

Graphical abstract
Keywords: MicroRNA, Extracellular vesicles, Host-parasite interactions, Trichinella spiralis
Highlights
-
•
Trichinella spiralis adults and larvae secrete abundant small (18–40 nt) RNAs including microRNAs (miRNAs).
-
•
Both adults and larvae secrete similar quantities of exosomes.
-
•
Adult secreted miRNAs are contained within exosomes.
-
•
Surprisingly, larvae, which live intracellularly within muscle cells secrete miRNAs which are not contained within exosomes.
-
•
Direct release of unencapsulated miRNAs by intracellular larvae might be a way to influence host cell gene expression.
Abstract
Many organisms, including parasitic nematodes, secrete small RNAs into the extracellular environment, largely encapsulated within small vesicles. Parasite-secreted material often contains microRNAs (miRNAs), raising the possibility that they might regulate host genes in target cells. Here we characterise secreted RNAs from the parasitic nematode Trichinella spiralis at two different life stages. We show that adult T. spiralis, which inhabit intestinal mucosa, secrete miRNAs within vesicles. Unexpectedly, T. spiralis muscle stage larvae, which live intracellularly within skeletal muscle cells, secrete miRNAs that appear not to be encapsulated. Notably, secreted miRNAs include a homologue of mammalian miRNA-31, which has an important role in muscle development. Our work therefore suggests that RNAs may be secreted without encapsulation in vesicles, with implications for the biology of T. spiralis infection.
1. Introduction
It is becoming increasingly clear that a variety of RNA molecules have a life outside the cell (Sarkies and Miska, 2014). RNA has been found in many different extracellular fluids (Liu et al., 2019). Often, RNA molecules are encapsulated in membrane-bound extracellular vesicles (EVs) such as exosomes and microvesicles, which protect them from nuclease digestion. Of the categories of RNA observed, one of the most abundant appears to be small (18–30 nucleotides (nt)) RNAs, including microRNAs (miRNAs) which, within cells, are critical regulators of gene expression (Bartel, 2018). Extracellular miRNAs are often represented in different proportions to miRNAs within the cells from which they originate, indicating that specific selection of certain miRNAs for export might occur (Turchinovich et al., 2016).
Whilst the existence of extracellular miRNAs is clear, most aspects of their generation and mode of transport are still poorly understood. It is not clear how specific miRNAs are targeted for export, which cytoplasmic proteins are responsible for sorting miRNAs into vesicles, or the protein factors that bind miRNAs outside cells (Sarkies and Miska, 2014). There is also little understanding of the function of extracellular miRNAs. In principle, miRNAs might be taken up by target cells and act to regulate gene expression, thus contributing to cell-to-cell communication (Turchinovich et al., 2016). However, significant challenges remain for this model. The density of miRNAs within EVs is somewhat low, meaning that high numbers might have to be delivered to target cells. Moreover, there is limited evidence that extracellular miRNAs are bound to the Argonaute proteins that are required for their operation (Gibbings et al., 2009), and it is thus unclear how they would be able to function in host gene expression pathways.
Recently, parasitic nematodes have emerged as an attractive system in which to study the potential roles of extracellular miRNAs (Buck et al., 2014, Coakley et al., 2015). Parasitic nematodes secrete abundant material in order to develop and complete their life cycle within their hosts, often involving manipulation of host immunity or physiology (Coakley et al., 2016). The secreted material from many nematode species is enriched for small RNAs including miRNAs (Buck et al., 2014). Communication with the host via secreted miRNAs might be a key mechanism in parasitic infection, as the high conservation of miRNA sequences and mechanism across animals (Bartel, 2018) would facilitate their incorporation into host gene expression pathways. Additionally, the crucial importance of many miRNA-target interactions in development and cellular homeostasis (Gebert and MacRae, 2019) would make it difficult for host cells to evolve resistance mechanisms. Many examples have been cited whereby exposure to parasite-derived EVs led to modified immune responses in mammalian cells (Coakley et al., 2017). EVs secreted by Heligmosomoides polygyrus containing miRNAs and other small RNAs were taken up by murine epithelial cells in vitro, resulting in altered gene expression, and intranasal delivery of parasite exosomes modified the pulmonary immune response to a co-administered fungal extract (Buck et al., 2014). Nevertheless, there is still little concrete evidence that these pathways occur in vivo, and particularly challenging is the question of whether secreted vesicles can be taken up by target cells at a high enough rate to ensure delivery of miRNAs at robust levels, although parasite-derived miRNAs were recently identified in macrophages from the pleural/peritoneal cavity of Mongolian jirds infected with the filarial nematode Litomosoides sigmodontis (Quintana et al., 2019).
