

Abstract
Cancer is driven by genetic mutations in oncogenes and tumor suppressor genes and by cellular events that develop a misregulated molecular microenvironment in the growing tumor tissue. The tumor microenvironment is guided by the excessive action of specific cytokines including transforming growth factor‐β (TGF‐β), which normally controls embryonic development and the homeostasis of young or adult tissues. As a consequence of the genetic alterations generating a given tumor, TGF‐β can preserve its homeostatic function and attempt to limit neoplastic expansion, whereas, once the tumor has progressed to an aggressive stage, TGF‐β can synergize with various oncogenic stimuli to facilitate tumor invasiveness and metastasis. TGF‐β signaling mechanisms via Smad proteins, various ubiquitin ligases, and protein kinases are relatively well understood. Such mechanisms regulate the expression of genes encoding proteins or non‐coding RNAs. Among non‐coding RNAs, much has been understood regarding the regulation and function of microRNAs, whereas the role of long non‐coding RNAs is still emerging. This article emphasizes TGF‐β signaling mechanisms leading to the regulation of non‐coding genes, the function of such non‐coding RNAs as regulators of TGF‐β signaling, and the contribution of these mechanisms in specific hallmarks of cancer.
Keywords: non‐coding RNA, signal transduction, Smad, transcription, transforming growth factor‐β
Transforming growth factor‐β (TGF‐β) can synergize with various oncogenic stimuli to facilitate tumor invasiveness and metastasis. TGF‐β receptor signaling via Smads regulates expression of genes encoding for long non‐coding RNAs. Some of these non‐coding RNAs act as regulators of TGF‐β signaling and others mediate diverse processes of cancer biology.
Abbreviations
- EMT
epithelial‐mesenchymal transition
- lncRNA
long non‐coding RNA
- TGF‐β
transforming growth factor β
1. INTRODUCTION
Cancer represents a large group of diseases that affects many organs. Cancers are characterized in their onset by genetic mutations in key genes that subsequently unleash a cohort of cell biological processes; the various pathological processes ultimately lead to the growth of malignant tissue in the form of tumors and even further, but infrequently, to disseminating cells into metastases in nearby or distant organs. 1 , 2 Secreted growth factors control the communication between cells and the organization of tissues. For these reasons, growth factor genes, upon mutagenic alteration, can act as initiators of the malignant process (oncogenes), but also as functional mediators of the malignant evolution through various stages. 1 , 3 Since its discovery, transforming growth factor β (TGF‐β) has been linked functionally with and continues to provide new lessons on mechanisms that govern cancer development. 4 , 5 , 6 , 7
2. TGF‐β SIGNALING IN CANCER
TGF‐β is the prototype member of a large family of polypeptide growth factors that has exhibited evolutionary conservation in all animals since the emergence of multicellularity. 8 TGF‐β signaling pathways participate in developmental morphogenetic programs and contribute to young or adult organismic homeostasis; the growth factors of this family regulate differentiation, proliferation, and motility. 5 , 8 TGF‐β (encompassing 3 isoforms, TGF‐β1/2/3) is secreted from many cells via a well controlled mechanism that delivers a latent, inactive form of the growth factor, together with other proteins, to the extracellular matrix. 8 Upon activation, TGF‐β signals via its receptors, serine‐threonine and weak tyrosine kinase enzymes, known as type II (TGFβRII) and type I (TGFβRI) receptors. 8 When TGF‐β binds to TGFβRII, recruitment of TGFβRI is followed by trans‐phosphorylation of serine and threonine residues in TGFβRI by TGFβRII kinase, conformational activation of TGFβRI, which subsequently phosphorylates Smad2 and Smad3, members of the Smad family of signal transducers and latent transcription factors. 8 The phosphorylated Smad2 and Smad3 interact with Smad4 to generate trimeric complexes that associate directly with DNA and many transcription factors that mediate the regulation of target gene expression. 5 A negative feedback mechanism is mediated by the inhibitory Smad7. TGF‐β induces Smad7 expression, which inhibits signaling via direct interaction of Smad7 with TGFβRI, Smad complexes, and several ubiquitin ligases that ubiquitylate and degrade either the receptor, upon its internalization, or the active Smad complexes. 5 The TGF‐β receptor complex also recruits ubiquitin ligases that then, via ubiquitylation, activate protein kinases that lead to downstream engagement of the mitogen‐activated protein (MAP) kinases. 8 The same ubiquitylation‐dependent mechanism also controls a cleavage and translocation of the cytoplasmic, protein kinase domain of TGFβRI, to the nucleus for further signaling in association with Smad and other transcriptional cofactors. 8 The coordinated activity of Smads, phosphorylation inputs generated by TGF‐β‐mediated MAP kinase activation and nuclear TGFβRI intracellular domain mediate the diverse biological actions of TGF‐β. 5
Similar to its actions in adult homeostasis, TGF‐β signaling limits the development of hyperplastic, pre‐malignant lesions in many organs. 4 , 5 , 6 , 7 Once tumorigenesis has progressed, TGF‐β cooperates with diverse oncogenic pathways and facilitates the development of aggressive, less differentiated, and invasive tumors. 4 , 5 , 6 , 7 TGF‐β also facilitates cancer metastasis. 4 , 5 , 6 , 7 Homeostatic signaling fighting against hyperplastic growth is exemplified by the ability of TGF‐β to induce the expression of cyclin‐dependent kinase inhibitors, including CDKN1A (p21CIP1), CDKN1B (p27KIP1), CDKN2B (p15INK4B). 5 , 7 These cell cycle inhibitors stall the epithelial, endothelial, lymphocytic, and erythropoietic cell cycle in the early G1 phase. 5 , 7 In hepatocytes, prostate, and other epithelial cell types, TGF‐β can also induce apoptosis via coordinated signaling actions: (i) Smad‐mediated induction of pro‐apoptotic genes (Bim, DAPK); (ii) activation of MAP kinases and cytochrome c release from mitochondria, leading to pro‐caspase activation. 5 , 6 , 7 Furthermore, the TGF‐β receptors and the Smad genes can be mutated in various tumors. 9 Genetic alterations cause either complete loss of responses or preferential loss of the cytostatic and pro‐apoptotic responses to TGF‐β by malignant cells. 5
Once malignancy progresses, TGF‐β secretion by cancer cells, cancer‐associated fibroblasts, or in some cases even from immune cells, is abundantly observed. 4 In carcinomas, EMT is potently induced by TGF‐β and contributes to the invasive and pro‐metastatic phases of tumor development. 6 , 10 TGF‐β inhibits the proliferation and differentiation of B and T lymphocytes, causing a local immune suppression that promotes expansive tumor growth and invasiveness. 4 TGF‐β can indirectly stimulate neo‐angiogenesis that feeds the growing malignancy and facilitates invasiveness and metastatic dissemination. 5 , 6 These multi‐faceted effects of TGF‐β have, in recent years, stimulated several clinical trials. As a combinatorial treatment, together with more classical chemo‐ or radio‐therapy, TGF‐β pathway inhibitors have shown ability to limit expansion of various tumors. 4 , 6
3. LONG NON‐CODING RNAs
The majority of the biological activities of TGF‐β can be explained by regulation of expression of a large cohort of mRNAs and their encoded proteins. In recent years, attention has been given to the functional roles of non‐protein‐coding RNAs. Among the various non‐coding RNAs, much focus has been given to microRNAs (miRNAs); TGF‐β signaling regulates miRNA gene expression and miRNA maturation from precursor transcripts, whereas various miRNAs can regulate TGF‐β signaling in the context of cancer. 11 , 12 , 13 Here, we focus exclusively on long non‐coding RNAs (lncRNAs), whose regulation by TGF‐β signaling and functional participation in multiple responses to TGF‐β form an emerging field. 6 , 12
lncRNAs are structurally identical to mRNAs. They are transcribed by RNA polymerase II into 250 nt or longer RNAs; they have 5′ modified caps and poly‐adenylated tails at their 3′‐end, and localize in the nucleus, cytoplasm or both (Figure 1). 14 , 15 , 16 lncRNAs almost universally carry open reading frames, which are small, initiating with non‐optimal start codons, embedded in the lncRNA sequence far away from the 5′‐end and are often considered incapable of encoding polypeptides. 14 , 15 , 16 The non‐coding capacity of lncRNAs is bioinformatically attested and only rarely experimentally tested. Examples of lncRNAs encoding for functional polypeptides exist. The putative lncRNA LOC100507537 encodes for the 34 amino acid‐long peptide “dwarf open reading frame,” which associates with and activates the sarcoplasmic reticulum calcium pump sarco/endoplasmic reticulum Ca2+‐ATPase (SERCA) in cardiomyocytes, thus regulating heart muscle contraction. 17 In the context of cancer, the homeobox B cluster antisense RNA 3 (HOXB‐AS3) encodes a 53 amino acid‐long polypeptide; the polypeptide binds to arginine‐rich sequences in the hnRNP A1 splicing factor regulating alternative splicing of the pyruvate kinase M. 18 By inducing expression of pyruvate kinase isoform M2, the HOXB‐AS3 polypeptide facilitates manifestation of the oncogenic Warburg effect in colorectal cancer. 18
FIGURE 1.
