

Abstract
The severe acute respiratory syndrome coronavirus‐2 (SARS‐COV‐2), a novel coronavirus responsible for the recent infectious pandemic, is known to downregulate angiotensin‐converting enzyme‐2 (ACE2). Most current investigations focused on SARS‐COV‐2‐related effects on the renin–angiotensin system and especially the resultant increase in angiotensin II, neglecting its effects on the kinin–kallikrein system. SARS‐COV‐2‐induced ACE2 inhibition leads to the augmentation of bradykinin 1‐receptor effects, as ACE2 inactivates des‐Arg9‐bradykinin, a bradykinin metabolite. SARS‐COV‐2 also decreases bradykinin 2‐receptor effects as it affects bradykinin synthesis by inhibiting cathepsin L, a kininogenase present at the site of infection and involved in bradykinin production. The physiologies of both the renin–angiotensin and kinin–kallikrein system are functionally related suggesting that any intervention aiming to treat SARS‐COV‐2‐infected patients by triggering one system but ignoring the other may not be adequately effective. Interestingly, the snake‐derived bradykinin‐potentiating peptide (BPP‐10c) acts on both systems. BPP‐10c strongly decreases angiotensin II by inhibiting ACE, increasing bradykinin‐related effects on the bradykinin 2‐receptor and increasing nitric oxide‐mediated effects. Based on a narrative review of the literature, we suggest that BPP‐10c could be an optimally effective option to consider when aiming at developing an anti‐SARS‐COV‐2 drug.
Keywords: COVID‐19, kinin–kallikrein system, snake‐derived bradykinin‐potentiating peptide
1. INTRODUCTION
A novel frightening coronavirus disease (COVID‐19) caused by the severe acute respiratory syndrome coronavirus‐2 (SARS‐CoV‐2) has spread worldwide since December 2019. COVID‐19 is mainly a contagious acute respiratory infection that can lead to life‐threatening presentations and fatalities (Phan et al., 2020). On March 2020, the World Health Organization declared that the outbreak is a pandemic (Habibzadeh & Lang, 2020). On mid‐June 2020, the total number of reported cases worldwide was ~7.7 million causing ~430,000 deaths (Worldometers.Info).
To date, an unbelievable number of research projects have been conducted. Full‐length genome analyses of SARS‐CoV‐2 were obtained concluding its 96% identity with bat coronaviruses (Zhou et al., 2020). Studies of the virus relative synonymous codon usage bias revealed closeness to the snake, suggesting that snake may be an animal reservoir (Ji, Wang, Zhao, Zai, & Li, 2020). Researchers are currently investigating infectivity, mode of transmission, mechanisms of disease and clinical effects aiming at identifying effective therapeutic and preventative options. Clinical trials are in process to test various anti‐SARS‐CoV‐2 therapies; but none is yet approved (Ahsan, Javed, Al Bratty, Alhazmi, & Najmi, 2020).
SARS‐CoV‐2, a positive‐stranded RNA virus, contains major structural proteins including the spike (S), envelope (E), membrane (M) and nucleocapsid proteins (Wu et al., 2020). SARS‐CoV‐2 spike protein interacts with an epithelial cell surface receptor identified as the angiotensin‐converting enzyme‐2 (ACE2) allowing the virus to enter cells and cause the infection (Walls et al., 2020). SARS‐CoV‐2/ACE2 interaction rapidly induces ACE2 downregulation (Hoffmann et al., 2020; Silhol, Sarlon, Deharo, & Vaïsse, 2020). ACE2 is a zinc‐containing metalloenzyme expressed predominantly at the membrane surface of vascular endothelia cells, type II pneumocytes and various epithelia cells (Sodhi et al., 2018). ACE2 expression correlates with sites of infection including lung and intestine (Hamming et al., 2004), consistent with COVID‐19‐related main respiratory and intestinal manifestations (Xu, Zhou, & Xu, 2020).
Almost all studies interpreted the COVID‐19‐attributed ACE2 downregulation and the subsequent inflammation as related to the renin–angiotensin system (RAS) dysfunction (Gurwitz, 2020; Hoffmann et al., 2020; Lu et al., 2020). Rare studies suggested COVID‐19‐associated kinin–kallikrein system (KKS) dysregulation as potential trigger for the observed inflammation mediated by the innate immune response (van de Veerdonk et al., 2020). The role of ACE2 is crucial since its altered activity affects both RAS and KKS (Gralinski et al., 2018; Kuba et al., 2005; Sampaio et al., 2007; Santos et al., 2003; Sodhi et al., 2018; Yang, Yin, Li, Zimmerman, & Schultz, 2011). Here we aimed to review the involvement of both systems in COVID‐19 pathogenesis and discuss the potential usefulness of the snake‐derived bradykinin‐potentiating peptides (BPPs), able to interact with both systems, as optimal anti‐COVID‐19 therapy.
2. THE RAS
The RAS regulates blood pressure, systemic vascular resistance and electrolyte balance. Briefly, the renal juxtaglomerular cells secrete renin when the renal blood flow decreases. Plasma renin converts angiotensinogen released by the liver to angiotensin I that subsequently convert angiotensin I to angiotensin II by ACE present at the vascular endothelial cell surface (Figure 1). Angiotensin II is a potent vasoconstrictor and stimulates aldosterone production from the adrenal cortex.
FIGURE 1.
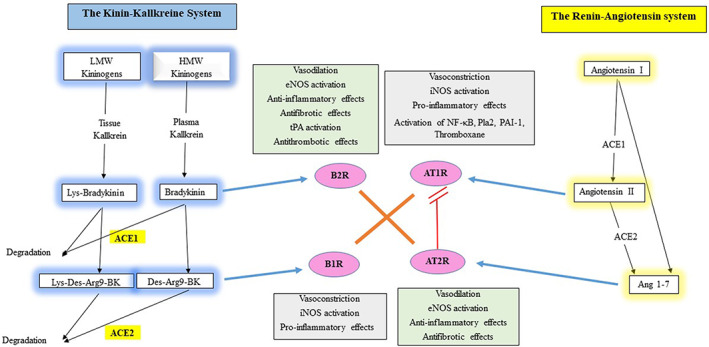
The physiology and functional relationships between the renin angiotensin and KKSs
ACE1 degrades angiotensin I to angiotensin II, which acts through the angiotensin I‐receptor (AT1R; Kuba et al., 2005). Angiotensin II/AT1R stimulates the Janus kinase–signal transducer and activator of transcription (JAK–STAT) signaling pathway to exert its vasoconstriction and hypertensive activity (Recinos et al., 2007). Additionally, angiotensin II/ATI‐R activates the nuclear factor kappa‐light‐chain‐enhancer of activated B cells (NF‐κB) signaling pathway, increases the production of reactive oxygen species (ROS), induces immune cell infiltration and increases the production of C‐reactive protein and proinflammatory cytokines such as the tumor necrosis factor‐alpha (TNF‐α), interleukins (IL)‐1, ‐6, and interferon (IFN)‐y in target tissues (Han, Liu, Liu, & Li, 2010; Lee et al., 2014). Angiotensin II/AT1R‐related stimulation of ROS generation results in nitric oxide (NO) breakdown leading to endothelial dysfunction (Griendling, Lassègue, Murphy, & Wayne Alexander, 1994). Angiotensin II activates mitogen‐activated protein kinase (MAPK) signaling cascade leading to phospholipase A2 (PlA2) activation (Kaschina & Unger, 2003). This activations enhance the secretion of arachidonic acid which mediates various signaling (generated by cytosolic PLA2) and inflammatory processes (generated by secretory PLA2; Baynes & Dominiczak, 2005). Arachidonic acid is metabolized to eicosanoids as prostaglandins, prostacyclin (by the action of cyclooxygenase 1 and 2), and leukotrienes and thromboxanes (by the action of prostaglandin G/H synthase‐1 and 2), which regulate the immunopathological processes of inflammatory responses (Harizi, Corcuff, & Gualde, 2008). Angiotensin II/AT1R causes vasoconstriction, increased vascular permeability, inflammation and fibrosis (Kuba et al., 2005). Angiotensin II enhances arterial vascular thrombosis in arterioles by modulating plasminogen activation mediated by enhanced plasminogen activator inhibitor type 1 (PAI‐1) expression (Senchenkova, Russell, Almeida‐Paula, Harding, & Granger, 2010) and thromboxane production (Smith & Song, 2002).
ACE2 degrades angiotensin II to generate angiotensin (1–7) which subsequently acts through the mas oncogene product (MAS) receptor to counteract angiotensin II/AT1R effects and stimulate NOS (Sampaio et al., 2007). Angiotensin 1–7 stimulates AT2R which has vasodilatatory, antiproliferative, antiinflammatory and antifibrotic effects (Calò, Rigato, & Bertoldi, 2019; Chappell & Al Zayadneh, 2017; Katovich, Grobe, & Raizada, 2008; Rodrigues Prestes, Rocha, Miranda, Teixeira, & Simoes‐e‐Silva, 2017). Angiotensin 1–7 can be inactivated by ACE to inactive angiotensin 1–5 (Santos, Ferreira, & Simões e Silva, 2008). Noteworthy, angiotensin (1–7) can be directly synthesized from angiotensin I by neprilysin (Chappell, 2016), thimet oligopeptidase (Wilson, Nautiyal, Gwathmey, Rose, & Chappell, 2016), prolylendopeptidase and metalloendopeptidase (Deddish et al., 1998).
ACE has two homologous N‐ and C‐domains, present at the N‐terminal half (N‐ACE) or the C‐terminal half (C‐ACE) of the single‐chain protein, respectively. Angiotensin (1–7) is both ACE substrate and inhibitor. It is degraded mainly by N‐ACE and more slowly by C‐ACE and inhibits C‐ACE, which mainly hydrolyzes bradykinin and angiotensin 1 (Deddish et al., 1998).
Collectively, ACE metabolizes angiotensin I to angiotensin II (which induces vasoconstriction, ROS production, immune cell infilteration, inflammatory response, vascular permeability, thrombosis and fibrosis), while ACE2 metabolizes angiotensin II to angiotensin (1–7) (which counteracts angiotensin II and presents vasodilatatory, antiproliferative, antiinflammatory and antifibrotic effects).
