

Abstract
TMPRSS2 is both the most frequently altered gene in primary prostate cancer and a critical factor enabling cellular infection by coronaviruses, including SARS-CoV-2. The modulation of its expression by sex steroids could contribute to the male predominance of severe infections and given that TMPRSS2 has no known indispensable functions, and inhibitors are available, it is an appealing target for prevention or treatment of respiratory viral infections.
The global COVID-19 pandemic, caused by the SARS-CoV-2 virus, has led to over 1,400,000 diagnosed cases and over 80,000 reported deaths as of April 8, 2020. These epidemiological statistics represent the tip of the iceberg, given ongoing significant transmission, high rates of subclinical infection, insufficient testing in multiple communities, and potential differences in attribution of cause of death in infected patients.
The global research community has coalesced on multiple fronts to understand the mechanisms of infection and the heterogeneity in the virulence of SARS-CoV-2, as well as the constellation of symptoms and risk factors for subsequent mortality. One key discovery in understanding the mechanism of SARS-CoV-2 infection involves the role of the transmembrane serine protease 2 (TMPRSS2), a cell surface protein that is expressed by epithelial cells of specific tissues including those in the aerodigestive tract. As one of the serendipities of science, many of the insights related to TMPRSS2 have come from cancer research. This overview summarizes the history of the connection of TMPRSS2 with coronaviruses as well as influenza viruses, provides insights derived from cancer research, and integrates what is known (and not known) concerning the potential roles of TMPRSS2 as a target for intervention or prevention of COVID-19.
TMPRSS2 and respiratory viruses
Coronaviruses as well as influenza viruses critically depend on TMPRSS2 for viral entry and spread in the host. As a first step enabling host cell entry, the viral hemagglutinin protein attaches to angiotensin-converting enzyme 2 (ACE2), encoded by the ACE2 gene, that is expressed on respiratory epithelial cells. In a second step, hemagglutinin is cleaved to activate internalization of the virus. This second step is dependent on proteases on the host cell, particularly the type 2 transmembrane serine protease, TMPRSS2 (1). Moreover, not only SARS-CoV-2, but also other types of coronaviruses and influenza viruses depend on TMPRSS2 for viral activation and cell entry, including SARS-CoV, the agent responsible for the 2003 SARS outbreak, as well as influenza H1N1, the agent responsible for the 1918 and 2009 influenza pandemics (2–4). These examples highlight the central and conserved role of TMPRSS2 in the pathogenesis of the illnesses caused by coronaviruses and influenza viruses.
In an in vitro study using cell lines and primary pulmonary cells, an inhibitor of the protease activity of TMPRSS2, camostat mesylate, partially inhibited the entry of SARS-CoV-2 into these lung epithelial cells (4). In a Tmprss2 knockout model, mice infected with the H1N1 influenza virus showed minimal initial infection and had a considerably attenuated disease course with protection from lung pathology, weight loss, and mortality compared to wild-type control mice (5). Given its central role in initiating SARS-CoV-2 and other respiratory viral infections, modulating TMPRSS2 expression or activity is hypothesized to represent a promising candidate for potential interventions against COVID-19.
TMPRSS2, a key regulator in prostate cancer
TMPRSS2 was first identified in prostate cancer shortly after the gene had been originally cloned. Prostate cancer cell lines strongly upregulated TMPRSS2 expression in response to androgens (6). TMPRSS2 is expressed on the luminal side of the prostate epithelium, and its expression is increased in prostate cancer tissue compared to non-cancerous prostate tissue (7). Notably, the TMPRSS2 gene is a partner in one of the most common gene fusion events in solid tumors: somatic gene rearrangements involving TMPRSS2 with a member of the ETS family of oncogenic transcription factors, most commonly ERG. This fusion occurs in approximately 50% of primary prostate cancers among men of European ancestry. While ERG is not normally regulated by androgen, the gene fusion juxtaposes the androgen receptor regulatory elements of TMPRSS2 with the ERG gene. The ERG gene is consequently controlled by androgen receptor signaling and expressed highly in prostate cancers harboring the TMPRSS2:ERG fusion.
