

Abstract
The natural history of allergic diseases suggests bidirectional and progressive relationships between allergic disorders of the skin, lung and gut indicative of mucosal organ cross-talk. However, impacts of local allergic inflammation on the cellular landscape of remote mucosal organs along the skin:lung:gut axis are not yet known. Eosinophils are tissue-dwelling innate immune leukocytes associated with allergic diseases. Emerging data suggest heterogeneous phenotypes of tissue-dwelling eosinophils contribute to multifaceted roles that favor homeostasis or disease. This study investigated the impact of acute local allergen exposure on the frequency and phenotype of tissue eosinophils within remote mucosal organs. Our findings demonstrate allergen challenge to skin, lung or gut elicited not only local eosinophilic inflammation, but also increased the number and frequency of eosinophils within remote, allergen non-exposed lung and intestine. Remote allergen-elicited lung eosinophils exhibited an inflammatory phenotype and their presence associated with enhanced susceptibility to airway inflammation induced upon subsequent inhalation of a different allergen. These data demonstrate, for the first time, a direct effect of acute allergic inflammation on the phenotype and frequency of tissue eosinophils within antigen non-exposed remote mucosal tissues associated with remote organ priming for allergic inflammation.
INTRODUCTION
Allergic diseases encompass a range of chronic, relapsing inflammatory disorders (e.g. atopic dermatitis, hay fever, asthma, food allergy) characterized by Th2 immunity. Their global prevalence has risen dramatically within the past several decades such that allergic diseases now affect approximately 20% of the worldwide population and represent the most common chronic conditions in childhood 1. Frequently encountered as sequential- or co-morbidities, the natural history of allergic diseases, as revealed through cross-sectional and longitudinal clinical studies and animal models, suggests progressive and causal relationships between allergic disorders of the skin, lung and gut collectively referred to as the “allergic march”, and often reflective of sensitization to multiple allergens 2-10. Despite strong epidemiologic evidence for causal and bidirectional relationships between cutaneous, respiratory and gastrointestinal allergic reactions, cellular mechanisms that link local allergic reactions to remote organs along the skin:lung:gut axis and prime for the development of subsequent allergic reactions in response to new allergens remain unknown.
Eosinophils have long been implicated in allergic diseases, and increased numbers of eosinophils have been observed within both the airways and intestinal tracts of patients with asthma or allergic rhinitis and within both skin and colon of patients with atopic eczema (reviewed in 11). Data from our lab and others’ have identified phenotypically distinct populations of tissue resident eosinophils, particularly within the intestinal tract 12, 13 and lung 14, 15, exhibiting putatively homeostatic or inflammatory phenotypes. We hypothesized that the frequency and/or functional phenotype of mucosal tissue eosinophils might be modulated in response to allergic inflammatory reactions occurring at remote organs, and that these cellular alterations might prime mucosal organs for allergic inflammation. To test this hypothesis, we modified acute models of allergic inflammation localized to the skin, airway or GI tract to model the allergic march in mice and investigated the frequency and phenotype of tissue-resident eosinophils within remote, allergen non-exposed organs along the skin:lung:gut axis. Our findings demonstrate that in allergen-sensitized mice direct challenge of the skin, lung or gut with allergen elicited not only a local eosinophilic inflammation at the site of allergen exposure, but also increased the number and frequency of eosinophils within allergen non-exposed lung and intestine. Remote allergen-recruited airway and intestinal eosinophils exhibited an activated phenotype. Moreover, the accumulation of inflammatory eosinophils within the allergen non-exposed lung was associated with mucous secretion and an exacerbated allergic airways response to subsequent inhalation of an antigenically distinct allergen, house dust mite (HDM). These data add to our understanding of the phenotypic diversity of mucosal tissue-resident eosinophils and reveal, for the first time, a direct relationship between local allergic inflammation and eosinophil frequency and phenotypes within remote tissues along the skin:lung:gut axis that prime for allergic inflammation. As such, these findings shed light on how local allergic manifestations of the skin, respiratory tract or gut may predispose individuals for concurrent and sequential development of allergenically distinct allergic diseases at remote loci. Our results are therefore relevant to atopic patients at risk for progressing along the “allergic march”.
RESULTS
Developing a mouse model to investigate the atopic march.
To investigate effects of local allergic inflammation on the number and phenotype of tissue-resident eosinophils at remote sites along the skin:lung:gut axis, we modified mouse models of local skin (atopic dermatitis), lung (allergic airways disease) and gut (food allergy) allergen exposure to achieve three acute models that elicited local eosinophilic inflammation in parallel with similar levels of peripheral blood eosinophilia (Fig 1a). Strong systemic Th2 immunity is the classic characteristic of patients at greatest risk of progressing along the atopic march. Therefore, as a foundation for each model we utilized a standard methodology of robust systemic Th2 sensitization in mice through intraperitoneal sensitization with the soluble protein antigen ovalbumin (OVA) admixed with alum adjuvant. Among other functions, alum has been shown to elicit release of IL-33 16 that could directly influence multiple aspects of the allergic march, including eosinophil hematopoiesis and recruitment 17. Therefore to avoid any potential confounding effects of alum, control mice throughout the study received sham sensitizations with PBS admixed with alum. Importantly, OVA-alum sensitization alone had no significant impact on eosinophil frequencies in bone marrow, spleen or intestinal tissues (Supplemental Fig 1).
Figure 1. Mouse models of acute local and systemic eosinophilic infiltration.