3. THE KKS
KKS is a poorly understood hormonal system consisting of blood proteins including bradykinin and kallidin and implicated in inflammation, blood pressure control, coagulation and pain (Figure 1). Kininogens are processed by two types of serine‐proteases called kallikreins and released as part of innate inflammation. Plasma kallikrein processes high‐molecular‐weight kininogen (HMWK) to bradykinin and tissue kallikrein processes kinins from low‐molecular‐weight kininogen (LMWK) to Lys‐bradykinin. Both bradykinin and Lys‐bradykinin interact with the bradykinin receptor‐B2 (B2R) on the endothelial cells. They can both be further processed to des‐Arg9‐bradykinin (from bradykinin degradation by carboxypeptidase M [CPM]) and Lys‐des‐Arg9‐bradykinin (from Lys‐bradykinin degradation by carboxypeptidase N [CPN]), which are ligands of the bradykinin receptor‐B1 (B1R), also present on the endothelial cells and upregulated under inflammatory conditions (Jurado‐Palomo & Caballero, 2017).
B2R agonists are considered as potent vasodilators and organoprotective peptides (Heitsch, 2002). B2R inhibits adenylate cyclase, stimulates phospholipase C that subsequently increases intracellular free calcium and provokes endothelium‐dependent vasodilation by inducing local NO production (Fernandes et al., 2001). B2R stimulation also results in prostacyclin production and increase in endothelium‐derived hyperpolarizing factor and tissue plasminogen activator (tPA; Ancion, Tridetti, Nguyen Trung, Oury, & Lancellotti, 2019). Endothelial NO production triggered by bradykinin plays an organoprotective role by limiting leukocyte adhesion/migration, platelet adhesion/aggregation and smooth muscle contraction/proliferation (Hällgren, Samuelsson, Laurent, & Modig, 1989). Deficient NO bioavailability resulting from reduced NO production by the endothelial NO synthase (eNOS) and increased NO breakdown by ROS are associated with endothelial dysfunction (Tomasian, 2000). Additionally, NO possesses an antiapoptotic activity (Baudin, Berard, Carrier, Legrand, & Drouet, 1997).
Bradykinin is considered to be a potent stimulator of tPA secretion, thus affecting fibrinolysis and thrombolysis (Brown, Nadeau, & Vaughan, 1997). t‐PA/PAI‐1 balance controls thrombi formation and degradation (Bentley et al., 2010). Increases in PAI‐1 and decreases in t‐PA are associated with thrombi formation (Wiman et al., 2000). Bradykinin has antithrombotic effects by increasing t‐PA by an independent direct pathway based on NO production (Rahman et al., 2014).
B1R, poorly expressed on endothelial cells in physiological conditions, is upregulated in proinflammatory conditions (Heitsch, 2002). B1R and B2R have different functions (Wu, Lin, Bernloehr, Hildebrandt, & Doods, 2012). B1R stimulation induced by cytosolic calcium ion contributes to the inflammatory responses (Gurusamy et al., 2016). B1R stimulation has detrimental effects by enhancing the proinflammatory response and causing vasoconstriction that contributes to organ injury including acute respiratory distress syndrome (ARDS; Murugesan et al., 2016; Qadri & Bader, 2018). Moreover, B1R stimulation results in prolonged inducible NO synthase (iNOS) activation which is responsible for prolonged high NO output and deleterious organ effect, whereas B2R stimulation results in eNOS activation which is responsible for short burst of NO and protective organ effects (Erdös, Tan, & Skidgel, 2010; Kuhr, Lowry, Zhang, Brovkovych, & Skidgel, 2010). Kinin‐B1R overexpression induces hypertensive response to Des‐Arg9‐bradykinin and susceptibility to inflammation (Ni et al., 2003).
Kinin degradation is controlled by ACE1 that inactivates bradykinin and ACE2 that inactivates Lys‐des‐Arg9‐bradykinin and des‐Arg9‐bradykinin to decrease their effects on B2R and B1R, respectively (Sodhi et al., 2018). ACE1 inhibition, which results in increased bradykinin effects on B2R, has been associated with systemic angioedema (Jurado‐Palomo & Caballero, 2017). ACE2 inhibition, which increases the effects of Lys des‐Arg9‐bradykinin and des‐Arg9‐bradykinin on B1R, leads to proinflammatory effects associated with pulmonary edema and acute lung injury (Imai et al., 2005; Sodhi et al., 2018).
Collectively, kallikreins processes kininogens to bradykinins. Bradykinins act through endothelial B2R (inducing vasodilation and organoprotective effects) and are further processed to des‐bradykinins which act on B1R which is upregulated in inflammatory conditions (inducing vasoconstriction and inflammatory response). ACE1 inactivates bradykinins and ACE2 inactivates des‐bradykinins.
4. FUNCTIONAL RELATION BETWEEN RAS AND KKS
Studies showed that angiotensin 1–7 and bradykinin actions are strongly interrelated. Angiotensin (1–7) potentiates bradykinin‐induced vasodilation on B2R by inhibiting ACE and releasing NO (Fernandes et al., 2001; Greco, Master Jr, F, Baber, & Kadowitz, 2006; Li, Chappell, Ferrario, & Brosnihan, 1997). Other studies reported that bradykinin/NO cascade mediates AT2R stimulation (Gohlke, Pees, & Unger, 1998; Siragy & Carey, 1996). There is a heterodimerization and strong functional interaction between MAS and AT2R suggesting that they depend on each other (Leonhardt et al., 2017). Bradykinin/NO system mediates counteraction between AT1R and AT2R (Searles & Harrison, 1999). Experimental studies correlated B2R‐attributed organoprotective role and AT1R‐attributed counteracting effects (Taddei & Bortolotto, 2016). B2R is considered as a physiological antagonist of angiotensin II‐related effects on AT1R (Alhenc‐Gelas et al., 2011). This indicates that the functional activities of angiotensin 1–7 (on MAS receptor and AT1R) and bradykinin (on B2R) depend on each other. Also, angiotensin (1–7) can indirectly potentiate B2R by binding to the active site of ACE, thus blocking bradykinin degradation (Deddish et al., 1998). Finally, the action of ATIR and B1R are functionally interrelated. ATIR can synergize BIR‐related effects by ROS induction in the endothelial cells (Ceravolo et al., 2014). The functions and relations between RAS and KKS are shown in Figure 1.
5. IMPLICATIONS OF RAS AND KKS IN COVID‐19
ACE2 inhibition affects both RAS and KKS as it decrease degradation of angiotensin II to angiotensin 1–7 and decreases degradation of Lys des‐Arg9‐bradykinin and des‐Arg9‐bradykinin, leading to increase in functionally interrelated angiotensin II (through effects on ATIR) as well as Lys des‐Arg9‐bradykinin and des‐Arg9‐bradykinin (through effects on B1R). ACE2 inhibition limits the functions of the functionally interrelated angiotensin 1–7 (through effects on MAS receptor and AT2R) and bradykinin (through effects on B2R). The effects of SARS‐COV‐2 on both RAS and KKS are shown in Figure 2.
FIGURE 2.
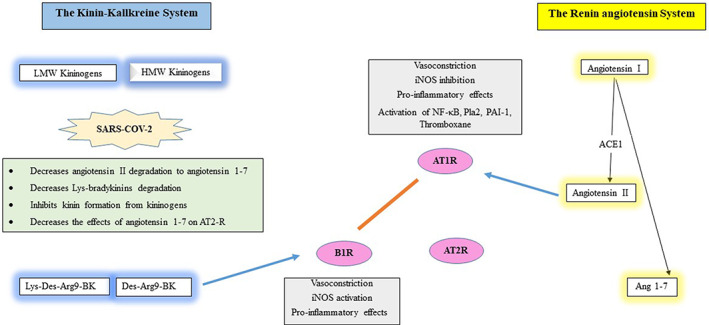
The pathophysiological effects of SARS‐COV‐2 on both the renin angiotensin and KKSs
It is important to consider the role of cathepsin L, a lysosomal cysteine endopeptidase that strongly enhances KKS involvement in COVID‐19 pathogenesis. Cathepsin L mediates SARS‐COV‐2 cell entry through the transmembrane protease serine‐2 (TMPRSS2; Hoffmann et al., 2020). Consistently, cathepsin L was found to be needed for viral cell entry with severe acute respiratory syndrome coronavirus (SARS; Huang et al., 2006; Simmons et al., 2005). Cathepsin L generates kinins from LMWK and HMWK at the infection site (Desmazes, Gauthier, & Lalmanach, 2001), thus acting as a kininogenase independently of kallikreins, which are unable to generate kinins under inflammatory conditions to restore kinin levels (Desmazes, Galineau, Gauthier, Brömme, & Lalmanach, 2003). Interestingly, cathepsin L exhibits hypotensive effects with increase in bradykinin release from kininogen in vivo, due to its kininogenase activity (Puzer et al., 2005). SARS‐COV‐2‐related effects on cathepsin L activity may modulate kinin and especially bradykinin levels, which may be at least partly responsible for some clinical aspects, supporting KKS role in COVID‐19 pathogenesis (Johnson et al., 2009). This is supported by previous studies reporting an increase in the risk of ARDS with increase in angiotensin II and decrease in angiotensin 1–7 (Kuba et al., 2005). Also, enhancing the effects of angiotensin 1–7 has been reported as likely to provide protection from ARDS induced by coronaviruses (Imai et al., 2005).
Moreover, suppressed eNOS and NO deficiency are associated in COVID‐19 patients with endothelial dysfunction leading to thrombotic events and organ dysfunctions (Green, 2020; Varga et al., 2020), thus supporting the hypotheses of deleterious suppressed B2R stimulation (which stimulate eNOS) and depressed NO synthesis.
