators of chloroquine‐induced resistance to mammary carcinogenesis. Cancer Res 67: 12026–12033 [DOI] [PubMed] [Google Scholar]
- Loehberg CR, Strissel PL, Dittrich R, Strick R, Dittmer J, Dittmer A, Fabry B, Kalender WA, Koch T, Wachter DL et al (2012) Akt and p53 are potential mediators of reduced mammary tumor growth by Chloroquine and the mTOR inhibitor RAD001. Biochem Pharmacol 83: 480–488 [DOI] [PubMed] [Google Scholar]
- van Loosdregt J, Rossetti M, Spreafico R, Moshref M, Olmer M, Williams GW, Kumar P, Copeland D, Pischel K, Lotz M et al (2016) Increased autophagy in CD4+ T cells of rheumatoid arthritis patients results in T‐cell hyperactivation and apoptosis resistance. Eur J Immunol 46: 2862–2870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lubberts E, Koenders M, van den Berg WB (2005) The role of T cell interleukin‐17 in conducting destructive arthritis: lessons from animal models. Arthritis Res Ther 7: 29–37 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macfarlane DE, Manzel L (1998) Antagonism of immunostimulatory CpG‐oligodeoxynucleotides by quinacrine, chloroquine, and structurally related compounds. J Immunol 160: 1122–1131 [PubMed] [Google Scholar]
- Maclean KH, Dorsey FC, Cleveland JL, Kastan MB (2008) Targeting lysosomal degradation induces p53‐dependent cell death and prevents cancer in mouse models of lymphomagenesis. J Clin Invest 118: 79–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maes H, Kuchnio A, Peric A, Moens S, Nys K, DeBock K, Quaegebeur A, Schoors S, Georgiadou M, Wouters J et al (2014) Tumor vessel normalization by chloroquine independent of autophagy. Cancer Cell 26: 190–206 [DOI] [PubMed] [Google Scholar]
- Mahdy AA, Raafat HA, El‐Fishawy HS, Gheita TA (2014) Therapeutic potential of hydroxychloroquine on serum B‐cell activating factor belonging to the tumor necrosis factor family (BAFF) in rheumatoid arthritis patients. Bull Fac Pharm Cairo Univ 52: 37–43 [Google Scholar]
- Mancias JD, Kimmelman AC (2011) Targeting autophagy addiction in cancer. Oncotarget 2: 1302–1306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manic G, Obrist F, Kroemer G, Vitale I, Galluzzi L (2014) Chloroquine and hydroxychloroquine for cancer therapy. Mol Cell Oncol 1: e29911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mantovani A, Marchesi F, Malesci A, Laghi L, Allavena P (2017) Tumour‐associated macrophages as treatment targets in oncology. Nat Rev Clin Oncol 14: 399–416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez‐Lopez N, Athonvarangkul D, Mishall P, Sahu S, Singh R (2013) Autophagy proteins regulate ERK phosphorylation. Nat Commun 4: 2799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuzawa Y, Hostetler KY (1980) Inhibition of lysosomal phospholipase A and phospholipase C by chloroquine and 4,4′‐bis(diethylaminoethoxy)α, β‐diethyldiphenylethane. J Biol Chem 255: 5190–5194 [PubMed] [Google Scholar]
- Mauthe M, Orhon I, Rocchi C, Zhou X, Luhr M, Hijlkema KJ, Coppes RP, Engedal N, Mari M, Reggiori F (2018) Chloroquine inhibits autophagic flux by decreasing autophagosome‐lysosome fusion. Autophagy 14: 1435–1455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maycotte P, Aryal S, Cummings CT, Thorburn J, Morgan MJ, Thorburn A (2012) Chloroquine sensitizes breast cancer cells to chemotherapy independent of autophagy. Autophagy 8: 200–212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McInnes IB, Schett G (2007) Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol 7: 429–442 [DOI] [PubMed] [Google Scholar]