(a) Experimental timeline. Mice were sensitized with 3 intraperitoneal injections of OVA-alum followed one week later by 3 consecutive challenges through epicutaneous, lower airways, or oral routes with OVA. Sham control mice were sensitized with PBS-alum followed by challenges with PBS. Mice were sacrificed 4 days after the final allergen challenge and eosinophils quantified from the site of allergen challenge (b-e) or peripheral blood (f-h). (b) Eosinophils were quantified from H&E stained sections of the dermis following allergen administration to tape-stripped skin. Following allergen administration to the lower airways lung tissue sections revealed cellular infiltrates (c) and induction of BAL fluid eosinophilia, as quantified by flow cytometry (d). (e) Oral gavage of allergen elicited increased numbers of eosinophils within intestinal jejunum, as quantified from αMBP immunohistochemistry. Allergen challenge to the skin (f), lung (g) or gut (h) also elicited an increase in peripheral eosinophils. Data shown are from 1 representative experiment (d) or combined data from 2 independent experiments (b,e,f-h). (b,e) Magnification 400x, scale bar 80 μm. (c) 200x, scale bar 200 μm. Error bars show standard deviation. **, p<0.001; ***, p<0.0001.
Following systemic sensitization to OVA-alum mice were challenged with OVA on three consecutive days by direct administration to either the skin, lung or gut, and tissue eosinophils within lung and gut compartments assessed four days after the final allergen challenge (Fig 1a). Sham control mice were sensitized with PBS-alum followed by challenge with PBS vehicle control. For skin challenge, OVA was applied to tape-stripped skin to mimic scratching of AD patients (see Methods). To deliver OVA directly to lower airways while avoiding direct allergen exposure to the gastrointestinal tract inherent in the standard methods of airway allergen exposures (i.e. aerosolization or intranasal instillation) mice received challenge doses of OVA aerosolized directly into the right lung using a microsprayer (see Methods). Of note, although the endotracheal spray method avoids direct allergen exposure to the gastrointestinal tract, it is plausible that self-clearing mechanisms associated with the respiratory mucociliary escalator may indirectly deliver a fraction of the allergen to intestinal tissues, as would also occur in humans. Delivery of OVA to the gut was achieved by oral gavage.
For each of the three acute allergic disease models, tissue recovered from the respective sites of allergen exposure (i.e. skin, lung or gut) four days after final allergen challenge revealed local eosinophilic infiltration. For the AD model this was manifest as an increase in dermis eosinophils (Fig 1b). In the allergic airway inflammation model OVA challenge elicited cellular infiltration surrounding blood vessels and airways (Fig 1c), an increase in total bronchoalveolar lavage (BAL) cells (not shown), and an increase in the frequency of BAL eosinophils (Fig 1d). Likewise, OVA administration to the gut elicited an increase in jejunum eosinophils (Fig 1e). Of note, these data are significant in themselves, as to our knowledge this is the first report of an acute model eliciting a local intestinal eosinophilic response. Increased numbers of jejunal eosinophils were observed both around crypts and within villi, although numbers of villus eosinophils rose more steeply (Supplemental Fig 2a). In addition to local eosinophilic inflammation, all three models elicited a similar level of peripheral eosinophilia with eosinophils accounting for approximately 7-10% of circulating leukocytes, compared to baseline levels of 1-3% (Fig 1f-h).
Epicutaneous or airway allergen exposure increases the number and frequency of eosinophils within allergen non-exposed intestine of allergic mice.
To explore a potential link between acute local skin or lung allergic inflammation on tissue eosinophils within remote intestinal tissues (i.e. skin:gut and lung:gut axes), eosinophils were quantified from tissue sections recovered from the center of the jejunum four days after the final allergen challenge. As shown in Fig 2a, allergen challenge of the skin elicited a near doubling of the total number of eosinophils within allergen non-exposed jejunum. Increased numbers of lamina propria eosinophils were observed both in areas surrounding crypts and along the length of the villi, and increased from an average of 2.16 ± 0.89 eosinophils per crypt:villus unit (cvu) to 3.65 ± 0.90 eosinophils per cvu in sham control versus remote allergen challenged mice, respectively. Likewise, allergen exposure to the lower airways more than doubled the total number of eosinophils within allergen non-exposed jejunum (2.36 ± 0.79 eosinophils per cvu in sham control versus 6.51 ± 2.54 eosinophils per cvu in remote allergen challenged mice, Fig 2b). To confirm jejunum tissues are broadly representative of small intestinal tissue eosinophil counts, in some experiments eosinophils were also quantified from ileal tissues (Supplemental Fig 3). Of note, despite similar levels of peripheral eosinophilia (see Fig 1f-h), allergen exposure to the lung elicited stronger eosinophilic accumulation within the gut than allergen exposure to the skin (1.7- versus 2.8-fold increases elicited by remote skin and airway allergen exposures, respectively). Similar to the pattern of eosinophilic infiltration observed following direct oral gavage of OVA, remote allergen-elicited intestinal eosinophil infiltration favored eosinophil infiltration of villi (Supplemental Fig 2b,c). Analyses of esophageal tissues at this time point did not reveal a substantial increase in eosinophils in either challenge model (not shown).
Figure 2. Epicutaneous or endotracheal allergen challenge elicits an increase in eosinophils within remote, allergen non-exposed intestine.

Eosinophils were quantified from anti-MBP-stained jejunum tissue sections recovered four days after the final allergen challenge to tape-stripped skin (a) or lower airways (b). Frequencies of eosinophils (c), CD4+ T cells (d), CD8+ T cells (e) and dendritic cells (f) were further quantified by flow cytometry of single cell suspensions of intraepithelial (IE, upper panels) or lamina propria (LP, lower panels) compartments recovered from whole small intestine using the following gating strategies: eosinophils, live, CD45+SSChiSiglecFhi; CD4+ T cells, live, CD45+CD3+CD4+cells; CD8+ T cells, live, CD45+CD3+CD8+ cells; and dendritic cells, live, CD45+SiglecF−CD11c+ cells and expressed as percentage of CD45+ leukocytes. (g) Gated eosinophils from IE and LP preparations were further assessed for their expression of surface CD11c (left panel) and CD11b (right panel) that distinguish resident IE and LP eosinophils. Data are expressed as average change in mean fluorescence intensity (MFI) of relevant over isotype control antibodies. (c-f) N≥4. Sham control mice were sensitized with PBS-alum and challenged with PBS. In (a) magnification 200x; scale bars, 100 μm. Error bars show standard deviation. *, p<0.01; **, p<0.001.
