

Abstract
WRKY transcription factors regulate diverse biological processes in plants, including abiotic and biotic stress responses, and constitute one of the largest transcription factor families in higher plants. Although the past decade has seen significant progress towards identifying and functionally characterizing WRKY genes in diverse species, little is known about the WRKY family in sorghum (Sorghum bicolor (L.) moench). Here we report the comprehensive identification of 94 putative WRKY transcription factors (SbWRKYs). The SbWRKYs were divided into three groups (I, II, and III), with those in group II further classified into five subgroups (IIa–IIe), based on their conserved domains and zinc finger motif types. WRKYs from the model plant Arabidopsis (Arabidopsis thaliana) were used for the phylogenetic analysis of all SbWRKY genes. Motif analysis showed that all SbWRKYs contained either one or two WRKY domains and that SbWRKYs within the same group had similar motif compositions. SbWRKY genes were located on all 10 sorghum chromosomes, and some gene clusters and two tandem duplications were detected. SbWRKY gene structure analysis showed that they contained 0–7 introns, with most SbWRKY genes consisting of two introns and three exons. Gene ontology (GO) annotation functionally categorized SbWRKYs under cellular components, molecular functions and biological processes. A cis-element analysis showed that all SbWRKYs contain at least one stress response-related cis-element. We exploited publicly available microarray datasets to analyze the expression profiles of 78 SbWRKY genes at different growth stages and in different tissues. The induction of SbWRKYs by different abiotic stresses hinted at their potential involvement in stress responses. qRT-PCR analysis revealed different expression patterns for SbWRKYs during drought stress. Functionally characterized WRKY genes in Arabidopsis and other species will provide clues for the functional characterization of putative orthologs in sorghum. Thus, the present study delivers a solid foundation for future functional studies of SbWRKY genes and their roles in the response to critical stresses such as drought.
Introduction
WRKY transcription factors (TFs), one of the largest TF families in plants, regulate various biological processes, including stress responses. WRKY proteins contain a conserved WRKYGQK motif at their N-terminus, along with a 60-amino-acid-long zinc finger motif at their C-terminus [1]. These two motifs are essential for the binding of WRKY TFs to the W-box cis-element [(T)TGAC(C/T)] located within the promoters of their target genes. WRKY proteins can be classified into three groups (I, II, and III) according to the number of WRKY domains and the type of zinc finger motif, i.e., C2H2 or C2HC [2]. Group I members have two WRKY domains and the C2H2-type zinc finger. Group II members have only one WRKY domain and a C2H2 zinc finger motif and can be further classified into five subgroups (IIa, IIb, IIc, IId, and IIe) based on the sequence of the DNA-binding domain. Finally, group III members have one WRKY domain and a C2HC-type zinc finger [3, 4].
Several studies have demonstrated that WRKY TFs regulate various biological processes and control gene expression via a combination of positive or negative regulation [5, 6]. WRKY TFs have been reported to be involved in responses to biotic stresses [7], developmental processes such as senescence, embryogenesis, and seed development, as well as abiotic stresses [8, 9]. For example, the wheat (Triticum aestivum) TaWRKY10 gene is considered to be a key regulator in salt and drought responses by regulating stress-responsive genes [10]. Heterologous expression of TaWRKY1 and TaWRKY33 enhanced drought and heat tolerance in Arabidopsis plants [11]. Also, Arabidopsis plants heterologously expressing the maize (Zea mays) ZmWRKY40 gene exhibited improved tolerance to drought [12]. Similarly, heterologous expression of TaWRKY13 in Arabidopsis increased root length and proline content, and reduced malondialdehyde content, thus improving salt stress tolerance [13]. Overexpression of WRKY13 in Arabidopsis enhanced cadmium tolerance of transgenic plants by inducing the expression of PLEIOTROPIC DRUG RESISTANCE 8 (PDR8), encoding an ATP-binding cassette transporter [14].
WRKY can however also act as a negative regulator of gene expression. Heterologous expression of the cotton (Gossypium hirsutum) GhWRKY33 gene reduced drought tolerance of transgenic Arabidopsis plants [15]. Likewise, heterologous expression of ZmWRKY17 impaired salt stress tolerance in transgenic Arabidopsis and lowered the abscisic acid (ABA) content by repressing ABA-dependent and stress-responsive genes [16].
Beyond the functional characterization of WRKY genes in Arabidopsis, the functions of many WRKY genes remain to be validated in non-model species. Indeed, few studies have tested the contribution of WKRY genes in their species of origin. Overexpression of TaWRKY2 in wheat enhanced tolerance to drought stress and increased yield [17]. Overexpression of OsWRKY11 enhanced tolerance to drought and heat in transgenic rice [18]. The grapevine (Vitis amurensis) VaWRKY12 gene enhanced cold tolerance in transgenic grapevine calli [19]. In wild sorghum (Sorghum propinquum), SpWRKY controls seed shattering but is unrelated to seed shattering genes selected during domestication, as it likely arose recently [20]. These studies confirm that WRKY TFs play important roles and suggest their potential use for crop improvement in terms of stress tolerance.
Since the identification of the first WRKY gene from sweet potato (Ipomoea batatas) [21], genome-wide analyses in different species have identified many WRKY genes, including 171 WRKY genes in wheat [22], 119 in maize [23], 103 in rice (Oryza sativa) [24], 71 in sesame (Sesamum indicum) [25], 70 in chickpea (Cicer arietinum) [26], and 79 in potato (Lycopersicum tuberosum) [27]. Sorghum is the fifth most important cereal crop in terms of production and dedicated arable land, and displays unique adaptations that allow it to withstand harsh conditions at different growth stages. Sorghum is also an excellent model for TF studies [28–30]. The availability of a complete genome assembly for sorghum now provides an opportunity for the genome-wide identification of SbWRKY genes. To gain insight into the roles of SbWRKYs in plant responses to stresses such as drought, we used a variety of approaches to identify and functionally characterize 94 putative members of the WRKY family in sorghum.
Material and methods
Identification of WRKY family genes in sorghum
We collected data from the following databases to identify putative SbWRKY genes. The Plant Transcription Factor Database version 4 (http://planttfdb.cbi.pku.edu.cn/) was used to download the amino acid sequences of sorghum WRKY proteins, and "WRKY" was used as a query to search against the Grassius Transcription Factor Database (https://grassius.org/grasstfdb.php). We used the WRKY domain ID (PF03106) to identify putative WRKY proteins encoded by the S. bicolor genome (v3.1) through the Joint Genome Institute (JGI) (https://phytozome.jgi.doe.gov/pz/portal.html#). We also used the keyword “WRKY” as a search query in the MOROCOSHI Sorghum Transcription Factor Database (http://sorghum.riken.jp/morokoshi/Home.html).
We employed CD-HIT suite (http://weizhong-lab.ucsd.edu/cdhit_suite/cgi-bin/index.cgi?cmd=cd-hit) to remove redundant and incomplete sequences, and Simple Modular Architecture Research Tool (SMART) (http://smart.embl-heidelberg.de/#) [31] to confirm that the sequences contained WRKY domain(s).
For all identified SbWRKY proteins, we obtained their predicted isoelectric point (pI) and molecular weight (MW) from the ExPASy proteomic server (http://web.expasy.org/protparam).
Chromosome mapping and tandem duplications of SbWRKY genes
Information about the chromosomal locations of all identified SbWRKY genes was obtained using the Phytozome BioMart tool (https://phytozome.jgi.doe.gov/biomart/martview/), and the genes were mapped onto sorghum chromosomes by MapChart (v.2.32). Tandem duplications of SbWRKYs were based on the following criteria: genes within a 100-kbp region on an individual chromosome with a sequence similarity ≥ 70% [32]. We calculated sequence similarities using EMBOSS Water, which uses the Smith-Waterman local pairwise alignment algorithm (https://www.ebi.ac.uk/Tools/psa/emboss_water/).
Classification and phylogenetic analysis of SbWRKY genes
Arabidopsis WRKY amino acid sequences obtained from the Arabidopsis Information Resource (TAIR) (https://www.arabidopsis.org/)) together with our SbWRKY sequences were used to construct a phylogenetic tree and classify SbWRKY genes. We performed multiple sequence alignments with ClustalW for AtWRKY and SbWRKY protein sequences and constructed a phylogenetic tree using MEGA v7.0 (https://www.megasoftware.net/) and the p-distance model. Pairwise deletion and 1,000 bootstrap replicates were used for the neighbour-joining method [33]. Based on the phylogenetic tree of AtWRKY and SbWRKY sequences, SbWRKY genes were classified into different groups and subgroups. The identification of putative sorghum WRKY orthologs in Arabidopsis was based on sequence alignment data and the phylogenetic tree [27].
Gene structure analysis of SbWRKY genes
The exon-intron structure of SbWRKYs was analyzed using the Gene Structure Display Server (GSDS v. 2.0) (http://gsds.cbi.pku.edu.cn/) from the Center for Bioinformatics at Peking University [34]. The genomic sequence and coding sequence (CDS) of each SbWRKY gene were used to predict the exon-intron pattern.
Conserved motif distribution analysis of SbWRKY genes
Conserved motifs in the SbWRKY proteins were identified using Multiple Em for Motif Elicitation (MEME) (v. 4.9.0; http://meme.nbcr.net/meme/)) with the following parameters: maximum motif number: 20; site distributions: any number of repetitions; minimum and maximum width: 10 and 50, respectively [35].
Gene ontology annotation and analysis of cis-acting elements
The gene ontology (GO) annotation analysis for SbWRKY proteins was performed using the Blast2GO tool with default parameters [36]. We screened protein sequences using the Basic Local Alignment Tool for Proteins (BlastP), followed by functional analysis, including mapping and annotation. Moreover, we collected information related to the biological process, molecular function, and cellular component associated with each SbWRKY. We also analyzed the cis-acting elements of SbWRKY genes by extracting 1,500 bp of upstream region for all SbWRKY genes and running the sequences through the online website PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/).
Digital expression pattern analysis of SbWRKY genes
To survey SbWRKY expression profiles, Affymetrix transcriptomic array data for sorghum were obtained from the Sorghum Functional Genomics Database (http://structuralbiology.cau.edu.cn/sorghum/pattern.php). The Genevestigator platform was used to analyze the expression profiles of SbWRKYs under different environmental stresses (drought, salt, ABA, and heat) with different samples stored in the Genevestigator platform [37]. The SbWRKY expression profiles heatmaps were generated using a hierarchical clustering analysis tool on the Genevestigator platform.