- McKee DL, Sternberg A, Stange U, Laufer S, Naujokat C (2020) Candidate drugs against SARS‐CoV‐2 and COVID‐19. Pharmacol Res 157: 104859 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mclachlan AJ, Tett SE, Cutler DJ, Day RO (1994) Bioavailability of hydroxychloroquine tablets in patients with rheumatoid arthritis. Rheumatology 33: 235–239 [DOI] [PubMed] [Google Scholar]
- Merrell MA, Ilvesaro JM, Lehtonen N, Sorsa T, Gehrs B, Rosenthal E, Chen D, Shackley B, Harris KW, Selander KS (2006) Toll‐like receptor 9 agonists promote cellular invasion by increasing matrix metalloproteinase activity. Mol Cancer Res 4: 437–447 [DOI] [PubMed] [Google Scholar]
- Mescher AL (2016) Junqueira's basic histology text & atlas, 14th edn, pp 295–304. New York: McGraw Hill; [Google Scholar]
- Michael RO, Williams GM (1974) Choloroquine inhibition of repair of DNA damage induced in mammalian cells by methyl methanesulfonate. Mutat Res, Fundam Mol Mech Mutagen 25: 391–396 [DOI] [PubMed] [Google Scholar]
- Michaelides M, Stover NB, Francis PJ, Weleber RG (2011) Retinal toxicity associated with hydroxychloroquine and chloroquine: risk factors, screening, and progression despite cessation of therapy. Arch Ophthalmol 129: 30–39 [DOI] [PubMed] [Google Scholar]
- Miranda S, Billoir P, Damian L, Thiebaut PA, Schapman D, Le Besnerais M, Jouen F, Galas L, Levesque H, Le Cam‐Duchez V et al (2019) Hydroxychloroquine reverses the prothrombotic state in a mouse model of antiphospholipid syndrome: role of reduced inflammation and endothelial dysfunction. PLoS ONE 14: e0212614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misra UK, Gawdi G, Pizzo SV (1997) Chloroquine, quinine and quinidine inhibit calcium release from macrophage intracellular stores by blocking inositol 1,4,5‐trisphosphate binding to its receptor. J Cell Biochem 64: 225–232 [DOI] [PubMed] [Google Scholar]
- Mizushima N, Levine B, Cuervo AM, Klionsky DJ (2008) Autophagy fights disease through cellular self‐digestion. Nature 451: 1069–1075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizushima N, Yoshimori T, Levine B (2010) Methods in mammalian autophagy research. Cell 140: 313–326 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mocholi E, Dowling SD, Botbol Y, Gruber RC, Ray AK, Vastert S, Shafit‐Zagardo B, Coffer PJ, Macian F (2018) Autophagy is a tolerance‐avoidance mechanism that modulates TCR‐mediated signaling and cell metabolism to prevent induction of T cell anergy. Cell Rep 24: 1136–1150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore JB, June CH (2020) Cytokine release syndrome in severe COVID‐19. Science 368: 473–474 [DOI] [PubMed] [Google Scholar]
- Müller‐Calleja N, Manukyan D, Canisius A, Strand D, Lackner KJ (2017) Hydroxychloroquine inhibits proinflammatory signalling pathways by targeting endosomal NADPH oxidase. Ann Rheum Dis 76: 891–897 [DOI] [PubMed] [Google Scholar]
- Mumcu G, Biçakçigil M, Yilmaz N, Ozay H, Karaçayliümit Cimilli H, Yavuz S (2013) Salivary and Serum B‐cell activating factor (BAFF) levels after hydroxychloroquine treatment in primary Sjögren's syndrome. Oral Heal Prev Dent 11: 229–234 [DOI] [PubMed] [Google Scholar]
- Muskardin TLW, Niewold TB (2018) Type I interferon in rheumatic diseases. Nat Rev Rheumatol 14: 214–228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy G, Koncz A, Telarico T, Fernandez D, Érsek B, Buzás E, Perl A (2010) Central role of nitric oxide in the pathogenesis of rheumatoid arthritis and sysemic lupus erythematosus. Arthritis Res Ther 12: 210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naksuk N, Lazar S, Peeraphatdit TB (2020) Cardiac safety of off‐label COVID‐19 drug therapy: a review and proposed monitoring protocol. Eur Heart J Acute Cardiovasc Care 9: 215–221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Brien RL, Olenick JG, Hahn FE (1966) Reactions of quinine, chloroquine, and