All ACE2 inhibition effects were observed in COVID‐19 patients who exhibit increased risk of vasoconstriction (Ruocco, Feola, & Palazzuoli, 2020), proinflammatory cytokine profile (Jamilloux et al., 2020), acute phase reactants including C‐reactive protein (Liu et al., 2020), pulmonary fibrosis (Spagnolo et al., 2020), coagulopathy with elevated plasma D‐dimer, pulmonary thrombosis and venous thromboembolic events (Frater, Zini, D'Onofrio, & Rogers, 2020; Thachil, 2020), disseminated intravascular coagulation (Lillicrap, 2020) and ARDS (Gattinoni, Chiumello, & Rossi, 2020; Jamilloux et al., 2020).
6. CURRENTLY PROPOSED ANTI‐COVID‐19 THERAPIES AIMING TO MODULATE RAS AND KKS
Many drugs have been used and thousands of clinical trials started to investigate the benefits of anti‐COVID‐19 therapies (Esposito, Noviello, & Pagliano, 2020). Of these, drugs theoretically effective in modulating the RAS and KKS are currently being trialed to alleviate the consequences of ACE2 downregulation. Some drugs target ACE2 activity, while most target consequential effects of ACE2 inhibition mainly regarding the RAS. Surprisingly, none considered the strong functional relationships between RAS and KKS and the influence of ACE2 inhibition on both systems.
Some studies recommended targeting ACE2 itself to competitively antagonize SARS‐COV‐2 binding (e.g., using soluble receptor‐binding domains or antibodies). ACE2 blockers may be useful during the initial phase of infection or as prophylaxis to inhibit SARS‐COV‐2 entry (Kruse, 2020). Meanwhile, ACE2 blockage may exacerbate COVID‐19 severity by exaggerating the consequences of ACE2 inhibition; accordingly, even its usage as prophylaxis may lead to ACE2 inhibition manifestations. Interestingly, one pilot study recommended restoring ACE2 downregulation by using recombinant human ACE2 (rhACE2, GSK2586881) aiming at decreasing angiotensin II and increasing angiotensin 1–7, but no improvement in physiological or clinical parameters was observed (Peng et al., 2013).
Other studies recommended angiotensin receptor blockers to block angiotensin II overactivation effects (Gurwitz, 2020; Silhol et al., 2020), neglecting the effects of ACE2 inhibition on the KKS. Although classical ACE inhibitors do not inhibit ACE2 (Zisman, 2005), some studies recommended ACE inhibitor use to decrease angiotensin II overexpression. These studies depended on observations of clinical improvements in COVID‐19 patients on antihypertensive ACE inhibitors evident by the lower prevalence of severe presentations, reduced hospitalization risk, lower viral load and plasma IL‐6 and increased CD3 and CD8 T‐cell counts (Khera et al., 2020; Meng et al., 2020). A meta‐analysis suggested that ACE inhibitors may reduce mortality (Ghosal, Mukherjee, Sinha, & Gangopadhyay, 2020), consistent with the association with reduced mortality in septic patients (Hsu et al., 2020). By contrast, other studies discouraged its use as possibly facilitating SARS‐CoV‐2 entrance into the cell (Gurwitz, 2020; Hoffmann et al., 2020) and leading to bradykinin escape (Ruocco et al., 2020). Although ACE inhibitor therapy was associated with decreased Th1/Th2 cytokine ratios and inflammatory cytokine production, no inhibition of ACE2 was observed (Gage et al., 2004). Additionally, ACE inhibition may prevent bradykinin inhibition leading to the accumulation of Lys des‐Arg9‐bradykini and des‐Arg9‐bradykinin (ACE2‐related reduced degradation), resulting in B1R overstimulation and detrimental clinical effects.
Icatibant, a selective peptidomimetic B2R antagonist of 10 amino acids, was proposed as useful drug to alleviate the inflammatory symptoms by inhibiting B2R effects and consequently angiotensin II/ATIR actions (van de Veerdonk et al., 2020). This hypothesis was based on the functional relationships between ATIR and B2R, although ATIR/B2R heterodimer formation is controversial (AbdAlla, Lother, el Massiery, & Quitterer, 2001). Arachidonic acid can inactivate SARS‐CoV‐2 as its eicosanoids metabolites may help in resolution of inflammation and regulate the phagocytic action of macrophage and other immunocytes (Das, 2020). Current laboratory data obtained in COVID‐19 cases (increased eicosanoids) contradict the hypothetical basis for this therapy. Interestingly, antagonistic maneuvers directed against some consequences of angiotensin II overexpression have been recommended. Of these, corticosteroids have been evaluated as a potent treatment to inhibit cytokine production by NF‐κB‐mediated transcription factor inhibition (Qin et al., 2020; Russell, Millar, & Baillie, 2020). Anticoagulants such as heparin have been suggested to reduce mortality (Thachil, 2020). Although these studies did not cover all SARS‐COV‐2‐induced pathological effects, they may be beneficial against specific consequences such as thrombosis and thromboembolic events, thus ameliorating COVID‐19 severity.
Inhaled NO with its potential vasodilatory and bronchodilatory properties is also currently being investigated aiming to decrease pulmonary hypertension, improve arterial oxygenation and reduce spread and density of lung infiltrates in COVID‐19 patients (Chen et al., 2004). Interestingly, NO has been shown able to inactivate viral replication in vitro by modifying proteins and nucleic acids (Croen, 1993) with many viruses (Klingström et al., 2006; Saura et al., 1999) including SARS coronavirus (Åkerström, Gunalan, Keng, Tan, & Mirazimi, 2009). Theoretically, this therapy is appealing as deficient NO bioavailability is associated with endothelial dysfunction (Tomasian, 2000). Deficient NO bioavailability may result from reduced NO production by eNOS which occurs from decreased B2R and AT2R stimulation and/or ROS‐related increased NO breakdown which increases with ATIR and B1R stimulation (Taddei, Ghiadoni, Virdis, Versari, & Salvetti, 2003).
Teicoplanin and glycopeptides have also been proposed to treat COVID‐19 patients by targeting cathepsin L in the late endosomes and thus blocking the virus replication cycle (Zhang et al., 2020). Targeting cathepsin L may be useful as a prophylactic strategy to prevent SARS‐CoV‐2 entry into the cells and/or limit the subsequent COVID‐19 severity before host defense impairment as observed with influenza A (Xu, Greenland, Gotts, Matthay, & Caughey, 2016). However, this targeting strategy may also aggravate COVID‐19 severity as cathepsin L plays a crucial pathogenic role by modulating bradykinin levels.
Interestingly, a new potential therapeutic approach to combat COVID‐19 by using Rho kinase inhibitors such as fasudil has been suggested (Abedi, Rezaee, & Karimi, 2020), as it showed a protective effect from lung injury through its ability to counteract the proinflammatory and immune responses of Rho kinase that mediates lung injury. Rho kinase inhibitors also shown to have antifibrotic effects and ability to upregulate ACE2 (Abedi, Hayes, Reiter, & Karimi, 2020; Zhang, Penninger, Li, Zhong, & Slutsky, 2020). This is consistent with other studies that showed that Rho pathway is activated by angiotensin II/AT1R (Kimura & Eguchi, 2009; Suzuki et al., 2009).
7. SNAKE VENOM‐DERIVED PEPTIDES
Snake venoms contain a mixture of amino acids, proteins, peptides, nucleotides, lipids, carbohydrates and metallic elements coupled to proteins (Calvete, Juárez, & Sanz, 2007; Georgieva, Arni, & Betzel, 2008). Although these components may be lethal, they constitute a rich biological resource of significant therapeutic value including promising antiviral drugs (Kang et al., 2011; Meenakshisundaram, Sweni, & Thirumalaikolundusubramanian, 2009; Pennington, Czerwinski, & Norton, 2018; Rivero et al., 2011). Several studies reported that snake venom components have antiviral activity against measles, Sendai, dengue, yellow fever and human immunodeficiency viruses (Borkow & Ovadia, 1999; Meenakshisundaram et al., 2009; Muller et al., 2012). Snake venom properties may constitute promising therapeutic options against defense mechanisms developed by viruses (Meenakshisundaram et al., 2009).
Interestingly, the venom‐derived peptides present properties adequate to counteract COVID‐19 consequences in the light of SARS‐CoV‐2‐related pathophysiological mechanisms. BPPs, isolated from Bothrops jararaca venom (Ferreira, Greene, Alabaster, Bakhle, & Vane, 1970), launched the discovery of bradykinin in the bitten patients (e Silva, Beraldo, & Rosenfeld, 1949), allowing understanding of the physiological roles of the KKS (Linz, Wiemer, Gohlke, Unger, & Schölkens, 1995). Peptide fraction analysis of Bothrops jararaca venoms contains various BPPs (9a, 10b, 10c, 11a, 11d, 11e, 12b, 12c, 13a, 13b, 14a), short proline‐rich peptides with remarkable functional differences (Camargo, Ianzer, Guerreiro, & Serrano, 2012; Morais et al., 2011). The first BBP to be sequenced was Pyr‐Lys‐Trp‐Ala‐Pro‐OH (Munawar et al., 2016).
BPP‐10c (Glu‐Asn‐Trp‐Pro‐His‐Pro‐Gln‐Ile‐Pro‐Pro) strongly decreases angiotensin II by inhibiting ACE, increasing bradykinin‐related effects on B2R, increasing NO‐attributed antioxidant, antiinflammatory and neuroprotective effects and exhibiting direct neural antihypertensive effects. Therefore, we hypothesized that BPP‐10c may be an excellent anti‐COVID‐19 treatment due to its ability to counteract most of the deleterious effects of SARS‐COV‐2 on both RAS and KKS.