Drought stress treatment and samples collection
Seeds of Sorghum bicolor genotype (SX44B) used in this study were provided by Professor Zhang Fuyao (Sorghum Institute, Shanxi Academy of Agricultural Sciences, Shanxi, China). The seeds were surface-sterilized with 75% (v/v) ethanol and 5% (v/v) sodium hypochlorite for 1–2 min, and then rinsed them three times with distilled water. Sterilized seeds were allowed to germinate on two layers of water-soaked paper and incubated at 25°C in darkness for 3 d. Seedlings were transferred to pots containing a soil mixture of vermiculite and peat moss in a 1:1 ratio. Seedlings were kept in a growth chamber with 60–70% relative humidity, 28°C/23°C day/night temperature cycles, and a 16-h light/8-h dark photoperiod. Seedlings were maintained under normal growth conditions for two weeks before exposure to drought treatment, seedlings were treated with PEG8000 20% [38]. The seedlings shoot samples were collected at 0, 3, 6, 12, and 24 h, all samples were frozen in liquid nitrogen, and then stored at –80°C until used for RNA extraction.
Gene expression analysis by quantitative Real-Time PCR (qRT-PCR)
RNAs were extracted from the samples using (Promega, China) according to the manufacturer’s specifications. The first-strand cDNAs were synthesized by reverse transcription of 100 μg total RNA which was generated using the Easy Script First-strand cDNA Synthesis SuperMix Kit (TransGen Biotech, China). Synthesized cDNA was diluted 1:10 with nuclease-free water for use in qRT-PCR. The expression levels of the genes were normalized to the sorghum housekeeping gene GLYCERALDEHYDE 3-PHOSPHATE DEHYDROGENASE (GAPDH) gene as an internal control. Gene-specific primers were designed using the National Center for Biotechnology Information (NCBI) Primer-BLAST tool and were synthesized by Sangon (Beijing, China). Primers used in this study are listed in Supplementary S7 File. qRT-PCR was carried out using TransStart Green qPCR SuperMix UDG (TransGen Biotech, China) following the manufacturer’s instructions, on a Bio-Rad CFX96TM real-time PCR detection system (Bio-Rad, USA). Reaction parameters for thermal cycling were as follows: 94°C for 10 min, 40 cycles of 94°C for 5 sec and 55°C for 15 sec, 72°C for 10 sec. We performed RT-qPCR on three biological replicates and used the 2–ΔΔCt method for quantification [39].
Results
Identification of WRKY family members in sorghum
Taking advantage of the availability of a complete genome assembly for sorghum, we identified SbWRKY family members using the keyword “WRKY” and the WRKY domain consensus sequence (PF03106) as queries in different databases. In this study, the presence of the WRKYGQK or WRKYGQK-like conserved domain was the basic criterion for the inclusion of genes in the SbWRKY family. We initially identified 134, 94, 99, and 97 transcripts from the TFDB, Grassius, MOROCOSHI, and JGI databases, respectively. We then used CD-HIT and multiple sequence alignments to remove redundant SbWRKY protein sequences and confirmed the presence of the WRKY domain in the remaining sequences by running all proteins through the SMART database. We thus removed all redundant sequences and those with an incomplete WRKY domain. A total of 94 non-redundant SbWRKY sequences were identified, and the following gene and protein data were summarized in Table 1. We named SbWRKYs according to their physical positions along sorghum chromosomes, starting with the upper arm of chromosome 1 and moving down to the lower arm (SbWRKY1 to SbWRKY94), as described previously [1] (Table 1). We used this set of SbWRKY protein sequences for further characterization. SbWRKY proteins ranged from 110 to 1,584 amino acids, with an average length of 390 amino acids. Their predicted MW and pI values ranged from 12.23 to 78.24 and from 4.75 to 10.06, respectively.
Table 1. Characteristics of the identified SbWRKY genes.
| Gene Name | Gene Locus ID | Chromosome Location | Gene Start | Gene End | pI | MW | Conserved Heptapeptide | Zinc Finger Type | Domain Number | Group | Protein Length (aa) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SbWRKY1 | Sobic.001G006600.1 | Chr01 | 661532 | 669834 | 6.10 | 42.50 | WRKYGQK | C2H2 | 1 | lle | 392 |
| SbWRKY2 | Sobic.001G055400.1 | Chr01 | 4145000 | 4147281 | 9.42 | 46.08 | WRKYGQK | C2H2 | 1 | lle | 434 |
| SbWRKY3 | Sobic.001G083000.1 | Chr01 | 6408709 | 6412370 | 6.76 | 45.19 | WRKYGQK | C2H2 | 2 | l | 424 |
| SbWRKY4 | Sobic.001G084000.1 | Chr01 | 6506824 | 6511079 | 6.2 | 34.37 | WRKYGQK | C2H2 | 1 | llc | 331 |
| SbWRKY5 | Sobic.001G095500.1 | Chr01 | 7340447 | 7343822 | 10.05 | 43.61 | WRKYGQK | C2H2 | 1 | lld | 406 |
| SbWRKY6 | Sobic.001G148000.1 | Chr01 | 11929277 | 11931246 | 8.89 | 15.12 | WRKYGQK | C2H2 | 1 | NG | 141 |
| SbWRKY7 | Sobic.001G162100.1 | Chr01 | 13349344 | 13352564 | 9.78 | 37.88 | WRKYGQK | C2H2 | 1 | lld | 352 |
| SbWRKY8 | Sobic.001G282400.1 | Chr01 | 55413200 | 55415329 | 5.97 | 46.31 | WRKYGQK | C2H2 | 1 | lle | 427 |
| SbWRKY9 | Sobic.001G332500.1 | Chr01 | 62096233 | 62103315 | 7.34 | 52.62 | WRKYGQK | C2HC | 2 | l | 498 |
| SbWRKY10 | Sobic.001G381300.1 | Chr01 | 66930715 | 66931988 | 5.60 | 33.95 | WRKYGQK | C2HC | 1 | lll | 328 |
| SbWRKY11 | Sobic.001G389000.1 | Chr01 | 67559616 | 67561361 | 8.71 | 24.20 | WRKYGEK | C2H2 | 1 | lll | 225 |
| SbWRKY12 | Sobic.002G008600.2 | Chr02 | 791610 | 795938 | 9.71 | 29.42 | WRKYGQK | C2H2 | 1 | llc | 272 |
| SbWRKY13 | Sobic.002G128400.2 | Chr02 | 17755739 | 17757847 | 5.75 | 29.09 | WRKYGQK | C2H2 | 1 | lll | 275 |
| SbWRKY14 | Sobic.002G168300.3 | Chr02 | 52688090 | 52696535 | 6.92 | 17.74 | WRKYGSK | C2HC | 2 | lll | 1584 |
| SbWRKY15 | Sobic.002G174200.1 | Chr02 | 55134836 | 55136822 | 5.67 | 32.89 | WRKYGQK | C2HC | 1 | lll | 310 |
| SbWRKY16 | Sobic.002G174300.1 | Chr02 | 55188598 | 55190855 | 5.99 | 34.60 | WRKYGQK | C2HC | 1 | lll | 328 |
| SbWRKY17 | Sobic.002G202700.1 | Chr02 | 59258884 | 59260622 | 6.46 | 34.72 | WRKYGQK | C2H2 | 1 | lla | 327 |
| SbWRKY18 | Sobic.002G202800.1 | Chr02 | 59275303 | 59277597 | 6.72 | 31.77 | WRKYGQK | C2H2 | 1 | lla | 295 |
| SbWRKY19 | Sobic.002G242500.1 | Chr02 | 63158788 | 63162729 | 6.94 | 64.54 | WRKYGQK | C2H2 | 1 | l | 602 |
| SbWRKY20 | Sobic.002G355000.1 | Chr02 | 71806701 | 71813179 | 6.24 | 65.56 | WRKYGQK | C2H2 | 2 | l | 611 |
| SbWRKY21 | Sobic.002G418500.1 | Chr02 | 76629585 | 76632298 | 9.17 | 37.05 | WRKYGQK | C2HC | 1 | lll | 354 |
| SbWRKY22 | Sobic.003G000600.1 | Chr03 | 65678 | 68474 | 8.62 | 59.48 | WRKYGQK | C2H2 | 1 | llb | 570 |
| SbWRKY23 | Sobic.003G037400.1 | Chr03 | 3517021 | 3519078 | 6.07 | 26.65 | WRKYGKK | C2H2 | 1 | llc | 260 |
| SbWRKY24 | Sobic.003G037500.1 | Chr03 | 3520321 | 3522720 | 7.27 | 56.13 | WRKYGQK | C2H2 | 1 | llb | 548 |
| SbWRKY25 | Sobic.003G040800.1 | Chr03 | 3787973 | 3794918 | 9.75 | 30.73 | WRKYGQK | C2H2 | 1 | llc | 295 |
| SbWRKY26 | Sobic.003G138400.1 | Chr03 | 13400561 | 13405829 | 6.77 | 60.58 | WRKYGQK | C2H2 | 1 | llb | 582 |
| SbWRKY27 | Sobic.003G199400.1 | Chr03 | 52648841 | 52650568 | 5.57 | 23.75 | WRKYGKK | C2H2 | 1 | llc | 216 |
| SbWRKY28 | Sobic.003G200700.1 | Chr03 | 52845063 | 52846563 | 9.17 | 33.94 | WRKYGQK | C2H2 | 1 | llb | 332 |
| SbWRKY29 | Sobic.003G226600.1 | Chr03 | 56293002 | 56295461 | 8.7 | 44.22 | WRKYGQK | C2H2 | 1 | lle | 413 |