Intriguingly, the prevalence of the TMPRSS2:ERG fusion is lower in prostate tumors of both black and Asian men. The relevance of this to the current COVID-19 pandemic is unclear. TMPRSS2:ERG fusion-positive prostate cancers also have a distinct set of risk factors related to hormonal signaling. For example, men with higher genetically determined transcriptional activity of the androgen receptor have a higher risk of TMPRSS2:ERG fusion-positive prostate cancer but not of fusion-negative prostate cancer (8). Moreover, in mice, prostate tumors arising in the presence of Tmprss2 have a higher capacity to metastasize (9). In men, tumors with the TMPRSS2:ERG fusion have higher insulin/insulin-like growth factor signaling, and TMPRSS2:ERG may modify how hormonal risk factors such as obesity influence the risk of metastasis (10). Together with the observation that TMPRSS2:ERG fusion-positive and fusion-negative prostate cancers may have distinct germline genetic risk factors (11), these findings from cancer research suggest potential drivers of differential TMPRSS2 expression.
TMPRSS2 and susceptibility to coronavirus and influenza infections
Understanding how TMPRSS2 protein expression in the lung varies in the population could reveal important insights into differential susceptibility to influenza and coronavirus infections. Immunohistochemical studies, with limited sample size, suggest that the TMPRSS2 protein is more heavily expressed in bronchial epithelial cells than in surfactant-producing type II alveolar cells and alveolar macrophages, and that there is no expression in type I alveolar cells that form the respiratory surface (Fig. 1A) (12).
Figure 1.
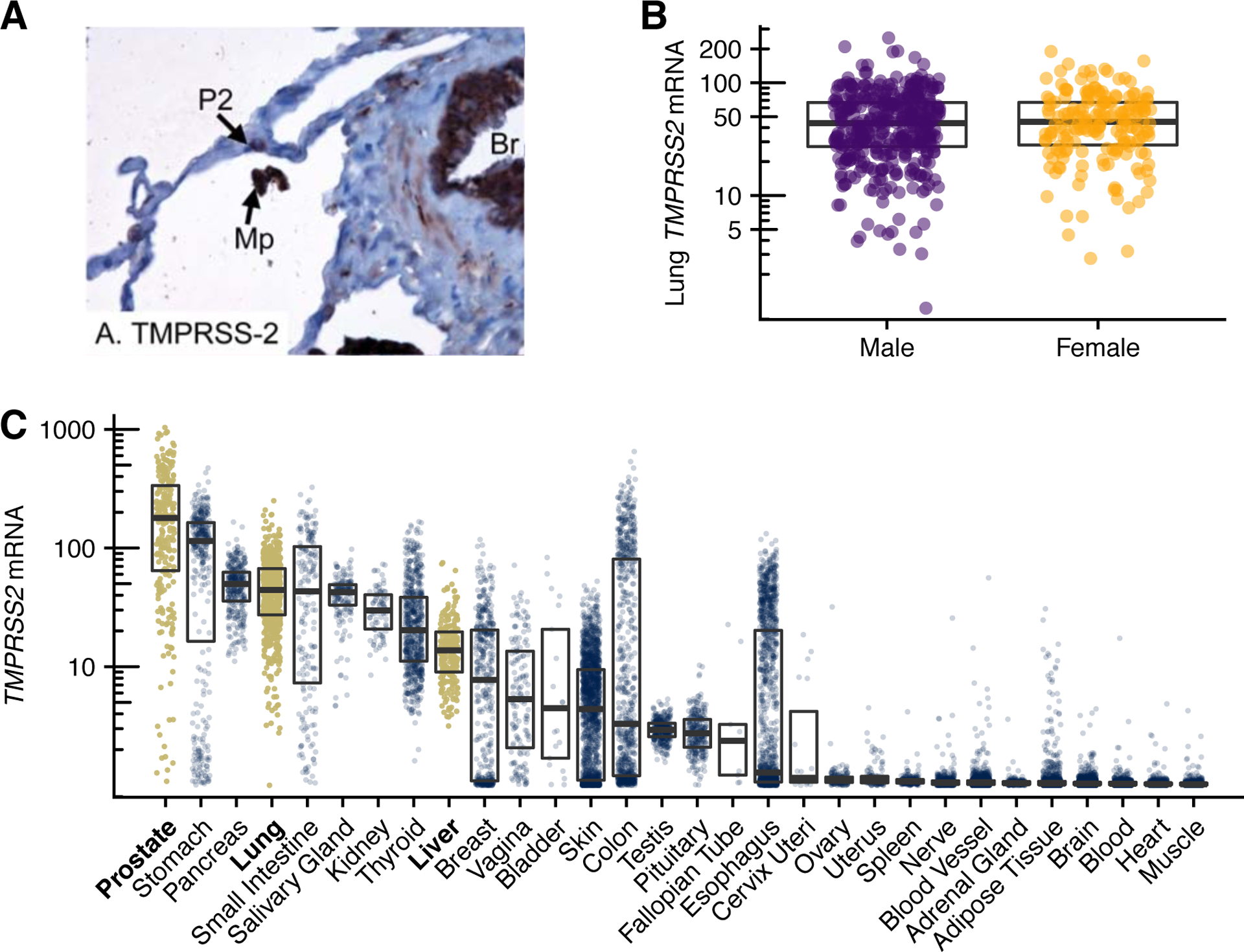
TMPRSS2 mRNA and TMPRSS2 protein expression in humans. A, TMPRSS2 lung expression is localized to type 2 alveolar cells (P2), alveolar macrophages (Mp) and bronchial epithelial cells (Br). B, TMPRSS2 expression is similar in men and women. C, TMPRSS2 is expressed in several human tissues. Levels in prostate, lung, and liver are highlighted. A is from (12), reused under the CC-BY license. Data for B and C are from the Genotype-Tissue Expression (GTEx) project, version 8, showing log10 of RNA transcripts per million (TPM) + 1; boxes span interquartile ranges, thick lines indicate medians.