quinacrine with DNA and their effects on the DNA and RNA polymerase reactions. Proc Natl Acad Sci USA 55: 1511–1517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohkuma S, Poole B (1978) Fluorescence probe measurement of the intralysosomal pH in living cells and the perturbation of pH by various agents. Proc Natl Acad Sci USA 75: 3327–3331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onorati AV, Dyczynski M, Ojha R, Amaravadi RK (2018) Targeting autophagy in cancer. Cancer 124: 3307–3318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostuni R, Kratochvill F, Murray PJ, Natoli G (2015) Macrophages and cancer: from mechanisms to therapeutic implications. Trends Immunol 36: 229–239 [DOI] [PubMed] [Google Scholar]
- Oxholm P, Prause JU, Schiødt M (1998) Rational drug therapy recommendations for the treatment of patients with Sjogren's syndrome. Drugs 56: 345–353 [DOI] [PubMed] [Google Scholar]
- Park TY, Jang Y, Kim W, Shin J, Toh HT, Kim CH, Yoon HS, Leblanc P, Kim KS (2019) Chloroquine modulates inflammatory autoimmune responses through Nurr1 in autoimmune diseases. Sci Rep 9: 15559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pascolo S (2016) Time to use a dose of Chloroquine as an adjuvant to anti‐cancer chemotherapies. Eur J Pharmacol 771: 139–144 [DOI] [PubMed] [Google Scholar]
- Pasquier B (2016) Autophagy inhibitors. Cell Mol Life Sci 73: 985–1001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastick KA, Okafor EC, Wang F, Lofgren SM, Skipper CP, Nicol MR, Pullen MF, Rajasingham R, McDonald EG, Lee TC et al (2020) Review: hydroxychloroquine and chloroquine for treatment of SARS‐CoV‐2 (COVID‐19). Open Forum Infect Dis 7: ofaa130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkett EA, Ornatowski W, Poschet JF, Deretic V (2006) Chloroquine normalizes aberrant transforming growth factor beta activity in cystic fibrosis bronchial epithelial cells. Pediatr Pulmonol 41: 771–778 [DOI] [PubMed] [Google Scholar]
- Picot S, Peyron F, Vuillez JP, Polack B, Ambroise‐Thomas P (1991) Chloroquine inhibits tumor necrosis factor production by human macrophages in vitro. J Infect Dis 164: 830 [DOI] [PubMed] [Google Scholar]
- Plantone D, Koudriavtseva T (2018) Current and future use of chloroquine and hydroxychloroquine in infectious, immune, neoplastic, and neurological diseases: a mini‐review. Clin Drug Invest 38: 653–671 [DOI] [PubMed] [Google Scholar]
- Pollard KM, Cauvi DM, Toomey CB, Morris KV, Kono DH (2013) Interferon‐γ and systemic autoimmunity. Discov Med 16: 123–131 [PMC free article] [PubMed] [Google Scholar]
- Ponticelli C, Moroni G (2017) Hydroxychloroquine in systemic lupus erythematosus (SLE). Expert Opin Drug Saf 16: 411–419 [DOI] [PubMed] [Google Scholar]
- Poole B, Ohkuma S (1981) Effect of weak bases on the intralysosomal pH in mouse peritoneal macrophages. J Cell Biol 90: 665–669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rainsford KD, Parke AL, Clifford‐Rashotte M, Kean WF (2015) Therapy and pharmacological properties of hydroxychloroquine and chloroquine in treatment of systemic lupus erythematosus, rheumatoid arthritis and related diseases. Inflammopharmacology 23: 231–269 [DOI] [PubMed] [Google Scholar]
- Randolph VB, Winkler G, Stollar V (1990) Acidotropic amines inhibit proteolytic processing of flavivirus prM protein. Virology 174: 450–458 [DOI] [PubMed] [Google Scholar]
- Rang HP, Ritter JM, Flower RJ, Henderson G (2016) Rang and dale's pharmacology, 8th edn London: Elsevier; [Google Scholar]
- Ratikan JA, Sayre JW, Schaue D (2013) Chloroquine engages the immune system to eradicate irradiated breast tumors in mice. Int J Radiat Oncol Biol Phys 87: 761–768 [DOI] [PubMed] [Google Scholar]
