

Abstract
A lithoautotrophic, Fe(II) oxidizing, nitrate-reducing bacterium, strain 2002 (ATCC BAA-1479; =DSM 18807), was isolated as part of a study on nitrate-dependent Fe(II) oxidation in freshwater lake sediments. Here we provide an in-depth phenotypic and phylogenetic description of the isolate. Strain 2002 is a gram-negative, non-spore forming, motile, rod-shaped bacterium which tested positive for oxidase, catalase, and urease. Analysis of the complete 16S rRNA gene sequence placed strain 2002 in a clade within the family Neisseriaceae in the order Nessieriales of the Betaproteobacteria 99.3% similar to Pseudogulbenkiania subflava. Similar to P. sublfava, predominant whole cell fatty acids were identified as 16:17c, 42.4%, and 16:0, 34.1%. Whole cell difference spectra of the Fe(II) reduced minus nitrate oxidized cyctochrome content revealed a possible role of c-type cytochromes in nitrate-dependent Fe(II) oxidation. Strain 2002 was unable to oxidize aqueous or solid-phase Mn(II) with nitrate as the electron acceptor. In addition to lithotrophic growth with Fe(II), strain 2002 could alternatively grow heterotrophically with long-chain fatty acids, simple organic acids, carbohydrates, yeast extract, or casamino acids. Nitrate, nitrite, nitrous oxide, and oxygen also served as terminal electron acceptors with acetate as the electron donor.
Keyword: Fe(II) oxidation, Anaerobic, Nitrate, Bioremediation, Uranium
Introduction
Microbial oxidation of Fe2+ and solid-phase Fe(II)-bearing minerals can significantly affect the geochemistry of saturated soils and sediments by the formation of reactive Fe(III) minerals, including mixed Fe(II)–Fe(III) minerals, which may result in the sorption or coprecipitation of metals and/or nutrients (Chaudhuri et al. 2001; Cornell & Schwertmann 2003; Lack et al. 2002b; Weber et al. 2001; Weber et al. 2006c). In recent years, the demonstration of anaerobic microbial Fe(II) oxidation, light-dependent or nitrate-dependent, has been identified as a mechanism in which biological re-oxidation of Fe(II) in an anoxic environment can occur (Straub et al. 1996; Weber et al. 2006a; Widdel et al. 1993). To date, several microorganisms have been identified that are capable of anaerobic Fe(II) oxidation (Bruce et al. 1999; Chaudhuri et al. 2001; Edwards et al. 2003; Finneran et al. 2002; Hafenbradl et al. 1996; Lack et al. 2002a; Straub et al. 1996; Straub et al. 1999; Widdel et al. 1993), however, few of these grow by the metabolism (Weber et al. 2006a). Given that light penetration of soil and particulate matter is only between 8 to 200 μm (Ciania et al. 2005), light independent reactions such as nitrate-dependent Fe(II) oxidation are more likely to dominate bio-oxidative processes in saturated and subsurface soil/sedimentary environments.
Nitrate-dependent Fe(II)-oxidizing microorganisms have been identified in both the Archaea and Bacteria (Weber et al. 2006a). Among the isolates currently described only a hyperthermophilic archaeum (Hafenbradl et al. 1996) and the Betaproteobacterium strain 2002 (Weber et al. 2006b) have been demonstrated to oxidize Fe(II) as the sole electron donor and couple this metabolism to autotrophic growth. For all other isolates described to date either growth was not demonstrated or a co-substrate (acetate or H2) was required under iron oxidizing conditions (Benz et al. 1998; Chaudhuri et al. 2001; Edwards et al. 2003; Finneran et al. 2002; Lack et al. 2002a; Sheloblina et al. 2003; Straub & Buchholz-Cleven 1998; Straub et al. 1996; Straub et al. 2004). To date, the only other microbial isolate in pure culture capable of autotrophic growth with Fe(II) as the sole electron donor and nitrate as a terminal electron acceptor is the hyperthermophilic archaeum, Ferroglobus placidus (Hafenbradl et al. 1996). It is worthwhile to note that under autotrophic growth conditions Thiobacillus denitrificans has also been demonstrated to oxidize metals coupled to nitrate reduction; however, metal oxidation is dependent on H2 as a co-electron donor (Beller 2005; Beller et al. 2009). It is unclear if Fe(II) could serve as the sole electron donor for nitrate reduction in this bacterium.
Strain 2002 was isolated from a nitrate-dependent Fe(II) oxidizing microbial community in freshwater lake sediments (Weber et al. 2006b). The natural abundance of strain 2002 within the freshwater lake sediment was estimated at 2.4 × 103 cells g-1 wet sediment based on most probable number enumeration studies (Weber et al. 2006b). Strain 2002 was described as a member of the family Nesseriaceae (Weber et al. 2006b). This family is comprised of organisms commonly identified as environmental bacteria as well as mammalian, avian, and invertebrate pathogens and commensals (Corpe 1951; Gillis & Logan 2005; Hungria et al. 2005; Thomsen et al. 2007; Weber et al. 2006c). From among the environmentally relevant genera of the Nesseriaceae, Chromobacterium spp. are frequently identified in freshwater aquatic environments, soils, and sediments (Corpe 1951; Gillis & Logan 2005; Hungria et al. 2005; Thomsen et al. 2007; Weber et al. 2006c). The metabolic versatility of Chromobacterium spp., specifically C. violaceum, has resulted in a variety of industrial applications (Vasconcelos et al. 2003). Related to metal biogeochemistry, the biogenic production of cyanide during aerobic growth of C. violaceum has been demonstrated to mobilize precious metals such as silver, gold, and platinum as a result of the formation of metal–cyanide aqueous complexes (Brandl et al. 2007; Reith et al. 2007). The ability of this microorganism to oxidize aqueous Fe(II) to an insoluble Fe(III) oxide mineral under anaerobic conditions at neutral pH in addition to strain 2002 was recently demonstrated (Weber et al. 2006b). However, similar to many other known Fe(II)-oxidizing organisms, C. violaceum did not grow by this metabolism (Weber et al. 2006b). The coprecipitation of heavy metal and radionuclide contaminants with biogenic Fe(III) oxides has been proposed as a bioremediation strategy (Lack et al. 2002b). Thus, implicating the role of members within the family Nesseriaceae to participate in metal biogeochemical cycling. Because of its associated growth, strain 2002 presents a novel opportunity to study nitrate- dependent metal oxidation and the environmental implications of this microbial metabolism in pure culture. However, it is first necessary to detail the taxonomic and physiological characteristics of this microorganism for further study. Here we further describe the physiology and phylogenetic placement of the novel lithoautotrophic, nitrate-dependent Fe(II) oxidizing bacterium, strain 2002, as a member of the recently described genus Pseudogulbenkiania in relation to closely related genera in the family Nesseriaceae.
