

Abstract
Background
Microbe-mediated redox transformation of arsenic (As) leading to its mobilization has become a serious environmental concern in various subsurface ecosystems especially within the alluvial aquifers. However, detailed taxonomic and eco-physiological attributes of indigenous bacteria from As impacted aquifer of Brahmaputra river basin has remained under-studied.
Results
A newly isolated As-resistant and -transforming facultative anaerobic bacterium IIIJ3–1 from As-contaminated groundwater of Jorhat, Assam was characterized. Near complete 16S rRNA gene sequence affiliated the strain IIIJ3–1 to the genus Bacillus and phylogenetically placed within members of B. cereus sensu lato group with B. cereus ATCC 14579(T) as its closest relative with a low DNA-DNA relatedness (49.9%). Presence of iC17:0, iC15:0 fatty acids and menaquinone 7 corroborated its affiliation with B. cereus group, but differential hydroxy-fatty acids, C18:2 and menaquinones 5 & 6 marked its distinctiveness. High As resistance [Maximum Tolerable Concentration = 10 mM As3+, 350 mM As5+], aerobic As3+ (5 mM) oxidation, and near complete dissimilatory reduction of As 5+ (1 mM) within 15 h of growth designated its physiological novelty. Besides O2, cells were found to reduce As5+, Fe3+, SO42−, NO3−, and Se6+ as alternate terminal electron acceptors (TEAs), sustaining its anaerobic growth. Lactate was the preferred carbon source for anaerobic growth of the bacterium with As5+ as TEA. Genes encoding As5+ respiratory reductase (arr A), As3+ oxidase (aioB), and As3+ efflux systems (ars B, acr3) were detected. All these As homeostasis genes showed their close phylogenetic lineages to Bacillus spp. Reduction in cell size following As exposure exhibited the strain’s morphological response to toxic As, while the formation of As-rich electron opaque dots as evident from SEM-EDX possibly indicated a sequestration based As resistance strategy of strain IIIJ3–1.
Conclusion
This is the first report on molecular, taxonomic, and ecophysiological characterization of a highly As resistant, As3+ oxidizing, and dissimilatory As5+ reducing Bacillus sp. IIIJ3–1 from As contaminated sites of Brahmaputra river basin. The strain’s ability to resist and transform As along with its capability to sequester As within the cells demonstrate its potential in designing bioremediation strategies for As contaminated groundwater and other ecosystems.
Keywords: Arsenic; Bacillus cereus group; Brahmaputra river basin; Ecophysiology, Dissimilatory reduction; Groundwater; Taxonomy
Background
Arsenic (As) contamination in groundwater of Bengal Delta Plain (BDP) (covering large parts of Bangladesh and India) has become an emergent health concern for millions of people over the decades [1, 2]. Consumption of As contaminated drinking water and food grains has been implicated with severe health crisis including arsenicosis and cancer affecting more than 100 million people in BDP [3, 4]. The predominant inorganic species of As in naturally (geogenic) contaminated alluvial aquifers are As3+ and As5+. As3+ is more mobile in aqueous, oxic environments while As5+ tends to remain adsorbed to the sediments in anoxic state [5]. Relative abundance of these two species which eventually affects As -mobility and –toxicity in aquifers depends mainly on the prevailing redox conditions and inhabitant microbial activity [6–9]. The resident microorganisms in contaminated aquifer involved in redox transformation of As are considered to be the most precarious factors for As-release into the groundwater [1, 10–13]. Eco-physiological, taxonomic, and molecular characterization of As transforming bacteria from As contaminated groundwater constitute an important component of subsurface geomicrobiology, particularly to better understand the potential of these organisms in geo-cycling of As and their natural attenuation [13–16].
In recent years, several geomicrobiological studies have unanimously agreed upon the wide physiological role of taxonomically diverse bacterial populations viz. Alpha-, Beta-, Gamma-proteobacteria, Firmicutes (Bacillus and relatives), Actinobacteria, etc. influencing As bio-geochemistry in alluvial groundwater [13, 17–23]. Arsenic transforming bacteria have been known to deploy an array of metabolic routes including lithotrophic to heterotrophic mechanisms of As-oxidation [24], -reduction [25], -respiration [26], and -methylation [27], affecting As -solubility, -speciation, and -mobilization. Heterotrophic As3+ oxidizing (HAO) and chemoautotrophic As3+ oxidizing (CAO) members have been described to use As3+ as their electron source [28] and dissimilatory As5+ respiring members (DARB) to use As5+ as electron acceptor [29]. Among these organisms, DARB have been identified to play the crucial role in As mobilization from As bearing host minerals in alluvial aquifers [8, 16, 30–32]. With respect to the taxonomic and physiological characterization of DARB from diverse habitats, till date 32 cultivable representatives have been studied [29, 32, 33]. Interestingly, only few members of Proteobacteria i.e. Desulfuromonas/ Pelobacter sp. WB-3 [32], Rhizobium arsenicireducens KAs 5-22T [16], Pseudoxanthomonas arseniciresistens KAs 5-3T [34]; Achromobacter sp. KAs 3-5 [35] and Firmicutes viz. B. arsenicus [36], B. indicus [37] have been isolated from BDP (West Bengal) and characterized thoroughly. Recently, an As tolerant siderophore producing Staphylococcus sp. strain TA6 has been isolated from upper Brahmaputra River Basin (BRB) aquifer and reported to have possible role in biogeochemical cycling of As therein [38]. However, except few recent documentations, the geomicrobiology of the vast As rich alluvial aquifers of BRB remain largely unexplored [38–40].
Using anaerobic microcosms of subsurface As bearing sediment of BRB we have recently demonstrated the prominent role of Bacillus and other members of Firmicutes in As/ Fe reduction and As mobilization [40]. Particularly, strain IIIJ3-1 has been recently shown to play prominent role in As mobilization from sediments of BRB aquifer under NO3- amended anaerobic microcosm by the process of oxalate mediated mineral weathering [41]. However, detailed molecular and taxonomic characterization of any of the As reducing taxa remained still elusive. In general, presence of Bacillus spp. in As-contaminated groundwater of Bengal basin and other alluvial aquifers as well in various polluted environment and their As transformation abilities have been well documented [17, 42–45]. Dissimilatory reduction of As5+ by Bacillus spp. from soda lake, mono lake, mine environment, and effluent plants (B. arsenicoselenatis, B. selenitireducens, B. macyae and B. selenatarsenatis) and As3+ oxidation (B. firmus L-148, Bacillus sp. PNKP-S2) have been described [36, 37, 46–49]. Metabolic versatility including the ability of dissimilatory reduction of Fe and As or even SO4 by the members of this taxon could have strong influence of subsurface As mobility. Bacillus strains isolated from As contaminated sites of West Bengal (India) [36, 37], or Datong basin (China) [43] and Hetao basins (Mongolia) [50] were characterized. However, till date, no pure culture Bacillus strain capable of dissimilatory As reduction has been isolated from the vast As contaminated area of North East India (covering the BRB) and studied for its taxonomic characterization, overall physiology and As biotransformation ability (including respiratory function). The present study was carried out to highlight the taxonomic description of an As5+- respiring strain IIIJ3-1isolated from As-contaminated groundwater of BRB, India.
In the present study, we have reported a novel bacterium strain IIIJ3-1, belonging to the B. cereus group isolated from As contaminated groundwater of Jorhat, Assam (India) capable of As3+ oxidation as well as dissimilatory As5+ reduction. Molecular phylogenetic analysis coupled with chemotaxonomic and ecophysiological characterization and study on As biotransformation properties have been performed to establish its taxonomic novelty within the group B. cereus sensu lato. Based on its ability to survive under both oxic and anoxic environments and high As resistance as well as As redox transformation, the strain has been chosen for further investigation and characterization of its eco-physiological property.
Results
Polyphasic taxonomic characterization of strain IIIJ3–1
16S rRNA gene phylogenetic analysis
Comparison of near complete 16S rRNA gene sequence (1474 bp) of strain IIIJ3-1 indicated high degrees of sequence similarities (98.2-99.9 %) to the members of the genus Bacillus, where type strain B. cereus ATCC 14579(T) was the most hit taxon (99.9 % similarity) at 99 % query coverage. The NJ phylogenetic reconstruction showed that strain IIIJ3-1 formed a coherent cluster of monophyletic pattern with the type strain of B. cereus ATCC 14579(T) and claded to the type members of Bacillus (Fig 1a), indicating its affiliation to the genus Bacillus. Both ML and ME phylogenetic reconstruction methods indicated a consistent tree topology clading strain IIIJ3-1 to the B. cereus ATCC 14579(T) as the nearest phylogenetic neighbor. Phylogenetic analysis involving As transforming Bacillus spp., from various habitat showed that strain IIIJ3-1 is taxonomically closest to the B. cereus AG27 (AY970345.1, an As-resistant bacterium from agricultural soil) [51], denoting its species level affiliation to the B. cereus members (Fig 1b). While, other Bacillus members having arsenate and selenite respiratory/ reductive activities (B. arseniciselenatis from mono lake and B. arsenicus from groundwater) coherently clustered with the clade comprising strain IIIJ3-1.
Fig. 1.

Unrooted Neighbour Joining (NJ) phylogenetic tree based on 16SrRNA gene sequences of strain IIIJ3–1 and related Bacillus spp., a. consensus phylogenetic tree of strain IIIJ3–1 and all validly described type strains of Bacillus using the Jukes-Cantor distance model considering a total of 1485 positions in the final dataset, b. Phylogenetic relationship of strain IIIJ3–1 and other As transforming Bacillus spp. isolated from diverse As-impacted habitat with a total of 1490 positions in the final dataset. The black solid circle indicates the consistent tree topology obtained through both NJ and ML methods. Bootstrap values (percentage of 1000 replications) greater than 60% are represented at the nodes. The GenBank accession numbers are mentioned in the parentheses. Scale bar denotes the rate of nucleotide substitution
Genomic analysis
Genomic G+C content of strain IIIJ3-1 was found to be 45.0 mol%. The value of ∆Tm for the heteroduplex, formed with the genomic DNA of test isolate IIIJ3-1 and its closest phylogenetic match B. cereus ATCC 14579(T) at optimal renaturation temperature (TOR) 69.95°C, and homoduplex of B. cereus ATCC 14579(T) was found to be 8°C (Supplementary Fig S1). Relative binding ratio indicating DNA-DNA relatedness of the two strains was found to be 49.9%.
Chemotaxonomic characteristics
Cellular FAME profile of strain IIIJ3-1 showed abundance of isoC17:0 (10.3%), isoC15:0 (8.8%) followed by C15:0 (6.8%), C17:0Δ (5.7%), and 2OH-C14:0 (5.2%) (Supplementary Fig. S2). Relatively less abundant 2-OH-C16:0(3.8%), C18:29,12 (3.2%), C19:0 (3.2%), C11:0 (3.1%), trans C18:19(2.9%), cis-C18:19 (1.8%), C16:0 (2.7%), isoC16:0 (2.4%), and anteiso-C15:0(2.4%) were detected. Trace amounts of 2OH-C10:0, 3OH-C12:0, 3OH-C14:0, C14:0 and unresolved peaks corresponding to 24% of total FAMEs were obtained. Detailed analysis of isoprenoid respiratory quinones revealed presence of several menaquinones (MKs) in strain IIIJ3-1 (Table 1). Abundance of MK-5 (55%), MK-6 (25%), and MK-7 (7.8%) fractionated by HPLC at 2.9, 4.4 and 8.4 minutes were determined (Supplementary Fig. S3a). MALDI-MS analysis followed by Pubchem database search revealed the molecular identity of the MKs. (Supplementary Fig. S3b). Derivatives of MK-5 were found to be MK-5-d7, MK-5 epoxide and demethyl MK-5 d7 with corresponding molecular masses of 519, 535, 505 respectively. MK-6 derivatives were found to be methyl substituted MK-6 d5, MK-6 d5 and deoxygenated MK-6 with molar mass of 599, 585 and 551.56, respectively. MK-7 d7 and MK7-d7 epoxide with molar mass of 656 and 672 were found to be the major MK-7 derivatives (Supplementary Fig. S3c).
Table 1.