- Retamozo S, Flores‐Chavez A, Consuegra‐Fernández M, Lozano F, Ramos‐Casals M, Brito‐Zerón P (2018) Cytokines as therapeutic targets in primary Sjögren syndrome. Pharmacol Ther 184: 81–97 [DOI] [PubMed] [Google Scholar]
- Rihl M, Ulbricht K, Schmidt RE, Witte T (2009) Treatment of sicca symptoms with hydroxychloroquine in patients with Sjögren's syndrome. Rheumatology 48: 796–799 [DOI] [PubMed] [Google Scholar]
- Rivinoja A, Hassinen A, Kokkonen N, Kauppila A, Kellokumpu S (2009) Elevated golgi pH impairs terminal N‐glycosylation by inducing mislocalization of golgi glycosyltransferases. J Cell Physiol 220: 144–154 [DOI] [PubMed] [Google Scholar]
- Rodrigo C, Fernando SD, Rajapakse S (2020) Clinical evidence for repurposing chloroquine and hydroxychloroquine as antiviral agents: a systematic review. Clin Microbiol Infect 10.1016/j.cmi.2020.05.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rönnblom L, Elkon KB (2010) Cytokines as therapeutic targets in SLE. Nat Rev Rheumatol 6: 339–347 [DOI] [PubMed] [Google Scholar]
- Roos JM, Aubry MC, Edwards WD (2002) Chloroquine cardiotoxicity: clinicopathologic features in three patients and comparison with three patients with Fabry disease. Cardiovasc Pathol 11: 277–283 [DOI] [PubMed] [Google Scholar]
- Rosenthal AR, Kolb H, Bergsma D, Huxsoll D, Hopkins JL (1978) Chloroquine retinopathy in the rhesus monkey. Invest Ophthalmol Vis Sci 17: 1158–1175 [PubMed] [Google Scholar]
- Ruiz‐Irastorza G, Ramos‐Casals M, Brito‐Zeron P, Khamashta MA (2010) Clinical efficacy and side effects of antimalarials in systemic lupus erythematosus: a systematic review. Ann Rheum Dis 69: 20–28 [DOI] [PubMed] [Google Scholar]
- Sanders M (2000) A review of controlled clinical trials examining the effects of antimalarial compounds and gold compounds on radiographic progression in rheumatoid arthritis. J Rheumatol 27: 523–529 [PubMed] [Google Scholar]
- Savarino A, Boelaert JR, Cassone A, Majori G, Cauda R (2003) Effects of chloroquine on viral infections: an old drug against today's diseases? Lancet Infect Dis. 3: 722–727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savarino A, Lucia MB, Rastrelli E, Rutella S, Golotta C, Morra E, Tamburrini E, Perno CF, Boelaert JR, Sperber K et al (2004) Anti‐HIV effects of chloroquine: inhibition of viral particle glycosylation and synergism with protease inhibitors. J Acquir Immune Defic Syndr 35: 223–232 [DOI] [PubMed] [Google Scholar]
- Schinnerling K, Aguillón JC, Catalán D, Soto L (2017) The role of interleukin‐6 signalling and its therapeutic blockage in skewing the T cell balance in rheumatoid arthritis. Clin Exp Immunol 189: 12–20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt RLJ, Jutz S, Goldhahn K, Witzeneder N, Gerner MC, Trapin D, Greiner G, Hoermann G, Steiner G, Pickl WF et al (2017) Chloroquine inhibits human CD4+ T‐cell activation by AP‐1 signaling modulation. Sci Rep 7: 42191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schrezenmeier E, Dörner T (2020) Mechanisms of action of hydroxychloroquine and chloroquine: implications for rheumatology. Nat Rev Rheumatol 16: 155–166 [DOI] [PubMed] [Google Scholar]
- Schug BS, Kalbhen DA (1995) Influence of chloroquine and other substances on the collagenolytic activity in human osteoarthritic cartilage in vitro. Arzneimittel‐Forschung/Drug Res 45: 285–289 [PubMed] [Google Scholar]
- Seglen PO, Grinde B, Solheim AE (1979) Inhibition of the lysosomal pathway of protein degradation in isolated rat hepatocytes by ammonia, methylamine, chloroquine and leupeptin. Eur J Biochem 95: 215–225 [DOI] [PubMed] [Google Scholar]