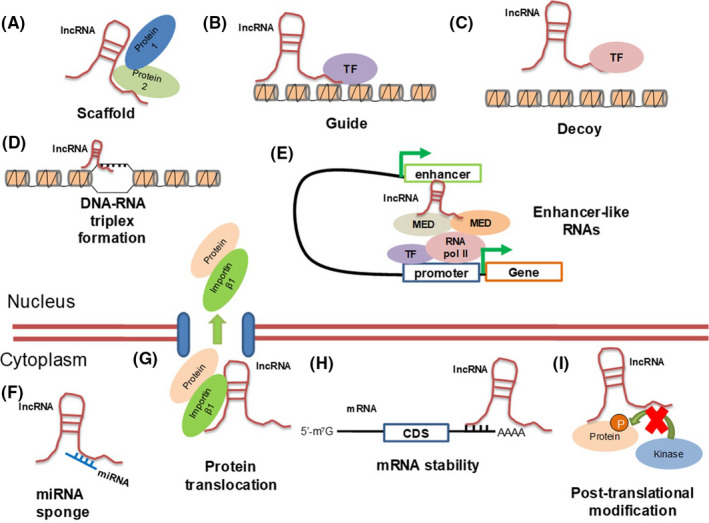
Molecular functions of lncRNAs. Nuclear lncRNAs act as scaffold (A), guide (B) or decoy (C) molecules. They also form triple helix formations by directly binding to DNA (D), and act as enhancer‐like RNAs, promoting gene transcription (E). Cytoplasmic lncRNAs function as sponges for miRNAs (F), facilitate nucleocytoplasmic protein translocation (G), affect mRNA stability (H) and interfere with post–translational modifications of proteins (I). CDS: coding sequence, MED: component of the mediator complex, RNA pol II: RNA polymerase II, TF: transcription factor
lncRNAs are classified based on the position of their gene relative to protein‐coding genes. Antisense lncRNAs are transcribed from the opposite DNA strand of a protein‐coding gene and partially overlap with mRNAs. 19 , 20 Intronic lncRNAs are completely embedded in the intron of a protein‐coding gene. 21 Divergent lncRNAs do not overlap with mRNAs but share promoter‐enhancer sequences with a protein‐coding gene and are transcribed in the opposite direction relative to the mRNA. 21 Enhancer RNAs (eRNAs) or ncRNA‐activating, are encoded by genes that overlap characterized enhancer sequences and regulate expression of the genes that are controlled by the enhancer. 22 , 23 Finally, intergenic lncRNAs map as independent genes far away from protein‐coding genes. 21
Similar to mRNAs, the biological functions of lncRNAs permeate all essential cell biological processes, and their actions are often linked to cancer development. These functions range from the control of stemness and differentiation, including genomic imprinting and the mechanism of X chromosome inactivation, to immunity and programmed cell death. 14 , 15 , 16 , 24 Mechanistically (Figure 1), lncRNAs regulate gene expression by acting as scaffolds, guides, or decoys or by base‐pairing with DNA, through formation of triple helices. 16 lncRNAs associate with nuclear proteins and affect nucleosome remodeling, 21 including histone modifications catalyzed by protein methyltransferases, such as the polycomb repressor complex 2 (PRC2). 16 , 25 lncRNAs can regulate mRNA splicing, stability, or translation. 21 A widely established function of lncRNAs, especially when they are located in the cytoplasm, is the “sponge” or competing endogenous (ce) RNA function; this indicates their ability to base‐pair with miRNAs (Figure 1), and thus shield the action of the miRNAs toward target mRNAs. 26 Most of these molecular mechanisms of action have been demonstrated in the context of cancer cell biology.
4. lncRNAs ACT AS EFFECTORS OF TGF‐β SIGNALING
The list of lncRNAs, described as effectors of TGF‐β signaling, is constantly growing (Figure 2). Table 1 summarizes TGF‐β‐regulated lncRNAs and their roles in different cancer types. Furthermore, lncRNAs acting as effectors of TGF‐β signaling have been reported in a plethora of different cancer types. One of the first lncRNAs, demonstrated to be modulated by TGF‐β, is the lncRNA‐activated by TGF‐β (lncRNA‐ATB) in hepatocellular carcinoma (HCC). 27 TGF‐β upregulates lncRNA‐ATB in order to favor EMT and establish a pro‐metastatic program. lncRNA‐ATB acts as a sponge for the epithelial‐specific miR‐200. 27 miR‐200 was previously established as a negative regulator of the EMT transcription factors ZEB1/2 and, accordingly, lncRNA‐ATB acts by enhancing ZEB1/2 expression. Moreover, lncRNA‐ATB stabilizes interleukin‐11 mRNA, leading to increased cytokine signaling mediated by STAT3, which potentiates tumor colonization in secondary tissues to ensure efficient metastasis. 27 Similar to HCC, lncRNA‐ATB is induced by TGF‐β in MCF7 breast cancer cells and was established as a marker of poor prognosis in breast cancer; lncRNA‐ATB promotes EMT by sponging miR‐200 and thus, upregulating Twist1 expression, the latter being another transcription factor of the EMT program. 28 In intrahepatic cholangiocarcinoma, the TGF‐β‐induced long noncoding RNA (TLINC) boosts a pro‐migratory phenotype and positively regulates interleukin‐8, reinforcing a pro‐inflammatory tumor microenvironment. 29 In pancreatic ductal adenocarcinoma (PDAC), TGF‐β induces the mir‐100‐let‐7a‐2‐mir‐125b‐1 cluster host gene (MIR100HG), a lncRNA that gives rise to mir‐100, let‐7a‐2, and mir‐125b‐1 miRNAs. miR‐100 and miR‐125b promote PDAC progression and EMT, by downregulating p53 and apoptotic pathways and upregulating the pro‐survival phosphatidylinositol 3´‐kinase/Akt signaling pathway. 30 In colorectal cancer, the taurine up‐regulated gene 1 (TUG1) lncRNA is a mediator of TGF‐β‐induced EMT in vitro and metastasis in vivo; TUG1 is enhanced in response to TGF‐β, in order to increase Twist1 expression, resulting in enhanced migration, invasion, and lung metastasis. 31
FIGURE 2.