Materials and methods
Source and culturing of microorganisms
Strain 2002 (ATCC BAA-1479; =DSM 18807) was previously isolated from a nitrate-dependent Fe(II) oxidizing most probable number enumeration series initiated from sediments collected from a freshwater lake, Southern Illinois University campus, Carbondale, Illinois (Weber et al. 2006b) and maintained on freshwater basal medium. Freshwater basal medium contained the following components (l−1): 0.25 g NH4Cl, 0.6 g NaH2PO4, 0.1 g KCl, 0.42 g NaNO3, and 2.52 g NaHCO3. The vitamin and trace mineral solution was added from sterile stock solution (10 ml l−1). The vitamin solution contained (l−1): 2 mg d-biotin, 2 mg folic acid, 10 mg pyridoxine HCl, 5 mg riboflavin, 5 mg thiamine, 5 mg nicotinic acid, 5 mg pantothenic acid, 0.1 mg vitamin B12, 5 mg p-amino benzoic acid, and 5 mg d,l-6,8-thiotic cid. The trace mineral solution was prepared by dissolving the following in a 1.5 g l−1 nitrilotriacetic acid disodium salt solution (l−1): 3 g MgSO4⋅7H2O, 0.5 g MnSO4, 1.0 g NaCl, 0.1 g FeSO4⋅7H2O, 0.1 g CaCl2⋅2H2O, 0.1 g CoCl2⋅6H2O, 0.13 g ZnCl, 0.01 g CuSO4⋅5H2O, 0.01 AlK(SO4)2⋅12H2O, 0.01 g H3BO3, 0.025 g Na2MoO4⋅2H2O, 0.024 g NiCl2⋅6H2O, 0.025 g Na2WO4⋅2H2O, 0.02 g Na2SeO4. Anoxic freshwater basal medium was prepared by heating the solution to 100°C and immediately cooling it to room temperature under an anoxic atmosphere (N2:CO2; 80:20). The anoxic basal medium was then dispensed into pressure tubes under an N2:CO2 atmosphere (80:20, v/v), sealed with blue butyl stoppers, and autoclaved. Fe(II) source was added from a concentrated anoxic sterile stock solution to achieve a final concentration of 10 mM. A concentrated soluble sterile Fe(II) stock solution (1 M) was prepared by adding FeCl2 to anoxic (100% N2 headspace) distilled deionized water and sealed with a blue butyl stopper under an anoxic atmosphere (100% N2 headspace). The solution was filter sterilized by passing the solution directly through a sterile 0.22 μm nylon filter and sterile needle into a sterile, anoxic (100% N2 headspace), stoppered serum bottle. For the purposes of this study, strain 2002T was routinely maintained under anaerobic conditions on basal medium with a soluble electron donor and acceptor, acetate (10 mM) and nitrate (10 mM) respectively. Standard anaerobic microbiological culture techniques were employed as previously described (Hungate 1969). Studies determining the optimal pH, temperature, and salinity were carried out on basal medium containing sodium acetate (10 mM) and sodium nitrate (10 mM).
Electron donors and acceptors
The ability of strain 2002 to utilize alternative electron donors and acceptors was tested by adding chemicals from anoxic, sterile stock solutions to freshwater basal medium as described above. Ferric iron was added from a filter sterilized 1 M soluble iron stock solution prepared as previously described by dissolving nitrilotriacetic acid disodium salt (NTA), sodium bicarbonate, and ferric chloride (Roden & Lovely 1993). Benzene, toluene, and hexadecane were added directly to culture medium. Growth after three successive transfers was recorded as positive. Growth was determined by changes in optical density at 600 nm. In cultures containing solid-phase electron donors and/or acceptors cell density was monitored by direct cell counts (Petroff-Hausser Counter, 0.02 mm depth). Samples collected for direct cell counts were immediately fixed in formaldehyde (final concentration 3.7%).
Analytical techniques
Ion chromatrography with conductivity detection (IonPac® AS9-HC analytical column, Dionex DX-500 system, Dionex Corp., Sunnyvale, CA). was used to analyze NO−3 and NO−2. Concentrations of NH+4 in growth experiments were determined spectrophotometrically according to Wetzel & Likens (1991). Batch culture headspace N2 was quantified using a gas chromatograph equipped with a thermal conductivity detector (Shimadzu GC-8A; Porapak N, 80–100 mesh, 12ft. × 1/8 in. column).
Electron microscopy
Cells for electron microscopy were grown anaerobically in freshwater basal medium (described above) amended with acetate (10 mM) and NO−3 (10 mM). Cells were harvested by centrifugation during log growth phase under an anaerobic atmosphere (N2:CO2; 80:20), washed twice with 0.1 M sodium cacodylate buffer pH 7.2, and fixed with glutaraldehyde (final concentration 3% v/v). Cells were then resuspended in 1% osmium tetroxide in sodium cacodylate buffer for 2 h and rinsed in sodium cacodylate buffer. Cells were dehydrated subsequently for 10 min in 35%, 50%, 70%, 80%, 95%, 100%, and 100% ethanol followed by critical point drying. Cells were mounted onto stubs and sputter coated with palladium/gold and viewed with a Hitachi S5000 scanning electron microscope at 20 kV.