- Shanks GD (2016) Historical review: problematic malaria prophylaxis with quinine. Am J Trop Med Hyg 95: 269–272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi TT, Yu XX, Yan LJ, Xiao HT (2017) Research progress of hydroxychloroquine and autophagy inhibitors on cancer. Cancer Chemother Pharmacol 79: 287–294 [DOI] [PubMed] [Google Scholar]
- Shiryaev SA, Mesci P, Pinto A, Fernandes I, Sheets N, Shresta S, Farhy C, Huang CT, Strongin AY, Muotri AR et al (2017) Repurposing of the anti‐malaria drug chloroquine for Zika Virus treatment and prophylaxis. Sci Rep 7: 15771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shukla S, Gultekin S, Saporta M (2019) Pearls & Oy‐sters: hydroxychloroquine‐induced toxic myopathy mimics Pompe disease: critical role of genetic test. Neurology 92: E742–E745 [DOI] [PubMed] [Google Scholar]
- da Silva JC, Mariz HA, da Rocha Júnior LF, de Oliveira PSS, Dantas AT, Duarte ALBP, da Pitta IR, Galdino SL, da Pitta MGR (2013) Hydroxychloroquine decreases Th17‐related cytokines in systemic lupus erythematosus and rheumatoid arthritis patients. Clinics 68: 766–771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith DA, Germolec DR (1999) Introduction to immunology and autoimmunity Environ health perspect 107: 661–665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smolen JS, Landewé R, Breedveld FC, Buch M, Burmester G, Dougados M, Emery P, Gaujoux‐Viala C, Gossec L, Nam J et al (2014) EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease‐modifying antirheumatic drugs: 2013 update. Ann Rheum Dis 73: 492–509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sourisseau M, Schilte C, Casartelli N, Trouillet C, Guivel‐Benhassine F, Rudnicka D, Sol‐Foulon N, Le Roux K, Prevost MC, Fsihi H et al (2007) Characterization of reemerging chikungunya virus. PLoS Pathog 3: 0804–0817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sousa CM, Biancur DE, Wang X, Halbrook CJ, Sherman MH, Zhang L, Kremer D, Hwang RF, Witkiewicz AK, Ying H et al (2016) Pancreatic stellate cells support tumour metabolism through autophagic alanine secretion. Nature 536: 479–483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperber K, Quraishi H, Kalb TH, Panja A, Stecher V, Mayer L (1993) Selective regulation of cytokine secretion by hydroxychloroquine: inhibition of interleukin 1 alpha (IL‐1‐α) and IL‐6 in human monocytes and T cells. J Rheumatol 20: 803–808 [PubMed] [Google Scholar]
- Sternglanz H, Yielding KL, Pruitt KM (1969) Nuclear magnetic resonance studies of the interaction of chloroquine diphosphate with adenosine 5′‐phosphate and other nucleotides. Mol Pharmacol 5: 376–381 [PubMed] [Google Scholar]
- Strohecker AM, Guo JY, Karsli‐Uzunbas G, Price SM, Chen GJ, Mathew R, McMahon M, White E (2013) Autophagy sustains mitochondrial glutamine metabolism and growth of BrafV600E ‐driven lung tumors. Cancer Discov 3: 1272–1285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun J, Lin Z, Feng J, Li Y, Shen B (2008) BAFF‐targeting therapy, a promising strategy for treating autoimmune diseases. Eur J Pharmacol 597: 1–5 [DOI] [PubMed] [Google Scholar]
- Sun X, Cheng G, Hao M, Zheng J, Zhou X, Zhang J, Taichman RS, Pienta KJ, Wang J (2010) CXCL12 / CXCR4 / CXCR7 chemokine axis and cancer progression. Cancer Metastasis Rev 29: 709–722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun L, Wu J, Du F, Chen X, Chen ZJ (2013) Cyclic GMP‐AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339: 786–791 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tishler M, Yaron I, Shirazi I, Yaron M (1999) Hydroxychloroquine treatment for primary Sjoegren's syndrome: its effect on salivary and serum inflammatory markers. Ann Rheum Dis 58: 253–256 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torigoe M, Sakata K, Ishii A, Iwata S, Nakayamada S, Tanaka Y (2018) Hydroxychloroquine efficiently suppresses inflammatory responses of human class‐switched memory B cells via Toll‐like receptor 9 inhibition. Clin Immunol 195: 1–7 [DOI] [PubMed] [Google Scholar]