Determinants of TMRPSS2 protein expression in the lung could overlap with risk factors of TMPRSS2:ERG fusion-positive prostate cancer, some of which are modifiable and may merit testing in trials as a means of reducing risk of respiratory viral infections—as well as of prostate cancers harboring the TMPRSS2:ERG fusion. It is important that studies also be conducted in lung tissue from individuals not affected by a respiratory virus, given that viral infection may in turn alter TMPRSS2 expression, just as SARS-CoV does with the ACE2 receptor (13).
Importantly, such insights can indeed help better understand the risk of respiratory viral infections. For example, patients who carried a single nucleotide polymorphism associated with higher TMPRSS2 expression were more susceptible to influenza virus infection in two separate patient cohorts (14).
Does TMPRSS2 explain the male predominance in deaths from COVID-19?
There are suggestions from epidemiological studies across diverse countries including China, Italy, and the United States that the incidence and severity of diagnosed COVID-19 as well as other TMPRSS2-dependent viral infections such as influenza may be higher in men than women. One explanation underlying these potential differences is cigarette smoking, which is generally more common among men, and to date the epidemiological investigations of COVID-19 have not sufficiently accounted for tobacco use in sex comparisons. Any sex-specific differences in the overall incidence should also take into account possible differences in laboratory testing for SARS-CoV-2. Still, the presence of TMPRSS2:ERG in prostate cancer as well as the strong regulation of TMPRSS2 by androgens have raised the hypothesis that the male predominance in the COVID-19 pandemic could partially be explained by TMPRSS2.