BPPs increase bradykinin‐induced hypotension and decrease angiotensin I‐related vasopressor effects by inhibiting ACE (Camargo et al., 2012; Lopes et al., 2014). They represent the first natural bradykinin agonists and ACE inhibitors (Camargo et al., 2012). BPPs augment bradykinin‐related effects by interacting directly on bradykinin receptors rather than inhibiting bradykinin degradation by ACE1 inhibition (Chi et al., 1985). BPP‐10c strongly potentiates bradykinin‐related effects on B2R and is additionally a strong selective ACE C‐domain inhibitor (400‐fold more selective than for the N‐domain; Camargo et al., 2012; Cotton et al., 2002). Angiotensin I is predominantly hydrolyzed by the C‐domain, whereas bradykinin is hydrolyzed by both active domains (Junot et al., 2001). Hence, a purely C‐domain selective inhibitor would be more beneficial as it mainly decreases angiotensin II by inhibiting its synthesis from angiotensin I by the C‐domain. BPPs only decrease bradykinin degradation while preventing its accumulation by preserving ACE N‐domain activity (Messerli & Nussberger, 2000). This property renders BPPs superior to classical ACE inhibitors that have the risk of developing bradykinin‐mediated angioedema. Besides its ability to inhibit ACE and directly activate bradykinin‐B2R, BPP‐10c exerts its antihypertensive effect by increasing free intracellular calcium in neuronal cells and releasing specific neurotransmitters in the central nervous system (Lameu et al., 2010; Querobino, Ribeiro, & Alberto‐Silva, 2018).
Additionally, BPP‐10c is reported to enhance argininosuccinate synthetase (AsS) activity leading to sustained increase in NO production (Camargo et al., 2012; Morais et al., 2011, 2013). BPP‐10c binding to AsS enhances adenosine triphosphate and citrulline (Guerreiro et al., 2009) leading to NO release from endothelial cells and vasodilatation (Morais et al., 2013). AsS enhances argininosuccinate synthesis via conjugation of aspartate with citrulline. Argininosuccinate is cleaved by argininosuccinate lyase resulting in fumarate and L‐arginine formation (Haines, Pendleton, & Eichler, 2011). This amino acid participates in the synthesis of neuroprotective molecules including agmatine and various polyamines such as spermine, spermidine and putrescine (Blantz, Satriano, Gabbai, & Kelly, 2000; Querobino et al., 2018). Polyamines could prevent alterations in mitochondrial membrane permeability, regulating calcium concentrations and NOS activity (Jamwal & Kumar, 2016). Agmatine is reported to exhibit antiinflammatory properties by inhibiting NF‐κB leading to iNOS suppression (Ahn et al., 2012), inhibiting TNF‐α (Hong, Kim, Lee, & Seong, 2009) and inducing neuroprotective and antioxidant actions (Freitas et al., 2016). L‐arginine can also be metabolized to NO (Maes, Galecki, Chang, & Berk, 2011). The importance of the arginine–citrulline cycle for endothelial NO production was supported by a report of two infants with a deficiency of argininosuccinate lyase, who were shown to be hypertensive (Fakler, Kaftan, & Nelin, 1995).
BPP‐10c reduces ROS production (Querobino et al., 2018; Zhou, Ai, Chen, & Li, 2019), increases NO synthesis (de Oliveira et al., 2010), reduces NF‐κΒ expression and reduces iNOS expression (Querobino et al., 2018). BPP‐10c has been reported to be safe and without cytotoxic effects (Querobino et al., 2018). It caused sustained reduction in blood pressure in hypertensive but not normotensive rats (Guerreiro et al., 2009). Other studies recommended its consideration as a potential therapeutic agent for various diseases related to NO deficiency (Morais et al., 2011).
8. CONCLUSION
SARS‐COV‐2 downregulates ACE2 and affects cathepsin L that significantly contributes to COVID‐19 pathophysiology by increasing the proinflammatory and organodestructive effects of angiotensin II and Lys‐bradykinins and decreasing the antiinflammatory and organoprotective effects of angiotensin 1–7, NO and bradykinin. Most investigations on anti‐COVID‐19 therapies did not consider the effects on both RAS and KKS. Snake venom‐derived BPP‐10c exhibits remarkable organoprotective effects targeting both systems and thus providing great value as a natural treatment option to alleviate COVID‐19 manifestations. Further trials are now required to evaluate any clinical benefits.
CONFLICT OF INTEREST
The authors declare no potential conflict of interest.
AUTHOR CONTRIBUTIONS
Ahmed S. Gouda: Writing – original draft (lead); writing – approval. Bruno Mégarbane: Writing – original draft (lead); writing – approval.
Gouda AS, Mégarbane B. Snake venom‐derived bradykinin‐potentiating peptides: A promising therapy for COVID‐19? Drug Dev Res. 2021;82:38–48. 10.1002/ddr.21732
Contributor Information
Ahmed S. Gouda, Email: salah1001@hotmail.com.
Bruno Mégarbane, Email: bruno.megarbane@lrb.aphp.fr.
REFERENCES
- AbdAlla, S. , Lother, H. , el Massiery, A. , & Quitterer, U. (2001). Increased AT1 receptor heterodimers in preeclampsia mediate enhanced angiotensin II responsiveness. Nature Medicine, 7(9), 1003–1009. 10.1038/nm0901-1003 [DOI] [PubMed] [Google Scholar]
- Abedi, F. , Hayes, A. W. , Reiter, R. , & Karimi, G. (2020). Acute lung injury: The therapeutic role of Rho kinase inhibitors. Pharmacological Research, 155, 104736. 10.1016/j.phrs.2020.104736 [DOI] [PubMed] [Google Scholar]
- Abedi, F. , Rezaee, R. , & Karimi, G. (2020). Plausibility of therapeutic effects of Rho kinase inhibitors against severe acute respiratory syndrome coronavirus 2 (COVID‐19). Pharmacological Research, 156, 104808. 10.1016/j.phrs.2020.104808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahn, S. K. , Hong, S. , Park, Y. M. , Choi, J. Y. , Lee, W. T. , Park, K. A. , & Lee, J. E. (2012). Protective effects of agmatine on lipopolysaccharide‐injured microglia and inducible nitric oxide synthase activity. Life Sciences, 91(25–26), 1345–1350. 10.1016/j.lfs.2012.10.010 [DOI] [PubMed] [Google Scholar]
- Ahsan, W. , Javed, S. , Al Bratty, M. , Alhazmi, H. A. , & Najmi, A. (2020). Treatment of SARS‐CoV‐2: How far have we reached? Drug Discoveries & Therapeutics, 14(2), 67–72. 10.5582/ddt.2020.03008 [DOI] [PubMed] [Google Scholar]
- Åkerström, S. , Gunalan, V. , Keng, C. T. , Tan, Y.‐J. , & Mirazimi, A. (2009). Dual effect of nitric oxide on SARS‐CoV replication: Viral RNA production and palmitoylation of the S protein are affected. Virology, 395(1), 1–9. 10.1016/j.virol.2009.09.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alhenc‐Gelas, F. , Bouby, N. , Richer, C. , Potier, L. , Roussel, R. , & Marre, M. (2011). Kinins as therapeutic agents in cardiovascular and renal diseases. Current Pharmaceutical Design, 17(25), 2654–2662. 10.2174/138161211797416002 [DOI] [PubMed] [Google Scholar]
- Ancion, A. , Tridetti, J. , Nguyen Trung, M.‐L. , Oury, C. , & Lancellotti, P. (2019). A review of the role of bradykinin and nitric oxide in the cardioprotective action of angiotensin‐converting enzyme inhibitors: Focus on perindopril. Cardiology and Therapy, 8(2), 179–191. 10.1007/s40119-019-00150-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baudin, B. , Berard, M. , Carrier, J. L. , Legrand, Y. , & Drouet, L. (1997). Vascular origin determines angiotensin I‐converting enzyme expression in endothelial cells. Endothelium, 5(1), 73–84. 10.3109/10623329709044160 [DOI] [PubMed] [Google Scholar]
- Baynes, J. W. , & Dominiczak, M. H. (2005). Medical biochemistry (2nd ed.). pp. 555. Philadelphia: Elsevier, ISBN 0‐7234‐3341‐0. [Google Scholar]
- Bentley, J. P. , Asselbergs, F. W. , Coffey, C. S. , Hebert, P. R. , Moore, J. H. , Hillege, H. L. , & van Gilst, W. H. (2010). Cardiovascular risk associated with interactions among polymorphisms in genes from the renin–angiotensin, bradykinin, and fibrinolytic systems. PLoS One, 5(9), e12757. 10.1371/journal.pone.0012757 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blantz, R. C. , Satriano, J. , Gabbai, F. , & Kelly, C. (2000). Biological effects of arginine metabolites. Acta Physiologica Scandinavica, 168(1), 21–25. 10.1046/j.1365-201x.2000.00646.x [DOI] [PubMed] [Google Scholar]
- Borkow, G. , & Ovadia, M. (1999). Selective lysis of virus‐infected cells by cobra snake cytotoxins: A sendai virus, human erythrocytes, and cytotoxin model. Biochemical and Biophysical Research Communications, 264(1), 63–68. 10.1006/bbrc.1999.1483 [DOI] [PubMed] [Google Scholar]
- Brown, N. J. , Nadeau, J. H. , & Vaughan, D. E. (1997). Selective stimulation of tissue‐type plasminogen activator (t‐PA) in vivo by infusion of bradykinin. Thrombosis and Haemostasis, 77(3), 522–525. 10.1055/s-0038-1656000 [DOI] [PubMed] [Google Scholar]
- Calò, L. A. , Rigato, M. , & Bertoldi, G. (2019). ACE2/angiotensin 1–7 protective anti‐inflammatory and antioxidant role in hyperoxic lung injury: Support from studies in Bartter's and Gitelman's syndromes. QJM: An International Journal of Medicine., 113, 440–441. 10.1093/qjmed/hcz319 [DOI] [PubMed] [Google Scholar]
