

Abstract
The present work assessed the effects of dietary ratios of essential fatty acids, arachidonic (ARA), eicosapentaenoic (EPA) and docosahexaenoic acid (DHA), on liver and intestine oxidative status, intestinal histomorphology and gut microbiota of gilthead sea bream. Four isoproteic and isolipidic plant-based diets were formulated containing a vegetable oil blend as the main lipid source. Diets were supplemented with ARA/EPA/DHA levels (%DM) equivalent to: 2%:0.2%:0.1% (Diet A); 1.0%:0.4%:0.4% (Diet B); 0%:0.6%:0.6% (Diet C); 0%:0.3%:1.5% (Diet D) and tested in triplicate groups for 56 days. Lipid peroxidation was higher in fish fed diets C and D while no differences were reported between diets regarding total, oxidized, and reduced glutathione, and oxidative stress index. Glutathione reductase was higher in fish fed diet A than diets C and D. No histological alterations were observed in the distal intestine. Lower microbiota diversity was observed in intestinal mucosa of fish fed diet C than A, while diets C and D enabled the proliferation of health-promoting bacteria from Bacteroidetes phylum (Asinibacterium sp.) and the absence of pathogenic species like Edwardsiella tarda. Overall, results suggest that a balance between dietary ARA/EPA + DHA promotes gilthead sea bream juveniles’ health however higher dietary content of n-3 LC-PUFA might limited the presence of microbial pathogens in intestinal mucosa.
Subject terms: Microbiology, Physiology, Zoology
Introduction
In marine fish species, dietary supply of linoleic (LA; 18:2n-6) and α-linolenic (ALA; 18:3n-3) acids cannot meet essential fatty acids (EFA) requirements due to the evolutionary loss or low capacity of long-chain polyunsaturated fatty acids (LC-PUFA) biosynthesis1–3. In gilthead sea bream (Sparus aurata), the leading finfish species in Mediterranean aquaculture, the limited capacity to synthesize LC-PUFAs from C-18 precursors is related to low activities of the Δ5 fatty acid desaturase and elongase 23,4. Thus, dietary supply of the main LC-PUFA, arachidonic acid (ARA; 20:4n-6), eicosapentaenoic acid (EPA; 20:5n-3), and docosahexaenoic acid (DHA; 20:6 n-3) is necessary to fulfill the physiological requirements of EFA of this species3,5.
For gilthead sea bream juveniles, total dietary EPA + DHA requirements vary with lipid level and DHA/EPA ratio and were estimated to be 0.9 or 1.9% with a DHA/EPA ratio of 1 or 0.5, respectively6–8. However individual EPA and DHA requirements have been reported to be 0.7 and 0.6% of dry matter (DM), respectively (reviewed by9). As for ARA, the only available study indicates that increasing the dietary levels (0, 0.6, 1.14, 1.7%DM) has no effect on growth performance10. However, in European sea bass juveniles, dietary ARA levels below 0.2% reduced growth performance11.
In marine aquafeeds, the main source of LC-PUFA has been fish oil (FO) but, due to its stagnant availability in the world market, its dietary replacement by alternative lipid sources is required for promoting further growth of the marine aquaculture industry.
Thus, in recent years, research focused on FO replacement by more available and sustainable oil sources, such as vegetable oils (VO)12–15, land animal oils13,16–18, or microalgae oils19–21. However, most of the commercially available alternative oil sources, namely VO and animal oils, are deficient in LC-PUFA and its dietary inclusion may jeopardize normal fish growth, health, intestinal histomorphology, and microbiota community5,22–24.
LC-PUFAs are highly susceptible to peroxidation, generating reactive oxygen species (ROS) causing tissue oxidative damage14,25,26.
ROS induce damage to lipids, proteins, carbohydrates, and nucleic acids, with the potential to induce cellular death and compromising tissue functionality and palatability of the final product14,25. LC-PUFA oxidation can compromise membrane structure, by decreasing its fluidity and increasing its permeability to harmful substances, with potential to be pathological to the cell or tissue27. The balance between generation and clearance of ROS is maintained through the action of the antioxidant system, involving radical scavenging enzymes (catalase, CAT; superoxide dismutase, SOD; glutathione reductase, GR; and glutathione peroxidase, GPX) and non-enzymatic antioxidants (Vitamin A, C, and E; glutathione, GSH; bilirubin and flavonoids)25,28. However, this balance may be highly affected by diet composition, including the level of LC-PUFA. For instance, increasing dietary levels of n-3 LC-PUFA were associated with increased liver and plasma oxidative stress of silvery black porgy (Sparidentex hasta)29. Also, high dietary content of n-3 LC-PUFA increased oxidative stress in the muscle of rainbow trout, Oncorhynchus mykiss30, the intestine of European sea bass (Dicentrarchus labrax)31 and liver and intestine of gilthead sea bream14. Increasing levels of ARA were also shown to increase liver oxidative stress in Synechogobius hasta juveniles32 and Japanese eel serum (Anguilla japonica)33.
Dietary DHA/EPA ratio also seems to affect oxidative stress, apparently in a species-specific manner. For instance, higher ratios caused increased oxidative damage in serum of black sea bream (Acanthopagrus schlegelii) while the opposite was observed in the plasma of silvery-black porgy juveniles2,34. In an in vitro study, it was shown that increasing concentrations of EPA, but not of DHA, increased lipid peroxidation levels in yellow croaker (Larmichthys crocea) macrophages35. Further, while EPA increased superoxide anion production the opposite was observed for DHA. Thus, EPA and DHA seem to have different capacities for modulating redox mechanisms.
Intestinal microbiota is vital to promote fish physiological balance and health, and its effects on fish feeding, digestion, metabolism, immunity, and disease resistance have been highlighted24,36,37. Fish intestinal microbiota is modulated by several factors including host genetic background, living environment, section of the gastrointestinal tract, stress, and diet24,36,37. Intestinal microbiota seems to be particularly sensitive to diet composition12,24,38,39. However, the impact of dietary PUFA, particularly dietary LC-PUFAs, on gut microbiota has been poorly explored booth in humans and aquatic species12,24,40.
Previously, Magalhães et al.41 showed that a diet with a balanced ARA/EPA/DHA (1/0.4/0.4) ratio that closely matches the estimated EFA requirements of gilthead seabream increased feed efficiency and protein efficiency ratio compared to diets with unbalanced ARA/EPA/DHA ratios (0/0.6/0.6 and 0/0.3/1.5). Dietary ARA/EPA/DHA ratios also affected lipid metabolism, with increased β-oxidation activity in fish fed high ARA/EPA/DHA (2.0/0.2/0.1) and decreased lipogenesis in fish fed high DHA (0/0.3/1.5). Furthermore, liver lipid content was higher in fish fed high dietary EPA and DHA levels.
The objective of this study was to further evaluate dietary ARA/EPA/DHA ratios of the fore mentioned diets on oxidative stress in target organs (liver and intestine), distal intestine histomorphology, and gut microbiota composition of gilthead sea bream juveniles. Therefore, the importance of dietary ARA or n-3 LC-PUFA on these indicators of wellbeing was compared to the EFA balanced diet.
Results
The results of the feeding trial are presented elsewhere41 as it was not the aim of this study.
Oxidative stress
Oxidative stress indicators, namely liver and intestine enzymatic activities and glutathione responded similarly to the diet treatments, as no organ × diet interaction in any parameter analyzed was observed (Tables 1 and 2).
Table 1.
Liver and intestine total glutathione (tGSH), oxidized glutathione (GSSG), reduced glutathione (GSH), oxidative stress index (OSI) and lipid peroxidation (LPO) levels of gilthead sea bream fed the experimental diets.
| Organ | Liver | Intestine | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diets | A | B | C | D | SEM | A | B | C | D | SEM |
| 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | |||
| tGSH | 1017 | 935 | 928 | 739 | 51.8 | 947 | 797 | 884 | 788 | 42.8 |
| GSSG | 39 | 29 | 36 | 28 | 3.8 | 111 | 114 | 135 | 133 | 7.5 |
| GSH | 977 | 906 | 893 | 711 | 49.0 | 837 | 675 | 748 | 655 | 42.3 |
| OSI1 | 7.19 | 6.12 | 7.25 | 7.00 | 0.5 | 23.7 | 27.7 | 33.9 | 34.3 | 2.2 |
| LPO | 13.5 | 14.9 | 18.7 | 19.5 | 0.8 | 69.6 | 62.6 | 90.6 | 79.2 | 3.0 |
| Variance source | Diets | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Organ | Interaction | A | B | C | D | ||||
| Two-way Anova | ||||||||||
| tGSH | Ns | Ns | ns | – | – | – | – | |||
| GSSG | Ns | 0.000 | ns | – | – | – | – | |||
| GSH | Ns | 0.043 | ns | – | – | – | – | |||
| OSI1 | Ns | 0.000 | ns | – | – | – | – | |||
| LPO | 0.000 | 0.000 | ns | a | a | b | b | |||
Values presented as means (n = 9 for liver and n = 6 for intestine)) and pooled standard error of the mean (SEM). LPO values expressed as nmols MDA g−1 tissue and GSH, tGSH, and GSSG as nmol g−1 tissue
Two-way ANOVA: ns: non-significant (P > 0.05).