Interestingly, at least on an mRNA level, constitutive expression of TMPRSS2 in lung tissue does not appear to differ between men and women (Fig. 1B). Yet, there is a wide variation among both sexes in terms of mRNA expression levels. Low levels of androgens present in women may suffice to sustain TMPRSS2 expression. In addition, TMPRSS2 (and tumors with the TMPRSS2:ERG fusion) may be responsive to estrogen signaling (9,15). It is tempting to speculate that androgen receptor-inhibitory therapies might reduce susceptibility to COVID-19 pulmonary symptoms and mortality.
Targeting TMPRSS2 expression
An important open question is to what degree susceptibility to viral infection could potentially be reduced by inhibiting androgen signaling. A subsequent question is to what extent TMPRSS2 protein expression in the lung or other sites of viral entry is regulated by androgen signaling.
Studies thus far suggest that androgen receptors are expressed in the epithelium of the respirator tract in mice and humans, especially in type 2 pneumocytes and in bronchial epithelial cells. Androgen administration to a lung adenocarcinoma cell line upregulated the TMPRSS2 transcript over two-fold, accompanied by an androgen-dependent loading of the androgen receptor protein onto the TMPRSS2 enhancer (16). However, whether antagonists of androgen receptor signaling can in turn abrogate TMPRSS2 expression in this cell line, or in non-cancerous human respiratory epithelium, in not known. Observational studies that assess TMPRSS2 protein expression in (non-tumor) lung tissue from men who have been treated for prostate cancer with androgen receptor signaling inhibitors, compared to those with normal androgen levels, could shed further light on this question.
If indeed observational studies further corroborate androgen receptor signaling inhibition as a viable strategy, then several therapeutics that effectively repress androgen receptor signaling activity could be rapidly repurposed to determine benefit in patients with COVID-19 infection. These medications have been used in the treatment of prostate cancer for decades. Commercially available antiandrogens such as enzalutamide, apalutamide, or darolutamide, or chemical gonadal ablation, could potentially downregulate expression of TMPRSS2, thereby attenuating symptom severity in patients who have contracted the SARS-CoV-2 virus. Such a trial could be conducted rapidly, given the immediate availability of these agents as well as their known safety profile in both men and women. Further, if TMPRSS2 is negatively regulated via estrogen receptor activity, then similar studies of estrogen receptor-modulating drugs could be considered.
Targeting TMPRSS2 protease activity
Besides the theoretical potential for androgen receptor-targeted therapies to modulate TMPRSS2 expression, an alternative strategy involves directly impairing the protease activity of TMPRSS2. Though not developed specifically for targeting TMPRSS2, the protease inhibitors camostat, nafamostat, and aerosolized aprotinin have been shown to attenuate TMPRSS2 protease activity and are approved for the unrelated indications in specific countries. At least one phase 1–2 clinical trial of camostat for COVID-19 (ClinicalTrials.gov, NCT04321096) has been recruiting since April 6, 2020. Whether camostat leads to clinical toxicity by inhibiting trypsin-like proteases other than TMPRSS2 remains to be seen.
Additional discoveries in cancer research point to other potentially promising TMPRSS2 antagonists. Preclinical studies suggested that TMPRSS2 promoted the metastatic spread of prostate cancer. A large-scale chemical library screen designed to select compounds capable of blocking TMPRSS2 activity identified bromhexine as a potent and TMPRSS2-specific protease inhibitor. Systemic administration of bromhexine reduced the frequency of prostate cancer metastases with no evidence of systemic toxicity (9).
Bromhexine has been in clinical use for decades as a mucolytic agent and expectorant. However, on a cautionary note, bromhexine’s long track record should not be taken as evidence for its efficacy or safety at dosing schedules needed to sufficiently inhibit TMPRSS2 in patients, given the paucity of modern randomized-controlled trials. Indeed, TMPRSS2 mRNA and protein are expressed in several tissues other than lung and prostate (7,12) (Fig. 1C), perhaps contributing one explanation for non-respiratory symptoms seen with influenza and coronaviruses. That TMPRSS2 is expressed in liver tissue, for example, merits attention in clinical trials of TMPRSS2 inhibitors, as the functional role of TMPRSS2 in the normal physiology of various cells and tissues is not completely understood. In the normal prostate, TMPRSS2 contributes to proteolytic cascades that result in the activation of prostate-specific antigen (PSA), itself a protease involved in ejaculate production (9).