- Calvete, J. J. , Juárez, P. , & Sanz, L. (2007). Snake venomics. Strategy and applications. Journal of Mass Spectrometry, 42(11), 1405–1414. 10.1002/jms.1242 [DOI] [PubMed] [Google Scholar]
- Camargo, A. C. M. , Ianzer, D. , Guerreiro, J. R. , & Serrano, S. M. T. (2012). Bradykinin‐potentiating peptides: Beyond captopril. Toxicon, 59(4), 516–523. 10.1016/j.toxicon.2011.07.013 [DOI] [PubMed] [Google Scholar]
- Ceravolo, G. S. , Montezano, A. C. , Jordão, M. T. , Akamine, E. H. , Costa, T. J. , Takano, A. P. , … Carvalho, M. H. C. (2014). An interaction of renin–angiotensin and kallikrein–kinin systems contributes to vascular hypertrophy in angiotensin II‐induced hypertension: in vivo and in vitro studies. PLoS One, 9(11), e111117. 10.1371/journal.pone.0111117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chappell, M. C. (2016). Biochemical evaluation of the renin–angiotensin system: The good, bad, and absolute? American Journal of Physiology‐Heart and Circulatory Physiology, 310(2), H137–H152. 10.1152/ajpheart.00618.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chappell, M. C. , & Al Zayadneh, E. M. (2017). Angiotensin‐(1–7) and the regulation of anti‐fibrotic signaling pathways. Journal of Cellular Signaling, 2(1), 134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, L. , Liu, P. , Gao, H. , Sun, B. , Chao, D. , Wang, F. , … Wang, C. G. (2004). Inhalation of nitric oxide in the treatment of severe acute respiratory syndrome: A rescue trial in Beijing. Clinical Infectious Diseases, 39(10), 1531–1535. 10.1086/425357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi, C. W. , Wang, S. Z. , Xu, L. G. , Wang, M. Y. , Lo, S. S. , & Huang, W. D. (1985). Structure‐function studies on the bradykinin potentiating peptide from Chinese snake venom (Agkistrodon Halys Pallas). Peptides, 6(Suppl. 3), 339–342. [PubMed] [Google Scholar]
- Cotton, J. , Hayashi, M. A. F. , Cuniasse, P. , Vazeux, G. , Ianzer, D. , De Camargo, A. C. M. , & Dive, V. (2002). Selective inhibition of the C‐domain of angiotensin I converting enzyme by bradykinin potentiating peptides. Biochemistry, 41(19), 6065–6071. 10.1021/bi012121x [DOI] [PubMed] [Google Scholar]
- Croen, K. D. (1993). Evidence for antiviral effect of nitric oxide. Inhibition of herpes simplex virus type 1 replication. Journal of Clinical Investigation, 91(6), 2446–2452. 10.1172/JCI116479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das, U. N. (2020). Can bioactive lipids inactivate coronavirus (COVID‐19)? Archives of Medical Research, 51(3), 282–286. 10.1016/j.arcmed.2020.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Oliveira, E. F. , Guerreiro, J. R. , Silva, C. A. , Benedetti, G. F. d. S. , Lebrun, I. , Ulrich, H. , … Camargo, A. C. M. (2010). Enhancement of the citrulline–nitric oxide cycle in astroglioma cells by the proline‐rich peptide‐10c from Bothrops jararaca venom. Brain Research, 1363, 11–19. 10.1016/j.brainres.2010.09.067 [DOI] [PubMed] [Google Scholar]
- Deddish, P. A. , Marcic, B. , Jackman, H. L. , Wang, H.‐Z. , Skidgel, R. A. , & Erdös, E. G. (1998). N‐domain–specific substrate and C‐domain inhibitors of angiotensin‐converting enzyme. Hypertension, 31(4), 912–917. 10.1161/01.HYP.31.4.912 [DOI] [PubMed] [Google Scholar]
- Desmazes, C. , Galineau, L. , Gauthier, F. , Brömme, D. , & Lalmanach, G. (2003). Kininogen‐derived peptides for investigating the putative vasoactive properties of human cathepsins K and L. European Journal of Biochemistry, 270(1), 171–178. 10.1046/j.1432-1033.2003.03382.x [DOI] [PubMed] [Google Scholar]
- Desmazes, C. , Gauthier, F. , & Lalmanach, G. (2001). Cathepsin L, but not cathepsin B, is a potential kininogenase. Biological Chemistry, 382(5), 811–815. 10.1515/BC.2001.098 [DOI] [PubMed] [Google Scholar]
- e Silva, M. R. , Beraldo, W. T. , & Rosenfeld, G. (1949). Bradykinin, a hypotensive and smooth muscle stimulating factor released from plasma globulin by snake venoms and by trypsin. American Journal of Physiology‐Legacy Content, 156(2), 261–273. 10.1152/ajplegacy.1949.156.2.261 [DOI] [PubMed] [Google Scholar]
- Erdös, E. G. , Tan, F. , & Skidgel, R. A. (2010). Angiotensin I–converting enzyme inhibitors are allosteric enhancers of Kinin B1 and B2 receptor function. Hypertension, 55(2), 214–220. 10.1161/HYPERTENSIONAHA.109.144600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esposito, S. , Noviello, S. , & Pagliano, P. (2020). Update on treatment of COVID‐19: Ongoing studies between promising and disappointing results. Le Infezioni in Medicina, 28(2), 198–211. [PubMed] [Google Scholar]
- Fakler, C. , Kaftan, H. , & Nelin, L. (1995). Two cases suggesting a role for the L‐arginine nitric oxide pathway in neonatal blood pressure regulation. Acta Paediatrica, 84(4), 460–462. 10.1111/j.1651-2227.1995.tb13673.x [DOI] [PubMed] [Google Scholar]
- Fernandes, L. , Fortes, Z. B. , Nigro, D. , Tostes, R. C. A. , Santos, R. A. S. , & Catelli de Carvalho, M. H. (2001). Potentiation of bradykinin by angiotensin‐(1–7) on arterioles of spontaneously hypertensive rats studied in vivo. Hypertension, 37(2), 703–709. 10.1161/01.HYP.37.2.703 [DOI] [PubMed] [Google Scholar]
- Ferreira, S. H. , Greene, L. J. , Alabaster, V. A. , Bakhle, Y. S. , & Vane, J. R. (1970). Activity of various fractions of bradykinin potentiating factor against angiotensin I converting enzyme. Nature, 225(5230), 379–380. 10.1038/225379a0 [DOI] [PubMed] [Google Scholar]
- Frater, J. L. , Zini, G. , D'Onofrio, G. , & Rogers, H. J. (2020). COVID‐19 and the clinical hematology laboratory. International Journal of Laboratory Hematology, 42, 11–18. 10.1111/ijlh.13229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freitas, A. E. , Egea, J. , Buendia, I. , Gómez‐Rangel, V. , Parada, E. , Navarro, E. , … Lopez, M. G. (2016). Agmatine, by improving neuroplasticity markers and inducing Nrf2, prevents corticosterone‐induced depressive‐like behavior in mice. Molecular Neurobiology, 53(5), 3030–3045. 10.1007/s12035-015-9182-6 [DOI] [PubMed] [Google Scholar]
- Gage, J. R. , Fonarow, G. , Hamilton, M. , Widawski, M. , Martínez‐Maza, O. , & Vredevoe, D. L. (2004). Beta blocker and angiotensin‐converting enzyme inhibitor therapy is associated with decreased Th1/Th2 cytokine ratios and inflammatory cytokine production in patients with chronic heart failure. Neuroimmunomodulation, 11(3), 173–180. 10.1159/000076766 [DOI] [PubMed] [Google Scholar]
- Gattinoni, L. , Chiumello, D. , & Rossi, S. (2020). COVID‐19 pneumonia: ARDS or not? Critical Care, 24(1), 154. 10.1186/s13054-020-02880-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Georgieva, D. , Arni, R. K. , & Betzel, C. (2008). Proteome analysis of snake venom toxins: Pharmacological insights. Expert Review of Proteomics, 5(6), 787–797. 10.1586/14789450.5.6.787 [DOI] [PubMed] [Google Scholar]
- Ghosal, S. , Mukherjee, J. J. , Sinha, B. , & Gangopadhyay, K. K. (2020). The effect of angiotensin converting enzyme inhibitors and angiotensin receptor blockers on death and severity of disease in patients with coronavirus disease 2019 (COVID‐19): A meta‐analysis. MedRxiv Preprint. 10.1101/2020.04.23.20076661 [DOI]
- Gohlke, P. , Pees, C. , & Unger, T. (1998). AT 2 receptor stimulation increases aortic cyclic GMP in SHRSP by a kinin‐dependent mechanism. Hypertension, 31(1), 349–355. 10.1161/01.HYP.31.1.349 [DOI] [PubMed] [Google Scholar]
- Gralinski, L. E. , Sheahan, T. P. , Morrison, T. E. , Menachery, V. D. , Jensen, K. , Leist, S. R. , … Baric, R. S. (2018). Complement activation contributes to severe acute respiratory syndrome coronavirus pathogenesis. MBio, 9(5), e01753–18. 10.1128/mBio.01753-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greco, A. J. , Master, R. G., Jr. , F, A. , Baber, S. R. , & Kadowitz, P. J. (2006). Angiotensin‐(1–7) potentiates responses to bradykinin but does not change responses to angiotensin I. Canadian Journal of Physiology and Pharmacology, 84(11), 1163–1175. 10.1139/y06-053 [DOI] [PubMed] [Google Scholar]
- Green, S. J. (2020). Covid‐19 accelerates endothelial dysfunction and nitric oxide deficiency. Microbes and Infection, 22(4–5), 149–150. 10.1016/j.micinf.2020.05.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griendling, K. K. , Lassègue, B. , Murphy, T. J. , & Wayne Alexander, R. (1994). Angiotensin II receptor. Advances in Pharmacology, 28, 269–306. 10.1016/S1054-3589(08)60498-6 [DOI] [PubMed] [Google Scholar]
- Guerreiro, J. R. , Lameu, C. , Oliveira, E. F. , Klitzke, C. F. , Melo, R. L. , Linares, E. , … Camargo, A. C. M. (2009). Argininosuccinate synthetase is a functional target for a snake venom anti‐hypertensive peptide. Journal of Biological Chemistry, 284(30), 20022–20033. 10.1074/jbc.M109.021089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gurusamy, M. , Nasseri, S. , Lee, H. , Jung, B. , Lee, D. , Khang, G. , … Wu, D. (2016). Kinin B1 receptor antagonist BI113823 reduces allergen‐induced airway inflammation and mucus secretion in mice. Pharmacological Research, 104, 132–139. 10.1016/j.phrs.2015.12.017 [DOI] [PubMed] [Google Scholar]