1OSI = 100 × (2 × GSSG/tGSH).
Table 2.
Liver and intestine antioxidant enzymes activity of gilthead sea bream fed the experimental diets.
| Organ | Liver | Intestine | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diets | A | B | C | D | SEM | A | B | C | D | SEM |
| 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | |||
| CAT | 56.7 | 51.8 | 42.1 | 48.5 | 1.68 | 153.8 | 92.4 | 76.6 | 158.6 | 16.0 |
| G6PDH | 136.6 | 132.1 | 127.8 | 110.0 | 5.25 | 24.6 | 25.8 | 31.0 | 20.2 | 1.66 |
| GR | 8.8 | 7.3 | 7.0 | 6.6 | 0.28 | 20.7 | 15.9 | 14.9 | 16.1 | 0.96 |
| SOD | 106.9 | 126.9 | 110.3 | 108.5 | 6.59 | 1,341.4 | 1,220.3 | 1,232.5 | 929.0 | 63.7 |
| GPX | 54.2 | 36.4 | 38.5 | 35.5 | 3.22 | 10.2 | 9.6 | 9.6 | 9.0 | 0.64 |
| Variance source | Diets | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Organ | Interaction | A | B | C | D | ||||
| Two-way Anova | ||||||||||
| CAT | 0.022 | 0.000 | ns | – | – | – | – | |||
| G6PDH | Ns | 0.000 | ns | – | – | – | – | |||
| GR | 0.008 | 0.000 | ns | b | ab | a | a | |||
| SOD | Ns | 0.000 | ns | – | – | – | – | |||
| GPX | Ns | 0.000 | ns | – | – | – | – | |||
Values presented as means (n = 9 for liver and n = 6 for intestine) and pooled standard error of the mean (SEM). Enzyme activities expressed as mU mg protein−1 for G6PDH, GR, and GPX and as U mg protein−1 for CAT and SOD
Two-way ANOVA: ns: non-significant (P ≥ 0.05).
Liver and intestinal glutathione and oxidative stress index (OSI) were not affected by dietary treatments (Table 1). Oxidized glutathione (GSSG) and OSI values were higher while reduced glutathione (GSH) was lower in the intestine compared to liver. However, total glutathione (tGSH) was similar in both tissues. Further, LPO was higher in the intestine than in the liver and in fish fed diets C and D than diets A and B (Table 1).
Regarding antioxidant enzymes, no organ × diet interaction was observed (Table 2). G6PDH and GPX activities were higher in the liver than in the intestine, while the opposite was true for CAT, GR, and SOD activities. Regarding dietary effects,
GR activity was higher in fish fed diet A than diets C and D, whereas G6PDH, SOD, and GPX activities were unaffected by dietary treatments. For CAT, although Anova indicates a significant dietary effect, that effect was not detected by the Tukey’s multiple range test.
Distal intestine histological evaluation
Distal intestine histomorphology parameters analyzed, namely enterocyte vacuolization, height of mucosal folds, width of lamina propria and leukocyte infiltration of the lamina propria and submucosa were not affected by dietary treatments (Table 3; Fig. 1).
Table 3.
Score-based histological evaluation of the distal intestine in gilthead sea bream juveniles fed the experimental diets Values presented as means (n = 6) and pooled standard error of the mean (SEM).
| Diets | A | B | C | D | SEM |
|---|---|---|---|---|---|
| 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | ||
| Fold height | 2.7 | 1.8 | 2.3 | 2.5 | 0.13 |
| Lamina propria | 2.5 | 2.2 | 1.7 | 2.7 | 0.17 |
| Submucosa | 1.0 | 1.0 | 1.0 | 1.0 | 0.00 |
| Intraepithelial leucocytes | 1.8 | 2.2 | 2.7 | 2.3 | 0.17 |
| Enterocytes | 1.5 | 2.0 | 1.7 | 2.8 | 0.26 |
| Mean score | 1.9 | 1.8 | 1.9 | 2.3 | 0.08 |
Absence of superscript letters indicates no significant differences between dietary treatments (P > 0.05).
Figure 1.
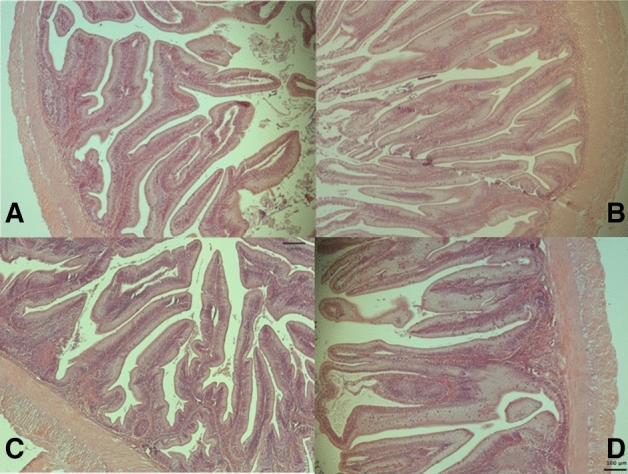
Histomorphology features of distal intestine of gilthead sea bream juveniles fed the experimental diets (A–D) recorded with Zen software (Blue edition). Normal height of mucosal folds, normal width lamina propria with normal leukocyte infiltration of the lamina propria and submucosa. No signs of inflammation. H-E staining.
Intestinal microbiota
Number of OTU’s, species richness, in both digesta and mucosa, and digesta diversity were not affected by dietary treatments, except for species diversity in the intestinal mucosa of fish fed diet C which was lower than in fish fed diet A (Table 4).
Table 4.
Ecological parameters obtained from PCR-DGGE fingerprints of the intestinal allochthonous (digesta) and autochthonous (mucosa) microbiota of gilthead sea bream fed with the experimental diets.
| Diets | Digesta | Mucosa | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | SEM | A | B | C | D | SEM | |
| 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | 2.0/0.2/0.1 | 1.0/0.4/0.4 | 0/0.6/0.6 | 0/0.3/1.5 | |||
| OTUs1 | 21.0 | 22.0 | 17.7 | 20.7 | 0.64 | 28.7 | 25.0 | 22.0 | 24.0 | 1.07 |
| Richness2 | 1.2 | 1.3 | 1.0 | 1.2 | 0.04 | 1.7 | 1.5 | 1.3 | 1.4 | 0.06 |
| Diversity3 | 2.9 | 3.0 | 2.8 | 2.9 | 0.03 | 3.3b | 3.2ab | 3.0a | 3.1ab | 0.05 |
| Similarity (%)4 | 80.7 | 85.8 | 77.8 | 85.4 | 1.37 | 83.9 | 76.6 | 73.2 | 83.5 | 2.31 |
Values presented as means (n = 3) and pooled standard error of the mean (SEM). Different superscript letters indicate significant differences between dietary treatments (P < 0.05).
1OTUs: Average number of operational taxonomic units.
2Margalef species richness: d = (S − 1)/log(N) where S is the number of species, and N is the total number of individuals in the sample.
3Shannons diversity index: H’ = − ∑(pi(lnpi)) where pi is the proportion of individuals belonging to the ith species present in the sample.
4SIMPER, similarity percentage within group replicates.
The Bray–Curtis dendrogram showed that for digesta samples the three dietary replicates failed to cluster (Fig. 2A). For mucosa samples, dendrogram showed that diets with and without ARA supplementation cluster separately. A clearer visualization of this result can be seen in the MDS plot of DGGE bands (Fig. 2B).
Figure 2.

A Dendrograms and PCR-DGGE fingerprints of the allochthonous (digesta) and autochthonous (mucosa) intestinal microbiota of gilthead sea bream fed the experimental diets. Numbers (1 to 13 on digesta; 1 to 22 on mucosa) indicate bands excised for sequence analysis, identified on Table 5. B Multidimensional scaling (MDS) plot of DGGE bands presence and abundances in intestinal mucosa samples from gilthead sea bream juveniles fed the same experimental diets depicted in A: Diet A ( ); Diet B (
); Diet B ( ); Diet C (
); Diet C ( ); Diet D (
); Diet D ( ). Dotted lines represent 65% similarity between samples.