Phospholipid fatty acids, DNA base composition, and phylogenetic analyses
Major phospholipid fatty acids of strain 2002T were determined using methods described by Singleton et al. (2003) when grown under heterotrophic (10 mM acetate and 10 mM nitrate) and lithoautotrophic (10 mM Fe(II) and 5 mM nitrate) conditions. Whole cell fatty acids of strain 2002 and pure culture relatives, Chromobacterium violaceum ATCC 12472T, Chromobacterium subtsugae PRAA4-1T, Aquitalea magnusonii TRO-001-1DR8T, and Gulbenkiania mobilis E4FC31T, grown aerobically on tryptic soy agar were determined as previously described (Komagata & Suzuki 1987; MIDI 2001). G + C content of strain 2002 was calculated from the ratio of deoxyguanosine (dG) and thymidine (dT) as previously described (Mesbah et al. 1989). Phylogenetic analysis was conducted using 1395 MUSCLE aligned characters from the 16S rRNA gene sequences of members from strain 2002 (AY609199), close relatives, and the family Neisseriaceae as follows: Pseudogulbenkiania subflava (EF626692), clone JH WH6 (EF492895), Paludimonas yongneupensis (AM396358), Gulbenkiania mobilis (AM295491), Chromobacterium violaceum (M22510), Chromobacterium str. MBIC3901 (AB017487), Chromobacterium str. MBIC3903 (AB017489), Chromobacterium subtsugae (AY344056), Aquitalea magnusonii (DQ018117), Aquitalea sp. LT 5 (EU287928), Aquitalea sp. H4 4 (AB277845), Aquitalea sp. PGP 1 (AB277847), (L06174), Alysiella filiformis (AB087263), Uruburuella suis (AJ586614), Conchiformibius steedae (AF328153), Neisseria gonorrhoeae (X07714), Bergeriella denitrificans (L06173), Simonsiella muelleri (AF328147), Eikenella corrodens (M22512), Kingella denitrificans (M22516), Vogesella indigofera (AB021385), Aquaspirillum serpens (AB074518), Microvirgula aerodenitrificans, (U89333), Laribacter hongkongensis (AF389085), Iodobacter fluviatilis (M22511), Chitinibacter tainanensis (AY264287), and Silvimonas terrae (AB194302) with Methylophilus methylotrophus (M29021) as the outgroup. Bayesian consensus analysis of aligned 16S rRNA gene sequences was performed using MRBAYES using GTR + gamma model of evolution for 4,000,000 generations with a tree sample frequency every 1,000 generations creating a posterior probability distribution (Huelsenbeck & Ronquist 2001). The alignments were also subjected to maximum-likelihood analysis in Bio-Edit version 7.0.9.0 using default parameters (Hall 1999).
Results
Cell morphology and growth conditions
Cells of strain 2002 as previously described (Weber et al. 2006b) were rod-shaped, gram-negative, non-spore forming, flagellated, and motile, occurring as single cells or in pairs or multiples (Fig. 1). Colonies of strain 2002 grown aerobically on tryptic soy agar (TSA) or solidified nutrient broth, appear pink–orange, smooth, convex, and circular with a 1–2 mm diameter. When grown anaerobically on acetate (10 mM) and NO−3 (10 mM), rod-shaped cells of strain 2002 were ca. 0.5 μm in diameter and 1.5 μm in length. Hemolytic activity was observed when strain 2002 was grown aerobically on TSA with 5% sheep’s blood (Environmental Microbiology Laboratory, Inc. Part #1643). Strain 2002 grew over a temperature range 15-40°C with an optimal growth temperature of 37°C. No growth was observed at 4°C and 45°C. Growth was observed over a pH range of 6.75–8.0 and salinity (NaCl) range of 0–1%. Optimal pH and salinity were determined to be 7.25 and 0% respectively.
Fig. 1.

Scanning electron micrograph of strain 2002 grown under heterotrophic nitrate reducing conditions, 10 mM acetate and 10 mM nitrate. Bar denotes image scale, 0.5 μm
Phylogenetic and chemotaxonomic comparisons
Pairwise comparison of 16S rRNA gene sequences of cultivated bacteria identified Pseudogulbenkiania subflava as the nearest cultured relative (99.3% 16S rRNA gene sequence identity) and an uncultured bacterial clone affiliated with iron-manganese nodules (GenBank accession# EF492895; (He et al. 2008)). Phylogenetic analysis of the complete 16S rRNA gene sequence of pure culture representatives placed strain 2002 in a clade with P. subflava and separated strain 2002 from the closely related genera Paludimonas, Gulbenkiania, Chromobacterium, and Aquitalea, as well as and other pure culture relatives of the family Nesseriaceae in the order Neisseriales within the Betaproteobacteria (Fig. 2). Commercial (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, DSMZ) determination of the G + C content (Mesbah et al. 1989) of the genomic DNA of strain 2002 gave a value of 64 mol%, within the range of the DNA base composition described within the family Neisseriaceae (44–69%; Tonjum 2005); Pseudogulbenkiania subflava (63%), Paludimonas spp. (63%; Kwon et al. 2008), Gulbenkiania spp. (59%; Vaz-Moreira et al. 2007), Chromobacterium spp. (65–68 mol%; Gillis & Logan 2005), and Aquitalea spp. (65%; Lau et al. 2006).
Fig. 2.