Differential phenotypic and biochemical characteristics of Bacillus cereus strain IIIJ3–1 and taxonomically and metabolically related species.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Colony colour/ nature | cream | ND | Red | Cream | Brown | Yellow–orange | cream | rhizoidal | white | White/grey | cream | rhizoidal | White | cream |
| As5+ tolerance (mM) | 350 | 0 | NA | 20 | 0 | 20 | nr | nr | nr | nr | nr | nr | nr | nr |
| As3+ tolerance (mM) | 10 | 0.3 | 0.3 | 0.3 | 0 | 0.3 | nr | nr | nr | nr | nr | nr | nr | nr |
| Catalase | + | + | + | + | nr | nr | + | + | + | + | nr | + | + | + |
| Oxidase | + | + | – | + | – | – | nr | – | + | – | nr | – | – | + |
| Temperature Range (°C) | 20–50 | nr | nr | 20–40 | 18–37 | 15–37 | 20–50 | 10–40 | 10–45 | 10–45 | 5–37 | 5–37 | 10–50 | 10–45 |
| Salinity (%) | 0–4 | 2.4–6 | 0–6 | 0–1 | 0–2 | 0–2 | nr | 0–2.5 | 0 | 0 | nr | 0–4 | nr | 0–5 |
| pH range | 6–10 | 8.5–10 | 8.5–10 | 5.5–8 | 7–9.5 | 06–7.1 | nr | 5–9.5 | 5–9.5 | 5–9.5 | nr | 5–9.5 | nr | 5–9.5 |
| (G + C)% | 45 | 40 | 49 | 35 | nr | 41.2 | 35.8 | 34–36 | 35.3 | 35.38 | 35.5 | 34.1 | 35.4 | 35.6 |
| Major quinone | MK5, MK6, MK7 | nr | nr | MK7 | MK7 | MK7 | nr | nr | MK7 |
MK8 MK7 MK3 |
nr | MK7 | MK7 | nr |
| Mannitol | – | nr | nr | + | – | – | – | – | – | – | – | – | – | – |
| Rhyzoidal colony | – | – | – | – | – | – | – | + | – | – | – | + | – | – |
| Starch hydrolysis | + | + | + | + | + | + | – | – | + | + | + | + | + | + |
| Glycerol | – | – | nr | – | + | – | – | + | – | – | – | |||
| Ribose | ND | nr | nr | – | + | + | + | + | + | – | + | + | + | + |
| Galactose | – | – | + | nr | – | nr | – | – | – | – | – | – | – | – |
| D-mannose | – | nr | nr | nr | + | + | + | – | – | + | – | – | – | – |
| N acetyl Glucosamine | + | nr | nr | nr | + | nr | + | + | + | + | + | + | + | + |
| Salicin | – | nr | nr | nr | – | nr | + | + | + | + | + | + | – | + |
| Cellobiose | – | nr | nr | – | – | + | + | – | + | + | + | + | – | + |
| Sucrose | + | nr | nr | + | – | + | – | – | + | + | – | + | + | + |
| Trehalose | + | nr | nr | nr | + | nr | – | + | + | + | + | + | + | + |
| Glycogen | ND | nr | nr | nr | + | nr | – | + | + | + | + | + | + | + |
| β-gentiobiose | – | nr | nr | nr | + | nr | – | – | – | – | – | – | – | – |
| Turanose | – | nr | nr | nr | nr | – | – | – | – | – | – | + | – |
Negative growth results were found for sugars- D-raffinose, mellibiose, D- mannose, D- galactose, β-gentiobiose, D-turanose, α-rhamnose, 3-methyl glucose, D-fucose and β–methyl–D–glucoside, sugar alcohols- D-sorbitol, D- mannitol, D-arabitol, myo-inositol, glycerol, sugar amine and amide- N-acetyl galactosamine and glucuronamide, amino acids- D-serine, α-glutamic acid, α-histidine glycyl-L- proline, L-alanine, L-arginine, sugar alcohols- glycerol, D- salicin, sugar acids- p-hydroxy phenyl acetic acid, L-aspartic acid, D-galactouronic acid, L-galactonic acid, D-glucuronic acid, D-lactic acid methyl ester, citric acid, γ-amino butyric acid, α-hydroxy butyric acid, β-hydroxy D-Lbutyric acid, α-keto butyric acid, N-acetyl neuraminic acid, D-aspartic acid, α- pyroglutamic acid, quinic acid, acetic acid, fusidic acid, D- saccharic acid, bromosuccinic acid, formic acid, nalidixic acid, sodium butyrate, mucic acid, α-ketoglutaric acid, D-malic acid, acetoacetic acid, propionic acid and Tween 4
B. inferioriaquae strain IIIJ3–1 (present study); 2. B. arsenicoselenatis; 3. B. selenatireducens (2,3) [46]; 4. B. arsenicus [36]; 5. B.barbaricus [52]; 6. B. indicus [37]; 7. B. cytotoxicus; 8. B. pseudomycoides; 9. B. cereus; 10. B. thuringiensis; 11. B. weihenstephanensis; 12. B. mycoides; 13. B.anthracis (7–13) [53]; 14. B. toyonensis [54]. Symbols; (+) = positive, (−) = negative; ND = not determined, nr = no report, MK = menaquinone
Phenotypic and biochemical characterization
The colonies of strain IIIJ3-1 were creamy white with undulating edges and pasty appearance. The colonies became rhizoidal after 48h of growth. Scanning electron micrograph confirmed the cells of strain IIIJ3-1 to be rod-shaped with cell size of 2.5 x 0.7 μm (Fig. 2). The strain was found to be Gram-positive, facultative anaerobic, non-motile, catalase-, and oxidase- positive, endospore-forming (terminal to sub-terminal), non-flagellated, capsulated, and capable of reducing As5+ via dissimilatory process. Differential physiological characteristics of strain IIIJ3-1 with those of phylogenetically related Bacillus spp. including those isolated from As contaminated groundwater have been summarized in Table 1. Ability to utilize diverse C-sources and TEA of strain IIIJ3-1 was ascertained and compared with other Bacillus spp. reported to be As resistant (Supplementary Table 1). Anaerobic growth of strain IIIJ3-1 with diverse TEAs yielded positive response with As5+, SO42- , Fe3+, NO3- and Se6+.
Fig. 2.

Scanning electron micrograph of strain IIIJ3–1(T) after 18 h of growth on LB agar plate
Arsenic transformation potential of strain IIIJ3–1
Aerobic As biotransformation assay
The activity of resting cells of strain IIIJ3-1 for substrates As5+ and As3+ was investigated. Using molybdenum blue method, the Km for As5+ reduction was estimated to be 10 mM (Supplementary Fig. S4) with a Vmax of 0.25 mM h-1 and those for As3+ oxidation were found to be 2.8mM and 0.2 mM h-1, respectively. Kinetics for As5+ reduction and As3+ oxidation by strain IIIJ3-1 demonstrated its considerably higher affinity towards As3+ (~3 times) than As5+ (Supplementary Fig. S4).
Anaerobic growth of strain IIIJ3–1
Anaerobic growth of strain IIIJ3-1 with varying TEAs revealed that As5+ was the most respiratory substrate preferred (with lactate as electron and C- source) followed by utilization of other TEAs in the following order: Fe3+ > Se6+> SO42-> NO3- (Fig. 3a). No growth could be detected with S2O32-. Mean generation time (g) and mean growth rate constant (k) of the strain with As5+ was 45 minutes and 0.220, respectively followed by increasing g and decreasing k for Fe3+, Se6+, NO3-, and SO42- (Supplementary Table 2) indicating fastest growth utilizing As5+ as compared to others. Electron donor utilization profile with As5+ as TEA revealed maximum cell yield with lactate followed by raffinose, citrate, inositol, tartarate, gluconate, starch, nitrite and pyruvate. Faint or negligible growth was found with mannose, fructose, acetate, arabinose, succinate, mannitol, glucose and glycerol (Fig. 3b). Reduction profiles for As5+, Fe3+, NO3- and SO42- by strain IIIJ3-1 indicated near complete transformation of As5+ within 15 h of growth (Fig. 3c). Linear regression and correlation analysis (Supplementary Fig. S5) of growth vs coupled reduction profile (Conc-1) showed that a high goodness of fit (R2= 0.8, r = -0.671) was achieved for SO42- reduction w.r.t time indicating the stoichiometric balance between reduction profile and energy generation by IIIJ3-1 for cellular growth. While, a high variance (R2=0.38, r = -0.915, R2=0.38, r = 0.691) was noted for both As5+ and Fe3+ indicated a partially uncoupled behavior of growth vs reduction profile (Conc-1). This observed stoichiometric imbalance between reduction profile and energy generation by IIIJ3-1 might be attributed to the cellular As uptake during reduction/respiration by IIIJ3-1. A relatively less variance (R2= 0.46, r = -0.705) was noticed for NO3- reduction w.r.t time indicated a partially coupled growth behavior of cells of IIIJ3-1, which might be attributed to the organization of denitrification pathway i.e. either only reduction of NO3- to NO2- or denitrification to NH4+ or N2. The overall observation indicated the versatile ability of the bacterium to cope up with wide redox fluctuation within its natural environment i.e., groundwater.
Fig. 3.
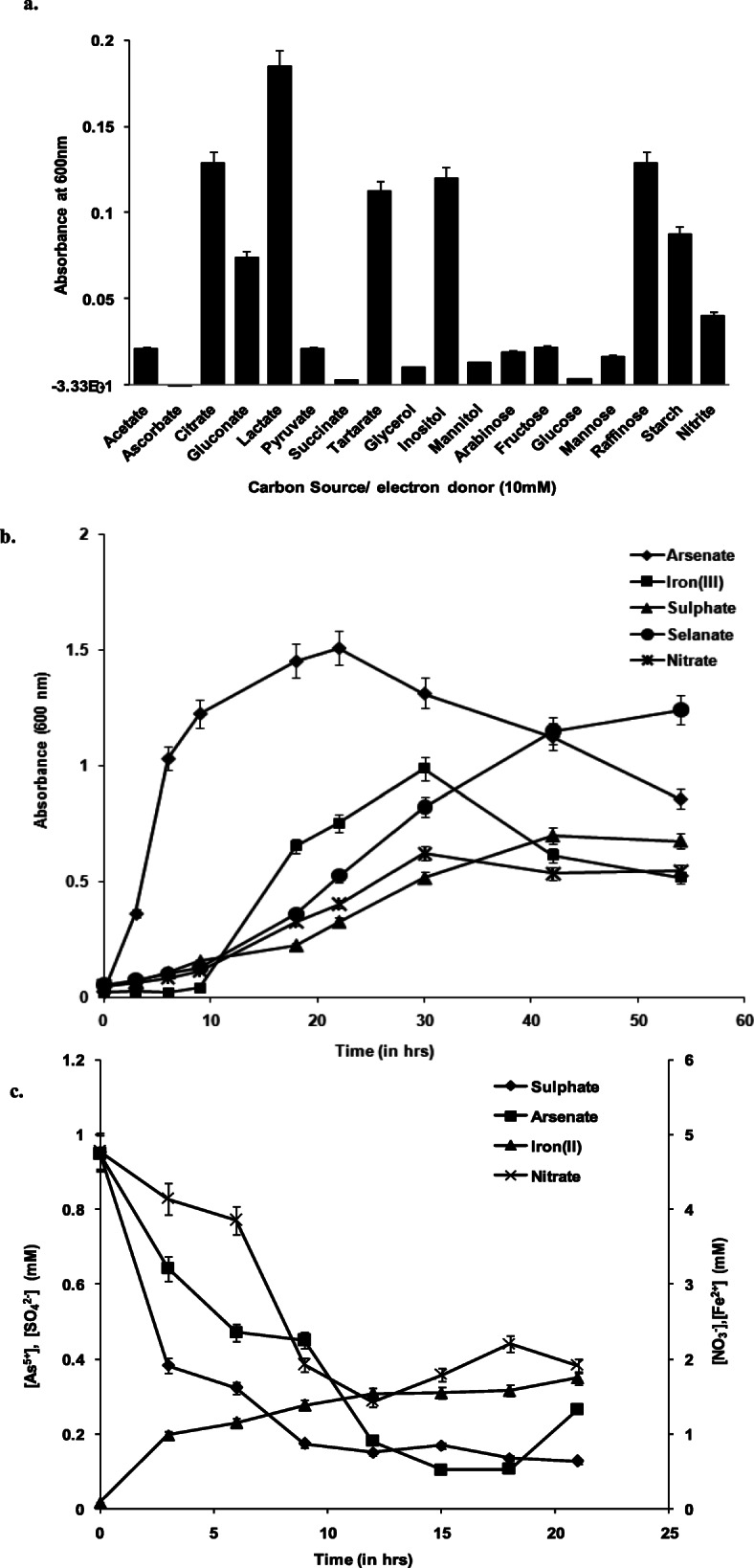
Anaerobic growth kinetics of strain IIIJ3–1 in presence of alternate carbon source and terminal electron acceptors. a. C- source/e− donor utilization profile with As5+ as TEA, b. anaerobic growth kinetics in presence of alternate e− acceptors, and c. reduction profile of alternate electron acceptors. Error bars indicate 5% percent of the value plotted
Molecular analysis
Presence of As homeostasis genes arrA, aioB, arsB and acr3 (1) were observed through PCR amplification with respective primers, sequencing and phylogenetic analysis. The arrA gene (147 deduced amino acids) from strain IIIJ3-1 revealed 99% identity with membrane proteins of B. cereus. Close phylogenetic relatedness of arr from strain IIIJ3-1 with the hypothetical membrane proteins of members of B. cereus group (Fig. 4a). Multiple alignment of the deduced amino acid sequences of putative Arr from strain IIIJ3-1 and Arr from other DARBs indicated 12 conserved amino acid residues and 6 residues replaced by amino acids bearing similar side chain or same functional group maintaining the expected role (Supplementary Fig. S6a).