- Tsai WPO, Nara PL, Kung HFU, Oroszlan S (1990) Inhibition of human immunodeficiency virus infectivity by chloroquine. AIDS Res Hum Retroviruses 6: 481–489 [DOI] [PubMed] [Google Scholar]
- Väisänen MR, Jukkola‐Vuorinen A, Vuopala KS, Selander KS, Vaarala MH (2013) Expression of Toll‐like receptor‐9 is associated with poor progression‐free survival in prostate cancer. Oncol Lett 5: 1659–1663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Den Borne BEEM, Dijkmans BAC, De Rooij HH, Le Cessie S, Verweij CL (1997) Chloroquine and hydroxychloroquine equally affect tumor necrosis factor‐α, interleukin 6, and interferon‐γ production by peripheral blood mononuclear cells. J Rheumatol 24: 55–60 [PubMed] [Google Scholar]
- Verbaanderd C, Maes H, Schaaf MB, Sukhatme VP, Pantziarka P, Sukhatme V, Agostinis P, Bouche G (2017) Repurposing drugs in oncology (ReDO) ‐ Chloroquine and hydroxychloroquine as anti‐cancer agents. Ecancermedicalscience 11: 781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vincent MJ, Bergeron E, Benjannet S, Erickson BR, Rollin PE, Ksiazek TG, Seidah NG, Nichol ST (2005) Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol J 2: 69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang SQ, Zhang LW, Wei P, Hua H (2017) Is hydroxychloroquine effective in treating primary Sjogren's syndrome: a systematic review and meta‐analysis. BMC Musculoskelet Disord 18: 186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weindel CG, Richey LJ, Bolland S, Mehta AJ, Kearney JF, Huber BT (2015) B cell autophagy mediates TLR7‐dependent autoimmunity and inflammation. Autophagy 11: 1010–1024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weyerhäuser P, Kantelhardt SR, Kim EL (2018) Re‐purposing chloroquine for glioblastoma: potential merits and confounding variables. Front Oncol 8: 335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- White NJ (2007) Cardiotoxicity of antimalarial drugs. Lancet Infect Dis 7: 549–558 [DOI] [PubMed] [Google Scholar]
- Willis R, Seif AM, McGwin G, Martinez‐Martinez LA, González EB, Dang N, Papalardo E, Liu J, Vilá LM, Reveille JD et al (2012) Effect of hydroxychloroquine treatment on pro‐inflammatory cytokines and disease activity in SLE patients: data from LUMINA (LXXV), a multiethnic US cohort. Lupus 21: 830–835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodward RB, Doering WE (1945) The total synthesis of quinine. J Am Chem Soc 67: 860–874 [Google Scholar]
- Wu DJ, Adamopoulos IE (2017) Autophagy and autoimmunity. Clin Immunol 176: 55–62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu SF, Bin Chang C, Hsu JM, Lu MC, Lai NS, Li C, Tung CH (2017) Hydroxychloroquine inhibits CD154 expression in CD4+ T lymphocytes of systemic lupus erythematosus through NFAT, but not STAT5, signaling. Arthritis Res Ther 19: 183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie X, Koh JY, Price S, White E, Mehnert JM (2015) Atg7 overcomes senescence and promotes growth of BrafV600E‐driven melanoma. Cancer Discov 5: 410–423 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu JC, Peng YB, Wei MY, Wu YF, Guo D, Qin G, Ji G, Shen J, Liu QH (2015) Chloroquine inhibits Ca signaling in murine CD4 + thymocytes. Cell Physiol Biochem 36: 133–140 [DOI] [PubMed] [Google Scholar]
- Xu C, Zhu L, Chan T, Lu X, Shen W, Madigan MC, Gillies MC, Zhou F (2016) Chloroquine and hydroxychloroquine are novel inhibitors of human organic anion transporting polypeptide 1A2. J Pharm Sci 105: 884–890 [DOI] [PubMed] [Google Scholar]