Bayesian consensus phylogenetic tree of 1,359 aligned characters from the 16S rRNA gene sequence of strain 2002 and members of the family Neisseriaceae after 4,000,000 generations with a sample frequency of 1,000. Results from Bayesian posterior probability are indicated at the nodes as follows: filled circle 1.00, empty circle >0.90, filled square, 0.80–0.90; empty square <0.80. Methylophilus methylotrophus, a member of the family Methylophilaceae within the Betaproteobacteria, served as the outgroup. The same topology was obtained with maximum-likelihood analysis
The major phospholipid fatty acids identified under lithoautotrophic and heterotrophic growth conditions were 16:0 (38 mol%), 16:1ω7c (24–33 mol%), Cy17:0 (16–21 mol%), and 18:1ω7c (12–15 mol%; Table 1). Similar phospholipids fatty acid profiles were obtained under both growth conditions. A subsequent analysis of the whole cell fatty acid content of strain 2002 identified the predominant fatty acids as 16:1ω7c, 42.4%, and 16:0, 34.1%, using previously described methods (Komagata & Suzuki 1987; MIDI 2001). Other fatty acids detected greater than 1 mol%, 3oh10:0, 12:0, 3oh12:0, 14:0, Cy17:0, and 18:1ω7c (Table 2) totaled 21.2 mol%. Principle components and cluster analyses (Minitab 15.1.1.0) of the whole cell fatty acid data indicated that strain 2002 was chemotaxonomically similar to P. subflava but different from other near relatives in pure culture, Paludimonas spp., Microvirgula spp., Gulbenkiania spp., Chromobacterium spp., Aquitalea spp. (Fig. 3a & b). Principal component analysis extracted two factors which accounted for 86% of the total variance (Fig. 3a), and clearly placed strain 2002 in a cluster with P. subflava (Fig. 3b).
Table 1.
Phopholipid fatty acid composition of strain 2002T grown under heterotrophic conditions (10 mM Acetate and 10 mM NO−3) and lithoautotrophic conditions (10 mM FeCl2 and 5 mM NO−3)
| Mol % heterotrophically grown | Mol % lithoautotrophically grown | |
|---|---|---|
| Saturated | ||
| 14:0 | 0.89 | 0 |
| 15:0 | 0.34 | 0 |
| 16:0 | 36.67 | 36.74 |
| 17:0 | 0.05 | 0 |
| 18:0 | 0.4 | 1.06 |
| Branched chain monounsaturated | ||
| br19:1 | 0.1 | 0 |
| Monounsaturated | ||
| 16:1ω9c | 0.18 | 0 |
| 16:1ω7c | 32.82 | 23.53 |
| 16:1ω7t | 0.19 | 0.98 |
| 16:1ω5c | 0.35 | 0.61 |
| 17:1 | 0.06 | 0 |
| Cy17:0 | 15.45 | 21.15 |
| 18:1ω7c | 11.82 | 14.73 |
| 18:1ω7t | 0 | 0.53 |
| 18:1ω5c | 0.13 | 0 |
| Cy19:0 | 0.47 | 0.68 |
| Hydroxy | ||
| 3OH16:0 | 0.09 | 0 |
Table 2.
Phenotypic and biochemical characteristics of strain 2002 (1) and relative cehtnothe type strains of the closely related genera within the family Neisseriaceae, Psuedogulbenkiania subflava BP-5T (2), Pauldibacterium yongneupense 5YN8-15T (3), Chromobacterium violaceum ATCC 12472T (4), Chromobacterium subtsugae PRAA4-1T (5), Aquitalea magnusonii TRO-001-1DR8T (6), and Gulbenkiania mobilis E4FC31T (7)
| Characteristic | 1 | 2a | 3b | 4c | 5f | 6g | 7h |
|---|---|---|---|---|---|---|---|
| Isolation source | Freshwater lake sediment | Stream water | Wetland peat | Soil/water | Soil | Humic lake | Municipal wastewater |
| Colony color | Pink–orange | Yellow | Violet | Violet | Tan | No pigment | |
| DNA G + C content (mol%) | 64 | 63 | 63 | 65–68 | 65 | 59 | 63 |
| Growth Temperature | 15–40 | 15–42 | 4–35 | 28–40 | 4–40 | 28–37 | 15–45 |
| pH Growth | 6.75–8 | 6–8 | 4–8 | 5–8 | 4.5–9.0 | 5–8 | 5.5–9 |
| NaCl tolerance (%) | <1 | <1 | <5 | <4 | <3 | <1.5 | <3 |
| Urease | + | − | − | −e | NT | − | − |
| Lithoautotrophic | + | NT | NT | − | NT | NT | − |
| Carbon utilization of | |||||||
| Maltose | + | + | + | − | NT | − | − |
| Glucose | + | + | + | + | + | + | − |
| Sucrose | + | + | NT | − | − | − | − |
| Trehalose | + | + | NT | + | + | − | − |
| Capric acid | + | + | − | +e | + | NT | + |
| Mannose | − | − | − | + | + | − | − |
| N-acetylglucosamine | − | − | + | + | + | + | − |
| Use of electron acceptors | |||||||
| Nitrous oxide | + | − | NT | −d | − | + | − |
| Fermentation of | |||||||
| Glucose | − | − | + | + | + | + | − |
| Ribose | − | − | − | + | NT | − | − |
| HCN production | − | NT | NT | + | NT | NT | − |
| Hydrolysis of Gelatin | − | NT | − | + | + | − | − |
NT Not tested
aCharacteristics of P. subflava obtained from Kwon et al. (2008) and Lin et al. (2008)
bCharacteristics of P. yongneupense obtained from Kwon et al. (2008)
cCharacteristics of C. violaceum obtained from Gillis & Logan (2005) unless otherwise noted
dCharacteristics of C. violaceum obtained from Bazylinski et al. (1986)
eCharacteristics of C. violaceum obtained from the Identification table of the bioMérieux API 20 NE kit
fCharacteristics of C. subtsugae obtained from Martin et al. (2007) unless otherwise noted
gCharacteristics of A. magnusonii obtained from Lau et al. (2006) unless otherwise noted
hCharacteristics of G. mobilis obtained from Vaz-Moreira et al. (2007) unless otherwise noted
Fig. 3.