Fig. 4.

Maximum Likelihood phylogenetic tree for As homeostasis genes based on their deduced amino acid sequences: Arr (a), AioB (b), Acr3 (c) and ArsB (d)
NCBI database search for the deduced amino acid sequences of putative AioB from strain IIIJ3-1 did not reveal sequence identity with any of the known Aio sequences. However, phylogenetic analysis of AioB sequence obtained from strain IIIJ3-1 indicated its distant relatedness with As3+oxidase reported from Listeria monocytogenes (Fig. 4b). Multiple alignments of 188 deduced amino acids of AioB from strain IIIJ3-1 with 12 other reported AioB sequences revealed 19 consensus amino acids with a conserved arginine residue (Supplementary Fig. S6b). The conserved regions FDHGG, GGGFEN and IHNRPAYNSE of known As3+ oxidases were found to be 40%, 17% and 20% conserved in the putative AioB sequence of IIIJ3-1 respectively.
Deduced amino acid sequences of acr3 and arsB genes showed a similarity match with arsenical resistance proteins and arsenite pump proteins (ArsB) of various As resistant strains, respectively. Phylogenetic analysis of the deduced amino acid sequences of Acr3 obtained from B.cereusIIIJ3-1 showed relatedness with the Acr3 sequence obtained from other members of B. cereus, Desulfosporosinus, Peptococcaceae, Lentisphaera, Pseudomonas and Aeromonas (Fig. 4c). However, a bootstrap value of only 67 showed significant sequence diversification of the putative Acr3 obtained from IIIJ3-1. Deduced amino acid residues of the putative ArsB from strain IIIJ3-1 revealed comparatively closer phylogenetic relatedness to those reported from Halanaeroarchaeum, followed by ArsB sequences of Stenotrophomonas, Acinetobacter, Desulfovovibrio, B. subtilis and B. anthracis than those obtained from members of B. cereus (i.e. B. thuringiensis, B. weihenstephanensis (Fig. 4d). Multiple alignments of the deduced amino acid sequences of acr3 (94a.a) and arsB (107a.a) showed that the sequences were 35% and 21% conserved, respectively (Supplementary Fig. S6c and d).
SEM and EDX analysis
Scanning electron microscopic images revealed distinct morphological changes in terms of cell shape and size of strain IIIJ3-1 following growth with As3+ or As5+ under aerobic and anaerobic conditions (Fig. 5a). Control cells were found to be ~ 3 μm x 1 μm with smooth cell surface (Fig. 5a i) whereas those grown with As showed reduction in cell size and exhibited rough and convoluted cell surface. Cells grown with As3+ (under both aerobic and anaerobic conditions) indicated electron opaque dots and reduction in length but increase in diameter (i.e 2.4 μm X 1.4 μm for aerobically grown cells and 2.2 μm X 1.2 μm for anaerobically grown cells). Cells under As5+ stress (both aerobic and anaerobic) did not show distinct As rich dots but cell shrinkage was prominent i.e 2 μm X 0.4 μm and 1.8 μm X 0.5 μm aerobically and anaerobically grown cells respectively. Moreover, the As stressed cells were found to be clustered together and covered with an exopolysaccharide layer which contributes to effectively lower the exposed cell surfaces through which the cells adhere to each other (Fig. 5; Table 2). Surface area: volume ratio for control cells is found to be 4.67 which eventually decrease for As3+ stressed cells to 3.7 and 4.2 for oxic and anoxic growth, respectively. However, the As5+ cells show an increase in surface area : volume ratio upto 6 and 5 for oxic and anoxic growth, respectively along with prominent surface constrictions and clustering of the cells. The pattern of As accumulation as determined from the EDX data is as follows (Fig. S7): As3+ grown aerobic cell (1.21 wt%) > As3+ grown anaerobic cells (0.94 wt%) > As5+ grown aerobic cells (0.36 wt %) > As5+ grown anaerobic cells (0.09 wt %) (Supplementary Fig. S7 a-d). This observation corroborated the stoichiometric imbalance between growth and reduction kinetics profile by the bacterium. No peak for As could be observed for control cells (Supplementary Fig. S7e). Point EDS analysis performed on electron opaque dots formed during incubation of the cells with As3+ showed prominent peak for As. Such a peak for As was found to be absent on any other position on the cell surface except on the dots formed (Fig. 5b). Carbon content of the As containing electron opaque dots was found to be much higher than that of the normal cell surface which might have occurred due to encapsulation of the dots with polymeric C containing exudates.
Fig. 5.

Cells grown (i) without As stress i.e. control (ii) aerobically with As3+,(iii) anaerobically with As3+, (iv) aerobically with As5+, (v) anaerobically with As5+(a). Analysis of electron dense particles (i) magnified image of anaerobically grown strain in presence of As3+ (ii) point EDX analysis on an electron opaque dot and on another point (as control) on cell surface of strain IIIJ3–1 (b)
Table 2.
Morphometric calculations of strain IIIJ3–1 grown with and without As stress; h = length, r = radius, V = volume and, A = area of the cells
| Growth condition | h (μm) | r (μm) | V (μm3) | A (μm2) | A/V |
|---|---|---|---|---|---|
| Control | 3.0 | 0.5 | 2.4 | 11.0 | 4.7 |
| Aerobic As(III) | 2.4 | 0.7 | 3.7 | 13.6 | 3.7 |
| Anaerobic As(III) | 2.2 | 0.6 | 2.5 | 10.6 | 4.2 |
| Aerobic As(V) | 2.0 | 0.4 | 1.0 | 6.0 | 6.0 |
| Anaerobic As(V) | 1.8 | 0.5 | 1.4 | 7.2 | 5.1 |
Discussion
Although B. cereus is a well-known facultative anaerobe, anaerobic respiration of As5+ in this group has never been documented. This is the first report on detailed taxonomic analysis of strain IIIJ3-1, phylogenetically belonging to B. cereus sensu lato group, isolated from As contaminated aquifers of Jorhat, showing remarkable metabolic and genomic potential to respire and resist As. Despite highest sequence similarity of strain IIIJ3-1 with B. cereus ATCC 14579(T), neighbor joining tree clearly delineated strain IIIJ3-1 to be a different taxon, phylogenetically distinct from other related members (Fig. 1a). Phylogenetic trees (NJ) (Fig. 1b) constructed with strain IIIJ3-1 and other Bacillus spp. resistant to As revealed a close taxonomic relatedness of strain IIIJ3-1 with B. cereus AG-27 (AY970345.1), an As resistant strain isolated from a thermal power plant soil at Kanpur (India) [51] with a bootstrap value (100 %). Other As resistant strains B. indicus and B. arsenicus isolated from As contaminated aquifers of BDP [36, 37], B. selenatiarsenatis, B. arseniciselenatis strains known for their ability to respire Se6+ as well as As5+ [46, 47], B. beveredgei, a facultative anaerobic haloalkaliphilic strain capable of As respiration [55], B. foraminis, isolated from a non-saline alkaline groundwater [56] and Bacillus sp. S1 (KF857225), an As resistant strain isolated from As contaminated aquifer of Jorhat (BRB) (data retrieved from NCBI) were found to be phylogenetically distant from strain IIIJ3-1. The phylogenetic analyses indicate that strain IIIJ3-1 is taxonomically distantly related to the previously reported members of B. cereus as well as other As resistant members of Bacillus sp. isolated from elsewhere sites. Interestingly, it is even found to be distantly related to the Bacillus sp. reported from Jorhat, similar region from where strain IIIJ3-1 has been isolated (Fig. 1b). Further genomic characterization of strain IIIJ3-1 for DNA-DNA relatedness with the closest NCBI match revealed a ΔTm of 8 °C (Supplementary Fig. S1) which remains well above the suggested limit of 5 oC for species delineation [57] and a characteristic low G+C content corroborated its affiliation to Bacillus cereus group.
Chemotaxonomic characterization through bacterial FAME and respiratory quinone analyses also indicated characteristic features of B. cereus group along with significant distinct features. Comparison of the fatty acid profile of strain IIIJ3-1 with its closest phylogenetic neighbors revealed prominent differences in its fatty acid composition (Table 3). Like all other members of B. cereus group, strain IIIJ3-1 is characterized by two fold abundant iC15:0 than aC15:0 and abundance of iC17:0 [58]. Similar to ATCC 14579 (T), presence of C18:1 in strain IIIJ3-1 was also noted [54]. However, presence of hydroxyl fatty acids (HFAs), C11:0 and C18:29,12 with simultaneous absence of iC13:0, iC14:0, iC16:2, aC13:0, aC17:0, aC17:1, nC16:2 and nC17:1 in strain IIIJ3-1 could be considered as distinct chemotaxonomic properties supporting its taxonomic uniqueness from its closest taxonomic relatives.
Table 3.
Fatty acid composition of Bacillus cereus strain IIIJ3-1 and taxonomically related Bacillus species
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hydroxy- fatty acids | ||||||||||||
| 2-OH C10:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| 2-OH C12:0 | – | – | – | – | – | – | – | – | – | – | – | – |
| 3-OH C12:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| 2-OH C14:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| 3-OH C14:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| 2-OHC15:0 | – | – | – | – | – | – | + | + | – | – | – | – |
| 2-OH C16:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| Branched chain iso fatty acids | ||||||||||||
| iC12:0 | – | + | – | – | – | – | – | – | – | + | + | + |
| iC13:0 | – | + | + | + | + | + | + | + | + | + | + | + |
| iC14:0 | – | + | – | – | – | + | + | + | + | + | + | + |
| iC15:0 | + | + | + | + | + | + | + | + | + | + | + | + |
| iC16:0 | + | – | – | – | – | + | + | + | + | + | + | + |
| iC17:0 | + | – | + | + | + | + | + | + | – | + | – | + |
| Branched chain anteiso fatty acids | ||||||||||||
| aC13:0 | – | + | + | + | + | – | + | + | + | + | + | + |
| aC15:0 | + | + | + | + | + | + | + | + | + | + | + | + |
| aC17:0 | – | + | + | + | + | – | + | + | + | + | + | + |
| aC17:1 | – | + | + | + | + | + | + | + | + | + | + | + |
| Saturated fatty acids (SFAs) | ||||||||||||
| C11:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| C12:0 | + | + | – | – | – | – | – | + | + | + | + | + |
| C14:0 | + | + | + | + | + | + | + | + | + | + | + | + |
| C15:0 | + | – | – | – | – | – | – | – | – | – | – | f |
| C16:0 | + | + | + | + | + | + | + | + | + | + | + | + |
| C17:0 | + | + | – | – | – | – | – | – | – | – | – | f |
| C18:0 | – | + | – | – | – | – | – | + | + | + | + | + |
| C19:0 | + | – | – | – | – | – | – | – | – | – | – | – |
| Unsaturated fatty acids (UFAs) | ||||||||||||
| C18:1 | + | + | – | – | – | – | – | – | – | – | – | f |
| C16:1ω7cOH | – | nr | – | – | – | – | + | – | + | – | + | – |
| C16:1ω11c | – | f | – | – | – | – | – | – | + | – | + | – |
| nC16:1 | – | + | – | – | – | – | – | – | – | – | – | + |
| iC16:2 | – | + | – | – | – | – | – | – | – | – | – | – |
| nC16:2 | – | + | – | – | – | – | – | – | – | – | – | – |
| nC17:1 | – | + | – | – | – | – | – | – | – | – | – | – |
| C18:29,12 | + | – | – | – | – | – | – | – | – | – | – | – |
Strains 1. IIIJ3–1 (present study); 2. B. cereus ATCC 14579 [54]; 3. B-17, 4. B-19, 5. B-82,(3,4,5) – [58, 59]; 6. B. toyonensis BCT-7112T; 7, B. cereus CECT 148T; 8. B. thuringiensis CECT 197T; 9. B. mycoides CECT 4128T; 10. B. pseudomycoides CECT 7065T;; 11. B. weihenstephanensis LMG 18989T;12. B. cytotoxicus) (6–12) [60]. (+ : presence, −: absence and f: faint peak for FAME obtained, nr: no report)
Study on respiratory menaquinone also supported its relatedness to the members of B. cereus group with simultaneous unique properties. The molecular identity of quinones provided a further clue to its novelty. While the presence of MK-7 could act as supporting feature for its chemotaxonomic affiliation to B. cereus group, presence and abundance of derivatives of MK-5 and MK-6, which has never been reported for members of B. cereus group differentiates it from the other members of this group.