). Dotted lines represent 65% similarity between samples.
Identification of the selected DGGE bands from the digesta and mucosa microbiota is shown in Table 5. Sequence analysis of 35 DGGE bands showed 20 sequences below 97% of sequence similarity, indicating a poor resolution for species identification. However, at the genus level identification, sequence analysis revealed that the dominant allochthonous (digesta) bacterial genus belonged to the Proteobacteria phylum, with three bands of Klebsiella genus detected, two bands of Pantoea, and one of Nitrosospira. Three bands of Lactobacillus (phylum Firmicutes) were also identified.
Table 5.
Closest relatives (BLAST) to the sequenced PCR-DGGE gel bands (from Fig. 2) of the intestinal communities of gilthead sea bream fed the experimental diets.
| Band | Closest known species (BLAST) | Phylum | ID (%) | Accession nr |
|---|---|---|---|---|
| Digesta | ||||
| 1 | Lactobacillus delbrueckii | Firmicutes | 89 | MF784102.1 |
| 2 | Lactobacillus aviarius | Firmicutes | 99 | LC071826.1 |
| 3 | Lactobacillus helveticus | Firmicutes | 99 | KX951719.1 |
| 4 | Pantoea sp. | Proteobacteria | 86 | KR029059.1 |
| 5 | Klebsiella pneumoniae | Proteobacteria | 100 | MG923524.1 |
| 6 | Klebsiella sp. | Proteobacteria | 98 | KT301470.1 |
| 7 | Uncultured bacterium | n/a | 97 | LC031369.1 |
| 8 | Uncultured bacterium clone OTU5449 | n/a | 84 | KT788917.1 |
| 9 | Pantoea sp. | Proteobacteria | 90 | KR029059.1 |
| 10 | Klebsiella pneumoniae | Proteobacteria | 89 | MG923524.1 |
| 11 | Nitrosospira sp. | Proteobacteria | 79 | AY293079.1 |
| 12 | Uncultured bacterium clone ac-25 16S rRNA | n/a | 92 | KY857639.1 |
| 13 | Klebsiella pneumoniae | Proteobacteria | 96 | MH266241.1 |
| Mucosa | ||||
| 1 | Uncultured bacterium clone SHFH766 | n/a | 94 | KT981153.1 |
| 2 | Asinibacterium sp. | Bacteroidetes | 96 | KP301113.1 |
| 3 | Lactobacillus helveticus | Firmicutes | 98 | KX951719.1 |
| 4 | Pseudomonas veronii | Proteobacteria | 96 | Kt302366.1 |
| 5 | Thalassomonas sp. | Proteobacteria | 90 | MG819691.1 |
| 6 | Klebsiella pneumoniae | Proteobacteria | 99 | MH150802.1 |
| 7 | Klebsiella pneumoniae | Proteobacteria | 97 | KU550763.1 |
| 8 | Janthinobacterium sp. | Proteobacteria | 91 | AB196254.1 |
| 9 | Bacillus sp. | Firmicutes | 98 | FJ195793.1 |
| 10 | Thiohalophilus sp. | Proteobacteria | 81 | KX599116.1 |
| 11 | Massilia sp. | Proteobacteria | 97 | KY912083.1 |
| 12 | Massilia sp. | Proteobacteria | 93 | FN386762.1 |
| 13 | Edwardsiella tarda | Proteobacteria | 96 | MF034724.1 |
| 14 | Bacillus sp. | Firmicutes | 78 | JX191913.1 |
| 15 | Ralstonia sp. | Proteobacteria | 95 | MG238553.1 |
| 16 | Klebsiella sp. | Proteobacteria | 86 | KT300944.1 |
| 17 | Planktothricoides sp. | Cyanobacteria | 95 | JX628815.1 |
| 18 | Bacillus halodurans | Firmicutes | 98 | MH299852.1 |
| 19 | Klebsiella pneumoniae | Proteobacteria | 99 | MH266241.1 |
| 20 | Klebsiella pneumoniae | Proteobacteria | 98 | MH266241.1 |
| 21 | Klebsiella pneumoniae | Proteobacteria | 100 | CP029226.1 |
| 22 | Klebsiella variicola | Proteobacteria | 100 | KR092086.1 |
Autochthonous (mucosa) bacteria analysis revealed one genus of the phylum Bacteroidetes (Asinibacterium sp.) and 4 bands of the phylum Firmicutes (1 Lactobacillus and 3 bands of the genus Bacillus). Nevertheless, the predominant phylum present in the mucosa was Proteobacteria with 15 bands identified belonging to the Pseudomonas, Thalassomonas, Klebsiella, Janthinobacterium, Thiohalophilus, Massila, Edwardsiella and, Ralstonia genera (Table 5).
Discussion
The diets in the presented study had different n6/n3 EFA ratios as well as total n-6 and n-3 LC-PUFA content. Thus, Diet A was rich in ARA and had low EPA and DHA levels; diet B had balanced ARA, EPA and DHA levels; diet C had no ARA and was balanced in EPA and DHA; Diet D had no ARA and had high levels of DHA than EPA.
It is well known that susceptibility of fatty acids (FA) to peroxidation increases with unsaturation, as the weak carbon-hydrogen bond of the bis-allylic methylene groups is particularly prone to oxidation25. In the present study, liver and intestine TBARS levels, a well-accepted physiological index of tissue LPO, were inversely related to the dietary n-6/n-3 ratio (diets C and D) and this may be related to increased levels of n-3 LC-PUFA in these tissues. Indeed, a positive correlation between LC-PUFA content in different tissues and lipid peroxidation was already demonstrated in several fish species14,29,31,42. Even though liver and intestine FA profiles were not determined in the present study, muscle EPA and particularly DHA contents were higher in fish fed diets C and D than the other diets, while ARA content showed only slight variations41 and it is expected that the FA concentrations in liver and intestine should follow a similar pattern. Therefore, a lower concentration of EPA and DHA might explain the lower LPO values in ARA rich diets. Previously it was already observed that the degree of unsaturation of dietary FA correlates with the FA unsaturation in tissues43,44 as well as its susceptibility to oxidation14,45.
The higher LPO content appeared to be related to elevated contents of DHA in diets C and D. One reason for this may relate to the tendency of DHA to accumulate in mitochondrial cardiolipin that would impair electron transport efficiency and increasing ROS production46. The fact that ARA and EPA are not incorporated into mitochondrial cardiolipin may explain the lower LPO content in fish fed diets A and B46.
The primary defense against oxidative stress includes antioxidant enzymes such as GR, SOD, CAT, and GPX25. GR is essential for maintenance of GSH level by reducing GSSG. The activity was higher in fish fed the high ARA, diet A, followed by diet B, while CAT, SOD and GPX activities were not affected in this experiment. This suggests that the glutathione system was important in the modulation of the redox environment of fish in relation to the dietary EFA composition. This is also supported by tGSH and GSH levels, which were higher (though not statistically higher), and the LPO levels, which were lower, in fish fed diets A and B than the other diets. Similar to the present results, other authors have also reported that higher dietary ARA levels improved the response of antioxidant enzymes in fish32,33,47.
As already reported in gilthead sea bream juveniles48,49 the antioxidant defense mechanism responded differently in liver and intestine. CAT, GR, SOD, and GSSG were higher in the intestine, while G6PDH and GPX activities were higher in the liver. Overall, LPO values were also considerably higher in the intestine than in the liver, which agrees with previously published data48. Higher intestine LPO and OSI values were expected due to the high enterocyte turnover rate that increases susceptibility to oxidation.
In the present study, the dietary effect on intestine histomorphology was accessed in the distal section as previous studies showed that this intestine portion is more sensitive to dietary treatments than the anterior and mid-intestine50,51. However, distal intestine histomorphology was not affected by the different dietary n-6/n-3 LC-PUFA levels used in the experimental diets. This confirms previous studies on mid- or distal- intestines of European seabass fed diets with 60–70% of VO blends52,53. Also, gilthead sea bream juveniles and Atlantic salmon fed diets with wild type or genetically modified Camelina oil (richer in ARA, EPA, and DHA) showed no differences in distal intestine morphology54–56. On the contrary, an accumulation of enterocytes lipid droplets with dietary VO incorporation, leading to lower dietary content of ARA, EPA, and DHA was reported in gilthead sea bream juveniles57.