| SbWRKY30 | Sobic.003G227300.1 | Chr03 | 56472263 | 56479136 | 6.62 | 38.87 | WRKYGQK | C2H2 | 1 | llc | 361 |
| SbWRKY31 | Sobic.003G242800.2 | Chr03 | 58197072 | 58201064 | 6.45 | 35.97 | WRKYGQK | C2HC | 1 | lll | 346 |
| SbWRKY32 | Sobic.003G248400.1 | Chr03 | 58718991 | 58722963 | 7.32 | 42.36 | WRKYGQK | C2H2 | 1 | llc | 410 |
| SbWRKY33 | Sobic.003G276000.1 | Chr03 | 61248940 | 61250384 | 8.64 | 23.95 | WRKYGKK | C2H2 | 1 | llc | 225 |
| SbWRKY34 | Sobic.003G285500.1 | Chr03 | 61880213 | 61882652 | 5.27 | 32.23 | WRKYGQK | C2H2 | 1 | lle | 319 |
| SbWRKY35 | Sobic.003G287200.1 | Chr03 | 62028638 | 62032127 | 7.08 | 26.67 | WRKYGQK | C2H2 | 1 | llc | 246 |
| SbWRKY36 | Sobic.003G296300.1 | Chr03 | 62835017 | 62837081 | 4.75 | 33.69 | WRKYGQK | C2H2 | 1 | lle | 310 |
| SbWRKY37 | Sobic.003G337500.1 | Chr03 | 66057329 | 66064893 | 5.85 | 26.64 | WRKYGQK | C2HC | 1 | lll | 236 |
| SbWRKY38 | Sobic.003G337600.1 | Chr03 | 66070688 | 66076206 | 5.98 | 29.56 | WRKYGQK | C2HC | 1 | lll | 264 |
| SbWRKY39 | Sobic.003G337700.1 | Chr03 | 66081053 | 66083714 | 5.87 | 40.92 | WRKYGQK | C2HC | 1 | lll | 377 |
| SbWRKY40 | Sobic.003G337800.1 | Chr03 | 66089525 | 66092348 | 5.17 | 36.43 | WRKYGQK | C2HC | 1 | lll | 333 |
| SbWRKY41 | Sobic.003G337900.1 | Chr03 | 66104667 | 66107290 | 5.91 | 29.63 | WRKYGQK | C2HC | 1 | lll | 277 |
| SbWRKY42 | Sobic.003G341100.1 | Chr03 | 66392943 | 66395715 | 6.32 | 59.35 | WRKYGQK | C2H2 | 2 | l | 556 |
| SbWRKY43 | Sobic.003G353000.1 | Chr03 | 67192266 | 67198503 | 6.13 | 39.68 | WRKYGQK | C2H2 | 1 | NG | 377 |
| SbWRKY44 | Sobic.003G444000.1 | Chr03 | 74207255 | 74210127 | 4.76 | 38.85 | WRKYGQK | C2H2 | 1 | llc | 354 |
| SbWRKY45 | Sobic.004G065900.1 | Chr04 | 5352084 | 5353418 | 8.45 | 38.95 | WRKYGQK | C2H2 | 1 | lla | 364 |
| SbWRKY46 | Sobic.004G117600.1 | Chr04 | 12353902 | 12355528 | 5.81 | 39.57 | WRKYGQK | C2H2 | 1 | lle | 375 |
| SbWRKY47 | Sobic.004G138400.2 | Chr04 | 38911863 | 38913035 | 9.78 | 32.03 | WRKYGQK | C2H2 | 1 | lld | 299 |
| SbWRKY48 | Sobic.004G271800.1 | Chr04 | 61583724 | 61588753 | 5.67 | 51.61 | WRKYGQK | C2H2 | 1 | lle | 497 |
| SbWRKY49 | Sobic.004G298400.1 | Chr04 | 63778460 | 63782424 | 8.43 | 26.10 | WRKYGQK | C2H2 | 1 | llc | 238 |
| SbWRKY50 | Sobic.004G312200.1 | Chr04 | 64900147 | 64902898 | 5.09 | 61.31 | WRKYGQK | C2H2 | 1 | llb | 578 |
| SbWRKY51 | Sobic.005G013400.1 | Chr05 | 1211003 | 1212487 | 5.69 | 36.49 | WRKYGEK | C2HC | 1 | lll | 334 |
| SbWRKY52 | Sobic.005G013500.1 | Chr05 | 1224392 | 1225924 | 6.31 | 38.95 | WRKYGEK | C2HC | 1 | lll | 361 |
| SbWRKY53 | Sobic.005G013600.2 | Chr05 | 1235265 | 1241922 | 8.85 | 25.24 | WRKYGEK | C2HC | 1 | lll | 227 |
| SbWRKY54 | Sobic.005G013800.1 | Chr05 | 1245144 | 1246547 | 5.88 | 30.67 | WRKYGQK | C2HC | 1 | lll | 271 |
| SbWRKY55 | Sobic.005G014000.1 | Chr05 | 1261347 | 1263314 | 6.27 | 33.04 | WRKYGQK | C2HC | 1 | lll | 289 |
| SbWRKY56 | Sobic.005G014200.1 | Chr05 | 1302596 | 1304021 | 6 | 30.80 | WRKYGQK | C2HC | 1 | lll | 272 |
| SbWRKY57 | Sobic.005G117400.2 | Chr05 | 51657060 | 51660442 | 9.18 | 24.24 | WRKYGQK | C2H2 | 1 | llc | 225 |
| SbWRKY58 | Sobic.006G051700.1 | Chr06 | 38157163 | 38162677 | 5.97 | 12.23 | WRKYGQK | C2HC | 1 | lll | 110 |
| SbWRKY59 | Sobic.006G115700.1 | Chr06 | 48356715 | 48362695 | 6.77 | 78.24 | WRKYGQK | C2H2 | 2 | l | 740 |
| SbWRKY60 | Sobic.006G166300.1 | Chr06 | 52377828 | 52383758 | 8.66 | 29.10 | WRKYGQK | C2H2 | 1 | llc | 269 |
| SbWRKY61 | Sobic.006G201000.1 | Chr06 | 55241047 | 55245365 | 7.01 | 55.79 | WRKYGQK | C2H2 | 1 | lle | 532 |
| SbWRKY62 | Sobic.006G206000.1 | Chr06 | 55572908 | 55574447 | 10.05 | 33.13 | WRKYGQK | C2H2 | 1 | lld | 315 |
| SbWRKY63 | Sobic.007G077466.1 | Chr07 | 8936871 | 8940145 | 7.7 | 49.41 | WRKYGQK | C2H2 | 1 | llc | 455 |
| SbWRKY64 | Sobic.007G085300.1 | Chr07 | 10754655 | 10756113 | 9.87 | 32.97 | WRKYGQK | C2H2 | 1 | lld | 318 |
| SbWRKY65 | Sobic.007G111600.1 | Chr07 | 41587770 | 41594141 | 6.05 | 61.78 | WRKYGQK | C2H2 | 2 | l | 569 |
| SbWRKY66 | Sobic.007G118301.1 | Chr07 | 51192936 | 51194468 | 6.48 | 35.62 | WRKYGQK | C2HC | 1 | lll | 340 |
| SbWRKY67 | Sobic.007G217700.3 | Chr07 | 64590499 | 64600679 | 6.89 | 74.52 | WRKYGQK | C2H2 | 2 | l | 685 |
| SbWRKY68 | Sobic.008G028600.1 | Chr08 | 2541872 | 2544263 | 5.35 | 36.20 | WRKYGQK | C2HC | 1 | lll | 341 |
| SbWRKY69 | Sobic.008G029000.2 | Chr08 | 2604929 | 2606770 | 6.72 | 33.68 | WRKYGQK | C2HC | 1 | lll | 294 |
| SbWRKY70 | Sobic.008G029200.1 | Chr08 | 2612734 | 2614762 | 5.99 | 30.33 | WRKYGQK | C2HC | 1 | lll | 267 |
| SbWRKY71 | Sobic.008G029400.1 | Chr08 | 2625199 | 2632242 | 8.94 | 25.19 | WRKYGEK | C2HC | 1 | lll | 225 |
| SbWRKY72 | Sobic.008G060300.1 | Chr08 | 6462595 | 6464519 | 5.15 | 35.55 | WRKYGQK | C2HC | 1 | lll | 334 |
| SbWRKY73 | Sobic.008G107500.1 | Chr08 | 50603514 | 50609969 | 7.71 | 52.42 | WRKYGQK | C2H2 | 2 | l | 496 |
| SbWRKY74 | Sobic.008G153600.1 | Chr08 | 58584243 | 58587317 | 10.06 | 39.29 | WRKYGQK | C2H2 | 1 | lld | 371 |
| SbWRKY75 | Sobic.008G174100.1 | Chr08 | 60851165 | 60856614 | 8.89 | 13.13 | WRKSGQR | C2HC | 1 | lll | 1163 |
| SbWRKY76 | Sobic.009G034800.1 | Chr09 | 3182138 | 3188861 | 5.79 | 58.46 | WRKYGQK | C2H2 | 1 | llb | 567 |
| SbWRKY77 | Sobic.009G068900.1 | Chr09 | 7567284 | 7568764 | 6.96 | 22.09 | WRKYGKK | C2H2 | 1 | llc | 206 |
| SbWRKY78 | Sobic.009G092100.1 | Chr09 | 20912936 | 20928927 | 6.39 | 40.83 | WRKYGQK | C2HC | 1 | III | 378 |
| SbWRKY79 | Sobic.009G100500.1 | Chr09 | 39826393 | 39831148 | 7.69 | 54.06 | WRKYGQK | C2H2 | 2 | l | 517 |
| SbWRKY80 | Sobic.009G171600.1 | Chr09 | 52696414 | 52699630 | 8.28 | 68.54 | WRKYGQK | C2H2 | 2 | l | 649 |
| SbWRKY81 | Sobic.009G174300.1 | Chr09 | 52963804 | 52966801 | 6.25 | 26.62 | WRKYGEK | C2HC | 1 | lll | 235 |
| SbWRKY82 | Sobic.009G206800.1 | Chr09 | 55463596 | 55464452 | 9.2 | 25.32 | WRKYGQK | C2H2 | 1 | llc | 241 |
| SbWRKY83 | Sobic.009G212800.1 | Chr09 | 55836119 | 55837874 | 6.08 | 22.71 | WRKYGKK | C2H2 | 1 | llc | 219 |
| SbWRKY84 | Sobic.009G234100.1 | Chr09 | 57342319 | 57345920 | 6.38 | 44.44 | WRKYGQK | C2H2 | 1 | llc | 424 |
| SbWRKY85 | Sobic.009G234900.1 | Chr09 | 57418453 | 57422639 | 6.52 | 65.84 | WRKYGQK | C2H2 | 1 | llb | 631 |
| SbWRKY86 | Sobic.009G238200.1 | Chr09 | 57628850 | 57630763 | 5.81 | 29.08 | WRKYGQK | C2HC | 1 | lll | 272 |
| SbWRKY87 | Sobic.009G247300.2 | Chr09 | 58261832 | 58265060 | 8.22 | 28.36 | WRKYGQK | C2H2 | 1 | llc | 262 |
| SbWRKY88 | Sb09g029810 | Chr09 | 58261882 | 58265547 | 6.07 | 36.12 | WRKYGQK | C2H2 | 1 | llc | 343 |
| SbWRKY89 | Sobic.009G247700.1 | Chr09 | 58309106 | 58310986 | 6.05 | 38.89 | WRKYGQK | C2H2 | 1 | lle | 364 |
| SbWRKY90 | Sobic.010G035300.2 | Chr10 | 2858141 | 2865169 | 6.79 | 65.39 | WRKYGQK | C2H2 | 1 | llb | 625 |
| SbWRKY91 | Sobic.010G045700.1 | Chr10 | 3566332 | 3570889 | 6.11 | 40.52 | WRKYGQK | C2HC | 1 | lll | 378 |
| SbWRKY92 | Sobic.010G148600.2 | Chr10 | 42567454 | 42569300 | 6.01 | 41.69 | WRKYGQK | C2H2 | 1 | lle | 385 |
| SbWRKY93 | Sobic.010G148800.1 | Chr10 | 42829856 | 42831874 | 7.58 | 37.43 | WRKYGQK | C2H2 | 1 | lle | 350 |
| SbWRKY94 | Sobic.010G209100.1 | Chr10 | 55261687 | 55263735 | 9.64 | 37.82 | WRKYGQK | C2H2 | 1 | lla | 348 |
pI, isoelectric point; MW, molecular weight; aa, amino acid.