Notably, TMPRSS2 does not appear to play an essential role in any organ, as other proteases may provide a degree of redundancy. A compelling argument that TMPRSS2 inhibition may have little on-target side effects again comes from the Tmprss2 knockout mouse model in cancer research, where Tmprss2 appeared entirely dispensable for normal development, growth, and organ function (17).
Conclusions
Taken together, cancer-focused studies have contributed critical elements to our understanding of TMPRSS2, a key factor in the pathogenesis of coronavirus and influenza infections. Cancer epidemiology studies point to potential explanations for differential susceptibility to such infections through androgen regulation and differential TMPRSS2 expression. It remains to be explored in well-designed observational studies whether TMPRSS2 polymorphisms or other determinants of differential expression contribute to viral susceptibility, disease progression and mortality. Approaches that merit further study and consideration for clinical testing could be the modulation of TMPRSS2 expression indirectly via androgen pathway blockade, or direct inhibition of TMPRSS2 function using protease inhibitors. As with all other potential novel treatments for COVID-19, such as those targeted at the ACE2 receptor, the existing data on TMPRSS2-targeted treatments can only be summarized as promising hypotheses. They require further nonrandomized studies and eventually rigorous testing for efficacy and safety by randomized-controlled trials in humans.
Funding
The work of the authors is supported by the Prostate Cancer Foundation, the National Institutes of Health (P30CA008748, P30CA006973, P30CA015704, P01CA163227, P01CA228696), and the Department of Defense (W81XWH-18-1-0330).
Footnotes
Conflicts of Interest
As of April 9, 2020, E.S.A. is a paid consultant/advisor to Janssen, Astellas, Sanofi, Dendreon, Pfizer, Amgen, AstraZeneca, Bristol-Myers Squibb, Clovis, and Merck; he has received research funding to his institution from Janssen, Johnson & Johnson, Sanofi, Dendreon, Genentech, Novartis, Tokai, Bristol Myers-Squibb, AstraZeneca, Clovis, and Merck; and he is the co-inventor of a biomarker technology that has been licensed to Qiagen. P.S. Nelson is a consultant/scientific advisory board member for Astellas and Janssen and reports other remuneration by Veneble/Fitzpatrick. P.W. Kantoff reports the following disclosures for the last 24-month period: he has investment interest in Context Therapeutics LLC, DRGT, Placon, Seer Biosciences, he is a company board member for Context Therapeutics LLC, and is a consultant/scientific advisory board member for Bavarian Nordic Immunotherapeutics, DRGT, GE Healthcare, Janssen, OncoCellMDX, Progenity, Seer Biosciences and Tarveda Therapeutics, and serves on data safety monitoring boards for Genentech/Roche and Merck. The remaining authors have no conflicts of interest to disclose.