- Gurwitz, D. (2020). Angiotensin receptor blockers as tentative SARS‐CoV‐2 therapeutics. Drug Development Research, 2020, 1–4. 10.1002/ddr.21656 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Habibzadeh, F. , & Lang, T. (2020). The coronavirus pandemic: “The show must not go on”. International Journal of Occupational and Environmental Medicine, 11(2), 63–64. 10.34172/ijoem.2020.1979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haines, R. J. , Pendleton, L. C. , & Eichler, D. C. (2011). Argininosuccinate synthase: At the center of arginine metabolism. International Journal of Biochemistry and Molecular Biology, 2(1), 8–23. [PMC free article] [PubMed] [Google Scholar]
- Hällgren, R. , Samuelsson, T. , Laurent, T. C. , & Modig, J. (1989). Accumulation of hyaluronan (hyaluronic acid) in the lung in adult respiratory distress syndrome. American Review of Respiratory Disease, 139(3), 682–687. 10.1164/ajrccm/139.3.682 [DOI] [PubMed] [Google Scholar]
- Hamming, I. , Timens, W. , Bulthuis, M. , Lely, A. , Navis, G. , & van Goor, H. (2004). Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. Journal of Pathology, 203(2), 631–637. 10.1002/path.1570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han, C. , Liu, J. , Liu, X. , & Li, M. (2010). Angiotensin II induces C‐reactive protein expression through ERK1/2 and JNK signaling in human aortic endothelial cells. Atherosclerosis, 212(1), 206–212. 10.1016/j.atherosclerosis.2010.05.020 [DOI] [PubMed] [Google Scholar]
- Harizi, H. , Corcuff, J.‐B. , & Gualde, N. (2008). Arachidonic‐acid‐derived eicosanoids: Roles in biology and immunopathology. Trends in Molecular Medicine, 14(10), 461–469. 10.1016/j.molmed.2008.08.005 [DOI] [PubMed] [Google Scholar]
- Heitsch, H. (2002). Non‐peptide antagonists and agonists of the bradykinin B2 receptor. Current Medicinal Chemistry, 9(9), 913–928. 10.2174/0929867024606722 [DOI] [PubMed] [Google Scholar]
- Hoffmann, M. , Kleine‐Weber, H. , Schroeder, S. , Krüger, N. , Herrler, T. , Erichsen, S. , … Pöhlmann, S. (2020). SARS‐CoV‐2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell, 181(2), 271–280.e8. 10.1016/j.cell.2020.02.052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong, S. , Kim, C. Y. , Lee, J. E. , & Seong, G. J. (2009). Agmatine protects cultured retinal ganglion cells from tumor necrosis factor‐alpha‐induced apoptosis. Life Sciences, 84(1–2), 28–32. 10.1016/j.lfs.2008.10.006 [DOI] [PubMed] [Google Scholar]
- Hsu, W.‐T. , Galm, B. P. , Schrank, G. , Hsu, T.‐C. , Lee, S.‐H. , Park, J. Y. , & Lee, C.‐C. (2020). Effect of renin–angiotensin–aldosterone system inhibitors on short‐term mortality after sepsis. Hypertension, 75(2), 483–491. 10.1161/HYPERTENSIONAHA.119.13197 [DOI] [PubMed] [Google Scholar]
- Huang, I.‐C. , Bosch, B. J. , Li, F. , Li, W. , Lee, K. H. , Ghiran, S. , … Choe, H. (2006). SARS coronavirus, but not human coronavirus NL63, utilizes Cathepsin L to infect ACE2‐expressing cells. Journal of Biological Chemistry, 281(6), 3198–3203. 10.1074/jbc.M508381200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai, Y. , Kuba, K. , Rao, S. , Huan, Y. , Guo, F. , Guan, B. , … Penninger, J. M. (2005). Angiotensin‐converting enzyme 2 protects from severe acute lung failure. Nature, 436(7047), 112–116. 10.1038/nature03712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamilloux, Y. , Henry, T. , Belot, A. , Viel, S. , Fauter, M. , El Jammal, T. , … Sève, P. (2020). Should we stimulate or suppress immune responses in COVID‐19? Cytokine and anti‐cytokine interventions. Autoimmunity Reviews, 19, 102567. 10.1016/j.autrev.2020.102567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamwal, S. , & Kumar, P. (2016). Spermidine ameliorates 3‐nitropropionic acid (3‐NP)‐induced striatal toxicity: Possible role of oxidative stress, neuroinflammation, and neurotransmitters. Physiology & Behavior, 155, 180–187. 10.1016/j.physbeh.2015.12.015 [DOI] [PubMed] [Google Scholar]
- Ji, W. , Wang, W. , Zhao, X. , Zai, J. , & Li, X. (2020). Cross‐species transmission of the newly identified coronavirus 2019‐nCoV. Journal of Medical Virology, 92(4), 433–440. 10.1002/jmv.25682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson, E. M. , Doyle, J. D. , Wetzel, J. D. , McClung, R. P. , Katunuma, N. , Chappell, J. D. , … Dermody, T. S. (2009). Genetic and pharmacologic alteration of Cathepsin expression influences reovirus pathogenesis. Journal of Virology, 83(19), 9630–9640. 10.1128/JVI.01095-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Junot, C. , Gonzales, M. F. , Ezan, E. , Cotton, J. , Vazeux, G. , Michaud, A. , … Dive, V. (2001). RXP 407, a selective inhibitor of the N‐domain of angiotensin I‐converting enzyme, blocks in vivo the degradation of hemoregulatory peptide acetyl‐Ser‐Asp‐Lys‐Pro with no effect on angiotensin I hydrolysis. Journal of Pharmacology and Experimental Therapeutics, 297(2), 606–611. [PubMed] [Google Scholar]
- Jurado‐Palomo, J. , & Caballero, T. (2017). Pathophysiology of bradykinin‐mediated angioedema: The role of the complement system. In A comprehensive review of urticaria and angioedema. London, UK: InTech. 10.5772/67704 [DOI] [Google Scholar]
- Kang, T. S. , Georgieva, D. , Genov, N. , Murakami, M. T. , Sinha, M. , Kumar, R. P. , … Kini, R. M. (2011). Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. FEBS Journal, 278(23), 4544–4576. 10.1111/j.1742-4658.2011.08115.x [DOI] [PubMed] [Google Scholar]
- Kaschina, E. , & Unger, T. (2003). Angiotensin AT1/AT2 receptors: Regulation, signalling and function. Blood Pressure, 12(2), 70–88. 10.1080/08037050310001057 [DOI] [PubMed] [Google Scholar]
- Katovich, M. J. , Grobe, J. L. , & Raizada, M. K. (2008). Angiotensin‐(1–7) as an antihypertensive, antifibrotic target. Current Hypertension Reports, 10(3), 227–232. 10.1007/s11906-008-0043-9 [DOI] [PubMed] [Google Scholar]
- Khera, R. , Clark, C. , Lu, Y. , Guo, Y. , Ren, S. , Truax, B. , Spatz, E. S. , Murugiah, K. , Lin, Z. , Omer, S. B. , Vojta, D. , & Krumholz, H. M. (2020). Association of angiotensin‐converting enzyme inhibitors and angiotensin receptor blockers with the risk of hospitalization and death in hypertensive patients with coronavirus disease‐19. MedRxiv Preprint. 10.1101/2020.05.17.20104943 [DOI] [PMC free article] [PubMed]
- Kimura, K. , & Eguchi, S. (2009). Angiotensin II type‐1 receptor regulates RhoA and Rho‐kinase/ROCK activation via multiple mechanisms. Focus on “Angiotensin II induces RhoA activation through SHP2‐dependent dephosphorylation of the RhoGAP p190A in vascular smooth muscle cells”. American Journal of Physiology‐Cell Physiology, 297(5), C1059–C1061. 10.1152/ajpcell.00399.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klingström, J. , Åkerström, S. , Hardestam, J. , Stoltz, M. , Simon, M. , Falk, K. I. , … Lundkvist, Å. (2006). Nitric oxide and peroxynitrite have different antiviral effects against hantavirus replication and free mature virions. European Journal of Immunology, 36(10), 2649–2657. 10.1002/eji.200535587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruse, R. L. (2020). Therapeutic strategies in an outbreak scenario to treat the novel coronavirus originating in Wuhan, China. F1000Research, 9, 72. 10.12688/f1000research.22211.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuba, K. , Imai, Y. , Rao, S. , Gao, H. , Guo, F. , Guan, B. , … Penninger, J. M. (2005). A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus–induced lung injury. Nature Medicine, 11(8), 875–879. 10.1038/nm1267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhr, F. , Lowry, J. , Zhang, Y. , Brovkovych, V. , & Skidgel, R. A. (2010). Differential regulation of inducible and endothelial nitric oxide synthase by kinin B1 and B2 receptors. Neuropeptides, 44(2), 145–154. 10.1016/j.npep.2009.12.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lameu, C. , Hayashi, M. A. F. , Guerreiro, J. R. , Oliveira, E. F. , Lebrun, I. , Pontieri, V. , … Ulrich, H. (2010). The central nervous system as target for antihypertensive actions of a proline‐rich peptide from Bothrops jararaca venom. Cytometry Part A, 77, 220–230. 10.1002/cyto.a.20860 [DOI] [PubMed] [Google Scholar]
- Lee, E. B. , Fleischmann, R. , Hall, S. , Wilkinson, B. , Bradley, J. D. , Gruben, D. , … van Vollenhoven, R. F. (2014). Tofacitinib versus methotrexate in rheumatoid arthritis. New England Journal of Medicine, 370(25), 2377–2386. 10.1056/NEJMoa1310476 [DOI] [PubMed] [Google Scholar]
- Leonhardt, J. , Villela, D. C. , Teichmann, A. , Münter, L.‐M. , Mayer, M. C. , Mardahl, M. , … Steckelings, U. M. (2017). Evidence for heterodimerization and functional interaction of the angiotensin type 2 receptor and the receptor MAS. Hypertension, 69(6), 1128–1135. 10.1161/HYPERTENSIONAHA.116.08814 [DOI] [PubMed] [Google Scholar]