Strain IIIJ3-1 is found to share a number of phenotypic and metabolic traits with B. cereus group. The physicochemical growth characteristics of strain IIIJ3-1 was found to be consistent with those of other members of genus Bacillus. The rhizoidal shape in ageing colonies as found for strain IIIJ3-1 has only been found in B. mycoides and B. pseudomycoides [52]. Ability to hydrolyze starch and utilize N-acetyl glucosamine, sucrose, trehalose and inability to assimilate mannitol, glycerol, galactose, β-gentiobiose and turanose aligns with the previous findings [36, 37, 46, 52, 53, 60]. However, inability to utilize salicin is a distinct metabolic character of strain IIIJ3-1. Anaerobic growth utilizing alternate TEAs are also in accordance with the previous reports for other members of Bacillus [36, 46, 52]. It was suggested that dissimilatory reduction of some metals or metalloids may be a specific character of spore-forming Gram-positive bacteria [61]. Recent geomicrobiological studies have also reported the versatile ability of As-rich groundwater bacteria (Rhizobium spp., Pseudoxanthomonas sp., Achromobacter sp., Escherichia spp.) from Bengal basin, India to couple electrons from hydrocarbons and organic acids (lactate, acetate, pyruvate, etc.) to multiple electron acceptors (TEAs) under anaerobic conditions, thus driving the reductive metabolism [16, 62, 63]. Alluvial aquifer of BRB is characterized to be oligotrophic, anoxic, and with fluctuating availability of redox equivalents, lowered derived carbon pool [64]. Considering the overall hydrogeochemistry of BRB groundwater, the metabolic versatility of the strain IIIJ3-1 seems highly justified for its competitive niche adaptation to cope up with fluctuating groundwater condition. Statistical analysis (UPGMA) based on overall chemotaxonomic and physiological properties of strain IIIJ3-1 with other 7 members of B. cereus group (Tables 1 and 3) clearly indicated the distinctiveness of this strain from its nearest taxonomic neighbors (Fig. 6). Linear regression and correlation analysis of growth vs coupled reduction profile with stoichiometric imbalance indicated the cellular As uptake during reduction by IIIJ3-1 and its cellular compartmentalization. The observed phenomena corroborated earlier reports on cellular and extracellular adsorption of As during growth by some As-transforming bacterial members from diverse habitat [35, 65–67].
Fig. 6.

UPGMA analysis for strain IIIJ3–1and its phylogenetic neighbors (members of B. cereus group) considering their biochemical and chemotaxonomic traits
A comparative analysis of kinetic parameters reveals that Km for As3+ oxidation (by oxidase) of strain IIIJ3-1 is about 5 times higher than that of Pseudomonas arsenoxydans but comparable to that of Alcaligenes strain (Supplementary Table 2). Km for As5+ reduction (by reductase) was comparable to that of E.coli plasmid R773 bearing arsC gene and is much lower than that of S. cereviseae having Acr2p, but 1-3 folds higher than Synechocystis sp. strain PCC 6803, Pseudomonas stutzeri and Staphylococcus aureus possessing arsC genotype [68–70] (Supplementary Table 2). Relatively higher Km towardsAs5+ reduction could be attributed to the nonspecific reduction by the cell and this perhaps corroborate well with absence of arsC gene encoding cytosolic As5+ specific reductase (discussed later).
Preference of lactate over other C-sources by strain IIIJ3-1 under anaerobic growth is in well accordance with the free energy values reported in the literature, where lactate has proved to be the highest energy yielding electron donor with diverse TEAs [68, 71]. Lactate is the readily available fermentation product by many of the naturally occurring bacteria possibly present in such aquifer systems. Therefore, capability to utilize lactate as a C-source by the As5+ reducing strain IIIJ3-1 might help in competitive nutrient acquisition and niche adaptation strategy over other bacteria unable to metabolize lactate. Dissimilatory As5+ reducing prokaryotes belonging to various genera such as Desulfitobacterium, Desulfuromonas, Desulfotomaculum, Sulfurospirillum, and Bacillus capable of utilizing lactate have been reported for their ability to metabolize various other inorganic respiratory substrates as well [32, 46, 47, 72–74]. However, a distinct preference of As5+ over other respiratory substrates is a novel characteristic found in strain IIIJ3-1.
Reduction of SO42-, NO3- and Fe3+ was relatively incomplete and their transformation rates varied considerably. Both As5+ and NO3- revealed similar patterns of reduction and interestingly after reaching the saturation level of reduction (i.e after 15 hours), oxidized substrates were detected in the aqueous phase indicating possible oxidation of the reduced products formed. The observed redox transformation might imply presence of an oxidoreductase system regulating As5+ and NO3- reduction which may act reversibly when the reduced species reached a threshold concentration. Involvement of a common, non-specific or constitutive oxidoreductase system in Sulfurospirillum barnesii SES-3 has been reported earlier [73]. The energetics reported in various studies for lactate oxidation with As5+, SO42- and NO3- has been tabulated in Supplementary Table 3 [46, 72]. Since, ∆Gf° denotes the free energy difference in reactants and the products, lower ∆Gf° value indicates higher amount of available energy for cell mass formation. Although NO3- was identified as a preferred substrate thermodynamically over As5+ and SO42-, the present observation on preference of As5+ by strain IIIJ3-1 could be attributed to the natural adaptation acquired from its provenance to a highly As contaminated aquifer which might have assisted in the evolution of such physiological characteristic. Relatively lower cell density with As5+ compared to NO3-was reported due to the toxic effect of accumulated As3+ formed while growing on As5+ [68]. Noticeably, such phenomenon was not evident with strain IIIJ3-1, indicating presence of strong As3+ detoxification system operating in the strain while it is growing on As5+. Presence of As3+ oxidase gene and its activity as present in the strain corroborated such interpretation.
Absence of gene encoding cytosolic As5+ reductase (arsC) in strain IIIJ3-1 was noted and this observation (presence of arsB but absence of arsC) corroborated with a similar finding in Exiguobacterium strain WK6 [74]. However, cytosolic As5+ reductase activity, very high Km for cytosolic As5+ reduction and absence of arsC gene suggested some arsC independent pathway present in strain IIIJ3-1 for cytosolic As5+ reduction. Despite the clear evidence for aerobic As5+ reduction by strain IIIJ3-1, an amplicon for As5+ reductase (arsC) could not be obtained. It might be due to high sequence diversification of arsC that the conventional primers for the gene could not target the one present in IIIJ3-1. The As5+ respiratory reductase (arr) and As3+ transporter genes arsB and acr3(1) appear to be vertically transferred maintaining their phylogenetic consistency. The phylogenetic discrepencies in case of aioB suggests possible horizontal transfer of this gene. Strain IIIJ3-1 cells showed alterations in cell morphology and indicated their ability to accumulate As forming electron opaque dots. The As accumulating As3+ stressed cells revealed lowering of surface area: volume ratio as a protective mechanism against toxicity of As so as to lower the available attachment/ uptake sites in effect [75, 76]. Relative decrease in cell surface to volume ratio plays the key role in the consequent reduction in attachment/uptake sites on the cell surface for the heavy metals like As in the case of strain IIIJ3-1 [76, 77]. Such stress responses upon exposure to toxic metals, metalloids and organics, other unfavourable conditions like highly acidic or alkaline pH, suboptimal temperatures, is believed to protect vital processes and restore cellular homeostasis, as well as help to enhance cellular resistance against subsequent stress challenges [75, 78, 79]. However, relative decrease in cellular dimensions but, increase in surface area: volume ration in As5+ stressed cells suggest an increase in exposure of various other pores or attachment sites to enable the cellular uptake of essential nutrients for cell survival. Cell constriction and appearance of rough cell surface might alter the cellular uptake sites. This might be suggestive of another resistance mechanism towards As stress. Expression of RND proteins (resistance, nodulation and division) for cell survival under stressed condition has been reported for gram negative bacteria [75, 80]. Arsenic accumulation studied through electron microscopy and EDX analysis within microbial cells belonging to Firmicutes, Actinobacteria and Proteobacteria have been reported in several studies previously [43, 66, 69, 81, 82]. Heavy metal (As, Cd, Zn, Hg, Pb) stress induced changes in cellular morphology have been noticed for many heterotrophic bacterial taxa viz. Acidocella sp. GS19h [83], Acidiphilium symbioticum H8 [75], Bacillus members (B. arsenicus, B. pumilus, B. indicus) [76] and Bacillus sp. strain XZM002 [43]. Substantial reduction in cell sizes has been ascribed to protective mechanism of the bacterial strain to cope up with metal toxicity. Overexpression of a metalloregulatory protein ArsR with high affinity for As3+ in E.coli showed increased bioaccumulation which allowed 100% removal of 5- ppb of As3+ from water [84], Another bacterium Pseudomonas stutzeri could accumulate As upto4 mg/gram of biomass (dry weight) [85]. Intracellular As accumulation has also been confirmed through TEM and EDAX analysis in isolates belonging to γ-Proteobacteria, Firmicutes and Kocuria [66]. However, none of the isolates have been shown to accumulate As during their anaerobic growth with As3+.Formation of small spheres composed of Se and encapsulated with polymeric material on the periphery of the cell envelope of B. selenitireducens on growth with Se4+ has been reported [46]. However, formation of electron opaque dots specifically following growth on the reduced species of As i.e. As3+ by strain IIIJ3-1 is a characteristic never reported earlier for any other bacteria. This could possibly indicate a novel mechanism to sustain the toxic effects of As3+ wherein, the bacteria complexes the inorganic ion in immobilized particles thus making it unavailable to the cellular machinery. It might also reflect that the cells upon As3+ oxidation are incapable of extruding As5+ and thus accumulate As within, whereas, while growing with As5+, As3+ formed from its reduction could be easily extruded out of the cell. Presence of As3+ extruding genes further supports this observation. Since the aio positive and arsC negative IIIJ3-1 bacterial cells were found to accumulate As only under As3+ stress, it proves that As3+ upon oxidation to As5+ cannot be further extruded out of the cell due to the lack of respective cellular machinery as also reported by Joshi et al. [86]. Moreover, As biotransformation assay for strain IIIJ3-1 has also shown very high Km for cytosolic As reduction implying presence of some non-specific reductase. This is also explained by Yang et al., [87] in Sinorhizobium meliloti, As5+ is reduced to As3+ via arsC, and AqpS confers resistance by downhill efflux of internally generated As3+. But in case of IIIJ3-1, the reduction of cytosolic As5+ does not occur/ or occur non-specifically due to absence of arsC gene. However, the threshold of As5+ accumulation has not been calculated and is an important aspect to be considered for our future scope of research. Several bacterial members have been known to accumulate As within their cellular components during growth under As5+ reducing and As3+ oxidizing condition upto varying extents. When incubated with 5 mg/L of As5+Marinomonas communis accumulated 2290 μg As/g dry weight, the highest value reported in bacteria [88]. Many other bacterial members reported to accumulate 110–765 μgAs/g dry wt. [84, 89, 90]. The highest amount of As accumulation reported is 765 μg/g dry wt. in E. coli without ars operon and this was ascribed to the inefficiency of this bacterium to extrude As out of cell during growth [84]. In one of the Bacillus sp. strain DJ-1, the lack of arsC gene and arsenate reductase activity was noted with 80% of As accumulation in cytoplasm, where a DNA protection during starvation (DPS) protein was reported to involve in As-binding in the cytoplasm to reduce the intracellular As toxicity [85]. It has been shown that for Marinomonas communis, ~45 % of total As was incorporated into the cytosol, 10 % lipid-bound in the membrane, and rest 45 % adsorbed onto the cell surface [88]. Although the details of the compartmentalization has not been studied in detail, the accumulation of As has been ascribed to be the result of higher uptake and lower efflux by the bacterium. Similarly approximately 80 % of the total accumulated As (11.5 % of total) was adsorbed onto the membrane instead of into cytoplasm of Bacillus sp. XZM002 from As-rich Datong basin, China [91]. However, EDX analysis shows maximum content of As under anaerobic growth with As3+. This might indicate that cells grown anaerobically with As3+ might have utilised As3+ as TEA as reduction of As3+ to As0 or As3- has been reported to be energetically favourable [68].