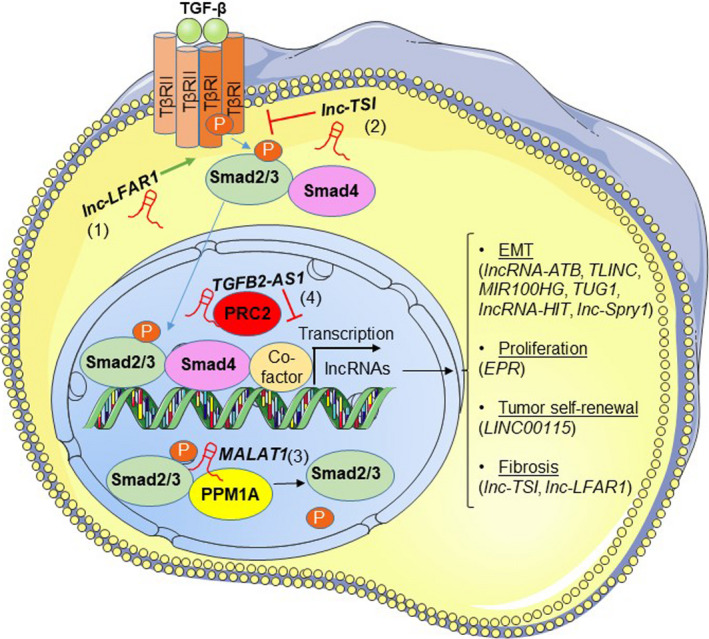
TGF‐β signaling regulates lncRNA expression and mechanisms of TGF‐β signaling control by lncRNAs. TGF‐β signals through Smad2/3/4 proteins to regulate the expression of lncRNAs, acting as effector molecules that mediate the physiological responses of the signaling pathway (listed on the right hand‐side). lncRNAs target TGF‐β signaling at different stages of the pathway. At the level of Smad2/3 activation by TGFβRI, lnc‐LFAR1 (1) enhances the association of Smad2/3 with TGFβRI, leading to increased activation of the pathway. lnc‐TSI (2) blocks the interaction of Smad3 with TGFβRI, attenuating the pathway. In the nucleus, MALAT1 (3) shifts the Smad2/3 levels toward the non‐phosphorylated state, by promoting the association of the phosphatase PPM1A to Smads and inhibiting TGF‐β‐mediated responses. TGFB2‐AS1 (4) epigenetically silences TGF‐β‐target genes by recruiting the PRC2 repressive complex to their promoters
TABLE 1.
TGF‐β‐regulated lncRNAs
| LncRNA | Type of regulation | Function | Mechanism of action | Cancer type/ cell line | Ref. |
|---|---|---|---|---|---|
| TUG1 | Up | Induces EMT in vitro; metastasis in vivo | Enhances Twist1 | Colorectal cancer | 31 |
| LINC00273 | Up | Promotes invasion and metastasis | Activates ZEB1 via sponging mir200a‐3p | A549 adenocarcinoma cells | 51 |
| LINC00115 | Up | Promotes cell self‐renewal | Upregulates ZEB1 and ZNF596, via sponging miR‐200 | Glioma stem‐like cells | 35 |
| EPR | Up (early) Down (late) | Inhibits cell proliferation | Positively regulates Cdkn1a | NMuMG breast epithelial cells | 34 |
| MIR155HG | Up | Promotes EMT | Regulates the miR‐155‐5p/ SOX10 axis | Laryngeal squamous cell carcinoma | 52 |
| MACC1‐AS1 | Up | Fatty acid oxidation‐dependent stemness and chemoresistance | De‐represses stemness and FAO genes, via sponging miR‐145‐5p | Gastric cancer | 53 |
| HCP5 | Up | Promotes EMT | Upregulates Snail and Slug by sponging miR‐203 | Lung adenocarcinoma | 54 |
| PTAF | Up | Promotes EMT and invasion | Enhances SNAI2 by targeting miR‐25 | Ovarian cancer | 55 |
| MIR100HG | Up | Hosts pro‐tumorigenic miRNAs | miR‐100 and miR‐125b downregulate p53 and apoptotic pathways and activate the PI3K pathway | PDAC | 30 |
| TLINC | Up | Promotes cell migration and pro‐inflammatory tumor microenvironment | Positively regulates pro‐inflammatory cytokines | Intrahepatic cholangiocarcinoma | 29 |
| MEG8 | Up | Induces EMT | Upregulates SNAI1 and SNAI2, by epigenetically suppressing miR‐34a and miR‐203 | A549, LC‐2/ad, Panc1 cells | 56 |
| UCA1 | Up | Promotes cell proliferation | Upregulates HXK2 | HCC | 57 |
| Up | Promotes EMT | Enhances Slug by targeting miR‐1 and miR‐203a | Glioma | 58 | |
| Up | Promotes EMT | Unknown | Gastric cancer | 59 | |
| NKILA | Up | Inhibits cell migration and invasion | Suppresses MMP14 by inhibiting the NF‐κB pathway | Esophageal squamous cell carcinoma | 60 |
| Up | Inhibits EMT | Blocks the NF‐κB pathway | MCF7 breast cancer cells | 61 | |
| LINP1 | Down | Inhibits EMT | Enhances CDH1 and represses mesenchymal genes (mechanism unknown) | Lung cancer | 62 |
| lncRNA‐ATB | Up | Promotes EMT | Upregulates Twist1 by sponging miR‐200 | MCF7 breast cancer cells | 28 |
| Up | Promotes EMT in vitro and metastasis in vivo | Enhances ZEB1/2, by sponging miR‐200 and stabilizes IL‐11 mRNA | HCC | 27 | |
| lnc‐MMP2‐2 | Up | Regulates cell migration and invasion | Promotes MMP2 expression | A549 lung adenocarcinoma exosomes | 63 |
| TBILA | Up | Promotes tumor progression in vitro and in vivo | Enhances RhoA and S100A7‐JAB1 pathway activation | Non‐small cell lung cancer | 64 |
| H19 | Down | Increases tumorigenic potential in vivo | Unknown | Tumor‐initiating hepatocytes | 65 |
| Up | Enhances cell invasion in vitro and metastasis in vivo | Upregulates Slug and inhibits CDH1 via miR‐675 | Hep3B HCC cells | 66 | |
| has2as | Up | Promotes EMT and cancer stemness | Induces has2, by facilitating Smad2/3 binding to its promoter | NMuMG breast epithelial cells | 67 |
| EPB41L4A‐AS2 | Down | Inhibits cell migration and invasion | Inhibits TGFBR1 expression | Head and neck squamous cell carcinoma | 68 |
| lnc‐Spry1 | Down | Suppresses EMT | Alternative splicing of FGFRs, via binding to U2AF65 | NMuMG breast epithelial cells | 33 |
| MEG3 | Up | Induces EMT | Represses CDH1 and miR‐200 by facilitating recruitment of JARID2 and EZH2 on their promoters | A549, LC‐2/ad cells | 69 |
| LINC01186 | Down | Inhibits EMT | Suppresses mesenchymal markers and induces CDH1, (mechanism unknown) | A549 lung adenocarcinoma cells | 70 |
| lncRNA‐LET | Down | Represses cancer cell stemness | Decreases NF90 stability leading to miR‐145 upregulation | Urinary bladder cancer | 71 |
| linc00673 | Up | Induces EMT | Upregulates ZEB1, by sponging miR‐150‐5p | Non‐small cell lung cancer | 72 |
| LINC01133 | Down | Inhibits EMT and metastasis | Blocks SRSF6 function | Colorectal cancer | 73 |
| lncRNA‐HIT | Up | Enhances EMT, migration, invasion | Represses CDH1 | NMuMG breast epithelial cells | 32 |
| MALAT1 | Up | Induces EMT | Represses CDH1, via binding to SUZ12 | Bladder cancer | 74 |
| lncRNA‐Smad7 | Up | Inhibits apoptosis | Unknown | NMuMG, JygMC(A) breast cancer cells | 75 |