Using standard methodologies for isolation of intestinal leukocytes, we previously demonstrated that eosinophils are recovered from both intraepithelial (IE) and lamina propria (LP) compartments 12. To further assess compartmentalization of eosinophils infiltrating intestinal tissues in response to remote (i.e. lung or skin) allergen challenge, LP and IE leukocytes were isolated from whole small intestine preparations four days after the final airway allergen challenge and analyzed by flow cytometry. In support of the histological eosinophil counts, remote allergen challenge increased the frequencies of both IE (Fig 2c, upper) and LP (Fig 2c, lower) eosinophils. In contrast, percentages of CD4+, CD8+, and CD11c+SiglecF− (dendritic) cells in both LP and IE compartments remained static following OVA challenge (Fig 2d-f).
Remote allergen-elicited intestinal eosinophils take on the phenotype of resident intestinal tissue eosinophils.
There is an increasing recognition that tissue resident eosinophils exist in phenotypically distinct subsets that may shed light on their diverse functional roles 12, 14, 15. We previously demonstrated that at baseline intestinal LP and IE eosinophil populations are phenotypically distinguishable, both from blood eosinophils and from each other, based on expression levels of surface markers including the adhesion molecules CD11b and CD11c 12. Analyses of small intestinal leukocytes from sham control and OVA-challenged mice revealed the expected phenotypes demarcating IE and LP eosinophils and demonstrated that remote allergen-elicited eosinophils had acquired tissue-specific surface markers appropriate to their respective localizations within intestinal tissues (Fig 2g,h).
Both epicutaneous and oral allergen exposure elicit eosinophil infiltration into the lung.
It is now recognized that low levels of eosinophils home to and reside within the healthy lung at steady-state 14. Therefore, we queried whether remote exposure of the skin or gut might impact basal numbers of lung tissue eosinophils (i.e. skin:lung and gut:lung axes). Similar to the effect of remote (i.e. skin or lung) allergen exposure on intestinal tissue eosinophils, allergen challenge of either the skin or gut elicited an increase in the number of eosinophils within lung tissue four days after final allergen challenge (Fig 3a,b). In contrast to the sparse distribution of eosinophils throughout the lung parenchyma at baseline (Fig 3a,b left panels), remote allergen-elicited lung eosinophils could be observed localized around venules (Fig 3a,b right upper panels), near airspaces (Fig 3a right lower panel), and in small clusters within the parenchyma (Fig 3b, right lower panel), and were not displaced by lung perfusion. Increased frequencies of eosinophils were detected within the lung as early as one day after allergen challenge (Fig 3c). Similar to the more robust effect elicited by the lung:gut axis compared to the skin:gut axis in promoting intestinal eosinophils, remote allergen challenge by oral gavage elicited a stronger lung eosinophil infiltration than allergen challenge to the skin (3.5 fold versus 1.8 fold increase over baseline following oral gavage or skin challenge, respectively).
Figure 3. Epicutaneous or intragastric allergen challenge elicits an increase in eosinophils within remote, allergen non-exposed lung.

(a,b) Eosinophils were quantified from anti-MBP-stained non-perfused (a) or PBS-perfused (b) lung tissue sections recovered four days after the final allergen challenge to tape-stripped skin (a) or gut (b). Combined data from 2 independent experiments are shown. (d) PBS-perfused lungs from mice 24 hours after final allergen challenge were digested and frequency of lung eosinophils determined by flow cytometry (c). Sham control mice were sensitized with PBS-alum and challenged with PBS. (a, b) Magnification 200x; scale bar, 100 μm. BV, blood vessel; Aw, airway. Error bars show standard deviation. *, p<0.05; **, p ≤ 0.005.
Intestinal tissue eosinophils, but not lung eosinophils, remain elevated at least 11 days post final skin allergen exposure
To begin to investigate the longevity of remote allergen-induced increases in tissue eosinophils, allergen sensitized and skin-challenged mice were sacrificed 11 days after the final allergen challenge and numbers of lung and jejunum tissue eosinophils quantified. As shown in Fig 4, mice continued to exhibit statistically higher numbers of jejunum eosinophils up to eleven days post final allergen exposure to the skin while numbers of lung eosinophils within the same animals had returned to baseline by this time point.
Figure 4. Numbers intestinal eosinophils, but not lung eosinophils, remain elevated up to 11 days following skin allergen challenge.
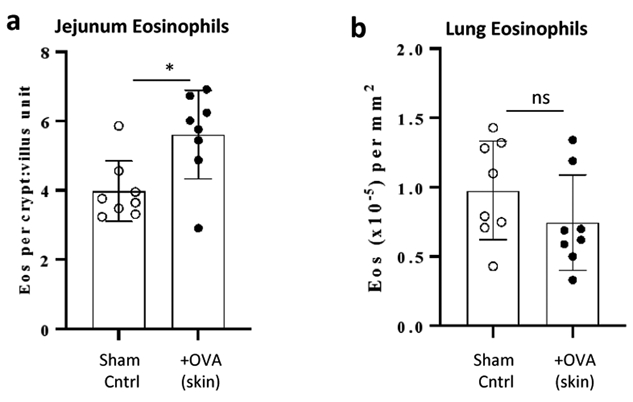
(a) PBS-alum (sham) or OVA-alum sensitized mice received three consecutive epicutaneous challenges with PBS (sham) or OVA as in figure 1A with the modification that all mice were sacrificed on day 34 (eleven days after the final skin allergen challenge). Intestinal (a) and lung (b) eosinophils were quantified from anti-MBP stained tissue sections. *, p<0.01.