Conclusion
Arsenic contamination in the Brahmaputra river basin is a natural calamity which has degraded the potability of groundwater of various areas in this region. Very few reports are available to understand the microbiology of these aquifers wherein, recently Das and Barooah [38] have documented the siderophore based role of an As resistant Staphylococcus strain TA6 in As mobilization in these valleys. However, immediate concern is imperative to remediate the groundwater of these areas to restore its potability. This study presents a detailed molecular, chemotaxonomic, biochemical and ecophysiological characterization of a novel member of B. cereus strain IIIJ3-1 capable of efficiently transforming and accumulating As3+. Evidently, based on its chemotaxonomic, genomic and metabolic properties, strain IIIJ3-1 represents a novel and non-clonal member of B. cereus group which can act as potential As3+ accumulator in As contaminated regions. Based on its capability to survive and accumulate the most toxic form of inorganic As (As3+) within its system, the bacterial strain IIIJ3-1 offers a novel mechanism of As3+ remediation in As- contaminated groundwater as well as heavily contaminated irrigational fields as found in West Bengal [92]. Further investigation is called for standardizing and designing of bioremedial procedures using this highly efficient As biosequestering strain Bacillus IIIJ3-1.
Methods
Bacterial strains and growth condition
Strain IIIJ3-1 (JCM 31241(T)) was previously isolated [93], sub-cultured and routinely maintained on Luria Bertani (LB) agar or Reasoner’s 2A (R2A) agar, unless otherwise indicated. Type strain B. cereus ATCC 14579 was obtained from ATCC, cultured on appropriate medium, and used for comparison of phenotypic, chemotaxonomic and genotypic characteristics.
16S rRNA gene sequencing and phylogenetic analysis of strain IIIJ3–1
The genomic DNA of strain IIIJ3–1 was extracted using Pure Link genomic DNA kit (Invitrogen). The 16S rRNA gene was amplified by PCR with bacterial universal primers (27Fand1492R) following PCR conditions as mentioned in [39]. The PCR master mix was prepared according to the manufacturer’s protocol (NEB). The PCR cycle composed of an initial denaturation at 95 °C for 5 min followed by 30 cycles of denaturation at 95 °C for 1 min, annealing at 58 °C for 1.5 min, an extension at 72 °C for 50 s and a final extension at 72 °C for 1 min. The PCR products were gel purified using a Qia-quick gel extraction kit (QIAGEN), cloned into pTZ57R/T vector (InsTA clone kit, Thermo scientific), and sequenced using internal primers: 27F, 341F, 811F and 1492R to obtain near complete sequence of 16S rRNA. Homology search for maximum similarity of the 16S rRNA gene sequence of strain IIIJ3–1 was carried out using identity tool of EzTaxon-e server (https://www.ezbiocloud.net/identify) and NCBI database. The Ez Taxon showed % similarity with reported type strains and similarity with type and non-type strains were obtained from NCBI database. Multiple alignments were performed with CLUSTALW package of MEGA 7.0 software [94] by removing gaps from the sequences. Phylogenetic reconstruction and validation were performed using neighbor-joining (NJ) method [95] based on bootstrap analysis with 1000 replications using Jukes-Cantor [96] distance model. Both maximum-likelihood (ML) [97] and minimum-evolution (ME) [98] methods were employed to test the robustness of the trees.
Determination of molar G + C % and DNA-DNA relatedness
Molar G+C content (%) of strain IIIJ3-1was determined using the thermal denaturation method [57, 99, 100] where Escherichia coli K-12 NCIM 2563 used as the internal standard. DNA-DNA relatedness of strain IIIJ3-1 with its nearest phylogenetic neighbor B.cereus ATCC 14579(T) was measured using the SyBr green binding fluorimetry based method and relative binding ratio (RBR) was calculated following the correlation equation: y = (-)5.051x + 90.329 relating DNA - DNA relatedness and ∆Tm [100].
Chemotaxonomic (fatty acid methyl esters and quinone content) analysis
Total cellular fatty acid methyl ester (FAME) profiles of strain IIIJ3-1 and its closest phylogenetic neighbors (type strains) were determined by the growing cells of respective strains on LB medium for 24h at 30 °C [101]. Cellular fatty acids were saponified, methyl-esterified, and extracted according to the protocol of the Sherlock Microbial Identification System (MIDI). The FAMEs were analyzed using Gas Chromatograph (GC, Clarus 500, PerkinElmer) and compared with standard bacterial acid methyl ester mix (BAME, Sigma) for their identity. Isoprenoid quinines were extracted following the protocol mentioned by Dispirito et al. [102] separated using a SB-C18 Zorbax reverse-phase column fixed to a High Pressure Liquid Chromatograph (1100 Series, Agilent) with a solvent system of methanol: isopropanol (75:25 v/v) maintaining a flow rate of 1ml/min. Fractions corresponding distinct peaks at 2.9, 4.3 and 8.3 minutes were collected and concentrated in an evaporator. Molecular masses of the constituent quinones were analyzed by 4800 MALDI TOF/TOF analyzer (Applied BiosystemsInc., USA) using sinapinic acid as matrix. Constituent menaquinones were analysed by comparing the molecular masses in NCBI PubChem (https://pubchem.ncbi.nlm.nih.gov/).
Phenotypic and metabolic characterization of strain IIIJ3–1
Culture characteristics of strain IIIJ3-1 were observed by growing cells on LB agar plates for 24 h at 30°C.Cellular morphology was examined by scanning electron microscope (SEM, Zeiss, Evo 60), after harvesting cells from mid-exponential growth phase. For SEM, cells were washed with phosphate buffer (1X), fixed with 0.25% glutaraldehyde (Sigma) and para-formaldehyde (4%, v/v) in PBS at 4 °C, serially dehydrated (30-100% ethanol, v/v), spotted onto poly-L-Lysine coated cover slips, dried, and viewed after gold coating. Motility was determined by the hanging-drop technique [103]. Gram staining was performed using a Gram stain kit according to the manufacturer’s instructions (Hi-Media). Catalase and oxidase tests were performed by testing bubble formation ability of the isolate on treatment with H2O2 (30%) and using oxidase reagent (Biomerieux) according to the manufacturer’s protocol, respectively. Sensitivity towards range of temperatures (5-50 °C, at the interval of 5 °C), pH (5.0-12.0, at the interval of 1 pH unit), NaCl concentrations (0-8%, at the interval of 0.5 %) was evaluated by growing cells of strain IIIJ3-1 in LB broth for 24-48 h. Cell growth under the varying temperature, pH and osmolarity was monitored by recording the culture absorbance (OD at 600 nm) using a UV-Visible spectrophotometer (Cary 50, Varian). Resistance towards As species (As3+ and As5+) was tested following growth of cells of strain IIIJ3-1 in LB broth amended with graded concentrations of As3+ (NaAsO2; 0.1-30 mM) and As5+ (Na2HAsO4; 1-600 mM). The highest concentration of As species, up to which growth was observed was considered as maximum tolerable concentration (MTC). Medium without As was treated as control. Assimilation of broad range carbon and nitrogen substrates, susceptibility towards various antibiotics, and ability to withstand metabolic inhibitors were tested using the Biolog system (GEN-III MicroPlate, Biolog) following the manufacturer’s instructions. Statistical analysis involving the phenotypic and biochemical properties of strain IIIJ3-1 and other Bacillus spp. were performed using hierarchical cluster analysis through unweighted paired group arithmetic mean (UPGMA) calculation using Euclidean distance matrix with complete linkage algorithm in Multi-variate statistical package (MVSP) software. Linear regression and correlation analyses involving cellular growth profile and substrate reduction kinetics were performed using Excel 2010 and Minitab Statistical software 17.0.
Test for arsenic transformation by resting cells of strain IIIJ3–1
Arsenic biotransformation assay
Kinetic assay for As3+ oxidase and cytosolic As5+ reductase activities of the strain IIIJ3-1 was performed under aerobic condition. Cells of strain IIIJ3-1 were grown in LB medium (300 ml) devoid of any As until mid-log phase (O.D600= 1.3). The cells were harvested and washed thrice with reaction buffer, 1X TE (Tris,1M; EDTA,100mM), re-suspended in 10 ml of reaction buffer with concentration gradient of As3+ or As5+(1-5 mM) and allowed to stand for24 h at 30°C. Concentrations of As3+ and As5+ were measured following molybdenum blue method [104] at regular intervals. All the experiments were repeated thrice with each set in duplicates and mean of all results were used to calculate the kinetic parameters. Kinetic parameters, Km, and Vmax were calculated by plotting respective Line-Weaver Burk plots.
Test for use of different electron acceptors by strain IIIJ3–1
Anaerobic growth with alternate terminal electron acceptors (TEAs) viz. As5+, Se6+, Fe3+, S2O32-, NO3-,and SO42- (1 mM each) in reduced LB medium was monitored by observing absorbance at 600 nm using a UV spectrophotometer (Cary win UV, Agilent Technologies) and colony forming units (CFU) per ml at definite intervals till 54 h. Cysteine-HCl (0.1% as reducing agent) and resazurin (0.1 mg/L as redox indicator) were added to the degassed medium and the vials were crimp sealed. This procedure was followed throughout the work for setting up anaerobic growth for various experiments. Utilization of various electron donors during anaerobic growth with As5+as TEA was tested by growing strain IIIJ3-1 in reduced minimal salt medium (MSM) [Composition (g/L): NH4Cl (0.535); KH2PO4 (0.136); MgCl2. 6H2O (0.204); CaCl2.2H2O (0.147); Na2MoO4 (0.01); Cys-HCl 0.1%, v/v] supplemented with alternate electron donors (10mM). Growth was monitored by measuring the culture turbidity at 660 nm after 15 days. Anaerobic reduction profile of strain IIIJ3-1 with alternate TEAs: As5+, SO42- (1 mM) and Fe3+, NO3- (5 mM) was also studied in MSM with lactate as the sole electron donor by measuring the concentrations of As5+, SO42-, NO3- and Fe2+ produced from Fe3+ reduction [104–107] at an interval of 3 h. A vial without any electron donor and acceptor was subjected to similar growth conditions which served as the control.
Scanning electron microscopy (SEM) and energy-dispersive X-ray spectroscopy (EDX)
For visualization of the structural changes of the cells of strain IIIJ3-1 while growing under different As amended conditions (24 hr), scanning electron microscopy (SEM) was performed following the same protocol mentioned before (as described in the phenotypic characterization). In order to confirm the cellular As-uptake and -accumulation, energy dispersive x-ray spectroscopy (EDX) were performed using EDX analyzer (Oxford Instruments) in conjunction with SEM microscope. The morphometric analysis of the strain IIIJ3-1 grown with and without As stress was done by calculating the cell volume (V) and surface area (A) using the following equations:
Where, A is the surface area (μm2), V is the volume (μm3), r is the radius and h is the length of the cell. The average dimensions of the non-dividing cells in SEM image presented have been calculated [108].