Principle components analysis of whole cell fatty acids of strain 2002 and closely related genera. The first two factors extracted account for 86% of the total variance. Inclusion of the third factor accounts for 97% of the total variance and further separates strain 2002 from Pauldibacterium, Gulbenkiania, Aqualitea and Chromobacterium strains. a Cluster analysis of the chemotaxonomic data set supported the separation of strain 2002 from Pauldibacterium, Gulbenkiania, Aqualitea and Chromobacterium strains and supported the similarities of the whole cell fatty acid content of strain 2002 with Pseudogulbenkiania subflava. b Clusters in the principle components analysis are denoted by the circles
Metabolic characterization
Phenotypic characterization was conducted with a commercial biochemical kit, API 20 NE (bioMérieux), according to the manufacturer’s instructions using supplemented freshwater basal medium. Strain 2002 tested positive for oxidase, catalase, urease, and arginine dihydrolase, and is capable of utilizing d-glucose, d-malate, potassium gluconate, capric acid, malic acid, and trisodium citrate. Strain 2002 cannot utilize l-arabinose, d-mannose, d-mannitol, N-acetylglucosamine, adipic acid, or phenylacetic acid and it does not produce hydrogen sulfide, gelatinase, β-glucosidase, or β-galactosidase (Table 2). Strain 2002 had been previously described as capable of anaerobically growing with Fe(II) as an electron donor coupled to the reduction of nitrate under autotrophic conditions (Weber et al. 2006b). Difference spectrum of the Fe(II)-reduced minus nitrate oxidized cytochrome content of whole cells of strain 2002 revealed an absorption spectrum consistent with c-type cytochromes (absorption maxima at a wavelength at 424, 521, and 553 nm; Fig. 4). In addition to the ability of strain 2002 to oxidize Fe(II), this bacterium has also previously been shown to oxidize uraninite, U(IV), in the presence of nitrate (Weber KA et al., 2009). In contrast to the oxidation of Fe(II) and U(IV), strain 2002 was incapable of anaerobically utilizing Mn(II) as an electron donor under these growth conditions (Table 3).
Fig. 4.

Whole cell difference spectrum of the Fe(II) reduced minus nitrate oxidized cytochrome content within intact live cells of strain 2002 suspended in anoxic (100% N2 atmosphere) PIPES buffer (20 mM, pH7.0). Arrows indicate absorption maxima with the corresponding wavelength denoted above the arrow. Vertical scale bar indicates a change of 0.025 absorbance units
Table 3.
Electron donors and accepters tested
| Electron donors | Electron acceptors |
|---|---|
| Utilized | Utilized |
| Fe(II) (10 mM) | O2 |
| Acetate (10 mM) | NO−3 |
| Fumarate (25 mM) | NO−2 |
| Lactate (10 mM) | N2O |
| Malate (10 mM) | |
| Succinate (10 mM) | |
| Citrate (10 mM) | |
| Glucose (10 mM) | |
| Sucrose (10 mM) | |
| Yeast Extract (1 g/L) | |
| Pyruvate (5 mM) | |
| Fructose (10 mM) | |
| Maltose (10 mM) | |
| Propionate (10 mM) | |
| Trehalose (10 mM) | |
| Ethanol (10 mM) | |
| Valeric acid (5 mM) | |
| Casamino acids (1 g L−1) | |
| Butyric acid (5 mM) | |
| Palmitic acid (1 mmol L−1) | |
| Stearic acid (1 mmol L−1) | |
| Lauric acid (1 mmol L−1) | |
| Behenic acid (1 mmol L−1) | |
| Lignoceric acid (1 mmol L−1) | |
| Tested not utilized | |
| Formate (10 mM) | SO2−4 (10 mM) |
| Glycerol (10 mM) | ClO−3 (10 mM) |
| Methanol (5 mM) | ClO−4 (10 mM) |
| Lactose (10 mM) | U(VI) (1 mM) |
| Benzoate (1 mM) | AQDS (10 mM) |
| Phenol (1 mM) | As(V) (0.5 mM) |
| Benzene (0.03 mM) | Fumarate (25 mM) |
| Toluene (0.03 mM) | Thiosulfate (10 mM) |
| Thiosulfate (10 mM) | Fe(III)-NTAa |
| n-eicosane (1 mM) | |
| n-tetracosane (1 mM) | |
| Mineral oil hexadecane | |
| Mn(II) (10 mM) | |
aStrain 2002 could not be consistently maintained under Fe(III) reducing conditions, however Fe(III) reduction was observed in initial transfers
In the absence of Fe(II), strain 2002 grew heterotrophically with several simple organic compounds including acetate, propionate, butyrate, ethanol, pyruvate, and succinate as the sole carbon and energy source (Table 3). In addition, strain 2002 is also capable of utilizing other short chain volatile fatty acids, a variety of simple dicarboxylic acids, hexoses, yeast extract, and casamino acids as well as the long-chain fatty acids palmitic acid, stearic acid, lauric acid, behenic acid, and lignoceric acid as the sole carbon and energy source with nitrate as the terminal electron acceptor (Table 3).
Strain 2002 was relatively limited in the range of alternative electron acceptors used and with acetate as the electron donor; it grew aerobically or anaerobically with nitrate, nitrite, or nitrous oxide. Denitrification by strain 2002 terminated with the production of dinitrogen gas (Fig. 5). Reduction of Fe(III) to Fe(II) was observed in the initial transfer of an anaerobic acetate oxidizing, nitrate-reducing culture into fresh anaerobic medium amended with Fe(III)-NTA as the sole electron acceptor, however strain 2002 could not be continuously cultured under Fe(III)-reducing conditions (Weber et al. 2006b). In contrast to Chromobacterium spp., strain 2002 is not capable of fermentation as no growth was observed with any of the above electron donors or in complex organic medium containing yeast extract (1 g L−1), casamino acids (1 g L−1), or glucose (10 mM) unless a suitable electron acceptor was added. Strain 2002 was capable of incorporating CO2 into biomass when grown under autotrophic nitrate-dependent Fe(II)-oxidizing conditions (Weber et al. 2006b).
Fig. 5.