Using a genome‐wide screen for identifying TGF‐β‐regulated lncRNAs in NMuMG mouse mammary epithelial cells, the homeobox A (HOXA) transcript induced by TGF‐β (lncRNA‐HIT) was shown to enhance EMT, migration, and invasion, by specifically inhibiting E‐cadherin expression. 32 TGF‐β signaling can additionally negatively regulate repressors of the EMT process, in order to elicit its pro‐tumorigenic role. For example, TGF‐β inhibits a lncRNA located in close proximity to the Spry1 gene, designated as lnc‐Spry1, in NMuMG cells. 33 lnc‐Spry1 interacts with the splicing factor U2AF65 and suppresses EMT, by affecting the alternative splicing of fibroblast growth factor receptors. 33 In NMuMG cells, TGF‐β initially upregulates and, upon sustained signaling, it downregulates the epithelial cell program regulator (EPR), an epithelial lncRNA that attenuates cell proliferation, by positively regulating the cell cycle inhibitor Cdkn1a, both transcriptionally and post‐transcriptionally. 34 At the transcriptional level, EPR directly binds to the Cdkn1a promoter and interacts with Smad3 during early TGF‐β signaling, thereby activating Cdkn1a transcription. 34 Upon sustained TGF‐β signaling Cdkn1a levels return to basal, a response that coincides with the delayed EPR downregulation. At the post–transcriptional level, EPR associates with the RNA‐binding protein KHSRP (KH‐type splicing regulatory protein) and prevents its binding to Cdkn1a mRNA, thereby increasing Cdkn1a stability. 34
TGF‐β regulates lncRNA expression not only in carcinomas but also in tumors of diverse tissue origin. For example, LINC00115 is overexpressed in glioblastoma and is upregulated by TGF‐β in glioma stem‐like cells, in order to facilitate their self‐renewal. 35 LINC00115 interferes with the binding of miR‐200 to its target mRNAs ZEB1 and ZNF596 (zinc finger protein 596), leading to increased expression of these proteins and potentiation of downstream pro‐tumorigenic signals that elicit tumor growth. 35 Table 1 presents additional lncRNAs which are not discussed here in the interest of space.
5. lncRNAs ACT AS REGULATORS OF TGF‐β SIGNALING
In addition to being effectors of TGF‐β signaling, lncRNAs modulate several components of the pathway, thereby affecting the magnitude of its response, during tumor progression (Figure 2). Several lncRNAs can regulate TGF‐β signaling in a wide range of cancers (Table 2). In HCC cells, metastasis‐associated lung adenocarcinoma transcript 1 (MALAT1) alters the balance between phosphorylated (p‐) and de‐phosphorylated Smad2 and Smad3 levels. 36 MALAT1 interacts with p‐Smad2 and p‐Smad3, through the protein known as SET domain containing 2 (SETD2), which serves as a scaffold that facilitates complex formation between the phosphatase PPM1A (protein phosphatase, Mg2+/Mn2+ dependent 1A) and p‐Smad2/3. Thus, MALAT1 promotes the termination of TGF‐β signaling, by inducing the PPM1A‐mediated de‐phosphorylation of activated p‐Smad2/3. 36 In colorectal cancer, the cancer susceptibility candidate 9 (CASC9) is a lncRNA that predicts poor survival for patients. 37 The pro‐tumorigenic function of CASC9 is due to the increased stabilization of TGFB2 levels, which lead to active TGF‐β2 signaling and enhanced p‐Smad3 levels. 37 The positive contribution of CASC9 to TGF‐β signaling depends on its binding to the protein cleavage and polyadenylation specific factor 3 (CPSF3), an mRNA‐processing factor, which is capable of directly interacting with TGFB2 mRNA. 37 Liver fibrosis‐associated lncRNA 1 (lnc‐LFAR1) potentiates TGF‐β signaling by enhancing TGFB1, Smad2, and Smad4 mRNA levels in the intrahepatic cholangiocarcinoma cell line QBC939. 38 Moreover, lnc‐LFAR1 exerts pro‐EMT functions, by enhancing vimentin and downregulating E‐cadherin protein levels and reinforces migration and invasion of QBC939 cells. 38 In colorectal cancer, the small nucleolar RNA host gene 6 (SNHG6) activates the TGF‐β pathway by reducing UPF1 (UPF1 RNA helicase and ATPase), a regulator of Smad7, leading to reduced Smad7 expression and, therefore, increased p‐Smad2/3 levels. 39 Thus, SNHG6 promotes cell proliferation, migration, and invasion in vitro and colorectal tumor growth in vivo. 39 In HCC, the nuclear enriched abundant transcript 1 (NEAT1) acts as a ceRNA for miR‐139‐5p, thereby protecting TGFB1 mRNA from miR‐139‐5p‐induced degradation. 40 Thus, NEAT1 is an activator of TGF‐β signaling and promotes HCC growth. 40 In ovarian carcinoma, the LINK‐A is frequently overexpressed and positively correlated to the TGF‐β1 protein levels. 41 Ectopic expression of LINK‐A leads to enhanced TGF‐β1 expression, as well as enhanced migratory and invasive behavior of ovarian cancer cells. 41 In lung adenocarcinoma cells, the NORAD (non‐coding RNA activated by DNA damage) is predominantly cytoplasmic and promotes the association between Smad3 and importin‐β1, thereby facilitating the TGF‐β‐induced nuclear translocation of Smad3. 42 NORAD positively regulates the activity of Smad‐responsive luciferase reporter genes, as well as the expression of TGF‐β‐target and EMT‐related genes, such as SERPINE1, SNAI1, and FN1. In addition, by activating the TGF‐β pathway, NORAD promotes the migration of A549 lung adenocarcinoma cells, in response to TGF‐β. 42 In invasive pituitary tumors, the oncogenic lnc‐SNHG1 upregulates TGFBR2, by preventing receptor degradation by the miR‐302/372/373/520 cluster, leading to enhanced Smad3 nuclear accumulation and increased migration and invasion, accompanied by a pro‐EMT phenotype of pituitary tumor cells. 43 In breast cancer cells, maternally expressed 3 (MEG3), in co‐operation with the co‐repressor complex PRC2, promotes epigenetic silencing of members of the TGF‐β pathway, such as TGFB2, TGFBR1, and Smad2, via formation of RNA‐DNA triple helical structures in GA‐rich, distal regulatory elements of these genes. 44 These cases clearly illustrate that essentially every component of the TGF‐β signaling pathway can be targeted for regulation by lncRNAs (Figure 2).