The dietary ARA, DHA, and EPA ratios used in this study, altered gilthead sea bream intestinal microbiota at the mucosal level. The MDS plot and the Bray–Curtis dendrogram representation of mucosal data showed a higher similarity between microbial communities of fish fed diets with elevated levels of EPA/DHA(diets C and D) than within fish fed diets with high ARA content (diets A and B). This shows the potential of dietary fatty acids to select for bacterial species affecting attachment sites of the intestinal mucosa resulting in establishment of different microbial communities24. In fact, the isolation of the genus Thalassomonas spp., Janthinobacterium spp., Thiohalophilus spp. and Massilia spp. were only isolated in diets A and B. Also, diet C contained 1.2% of EPA + DHA showed lower microbial diversity (Shannon index) at the intestinal mucosa than the other diets. Previous studies reported that 58% and 84% substitution of dietary FO by VO lowered Shannon’s diversity index in gilthead sea bream juveniles59. Similarly, other PUFA, such as 2.5% of LA (18:2n-6), were also shown to reduce the culturable intestinal microbiota diversity of Arctic charr (Salvelinus alpinus)60. On the other hand, the dietary inclusion of 0.5% 1-monoglycerides of short- and medium-chain FA (from C3 to C12) did not influence the same diversity index in gilthead sea bream juveniles61. The mechanism for dietary PUFA to reduce fish gut microbiota is not fully understood. It is however interesting to note that reduced bacterial diversity also seems to occur in humans with increase intake of n-3 LC-PUFA40. Here the reduction of gut microbial diversity was accompanied by an increase of potential butyrate-producing bacterial species (e.g., Asinibacterium sp.), a short-chain fatty acid (SCFA) known to promote gastrointestinal health and host immunity40. Accordingly, in Arctic charr fed casein-based diets supplemented with LA, ALA (18:3n-3), or EPA and DHA, the frequency of lactic acid bacteria (LAB, commonly used as probiotics and associated with benefits to host), was higher in diets supplemented with 7% of ALA or 4% of EPA + DHA58. Despite this, in our study, the main SCFA-producing species such as Bacillus spp. and Lactobacillus spp. (LAB)62 were present in both digesta and mucosa samples without significant changes between diets.
As reported for other fish species36, Proteobacteria and Firmicutes were the dominant phyla in both digesta and mucosa microbiota of gilthead sea bream juveniles in this study, and appear to be predominant in many fish species regardless of dietary treatment59,61,63–65. While digesta microbial community is considered transient, mucosal community is assumed to be more stable36. The colonization of gut mucosal epithelium is essential for establishing health-promoting bacterial species as autochthonous microbiota and, as consequence, reduce the establishment of opportunistic bacteria36,37. The presence of pathogenic species such as Klebsiella spp.66, despite the different intensity of the bands, was equally distributed among all dietary treatments, and previously isolated from different fish species, suggesting that these microorganisms are part of normal fish microbiota67–69 or an opportunistic pathogen attacking stressed animals70.
The isolation of one bacterial species from Bacteroidetes phylum (Asinibacterium sp.) in diets C and D might highlight the influence of n-3 LC-PUFA in autochthonous microbiota. Although Asinibacterium sp. is a recently described genus71 whose metabolic potential is not yet totally known, bacteria from Bacteroidetes phylum are also (adding to the Firmicutes, in particular Bacillus spp. and Lactobacillus spp.62), related to the fermentation of dietary non-starch polysaccharides, starch, and sugars into SCFAs as acetate and propionate72. These SCFA are absorbed and used as energy sources by the enterocytes and have anti-inflammatory and bactericidal activities61. In fact, the isolation of a fish-pathogen such as Edwardsiella tarda, causative agent of edwardsiellosis with severe losses in aquaculture73, in fish fed with lower levels of n-3 LC-PUFA (diets A and B) might be another indication of the positive effect n-3 LCPUFA had on gilthead sea bream mucosal microbiota.
Increasing bactericidal activity was already associated with long carbon chain length and degree of unsaturation of dietary FA58,74. Thus, the higher content of n-3 LC-PUFA might have limited the presence of pathogens such as E. tarda in the mucosal microbial community of fish fed diets C and D. Accordingly, a very recent study with golden pompano (Trachinotus ovatus) showed that FO substitution by VO increased the abundance of intestinal pathogenic bacteria such as Mycoplasma and Vibrio75. As recently reviewed76, gut microbial community, host immune system, and dietary n3-PUFA are interdependent pieces that together control intestinal wall integrity, and thus pathogen proliferation. Furthermore, the host-derived ROS was previously associated with bactericidal activity against pathogens such as Salmonella in mice77. Also, ROS was associated with gut epithelial response to microbial signals and stimulating immune responses against bacteria78. Thus, more studies are required to confirm if increasing ROS production may impair pathogenic bacterial proliferation as it was reported in the present study for fish fed diets C and D.
We report significant modifications in the oxidative status and mucosa microbiota with different dietary n-6/n-3 LC-PUFA content, revealing the importance of correct EFA ratios for gilthead sea bream health. This is especially important with increasing VO incorporation in aquafeeds. The impairment of mucosa colonization by pathogenic bacteria reduces the risk of fish infection and consequently the need for antibiotics utilization, leading to a better image of fish produced in aquaculture to the consumers.
Material and methods
Diets composition
Four plant-based diets were formulated as described in Magalhães et al.41. Shortly, the experimental diets were isoproteic (47% crude protein; 74% protein from plant feedstuffs and 26% protein from fish meal) and isolipidic (18% crude lipids) with a vegetable oil blend (20:50:30 rapeseed, linseed, and palm oils) as main lipid source. Mortierella alpine oil (Vevodar, DSM Food Specialties, the Netherlands); krill oil (Euphausia superba; SuperbaKrill Oil, Solchem); and tuna oil (70% DHA; BrudyTechnology) were used to adjust the ARA, EPA, and DHA levels of the diets, to obtain a final ARA/EPA/DHA level of 2%:0.2%:0.1% (Diet A); 1.0%:0.4%:0.4% (Diet B); 0%:0.6%:0.6% (Diet C); 0%:0.3%:1.5% (Diet D). The fatty acids profile of the oils was confirmed before diets were formulated. Ingredients, proximate composition and fatty acids composition of the experimental diets are presented in supplementary file 1—Table S1, and dietary FA composition is presented in additional file 1—Table S2.
Experimental trial
The growth trial was conducted in CIIMAR, Matosinhos, Portugal. Gilthead sea bream (Sparus aurata) juveniles were obtained from a commercial fish farm (Maresa S.A., Ayamonte, Huelva, Spain). Fish were moved to the experimental system after a quarantine period of 1 month and allowed to adapt to the experimental conditions for 15 days. The trial was conducted in a thermo-regulated recirculating marine water system (23.0 ± 1.0 °C; 35 ± 1 g L−1 salinity; 7 mg L−1 oxygen) equipped with 12 cylindrical fiberglass tanks. Tanks (100 L water capacity) were supplied with filtered seawater (flow of 2.5–3.5 L min−1) and kept under controlled photoperiod (12:12 h of light:dark). At the beginning of the experiment, a total of 240 gilthead sea bream juveniles with an initial mean body weight of 15 g were utilized and groups of 20 fish were distributed into the 12 experimental tanks. Each experimental diet was randomly assigned to triplicate groups. The trial was carried on during 56 days and fish were fed by hand, until apparent visual satiation, two times a day, 6 days a week. Extreme care was taken to minimize feed waste.
Sampling
At the end of the growth trial, fish were randomly sampled 4 h after the morning meal, euthanized by decapitation and dissected on chilled trays. The adjacent adipose and connective tissues of the intestine was removed from 2 fish per tank, and a small portion of the distal intestine (DI, differentiated by a darker mucosa and enlarged diameter from the mid intestine) was collected for histomorphology assessment. Therefore, DI samples were cleaned in phosphate-buffered saline (PBS), carefully absorbed with a paper towel and promptly fixed in phosphate-buffered formalin (4%, pH 7.4) for 24 h and immediately changed to ethanol (70%) until further processing according to Couto et al.51. The rest of the intestine was stored at − 80 °C until quantification of oxidative stress enzymatic and non-enzymatic indicators and lipid peroxidation levels. Liver from 3 fish was also sampled for the same purpose. Two other fish per tank were sampled under aseptic conditions (working with an open flame using sterilized solutions, collecting tubes and tools) for allochthonous (digesta) and autochthonous (mucosa) microbiota characterization. Mucosa samples were collected by scraping the internal intestinal mucosal surface, after opening the intestine in its longitudinal axis. Digesta samples were obtained by squeezing the entire intestinal content into a sterile tube. Samples were instantly frozen with liquid nitrogen and stored at − 80 °C until analyzed.