References
- 1.Bottcher E, Matrosovich T, Beyerle M, Klenk HD, Garten W, Matrosovich M. Proteolytic activation of influenza viruses by serine proteases TMPRSS2 and HAT from human airway epithelium. J Virol 2006;80(19):9896–8 doi 10.1128/JVI.01118-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chaipan C, Kobasa D, Bertram S, Glowacka I, Steffen I, Tsegaye TS, et al. Proteolytic activation of the 1918 influenza virus hemagglutinin. J Virol 2009;83(7):3200–11 doi 10.1128/JVI.02205-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Matsuyama S, Nagata N, Shirato K, Kawase M, Takeda M, Taguchi F. Efficient activation of the severe acute respiratory syndrome coronavirus spike protein by the transmembrane protease TMPRSS2. J Virol 2010;84(24):12658–64 doi 10.1128/JVI.01542-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hoffmann M, Kleine-Weber H, Schroeder S, Kruger N, Herrler T, Erichsen S, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020. doi 10.1016/j.cell.2020.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hatesuer B, Bertram S, Mehnert N, Bahgat MM, Nelson PS, Pohlmann S, et al. Tmprss2 is essential for influenza H1N1 virus pathogenesis in mice. PLoS Pathog 2013;9(12):e1003774 doi 10.1371/journal.ppat.1003774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lin B, Ferguson C, White JT, Wang S, Vessella R, True LD, et al. Prostate-localized and androgen-regulated expression of the membrane-bound serine protease TMPRSS2. Cancer Res 1999;59(17):4180–4. [PubMed] [Google Scholar]
- 7.Lucas JM, True L, Hawley S, Matsumura M, Morrissey C, Vessella R, et al. The androgen-regulated type II serine protease TMPRSS2 is differentially expressed and mislocalized in prostate adenocarcinoma. J Pathol 2008;215(2):118–25 doi 10.1002/path.2330. [DOI] [PubMed] [Google Scholar]
- 8.Yoo S, Pettersson A, Jordahl KM, Lis RT, Lindstrom S, Meisner A, et al. Androgen Receptor CAG Repeat Polymorphism and Risk of TMPRSS2:ERG-Positive Prostate Cancer. Cancer Epidemiol Biomarkers Prev 2014;23(10):2027–31 doi 10.1158/1055-9965.EPI-14-0020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lucas JM, Heinlein C, Kim T, Hernandez SA, Malik MS, True LD, et al. The androgen-regulated protease TMPRSS2 activates a proteolytic cascade involving components of the tumor microenvironment and promotes prostate cancer metastasis. Cancer Discov 2014;4(11):1310–25 doi 10.1158/2159-8290.CD-13-1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pettersson A, Lis RT, Meisner A, Flavin R, Stack EC, Fiorentino M, et al. Modification of the association between obesity and lethal prostate cancer by TMPRSS2:ERG. J Natl Cancer Inst 2013;105(24):1881–90 doi 10.1093/jnci/djt332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Penney KL, Pettersson A, Shui IM, Graff RE, Kraft P, Lis RT, et al. Association of Prostate Cancer Risk Variants with TMPRSS2:ERG Status: Evidence for Distinct Molecular Subtypes. Cancer Epidemiol Biomarkers Prev 2016;25(5):745–9 doi 10.1158/1055-9965.EPI-15-1078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bertram S, Heurich A, Lavender H, Gierer S, Danisch S, Perin P, et al. Influenza and SARS-coronavirus activating proteases TMPRSS2 and HAT are expressed at multiple sites in human respiratory and gastrointestinal tracts. PLoS One 2012;7(4):e35876 doi 10.1371/journal.pone.0035876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kuba K, Imai Y, Rao S, Gao H, Guo F, Guan B, et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat Med 2005;11(8):875–9 doi 10.1038/nm1267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cheng Z, Zhou J, To KK, Chu H, Li C, Wang D, et al. Identification of TMPRSS2 as a Susceptibility Gene for Severe 2009 Pandemic A(H1N1) Influenza and A(H7N9) Influenza. J Infect Dis 2015;212(8):1214–21 doi 10.1093/infdis/jiv246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Setlur SR, Mertz KD, Hoshida Y, Demichelis F, Lupien M, Perner S, et al. Estrogen-dependent signaling in a molecularly distinct subclass of aggressive prostate cancer. J Natl Cancer Inst 2008;100(11):815–25 doi 10.1093/jnci/djn150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mikkonen L, Pihlajamaa P, Sahu B, Zhang FP, Janne OA. Androgen receptor and androgen-dependent gene expression in lung. Mol Cell Endocrinol 2010;317(1–2):14–24 doi 10.1016/j.mce.2009.12.022. [DOI] [PubMed] [Google Scholar]
- 17.Kim TS, Heinlein C, Hackman RC, Nelson PS. Phenotypic analysis of mice lacking the Tmprss2-encoded protease. Mol Cell Biol 2006;26(3):965–75 doi 10.1128/MCB.26.3.965-975.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]