- Li, P. , Chappell, M. C. , Ferrario, C. M. , & Brosnihan, K. B. (1997). Angiotensin‐(1–7) augments bradykinin‐induced vasodilation by competing with ACE and releasing nitric oxide. Hypertension, 29, 394–398. 10.1161/01.HYP.29.1.394 [DOI] [PubMed] [Google Scholar]
- Lillicrap, D. (2020). Disseminated intravascular coagulation in patients with 2019‐nCoV pneumonia. Journal of Thrombosis and Haemostasis, 18(4), 786–787. 10.1111/jth.14781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linz, W. , Wiemer, G. , Gohlke, P. , Unger, T. , & Schölkens, B. A. (1995). Contribution of kinins to the cardiovascular actions of angiotensin‐converting enzyme inhibitors. Pharmacological Reviews, 47(1), 25–49. [PubMed] [Google Scholar]
- Liu, Y. , Yang, Y. , Zhang, C. , Huang, F. , Wang, F. , Yuan, J. , … Liu, L. (2020). Clinical and biochemical indexes from 2019‐nCoV infected patients linked to viral loads and lung injury. Science China Life Sciences, 63(3), 364–374. 10.1007/s11427-020-1643-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes, D. M. , Junior, N. E. G. , Costa, P. P. C. , Martins, P. L. , Santos, C. F. , Carvalho, E. D. F. , … Carvalho, K. M. (2014). A new structurally atypical bradykinin‐potentiating peptide isolated from Crotalus durissus cascavella venom (South American rattlesnake). Toxicon, 90, 36–44. 10.1016/j.toxicon.2014.07.011 [DOI] [PubMed] [Google Scholar]
- Lu, R. , Zhao, X. , Li, J. , Niu, P. , Yang, B. , Wu, H. , … Tan, W. (2020). Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet, 395(10224), 565–574. 10.1016/S0140-6736(20)30251-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maes, M. , Galecki, P. , Chang, Y. S. , & Berk, M. (2011). A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Progress in Neuro‐Psychopharmacology and Biological Psychiatry, 35(3), 676–692. 10.1016/j.pnpbp.2010.05.004 [DOI] [PubMed] [Google Scholar]
- Meenakshisundaram, R. , Sweni, S. , & Thirumalaikolundusubramanian, P. (2009). Hypothesis of snake and insect venoms against human immunodeficiency virus: A review. AIDS Research and Therapy, 6(1), 25. 10.1186/1742-6405-6-25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng, J. , Xiao, G. , Zhang, J. , He, X. , Ou, M. , Bi, J. , … Zhang, G. (2020). Renin‐angiotensin system inhibitors improve the clinical outcomes of COVID‐19 patients with hypertension. Emerging Microbes & Infections, 9(1), 757–760. 10.1080/22221751.2020.1746200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messerli, F. H. , & Nussberger, J. (2000). Vasopeptidase inhibition and angio‐oedema. Lancet, 356(9230), 608–609. 10.1016/S0140-6736(00)02596-4 [DOI] [PubMed] [Google Scholar]
- Morais, K. L. P. , Hayashi, M. A. F. , Bruni, F. M. , Lopes‐Ferreira, M. , Camargo, A. C. M. , Ulrich, H. , & Lameu, C. (2011). Bj‐PRO‐5a, a natural angiotensin‐converting enzyme inhibitor, promotes vasodilatation mediated by both bradykinin B2 and M1 muscarinic acetylcholine receptors. Biochemical Pharmacology, 81(6), 736–742. 10.1016/j.bcp.2010.12.016 [DOI] [PubMed] [Google Scholar]
- Morais, K. L. P. , Ianzer, D. , Miranda, J. R. R. , Melo, R. L. , Guerreiro, J. R. , Santos, R. A. S. , … Lameu, C. (2013). Proline rich‐oligopeptides: Diverse mechanisms for antihypertensive action. Peptides, 48, 124–133. 10.1016/j.peptides.2013.07.016 [DOI] [PubMed] [Google Scholar]
- Muller, V. D. M. , Russo, R. R. , Oliveira Cintra, A. C. , Sartim, M. A. , De Melo Alves‐Paiva, R. , Figueiredo, L. T. M. , … Aquino, V. H. (2012). Crotoxin and phospholipases A2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon, 59(4), 507–515. 10.1016/j.toxicon.2011.05.021 [DOI] [PubMed] [Google Scholar]
- Munawar, A. , Zahid, A. , Negm, A. , Akrem, A. , Spencer, P. , & Betzel, C. (2016). Isolation and characterization of bradykinin potentiating peptides from Agkistrodon bilineatus venom. Proteome Science, 14(1), 1. 10.1186/s12953-016-0090-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murugesan, P. , Jung, B. , Lee, D. , Khang, G. , Doods, H. , & Wu, D. (2016). Kinin B1 receptor inhibition with BI113823 reduces inflammatory response, mitigates organ injury, and improves survival among rats with severe sepsis. Journal of Infectious Diseases, 213(4), 532–540. 10.1093/infdis/jiv426 [DOI] [PubMed] [Google Scholar]
- Ni, A. , Yin, H. , Agata, J. , Yang, Z. , Chao, L. , & Chao, J. (2003). Overexpression of kinin B 1 receptors induces hypertensive response to Des‐Arg 9‐bradykinin and susceptibility to inflammation. Journal of Biological Chemistry, 278(1), 219–225. 10.1074/jbc.M209490200 [DOI] [PubMed] [Google Scholar]
- Peng, L. , Liu, H. , Ruan, H. , Tepp, W. H. , Stoothoff, W. H. , Brown, R. H. , … Dong, M. (2013). Cytotoxicity of botulinum neurotoxins reveals a direct role of syntaxin 1 and SNAP‐25 in neuron survival. Nature Communications, 4(1), 1472. 10.1038/ncomms2462 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pennington, M. W. , Czerwinski, A. , & Norton, R. S. (2018). Peptide therapeutics from venom: Current status and potential. Bioorganic & Medicinal Chemistry, 26(10), 2738–2758. 10.1016/j.bmc.2017.09.029 [DOI] [PubMed] [Google Scholar]
- Phan, L. T. , Nguyen, T. V. , Luong, Q. C. , Nguyen, T. V. , Nguyen, H. T. , Le, H. Q. , … Pham, Q. D. (2020). Importation and human‐to‐human transmission of a novel coronavirus in Vietnam. New England Journal of Medicine, 382(9), 872–874. 10.1056/NEJMc2001272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puzer, L. , Vercesi, J. , Alves, M. F. M. , Barros, N. M. T. , Araujo, M. S. , Aparecida Juliano, M. , … Carmona, A. K. (2005). A possible alternative mechanism of kinin generation in vivo by cathepsin L. Biological Chemistry, 386(7), 699–704. 10.1515/BC.2005.081 [DOI] [PubMed] [Google Scholar]
- Qadri, F. , & Bader, M. (2018). Kinin B1 receptors as a therapeutic target for inflammation. Expert Opinion on Therapeutic Targets, 22(1), 31–44. 10.1080/14728222.2018.1409724 [DOI] [PubMed] [Google Scholar]
- Qin, C. , Zhou, L. , Hu, Z. , Zhang, S. , Yang, S. , Tao, Y. , … Tian, D.‐S. (2020). Dysregulation of immune response in patients with coronavirus 2019 (COVID‐19) in Wuhan, China. Clinical Infectious Diseases. 2020 Mar 12, ciaa248. 10.1093/cid/ciaa248. Online ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Querobino, S. M. , Ribeiro, C. A. J. , & Alberto‐Silva, C. (2018). Bradykinin‐potentiating PEPTIDE‐10C, an argininosuccinate synthetase activator, protects against H2O2‐induced oxidative stress in SH‐SY5Y neuroblastoma cells. Peptides, 103, 90–97. 10.1016/j.peptides.2018.03.017 [DOI] [PubMed] [Google Scholar]
- Rahman, A. M. , Murrow, J. R. , Ozkor, M. A. , Kavtaradze, N. , Lin, J. , De Staercke, C. , … Quyyumi, A. A. (2014). Endothelium‐derived hyperpolarizing factor mediates bradykinin‐stimulated tissue plasminogen activator release in humans. Journal of Vascular Research, 51(3), 200–208. 10.1159/000362666 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Recinos, A. , LeJeune, W. S. , Sun, H. , Lee, C. Y. , Tieu, B. C. , Lu, M. , … Brasier, A. R. (2007). Angiotensin II induces IL‐6 expression and the Jak‐STAT3 pathway in aortic adventitia of LDL receptor‐deficient mice. Atherosclerosis, 194(1), 125–133. 10.1016/j.atherosclerosis.2006.10.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivero, J. V. , de Castro, F. O. F. , Stival, A. S. , Magalhães, M. R. , Carmo Filho, J. R. , & Pfrimer, I. A. H. (2011). Mechanisms of virus resistance and antiviral activity of snake venoms. Journal of Venomous Animals and Toxins Including Tropical Diseases, 17(4), 387–393. 10.1590/S1678-91992011000400005 [DOI] [Google Scholar]
- Rodrigues Prestes, T. R. , Rocha, N. P. , Miranda, A. S. , Teixeira, A. L. , & Simoes‐e‐Silva, A. C. (2017). The anti‐inflammatory potential of ACE2/angiotensin‐(1–7)/mas receptor Axis: Evidence from basic and clinical research. Current Drug Targets, 18(11), 1301–1313. 10.2174/1389450117666160727142401 [DOI] [PubMed] [Google Scholar]
- Ruocco, G. , Feola, M. , & Palazzuoli, A. (2020). Hypertension prevalence in human coronavirus disease: The role of ACE system in infection spread and severity. International Journal of Infectious Diseases, 95, 373–375. 10.1016/j.ijid.2020.04.058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell, C. D. , Millar, J. E. , & Baillie, J. K. (2020). Clinical evidence does not support corticosteroid treatment for 2019‐nCoV lung injury. Lancet, 395(10223), 473–475. 10.1016/S0140-6736(20)30317-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sampaio, W. O. , Souza dos Santos, R. A. , Faria‐Silva, R. , da Mata Machado, L. T. , Schiffrin, E. L. , & Touyz, R. M. (2007). Angiotensin‐(1–7) through receptor Mas mediates endothelial nitric oxide synthase activation via Akt‐dependent pathways. Hypertension, 49(1), 185–192. 10.1161/01.HYP.0000251865.35728.2f [DOI] [PubMed] [Google Scholar]