Analysis of functional genes related to as transformation
Genes responsible for As3+ oxidation (aioB), cytosolic As5+ reduction (arsC), respiratory As5+ reduction (arrA), arsenite efflux pumps (arsB) and (acr3) were targeted for PCR amplification from genome of strain IIIJ3-1 by colony lysis method using reported sets of primers and PCR conditions [13]. The desired amplified fragments were cloned in pTZRT57 (Insta cloning kit, Fermentas), sequenced using M13F and T7R primers, and the obtained nucleotide sequences were searched for similarity level using BLASTN. The corresponding nucleotides were translated to amino acids in ExPasy tool (https://web.expasy.org/translate/) using appropriate open reading frames (ORFs) and searched in BLASTP, (nr database) excluding options for uncultured/environmental sequences. Conserved domain was predicted through CDD database and phylogeny was inferred through Maximum Likelihood tree construction using MEGA 7.0 considering the translated amino acid sequence of strain IIIJ3-1 and similar sequences retrieved from NCBI Multiple alignments of deduced amino acid sequences was done using Clustal Omega and ESPript [109] to decipher the conserved/consensus active site residues for each functional gene.
Nucleotide sequence accession numbers
The nucleotide sequences of all the functional genes of strain IIIJ3-1 have been submitted to NCBI-Genbank under accession numbers: KJ769230 (16S rRNA gene), and KY024786- KY024788 (As-responsive genes), respectively.
Supplementary information
Acknowledgements
Authors are thankful to Prof. Madhumita Barooah, Assam Agricultural University, Jorhat (Assam) for her kind help during sample collection.
Abbreviations
- BAME
Bacterial Acid Methyl Ester
- BDP
Bengal Delta Plain
- BLAST
Basic Local Alignment Search Tool
- BRB
Brahmaputra River Basin
- CFU
Colony Forming Unit
- DARB
Dissimilatory As5+ Respiring Bacteria
- DPS
DNA Protection during Starvation
- EDX
Energy Dispersive X-ray
- FAME
Fatty Acid Methyl Ester
- GC
Gas Chromatography
- LB
Luria Bertani
- MALDI TOF-MS
Matrix Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry
- MCC
Microbial Culture Collection
- ME
Minimum Evolution
- MEGA
Molecular Evolutionary and Genetic Analysis
- MK
Menaquinone
- ML
Maximum Likelihood
- MTC
Maximum Tolerable Concentration
- MTCC
Microbial Type Culture Collection
- MVSP
Multivariate Statistical Package
- NCBI
National Centre for Biotechnology Information
- NJ
Neighbour Joining
- OD
Optical Density
- ORF
Open Reading Frame
- PCR
Polymerase Chain Reaction
- R2A
Reasoner’s 2A
- RBR
Relative Binding Ratio
- RND
Resistance, Nodulation and Division
- SEM
Scanning Electron Microscopy
- TEA
Terminal Electron Acceptor
- TEM
Transmission Electron Microscopy
- UPGMA
Unweighted Paired Group Arithmetic Mean
Authors’ contributions
Conceptualization of the project: PS; Investigation: SG; Formal Analysis: SG and BM,Draft Preparation, Review and Edit: SG, BM, PS and TS; Supervision: PS. All the authors have read and approved the final manuscript.
Authors’ information
Soma Ghosh, Ph. D, is currently working as DBT BioCaRe Scientist at CSIR-National Environmental Engineering Research Institute, Kolkata Zonal Centre, India. Balaram Mohapatra, Ph. D, is a post-doctoral fellow at Department of Biosciences and Bioengineering, Indian Institute of Technology Bombay, Mumbai, India. Tulasi Satyanarayana, Ph. D, is presently affiliated to Department of Biological Sciences and Engineering, Netaji Subhas University of Technology, Sector 3,Dwarka, New Delhi, India. Pinaki Sar, Ph. D, is affiliated to Departrment of Biotechnology, Indian Institute of Technology Kharagpur, Kharagpur, India.
Funding
This work was financially supported by NER Twinning Project (EMA) (Ref No.BT/19/NE/TBP/2010), Department of Biotechnology (DBT), Govt. of India. Financial support to Soma Ghosh by IIT Kharagpur (IIT/ACAD/PGSandR/F.112//11/BT/92P03) institutional fellowship and the fellowship to Balaram Mohapatra provided by INSPIRE fellowship scheme of Department of Science and Technology (DST), Govt. of India, fellowship number IF120832 is duly acknowledged.
Availability of data and materials
The nucleotide sequences of the genes analysed are submitted in the NCBI Genbank. Strain IIIJ3-1 has been submitted to three culture collections- MCC 2980, LMG 29433, JCM 31241.
Ethics approval and consent to participate
Not Applicable.
Consent for publication
Not Applicable.
Competing interests
All the authors declare no competing interest.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information accompanies this paper at 10.1186/s12866-020-01893-6.
References
- 1.Fendorf S, Michael HA, van Geen A. Spatial and temporal variations of groundwater arsenic in south and Southeast Asia. Science. 2010;328(5982):1123–1127. doi: 10.1126/science.1172974. [DOI] [PubMed] [Google Scholar]
- 2.Stuckey JW, Schaefer MV, Kocar BD, Benner SG, Fendorf S. Arsenic release metabolically limited to permanently water-saturated soil in Mekong Delta. Nat Geosci. 2016;9(1):70–76. [Google Scholar]
- 3.Saha D, Sahu S. A decade of investigations on groundwater arsenic contamination in middle ganga plain, India. Environ Geochem Health. 2016;38(2):315–337. doi: 10.1007/s10653-015-9730-z. [DOI] [PubMed] [Google Scholar]
- 4.Chakraborti D, Rahman MM, Das B, Chatterjee A, Das D, Nayak B, Pal A, Chowdhury UK, Ahmed S, Biswas BK, Sengupta MK. Groundwater arsenic contamination and its health effects in India. Hydrogeol J. 2017;25(4):1165–1181. [Google Scholar]
- 5.Smedley PL, Kinniburgh DG. A review of the source, behaviour and distribution of arsenic in natural waters. Appl Geochem. 2002;17(5):517–568. [Google Scholar]
- 6.Malasarn D, Keeffe JR, Newman DK. Characterization of the arsenate respiratory reductase from Shewanella sp. strain ANA-3. J Bacteriol. 2008;190(1):135–142. doi: 10.1128/JB.01110-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Blum JS, Han S, Lanoil B, Saltikov C, Witte B, Tabita FR, Langley S, Beveridge TJ, Jahnke L, Oremland RS. Ecophysiology of “Halarsenatibacter silvermanii” strain SLAS-1T, gen. Nov., sp. nov., a facultative chemoautotrophic arsenate respirer from salt-saturated Searles Lake, California. Appl Environ Microbiol. 2009;75(7):1950–1960. doi: 10.1128/AEM.02614-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ohtsuka T, Yamaguchi N, Makino T, Sakurai K, Kimura K, Kudo K, Homma E, Dong DT, Amachi S. Arsenic dissolution from Japanese paddy soil by a dissimilatory arsenate-reducing bacterium Geobacter sp. OR-1. Environ Sci Technol. 2013;47(12):6263–6271. doi: 10.1021/es400231x. [DOI] [PubMed] [Google Scholar]
- 9.Zhu YG, Yoshinaga M, Zhao FJ, Rosen BP. Earth abides arsenic biotransformations. Annu Rev Earth Planet Sci. 2014;42:443–467. doi: 10.1146/annurev-earth-060313-054942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Saunders JA, Lee MK, Uddin A, Mohammad S, Wilkin RT, Fayek M, Korte NE. Natural arsenic contamination of Holocene alluvial aquifers by linked tectonic, weathering, and microbial processes. Geochem Geophys Geosyst. 2005;6(4). 10.1029/2004GC000803.
- 11.Mukherjee A, Bhattacharya P, Savage K, Foster A, Bundschuh J. Distribution of geogenic arsenic in hydrologic systems: controls and challenges. 2008. pp. 1–7. [DOI] [PubMed] [Google Scholar]
- 12.Neidhardt H, Berner ZA, Freikowski D, Biswas A, Majumder S, Winter J, Gallert C, Chatterjee D, Norra S. Organic carbon induced mobilization of iron and manganese in a West Bengal aquifer and the muted response of groundwater arsenic concentrations. Chem Geol. 2014;367:51–62. [Google Scholar]
- 13.Paul D, Kazy SK, Gupta AK, Pal T, Sar P. Diversity, metabolic properties and arsenic mobilization potential of indigenous bacteria in arsenic contaminated groundwater of West Bengal, India. PloS One. 2015;10(3):e0118735. doi: 10.1371/journal.pone.0118735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huang JH. Impact of microorganisms on arsenic biogeochemistry: a review. Water Air Soil Pollution. 2014;225(2):1848. [Google Scholar]
- 15.Crognale S, Amalfitano S, Casentini B, Fazi S, Petruccioli M, Rossetti S. Arsenic-related microorganisms in groundwater: a review on distribution, metabolic activities and potential use in arsenic removal processes. Rev Environ Sci Biotechnol. 2017;16(4):647–665. [Google Scholar]
- 16.Mohapatra B, Sarkar A, Joshi S, Chatterjee A, Kazy SK, Maiti MK, Satyanarayana T, Sar P. An arsenate-reducing and alkane-metabolizing novel bacterium, Rhizobium arsenicireducens sp. nov., isolated from arsenic-rich groundwater. Arch Microbiol. 2017;199(2):191–201. doi: 10.1007/s00203-016-1286-5. [DOI] [PubMed] [Google Scholar]
- 17.Liao VH, Chu YJ, Su YC, Hsiao SY, Wei CC, Liu CW, Liao CM, Shen WC, Chang FJ. Arsenite-oxidizing and arsenate-reducing bacteria associated with arsenic-rich groundwater in Taiwan. J Contam Hydrol. 2011;123(1–2):20–29. doi: 10.1016/j.jconhyd.2010.12.003. [DOI] [PubMed] [Google Scholar]
- 18.Davolos D, Pietrangeli B. A molecular study on bacterial resistance to arsenic-toxicity in surface and underground waters of Latium (Italy) Ecotoxicol Environ Saf. 2013;96:1–9. doi: 10.1016/j.ecoenv.2013.05.039. [DOI] [PubMed] [Google Scholar]
- 19.Sarkar A, Kazy SK, Sar P. Characterization of arsenic resistant bacteria from arsenic rich groundwater of West Bengal, India. Ecotoxicology. 2013;22(2):363–376. doi: 10.1007/s10646-012-1031-z. [DOI] [PubMed] [Google Scholar]
- 20.Corsini A, Zaccheo P, Muyzer G, Andreoni V, Cavalca L. Arsenic transforming abilities of groundwater bacteria and the combined use of Aliihoeflea sp. strain 2WW and goethite in metalloid removal. J Hazard Mater. 2014;269:89–97. doi: 10.1016/j.jhazmat.2013.12.037. [DOI] [PubMed] [Google Scholar]
- 21.Li P, Wang Y, Dai X, Zhang R, Jiang Z, Jiang D, Wang S, Jiang H, Wang Y, Dong H. Microbial community in high arsenic shallow groundwater aquifers in Hetao Basin of Inner Mongolia, China. PLoS One. 2015;10(5):e0125844. doi: 10.1371/journal.pone.0125844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fazi S, Amalfitano S, Casentini B, Davolos D, Pietrangeli B, Crognale S, Lotti F, Rossetti S. Arsenic removal from naturally contaminated waters: a review of methods combining chemical and biological treatments. RendicontiLincei. 2016;27(1):51–58. [Google Scholar]
- 23.Das S, Bora SS, Yadav RN, Barooah M. A metagenomic approach to decipher the indigenous microbial communities of arsenic contaminated groundwater of Assam. Genomics data. 2017;12:89–96. doi: 10.1016/j.gdata.2017.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Santini JM, Stolz JF, Macy JM. Isolation of a new arsenate- respiring bacterium--physiological and phylogenetic studies. Geomicrobiol J. 2002;19(1):41–52. [Google Scholar]