Oral allergen-elicited lung-infiltrating eosinophils exhibit an inflammatory phenotype and are associated with airway mucus production
As described in the Introduction the phenotype of resident lung eosinophils resembles blood eosinophils and they are believed to serve homeostatic functions at baseline 14, 18. Within the context of an allergic airways response, recruited lung eosinophils take on an inflammatory phenotype, including the induction of CD11c expression and upregulation of the sialic acid binding lectin Siglec F 19. To determine the phenotype of lung-infiltrating eosinophils elicited by remote allergen exposure PBS-perfused lungs were digested and eosinophils phenotyped by flow cytometry. Confirming our histological findings, oral allergen exposure increased the frequency of eosinophils within lung tissues (Fig 5a,b). Control mice exhibited the expected single population of resident, homeostatic eosinophils (hEos), defined as CD45+SSChiSiglecFloCD11c− cells (Fig 5a, left panel). In contrast, following intragastric allergen exposure two distinct populations of lung tissue eosinophils were observed, the first identical to resident homeostatic lung eosinophils of control mice, and the second exhibiting the surface marker phenotype of activated, “inflammatory” lung eosinophils (iEos), i.e. CD45+SSChiSiglecFintCD11cint (Fig 5a, right panel). Of note, the appearance of iEos in the remote allergen challenged mice was not accompanied by a loss of hEos, suggesting hEos are not converting to iEos within the lung tissue (Fig 5c).
Figure 5. Oral allergen-elicited lung eosinophils exhibit an inflammatory phenotype and their accumulation is accompanied by increased mucus production.

Four days after final oral challenge mice were sacrificed and lungs perfused with PBS. Lung tissue was digested and disaggregated and frequencies and phenotypes of lung tissue eosinophils analyzed by flow cytometry (a-c) or formalin fixed, paraffin embedded and stained with PAS to assess mucus secretion (d-f). (a-c) Lung eosinophils were identified as live, CD45+SSChiSiglecF+ cells after gating out CD11chi alveolar macrophages. Within the gated population eosinophils were further categorized as homeostatic (hEos, SiglecFloCD11c−) or inflammatory (iEos, SiglecFhiCD11cint) (a,c). Sham control mice were sensitized with PBS-alum and challenged with PBS. Magnification 100x (d,e); 200x (di, ei). Scale bar, 200 μm. *, p<0.05; **, p ≤ 0.005, ***, p ≤ 0.0005. P-values determined by student’s ttest (b, f) or two-way ANOVA with Tukey’s multiple comparisons test (c).
Inflammatory eosinophils infiltrating the lungs within the context of allergic airway inflammation promote goblet cell hyperplasia and enhanced mucus production 20, 21. Therefore, we queried whether oral allergen-elicited inflammatory lung eosinophils might be accompanied by an increase in mucus secretion. To test this hypothesis tissue sections were stained with periodic acid-Schiff (PAS) stain to detect carbohydrates. Although none of the lung sections recovered from sham control mice (N=16) exhibited PAS staining, lung sections from 12 of 16 (75%) of the oral allergen challenged mice exhibited evidence of airway mucus secretion (Fig 5d-f). Quantification of mucus positive airways across all mice confirms a statistically significant induction of airway mucus in oral allergen-challenged mice (Fig 5f). Intriguingly, airway mucus secretion was not readily detected in lungs of mice following skin challenge (not shown).
Oral or epicutaneous allergen exposure primes the airways for a more robust allergic response to inhaled house dust mite
Our findings that oral allergen challenge elicits inflammatory eosinophils within the lung associated with mucus secretion suggested that remote allergen exposure might predispose for exacerbated airways inflammatory reactions to inhaled allergens. To test this hypothesis we utilized the acute oral allergen challenge model with the modification that five days after the final oral OVA challenge mice received intranasal administrations of house dust mite (HDM), a common allergen implicated in human asthma (Fig 6a). Compared to sham-treated controls, OVA-sensitized mice challenged with intragastric OVA exhibited more robust allergic airways inflammation in response to subsequent intranasal administrations of house dust mite, as evidenced histologically by cellular infiltrations (Fig 6b) and an overall increase in the total number of cells recovered from the bronchoalveolar lavage (BAL), comprised of a higher frequency and total number of eosinophils (Fig 6c).
Figure 6. Remote allergen challenge primes for respiratory allergic disease.

(a) Experimental timeline. OVA- or sham-sensitized mice were challenged with OVA or PBS, respectively, by oral gavage (b,c) or via skin (d) as described in Fig 1a. Beginning four days after final allergen (or PBS) challenge, mice received 6 intranasal instillations of house dust mite (HDM) or PBS vehicle control, spaced over 2 weeks as indicated. Mice were sacrificed one day following the final HDM instillation. (b) Representative lung histology sections stained with H&E. Magnification 200x, scale bar 200 μm. (c,d) Assessment of BAL fluid revealed an increase in total BAL cells, and in BAL eosinophils (expressed as a percentage of total BAL leukocytes and as total eosinophils) in mice receiving prior OVA challenge to the gut (c) or skin (d). *, p ≤ 0.05; **, p ≤ 0.005.
Although parallel experiments using india ink confirm dye is confined to the stomach following the oral gavage technique (Supplemental Fig 4c), we cannot completely rule out the possibility of microaspiration in individual mice during oral gavage, thereby inadvertently delivering allergen to the airways 22. Therefore, to confirm that the remote allergen-elicited increased sensitivity to a subsequent inhaled allergen was not caused by microaspiration, and to confirm applicability to both the gut:lung axis as well as the skin:lung axis, we subjected the acute skin challenge model to subsequent HDM exposure. Similar to orally-challenged mice, OVA skin-challenged mice exhibited a more robust eosinophilic airway response to inhaled HDM than sham treated control mice (Fig 6d).