- Santos, R. A. S. , e Silva, A. C. S. , Maric, C. , Silva, D. M. R. , Machado, R. P. , de Buhr, I. , … Walther, T. (2003). Angiotensin‐(1–7) is an endogenous ligand for the G protein‐coupled receptor Mas. Proceedings of the National Academy of Sciences, 100(14), 8258–8263. 10.1073/pnas.1432869100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos, R. A. S. , Ferreira, A. J. , & Simões e Silva, A. C. (2008). Recent advances in the angiotensin‐converting enzyme 2‐angiotensin(1–7)‐Mas axis. Experimental Physiology, 93(5), 519–527. 10.1113/expphysiol.2008.042002 [DOI] [PubMed] [Google Scholar]
- Saura, M. , Zaragoza, C. , McMillan, A. , Quick, R. A. , Hohenadl, C. , Lowenstein, J. M. , & Lowenstein, C. J. (1999). An antiviral mechanism of nitric oxide. Immunity, 10(1), 21–28. 10.1016/S1074-7613(00)80003-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Searles, C. D. , & Harrison, D. G. (1999). The interaction of nitric oxide, bradykinin, and the angiotensin II type 2 receptor: Lessons learned from transgenic mice. Journal of Clinical Investigation, 104(8), 1013–1014. 10.1172/JCI8507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Senchenkova, E. Y. , Russell, J. , Almeida‐Paula, L. D. , Harding, J. W. , & Granger, D. N. (2010). Angiotensin II–mediated microvascular thrombosis. Hypertension, 56(6), 1089–1095. 10.1161/HYPERTENSIONAHA.110.158220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silhol, F. , Sarlon, G. , Deharo, J.‐C. , & Vaïsse, B. (2020). Downregulation of ACE2 induces overstimulation of the renin–angiotensin system in COVID‐19: Should we block the renin–angiotensin system? Hypertension Research, 43, 854–856. 10.1038/s41440-020-0476-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmons, G. , Gosalia, D. N. , Rennekamp, A. J. , Reeves, J. D. , Diamond, S. L. , & Bates, P. (2005). Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proceedings of the National Academy of Sciences, 102(33), 11876–11881. 10.1073/pnas.0505577102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siragy, H. M. , & Carey, R. M. (1996). The subtype‐2 (AT2) angiotensin receptor regulates renal cyclic guanosine 3′, 5′‐monophosphate and AT1 receptor‐mediated prostaglandin E2 production in conscious rats. Journal of Clinical Investigation, 97(8), 1978–1982. 10.1172/JCI118630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, W. L. , & Song, I. (2002). The enzymology of prostaglandin endoperoxide H synthases‐1 and ‐2. Prostaglandins & Other Lipid Mediators, 68–69, 115–128. 10.1016/S0090-6980(02)00025-4 [DOI] [PubMed] [Google Scholar]
- Sodhi, C. P. , Wohlford‐Lenane, C. , Yamaguchi, Y. , Prindle, T. , Fulton, W. B. , Wang, S. , … Jia, H. (2018). Attenuation of pulmonary ACE2 activity impairs inactivation of des‐Arg 9 bradykinin/BKB1R axis and facilitates LPS‐induced neutrophil infiltration. American Journal of Physiology‐Lung Cellular and Molecular Physiology, 314(1), L17–L31. 10.1152/ajplung.00498.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spagnolo, P. , Balestro, E. , Aliberti, S. , Cocconcelli, E. , Biondini, D. , Casa, G. D. , … Maher, T. M. (2020). Pulmonary fibrosis secondary to COVID‐19: A call to arms? Lancet Respiratory Medicine. S2213‐2600(20), 30222–8. 10.1016/S2213-2600(20)30222-8. Online ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki, H. , Kimura, K. , Shirai, H. , Eguchi, K. , Higuchi, S. , Hinoki, A. , … Eguchi, S. (2009). Endothelial nitric oxide synthase inhibits G 12/13 and rho‐kinase activated by the angiotensin II type‐1 receptor. Arteriosclerosis, Thrombosis, and Vascular Biology, 29(2), 217–224. 10.1161/ATVBAHA.108.181024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taddei, S. , & Bortolotto, L. (2016). Unraveling the pivotal role of bradykinin in ACE inhibitor activity. American Journal of Cardiovascular Drugs, 16(5), 309–321. 10.1007/s40256-016-0173-4 [DOI] [PubMed] [Google Scholar]
- Taddei, S. , Ghiadoni, L. , Virdis, A. , Versari, D. , & Salvetti, A. (2003). Mechanisms of endothelial dysfunction: Clinical significance and preventive non‐pharmacological therapeutic strategies. Current Pharmaceutical Design, 9(29), 2385–2402. 10.2174/1381612033453866 [DOI] [PubMed] [Google Scholar]
- Thachil, J. (2020). The versatile heparin in COVID‐19. Journal of Thrombosis and Haemostasis, 18(5), 1020–1022. 10.1111/jth.14821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomasian, D. (2000). Antioxidants and the bioactivity of endothelium‐derived nitric oxide. Cardiovascular Research, 47(3), 426–435. 10.1016/S0008-6363(00)00103-6 [DOI] [PubMed] [Google Scholar]
- van de Veerdonk, F. L. , Netea, M. G. , van Deuren, M. , van der Meer, J. W. , de Mast, Q. , Brüggemann, R. J. , & van der Hoeven, H. (2020). Kallikrein–kinin blockade in patients with COVID‐19 to prevent acute respiratory distress syndrome. eLife, 9, e57555. 10.7554/eLife.57555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varga, Z. , Flammer, A. J. , Steiger, P. , Haberecker, M. , Andermatt, R. , Zinkernagel, A. S. , … Moch, H. (2020). Endothelial cell infection and endotheliitis in COVID‐19. Lancet, 395(10234), 1417–1418. 10.1016/S0140-6736(20)30937-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walls, A. C. , Park, Y.‐J. , Tortorici, M. A. , Wall, A. , McGuire, A. T. , & Veesler, D. (2020). Structure, function, and antigenicity of the SARS‐CoV‐2 spike glycoprotein. Cell, 181(2), 281–292.e6. 10.1016/j.cell.2020.02.058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, B. A. , Nautiyal, M. , Gwathmey, T. M. , Rose, J. C. , & Chappell, M. C. (2016). Evidence for a mitochondrial angiotensin‐(1–7) system in the kidney. American Journal of Physiology‐Renal Physiology, 310(7), F637–F645. 10.1152/ajprenal.00479.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiman, B. , Andersson, T. , Hallqvist, J. , Reuterwall, C. , Ahlbom, A. , & DeFaire, U. (2000). Plasma levels of tissue plasminogen activator/plasminogen activator inhibitor‐1 complex and von Willebrand factor are significant risk markers for recurrent myocardial infarction in the Stockholm heart epidemiology program (SHEEP) study. Arteriosclerosis, Thrombosis, and Vascular Biology, 20(8), 2019–2023. 10.1161/01.ATV.20.8.2019 [DOI] [PubMed] [Google Scholar]
- Wu, D. , Lin, X. , Bernloehr, C. , Hildebrandt, T. , & Doods, H. (2012). Effects of a novel bradykinin B1 receptor antagonist and angiotensin II receptor blockade on experimental myocardial infarction in rats. PLoS One, 7(12), e51151. 10.1371/journal.pone.0051151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu, F. , Zhao, S. , Yu, B. , Chen, Y.‐M. , Wang, W. , Song, Z.‐G. , … Zhang, Y.‐Z. (2020). A new coronavirus associated with human respiratory disease in China. Nature, 579(7798), 265–269. 10.1038/s41586-020-2008-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, P. , Zhou, Q. , & Xu, J. (2020). Mechanism of thrombocytopenia in COVID‐19 patients. Annals of Hematology, 99(6), 1205–1208. 10.1007/s00277-020-04019-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, X. , Greenland, J. R. , Gotts, J. E. , Matthay, M. A. , & Caughey, G. H. (2016). Cathepsin L helps to defend mice from infection with influenza A. PLoS One, 11(10), e0164501. 10.1371/journal.pone.0164501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, R.‐F. , Yin, J.‐X. , Li, Y.‐L. , Zimmerman, M. C. , & Schultz, H. D. (2011). Angiotensin‐(1–7) increases neuronal potassium current via a nitric oxide‐dependent mechanism. American Journal of Physiology‐Cell Physiology, 300(1), C58–C64. 10.1152/ajpcell.00369.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, H. , Penninger, J. M. , Li, Y. , Zhong, N. , & Slutsky, A. S. (2020). Angiotensin‐converting enzyme 2 (ACE2) as a SARS‐CoV‐2 receptor: Molecular mechanisms and potential therapeutic target. Intensive Care Medicine, 46(4), 586–590. 10.1007/s00134-020-05985-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, J. , Ma, X. , Yu, F. , Liu, J. , Zou, F. , Pan, T. , & Zhang, H. (2020). Teicoplanin potently blocks the cell entry of 2019‐nCoV. BioRxiv Preprint. 10.1101/2020.02.05.935387 [DOI]
- Zhou, P. , Yang, X.‐L. , Wang, X.‐G. , Hu, B. , Zhang, L. , Zhang, W. , … Shi, Z.‐L. (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature, 579(7798), 270–273. 10.1038/s41586-020-2012-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou, X. , Ai, S. , Chen, Z. , & Li, C. (2019). Probucol promotes high glucose‐induced proliferation and inhibits apoptosis by reducing reactive oxygen species generation in Müller cells. International Ophthalmology, 39(12), 2833–2842. 10.1007/s10792-019-01130-8 [DOI] [PubMed] [Google Scholar]
- Zisman, L. S. (2005). ACE and ACE2: A tale of two enzymes. European Heart Journal, 26(4), 322–324. 10.1093/eurheartj/ehi043 [DOI] [PMC free article] [PubMed] [Google Scholar]