Chromosome mapping of the SbWRKY genes and tandem duplication analysis
MapChart was used to determine the locations of the 94 SbWRKY genes, and they were distributed on all 10 sorghum chromosomes (Chr) (Fig 1). Chr 3 counted the highest number of SbWRKYs with 23 genes, corresponding to 24.5% of the entire gene family, followed by 14 genes on Chr 9, 11 genes on Chr 1, and 10 genes on Chr2. Chromosomes 6, 7, and 10 had the fewest number of genes, with only five SbWRKYs each. The remaining SbWRKY genes were located on Chr4 (six genes), Chr5 (seven genes), and Chr8 (eight genes).
Fig 1. Distribution of 94 SbWRKY genes on sorghum chromosomes.

Chr01-Chr10 above the colored bars indicates chromosome (Chr) numbers. The physical location of each SbWRKY gene is shown, and the gene name is indicated on the right side of each bar as SbWRKY#. Red boxes indicate tandem duplications, and green lines denote gene clusters.
Tandem repeats were identified based on previously reported criteria [40]; namely, two or more genes should be located within a 100-kbp window and display a sequence similarity of at least 70%. Gene cluster events were observed on six chromosomes. Specifically, there were five clusters on Chr3; two each on Chr2 and Chr9; and one cluster each on Chr1, Chr5, and Chr8. No clusters were found on Chr4, Chr6, Chr7, and Chr10. High-density clusters were detected on Chr5 and Chr3, and identified tandem repeats on two chromosomes: two group III genes (SbWRKY51 and SbWRKY52) on Chr5 and two group-IIc genes (SbWRKY87 and SbWRKY88) on Chr9 (Fig 1). These tandem-duplicated genes clustered together in the phylogenetic tree within their respective clades, and the sequence similarity metrics for the gene pairs are provided in S1 File.
Classification and phylogenetic analysis of SbWRKYs
To investigate the evolution of SbWRKY family members, an unrooted phylogenetic tree was constructed based on multiple sequence alignment between full-length protein sequences of 65 AtWRKYs and 94 SbWRKYs, using the neighbour-joining method in MEGA7.0 (Fig 2). The constructed phylogenetic tree was used to classify the SbWRKYs into three major groups (I, II, and III), according to the classification in Arabidopsis [1] (Fig 2). Of the 11 SbWRKYs in group I, 10 had two WRKYGQK motifs and two C2H2-type zinc finger motifs (C-X3-4-C-X22-23-H-X1-H), corresponding to two full WRKY domains. Although the protein encoded by SbWRKY19 had only one WRKY domain, it belonged to group I on the phylogenetic tree. Fifty protein sequences with one WRKY domain and the C2H2-type zinc finger motif (C-X4-5-C-X23-H-X1-H) were classified into group II. This group was further divided into five subgroups, IIa, IIb, IIc, IId, and IIe, with 4, 8, 20, 6, and 12 members, respectively. Group III contained 31 members with one WRKY domain and the C2HC-type zinc finger motif (C-X7-C-X23-H-X1-C). SbWRKY14 was unique in that it comprised of two WRKY domains with the C2HC-type zinc finger motif (C-X7-C-X23-H-X1-C). Thus, it had features associated with both group I and group III WRKYs, but was classified into group III based on its position in the phylogenetic tree. SbWRKY6 and SbWRKY43 did not belong to any group (Fig 2 and Table 1). Group II was the largest group and accounted for 53.2% of all putative SbWRKYs, which is similar to reports in wheat, soybean (Glycine max), and pepper (Capsicum annuum). Overall, the classification of SbWRKYs confirms their diversification, which suggests that different family members may have varied functions.
Fig 2. Phylogenetic tree of WRKY members in sorghum and Arabidopsis.

SbWRKY and AtWRKY protein sequences were aligned with ClustalW, and a phylogenetic tree was constructed with MEGA7.0 using the neighbour-joining method and 1,000 bootstrap replicates. The members were divided into groups I, II, and III, and group II was further divided into subgroups IIa, IIb, IIc, Ild, and Ile.
The highly conserved heptapeptide motif WRKYGQK was present in 81 SbWRKY proteins. We observed several heptapeptide variants in the remaining 13 proteins as follows: WRKYGEK in six proteins (SbWRKY11, SbWRKY51, SbWRKY52, SbWRKY53, SbWRKY71, and SbWRKY81); WRKYGKK in five proteins (SbWRKY23, SbWRKY27, SbWRKY33, SbWRKY77, and SbWRKY83); WRKYGSK in SbWRKY14; and WRKSGQR in SbWRKY75. Among the 94 identified SbWRKYs, only 11 had two WRKY domains, whereas the remaining members had one WRKY domain. SbWRKY protein sequences, genomic sequences, and CDS are provided in S2, S3, and S4 Files, respectively. AtWRKY protein sequences are provided in S5 File.
SbWRKY gene structure analysis
To obtain additional clues about the evolution of SbWRKY family members and their specific features, SbWRKY exon-intron structures were analyzed. The intron number of SbWRKY genes ranged from zero to seven, whereas their size varied. Among the 94 SbWRKY genes identified, 10 contained two exons (and one intron), 58 had three exons (two introns), eight had four exons (three introns), eight had five exons (four introns), five had six exons (five introns); the SbWRKY14 gene had eight exons and seven introns (Fig 3). Four genes (SbWRKY51, SbWRKY64, SbWRKY66, and SbWRKY88) lacked introns.
Fig 3. Exon–intron structures of SbWRKY genes.
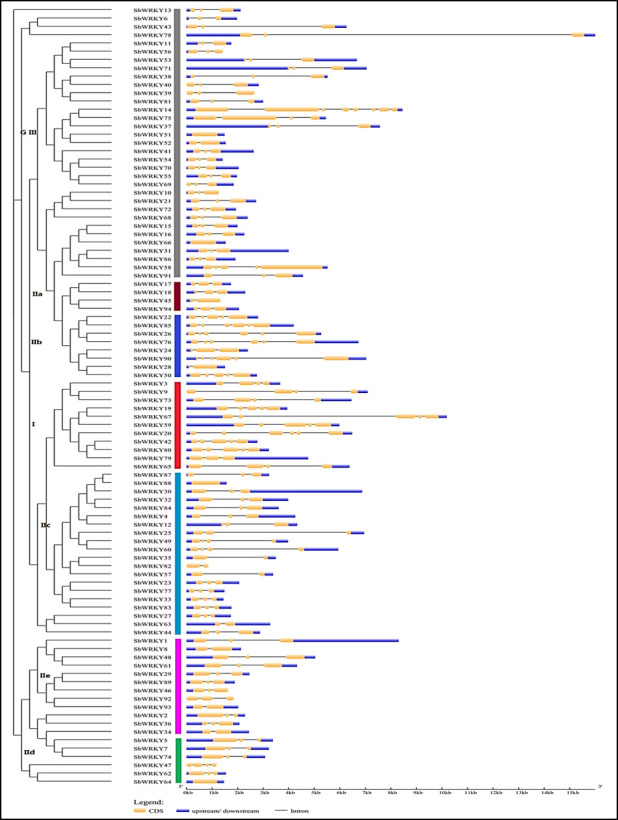
We used the Gene Structure Display Server (GSDS) for gene structure analysis and constructed the phylogenetic tree using MEGA v7.0. In the gene diagrams, blue bars indicate upstream and downstream UTRs, yellow bars indicate coding sequences (CDS), and black lines indicate introns.
The exon–intron distribution patterns showed some similarities in terms of their numbers and positions within the same group. However, there were also differences within groups. For instance, all genes in group II had zero to five introns, all SbWRKY genes in subgroup IIb had five introns except SbWRKY22 and SbWRKY50, which had four, SbWRKY24 had two, and SbWRKY28 had one intron. Intron numbers in group III ranged from zero to seven, with SbWRKY14 being the only gene with seven introns among all SbWRKYs. These results indicate that there is considerable structural variation among SbWRKYs, which may correspond to functional diversification between closely related members (Fig 3).
Motif composition analysis of SbWRKYs
MEME (version 4.11) was used to analyze all SbWRKY protein sequences for conserved motifs, resulting in the identification of 20 distinct conserved motifs, ranging from 6 to 50 amino acids in length (Fig 4). Motifs 1, 2, 3, and 4 corresponded to the WRKY domain located at the C-terminus of sorghum WRKY proteins. Most SbWRKY members within the same group or subgroup shared a similar motif composition. Motifs 12, 15, and 20 were unique to group III. Motifs 7, 10, 11, and 18 were unique to group IIb, and motif 14 was exclusively detected in group I. All group I members had two WRKY domains except for SbWRKY19, as mentioned earlier, suggesting it may have lost its N-terminal WRKY domain. Motif 13 was unique to group IIe. Examples of motifs shared by different groups included motif 8, shared by groups IIe and IId, and motif 5, shared by groups I and IIc. Although SbWRKY6 and SbWRKY43 clustered with group III, they are not associated with any group; they contained motifs 1, 2, 6, 3, and 17 (Fig 4). Groups IIe and IId were two close subgroups in the phylogenetic tree, and the vast majority of their members contained motifs 8, 2, 4, 3, and 17. Both subgroups had a similar domain arrangement, which may be indicative of functional similarity. Some motifs occurred only in a few SbWRKY members, such as motifs 12 and 20, which were only present in SbWRKY14, SbWRKY75, and SbWRKY58.
Fig 4. Motif analysis of SbWRKYs.

(a) The distribution of 20 conserved motifs identified by MEME in the different groups of SbWRKYs. (b) Each motif is indicated by a different color. (c) Sequence logos for motifs 1–20.
Gene ontology annotation and analysis of cis-acting elements
Gene ontology (GO) annotations of 94 SbWRKY proteins were analyzed using the Blast2GO tool. The SbWRKY target genes were categorized into different functional groups under three main categories including biological processes, molecular functions, and cellular components (Fig 5). Under the larger umbrella of biological processes, most SbWRKYs were identified as being involved in the regulation of cellular processes, biosynthetic processes and different metabolic processes, as well as response to different stimuli, signalling, cell communication, and responses to other organisms, chemicals and stress. The molecular functions of SbWRKYs were associated mostly with DNA-binding, DNA-binding transcription factor activity, catalytic activity, acting on a protein, and hydrolase activity. The cellular component of this protein family included organelle and intracellular organelle (Fig 5). In addition, all SbWRKY proteins were predicted to be localized in the nucleus.
Fig 5. Gene ontology analysis of identified SbWRKYs.

The enrichment analysis shows the involvement of SbWRKY in biological processes, molecular functions, and cellular components.