- 25.Dowdle PR, Laverman AM, Oremland RS. Bacterial dissimilatory reduction of arsenic (V) to arsenic (III) in anoxic sediments. Appl Environ Microbiol. 1996;62(5):1664–1669. doi: 10.1128/aem.62.5.1664-1669.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Saltikov CW, Olson BH. Homology of Escherichia coli R773 arsA, arsB, and arsC genes in arsenic-resistant bacteria isolated from raw sewage and arsenic-enriched creek waters. Appl Environ Microbiol. 2002;68(1):280–288. doi: 10.1128/AEM.68.1.280-288.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bentley R, Chasteen TG. Microbial methylation of metalloids: arsenic, antimony, and bismuth. Microbiol Mol Biol Rev. 2002;66(2):250–271. doi: 10.1128/MMBR.66.2.250-271.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Oremland RS, Stolz JF. The ecology of arsenic. Science. 2003;300(5621):939–944. doi: 10.1126/science.1081903. [DOI] [PubMed] [Google Scholar]
- 29.Saltikov CW, Cifuentes A, Venkateswaran K, Newman DK. The ars detoxification system is advantageous but not required for as (V) respiration by the genetically tractable Shewanella species strain ANA-3. Appl Environ Microbiol. 2003;69(5):2800–2809. doi: 10.1128/AEM.69.5.2800-2809.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Newman DK, Ahmann D, Morel FM. A brief review of microbial arsenate respiration. Geomicrobiol J. 1998;15(4):255–268. [Google Scholar]
- 31.Lukasz D, Liwia R, Aleksandra M, Aleksandra S. Dissolution of arsenic minerals mediated by dissimilatory arsenate reducing bacteria: estimation of the physiological potential for arsenic mobilization. Biomed Res Int. 2014;2014:841892. doi: 10.1155/2014/841892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Osborne TH, McArthur JM, Sikdar PK. SantiniJM. Isolation of an arsenate-respiring bacterium from a redox front in an arsenic-polluted aquifer in West Bengal, Bengal Basin. Environ Sci Technol. 2015;49(7):4193–4199. doi: 10.1021/es504707x. [DOI] [PubMed] [Google Scholar]
- 33.Kudo K, Yamaguchi N, Makino T, Ohtsuka T, Kimura K, Dong DT, Amachi S. Release of arsenic from soil by a novel dissimilatory arsenate-reducing bacterium, Anaeromyxobacter sp. strain PSR-1. Appl Environ Microbiol. 2013;79(15):4635–4642. doi: 10.1128/AEM.00693-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mohapatra B, Kazy SK, Sar P. Comparative genome analysis of arsenic reducing, hydrocarbon metabolizing groundwater bacterium Achromobacter sp. KAs 3-5T explains its competitive edge for survival in aquifer environment. Genomics. 2019;111(6):1604–1619. doi: 10.1016/j.ygeno.2018.11.004. [DOI] [PubMed] [Google Scholar]
- 35.Mohapatra B, Sar P, Kazy SK, Maiti MK, Satyanarayana T. Taxonomy and physiology of Pseudoxanthomonas arseniciresistens sp. nov., an arsenate and nitrate-reducing novel gamma-proteobacterium from arsenic contaminated groundwater, India. PLoS One. 2018;13(3):e0193718. doi: 10.1371/journal.pone.0193718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shivaji S, Suresh K, Chaturvedi P, Dube S, Sengupta S. Bacillus arsenicus sp. nov., an arsenic-resistant bacterium isolated from a siderite concretion in West Bengal, India. Int J Syst Evol Microbiol. 2005;55(3):1123–1127. doi: 10.1099/ijs.0.63476-0. [DOI] [PubMed] [Google Scholar]
- 37.Suresh K, Prabagaran SR, Sengupta S, Shivaji S. Bacillus indicus sp. nov., an arsenic-resistant bacterium isolated from an aquifer in West Bengal, India. Int J Syst Evol Microbiol. 2004;54(4):1369–1375. doi: 10.1099/ijs.0.03047-0. [DOI] [PubMed] [Google Scholar]
- 38.Das S, Barooah M. Characterization of siderophore producing arsenic-resistant Staphylococcus sp. strain TA6 isolated from contaminated groundwater of Jorhat, Assam and its possible role in arsenic geocycle. BMC Microbiol. 2018;18(1):104. doi: 10.1186/s12866-018-1240-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ghosh S, Sar P. Identification and characterization of metabolic properties of bacterial populations recovered from arsenic contaminated ground water of north East India (Assam) Water Res. 2013;47(19):6992–7005. doi: 10.1016/j.watres.2013.08.044. [DOI] [PubMed] [Google Scholar]
- 40.Ghosh S, Gupta A, Sarkar J, Verma S, Mukherjee A, Sar P. Enrichment of indigenous arsenate reducing anaerobic bacteria from arsenic rich aquifer sediment of Brahmaputra river basin and their potential role in as mobilization. J Environ Sci Health A. 2019;54(7):635–647. doi: 10.1080/10934529.2019.1579524. [DOI] [PubMed] [Google Scholar]
- 41.Ghosh S, Sar P. Microcosm based analysis of arsenic release potential of Bacillus sp. strain IIIJ3–1 under varying redox conditions. World J Microbiol Biotechnol. 2020;36(87):87. doi: 10.1007/s11274-020-02860-z. [DOI] [PubMed] [Google Scholar]
- 42.Yamamura S, Yamashita M, Fujimoto N, Kuroda M, Kashiwa M, Sei K, Fujita M, Ike M. Bacillus selenatarsenatissp. Nov., a selenate-and arsenate-reducing bacterium isolated from the effluent drain of a glass-manufacturing plant. Int J Syst Evol Microbiol. 2007;57(5):1060–1064. doi: 10.1099/ijs.0.64667-0. [DOI] [PubMed] [Google Scholar]
- 43.Xie Z, Wang Y, Duan M, Xie X, Su C. Arsenic release by indigenous bacteria Bacillus cereus from aquifer sediments at Datong Basin, northern China. Front Earth Sci. 2011;5(1):37–44. [Google Scholar]
- 44.Majumder A, Bhattacharyya K, Bhattacharyya S, Kole SC. Arsenic-tolerant, arsenite-oxidising bacterial strains in the contaminated soils of West Bengal, India. Sci Total Environ. 2013;463:1006–1014. doi: 10.1016/j.scitotenv.2013.06.068. [DOI] [PubMed] [Google Scholar]
- 45.Dey U, Chatterjee S, Mondal NK. Isolation and characterization of arsenic-resistant bacteria and possible application in bioremediation. Biotechnol Rep. 2016;10:1–7. doi: 10.1016/j.btre.2016.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Blum JS, Bindi AB, Buzzelli J, Stolz JF. OremlandRS. Bacillus arsenicoselenatis, sp. nov., and Bacillus selenitireducens, sp. nov.: two haloalkaliphiles from mono Lake, California that respire oxyanions of selenium and arsenic. Arch Microbiol. 1998;171(1):19–30. doi: 10.1007/s002030050673. [DOI] [PubMed] [Google Scholar]
- 47.Yamamura S, Yamashita M, Fujimoto N, Kuroda M, Kashiwa M, Sei K, Fujita M, Ike M. Bacillus selenatarsenatis sp. nov., a selenate-and arsenate-reducing bacterium isolated from the effluent drain of a glass-manufacturing plant. Int J Syst Evol Microbiol. 2007;57(5):1060–1064. doi: 10.1099/ijs.0.64667-0. [DOI] [PubMed] [Google Scholar]
- 48.Pattanapipitpaisal P, Yodsing N, Santhaweesuk R, Wamakhan P. Arsenite oxidation and arsenite resistance by Bacillus sp. PNKP-S2. Environment Asia. 2015;8(1):9–15. [Google Scholar]
- 49.Bagade A, Nandre V, Paul D, Patil Y, Sharma N, Giri A, Kodam K. Characterisation of hyper tolerant Bacillus firmus L-148 for arsenic oxidation. Environ Pollut. 2020;261:114124. doi: 10.1016/j.envpol.2020.114124. [DOI] [PubMed] [Google Scholar]
- 50.Guo H, Liu Z, Ding S, Hao C, Xiu W, Hou W. Arsenate reduction and mobilization in the presence of indigenous aerobic bacteria obtained from high arsenic aquifers of the Hetao basin, Inner Mongolia. Environ Pollut. 2015;203:50–59. doi: 10.1016/j.envpol.2015.03.034. [DOI] [PubMed] [Google Scholar]
- 51.Jain S, Saluja B, Gupta A, Marla SS, Goel R. Validation of arsenic resistance in Bacillus cereus strain AG27 by comparative protein modeling of arsC gene product. Protein J. 2011;30(2):91–101. doi: 10.1007/s10930-011-9305-5. [DOI] [PubMed] [Google Scholar]
- 52.Jiménez G, Blanch AR, Tamames J, Rosselló-Mora R. Complete genome sequence of Bacillus toyonensis BCT-7112T, the active ingredient of the feed additive preparation Toyocerin. Genome Announc. 2013;1(6):e01080. doi: 10.1128/genomeA.01080-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Täubel M, Kämpfer P, Buczolits S, Lubitz W, Busse HJ. Bacillus barbaricus sp. nov., isolated from an experimental wall painting. Int J Syst Evol Microbiol. 2003;53(3):725–730. doi: 10.1099/ijs.0.02304-0. [DOI] [PubMed] [Google Scholar]
- 54.Cifré LC, Alemany M, de Mendoza D, Altabe S. Exploring the biosynthesis of unsaturated fatty acids in Bacillus cereus ATCC 14579 and functional characterization of novel acyl-lipid desaturases. Appl Environ Microbiol. 2013;79(20):6271–6279. doi: 10.1128/AEM.01761-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Baesman SM, Stolz JF, Kulp TR, Oremland RS. Enrichment and isolation of Bacillus beveridgei sp. nov., a facultative anaerobic haloalkaliphile from mono Lake, California, that respires oxyanions of tellurium, selenium, and arsenic. Extremophiles. 2009;13(4):695–705. doi: 10.1007/s00792-009-0257-z. [DOI] [PubMed] [Google Scholar]
- 56.Tiago I, Pires C, Mendes V, Morais PV, Da Costa MS, Veríssimo A. Bacillus foraminis sp. nov., isolated from a non-saline alkaline groundwater. Int J Syst Evol Microbiol. 2006;56(11):2571–2574. doi: 10.1099/ijs.0.64281-0. [DOI] [PubMed] [Google Scholar]
- 57.Wayne LG, Brenner DJ, Colwell RR, Grimont PA, Kandler O, Krichevsky MI, Moore LH, Moore WE, Murray R, Stackebrandt ES, Starr MP. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int J Syst Evol Microbiol. 1987;37(4):463–464. [Google Scholar]
- 58.Niskanen A, Kiutamo T, Räisänen S, Raevuori M. Determination of fatty acid compositions of Bacillus cereus and related bacteria: a rapid gas chromatographic method using a glass capillary column. Appl Environ Microbiol. 1978;35(2):453–455. doi: 10.1128/aem.35.2.453-455.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kaneda T. Fatty acids in the genus Bacillus II. Similarity in the fatty acid compositions of Bacillus thuringiensis, Bacillus anthracis, and Bacillus cereus. J Bacteriol. 1968;95(6):2210–2216. doi: 10.1128/jb.95.6.2210-2216.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Guinebretière MH, Auger S, Galleron N, Contzen M, De Sarrau B, De Buyser ML, Lamberet G, Fagerlund A, Granum PE, Lereclus D, De Vos P. Bacillus cytotoxicus sp. nov.is a novel thermotolerant species of the Bacillus cereus group occasionally associated with food poisoning. Int J Syst Evol Microbiol. 2013;63(1):31–40. doi: 10.1099/ijs.0.030627-0. [DOI] [PubMed] [Google Scholar]
- 61.Tebo BM, Obraztsova AY. Sulfate-reducing bacterium grows with Cr (VI), U (VI), Mn (IV), and Fe (III) as electron acceptors. FEMS Microbiol Lett. 1998;162(1):193–199. [Google Scholar]
- 62.McArthur JM, Banerjee DM, Hudson-Edwards KA, Mishra R, Purohit R, Ravenscroft P, Cronin A, Howarth RJ, Chatterjee A, Talukder T, Lowry D. Natural organic matter in sedimentary basins and its relation to arsenic in anoxic ground water: the example of West Bengal and its worldwide implications. Appl Geochem. 2004;19(8):1255–1293. [Google Scholar]