TABLE 2.
lncRNAs that modulate TGF‐β signaling
| lncRNA | Type of regulation | Mechanism of action | Cancer type/cell line | Ref. |
|---|---|---|---|---|
| MALAT1 | Negative | Promotes de‐phosphorylation of Smad2/3 by PPM1A | Hep3B, PLC/PRF/5, SK‐Hep1 HCC cells | 36 |
| CASC9 | Positive | Increased stabilization of TGFB2 mRNA | Colorectal cancer | 37 |
| HAND2‐AS1 | Negative | Downregulates TGF‐β1 | Non‐small cell lung cancer | 76 |
| lnc‐LFAR1 | Positive | Enhances TGFB1, Smad2, and Smad4 mRNAs | Intrahepatic cholangiocarcinoma cells (QBC939) | 38 |
| AWPPH | Positive | Upregulates TGF‐β1 | Non‐small cell lung cancer cells (H1581, H1993) | 77 |
| POU3F3 | Positive | Upregulates TGF‐β1 | Nasopharyngeal carcinoma cells (HTB‐43, C666‐1) | 78 |
| SNHG6 | Positive | Reduces Smad7 expression | Colorectal cancer cells (RKO) | 79 |
| SBF2‐AS1 | Positive | Enhances TGFBR1 through sponging miR‐140‐5p | HCC | 80 |
| NORAD | Positive | Facilitates Smad3 nuclear transport | A549 lung adenocarcinoma cells | 42 |
| LINK‐A | Positive | Enhances TGF‐β1 expression | Ovarian carcinoma | 41 |
| lnc‐SNHG1 | Positive | Upregulates TGFBR2, by sponging the miR‐302/372/373/520 cluster | Pituitary tumor cells (GH1, RC‐4B/C) | 43 |
| NEAT1 | Positive | Prevents TGFB1 mRNA degradation by sponging miR‐139‐5p | HCC | 40 |
| UCA1 | Positive | Positively regulates TGF‐β | Multiple myeloma | 81 |
| LINC00978 | Positive | Enhances TGF‐β and p‐Smad2 levels | Gastric cancer cells (MGC‐803, SGC‐7901) | 82 |
| SPRY4‐IT1 | Positive | Positively regulates TGF‐β1 and p‐Smad2/3 levels | Thyroid squamous cell carcinoma (SW579), papillary thyroid carcinoma (TPC‐1) cells | 83 |
| EPB41L4AAS2 | Negative | Suppresses TGFBR1 | Head and neck squamous cell carcinoma | 68 |
| CCAT2 | Positive | Positively regulates TGF‐β and Smad2 | Breast cancer | 84 |
| XIST | Positive | Upregulates TGF‐β1 via targeting miR‐185 | Gastric cancer | 85 |
| TUG1 | Positive | Promotes p‐Smad2/3 | PDAC | 86 |
| BX357664 | Negative | Suppresses TGF‐β1 | Renal cell carcinoma | 87 |
| ANRIL | Negative | Inhibits TGF‐β1 | Thyroid cancer | 88 |
| Negative | Inhibits TGF‐β1 | Esophageal squamous cell carcinoma | 89 | |
| MEG3 | Negative | Epigenetically silences TGFB2, TGFBR1, and Smad2, via RNA‐DNA triplex formation | Breast cancer | 44 |
6. lncRNAs FORM FEEDBACK LOOPS WITH TGF‐β SIGNALING
Some of the lncRNAs whose expression is regulated by the TGF‐β pathway, as described above, potentiate or diminish the responses of the pathway itself. We categorize these lncRNAs into 2 subclasses: first, lncRNAs that are transcriptionally upregulated by TGF‐β, which then enhance TGF‐β signaling output, forming positive feedback loops; second, TGF‐β‐induced lncRNAs, with inhibitory roles on TGF‐β responses, thereby belonging to negative feedback loops. Examples of lncRNAs that form positive feedback loops with TGF‐β are the lncRNAs PCAT7, ELIT1, HOTAIR, lincRNA‐p21, MALAT1 and lncRNA‐ATB (Table 3). In prostate cancer, PCAT7 (prostate cancer‐associated transcript‐7) is upregulated by TGF‐β via the transcriptional complex of Smad3 with Sp1 and then positively regulates TGF‐β signaling by sponging miR‐324‐5p, leading to enhanced TGFBR1 expression, as TGFBR1 is downregulated by miR‐324‐5p. 45 In endothelial progenitor cells, the TGF‐β‐induced MALAT1 described earlier, is required for the induction of endothelial‐to‐mesenchymal transition, a process similar to the EMT that has been implicated in the dissemination of tumor cells to metastatic sites. Mechanistically, MALAT1 binds to the tumor suppressor miR‐145 and sequesters it away from its target mRNAs TGFBR2 and Smad3, resulting in increased TGF‐β activation. 46 In hepatocytes, lincRNA‐p21 is involved in a positive feedback loop, whereby TGF‐β induces its expression, in order to strengthen the magnitude of the pathway, by sponging miR‐30, leading to increased KLF11 levels, as miR‐30 downregulates KLF11; the transcription factor KLF11 represses Smad7, and thus enhances TGF‐β signaling, which promotes liver fibrosis. 47 In keloid fibroblasts, the TGF‐β‐induced lncRNA‐ATB described earlier, facilitates TGF‐β‐dependent responses, by acting as a ceRNA for miR‐200c, leading to ZNF217 upregulation and increased secretion of TGFβ2. 48
TABLE 3.
List of lncRNAs that form feedback loops with TGF‐β signaling
| lncRNA | Feedback loop | Mechanism of action | Cancer type/cell line | Ref. |
|---|---|---|---|---|
| PCAT7 | Positive | Upregulation of TGFBR1 via sponging miR‐324‐5p | Prostate cancer | 45 |
| TGFB2‐AS1 | Negative | Epigenetic silencing of TGF‐β‐target gene promoters via PRC2 | HaCaT keratinocytes, A549 lung adenocarcinoma cells | 49 |
| ELIT‐1 | Positive | Binds to Smad3 and facilitates its recruitment to promoters of EMT‐related genes | Huh7 HCC, A549 lung adenocarcinoma cells | 90 |
| lnc‐TSI | Negative | Blocking the interaction of Smad3 with TGFβRI | Renal tubular epithelial cells | 50 |
| lincRNA‐p21 | Positive | Interaction with miR‐30 leading to KLF11‐mediated suppression of Smad7 | AML12 hepatocytes | 47 |
| MALAT1 | Positive | Upregulation of TGFBR2 and Smad3 via sponging miR‐145 | Endothelial progenitor cells | 46 |
| lncRNA‐ATB | Positive | Increased secretion of TGFβ2 via sponging miR‐200c and induction of ZNF217 | Keloid fibroblasts | 48 |
Conversely, TGFB2‐AS1 and lnc‐TSI participate in negative feedback loops with TGF‐β signaling (Table 3). TGF‐β induces the expression of TGFB2 antisense RNA 1 (TGFB2‐AS1) in human immortalized keratinocytes and lung adenocarcinoma cells. TGFB2‐AS1, in turn inhibits Smad‐mediated transcriptional responses, via interaction with the PRC2 complex, leading to epigenetic silencing of TGF‐β target genes. 49 During renal fibrosis, the kidney‐specific TGF‐β/Smad3‐interacting long noncoding RNA (lnc‐TSI) is upregulated by TGF‐β and forms a negative loop, by binding to the MH2 domain of Smad3, thereby blocking the association of Smad3 with TGFβRI and inhibiting receptor signaling. 50 This mechanism seems to not involve the function of Smad7, and results in lower TGF‐β signaling and therefore diminished TGF‐β‐induced renal fibrogenesis. 50 Although some of these examples do not stem from studies of cancer biology, they are useful as they illustrate the importance of feedback control of the TGF‐β signaling pathway, a mechanism whereby previously well established proteins are now demonstrated to cooperate with lncRNAs in order to elicit their full action.