DISCUSSION
Bidirectional, progressive relationships link allergic diseases of the skin (e.g. atopic dermatitis), respiratory tract (e.g. asthma, allergic rhinitis), and gut (e.g. food allergy, eosinophilic gastrointestinal diseases (EGIDs)). However, impacts of local allergic inflammation on the cellular landscape of remote mucosal organs along the skin:lung:gut axes are unknown. Here we utilized acute mouse models of allergic dermatitis, allergic airway inflammation and food allergy to investigate impacts of acute allergic reactions on the frequency and phenotype of tissue eosinophils within remote, allergen non-exposed organs. Our data demonstrate that in allergen-sensitized mice epicutaneous, airway or intragastric allergen exposure elicits not only a local eosinophilic inflammation at the site of allergen challenge, but also increases the number and frequency of activated tissue eosinophils within allergen non-exposed intestine and lung. Accumulation of activated eosinophils within the lung following remote epicutaneous or oral allergen challenge was associated with an exacerbated allergic airways response to subsequent inhaled HDM. These data demonstrate for the first time that local allergic inflammatory reactions alter the composition of tissue eosinophil compartments within remote mucosal organs, offering new insights into the phenotype and plasticity of mucosal tissue eosinophils and the cross-talk between organs along the skin:lung:gut axes that drives progression of allergic diseases.
Administration of OVA either to tape-stripped skin or directly into the lower airways of OVA-sensitized mice increased both the number and frequency of intestinal tissue eosinophils. Both acute disease models induced a concurrent increase in circulating eosinophils. Since eosinophils exiting the bone marrow express the gut-homing integrin α4β7 and home naturally to the intestinal tract at baseline 23, it is tempting to speculate an increased level of circulating eosinophils as sufficient for remote allergen-induced intestinal eosinophil recruitment. However, levels of circulating eosinophils do not directly correlate with tissue-accumulated eosinophils since, despite similar levels of circulating eosinophils, allergen exposure to the lung elicited stronger eosinophil accumulation within the jejunum than did epicutaneous allergen exposure. Moreover, peripheral eosinophilia by itself does not always correspond with intestinal eosinophil numbers as demonstrated by Mishra et al wherein eosinophilic airway inflammation elicited by repeated inhalation of Aspergillus fumigatus was accompanied by an increase in peripheral eosinophilia with no impact on numbers of intestinal eosinophils 24. Of note, the Mishra et al studies were performed in mice on the 129 SvEv background. Further studies are warranted to define mechanisms that regulate eosinophil infiltration and accumulation within remote organs, including studies that parse relative contributions of mouse strain, nature of the allergen and mode of sensitization (i.e. sensitization via mucosal exposure versus systemic sensitization). Indeed our findings are in agreement with Bui et al who reported an increase in intestinal eosinophils following aerosolized OVA challenge of systemically-sensitized BALB/c mice (although ingestion of inhaled allergen can’t be ruled out in the Bui et al study) 25. Further comprehensive studies are underway in our mouse models of the allergic march to identify systemic networks of cytokines and their tissue targets that shape resident tissue eosinophil compartments in remote organs. Recent studies suggest signaling axes involving epithelial cell-derived TSLP, IL-33 and/or IL-25 and mucosal tissue ILC2 and Tuft cells may play important, yet complex roles 26, 27.
Biological significance of the remote allergen-induced influx of intestinal eosinophils remains to be determined. Approximately 64% of patients with eosinophilic esophagitis (EoE) 28, and 39-42% of patients with other (non-EoE) eosinophilic gastrointestinal diseases 29 (EGlDs; i.e. eosinophilic gastroenteritis, eosinophilic colitis) exhibit a co-existing or previous atopic disease; it is therefore plausible that remote allergen-elicited impacts on intestinal eosinophil populations might prime for the development of EGlDs. The presence of intraepithelial eosinophils is considered a negative prognostic indicator in EGlDs; therefore the predominance of remote allergen-elicited villus-migrated eosinophils in the lamina propria (Supplemental Fig 2) taken together with an increased frequency of IE-associated eosinophils (Fig 2c) might further support a connection between remote allergen exposure and EGID susceptibility and/or severity. A similar scenario may also help to explain the observed increased incidence of EoE in food allergy patients receiving repetitive allergen administrations through oral immunotherapy (OIT) 30, 31. Of note, in our acute models of skin or lung allergen challenge we did not observe substantial numbers of eosinophils infiltrating the esophagus. It will be interesting to determine whether a chronic model of allergen exposure to the skin or lungs might also elicit esophageal eosinophils. Indeed, epicutaneous routes of repetitive allergen exposure have proven a successful mechanism of sensitization for mouse models of eosinophilic esophagitis (EoE) 32, 33.
Here we also show that epicutaneous or intragastric allergen exposure modulates lung tissue eosinophil populations as early as one day post final allergen challenge, demonstrating crosstalk along the skin:lung and gut:lung axes. Notably, there were distinct differences in the longevity of remote allergen-elicited eosinophils within airway and intestinal tissues. Analyses of intestinal and lung eosinophil populations eleven days after three consecutive skin challenges revealed a sustained increase in intestinal eosinophils, in contrast to lung eosinophil numbers in the same animals, which had returned to baseline by this time point.
At baseline resident lung eosinophils express a phenotype resembling blood eosinophils (i.e. CD11c−CD62L+SiglecFlo/int) and are believed to function in homeostatic roles 14, 18. Our data shown here demonstrate that eosinophils infiltrating the lung in response to skin or gut allergen challenge exhibit an activated phenotype, including induced expression of CD11c and upregulation of SiglecF, thereby donning the phenotype exhibited by inflammatory lung tissue or BAL eosinophils recovered in active allergic airways disease 14, 19. Moreover, accumulation of oral allergen-elicited lung eosinophils was accompanied by airway mucus production. These observations may offer mechanistic insights into clinical observations of respiratory manifestations of gastrointestinal allergies, e.g. wheezing and bronchospasm 34-36.