Chemical analysis
The Association of Official Analytical Chemists methods AOAC79 were used to perform the chemical analysis of experimental diets. Dietary starch was quantified as described by Beutler80; The Folch et al.81 method was utilized for total lipids determination using dichloromethane instead of chloroform. FA methyl esters of oil ingredients and diets were prepared by transmethylation of total lipids extract and analyzed by chromatography using Shimadzu GC-2010 Plus gas chromatograph (Shimadzu Europe GmbH, Germany) equipped with a flame-ionization detector (GC-FID) and an Omegawax 250 capillary column (30 m × 0.25 mm i.d. × 0.25 µm film thickness; Supelco, Bellefonte, USA). FA were identified by comparison with known standard mixtures as described by Magalhães et al.41.
Enzyme activity
Liver and intestine samples were homogenized (dilution 1:7 and 1:5, respectively) in ice-cold 100 mM Tris–HCl buffer, containing 0.1 mM EDTA and 0.1% (v/v) Triton X-100, pH 7.8. All the procedures were performed on ice. Homogenates were centrifuged at 30,000×g for 30 min at 4 ºC and supernatants were divided into aliquots and stored at − 80 ºC until use. All assays were carried out at 37 °C in a Multiskan GO microplate reader (Model 5111 9200; Thermo Scientific, Nanjing, China). The specific assay conditions for each enzyme were as follows:
Glucose-6-phosphate dehydrogenase (G6PDH; EC 1.1.1.49) activity was analyzed by measuring the reduction of NADP+ 82.
Superoxide dismutase (SOD; EC 1.15.1.1) activity was measured by the ferricytochrome method using xanthine/xanthine oxidase as the source of superoxide radicals83. One unit of activity was defined as the amount of enzyme necessary to produce a 50% inhibition of the ferricytochrome c reduction rate.
Catalase (CAT; EC 1.11.1.6) activity was assessed by measuring the decrease of hydrogen peroxide concentration84.
Glutathione peroxidase (GPX; EC 1.11.1.9) activity was determined by measuring the NADPH consumption rate generated by the oxidized glutathione (GSSG) produced by GPX activity and reduced by glutathione reductase (GR)85.
Glutathione reductase (GR; EC 1.6.4.2) activity was analyzed by measuring the oxidation of NADPH, according to Morales et al.86.
Soluble protein concentration was determined according to Bradford87 using Sigma-Aldrich protein assay kit and bovine serum albumin as standard.
Except for SOD and CAT which are expressed as units per mg of soluble protein, the activities of the other enzymes are expressed as milliunits per mg of soluble protein. One unit of the enzyme was defined as the amount of enzyme required to transform 1 mmol of substrate per min under the assay conditions.
Lipid peroxidation
Malondialdehyde (MDA) concentration was used as a marker of lipid peroxidation (LPO) level in the liver and intestine following the methodology described by Buege and Aust88. In the presence of thiobarbituric acid, MDA reacts producing colored thiobarbituric acid reacting substances (TBARS) that were measured using a spectrophotometer at 535 nm. Results were calculated from an MDA calibration curve.
Total and oxidized glutathione
Liver and intestine samples were homogenized (1:10 and 1:5, respectively) in ice-cold solution containing 1.3% 5-sulfosalicylic acid (w/v) and 10 mM HCl, and the whole procedure was done in ice to avoid glutathione oxidation. Homogenates were centrifuged at 14,000g for 10 min at 4 °C and the supernatants stored at − 80 °C. Total glutathione (tGSH) and oxidized glutathione (GSSG) were determined accordingly Griffith89and Vandeputte et al.90 with some modifications according to Castro et al.14. Standard curves of reduced glutathione (GSH) and GSSG were used for tGSH and GSSG calculations, respectively. GSH level was calculated by subtracting GSSG from tGSH values.
Histological processing and morphological evaluation
Distal intestine samples were processed and sectioned using standard histological techniques and stained with hematoxylin and eosin. Blind evaluation of histological preparations was performed, analyzing mucosal folds height, width, and cellularity of the lamina propria and submucosa, number of intraepithelial lymphocytes, nucleus position size, and variation of enterocyte vacuolization91,92. A scale scoring system ranging from 1 (normal) to 5 (highly modified) was used as described in Penn et al.93. The overall score of histomorphology alterations was calculated by averaging scores of all the parameters. Images were acquired with Zen software (Blue edition; Zeiss, Jena, Germany).
Microbial diversity
Samples of 2 fish per tank were pooled to reduce variability. The extraction of bacterial DNA from fish intestinal digesta and mucosa was done by weighting around 300 mg of sample to a 2 mL bead-beater (Sigma-Aldrich, Buchs, Switzerland) tube previously prepared with 500 μL STE buffer (0.1 M NaCl, 10 mM Tris, 1 mM EDTA, pH 8) and 0.4 g of glass beads (Sigma-Aldrich G8772). Samples were then homogenized twice for 30 s in the BeadBug bead-beater (Benchmark Scientific, Edison, NJ, USA) at 2500 speed with an interval of at least 30 s on ice. Following 15 min incubation at 75 °C, with gentle agitation every 5 min, tubes were centrifuged for 1 min at 13,000g and 500 μL of supernatant was transferred to new sterile 2 mL microcentrifuge tubes. From this point, the protocol used for bacterial DNA extraction was based on the method of Pitcher et al.94.
Bacterial 16S rRNA gene fragments were amplified using a touchdown PCR on a T100Thermal Cycler (Bio-Rad Laboratories Lda., Amadora, Portugal), using oligonucleotide primers 16S-358F (which contained a GC clamp at the 5′ end) and 16S-517R95. 300 ng of each PCR product was resolved on 8% polyacrylamide gel composed by a denaturing gradient of 40–60% 7 M urea/40% formamide. DGGE was performed using a DCode universal mutation detection system (Bio-Rad Laboratories Lda.) during 16 h at 60 °C, 65 V in 1 × TAE buffer. Gels were stained for 1 h with SYBR-Gold Nucleic Acid Gel Stain (Thermo Fisher Scientific, Waltham, Massachusetts, EUA) and imaged on a Gel Doc EZ System (Bio-Rad Laboratories Lda., Amadora, Portugal). Distinct bands were excised from the gel and eluted in 20 µL ultrapure water prior to DNA re-amplification using the same oligonucleotide primers as above, but without the GC clamp95. Amplicons were sequenced to identify microbiota OTUs (Operational Taxonomic Units). Phylogenetic analysis, to identify the closest known species, was done by comparison with sequences in the GenBank non-redundant nucleotide database using BLAST (https://www.ncbi.nlm.nih.gov) (Macrogen Europe, Amsterdam, The Netherlands). Only sequences higher than 100 bp reads and 80–100% query coverage were considered valid identification.
Statistical analysis
Data are presented as the mean and pooled standard error. Normality and homogeneity of variances were tested by the Shapiro–Wilk and Levene tests, respectively, and normalized when appropriate. Statistical evaluation of the data was done by one-way or two-way ANOVA. When p values were significant (p < 0.05), the means were compared with Tukey’s multiple range test. Histological data were analysed by the Kruskal–Wallis non-parametric test because the data were neither normal nor homogeneous and could not be normalized. All statistical analyses were performed using SPSS 24.0 software package for Windows (IBM SPSS Statistics, New York, USA).
Microbiota data analysis was done according to Serra et al.69 using the DGGE banding patterns, the band's intensity was measured with Quantity One 1-D Analysis Software v4.6.9 (Bio-Rad Laboratories Lda., Amadora, Portugal) and converted into absence/presence matrices. The calculation of the relative similarities between experimental groups and replicates was done with Primer software v7.0.5. 5 (PRIMER-E, Ivybridge, UK). Non-metric multidimensional scaling (MDS) was based on Bray–Curtis similarities using relative band abundances. Data representation from MDS was considered reliable considering the Kruskal stress value (< 0.2)96. Species Richness was established with the use of Margalef’s diversity index, while Shannon–Weaver index was utilized to establish species diversity. Similarity percentages (SIMPER) were utilized to represent the relative similarities between studied groups.
Ethics approval and consent to participate
The experiment was approved by CIIMAR ethical committee for Managing Animal Welfare (ORBEA), in compliance with the European Union directive 2010/63/EU and the Portuguese Law (DL 113/2013).