Cis-acting elements within promoters are the binding sites through which transcriptional regulation is enacted. We therefore, extracted the 1.5-kbp promoter regions upstream of all SbWRKY genes from the sorghum genome assembly to identify cis-acting elements using the online tool PlantCARE. Thus, various cis-acting regulatory elements were found in all SbWRKY genes promoter regions. Featuring prominently in our list of cis-elements were stress-responsive elements, including MBS (MYB transcription factor binding site involved in drought inducibility), LTR (low-temperature responsive element), ARE (anaerobic induction responsive element), TC-rich repeats (defense-responsive and stress-responsive elements, WUN-motif (wound-responsive elements), and GC-motif (anoxic specific inducibility element). Phytohormone-responsive elements: ABA-responsive element (ABRE), methyl jasmonate (MeJA) responsive element (TGACG-motif and CGTCA-motif), auxin-responsive elements (AuxRR-core and TGA-element), salicylic acid-responsive element (TCA-element), and gibberellin-responsive element (GARE-motif). Multiple light-responsive elements were present in the promoters of SbWRKY genes, including Sp1, TCT-motif, GT1-motif, GATA-motif, GA-motif, BoX II, and G-box, as well as elements associated with development, including the CAT-box (element related to meristem expression), and the o2-site (metabolism regulation). The promoter related and binding sites elements were found included TATA-box, CAAT-box, A-box, HD-Zip, and W-box (a classic WRKY DNA-binding motif). Many unknown functions were detected included AAGAA-motif, DRE core, and MYB. The most common cis-acting regulatory elements in the SbWRKY promoter regions were TGACG-motif, ABRE, CGTCA-motif, CAAT-motif, MYB, TATA-box, and G-box. We identified W-box elements in the promoters of 38 SbWRKY genes. All SbWRKY genes contained at least one stress-responsive element along with other cis-elements, reflecting their potential functional variation (S6 File).
Digital expression analysis of SbWRKY genes at different growth stages and in different tissues
As a preliminary survey of the potential roles of SbWRKY genes during sorghum growth and development, the temporal and spatial expression profiles of SbWRKY genes were investigated using microarray data available from SorghumFDB. We used the Genevestigator platform for the analysis, and present the results as heatmaps. The sorghum accessions represented in the microarray datasets included R159, Fremont, Atlas, PI455230, PI152611, and AR2400 [41]. The microarray datasets comprised 37 samples representing leaves, roots, shoots, stems (pith and rind), and internodes. Seventy-eight of the 94 SbWRKY genes were represented in the microarray data and displayed distinct expression patterns across all tested tissues. For example, all 78 genes were expressed in leaves, 76 in roots, 77 in shoots and pith, 76 in internodes, and 75 in rind. Thirty-two of the 78 genes exhibited high expression levels in at least one tissue (Fig 6A). The number of genes with high expression levels (>65% expression) varied between tissues, although roots showed the highest number of highly expressed SbWRKY genes with 17 members, followed by nine in pith, seven in leaves, six in rind, five in internodes and four in shoots. The most highly expressed SbWRKY genes were SbWRKY19, SbWRKY83, SbWRKY45, SbWRKY79, SbWRKY5, SbWRKY42, SbWRKY73, SbWRKY22, SbWRKY34, SbWRKY72, SbWRKY25, and SbWRKY70 in different tissues (Fig 6A). Notably, SbWRKY72 was highly expressed in all tissues. By contrast, SbWRKY70 expression was only detected in leaves and shoot. Clustering analysis of SbWRKY expression patterns grouped rind and pith, this is consistent with their biological features.
Fig 6. Heatmaps of SbWRKY gene expression.

SbWRKY expression levels in different tissues (a) and at different growth stages (b). (c) Hierarchical clustering of SWRKY gene expression patterns under different environmental conditions, including drought, ABA, heat, salt and combination stress.
SbWRKY expression patterns at different growth and developmental stages (seedling, stem elongation, flowering, boot, and dough stage) were also analyzed (Fig 6B). SbWRKY genes were expressed differently (up- or down-regulated) at all stages, and those with high expression at different stages included SbWRKY74, SbWRKY75, SbWRKY19, SbWRKY5, SbWRKY45, SbWRKY79, SbWRKY25, and SbWRKY72. SbWRKY expression was slightly higher during the seedling, flowering, and dough stages, suggesting that they may be involved in stress responses during sensitive developmental stages to improve plant tolerance (Fig 6B).
Hierarchical clustering analysis of the expression patterns of 78 SbWRKY genes under different environmental stresses was performed in Genevestigator. Two major clusters were obtained, which divided SbWRKY genes into two groups. The first major cluster consisted of highly expressed genes, including SbWRKY45, SbWRKY79, SbWRKY83, and SbWRKY16, under various abiotic stress conditions such as drought, salt and ABA (Fig 6C). The second major cluster contained several sub-clusters of SbWRKY genes with different expression patterns, i.e., up- or downregulated at least 2.5-fold (in absolute terms), in response to drought, salt, ABA in different sorghum tissues (Fig 6C). Several genes were found to have a stable expression level across different tissues and may therefore be considered constitutively expressed. We hypothesize that other SbWRKY genes with low expression levels may work cooperatively with other proteins throughout plant development.
qRT-PCR expression analysis of SbWRKY genes in response to drought stress
Previous studies proved that WRKY genes involved in plant responses to drought stress in several crops such as maize, wheat, and rice [42]. To investigate the role of SbWRKY genes in drought responses in sorghum, we selected five genes (SbWRKY45, SbWRKY72, SbWRKY74, SbWRKY75, and SbWRKY79) for expression analysis by qRT-PCR in the shoot of sorghum seedlings subjected to drought stress. qRT-PCR results revealed that transcripts levels for these five SbWRKY genes were remarkably increased under drought stress at different time points (Fig 7) suggesting that these genes may function in this process. The relative expression of our selected SbWRKY genes peaked at different time points. SbWRKY75 exhibited the highest expression level, with an 89-fold change in expression after 6 h of drought stress, with a final decline after 24 h. The peak expression of SbWRKY74 occurred 12 h after the onset of drought stress, whereas SbWRKY45 and SbWRKY79 expression peaked 24 h into drought stress (Fig 7). Although the peak expression of SbWRKY72 and SbWRKY75 occurred at 6 h, the expression of SbWRKY72 gradually increased to this level, then decreased gradually at later time points. SbWRKY79 showed the least induction during drought stress relative to the other genes tested here. Overall, the expression pattern of these selected SbWRKY genes under drought stress conditions suggests that different SbWRKY genes may play an essential role in drought stress tolerance.
Fig 7. Relative expression of selected SbWRKY genes in response to drought stress.

Relative expression levels of five SbWRKY genes under drought stress. Genes expression was analyzed by RT-qPCR, with the 0 h sample used as untreated control (expression = 1). Error bars represent standard errors; data were calculated using the 2–ΔΔCt method.
Discussion
WRKY TFs are key regulators of many processes in plants, including responses to abiotic and biotic stresses. In both model and non-model plants, considerable progress has been made towards identifying and functionally characterizing WRKY TFs, and many WRKY genes have been found to promote stress tolerance [42]. The completed genome assembly of sorghum now makes it possible to perform a genome-wide analysis of the SbWRKY gene family. We identified a total of 94 SbWRKY genes in S. bicolor, which is slightly higher than in other species; for example, there are 75 WRKY genes in Arabidopsis [1], 71 in sesame [25], 79 in potato, 85 in cassava (Manihot essculenta) [43], and 59 in grapevine [44]. In contrast, sorghum contains fewer WRKY genes than maize, soybean, or rice [23, 45, 46]. The present findings in sorghum, an important cereal crop and a model plant for drought tolerance, add to the recent identification of WRKY genes in a variety of plant species, including chickpea [26], Chinese jujube (Ziziphus jujube) [47], sugar beet (Beta vulgaris) [48], coffee (Coffea arabica) [49], pepper [50], eggplant (Solanum melongena) [51], Asian legume crops [52], sweet potato (Ipomoea batatas) [53], and pearl millet (Pennisetum glaucum) [54].
SbWRKY genes are distributed on all 10 sorghum chromosomes. Previous studies mapped many traits, such as stay-green phenotypes, lodging tolerance, pre-flowering drought tolerance, and yield-related components, to various chromosomal locations in sorghum [55, 56, 57]. Several SbWRKY genes map within these candidate regions; for example, SbWRKY26 is located on chromosome 3, close to a mapping interval for stay-green and pre-flowering drought tolerance Quantitative Trait Loci (QTL) in sorghum. Likewise, SbWRKY67 maps to chromosome 7, in a genomic region associated with a lodging tolerance QTL, whereas SbWRKY59 maps to the same region associated with pre-flowering drought tolerance, flowering time, and stay-green QTLs. These results suggest that several SbWRKY genes might control or contribute to stay-green, pre-flowering drought tolerance, or other traits related to stresses tolerance.
Gene duplication events affect genome expansion, family size, and the distribution of genes on chromosomes. Distinct types of duplication events, such as tandem and segmental duplication, differ in terms of the resulting number of gene copies and their distribution, and these factors are important for functional prediction. Tandem duplication of chromosome regions can also give rise to a cluster of family members, with subsequent structural and functional divergence over time leading to the expansion and evolution of the gene family [58]. Our analysis identified two tandem duplications of SbWRKY genes, which is a smaller number than that reported for potato [27], rice [24], wheat [22], or Arabidopsis [59]. Despite these differences, tandem duplications may have shaped the evolution of the SbWRKY gene family in sorghum.
Both tandem duplications and segmental duplications have played essential roles in the evolution and diversification of the WRKY gene family in plant species [60]. These evolutionary events were assigned to three types of gene duplication: tandem, segmental and whole-genome duplications [61]. Duplication events are significant for WRKY diversification, as duplicated WRKY genes may acquire new functions. In this study, we focused on tandem duplication events. We hypothesize that tandem duplications have played an important role in the evolution and diversification of WRKY genes in sorghum, although the expansion of this gene family likely arose mainly through other events. Tandem duplications were a critical but recent gene duplication contributor in Arabidopsis [62]. Previous studies have shown that gene duplication greatly accounts for new genes [63]. Gene duplication may result in sub-functionalization: for example, expansion of the functions among the wheat WRKY gene family members has occurred through tandem duplication and whole-genome duplication [64]. Tandem duplications and gene clusters have been previously described for multiple WRKY genes in rice within the same intergenic region [65].
The presence of the conserved WRKY domain, which binds to the W-box motif in the promoters of WRKY target genes, is the most essential characteristic of the WRKY family [66, 67]. We performed a classification of SbWRKY genes here according to the approach used in other crop species, based on phylogenetic tree topology. We also adopted the divisions of WRKY family members into the same groups and subgroups described in Arabidopsis: groups I, II, and III according to the number of WRKY domains and the type of zinc finger motif, along with further subdivision of group II into subgroups IIa, IIb, IIc, IId, and IIe [1]. In this study, 11, 50, and 31 SbWRKY genes were classified into groups I, II, and III, respectively; two genes did not belong to any group. Group II was the largest with 50 members and accounted for 53.2% of all SbWRKY genes. These results are consistent with WRKY group sizes in sesame, Arabidopsis, and sugar beet [25, 1, 46]. Among the subgroups, subgroup IIc was the largest with 20 SbWRKY genes, or 40% of the genes assigned to group II (Fig 2), which is similar to results in sugar beet [48], Arabidopsis [1], and soybean [68].