7. CONCLUDING REMARKS
The large spectrum of biological actions engaging the TGF‐β signaling pathway during cancer development has necessitated the elucidation of many target genes of this pathway, and their functions. Whereas the first 35 y of TGF‐β signaling research focused on protein‐coding genes, the past 5 y have demonstrated the important function of lncRNAs. Most of the studied lncRNAs act either as regulators of chromatin modifications and transcriptional control or as sponges that limit the abundance of miRNAs. We anticipate the elucidation of completely new mechanisms of action of lncRNAs downstream of TGF‐β in cancer. Equally interesting is the large number of lncRNAs that regulate specific steps of TGF‐β signaling. Whereas TGF‐β ligand expression is a frequent target for regulation by lncRNAs, examples of very intricate mechanisms, such as regulation of Smad phosphorylation or Smad translocation to the nucleus, have been described to engage lncRNAs. In cancer, differentially expressed oncogenic lncRNAs that modulate TGF‐β signaling could serve as biomarkers to stratify patients that may benefit from anti‐TGF‐β‐based therapies. Placing such lncRNAs together with protein‐based mechanisms into the biology of specific tumors is a challenging task. Completion of this task promises a more coherent understanding of the mistakes made as cancer cells aim to survive and spread their biological potential in multiple organs of the afflicted patients.
DISCLOSURE
The authors declare no conflict of interest for this article.
ACKNOWLEDGMENTS
We express our appreciation for the support of Dr. Carl‐Henrik Heldin through the years of our research program. We thank past and present members of our group for their scientific contributions. Due to space limitations not all relevant publications are discussed and we apologize to their authors. We acknowledge funding by Barncancerfonden (grant number 2018‐0091), Cancerfonden (grant number CAN 2018/469), Ludwig Institute for Cancer Research (grant number Uppsala Branch), and Vetenskapsrådet (grant number 2018‐02757).
Papoutsoglou P, Moustakas A. Long non‐coding RNAs and TGF‐β signaling in cancer. Cancer Sci. 2020;111:2672–2681. 10.1111/cas.14509
REFERENCES
- 1. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646‐674. [DOI] [PubMed] [Google Scholar]
- 2. Massagué J, Obenauf AC. Metastatic colonization by circulating tumour cells. Nature. 2016;529:298‐306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Heldin C‐H, Lu B, Evans R, Gutkind JS. Signals and receptors. Cold Spring Harb Perspect Biol. 2016;8:a005900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Batlle E, Massagué J. Transforming Growth Factor‐β Signaling in Immunity and Cancer. Immunity. 2019;50:924‐940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. David CJ, Massagué J. Contextual determinants of TGFβ action in development, immunity and cancer. Nat Rev Mol Cell Biol. 2018;19:419‐435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Hao Y, Baker D, ten Dijke P. TGF‐β‐Mediated Epithelial‐Mesenchymal Transition and Cancer Metastasis. Int J Mol Sci. 2019;20:2767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Pardali K, Moustakas A. Actions of TGF‐β as tumor suppressor and pro‐metastatic factor in human cancer. Biochim Biophys Acta. 2007;1775:21‐62. [DOI] [PubMed] [Google Scholar]
- 8. Tzavlaki K, Moustakas A. TGF‐β signaling. Biomolecules. 2020;10:E487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Korkut A, Zaidi S, Kanchi RS, et al. A pan‐cancer analysis reveals high‐frequency genetic alterations in mediators of signaling by the TGF‐β superfamily. Cell Syst. 2018;7:422‐437.e427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Derynck R, Weinberg RA. EMT and cancer: more than meets the eye. Dev Cell. 2019;49:313‐316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Blahna MT, Hata A. Regulation of miRNA biogenesis as an integrated component of growth factor signaling. Curr Opin Cell Biol. 2013;25:233‐240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Grelet S, McShane A, Geslain R, Howe PH. Pleiotropic roles of non‐coding RNAs in TGF‐β‐mediated epithelial‐mesenchymal transition and their functions in tumor progression. Cancers. 2017;9:E75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Janakiraman H, House RP, Gangaraju VK, et al. The long (lncRNA) and short (miRNA) of It: TGFβ‐mediated control of RNA‐binding proteins and noncoding RNAs. Mol Cancer Res. 2018;16:567‐579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Hu W, Alvarez‐Dominguez JR, Lodish HF. Regulation of mammalian cell differentiation by long non‐coding RNAs. EMBO Rep. 2012;13:971‐983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Tordonato C, Di Fiore PP, Nicassio F. The role of non‐coding RNAs in the regulation of stem cells and progenitors in the normal mammary gland and in breast tumors. Front Genet. 2015;6:72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Akhade VS, Pal D, Kanduri C. Long noncoding RNA: genome organization and mechanism of action. Adv Exp Med Biol. 2017;1008:47‐74. [DOI] [PubMed] [Google Scholar]
- 17. Nelson BR, Makarewich CA, Anderson DM, et al. A peptide encoded by a transcript annotated as long noncoding RNA enhances SERCA activity in muscle. Science. 2016;351:271‐275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Huang J‐Z, Chen M, Chen DE, et al. A peptide encoded by a putative lncRNA HOXB‐AS3 suppresses colon cancer growth. Mol Cell. 2017;68:171‐184.e176. [DOI] [PubMed] [Google Scholar]
- 19. Pelechano V, Steinmetz LM. Gene regulation by antisense transcription. Nat Rev Genet. 2013;14:880‐893. [DOI] [PubMed] [Google Scholar]
- 20. Villegas VE, Zaphiropoulos PG. Neighboring gene regulation by antisense long non‐coding RNAs. Int J Mol Sci. 2015;16:3251‐3266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Rinn JL, Chang HY. Genome regulation by long noncoding RNAs. Annu Rev Biochem. 2012;81:145‐166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Lai F, Orom UA, Cesaroni M, et al. Activating RNAs associate with Mediator to enhance chromatin architecture and transcription. Nature. 2013;494:497‐501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Chen H, Du G, Song X, Li L. Non‐coding transcripts from enhancers: new insights into enhancer activity and gene expression regulation. Genomics Proteomics Bioinformatics. 2017;15:201‐207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Chen YG, Satpathy AT, Chang HY. Gene regulation in the immune system by long noncoding RNAs. Nat Immunol. 2017;18:962‐972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Davidovich C, Cech TR. The recruitment of chromatin modifiers by long noncoding RNAs: lessons from PRC2. RNA. 2015;21:2007‐2022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Salmena L, Poliseno L, Tay Y, et al. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146:353‐358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Yuan JH, Yang F, Wang F, et al. A long noncoding RNA activated by TGF‐β promotes the invasion‐metastasis cascade in hepatocellular carcinoma. Cancer Cell. 2014;25:666‐681. [DOI] [PubMed] [Google Scholar]