Denitrification by strain 2002. Error bars indicate standard deviation of triplicate samples. filled inverted triangle Optical density, filled circle NO−3; filled upright triangle Live NO−2, filled diamond N2
Discussion
Strain 2002 was previously described as capable of anaerobically growing with Fe(II) as an electron donor coupled to the reduction of nitrate under autotrophic conditions (Weber et al. 2006b). To date, strain 2002 remains the only mesophilic isolate in pure culture described as capable of autotrophic growth with Fe(II) serving as the sole electron donor. Isolation of strain 2002 from freshwater lake sediments harboring an abundant nitrate-dependent Fe(II)-oxidizing microbial community suggests that microorganisms such as this could play a significant role in iron biogeochemical cycling and iron mineral formation in freshwater sediments. Given the ability of strain 2002 to utilize not only Fe(II) but also simple dicarboxylic acids, short chain volatile fatty acids, hexoses, yeast extract, casamino acids and long-chain fatty acids, it is thus likely that strain 2002 also plays a significant role in the degradation of organic matter in freshwater soils and sedimentary environments. Strain 2002 represents a recently recognized lineage within the family Neisseriaceae as determined by phylogenetic and chemotaxonomic analyses (Figs. 2 and 3). The recent identification of P. subflava, strain 2002, and uncultured bacterial clones affiliated with rice roots, iron-manganese nodules, and freshwater lake sediments (He et al. 2008; Scheid et al. 2004; Tamaki et al. 2005; Weber et al. 2006b) indicates that this genus is present in various surface soil/sedimentary environments and could potentially play a role in metal redox cycling. Given the significance of biogenic Fe(III) oxide precipitation to iron biogeochemical cycling and metal mobility (Weber et al. 2006a), it is necessary to understand the phylogenetic and metabolic characters of strain 2002.
Strain 2002 was capable of autotrophic growth with Fe(II) as the sole electron donor (Weber et al. 2006b). Consistent with previously published reports demonstrating the metabolic role of c-type cytochromes in anaerobic iron oxidation (Chaudhuri et al. 2001; Croal et al. 2007; Jiao et al. 2007), the difference spectrum of the Fe(II) reduced cytochrome content in cells of strain 2002 suggested that c-type cytochromes in strain 2002 may also be involved in Fe(II) oxidation (Fig. 4). While strain 2002 has been demonstrated to oxidize Fe(II) as well U(IV) (Weber KA et al., 2009), physiological characterization revealed that Mn(II) could not serve as an electron donor. While the net reaction of neutrophilic, anaerobic Mn(II) oxidation may be energetically favorable according to Eq. 1, the initial nitrate reduction reaction to nitrite would consume energy (Eq. 2). As such, the unfavorable thermodynamics of this initial reaction step may preclude strain 2002 from using Mn(II) as an electron donor under conditions tested in this study.
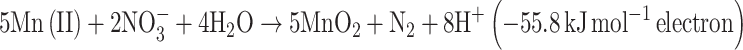 |
1 |
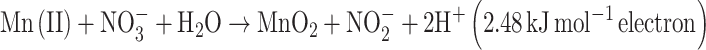 |
2 |
Strain 2002 does not have the ability to produce cyanide. As such, strain 2002 could not mobilize metals via the formation of metal–cyanide complexes, in a similar manner to C. violaceum (Brandl et al. 2007; Reith et al. 2007). However, the ability of strain 2002 to directly oxidize metals can directly influence metal mobility in the environment (Chaudhuri et al. 2001; Cornell & Schwertmann 2003; Lack et al. 2002b; Weber et al. 2001; Weber et al. 2006c). A recent study demonstrated the potential bioremediative applicability of this metabolism for the attenuation of soluble contaminating heavy metals and radionuclides in aquifer systems (Lack et al. 2002b). In that study, stimulation of nitrate-dependent Fe(II) oxidation resulted in the rapid precipitation and immobilization of over 50% of the uranium and 80% of the cobalt in solution within five days.
To date, C. violaceum and strain 2002, represent the members within the family Neisseriaceae capable of metabolically influencing metal biogeochemical cycling. While there are many metabolic dissimilarities from strain 2002 there are several metabolic similarities with the nearest pure culture relative P. subflava. These two organisms are capable of using the following carbon sources: maltose, glucose, sucrose, trehalose, and capric acid (Table 2). In contrast to strain 2002, P. subflava tested negative for urease and nitrous oxide reduction (Table 2). Together, the phylogenetic, chemotaxonomic, and phenotypic distinctions between strain 2002 and other members of the family Neisseriaceae support placement as a member of the Psuedogulbenkiania genus (=ATCC BAA-1479 = DSM 18807).
Acknowledgments
This research was supported by the Department of Energy, Natural and Accelerated Bioremediation Program, through grants #DE-FG02-98ER63592 to JDC and LAA. We sincerely thank Dr. Gordon Vrdoljak for the scanning electron micrograph of strain 2002.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Footnotes
The GenBank accession number for the 16S rRNA gene sequence of strain 2002 is AY609199
Strain 2002 was deposited in the American Type Culture Collection as well as the Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH under the following accession numbers: ATCC BAA-1479; =DSM 18807.