Supplementary information
Acknowledgements
This work was supported by the Fundação para a Ciência e a Tecnologia, Portugal (FCT) and Fundo Europeu de Desenvolvimento Regional (FEDER), from COMPETE 2020-Programa Operacional Competitividade e Internacionalização (POCI) (Project Eicobream: PTDC/MAR-BIO/1949/2014). Magalhães, R.,Guerreiro, I., Santos, R.A., were supported by an FCT Grant (SFRH/BD/115870/2016, SFRH/BPD/114959/2016 and SFRH/BD/131069/2017, respectively). Coutinho, F. was recipient of a grant within the Project ZEBRALGRE (PTDC/CVT-WEL/5207/2014). Serra, C.R. and Couto, A. have scientific employment contracts supported by national funds through FCT. This research was partially supported by the Strategic Funding to UID/Multi/04423/2019 (POCI-01-0145-FEDER-007621) through national funds provided by FCT and European Regional Development Fund (ERDF), in the framework of the programme PT2020. We would like to express our thanks to P. Correia for assistance during the growth trial.
Author contributions
R. E. O., H. P., and A.O.T. designed the study, R. M. and I. G. conducted the growth trial and sampling, R. M., H. P., F. C., R.A.S, C.R.S, and A.C. performed the experimental analysis and analyzed the data. R. M., H. P., and A. O. T. wrote the paper. All authors reviewed and approved the final manuscript.
Data availability
The data generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-020-70716-5.
References
- 1.NRC . Nutrient Requirements of Fish and Shrimp. Washington: The National Academies Press; 2011. [Google Scholar]
- 2.Jin M, et al. Dietary DHA/EPA ratio affected tissue fatty acid profiles, antioxidant capacity, hematological characteristics and expression of lipid-related genes but not growth in juvenile black seabream (Acanthopagrus schlegelii) PLoS ONE. 2017;12:e0176216. doi: 10.1371/journal.pone.0176216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tocher DR. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture. 2015;449:94–107. doi: 10.1016/j.aquaculture.2015.01.010. [DOI] [Google Scholar]
- 4.Castro C, et al. Regulation of glucose and lipid metabolism by dietary carbohydrate levels and lipid sources in gilthead sea bream juveniles. Brit. J. Nutr. 2016;116:19–34. doi: 10.1017/S000711451600163X. [DOI] [PubMed] [Google Scholar]
- 5.Tocher DR. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquacult. Res. 2010;41:717–732. [Google Scholar]
- 6.Ibeas C, Izquierdo MS, Lorenzo A. Effect of different levels of N-3 highly unsaturated fatty acids on growth and fatty acid composition of juvenile Gilthead Seabream (Sparus Aurata) Aquaculture. 1994;127:177–188. [Google Scholar]
- 7.Kalogeropoulos N, Alexis M, Henderson RJ. Effect of dietary soybean and cod-liver oil levels on growth and body composition of gilthead bream (Sparus aurata) Aquaculture. 1992;104:293–308. [Google Scholar]
- 8.Houston SJS, et al. The compositional and metabolic responses of gilthead seabream (Sparus aurata) to a gradient of dietary fish oil and associated n-3 long-chain PUFA content. Brit. J. Nutr. 2017;118:1010–1022. doi: 10.1017/S0007114517002975. [DOI] [PubMed] [Google Scholar]
- 9.Izquierdo M. Essential fatty acid requirements in Mediterranean fish species. Cahiers Options Mediterranéennes. 2005;63:91–102. [Google Scholar]
- 10.Fountoulaki E, Alexis MN, Nengas I, Venou B. Effects of dietary arachidonic acid (20: 4n–6), on growth, body composition, and tissue fatty acid profile of gilthead bream fingerlings (Sparus aurata L.) Aquaculture. 2003;225:309–323. [Google Scholar]
- 11.Torrecillas S, et al. Supplementation of arachidonic acid rich oil in European sea bass juveniles (Dicentrarchus labrax) diets: effects on growth performance, tissue fatty acid profile and lipid metabolism. Fish Physiol. Biochem. 2018;44:283–300. doi: 10.1007/s10695-017-0433-5. [DOI] [PubMed] [Google Scholar]
- 12.Torrecillas S, et al. Effect of fishmeal and fish oil replacement by vegetable meals and oils on gut health of European sea bass (Dicentrarchus labrax) Aquaculture. 2017;468:386–398. [Google Scholar]
- 13.Rombenso AN, Trushenski JT, Schwarz MH. Fish oil replacement in feeds for juvenile Florida Pompano: composition of alternative lipid influences degree of tissue fatty acid profile distortion. Aquaculture. 2016;458:177–186. [Google Scholar]
- 14.Castro C, et al. Liver and intestine oxidative status of gilthead sea bream fed vegetable oil and carbohydrate rich diets. Aquaculture. 2016;464:665–672. [Google Scholar]
- 15.Yildiz M, Eroldogan TO, Ofori-Mensah S, Engin K, Baltaci MA. The effects of fish oil replacement by vegetable oils on growth performance and fatty acid profile of rainbow trout: re-feeding with fish oil finishing diet improved the fatty acid composition. Aquaculture. 2018;488:123–133. [Google Scholar]
- 16.Bowyer JN, Qin JG, Smullen RP, Stone DAJ. Replacement of fish oil by poultry oil and canola oil in yellowtail kingfish (Seriola Lalandi) at optimal and suboptimal temperatures. Aquaculture. 2012;356–57:211–222. [Google Scholar]
- 17.Hatlen B, et al. Growth, feed utilization and endocrine responses in Atlantic salmon (Salmo salar) fed diets added poultry by-product meal and blood meal in combination with poultry oil. Aquacult. Nutr. 2015;21:714–725. [Google Scholar]
- 18.Monteiro M, Matos E, Ramos R, Campos I, Valente LMP. A blend of land animal fats can replace up to 75% fish oil without affecting growth and nutrient utilization of European seabass. Aquaculture. 2018;487:22–31. doi: 10.1016/j.aquaculture.2017.12.043. [DOI] [Google Scholar]
- 19.Eryalcin KM, et al. Fish oil replacement by different microalgal products in microdiets for early weaning of gilthead sea bream (Sparus aurata L.) Aquacult. Res. 2013;44:819–828. [Google Scholar]
- 20.Ryckebosch E, et al. Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem. 2014;160:393–400. doi: 10.1016/j.foodchem.2014.03.087. [DOI] [PubMed] [Google Scholar]
- 21.Qiao H, et al. Effects of dietary fish oil replacement by microalgae raw materials on growth performance, body composition and fatty acid profile of juvenile olive flounder Paralichthys olivaceus. Aquacult. Nutr. 2014 doi: 10.1111/anu.12127. [DOI] [Google Scholar]
- 22.Montero D, Izquierdo M. Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds 439–485. Boca Raton: CRC Press; 2010. [Google Scholar]
- 23.Caballero MJ, et al. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout Oncorhynchus mykiss. Aquaculture. 2002;214:253–271. [Google Scholar]
- 24.Ringo E, et al. Effect of dietary components on the gut microbiota ofaquatic animals. A never-ending story? Aquacult Nutr. 2016;22:219–282. [Google Scholar]
- 25.Mostafa Abd El-Aal, H. A. H. Lipid Peroxidation End-Products as a Key of Oxidative Stress: Effect of Antioxidant on Their Production and Transfer of Free Radicals, Lipid Peroxidation, Angel Catala. 10.5772/45944 (IntechOpen, 2012).
- 26.Jin M, et al. Regulation of growth, antioxidant capacity, fatty acid profiles, hematological characteristics and expression of lipid related genes by different dietary n-3 highly unsaturated fatty acids in juvenile black seabream (Acanthopagrus schlegelii) Aquaculture. 2017;471:55–65. doi: 10.1371/journal.pone.0176216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sargent, J. R., Tocher, D. R. & Bell, J. G. in Fish Nutrition 3rd edn (eds Halver, J. E. & Hardy, R. W.) 181–257 (Academic Press, Cambridge, 2002).