- 28. Li R‐H, Chen M, Liu J, et al. Long noncoding RNA ATB promotes the epithelial‐mesenchymal transition by upregulating the miR‐200c/Twist1 axe and predicts poor prognosis in breast cancer. Cell Death Dis. 2018;9:1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Merdrignac A, Angenard G, Allain C, et al. A novel transforming growth factor β‐induced long noncoding RNA promotes an inflammatory microenvironment in human intrahepatic cholangiocarcinoma. Hepatol Commun. 2018;2:254‐269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Ottaviani S, Stebbing J, Frampton AE, et al. TGF‐β induces miR‐100 and miR‐125b but blocks let‐7a through LIN28B controlling PDAC progression. Nat Commun. 2018;9:1845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Shen X, Hu X, Mao J, et al. The long noncoding RNA TUG1 is required for TGF‐β/TWIST1/EMT‐mediated metastasis in colorectal cancer cells. Cell Death Dis. 2020;11:65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Richards EJ, Zhang G, Li ZP, et al. Long non‐coding RNAs (LncRNA) regulated by transforming growth factor (TGF) β: LncRNA‐hit‐mediated TGFβ‐induced epithelial to mesenchymal transition in mammary epithelia. J Biol Chem. 2015;290:6857‐6867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Rodriguez‐Mateo C, Torres B, Gutierrez G, et al. Downregulation of Lnc‐Spry1 mediates TGF‐β‐induced epithelial‐mesenchymal transition by transcriptional and posttranscriptional regulatory mechanisms. Cell Death Differ. 2017;24:785‐797.28186499 [Google Scholar]
- 34. Rossi M, Bucci G, Rizzotto D, et al. LncRNA EPR controls epithelial proliferation by coordinating Cdkn1a transcription and mRNA decay response to TGF‐β. Nat Commun. 2019;10:1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Tang J, Yu B, Li Y, et al. TGF‐β‐activated lncRNA LINC00115 is a critical regulator of glioma stem‐like cell tumorigenicity. EMBO Rep. 2019;20:e48170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Zhang J, Han C, Song K, et al. The long‐noncoding RNA MALAT1 regulates TGF‐β/Smad signaling through formation of a lncRNA‐protein complex with Smads, SETD2 and PPM1A in hepatic cells. PLoS One. 2020;15:e0228160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Luo K, Geng J, Zhang Q, et al. LncRNA CASC9 interacts with CPSF3 to regulate TGF‐β signaling in colorectal cancer. J Exp Clin Cancer Res. 2019;38:249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Chen C, Li H, Wang X, et al. Lnc‐LFAR1 affects intrahepatic cholangiocarcinoma proliferation, invasion, and EMT by regulating the TGFβ/Smad signaling pathway. Int J Clin Exp Pathol. 2019;12:2455‐2461. [PMC free article] [PubMed] [Google Scholar]
- 39. Wang X, Lai Q, He J, et al. LncRNA SNHG6 promotes proliferation, invasion and migration in colorectal cancer cells by activating TGF‐β/Smad signaling pathway via targeting UPF1 and inducing EMT via regulation of ZEB1. Int J Med Sci. 2019;16:51‐59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Tu J, Zhao Z, Xu M, et al. NEAT1 upregulates TGF‐β1 to induce hepatocellular carcinoma progression by sponging hsa‐mir‐139‐5p. J Cell Physiol. 2018;233:8578‐8587. [DOI] [PubMed] [Google Scholar]
- 41. Ma J, Xue M. LINK‐A lncRNA promotes migration and invasion of ovarian carcinoma cells by activating TGF‐β pathway. Biosci Rep. 2018;38:BSR20180936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Kawasaki N, Miwa T, Hokari S, et al. Long noncoding RNA NORAD regulates transforming growth factor‐β signaling and epithelial‐to‐mesenchymal transition‐like phenotype. Cancer Sci. 2018;109:2211‐2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Wang H, Wang G, Gao Y, et al. Lnc‐SNHG1 activates the TGFBR2/SMAD3 and RAB11A/Wnt/β‐catenin pathway by sponging MiR‐302/372/373/520 in invasive pituitary tumors. Cell Physiol Biochem. 2018;48:1291‐1303. [DOI] [PubMed] [Google Scholar]
- 44. Mondal T, Subhash S, Vaid R, et al. MEG3 long noncoding RNA regulates the TGF‐β pathway genes through formation of RNA‐DNA triplex structures. Nat Commun. 2015;6:7743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Lang C, Dai Y, Wu Z, et al. SMAD3/SP1 complex‐mediated constitutive active loop between lncRNA PCAT7 and TGF‐β signaling promotes prostate cancer bone metastasis. Mol Oncol. 2020;14:808‐828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Xiang Y, Zhang Y, Tang Y, et al. MALAT1 modulates TGF‐ββ1‐induced endothelial‐to‐mesenchymal transition through downregulation of miR‐145. Cell Physiol Biochem. 2017;42:357‐372. [DOI] [PubMed] [Google Scholar]
- 47. Tu X, Zhang Y, Zheng X, et al. TGF‐β‐induced hepatocyte lincRNA‐p21 contributes to liver fibrosis in mice. Sci Rep. 2017;7:2957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Zhu HY, Bai WD, Li C, et al. Knockdown of lncRNA‐ATB suppresses autocrine secretion of TGF‐β2 by targeting ZNF217 via miR‐200c in keloid fibroblasts. Sci Rep. 2016;6:24728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Papoutsoglou P, Tsubakihara Y, Caja L, et al. The TGFB2‐AS1 lncRNA regulates TGF‐β signaling by modulating corepressor activity. Cell Rep. 2019;28:3182‐3198.e3111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Wang P, Luo ML, Song E, et al. Long noncoding RNA lnc‐TSI inhibits renal fibrogenesis by negatively regulating the TGF‐β/Smad3 pathway. Sci Transl Med. 2018;10:eaat2039. [DOI] [PubMed] [Google Scholar]
- 51. Sarkar A, Rahaman A, Biswas I, et al. TGFβ mediated LINC00273 upregulation sponges mir200a‐3p and promotes invasion and metastasis by activating ZEB1. J Cell Physiol. 2020; 10.1002/jcp.29614 [DOI] [PubMed] [Google Scholar]
- 52. Cui W, Meng W, Zhao L, et al. TGF‐β‐induced long non‐coding RNA MIR155HG promotes the progression and EMT of laryngeal squamous cell carcinoma by regulating the miR‐155‐5p/SOX10 axis. Int J Oncol. 2019;54:2005‐2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. He W, Liang B, Wang C, et al. MSC‐regulated lncRNA MACC1‐AS1 promotes stemness and chemoresistance through fatty acid oxidation in gastric cancer. Oncogene. 2019;38:4637‐4654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Jiang L, Wang R, Fang LI, et al. HCP5 is a SMAD3‐responsive long non‐coding RNA that promotes lung adenocarcinoma metastasis via miR‐203/SNAI axis. Theranostics. 2019;9:2460‐2474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Liang H, Zhao X, Wang C, et al. Systematic analyses reveal long non‐coding RNA (PTAF)‐mediated promotion of EMT and invasion‐metastasis in serous ovarian cancer. Mol Cancer. 2018;17:96. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 56. Terashima M, Ishimura A, Wanna‐Udom S, et al. MEG3 long noncoding RNA contributes to epigenetic progression of the epithelial‐mesenchymal transition of lung and pancreatic cancer cells. J Biol Chem. 2018;293:18016‐18030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Hu ML, Wang XY, Chen WM. TGF‐β1 upregulates the expression of lncRNA UCA1 and its downstream HXK2 to promote the growth of hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 2018;22:4846‐4854. [DOI] [PubMed] [Google Scholar]