Whereas most SbWRKY proteins contained the conserved WRKYGQK motif, other heptapeptide variants were identified in 13 SbWRKY proteins (SbWRKY11, SbWRKY14, SbWRKY23, SbWRKY27, SbWRKY33, SbWRKY51, SbWRKY52, SbWRKY53, SbWRKY71, SbWRKY75, SbWRKY77, SbWRKY81, and SbWRKY83; Table 1). Similar variations in the heptapeptide motif were also identified in potato [27]. Several examples indicate that these differences may affect the binding ability of WRKY TFs to the W-box element. For example, two soybean WRKYs with the WRKYGKK motif variant were unable to bind to the W-box element [66]. In tobacco (Nicotiana tabacum), NtWRKY12 with the WRKYGKK motif bound to TTTTCCAC instead of the W-box consensus sequence (TTGACT/C) [69]. Therefore, further investigation is warranted to identify the preferred DNA-binding sequences associated with different WRKYGQK-like motifs.
Group-specific patterns were detected in their exon–intron structure. Indeed, SbWRKY genes within the same group had similar exon–intron patterns. The number of introns in SbWRKY genes ranged from 0 to 7, which is similar to that reported in chickpea [26]. Four SbWRKY genes lacked an intron, indicating that intron loss may have occurred during the evolution of this gene family; a similar observation was reported in the rice WRKY gene family [46]. Intron-less genes within plant gene families may imply their close relationship. Intron-less genes is not a specific feature of the SbWRKY gene family, as they have been reported in other gene families like GRAS-domain TFs, F-box TFs [70], small auxin-up RNAs [71], and DEAD-box RNA helicases [72]. Intron-less genes may arise from one of three major mechanisms: retroposition (the integration of a sequence derived from RNA into the genome), duplication of existing intron-less genes, and horizontal gene transfer [73].
The variations in intron sizes within and between SbWRKY groups may have resulted from duplication, inversion, and/or fusion events [74]. These results were similar to findings in wheat [22], carrot (Daucus carota) [74], and cassava [43]. Overall, the diversification of the exon-intron pattern will provide important clues about the evolution of SbWRKY genes.
Among 20 identified functional motifs in SbWRKY proteins, motifs 1, 2, 3, and 4 corresponded to WRKY domains containing zinc finger domains that are present in most SbWRKY members. Motif 8 represented the nuclear localization signal (NLS), mainly distributed in subgroups IId and IIe. As described previously, members of subgroup IId possess an NLS and a conserved calmodulin-binding domain. Interestingly, three members of subgroup IId (SbWRKY74, SbWRKY62, and SbWRKY64) contained the conserved HARF motif (RTGHARFRR [A/G] P), which was also identified in Arabidopsis and poplar (Populus trichocarpa) WRKY subgroup IId, although the function of this motif is unknown [75, 1]. Some motifs were located nearby the WRKY domain, for instance, motifs 6, 9, and 17. Just as phylogenetic analysis divided SbWRKY genes into groups I-III and subgroups IIa-e, the presence or absence of shared motifs between SbWRKY proteins followed the same general separation into groups, consistent with previous studies [76–79]. Indeed, each of these motifs occurs in most subgroups, and each subgroup can be distinguished based on the motifs they present. Group-specific motifs might be involved in responses to a given biological process [80] and may provide clues about their potential function. The functions of other motifs identified by MEME are yet to be elucidated.
The promoter regions of 94 SbWRKY genes exhibited various conserved cis-acting regulatory elements involved in various functions, such as abiotic and biotic stress responses (MBS, LTR, ARE, TC-rich repeat, and GC-motif) and phytohormone regulation (ABRE, TCA-element, TGA-element, TGACG-motif, CGTCA-element, AuxRR-core, and GARE-motif). The presence of many cis-acting elements mediating responses to environmental stress and phytohormones indicates their involvement in different biological processes. Regulation of WRKY expression may occur via binding of a WRKY TF to W-box or by the binding of another TF to a different cis-element along WRKY promoters [81]. The wheat TaWRKY2 and TaWRKY19 exert their regulation of gene expression by binding to the promoter regions of target genes when overexpressed in Arabidopsis [82]. WRKY TFs may regulate the expression of their encoding genes by binding to their promoter or may regulate other WRKY TFs by cross-regulation [83]. Consistent with this hypothesis, the promoters of 38 SbWRKY genes had one or more W-boxes.
The essential roles of WRKY TFs in plant growth, development, and stress tolerance are supported by WRKY gene expression data from several species. From extensive studies in the model plant Arabidopsis, many AtWRKYs have been functionally characterized. Therefore, identifying the closest Arabidopsis homologue(s) of individual SbWRKYs may provide a hint as to their potential functions. For example, the highly expressed SbWRKY72 gene is most closely related to the Arabidopsis AtWRKY70 and AtWRKY54, which have been reported to modulate osmotic stress tolerance by regulating stomatal aperture [84].
SbWRKY79, SbWRKY80, and SbWRKY42 are putative orthologs of AtWRKY25, AtWRKY26, and AtWRKY33, which regulate heat shock proteins and the heat-induced ethylene-dependent response [85]. AtWRKY53 and AtWRKY70 belong to group III and both play important roles in leaf senescence [86]. SbWRKY72 and SbWRKY75 also belong to group III and are highly expressed in leaves. In addition, AtWRKY70 plays a critical role in osmotic stress signalling and plant defense responses in Arabidopsis [87]. SbWRKY19 and SbWRKY73, members of group I, are expressed in root tissues, although their putative Arabidopsis counterpart AtWRKY34 is involved in regulating gene expression during tapetum formation [88]. Several group II-a WRKY genes have documented roles in biotic stress responses in Arabidopsis [42]. Moreover, AtWRKY8, AtWRKY50 and AtWRKY57 are involved in phytohormones signalling and pathogen responses [89]. The highly expressed sorghum gene SbWRKY45 is the putative ortholog of AtWRKY18, AtWRKY40, and AtWRKY60, which are involved in abscisic acid signalling and abiotic stress [90]. Additionally, SbWRKY45 is the putative ortholog of maize ZmWRKY40, which confers drought resistance when expressed in transgenic Arabidopsis [12]. A putative ortholog of SbWRKY8 is Arabidopsis WRKY57, which can improve drought tolerance through elevated abscisic acid levels [52].
Based on the available sorghum transcriptomic data, the present analysis of SbWRKY genes revealed their different expression patterns at different growth stages and in different tissues. As indicated by the above examples, the known functions of putative Arabidopsis WRKY orthologs, together with expression data, will guide future functional analyses of SbWRKY genes, with a focus on their roles in responses to environmental stress. Highly expressed SbWRKY genes identified during our digital expression analysis of published sorghum microarrays were validated by qRT-PCR, which also provided general clues on SbWRKY responses to drought stress. We tested five genes that belonged to group I (SbWRKY79), IIa (SbWRKY45), IId (SbWRKY74), and III (SbWRKY72 and SbWRKY75). All selected genes were upregulated in response to drought. WRKY genes from the same groups were reported to play important roles in other plants. For example, the expression of sweet potato group I members ItfWRKY66, ItfWRKY69, and ItfWRKY80 was induced in response to drought, cold and salt stresses [53]. Furthermore, AtWRKY25 overexpression improved heat and salt stress tolerance in Arabidopsis [91]. Its putative ortholog SbWRKY79 was induced by drought as shown by RT-qPCR. Several putative SbWRKYs orthologs have been confirmed to be involved in drought stress tolerance in other crops. For example, SbWRKY45 was orthologous to maize ZmWRKY40, which is itself involved in drought stress tolerance [12].
Since we only tested a small fraction of SbWRKY genes, other SbWRKY genes may also be involved in drought stress responses. Therefore, further investigation into SbWRKY expression under other abiotic stress conditions (cold, salinity, and heat) is necessary. Our results provide several promising SbWRKY candidates for these future studies.
Conclusion
This study identified 94 WRKY genes in sorghum, and the following analyses were performed: characterization and classification, gene structure analysis, chromosome mapping, and conserved motif analysis. SbWRKY gene expression profiles indicated that SbWRKY genes may be important in different tissues and at different developmental stages. Several SbWRKY genes displayed tissue-specific expression. Besides, several SbWRKY genes were highly expressed in response to environmental stresses. qRT-PCR analysis revealed several SbWRKY genes induced by drought stress. In Arabidopsis, many AtWRKYs regulate abiotic and biotic stress responses, and the available information about specific WRKY members will facilitate functional validation and characterization of their putative orthologs in sorghum. Overall, the present findings provide a foundation for future functional analyses of SbWRKY genes in response to abiotic and biotic stress in sorghum.
Supporting information
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(DOCX)
(XLSX)
(DOCX)
Acknowledgments
The authors sincerely thank Professor Fuyao Zhang, Sorghum Institute, Shanxi Academy of Agriculture Sciences, Shanxi, for providing sorghum seeds used in this research. We thank Anfal for helpful suggestions and advice.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This project was supported by The National Key Research and Development Program of China (2016YFD0100605), the Innovation Academy for Seed Design, Chinese Academy of Sciences; special exchange programme of the Chinese academy of sciences (category A).The authors sincerely thank the Center for Agricultural Resources Research, Institute of Genetics and Developmental Biology (CAS) for providing the facilities‛.