Finally, we were interested in understanding whether acute local allergen exposure might prime remote mucosal organs for enhanced susceptibility to new allergic diseases. Therefore, we exposed epicutaneous or oral allergen challenged mice to inhaled HDM, an allergen antigenically distinct from OVA and a significant cause of human asthma. Unlike sensitization with a soluble protein antigen such as OVA which requires an adjuvant to elicit an immune response, HDM is inherently immunogenic, presumably owing to its protease activity. Shown here, prior intragastric or epicutaneous exposure of sensitized mice to OVA primed the airways for enhanced susceptibility to HDM, as evidenced by cellular infiltrations surrounding blood vessels and airways, and BAL eosinophilia. These data may offer mechanistic insights into polysensitization and progressive allergic disease manifestations associated with patients undergoing the allergic march.
Data shown here demonstrate bidirectional relationships between local acute gastrointestinal, skin or respiratory allergic reactions and the phenotype and frequency of eosinophils within remote mucosal tissues that were further associated with organ priming for allergic inflammation in response to an antigenically distinct allergen. Collectively these findings provide new insights into the plasticity of mucosal tissue eosinophil populations and organ cross-talk relevant to allergic disease progressions.
MATERIALS AND METHODS
Animals
Studies received prior approvals from BIDMC or CUSOM Institutional Animal Care and Use Committees. 6-10 week old BALB/c mice were from Charles River Laboratory (Wilmington, MA) and maintained under conventional specific pathogen free housing conditions with food and water provided ad libitum. Experiments were performed with age- and gender-matched female and male mice.
OVA Allergen Sensitization
Mice were sensitized by intraperitoneal injections on days 0, 7 and 14 with 50ug OVA (Ovalbumin, Sigma #A2512) and 0.4 mg alum (Imject Alum, Thermo Scientific #77161) in 100μL sterile PBS. Control mice were sham-sensitized with alum in PBS. Following sensitization mice were challenged according to one of the methods below.
Allergen Challenge Methods
Pulmonary allergen challenges were performed using a non-invasive endotracheal spray application originally developed for vaccine delivery systems 37 and modified here for allergen delivery. Mice were anesthetized with ketamine (50 mg/mL) and xylazine (5 mg/mL) before positioning on a Mouse Intubation Platform (MIP). A MicroSprayer® aerosolizer (model 1A-1C) with high pressure syringe (Penn Century®, Wyndmoor, PA) was inserted approximately 1.25 cm to intubate mice and deliver 50 μL of OVA (20 μg) in sterile PBS or PBS alone as an air-free aerosol directly into the trachea at approximately the level of the carina (Supplemental Fig 4a). In some experiments 0.6-6.0 % India ink solutions were substituted for the antigen and lungs harvested for evaluation of the depth and pattern of ink staining 20 min after inoculation (Supplemental Fig 4b). To confirm administered aerosol was restricted to lower airways, the oral cavity, esophagus, stomach (including contents), and small bowel were examined and found to be void of India ink (not shown). Of note, it is plausible that self-clearing mechanisms associated with the respiratory mucociliary escalator may, as in humans, deliver a fraction of the airway-deposited allergen to intestinal tissues over time.
Epicutaneous allergen challenge was performed as described 33. Briefly, dorsal skin of anesthetized mice was shaved and tape-stripped 6 times with Tegaderm (3m Healthcare, St. Paul, MN). 100μg of OVA in 100μL of PBS or PBS alone was applied directly onto tape-stripped skin and covered with a piece of sterile gauze (Covidien, Mansfield, MA) secured to the skin with Tegaderm. Mice were monitored 24 hours after allergen exposure to ensure gauze and Tegaderm strips remained intact, at which point the gauze was removed to avoid mice ingesting allergen-coated gauze during grooming.
Intragastric allergen challenged was achieved by oral gavage (22-gauge needle) with 50 mg of OVA (Sigma-Aldrich, St. Louis, MO) in 250μL of sterile PBS. Parallel experiments with india ink dye confirm dye is confined to the stomach following the oral gavage technique, with no evidence of dye staining along the length of the esophagus (Supplemental Fig 4c).
Recovery of tissues
At indicated time points mice were euthanized, cardiac blood collected for cell differential counts and lung, small intestine and skin samples collected into ice cold PBS. For histological analyses skin samples from the mid-dorsal back, longitudinal sections from the middle of the left lung lobe and rings of small intestinal tissues were fixed in 10% neutral buffered formalin and embedded in paraffin. 4 μm sections were stained with H&E for routine analyses or PAS (lung tissues) for analysis of mucus.
Eosinophil Enumeration by histology
Eosinophils were identified within lung and intestinal tissues by immunohistochemistry staining after antigen retrieval with an eosinophil specific rat anti-mouse major basic protein monoclonal antibody (MBP, provided by Dr. Beth Jacobsen and the Lee Laboratory, Mayo Clinic, Arizona) as previously described 38, followed by counterstaining with methyl green. In some parallel experiments eosinophils were enumerated from slides stained with Fast Green (0.2% in 70% ethanol; Sigma-Aldrich, St. Louis, MO) and Neutral Red (0.5% in water, Fluka, St. Louis, MO). Preliminary studies confirmed that although MBP immunohistochemistry increased the overall detection of eosinophils across all groups, statistical comparisons between groups were similar regardless of staining method used (Supplemental Fig 5). For all studies eosinophils were quantified from viewer-blinded slides.