- 58. Li Z, Liu H, Zhong Q, et al. LncRNA UCA1 is necessary for TGF‐β‐induced epithelial‐mesenchymal transition and stemness via acting as a ceRNA for Slug in glioma cells. FEBS Open Bio. 2018;8:1855‐1865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Zuo ZK, Gong Y, Chen XH, et al. TGFβ1‐induced LncRNA UCA1 upregulation promotes gastric cancer invasion and migration. DNA Cell Biol. 2017;36:159‐167. [DOI] [PubMed] [Google Scholar]
- 60. Lu Z, Chen Z, Li Y, et al. TGF‐β‐induced NKILA inhibits ESCC cell migration and invasion through NF‐κB/MMP14 signaling. J Mol Med. 2018;96:301‐313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Wu W, Chen F, Cui X, et al. LncRNA NKILA suppresses TGF‐β‐induced epithelial‐mesenchymal transition by blocking NF‐κB signaling in breast cancer. Int J Cancer. 2018;143:2213‐2224. [DOI] [PubMed] [Google Scholar]
- 62. Zhang C, Hao Y, Wang Y, et al. TGF‐β/SMAD4‐regulated LncRNA‐LINP1 inhibits epithelial‐mesenchymal transition in lung cancer. Int J Biol Sci. 2018;14:1715‐1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Wu DM, Deng SH, Liu T, et al. TGF‐β‐mediated exosomal lnc‐MMP2‐2 regulates migration and invasion of lung cancer cells to the vasculature by promoting MMP2 expression. Cancer Med. 2018;7:5118‐5129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Lu Z, Li Y, Che Y, et al. The TGFβ‐induced lncRNA TBILA promotes non‐small cell lung cancer progression in vitro and in vivo via cis‐regulating HGAL and activating S100A7/JAB1 signaling. Cancer Lett. 2018;432:156‐168. [DOI] [PubMed] [Google Scholar]
- 65. Zhang J, Han C, Ungerleider N, et al. A transforming growth factor‐β and H19 signaling axis in tumor‐initiating hepatocytes that regulates hepatic carcinogenesis. Hepatology. 2019;69:1549‐1563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Matouk IJ, Raveh E, Abu‐lail R, et al. Oncofetal H19 RNA promotes tumor metastasis. Biochim Biophys Acta. 2014;1843:1414‐1426. [DOI] [PubMed] [Google Scholar]
- 67. Kolliopoulos C, Lin CY, Heldin C‐H, et al. Has2 natural antisense RNA and Hmga2 promote Has2 expression during TGFβ‐induced EMT in breast cancer. Matrix Biol. 2019;80:29‐45. [DOI] [PubMed] [Google Scholar]
- 68. Huang T, Huang W, Lu H, et al. Identification and validation a TGF‐β‐associated long non‐coding RNA of head and neck squamous cell carcinoma by bioinformatics method. J Transl Med. 2018;16:46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Terashima M, Tange S, Ishimura A, et al. MEG3 long noncoding RNA contributes to the epigenetic regulation of epithelial‐mesenchymal transition in lung cancer cell lines. J Biol Chem. 2017;292:82‐99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Hao Y, Yang X, Zhang D, et al. Long noncoding RNA LINC01186, regulated by TGF‐β/SMAD3, inhibits migration and invasion through Epithelial‐Mesenchymal‐Transition in lung cancer. Gene. 2017;608:1‐12. [DOI] [PubMed] [Google Scholar]
- 71. Zhuang J, Shen L, Yang L, et al. TGFβ1 promotes gemcitabine resistance through regulating the LncRNA‐LET/NF90/miR‐145 signaling axis in bladder cancer. Theranostics. 2017;7:3053‐3067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Lu W, Zhang H, Niu Y, et al. Long non‐coding RNA linc00673 regulated non‐small cell lung cancer proliferation, migration, invasion and epithelial mesenchymal transition by sponging miR‐150‐5p. Mol Cancer. 2017;16:118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Kong J, Sun W, Li C, et al. Long non‐coding RNA LINC01133 inhibits epithelial‐mesenchymal transition and metastasis in colorectal cancer by interacting with SRSF6. Cancer Lett. 2016;380:476‐484. [DOI] [PubMed] [Google Scholar]
- 74. Fan Y, Shen B, Tan M, et al. TGF‐β‐induced upregulation of malat1 promotes bladder cancer metastasis by associating with suz12. Clin Cancer Res. 2014;20:1531‐1541. [DOI] [PubMed] [Google Scholar]
- 75. Arase M, Horiguchi K, Ehata S, et al. Transforming growth factor‐β‐induced lncRNA‐Smad7 inhibits apoptosis of mouse breast cancer JygMC(A) cells. Cancer Sci. 2014;105:974‐982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Miao F, Chen J, Shi M, et al. LncRNA HAND2‐AS1 inhibits non‐small cell lung cancer migration, invasion and maintains cell stemness through the interactions with TGF‐β1. Biosci Rep. 2019;39:BSR20181525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Huo Y, Li A, Wang Z. LncRNA AWPPH participates in the metastasis of non‐small cell lung cancer by upregulating TGF‐β1 expression. Oncol Lett. 2019;18:4246‐4252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Li W, Wu X, She W. LncRNA POU3F3 promotes cancer cell migration and invasion in nasopharyngeal carcinoma by up‐regulating TGF‐β1. Biosci Rep. 2019;39:BSR20181632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Wang X, Lan Z, He J, et al. LncRNA SNHG6 promotes chemoresistance through ULK1‐induced autophagy by sponging miR‐26a‐5p in colorectal cancer cells. Cancer Cell Int. 2019;19:234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Li YU, Liu G, Li X, et al. Long non‐coding RNA SBF2‐AS1 promotes hepatocellular carcinoma progression through regulation of miR‐140‐5p‐TGFBR1 pathway. Biochem Biophys Res Commun. 2018;503:2826‐2832. [DOI] [PubMed] [Google Scholar]
- 81. Zhang ZS, Wang J, Zhu BQ, et al. Long noncoding RNA UCA1 promotes multiple myeloma cell growth by targeting TGF‐β. Eur Rev Med Pharmacol Sci. 2018;22:1374‐1379. [DOI] [PubMed] [Google Scholar]
- 82. Fu M, Huang Z, Zang X, et al. Long noncoding RNA LINC00978 promotes cancer growth and acts as a diagnostic biomarker in gastric cancer. Cell Prolif. 2018;51(1):e12425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Zhou H, Sun Z, Li S, et al. LncRNA SPRY4‐IT was concerned with the poor prognosis and contributed to the progression of thyroid cancer. Cancer Gene Ther. 2018;25:39‐46. [DOI] [PubMed] [Google Scholar]
- 84. Wu ZJ, Li Y, Wu YZ, et al. Long non‐coding RNA CCAT2 promotes the breast cancer growth and metastasis by regulating TGF‐β signaling pathway. Eur Rev Med Pharmacol Sci. 2017;21:706‐714. [PubMed] [Google Scholar]
- 85. Zhang Q, Chen B, Liu P, et al. XIST promotes gastric cancer (GC) progression through TGF‐β1 via targeting miR‐185. J Cell Biochem. 2018;119:2787‐2796. [DOI] [PubMed] [Google Scholar]
- 86. Qin CF, Zhao FL. Long non‐coding RNA TUG1 can promote proliferation and migration of pancreatic cancer via EMT pathway. Eur Rev Med Pharmacol Sci. 2017;21:2377‐2384. [PubMed] [Google Scholar]
- 87. Liu Y, Qian J, Li X, et al. Long noncoding RNA BX357664 regulates cell proliferation and epithelial‐to‐mesenchymal transition via inhibition of TGF‐β1/p38/HSP27 signaling in renal cell carcinoma. Oncotarget. 2016;7:81410‐81422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Zhao JJ, Hao S, Wang LL, et al. Long non‐coding RNA ANRIL promotes the invasion and metastasis of thyroid cancer cells through TGF‐β/Smad signaling pathway. Oncotarget. 2016;7:57903‐57918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Chen D, Zhang Z, Mao C, et al. ANRIL inhibits p15INK4b through the TGFβ1 signaling pathway in human esophageal squamous cell carcinoma. Cell Immunol. 2014;289:91‐96. [DOI] [PubMed] [Google Scholar]
- 90. Sakai S, Ohhata T, Kitagawa K, et al. Long noncoding RNA ELIT‐1 acts as a Smad3 cofactor to facilitate TGF‐β/Smad signaling and promote epithelial‐mesenchymal transition. Cancer Res. 2019;79:2821‐2838. [DOI] [PubMed] [Google Scholar]