- 63.Héry M, Van Dongen BE, Gill F, Mondal D, Vaughan DJ, Pancost RD, Polya DA, Lloyd JR. Arsenic release and attenuation in low organic carbon aquifer sediments from West Bengal. Geobiology. 2010;8(2):155–168. doi: 10.1111/j.1472-4669.2010.00233.x. [DOI] [PubMed] [Google Scholar]
- 64.Verma S, Mukherjee A, Choudhury R, Mahanta C. Brahmaputra river basin groundwater: solute distribution, chemical evolution and arsenic occurrences in different geomorphic settings. J Hydrol. 2015;4:131–153. [Google Scholar]
- 65.Focardi S, Pepi M, Ruta M, Marvasi M, Bernardini E, Gasperini S, Focardi SE. Arsenic precipitation by an anaerobic arsenic-respiring bacterial strain isolated from the polluted sediments of Orbetello lagoon, Italy. Lett Appl Microbiol. 2010;51(5):578–585. doi: 10.1111/j.1472-765X.2010.02938.x. [DOI] [PubMed] [Google Scholar]
- 66.Banerjee S, Datta S, Chattyopadhyay D, Sarkar P. Arsenic accumulating and transforming bacteria isolated from contaminated soil for potential use in bioremediation. J Environ Sci Health A. 2011;46(14):1736–1747. doi: 10.1080/10934529.2011.623995. [DOI] [PubMed] [Google Scholar]
- 67.Mallick I, Hossain ST, Sinha S, Mukherjee SK. Brevibacillus sp. KUMAs2, a bacterial isolate for possible bioremediation of arsenic in rhizosphere. Ecotoxicol Environ Saf. 2014;107:236–244. doi: 10.1016/j.ecoenv.2014.06.007. [DOI] [PubMed] [Google Scholar]
- 68.Laverman AM, Blum JS, Schaefer JK, Phillips E, Lovley DR, Oremland RS. Growth of strain SES-3 with arsenate and other diverse electron acceptors. Appl Environ Microbiol. 1995;61(10):3556–3561. doi: 10.1128/aem.61.10.3556-3561.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Salmassi TM, Venkateswaren K, Satomi M, Newman DK, Hering JG. Oxidation of arsenite by Agrobacterium albertimagni, AOL15, sp. nov., isolated from Hot Creek, California. Geomicrobiol J. 2002;19(1):53–66. [Google Scholar]
- 70.Li R, Haile JD, Kennelly PJ. An arsenate reductase from Synechocystissp.Strain PCC 6803 exhibits a novel combination of catalytic characteristics. J Bacteriol. 2003;185(23):6780–6789. doi: 10.1128/JB.185.23.6780-6789.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lever MA. Acetogenesis in the energy-starved deep biosphere–a paradox? Front Microbiol. 2012;2:284. doi: 10.3389/fmicb.2011.00284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Newman DK, Kennedy EK, Coates JD, Ahmann D, Ellis DJ, Lovley DR, Morel FM. Dissimilatory arsenate and sulfate reduction in Desulfotomaculumauripigmentum sp. nov. Arch Microbiol. 1997;168(5):380–388. doi: 10.1007/s002030050512. [DOI] [PubMed] [Google Scholar]
- 73.Stolz JF, Ellis DJ, Blum JS, Ahmann D, Lovley DR, Oremland RS. Note: Sulfurospirillumbarnesii sp. nov.andSulfurospirillumarsenophilum sp. nov., new members of the Sulfurospirillum clade of the ε-Proteobacteria. Int J Syst Evol Microbiol. 1999;49(3):1177–1180. doi: 10.1099/00207713-49-3-1177. [DOI] [PubMed] [Google Scholar]
- 74.Niggemyer A, Spring S, Stackebrandt E, Rosenzweig RF. Isolation and characterization of a novel as (V)- reducing bacterium: implications for arsenic mobilization and the genus Desulfitobacterium. Appl Environ Microbiol. 2001;67(12):5568–5580. doi: 10.1128/AEM.67.12.5568-5580.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chakravarty R, Banerjee PC. Morphological changes in an acidophilic bacterium induced by heavy metals. Extremophiles. 2008;12(2):279–284. doi: 10.1007/s00792-007-0128-4. [DOI] [PubMed] [Google Scholar]
- 76.Nithya C, Devi MG, Karutha PS. A novel compound from the marine bacterium Bacillus pumilus S6-15 inhibits biofilm formation in gram-positive and gram-negative species. Biofouling. 2011;27(5):519–528. doi: 10.1080/08927014.2011.586127. [DOI] [PubMed] [Google Scholar]
- 77.Campbell KM, Malasarn D, Saltikov CW, Newman DK, Hering JG. Effect of sorbed arsenic species on bacterial reduction of HFO. GeCAS. 2005;69(10):A357. [Google Scholar]
- 78.Ramos JL, Gallegos MT, Marqués S, Ramos-González MI, Espinosa-Urgel M, Segura A. Responses of gram-negative bacteria to certain environmental stressors. Curr Opin Microbiol. 2001;4(2):166–171. doi: 10.1016/s1369-5274(00)00183-1. [DOI] [PubMed] [Google Scholar]
- 79.Young KD. The selective value of bacterial shape. Microbiol Mol Biol Rev. 2006;70(3):660–703. doi: 10.1128/MMBR.00001-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Nies DH. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol Rev. 2003;27(2–3):313–339. doi: 10.1016/S0168-6445(03)00048-2. [DOI] [PubMed] [Google Scholar]
- 81.Giri AK, Patel RK, Mahapatra SS, Mishra PC. Biosorption of arsenic (III) from aqueous solution by living cells of Bacillus cereus. Environ Sci Pollut Res. 2013;20(3):1281–1291. doi: 10.1007/s11356-012-1249-6. [DOI] [PubMed] [Google Scholar]
- 82.Pandey N, Bhatt R. Arsenic resistance and accumulation by two bacteria isolated from a natural arsenic contaminated site. J Basic Microbiol. 2015;55(11):1275–1286. doi: 10.1002/jobm.201400723. [DOI] [PubMed] [Google Scholar]
- 83.Chakravarty R, Manna S, Ghosh AK, Banerjee PC. Morphological changes in an Acidocella strain in response to heavy metal stress. Res J Microbiol. 2007;2(10):742–748. [Google Scholar]
- 84.Kostal J, Yang R, Wu CH, Mulchandani A, Chen W. Enhanced arsenic accumulation in engineered bacterial cells expressing ArsR. Appl Environ Microbiol. 2004;70(8):4582–4587. doi: 10.1128/AEM.70.8.4582-4587.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Joshi DN, Patel JS, Flora SJ, Kalia K. Arsenic accumulation by Pseudomonas stutzeri and its response to some thiol-chelators. Environ Health Prev Med. 2008;13(5):257. doi: 10.1007/s12199-008-0038-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Joshi A, Dang HQ, Vaid N, Tuteja N. Pea lectin receptor-like kinase promotes high salinity stress tolerance in bacteria and expresses in response to stress in planta. Glycoconj J. 2010;27(1):133–150. doi: 10.1007/s10719-009-9265-6. [DOI] [PubMed] [Google Scholar]
- 87.Yang HC, Cheng J, Finan TM, Rosen BP, Bhattacharjee H. Novel pathway for arsenic detoxification in the legume symbiont Sinorhizobiummeliloti. J Bacteriol. 2005;187(20):6991–6997. doi: 10.1128/JB.187.20.6991-6997.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Takeuchi M, Kawahata H, Gupta LP, Kita N, Morishita Y, Ono Y, Komai T. Arsenic resistance and removal by marine and non-marine bacteria. J Biotechnol. 2007;127(3):434–442. doi: 10.1016/j.jbiotec.2006.07.018. [DOI] [PubMed] [Google Scholar]
- 89.Silver S, Budd K, Leahy KM, Shaw WV, Hammond D, Novick RP, Willsky GR, Malamy MH, Rosenberg H. Inducible plasmid-determined resistance to arsenate, arsenite, and antimony (III) in Escherichia coli and Staphylococcus aureus. J Bacteriol. 1981;146(3):983–996. doi: 10.1128/jb.146.3.983-996.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Sauge-Merle S, Cuiné S, Carrier P, Lecomte-Pradines C, Luu DT, Peltier G. Enhanced toxic metal accumulation in engineered bacterial cells expressing Arabidopsis thalianaphytochelatin synthase. Appl Environ Microbiol. 2003;69(1):490–494. doi: 10.1128/AEM.69.1.490-494.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Xie Z, Zhou Y, Wang Y, Luo Y, Sun X. Influence of arsenate on lipid peroxidation levels and antioxidant enzyme activities in Bacillus cereus strain XZM002 isolated from high arsenic aquifer sediments. Geomicrobiol J. 2013;30(7):645–652. [Google Scholar]
- 92.Chowdhury NR, Das R, Joardar M, Ghosh S, Bhowmick S, Roychowdhury T. Arsenic accumulation in paddy plants at different phases of pre-monsoon cultivation. Chemosphere. 2018;210:987–997. doi: 10.1016/j.chemosphere.2018.07.041. [DOI] [PubMed] [Google Scholar]
- 93.Ghosh S. Characterization of Bacterial Communities in Arsenic Contaminated Groundwater of Brahmaputra River Basin. 2018. [Google Scholar]
- 94.Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 96.Jukes T, Cantor CR. Evolution of protein molecules. In: Munro HN, editor. Mammalian Protein Metabolism. New York: Academic Press; 1969. p. 132. [Google Scholar]
- 97.Felsenstein J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol. 1981;17:368–376. doi: 10.1007/BF01734359. [DOI] [PubMed] [Google Scholar]
- 98.Takahashi K, Nei M. Efficiencies of fast algorithms of phylogenetic inference under the criteria of maximum parsimony, minimum evolution, and maximum likelihood when a large number of sequences are used. Mol Biol Evol. 2000;17:1251–1258. doi: 10.1093/oxfordjournals.molbev.a026408. [DOI] [PubMed] [Google Scholar]
- 99.Ley JD, Cattoir H, Reynaerts A. The quantitative measurement of DNA hybridization from renaturation rates. FEBS J. 1970;12(1):133–142. doi: 10.1111/j.1432-1033.1970.tb00830.x. [DOI] [PubMed] [Google Scholar]
- 100.Rosselló-Mora R, Amann R. The species concept for prokaryotes. FEMS Microbiol Rev. 2001;25(1):39–67. doi: 10.1111/j.1574-6976.2001.tb00571.x. [DOI] [PubMed] [Google Scholar]
- 101.Miller LT. Single derivatization method for routine analysis of bacterial whole-cell fatty acid methyl esters, including hydroxyacids. J Clin Microbiol. 1982;16(3):584–586. doi: 10.1128/jcm.16.3.584-586.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.DiSpirito AA, Loh WH, Tuovinen OH. A novel method for the isolation of bacterial quinones and its application to appraise the ubiquinone composition of Thiobacillusferrooxidans. Arch Microbiol. 1983;135(1):77–80. [Google Scholar]
- 103.Tittsler RP, Sandholzer LA. The use of semi-solid agar for the detection of bacterial motility. J Bacteriol. 1936;31(6):575. doi: 10.1128/jb.31.6.575-580.1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Bachate SP, Khapare RM, Kodam KM. Oxidation of arsenite by two β-proteobacteria isolated from soil. Appl Microbiol Biotechnol. 2012;93(5):2135–2145. doi: 10.1007/s00253-011-3606-7. [DOI] [PubMed] [Google Scholar]
- 105.Cataldo DA, Maroon M, Schrader LE, Youngs VL. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun Soil Sci Plant Anal. 1975;6(1):71–80. [Google Scholar]
- 106.Tabatabai MA. A rapid method for determination of sulfate in water samples. Environ Lett. 1974;7(3):237–243. [Google Scholar]
- 107.Viollier E, Inglett PW, Hunter K, Roychoudhury AN, Van Cappellen P. The ferrozine method revisited: Fe (II)/Fe (III) determination in natural waters. Appl Geochem. 2000;15(6):785–790. [Google Scholar]
- 108.Anderson CR, Cook GM. Isolation and characterization of arsenate-reducing bacteria from arsenic-contaminated sites in New Zealand. Curr Microbiol. 2004;48(5):341–347. doi: 10.1007/s00284-003-4205-3. [DOI] [PubMed] [Google Scholar]
- 109.Robert X, Gouet P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014;42(W1):W320–W324. doi: 10.1093/nar/gku316. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The nucleotide sequences of the genes analysed are submitted in the NCBI Genbank. Strain IIIJ3-1 has been submitted to three culture collections- MCC 2980, LMG 29433, JCM 31241.