- Yang J, Yang X, Zou H, Chu Y, Li M (2011a) Recovery of the immune balance between Th17 and regulatory T cells as a treatment for systemic lupus erythematosus. Rheumatology 50: 1366–1372 [DOI] [PubMed] [Google Scholar]
- Yang S, Wang X, Contino G, Liesa M, Sahin E, Ying H, Bause A, Li Y, Stomme JM, Dell'Antonio G et al (2011b) Pancreatic cancers require autophagy for tumor growth. Genes Dev 25: 717–729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang J, Yang X, Yang J, Li M (2018) Hydroxychloroquine inhibits the differentiation of Th17 cells in systemic lupus erythematosus. J Rheumatol 45: 818–826 [DOI] [PubMed] [Google Scholar]
- Yao X, Ye F, Zhang M, Cui C, Huang B, Niu P, Liu X, Zhao L, Dong E, Song C et al (2020) In vitro antiviral activity and projection of optimized dosing design of hydroxychloroquine for the treatment of severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2). Dis Clin Infect 10.1093/cid/ciaa237 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yavuz S, Asfuroğlu E, Bicakcigil M, Toker E (2011) Hydroxychloroquine improves dry eye symptoms of patients with primary Sjogren's syndrome. Rheumatol Int 31: 1045–1049 [DOI] [PubMed] [Google Scholar]
- Yi AK, Tuetken R, Redford T, Waldschmidt M, Kirsch J, Krieg AM (1998) CpG motifs in bacterial DNA activate leukocytes through the pH‐dependent generation of reactive oxygen species. J Immunol 160: 4755–4761 [PubMed] [Google Scholar]
- Yin F, Guo M, Yao S (2003) Kinetics of DNA binding with chloroquine phosphate using capacitive sensing method. Biosens Bioelectron 19: 297–304 [DOI] [PubMed] [Google Scholar]
- Yogasundaram H, Putko BN, Tien J, Paterson DI, Cujec B, Ringrose J, Oudit GY (2014) Hydroxychloroquine‐induced cardiomyopathy: case report, pathophysiology, diagnosis, and treatment. Can J Cardiol 30: 1706–1715 [DOI] [PubMed] [Google Scholar]
- Yoon YH, Cho KS, Hwang JJ, Lee SJ, Choi JA, Koh JY (2010) Induction of lysosomal dilatation, arrested autophagy, and cell death by chloroquine in cultured ARPE‐19 cells. Invest Ophthalmol Vis Sci 51: 6030–6037 [DOI] [PubMed] [Google Scholar]
- Yoon CH, Lee HJ, Lee EY, Lee EB, Lee WW, Kim MK, Wee WR (2016) Effect of hydroxychloroquine treatment on dry eyes in subjects with primary Sjögren's syndrome: a double‐blind randomized control study. J Korean Med Sci 31: 1127–1135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yusuf IH, Sharma S, Luqmani R, Downes SM (2017) Hydroxychloroquine retinopathy. Eye 31: 828–845 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Li Y, Li Y, Li R, Ma Y, Wang H, Wang Y (2015) Chloroquine inhibits MGC803 gastric cancer cell migration via the Toll‐like receptor 9/nuclear factor kappa B signaling pathway. Mol Med Rep 11: 1366–1371 [DOI] [PubMed] [Google Scholar]
- Zhang Y, Cao Y, Sun X, Feng Y, Du Y, Liu F, Yu C, Jin F (2017) Chloroquine (CQ) exerts anti‐breast cancer through modulating microenvironment and inducing apoptosis. Int Immunopharmacol 42: 100–107 [DOI] [PubMed] [Google Scholar]
- Zhang J, Chen L, Xiong F, Zhang S, Huang K, Zhang Z, Wang CY (2019) Autophagy in regulatory T cells: a double‐edged sword in disease settings. Mol Immunol 109: 43–50 [DOI] [PubMed] [Google Scholar]
- Zhou Q, McCracken MA, Strobl JS (2002) Control of mammary tumor cell growth in vitro by novel cell differentiation and apoptosis agents. Breast Cancer Res Treat 75: 107–117 [DOI] [PubMed] [Google Scholar]
- Ziegler HK, Unanue ER (1982) Decrease in macrophage antigen catabolism caused by ammonia and chloroquine is associated with inhibition of antigen presentation to T cells. Proc Natl Acad Sci USA 79: 175–178 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table EV1