Determining eosinophil frequency by flow cytometry
To determine frequency of eosinophils within whole tissues, lung or small intestine were digested to achieve single cell suspensions and assessed by flow cytometry. For lung digests, lungs were perfused with 10 mL of PBS injected through the right ventricle prior to recovery. Minced lung tissues were digested with 175 U/mL collagenase IV (Worthington Biomedical Corporation), mechanically disrupted and passed through 40 μm filters. Residual red blood cells were lysed with BD PharmLyse (BD Biosciences) per manufacturer’s instructions. For intestinal tissue digests, small intestine was harvested as described 12. Briefly, intestinal fragments (approximately 5 mm) were incubated in DTE buffer (10% fetal bovine serum, 1mM HEPES, 2.5mM NaHCO3, 1mM DTT in HBSS−/−) at 37°C with shaking to release IE cells. Tissue fragments were then incubated in EDTA-containing buffer (1X penicillin/streptomycin, 1.3mM EDTA in HBSS−/−) at 37°C with shaking to release epithelial cells, which were discarded. Tissue fragments were then washed and incubated in 1 U/mL collagenase 1 at 37°C for 30 minutes with shaking. Digested tissue was passed through a 70μm strainer and collected as LP fraction. IE and LP cell pellets were resuspended in 44% Percoll, underlayed with 67% Percoll, and centrifuged for 20 min at 1000g. Enriched viable IE or LP cells were collected from the interphase and stained for flow cytometry. LIVE/DEAD Fixable Aqua Dead Cell Stain Kit (Invitrogen, Grand Island, NY), CD45-PE-Cy7 (30-F11), CD4-APC (RM 4-5) and Siglec F-PE (E50-2440) from BD Biosciences (San Jose, CA), and CD11c-FITC or CD11c-APC (N418) and CD8-FITC (53-6.7) from BioLegend (San Diego, CA) were used. Data were acquired using LSR II (BD Biosciences, San Jose, CA) or Gallios flow cytometers and analyzed with FlowJo analysis software (Tree Star, Ashland, OR).
House Dust Mite-Induced Allergic Airway Inflammation
Following OVA sensitization and challenge mice received six intranasal exposures spread over two weeks (see Fig 6a) to D. pteronyssinus HDM extract (Stallergenes Greer Laboratories, Lenoir, NC), each intranasal inoculum containing 3 μg of HDM (based on protein weight) in 35μL sterile PBS. Intranasal HDM was administered to anesthetized mice which were held in the supine position for 10 seconds following inhalation. Mice were sacrificed one day after final HDM administration, and BAL fluid collected into cold PBS containing 2.5% bovine serum albumin (BSA, Sigma #A7030).
Statistical Analyses
Data are presented as mean ± standard deviation for each experimental group. Statistical analyses were performed using one-way analysis of variance (ANOVA) or two-tailed Student’s ttest as indicated.
Supplementary Material
ACKNOWLEDGEMENTS
Authors thank Michiko Oyoshi, PhD, Boston Children’s Hospital for technical advice in establishing the skin challenge model. This work was supported by NIH R01AI121186 to LAS.
Footnotes
DISCLOSURE
Authors have no conflicts of interest to declare.
References
- 1.Bantz SK, Zhu Z, Zheng T. The Atopic March: Progression from Atopic Dermatitis to Allergic Rhinitis and Asthma. J Clin Cell Immunol. 2014;5(2). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Guilbert TW, Morgan WJ, Zeiger RS, Bacharier LB, Boehmer SJ, Krawiec M, et al. Atopic characteristics of children with recurrent wheezing at high risk for the development of childhood asthma. J Allergy Clin Immunol. 2004;114(6):1282–7. [DOI] [PubMed] [Google Scholar]
- 3.Gustafsson D, Sjoberg O, Foucard T. Development of allergies and asthma in infants and young children with atopic dermatitis--a prospective follow-up to 7 years of age. Allergy. 2000;55(3):240–5. [DOI] [PubMed] [Google Scholar]
- 4.Kapoor R, Menon C, Hoffstad O, Bilker W, Leclerc P, Margolis DJ. The prevalence of atopic triad in children with physician-confirmed atopic dermatitis. J Am Acad Dermatol. 2008;58(1):68–73. [DOI] [PubMed] [Google Scholar]
- 5.Kulig M, Bergmann R, Klettke U, Wahn V, Tacke U, Wahn U. Natural course of sensitization to food and inhalant allergens during the first 6 years of life. J Allergy Clin Immunol. 1999;103(6):1173–9. [DOI] [PubMed] [Google Scholar]
- 6.Martinez FD, Wright AL, Taussig LM, Holberg CJ, Halonen M, Morgan WJ. Asthma and wheezing in the first six years of life. The Group Health Medical Associates. N Engl J Med. 1995;332(3):133–8. [DOI] [PubMed] [Google Scholar]
- 7.Novembre E, Cianferoni A, Lombardi E, Bernardini R, Pucci N, Vierucci A. Natural history of "intrinsic" atopic dermatitis. Allergy. 2001;56(5):452–3. [DOI] [PubMed] [Google Scholar]
- 8.Ricci G, Patrizi A, Baldi E, Menna G, Tabanelli M, Masi M. Long-term follow-up of atopic dermatitis: retrospective analysis of related risk factors and association with concomitant allergic diseases. J Am Acad Dermatol. 2006;55(5):765–71. [DOI] [PubMed] [Google Scholar]
- 9.Spergel JM. From atopic dermatitis to asthma: the atopic march. Ann Allergy Asthma Immunol. 2010;105(2):99–106; quiz 7-9, 17. [DOI] [PubMed] [Google Scholar]
- 10.van der Hulst AE, Klip H, Brand PL. Risk of developing asthma in young children with atopic eczema: a systematic review. J Allergy Clin Immunol. 2007;120(3):565–9. [DOI] [PubMed] [Google Scholar]
- 11.Powell N, Walker MM, Talley NJ. Gastrointestinal eosinophils in health, disease and functional disorders. Nat Rev Gastroenterol Hepatol. 2010;7(3):146–56. [DOI] [PubMed] [Google Scholar]
- 12.Xenakis JJ, Howard ED, Smith KM, Olbrich CL, Huang Y, Anketell D, et al. Resident intestinal eosinophils constitutively express antigen presentation markers and include two phenotypically distinct subsets of eosinophils. Immunology. 2018;154(2):298–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Carlens J, Wahl B, Ballmaier M, Bulfone-Paus S, Forster R, Pabst O. Common gamma-chain-dependent signals confer selective survival of eosinophils in the murine small intestine. J Immunol. 2009;183(9):5600–7. [DOI] [PubMed] [Google Scholar]