References
- 1.Eulgem T, Rushton PJ, Robatzek S, Somssich IE. The WRKY superfamily of plant transcription factors. Trends in Plant Sci. 2000; 5(5): 199–206. 10.1016/S1360-1385(00)01600-9 [DOI] [PubMed] [Google Scholar]
- 2.Rushton PJ, Somssich IE, Ringler P, Shen QJ. WRKY transcription factors. Trends in Plant Sci. 2010; 15(5): 247–258. 10.1016/j.tplants.2010.02.006 [DOI] [PubMed] [Google Scholar]
- 3.Rinerson CI, Rabara RC, Tripathi P, Shen QJ, Rushton PJ. The evolution of WRKY transcription factors. BMC plant Biol. 2015; 15: 1–18. 10.1186/s12870-014-0410-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bakshi M, Oelmüller R. WRKY transcription factors, Plant Signalling and Behavior. J. Plant Signal. Behav. 2014; 9, e27700 10.4161/psb.27700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Finatto T, Viana VE, Woyann LG, Busanello C, Maia LCD, Oliveira AC. Can WRKY transcription factors help plants to overcome environmental challenges?. Genet Mol Biol. 41(3) 2018:533‐544. 10.1590/1678-4685-GMB-2017-0232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Liu H, Qin J, Fan H, Cheng J, Li L, Liu Z. Genome-wide identification, phylogeny and expression analyses of SCARECROW—LIKE (SCL) genes in millet (Setaria italica). Phys and Moleo Biol of Plants. 2017; 23(3): 629–640. 10.1007/s12298-017-0455-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pandey SP, Somssich IE. The role of WRKY transcription factors in plant immunity. Plant Physiol. 2009; 150(4): 1648–1655. 10.1104/pp.109.138990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ulker B, Somssich IE. WRKY transcription factors: from DNA binding towards biological function. Curr Opin Plant Biol. 2004; 7(5):491–498. 10.1016/j.pbi.2004.07.012 [DOI] [PubMed] [Google Scholar]
- 9.Chen L, Song Y, Li S, Zhang L, Zou C, Yu D. The role of WRKY transcription factors in plant abiotic stresses. Biochim Biophys Acta. 2012; 1819(2):120–128. 10.1016/j.bbagrm.2011.09.002 [DOI] [PubMed] [Google Scholar]
- 10.Wang C, Deng P, Chen L, et al. A wheat WRKY transcription factor TaWRKY10 confers tolerance to multiple abiotic stresses in transgenic tobacco. PLoS One. 2013; 8(6):e65120 10. 10.1371/journal.pone.0065120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.He GH, Xu JY, Wang YX, Liu JM, Li PS, Chen M, et al. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis. BMC Plant Biol. 2016; 16: 1–16, 10.1186/s12870-015-0700-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang CT, Ru JN, Liu YW, Yang JF, Li M, Xu ZS, et al. The Maize WRKY transcription factor ZmWRKY40 confers drought resistance in transgenic Arabidopsis. Inter J of Mole Sci. 2018; 19(9): 2580 10.3390/ijms19092580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhou S, Zheng WJ, Liu BH, et al. Characterizing the Role of TaWRKY13 in salt tolerance. Int J Mol Sci. 2019;20(22):5712 10.3390/ijms20225712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sheng Y, Yan X, Huang Y, Han Y, Zhang C, Ren Y, et al. The WRKY transcription factor, WRKY13, activates PDR8 expression to positively regulate cadmium tolerance in Arabidopsis. Plant Cell and Envir. 2019; 42(3):891–903. 10.1111/pce.13457 [DOI] [PubMed] [Google Scholar]
- 15.Cao S, Wu X, Ouyang J, Jiang L, Zhu X, Fan T, et al. WRKY12 represses GSH1 expression to negatively regulate cadmium tolerance in Arabidopsis. Plant Mol. Biol. 2019; 99: 149–159, 10.1007/s11103-018-0809-7 [DOI] [PubMed] [Google Scholar]
- 16.Cai R, Dai W, Zhang C, Wang Y, Wu M, Zhao Y, et al. Cheng, B, The maize WRKY transcription factor ZmWRKY17 negatively regulates salt stress tolerance in transgenic Arabidopsis plants. Planta. 2017; 246: 1215–1231, 10.1007/s00425-017-2766-9 [DOI] [PubMed] [Google Scholar]
- 17.Gao H, Wang Y, Xu P, Zhang Z. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant Sci. 2018; 9: 997 10.3389/fpls.2018.00997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wu X, Shiroto Y, Kishitani S, Ito Y, Toriyama K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009; 28(1): 21‐30. 10.1007/s00299-008-0614-x [DOI] [PubMed] [Google Scholar]
- 19.Zhang L, Zhao T, Sun X, et al. Overexpression of VaWRKY12, a transcription factor from Vitis amurensis with increased nuclear localization under low temperature, enhances cold tolerance of plants. Plant Mol Biol. 2019; 100(1–2): 95‐110. 10.1007/s11103-019-00846-6 [DOI] [PubMed] [Google Scholar]
- 20.Tang H, Cuevas HE, Das S, Sezen UU, Zhou C, Guo H, et al. Seed shattering in a wild sorghum is conferred by a locus unrelated to domestication. Proceedings of the National Academy of Sciences. 2013; 110:15824 10.1073/pnas.1305213110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ishiguro S, Nakamura K. Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5' upstream regions of genes coding for sporamin and beta-amylase from sweet potato. Mol Gen Genet. 1994; 244(6): 563–571. 10.1007/BF00282746 [DOI] [PubMed] [Google Scholar]
- 22.Ning P, Liu C, Kang J, Lv J. Genome-wide analysis of WRKY transcription factors in wheat (Triticum aestivum L.) and differential expression under water deficit condition. PeerJ. 2017; 5, e3232 10.7717/peerj.3232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wei KF, Chen J, Chen YF, Wu LJ, Xie DX. Molecular phylogenetic and expression analysis of the complete WRKY transcription factor family in maize. DNA Research. 2012; 19(2): 153–164. 10.1093/dnares/dsr048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ramamoorthy R, Jiang SY, Kumar N, Venkatesh PN, Ramachandran S. A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments. Plant and Cell Physio. 2008; 49(6): 865–879. 10.1093/pcp/pcn061 [DOI] [PubMed] [Google Scholar]
- 25.Li D, Liu P, Yu J, Wang L, Dossa K, Zhang Y, et al. Genome-wide analysis of WRKY gene family in the sesame genome and identification of the WRKY genes involved in responses to abiotic stresses. BMC Plant Biol. 2017; 17(1): 1–19. 10.1186/s12870-016-0951-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Waqas M, Azhar MT, Rana IA, Azeem F, Ali MA, Nawaz MA, et al. , Genome-wide identification and expression analyses of WRKY transcription factor family members from chickpea (Cicer arietinum L.) reveal their role in abiotic stress-responses. Genes and Genomics. 2019; 0(0): 0. 10.1007/s13258-018-00780-9 [DOI] [PubMed] [Google Scholar]
- 27.Zhang C, Wang D, Yang C, Kong N, Shi Z, Zhao P, et al. Genome-wide identification of the potato WRKY transcription factor family. PLoS ONE. 2017; 12(7): 1–20. 10.1371/journal.pone.0181573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kumar A, Reddy B, Sharma H, Hash C, Rao P, Ramiah B, et al. Recent advances in Sorghum Genetic Enhancement Research at ICRISAT. American J of plant Sci. 2011; 02(04): 589–600. 10.4236/ajps.2011.24070 [DOI] [Google Scholar]
- 29.Woldesemayat AA, Ntwasa M. Pathways and Network Based Analysis of Candidate Genes to Reveal Cross-Talk and Specificity in the Sorghum (Sorghum bicolor (L.) Moench) Responses to Drought and It’s Co-occurring Stresses. Frontiers in Genetics. 2018; 9(11): 1–22 10.3389/fgene.2018.00557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Woldesemayat AA, Heusden PVan, Ndimba BK, Christoffels A. An integrated and comparative approach towards identification, characterization and functional annotation of candidate genes for drought tolerance in sorghum (Sorghum bicolor (L.) Moench). BMC Genetics. 2017; 18 (1): 1–24. 10.1186/s12863-016-0468-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Letunic I, Bork P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Research. 2018; 46(D1): 493–D496. 10.1093/nar/gkx922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ren A, Ahmed R, Chen H, Han L, Sun J, Ding A, et al. Genome-Wide Identification, Characterization and Expression Patterns of the Pectin Methylesterase Inhibitor Genes in Sorghum bicolor. Genes. 2019; 10(10): 755 10.3390/genes10100755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Molec Biol and Evolution. 2016; 33(7): 1870–1874. 10.1093/molbev/msw054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hu B, Jin J, Guo AY, Zhang H, Luo J, Gao G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics. 2015; 31(8): 1296–1297. 10.1093/bioinformatics/btu817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zou Z, Yang L, Wang D, Huang Q, Mo Y, Xie G. Gene structures, evolution and transcriptional profiling of the WRKY gene family in castor bean (Ricinus communis L.). PLoS ONE. 2016; 11(2): 1–23. 10.1371/journal.pone.0148243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Götz S, García-Gómez JM, Terol J, et al. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008; 36(10): 3420–3435. 10.1093/nar/gkn176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gernnam AK. Genevestigator Impact Facilitating Analysis. Plant physio. 2006; 141(4): 1164–1166. 10.1104/pp.104.900198.1164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Osmolovskaya N, Shumilina J, Kim A, et al. Methodology of drought stress research: experimental setup and physiological characterization. Int J Mol Sci. 2018; 19(12): 4089 https://doi:10.3390/ijms19124089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Livak KJ, Schmittgen TD, Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Elsevier Science. 2001; 25: 402–408, 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- 40.Cheng Y, Yao ZP, Ruan MY, Ye QJ, Wang RQ, Zhou GZ. In silico identification and characterization of the WRKY gene superfamily in pepper (Capsicum annuum L.), Genet Mol Res. 2016; 15(3): 1–12. http://doi.10.4238/gmr.15038675 [DOI] [PubMed] [Google Scholar]
- 41.Shakoor N, Nair R, Crasta O, Morris G, Feltus A, Kresovich S, A Sorghum bicolor expression atlas reveals dynamic genotype-specific expression profiles for vegetative tissues of grain, sweet and bioenergy sorghums. BMC Plant Biol. 2014; 14(1): 10.1186/1471-2229-14-35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Baillo EH, Kimotho RJ, Zhang ZB, Xu P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes. 2019; 10(10): 771 10.3390/genes10100771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wei Y, Shi H, Xia Z, Tie W, Ding Z, Yan Y, et al. Genome-Wide Identification and Expression Analysis of the WRKY Gene Family in Cassava. Front in Plant Sci. 2016; 7(2): 1–18. 10.3389/fpls.2016.00025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wang M, Vannozzi A, Wang G, Liang YH, Tornielli GB, Zenoni S, et al. , Genome and transcriptome analysis of the grapevine (Vitis vinifera L.) WRKY gene family. Horticulture Research. 2014; 1(2): 4016 10.1038/hortres.2014.16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yu Y, Wang N, Hu R, Xiang F. Genome‑wide identification of soybean WRKY transcription factors in response to salt stress. SpringerPlus. 2016; 5: 920 10.1186/s40064-016-2647-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ross CA, Liu Y, Shen QJ. The WRKY Gene family in Rice (Oryza sativa), J of integr Pl Biol. 2007; 49(6): 827–842. 10.1111/j.1672-9072.2007.00504.x [DOI] [Google Scholar]