- 28.Carvan MJ, Di Giulio RT. Studies on Experimental Toxicology and Pharmacology. Totowa: Humana Press; 2015. pp. 481–493. [Google Scholar]
- 29.Mozanzadeh MT, et al. Dietary n-3 LC-PUFA requirements in silvery-black porgy juveniles (Sparidentex hasta) Aquaculture. 2015;448:151–161. [Google Scholar]
- 30.Alvarez MJ, et al. Dietary fish oil and digestible protein modify susceptibility to lipid peroxidation in the muscle of rainbow trout (Oncorhynchus mykiss) and sea bass (Dicentrarchus labrax) Brit. J. Nutr. 1998;80:281–289. [PubMed] [Google Scholar]
- 31.Castro C, et al. Dietary carbohydrate and lipid sources affect differently the oxidative status of European sea bass (Dicentrarchus labrax) juveniles. Brit. J. Nutr. 2015;114:1584–1593. doi: 10.1017/S0007114515003360. [DOI] [PubMed] [Google Scholar]
- 32.Luo Z, Tan XY, Li XD, Yin GJ. Effect of dietary arachidonic acid levels on growth performance, hepatic fatty acid profile, intermediary metabolism and antioxidant responses for juvenile Synechogobius hasta. Aquacult. Nutr. 2012;18:340–348. [Google Scholar]
- 33.Shahkar E, et al. Evaluation of the optimum dietary arachidonic acid level and its essentiality based on growth and non-specific immune responses in Japanese eel Anguilla japonica. Aquaculture. 2016;452:209–216. doi: 10.1016/j.aquaculture.2015.10.034. [DOI] [Google Scholar]
- 34.Mozanzadeh MT, et al. Dietary docosahexaenoic acid to eicosapentaenoic acid ratios effects on hemato-immunological and plasma biochemical parameters in silvery-black porgy (Sparidentex hasta) juveniles. Comp. Clin. Pathol. 2016;25:1107–1114. doi: 10.1007/s00580-016-2307-0. [DOI] [Google Scholar]
- 35.Li QF, Ai QH, Mai KS, Xu W, Zheng YF. A comparative study: In vitro effects of EPA and DHA on immune functions of head-kidney macrophages isolated from large yellow croaker (Larmichthys crocea) Fish Shellfish Immunol. 2013;35:933–940. doi: 10.1016/j.fsi.2013.07.004. [DOI] [PubMed] [Google Scholar]
- 36.Egerton S, Culloty S, Whooley J, Stanton C, Ross RP. The Gut microbiota of marine fish. Front Microbiol. 2018;9:873. doi: 10.3389/fmicb.2018.00873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Butt RL, Volkoff H. Gut Microbiota and Energy Homeostasis in Fish. Frontiers Endocrinol. 2019 doi: 10.3389/fendo.2019.00009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zarkasi KZ, et al. Atlantic Salmon (Salmo salar L.) Gastrointestinal microbial community dynamics in relation to digesta properties and diet. Microb. Ecol. 2016;71:589–603. doi: 10.1007/s00248-015-0728-y. [DOI] [PubMed] [Google Scholar]
- 39.Zhou Z, Ringø E, Olsen RE, Song SK. Dietary effects of soybean products on gut microbiota and immunity of aquatic animals: a review. Aquacult. Nutr. 2017 doi: 10.1111/anu.12532. [DOI] [Google Scholar]
- 40.Noriega BS, Sanchez-Gonzalez MA, Salyakina D, Coffman J. Understanding the impact of Omega-3 rich diet on the Gut microbiota. Case Rep Med. 2016;2016:3089303. doi: 10.1155/2016/3089303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Magalhães R, et al. Effect of dietary ARA/EPA/DHA ratios on growth performance and intermediary metabolism of gilthead sea bream (Sparus aurata) juveniles. Aquaculture. 2020;516:734644. doi: 10.1016/j.aquaculture.2019.734644. [DOI] [Google Scholar]
- 42.Yu DH, Chang JZ, Dong GF, Liu J. Replacement of fish oil with soybean oil in diets for juvenile Chinese sucker (Myxocyprinus asiaticus): effects on liver lipid peroxidation and biochemical composition. Fish Physiol. Biochem. 2017;43:1413–1420. doi: 10.1007/s10695-017-0381-0. [DOI] [PubMed] [Google Scholar]
- 43.Benedito-Palos L, et al. High levels of vegetable oils in plant protein-rich diets fed to gilthead sea bream (Sparus aurata L.): growth performance, muscle fatty acid profiles and histological alterations of target tissues. Brit. J. Nutr. 2008;100:992–1003. doi: 10.1017/S0007114508966071. [DOI] [PubMed] [Google Scholar]
- 44.Izquierdo MS, et al. Dietary lipid sources for seabream and seabass: growth performance, tissue composition and flesh quality. Aquacult. Nutr. 2003;9:397–407. [Google Scholar]
- 45.Álvarez A, Fontanillas R, García-García B, Hernández MD. Impact of dietary oil source on the shelf-life of Gilthead Seabream (Sparus aurata) J. Aquat. Food Prod. Technol. 2018;27:680–697. doi: 10.1080/10498850.2018.1484543. [DOI] [Google Scholar]
- 46.Leonardi F, et al. Effect of arachidonic, eicosapentaenoic and docosahexaenoic acids on the oxidative status of C6 glioma cells. Free Radical Res. 2005;39:865–874. doi: 10.1080/10715760500145069. [DOI] [PubMed] [Google Scholar]
- 47.Xu HG, et al. Effects of dietary arachidonic acid on growth performance, survival, immune response and tissue fatty acid composition of juvenile Japanese seabass Lateolabrax japonicus. Aquaculture. 2010;307:75–82. [Google Scholar]
- 48.Coutinho F, et al. Dietary glutamine supplementation effects on amino acid metabolism, intestinal nutrient absorption capacity and antioxidant response of gilthead sea bream (Sparus aurata) juveniles. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2016;191:9–17. doi: 10.1016/j.cbpa.2015.09.012. [DOI] [PubMed] [Google Scholar]
- 49.Coutinho F, et al. Dietary arginine surplus does not improve intestinal nutrient absorption capacity, amino acid metabolism and oxidative status of gilthead sea bream (Sparus aurata) juveniles. Aquaculture. 2016;464:480–488. [Google Scholar]
- 50.Couto A, et al. Dietary saponins and phytosterols do not affect growth, intestinal morphology and immune response of on-growing European sea bass (Dicentrarchus labrax) Aquacult. Nutr. 2015;21:970–982. [Google Scholar]
- 51.Couto A, et al. Effects of dietary phytosterols and soy saponins on growth, feed utilization efficiency and intestinal integrity of gilthead sea bream (Sparus aurata) juveniles. Aquaculture. 2014;432:295–303. doi: 10.1016/j.aquaculture.2014.05.009. [DOI] [Google Scholar]
- 52.Castro C, et al. Effects of fish oil replacement by vegetable oil blend on digestive enzymes and tissue histomorphology of European sea bass (Dicentrarchus labrax) juveniles. Fish Physiol. Biochem. 2016;42:203–217. doi: 10.1007/s10695-015-0130-1. [DOI] [PubMed] [Google Scholar]
- 53.Mourente G, Good JE, Thompson KD, Bell JG. Effects of partial substitution of dietary fish oil with blends of vegetable oils, on blood leucocyte fatty acid compositions, immune function and histology in European sea bass (Dicentrarchus labrax L.) Brit. J. Nutr. 2007;98:770–779. doi: 10.1017/S000711450773461X. [DOI] [PubMed] [Google Scholar]
- 54.Betancor MB, et al. Evaluation of a high-EPA oil from transgenic Camelina sativa in feeds for Atlantic salmon (Salmo salar L.): Effects on tissue fatty acid composition, histology and gene expression. Aquaculture. 2015;444:1–12. doi: 10.1016/j.aquaculture.2015.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Betancor M, Montero SM. D3, Usher S4, Sayanova O4, Campbell PJ5, Napier JA4, Caballero MJ3, Izquierdo M3, Tocher DR2 replacement of marine fish oil with de novo omega-3 Oils from Transgenic Camelina sativa in Feeds For Gilthead Sea Bream (Sparus aurata L.) Lipids. 2016;52:563. doi: 10.1007/s11745-016-4191-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Betancor MB, et al. An oil containing EPA and DHA from transgenic Camelina sativa to replace marine fish oil in feeds for Atlantic salmon (Salmo salar L.): Effects on intestinal transcriptome, histology, tissue fatty acid profiles and plasma biochemistry. PLoS ONE. 2017;12:e0175415. doi: 10.1371/journal.pone.0175415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Caballero MJ, et al. Morphological aspects of intestinal cells from gilthead seabream (Sparus aurata) fed diets containing different lipid sources. Aquaculture. 2003;225:325–340. [Google Scholar]