References
- Bazylinski DA, Palome E, Blakemore NA, Blakemore RP. Denitrification by Chromobacterium violaceum. Appl Environ Microbiol. 1986;52:696–699. doi: 10.1128/aem.52.4.696-699.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beller HR. Anaerobic, Nitrate-Dependent Oxidation of U(IV) Oxide Minerals by the Chemolithoautotrophic Bacterium Thiobacillus denitrificans. Appl Environ Microbiol. 2005;71:2170–2174. doi: 10.1128/AEM.71.4.2170-2174.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beller HR, Legler TC, Bourguet F, Letain TE, Kane SR, Coleman MA. Identification of c-type cytochromes involved in anaerobic, bacterial U(IV) oxidation. Biodegradation. 2009;20:45–53. doi: 10.1007/s10532-008-9198-y. [DOI] [PubMed] [Google Scholar]
- Benz M, Brune A, Schink B. Anaerobic and aerobic oxidation of ferrous iron at neutral pH by chemoheterotrophic nitrate-reducing bacteria. Arch Microbiol. 1998;169:159–165. doi: 10.1007/s002030050555. [DOI] [PubMed] [Google Scholar]
- Brandl H, Lehmann S, Faramarzi MA. Mobilization of silver, gold, and platinum from solid materials by HCN-forming microorganisms. Adv Mat Res. 2007;20–21:50–53. [Google Scholar]
- Bruce RA, Achenbach LA, Coates JD. Reduction of (per) chlorate by a novel organism isolated from paper mill waste. Environ Microbiol. 1999;1:319–329. doi: 10.1046/j.1462-2920.1999.00042.x. [DOI] [PubMed] [Google Scholar]
- Chaudhuri SK, Lack JG, Coates JD. Biogenic magnetite formation through anaerobic biooxidation of Fe(II) Appl Environ Microbiol. 2001;67:2844–2848. doi: 10.1128/AEM.67.6.2844-2848.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciania A, Gossa K-U, Schwarzenbach RP. Light penetration in soil and particulate minerals. Eur J Soil Sci. 2005;56:561–574. [Google Scholar]
- Cornell RM, Schwertmann U. The iron oxides: structure, properties, reactions, occurrences and uses. 2. Weinheim: WILEY-VCH; 2003. [Google Scholar]
- Corpe WA. A study of the wide spread distribution of chromobacterium species in soil by a simple technique. J Bacteriol. 1951;62:515–517. doi: 10.1128/jb.62.4.515-517.1951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croal L, Jiao Y, Newman D. The fox operon from Rhodobacter strain SW2 promotes phototrophic Fe(II) oxidation in Rhodobacter capsulatus SB1003. J Bacteriol. 2007;189:1774–1782. doi: 10.1128/JB.01395-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards KJ, Rogers DR, Wirsen CO, McCollom TM. Isolation and characterization of novel psychrophilic, neutrophilic, Fe-oxidizing, chemolithoautotrohic α- and γ- Proteobacteria from the deep sea. Appl Environ Microbiol. 2003;69:2906–2913. doi: 10.1128/AEM.69.5.2906-2913.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finneran KT, Housewright ME, Lovley DR. Multiple influences of nitrate on uranium solubility during bioremediation of uranium-contaminated subsurface sediments. Environ Microbiol. 2002;4:510–516. doi: 10.1046/j.1462-2920.2002.00317.x. [DOI] [PubMed] [Google Scholar]
- Gillis M, Logan NA. Genus IV. Chromobacterium Bergonzini 1881, 153AL. In: Brenner DJ, Kreig NR, Staley JT, editors. Bergey’s manual of systematic bacteriology. New York: Springer; 2005. pp. 824–827. [Google Scholar]
- Hafenbradl D, et al. Ferroglobus placidus gen. nov., sp. nov. a novel hyperthermophilic archaeum that oxidizes Fe2+ at neutral pH under anoxic conditions. Arch Microbiol. 1996;166:308–314. doi: 10.1007/s002030050388. [DOI] [PubMed] [Google Scholar]
- Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–98. [Google Scholar]
- He J, Zhang L, Jin S, Zhu Y, Liu F. Bacterial communities inside and surrounding soil iron-manganese nodules. Geomicrobiol J. 2008;25:14–24. [Google Scholar]
- Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001;17:754–755. doi: 10.1093/bioinformatics/17.8.754. [DOI] [PubMed] [Google Scholar]
- Hungate RE. A roll tube method for cultivation of strict anaerobes. Methods Microbiol. 1969;3B:117–132. [Google Scholar]
- Hungria M, et al. Genetic characterization of Chromobacterium isolates from black water environments in the Brazilian Amazon. Lett Appl Microbiol. 2005;41:17–23. doi: 10.1111/j.1472-765X.2005.01724.x. [DOI] [PubMed] [Google Scholar]
- Jiao YJ, Yongqin, Newman DN, Dianne K. The pio operon is essential for phototrophic Fe(II) oxidation in Rhodopseudomonas palustris TIE-1. J Bacteriol. 2007;189:1765–1773. doi: 10.1128/JB.00776-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Komagata K, Suzuki K. Lipid and cell-wall analysis in bacterial systematics. Methods Microbiol. 1987;19:161–208. [Google Scholar]
- Kwon S-W, Byung-Yong Kim W-GK, Kwan-Hee Yoo, Seung-Hee Yoo, Jung-A Son, Hang- Yeon Weon Paludibacterium yongneupense gen. nov., sp. nov., isolated from a wetland, Yongneup, in Korea. Int J Syst Evol Microbiol. 2008;58:190–194. doi: 10.1099/ijs.0.64831-0. [DOI] [PubMed] [Google Scholar]
- Lack JG, Chaudhuri SK, Chakraborty R, Achenbach LA, Coates JD. Anaerobic biooxidation of Fe(II) by Dechlorosoma suillum. Microb Ecol. 2002;43:424–431. doi: 10.1007/s00248-001-1061-1. [DOI] [PubMed] [Google Scholar]
- Lack JG, Chaudhuri SK, Kelly SD, Kemner KM, O'Connor SM, Coates JD. Immobilization of radionuclides and heavy metals through anaerobic bio-oxidation of Fe(II) Appl Environ Microbiol. 2002;68:2704–2710. doi: 10.1128/AEM.68.6.2704-2710.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau H-T, Faryna J, Triplett EW. Aquitalea magnusonii gen. nov., sp. nov., a novel Gram-negative bacterium isolated from a humic lake. Int. J. Syst. Evol. Microbiol. 2006;56:867–871. doi: 10.1099/ijs.0.64089-0. [DOI] [PubMed] [Google Scholar]