- 47.Chen X, Chen R, Wang Y, Wu C, Huang J. Genome-Wide Identification of WRKY Transcription Factors in Chinese jujube (Ziziphus jujuba Mill.) and Their Involvement in Fruit Developing, Ripening, and Abiotic Stress. Genes. 2019; 10: 360 10.3390/genes10050360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wu G, Li Z. W, Cao H, Wang J. Genome-wide identification and expression analysis of the WRKY genes in sugar beet (Beta vulgaris L.) under alkaline stress. PeerJ. 2019; 18 (6): e7817 10.7287/peerj.preprints.27810v1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Dong X, Yang Y, Zhang Z, Xiao Z, Bai X, Gao J, Genome-Wide Identification of WRKY genes and their Rresponse to cold stress in Coffea canephora. Forests. 2019; 10 (335): 1–15. 10.3390/f10040335 [DOI] [Google Scholar]
- 50.Zheng J, Liu F, Zhu C, et al. Identification, expression, alternative splicing and functional analysis of pepper WRKY gene family in response to biotic and abiotic stresses. PLoS One. 2019; 14(7): e0219775 10.1371/journal.pone.0219775 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yang Y, Liu J, Zhou X, Liu S, Zhuang Y. Identification of WRKY gene family and characterization of cold stress-responsive WRKY genes in eggplant. PeerJ. 2020; 8: e8777 10.7717/peerj.8777 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Srivastava R, Kumar S, Kobayashi Y, et al. Comparative genome-wide analysis of WRKY transcription factors in two Asian legume crops: Adzuki bean and Mung bean. Sci Rep. 2018; 8(1):16971 10.1038/s41598-018-34920-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li Y, Zhang L, Zhu P, et al. Genome-wide identification, characterisation and functional evaluation of WRKY genes in the sweet potato wild ancestor Ipomoea trifida (H.B.K.) G. Don. under abiotic stresses. BMC Genet. 2019;20(1):90 10.1186/s12863-019-0789-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chanwala J, Satpati S, Dixit A, Parida A, Giri MK, Dey N. Genome-wide identification and expression analysis of WRKY transcription factors in pearl millet (Pennisetum glaucum) under dehydration and salinity stress. BMC Genomics. 2020;21(1):231 10.1186/s12864-020-6622-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Rama Reddy NR, Ragimasalawada M, Sabbavarapu MM, Nadoor S, Patil JV. Detection and validation of stay-green QTL in post-rainy sorghum involving widely adapted cultivar, M35-1 and a popular stay-green genotype B35. BMC Genomics. 2014;15(1):909 10.1186/1471-2164-15-909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kamal NM, Gorafi YSA, Tsujimoto H, Ghanim AMA. Stay-Green QTLs Response in Adaptation to Post-Flowering Drought Depends on the Drought Severity. Biomed Res Int. 2018;2018:7082095 Published 2018 Nov 18. 10.1155/2018/7082095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sanchez AC, Subudhi PK, Rosenow DT, Nguyen HT. Mapping QTLs associated with drought resistance in sorghum (Sorghum bicolor L. Moench). Plant Mol Biol. 2002;48(5–6):713–726. 10.1023/a:1014894130270 [DOI] [PubMed] [Google Scholar]
- 58.Grassi ADe, Lanave C, Saccone C. Genome duplication and gene-family evolution: The case of three OXPHOS gene families. BMC Evol Biol. 2008; 421: 1–6. 10.1016/j.gene.2008.05.011 [DOI] [PubMed] [Google Scholar]
- 59.Cannon SB, Mitra A, Baumgarten A, Young ND, May G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC plant Biol. 2004; 21: 1–21. 10.1186/1471-2229-4-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhu Y, Wu N, Song W, et al. Soybean (Glycine max) expansin gene superfamily origins: segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014;14:93 Published 2014 Apr 11. 10.1186/1471-2229-14-93 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kong H, Landherr LL, Frohlich MW, Leebens-Mack J, Ma H, dePamphilis CW: Patterns of gene duplication in the plant SKP1 gene family in angiosperms: evidence for multiple mechanisms of rapid gene birth. Plant J. 2007; 50: 873–885. 10.1111/j.1365-313X.2007.03097.x [DOI] [PubMed] [Google Scholar]
- 62.Yang L, Gaut B. Factors that contribute to variation in evolutionary rate among Arabidopsis genes. Molecular Biology and Evolution. 2011; 28(8):2359–2369. 10.1093/molbev/msr058 [DOI] [PubMed] [Google Scholar]
- 63.Yin G, Xu H, Xiao S, et al. The large soybean (Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups. BMC Plant Biol. 2013;13:148 10.1186/1471-2229-13-148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hassan S, Lethin J, Blomberg R, Mousavi H, Aronsson H. In silico based screening of WRKY genes for identifying functional genes regulated by WRKY under salt stress. Comput Biol Chem. 2019;83:107131 10.1016/j.compbiolchem.2019.107131 [DOI] [PubMed] [Google Scholar]
- 65.Xu H, Watanabe KA, Zhang L, Shen QJ. WRKY transcription factor genes in wild rice Oryza nivara. DNA Res. 2016;23(4):311–323. 10.1093/dnares/dsw025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bi C, Xu Y, Ye Q, Yin T, Ye N. Genome-wide identification and characterization of WRKY gene family in Salix suchowensis. PeerJ. 2016; 4, e2437 10.7717/peerj.2437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ding M, Chen J, Jiang Y, Lin L, Cao Y, Wang M. Genome‑wide investigation and transcriptome analysis of the WRKY gene family in Gossypium. Mol Genet Genomics. 2014; 5 (9): 4–7 10.1007/s00438-014-0904-7 [DOI] [PubMed] [Google Scholar]
- 68.Luo X, Bai X, Sun X, Zhu D, Liu B, Ji W, et al. Expression of wild soybean WRKY20 in Arabidopsis enhances drought tolerance and regulates ABA signalling. J of Exp Bot. 2013; 6 (12): 64(8): 2155–69. 10.1093/jxb/ert073 [DOI] [PubMed] [Google Scholar]
- 69.Verk Van MC, Pappaioannou D, Neeleman L, Bol JF, Linthorst HJM. A Novel WRKY transcription factor is required for induction of PR-1a gene expression by salicylic acid. Plant physiol. 2008; 146(4): 1983–1995. 10.1104/pp.107.112789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Jain M, Nijhawan A, Arora R, et al. F-box proteins in rice. Genome-wide analysis, classification, temporal and spatial gene expression during panicle and seed development, and regulation by light and abiotic stress. Plant Physiol. 2007;143(4):1467–1483. 10.1104/pp.106.091900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Jain M, Tyagi AK, Khurana JP. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics. 2006;88(3):360–371. 10.1016/j.ygeno.2006.04.008 [DOI] [PubMed] [Google Scholar]
- 72.Aubourg S, Kreis M, Lecharny A. The DEAD box RNA helicase family in Arabidopsis thaliana. Nucleic Acids Res. 1999;27(2):628–636. 10.1093/nar/27.2.628 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zou M, Guo B, He S. The roles and evolutionary patterns of intronless genes in deuterostomes. Comp Funct Genomics. 2011:680673 10.1155/2011/680673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li MY, Xu ZS, Tian C, Huang Y, Wang F, Xiong AS. Genomic identification of WRKY transcription factors in carrot (Daucus carota) and analysis of evolution and homologous groups for plants. Scientific Reports. 2016; 6(7): 1–17. 10.1038/srep23101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.He H, Dong Q, Shao Y, et al. Genome-wide survey and characterization of the WRKY gene family in Populus trichocarpa. Plant Cell Rep. 2012; 31(7): 1199–1217. 10.1007/s00299-012-1241-0 [DOI] [PubMed] [Google Scholar]
- 76.He C, Teixeira da Silva JA, Tan J, et al. A Genome-wide identification of the WRKY family genes and a survey of potential WRKY target genes in Dendrobium officinale. Sci Rep. 2017; 7(1): 9200 10.1038/s41598-017-07872-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Guo H, Zhang Y, Wang Z, et al. Genome-Wide Identification of WRKY Transcription Factors in the Asteranae. Plants. 2019; 8(10): 393 10.3390/plants8100393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Chen P, Liu QZ. Genome-wide characterization of the WRKY gene family in cultivated strawberry (Fragaria ananassa Duch.) and the importance of several group III members in continuous cropping. Sci Rep. 2019;9(1):8423 10.1038/s41598-019-44479-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Nan H, Gao LZ. Genome-wide analysis of WRKY genes and their response to hormone and mechanic stresses in carrot. Front Genet. 2019; 10: 363 10.3389/fgene.2019.00363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Goel R, Pandey A, Trivedi PK, Asif MH. Genome-wide analysis of the musa WRKY gene family: fvolution and differential expression during development and stress. Front Plant Sci. 2016;7:299 10.3389/fpls.2016.00299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Phukan UJ, Jeena GS, Shukla RK. WRKY Transcription factors: molecular regulation and Stress responses in rlants. Front Plant Sci. 2016;7:760 10.3389/fpls.2016.00760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Niu CF, Wei W, Zhou QY, et al. Wheat WRKY genes TaWRKY2 and TaWRKY19 regulate abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Environ. 2012;35(6):1156–1170. 10.1111/j.1365-3040.2012.02480.x [DOI] [PubMed] [Google Scholar]
- 83.Tao Z, Kou Y, Liu H, Li X, Xiao J, Wang S. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice. J Exp Bot. 2011;62(14):4863–4874. 10.1093/jxb/err144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Li J, Besseau S, Petri T. Defense-related transcription factors WRKY70 and WRKY54 modulate osmotic stress tolerance by regulating stomatal aperture in Arabidopsis. New Phytol. 2013; 4: 57–472. 10.1111/nph.12378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Li S, Fu Q, Chen L. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermotolerance. Planta. 2011; 1:237–1252. 10.1007/s00425-011-1375-2 [DOI] [PubMed] [Google Scholar]
- 86.Chen H, Lai Z, Shi J, Xiao Y, Chen Z, Xu X. Roles of arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biology. 2010; 10(1): 281 10.1186/1471-2229-10-281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ay N, Irmler K, Fischer A, Uhlemann R, Reuter G, Humbeck K. Epigenetic programming via histone methylation at WRKY53 controls leaf senescence in Arabidopsis thaliana. Plant J. 2009; 58(2): 333‐346. 10.1111/j.1365-313X.2008.03782.x [DOI] [PubMed] [Google Scholar]
- 88.Honys D, Oh SA, Renák D, et al. Identification of microspore-active promoters that allow targeted manipulation of gene expression at early stages of microgametogenesis in Arabidopsis. BMC Plant Biol. 2006;6:31 10.1186/1471-2229-6-31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jiang Y, Yu D. WRKY transcription factors: links between phytohormones and plant processes. Sci China Life Sci. 2015; 58(5): 501‐502. 10.1007/s11427-015-4849-9 [DOI] [PubMed] [Google Scholar]
- 90.Jiang Y, Liang G, Yu D. Activated expression of WRKY57 confers drought tolerance in Arabidopsis. Molecular Plant. 1 2012; 5(6): 375–1388. 10.1093/mp/sss080 [DOI] [PubMed] [Google Scholar]
- 91.Fu QT, Yu DQ. Expression profiles of AtWRKY25, AtWRKY26 and AtWRKY33 under abiotic stresses. Hereditas. 2010; 32: 848–56. 10.3724/sp.j.1005.2010.00848 [DOI] [PubMed] [Google Scholar]