- 14.Mesnil C, Raulier S, Paulissen G, Xiao X, Birrell MA, Pirottin D, et al. Lung-resident eosinophils represent a distinct regulatory eosinophil subset. J Clin Invest. 2016;126(9):3279–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Percopo CM, Brenner TA, Ma M, Kraemer LS, Hakeem RM, Lee JJ, et al. SiglecF+Gr1hi eosinophils are a distinct subpopulation within the lungs of allergen-challenged mice. J Leukoc Biol. 2017;101(1):321–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rose WA 2nd, Okragly AJ, Patel CN, Benschop RJ. IL-33 released by alum is responsible for early cytokine production and has adjuvant properties. Sci Rep. 2015;5:13146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Johnston LK, Bryce PJ. Understanding Interleukin 33 and Its Roles in Eosinophil Development. Front Med (Lausanne). 2017;4:51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marichal T, Mesnil C, Bureau F. Homeostatic Eosinophils: Characteristics and Functions. Front Med (Lausanne). 2017;4:101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Abdala Valencia H, Loffredo LF, Misharin AV, Berdnikovs S. Phenotypic plasticity and targeting of Siglec-F(high) CD11c(low) eosinophils to the airway in a murine model of asthma. Allergy. 2016;71(2):267–71. [DOI] [PubMed] [Google Scholar]
- 20.McBrien CN, Menzies-Gow A. The Biology of Eosinophils and Their Role in Asthma. Front Med (Lausanne). 2017;4:93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dunican EM, Elicker BM, Gierada DS, Nagle SK, Schiebler ML, Newell JD, et al. Mucus plugs in patients with asthma linked to eosinophilia and airflow obstruction. J Clin Invest. 2018;128(3):997–1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hoggatt AF, Hoggatt J, Honerlaw M, Pelus LM. A spoonful of sugar helps the medicine go down: a novel technique to improve oral gavage in mice. J Am Assoc Lab Anim Sci. 2010;49(3):329–34. [PMC free article] [PubMed] [Google Scholar]
- 23.Jung Y, Rothenberg ME. Roles and regulation of gastrointestinal eosinophils in immunity and disease. J Immunol. 2014;193(3):999–1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mishra A, Hogan SP, Lee JJ, Foster PS, Rothenberg ME. Fundamental signals that regulate eosinophil homing to the gastrointestinal tract. J Clin Invest. 1999;103(12):1719–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bui LK, Hayashi T, Nakashima T, Horii Y. Eosinophilic venulitis in the small intestines in a mouse model of late asthma. Inflammation. 2011;34(5):499–508. [DOI] [PubMed] [Google Scholar]
- 26.Leyva-Castillo JM, Galand C, Kam C, Burton O, Gurish M, Musser MA, et al. Mechanical Skin Injury Promotes Food Anaphylaxis by Driving Intestinal Mast Cell Expansion. Immunity. 2019;50(5):1262–75 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li M Current evidence of epidermal barrier dysfunction and thymic stromal lymphopoietin in the atopic march. Eur Respir Rev. 2014;23(133):292–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Benninger MS, Strohl M, Holy CE, Hanick AL, Bryson PC. Prevalence of atopic disease in patients with eosinophilic esophagitis. Int Forum Allergy Rhinol. 2017;7(8):757–62. [DOI] [PubMed] [Google Scholar]
- 29.Jensen ET, Martin CF, Kappelman MD, Dellon ES. Prevalence of Eosinophilic Gastritis, Gastroenteritis, and Colitis: Estimates From a National Administrative Database. J Pediatr Gastroenterol Nutr. 2016;62(1):36–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lucendo AJ, Arias A, Tenias JM. Relation between eosinophilic esophagitis and oral immunotherapy for food allergy: a systematic review with meta-analysis. Ann Allergy Asthma Immunol. 2014;113(6):624–9. [DOI] [PubMed] [Google Scholar]
- 31.Semancik E, Sayej WN. Oral immunotherapy for peanut allergy induces eosinophilic esophagitis: three pediatric case reports. Pediatr Allergy Immunol. 2016;27(5):539–41. [DOI] [PubMed] [Google Scholar]
- 32.Akei HS, Mishra A, Blanchard C, Rothenberg ME. Epicutaneous antigen exposure primes for experimental eosinophilic esophagitis in mice. Gastroenterology. 2005;129(3):985–94. [DOI] [PubMed] [Google Scholar]
- 33.Venturelli N, Lexmond WS, Ohsaki A, Nurko S, Karasuyama H, Fiebiger E, et al. Allergic skin sensitization promotes eosinophilic esophagitis through the IL-33-basophil axis in mice. J Allergy Clin Immunol. 2016;138(5):1367–80 e5. [DOI] [PubMed] [Google Scholar]
- 34.Liu AH, Jaramillo R, Sicherer SH, Wood RA, Bock SA, Burks AW, et al. National prevalence and risk factors for food allergy and relationship to asthma: results from the National Health and Nutrition Examination Survey 2005-2006. J Allergy Clin Immunol. 2010;126(4):798–806 e13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sampson HA. Update on food allergy. J Allergy Clin Immunol. 2004;113(5):805–19; quiz 20. [DOI] [PubMed] [Google Scholar]
- 36.James JM. Respiratory manifestations of food allergy. Pediatrics. 2003;111(6 Pt 3):1625–30. [PubMed] [Google Scholar]
- 37.Bivas-Benita M, Zwier R, Junginger HE, Borchard G. Non-invasive pulmonary aerosol delivery in mice by the endotracheal route. Eur J Pharm Biopharm. 2005;61(3):214–8. [DOI] [PubMed] [Google Scholar]
- 38.Ochkur SI, Jacobsen EA, Protheroe CA, Biechele TL, Pero RS, McGarry MP, et al. Coexpression of IL-5 and eotaxin-2 in mice creates an eosinophil-dependent model of respiratory inflammation with characteristics of severe asthma. J Immunol. 2007;178(12):7879–89. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.