- Lin M-C, Chou J-H, Arun AB, Young C-C, Chen W-M. Pseudogulbenkiania subflava gen. nov., sp. nov., isolated from a cold spring. Int J Syst Evol Microbiol. 2008;58:2384–2388. doi: 10.1099/ijs.0.65755-0. [DOI] [PubMed] [Google Scholar]
- Martin PAW, Gundersen-Rindal D, Blackburn M, Buyer J. Chromobacterium subtsugae sp. nov., a betaproteobacterium toxic to Colorado potato beetle and other insect pests. Int J Syst Evol Microbiol. 2007;57:993–999. doi: 10.1099/ijs.0.64611-0. [DOI] [PubMed] [Google Scholar]
- Mesbah M, Premachandran U, Whitman W. Precise measurement of the G + C content of deoxyribonucleic acid by high performance liquid chromatography. Int J Syst Bacteriol. 1989;39:159–167. [Google Scholar]
- MIDI I (2001) Sherlock Microbial Identification System References
- Reith F, Lengke MF, Falconer D, Craw D, Southam G. Geomicrobiology of gold. ISME J. 2007;2007:567–584. doi: 10.1038/ismej.2007.75. [DOI] [PubMed] [Google Scholar]
- Roden EE, Lovely DR. Dissimilatroy Fe(III) Reduction by the marine microorganism Desulfuromonas acetoxidans. Appl Environ Microbiol. 1993;59:734–742. doi: 10.1128/aem.59.3.734-742.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheid D, Stubner S, Conrad R. Identification of rice root associated nitrate, sulfate and ferric iron reducing bacteria during root decomposition. FEMS Microbiol Ecol. 2004;50:101–110. doi: 10.1016/j.femsec.2004.06.001. [DOI] [PubMed] [Google Scholar]
- Sheloblina ES, VanPraagy CG, Lovley DR. Use of ferric and ferrous iron containing minerals for respiration by Desulfitobacterium frappieri. Geomicrobiol J. 2003;20:143–156. [Google Scholar]
- Singleton DR, Furlong MA, Peacock AD, White DC, Coleman DC, Whitman WB. Solirubrobacter pauli gen. nov., sp. nov., a mesophilic bacterium within the Rubrobacteridae related to common soil clones. Int J Syst Bacteriol. 2003;53:485–490. doi: 10.1099/ijs.0.02438-0. [DOI] [PubMed] [Google Scholar]
- Straub KL, Buchholz-Cleven BEE. Enumeration and detection of anaerobic ferrous iron- oxidizing, nitrate-reducing bacteria from diverse European sediments. Appl Environ Microbiol. 1998;64:4846–4856. doi: 10.1128/aem.64.12.4846-4856.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straub KL, Benz M, Schink B, Widdel F. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl Environ Microbiol. 1996;62:1458–1460. doi: 10.1128/aem.62.4.1458-1460.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straub KL, Rainey FA, Widdel F. Rhodovulum iodosum sp. nov, and Rhodovulum robiginosum sp. nov., two new marine phototrophic ferrous-iron-oxidizing purple bacteria. Int J Syst Bacteriol. 1999;49:729–735. doi: 10.1099/00207713-49-2-729. [DOI] [PubMed] [Google Scholar]
- Straub KL, Schonhuber W, Buchholz-Cleven B, Schink B. Diversity of ferrous iron- oxidizing, nitrate-reducing bacteria and their involvement in oxygen-independent iron cycling. Geomicrobiol J. 2004;21:371–378. [Google Scholar]
- Tamaki H, et al. Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques. Appl Environ Microbiol. 2005;71:2162–2169. doi: 10.1128/AEM.71.4.2162-2169.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomsen T, Kong Y, Nielsen P. Ecophysiology of abundant denitrifying bacteria in activated sludge. FEMS Microbiol Ecol. 2007;60:370–382. doi: 10.1111/j.1574-6941.2007.00309.x. [DOI] [PubMed] [Google Scholar]
- Tonjum T. Neisseriaceae Prevot 1933, 119AL emnd. Dewhirst, Paster and Bright 1989, 265. In: Brenner DJ, Kreig NR, Staley JT, editors. Bergey’s manual of systematic bacteriology. New York: Springer; 2005. pp. 775–776. [Google Scholar]
- Vasconcelos ATR, Almeida DF, Almeida FC, Almeida LGP, Almeida R, Alves-Gomes JA, Andrade, et al. The complete genome sequence of Chromobacterium violaceum reveals remarkable and exploitable bacterial adaptability. Proc Natl Acad Sci USA. 2003;100:11660–11665. doi: 10.1073/pnas.1832124100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaz-Moreira I, Nobre MF, Nunes OC, Manaia CM. Gulbenkiania mobilis gen. nov., sp. nov., isolated from treated municipal wastewater. Int J Syst Evol Microbiol. 2007;57:1108–1112. doi: 10.1099/ijs.0.64726-0. [DOI] [PubMed] [Google Scholar]
- Weber KA et al. (2009) Significance of microbially-mediated anaerobic oxidative dissolution of uraninite: a means of survival mobilizing a radionuclide (submitted)
- Weber KA, Picardal FW, Roden EE. Microbially catalyzed nitrate-dependent oxidation of biogenic solid-phase Fe(II) compounds. Environ Sci Technol. 2001;35:1644–1650. doi: 10.1021/es0016598. [DOI] [PubMed] [Google Scholar]
- Weber KA, Achenbach LA, Coates JD. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction. Nat Rev Microbiol. 2006;4:752–764. doi: 10.1038/nrmicro1490. [DOI] [PubMed] [Google Scholar]
- Weber KA, Pollock J, Cole KA, O'Connor SM, Achenbach LA, Coates JD. Anaerobic nitrate-dependent Iron(II) bio-oxidation by a novel, lithoautotrophic, Betaproteobacterium, strain 2002. Appl Environ Microbiol. 2006;72:686–694. doi: 10.1128/AEM.72.1.686-694.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber KA, Urrutia MM, Churchill PF, Kukkadapu RK, Roden EE. Anaerobic redox cycling of iron by freshwater sediment microorganisms. Environ Microbiol. 2006;8:100–113. doi: 10.1111/j.1462-2920.2005.00873.x. [DOI] [PubMed] [Google Scholar]
- Wetzel RG, Likens GE. Limnological analyses. New York: Springer; 1991. [Google Scholar]
- Widdel F, Schnell S, Heising S, Ehrenreich A, Assmus B, Schink B. Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature. 1993;362:834–836. [Google Scholar]