- 58.Ringo E, Bendiksen HR, Gausen SJ, Sundsfjord A, Olsen RE. The effect of dietary fatty acids on lactic acid bacteria associated with the epithelial mucosa and from faecalia of Arctic charr, Salvelinus alpinus (L.) J. Appl. Microbiol. 1998;85:855–864. doi: 10.1046/j.1365-2672.1998.00595.x. [DOI] [PubMed] [Google Scholar]
- 59.Piazzon MC, et al. Under control: how a dietary additive can restore the gut microbiome and proteomic profile, and improve disease resilience in a marine teleostean fish fed vegetable diets. Microbiome. 2017;5:164. doi: 10.1186/s40168-017-0390-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ringo E. Does dietary linoleic acid affect intestinal microflora in Arctic charr Salvelinus alpinus L. Aquacult. Fish. Manag. 1993;24:133–135. [Google Scholar]
- 61.Rimoldi S, Gliozheni E, Ascione C, Gini E, Terova G. Effect of a specific composition of short- and medium-chain fatty acid 1-Monoglycerides on growth performances and gut microbiota of gilthead sea bream (Sparus aurata) PeerJ. 2018;6:e5355. doi: 10.7717/peerj.5355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ilinskaya ON, Ulyanova VV, Yarullina DR, Gataullin IG. Secretome of Intestinal Bacilli: a natural guard against pathologies. Front Microbiol. 2017;8:1666. doi: 10.3389/fmicb.2017.01666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Estruch G, et al. Impact of fishmeal replacement in diets for Gilthead Sea Bream (Sparus aurata) on the Gastrointestinal microbiota determined by pyrosequencing the 16S rRNA Gene. PLoS ONE. 2015;10:e0136389. doi: 10.1371/journal.pone.0136389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Larsen AM. Studies on the Microbiota of Fishes and the Factors Influencing Their Composition. Auburn: Auburn University; 2014. [Google Scholar]
- 65.Parma L, et al. Next-generation sequencing characterization of the gut bacterial community of gilthead sea bream (Sparus aurata L.) fed low fishmeal based diets with increasing soybean meal levels. Anim. Feed Sci. Technol. 2016;222:204–216. [Google Scholar]
- 66.Das A, et al. Isolation, identification and characterization of Klebsiella pneumoniae from infected farmed Indian Major Carp Labeo rohita (Hamilton 1822) in West Bengal India. Aquaculture. 2017 doi: 10.1016/j.aquaculture.2017.08.037(2017). [DOI] [Google Scholar]
- 67.Floris R, et al. Intestinal bacterial flora of Mediterranean gilthead sea bream (Sparus aurata Linnaeus) as a novel source of natural surface active compounds. Aquacult. Res. 2018;49:1262–1273. [Google Scholar]
- 68.Oliveira RV, Pelli A. Disease infection by Enterobacteriaceae family in fishes: a review. J. Microbiol. Exp. 2017;4:128. doi: 10.15406/jmen.2017.04.00128. [DOI] [Google Scholar]
- 69.Serra CR, Magalhaes F, Couto A, Oliva-Teles A, Enes P. Gut microbiota and gut morphology of gilthead sea bream (Sparus aurata) juveniles are not affected by chromic oxide as digestibility marker. Aquacult. Res. 2018;49:1347–1356. [Google Scholar]
- 70.Oliveira R, et al. Klebsiella pneumoniae as a main cause of infection in nishikigoi Cyprinus carpio (carp) by inadequate handling. Braz. J. Vet. Pathol. 2014;7:86–88. [Google Scholar]
- 71.Lee DG, et al. Asinibacterium lactis gen. nov., sp. Nov., a member of the family Chitinophagaceae, isolated from donkey (Equus asinus) milk. Int. J. Syst. Evol. Microbiol. 2013;63:3180–3185. doi: 10.1099/ijs.0.047639-0. [DOI] [PubMed] [Google Scholar]
- 72.den Besten G, et al. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013;54:2325–2340. doi: 10.1194/jlr.R036012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zeng ZH, et al. Glucose enhances tilapia against Edwardsiella tarda infection through metabolome reprogramming. Fish Shellfish Immunol. 2017;61:34–43. doi: 10.1016/j.fsi.2016.12.010. [DOI] [PubMed] [Google Scholar]
- 74.Kanai K, Kondo E. Antibacterial and cytotoxic aspects of long-chain fatty acids as cell surface events: selected topics. Jpn. J. Med. Sci. Biol. 1979;32:135–174. doi: 10.7883/yoken1952.32.135. [DOI] [PubMed] [Google Scholar]
- 75.You C, et al. Effects of dietary lipid sources on the intestinal microbiome and health of golden pompano (Trachinotus ovatus) Fish Shellfish Immunol. 2019;89:187–197. doi: 10.1016/j.fsi.2019.03.060. [DOI] [PubMed] [Google Scholar]
- 76.Parolini C. Effects of Fish n-3 PUFAs on Intestinal Microbiota and Immune System. Marine Drugs. 2019;17:374. doi: 10.3390/md17060374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Fang FC. Antimicrobial actions of reactive oxygen species. mBio. 2011 doi: 10.1128/mBio.00141-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.78Marciano, F. & Vajro, P. in Gastrointestinal Tissue (eds Jordi Gracia-Sancho & Josepa Salvadó) 113–123 (Academic Press, Boca Raton, 2017).
- 79.Horwitz W. Official Methods of Analysis of AOAC International. Rockville: AOAC International; 2000. [Google Scholar]
- 80.Beutler, H. in Methods of Enzymatic Analysis, Vol. 6 (ed Bergmeyer, H. U.) 2–10 (Verlag, Chemie, Weinheim, 1984).
- 81.Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957;226:497–509. [PubMed] [Google Scholar]
- 82.Morales AE, Garciarejon L, Delahiguera M. Influence of handling and/or anesthesia on stress response in rainbow-trout: effects on liver primary metabolism. Comp. Biochem. Physiol. Physiol. 1990;95:87–93. [Google Scholar]
- 83.McCord JM, Fridovich I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein) J. Biol. Chem. 1969;244:6049–6055. [PubMed] [Google Scholar]
- 84.Aebi H. Catalase in vitro. Methods Enzymol. 1984;105:121–126. doi: 10.1016/s0076-6879(84)05016-3. [DOI] [PubMed] [Google Scholar]
- 85.Flohé, L. & Günzler, W. A. Methods in Enzymology, Vol. 105, 114–120 (Academic Press, Boca Raton, 1984). [DOI] [PubMed]
- 86.Morales AE, Perez-Jimenez A, Hidalgo MC, Abellan E, Cardenete G. Oxidative stress and antioxidant defenses after prolonged starvation in Dentex dentex liver. Comp. Biochem. Physiol. C-Toxicol. Pharmacol. 2004;139:153–161. doi: 10.1016/j.cca.2004.10.008. [DOI] [PubMed] [Google Scholar]
- 87.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 88.Buege JA, Aust SD. Microsomal lipid peroxidation. Methods Enzymol. 1978;52:302–310. doi: 10.1016/s0076-6879(78)52032-6. [DOI] [PubMed] [Google Scholar]
- 89.Griffith OW. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980;106:207–212. doi: 10.1016/0003-2697(80)90139-6. [DOI] [PubMed] [Google Scholar]
- 90.Vandeputte C, Guizon I, Genestie-Denis I, Vannier B, Lorenzon G. A microtiter plate assay for total glutathione and glutathione disulfide contents in cultured/isolated cells: performance study of a new miniaturized protocol. Cell Biol. Toxicol. 1994;10:415–421. doi: 10.1007/BF00755791. [DOI] [PubMed] [Google Scholar]
- 91.Baeverfjord G, Krogdahl A. Development and regression of soybean meal induced enteritis in atlantic salmon, Salmo Salar L, distal intestine—a comparison with the intestines of fasted fish. J. Fish Dis. 1996;19:375–387. [Google Scholar]
- 92.Krogdahl A, Bakke-McKellep AM, Baeverfjord G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmo salar L.) Aquacult. Nutr. 2003;9:361–371. [Google Scholar]
- 93.Penn MH, Bendiksen EA, Campbell P, Krogdahl A. High level of dietary pea protein concentrate induces enteropathy in Atlantic salmon (Salmo salar L.) Aquaculture. 2011;310:267–273. [Google Scholar]
- 94.Pitcher DG, Saunders NA, Owen RJ. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 1989;8:151–156. doi: 10.1111/j.1472-765X.1989.tb00262.x. [DOI] [Google Scholar]
- 95.Muyzer G, de Waal EC, Uitterlinden AG. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993;59:695–700. doi: 10.1128/aem.59.3.695-700.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Clarke KR, Warwick RM, Laboratory PM. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. Ivybridge: PRIMER-E Limited; 2001. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data generated during and/or analysed during the current study are available from the corresponding author on reasonable request.