Here we describe the nematode Trichinella spiralis as a new model to study extracellular miRNAs. Trichinella spiralis is a parasite that infects many mammalian species including humans. Its life cycle is unusual amongst parasitic nematodes because it has both intracellular and extracellular stages. In the intestinal phase, infective larvae penetrate and migrate through epithelial cell sheets (ManWarren et al., 1997, Wright, 1979), and following development to adult worms, release L1s which migrate via the lymphatics and vascular system to skeletal muscle, where they invade myofibres and develop to form a specialised intracellular niche known as the nurse cell. Interestingly, nurse cell formation involves perturbation of the normal gene expression programme of muscle cells, which results in cell cycle re-entry, arrest at apparent G2/M phase and downregulation of several key markers of differentiated muscle (Jasmer, 1993). How the parasite influences gene expression and subsequent alterations in skeletal muscle phenotype is poorly understood.
We reasoned that the existence of Trichinella in such radically different environments provided an interesting opportunity to study the potential role of RNA in extracellular communication. In particular, release of RNA from intracellular parasites might be expected to facilitate a high local concentration of parasite miRNAs which otherwise would be difficult to achieve through uptake of vesicles by target cells. We therefore isolated secreted material from adult and larval T. spiralis, and investigated the RNA content. Whilst adult T. spiralis releases miRNAs protected from RNase digestion, larvae isolated from muscle cells secrete unprotected small RNAs, suggesting they might be released directly into target cells. Amongst the miRNAs secreted by muscle stage larvae (MSL), we identified a homologue of the mammalian miR-31, which plays a key role in development and regeneration of skeletal muscle. Our results suggest that secretion of miRNAs without vesicular protection may be quite prevalent, and could be particularly relevant for regulation of host gene expression by intracellular pathogens such as T. spiralis.
2. Materials and methods
2.1. Parasite isolation and culture
This study was licensed by and performed under the UK Home Office Animals (Scientific Procedures) Act Personal Project Licence number 70/8193: ‘Immunomodulation by helminth parasites’. Adult parasites were recovered from Sprague-Dawley rats 6–7 days p.i. by sedimentation in a Baermann apparatus containing segments of small intestine, and (infective) MSL were recovered from digested muscle 2 months p.i. as previously described (Arden et al., 1997). Parasites were cultured in serum-free medium for up to 72 h as described (Arden et al., 1997), secreted products were collected daily and centrifuged at 2000g for 10 min, then supernatants were cleared through 0.2 µm filters and pooled.
2.2. Isolation and analysis of extracellular vesicles
Culture medium was first centrifuged at 10,000g for 30 min to clear cell debris, apoptotic bodies and large vesicles. The supernatant was then centrifuged at 100,000g for 90 min in polyallomer tubes at 4 °C in a SW40 rotor; the pellet was washed in PBS and recentrifuged under the same conditions. For Nanoparticle Tracking Analysis (NTA), the pellet was resuspended in 400 µl of PBS and analysed as previously described (Faruqu et al., 2018). For electron microscopy, the pellet was fixed in 2% paraformaldehyde, adsorbed onto copper Electron Microscopy (EM) grids, washed with PBS, stained with uranyl acetate and viewed in a Tecnai T12 Spirit Electron Microscope with images captured on a TVIPS TemCam-F216 CCD camera.
2.3. RNase protection assay
Parasite-secreted products were passed through a 0.2 µm filter and concentrated in 3 kDa molecular weight cutoff vivaspin columns, washed once in PBS, then thrice in RNase buffer (PBS, 5 mM EDTA, 300 mM NaCl, 10 mM Tris–Cl pH 7.5). Aliquots (150 µl) of concentrated secreted products were digested with different concentrations of RNase A/RNase T1 cocktail (2 µg of RNase A/5 U of RNase T1, 40 ng of RNase A/0.1 U RNase T1, no enzyme control) for 1 h at 37 °C, and digestion was terminated by addition of 700 µl of Trizol. RNA was isolated from each sample and resolved on a 2100 Bioanalyser using the Agilent small RNA kit in accordance with the manufacturer’s instructions.
2.4. Small RNA sequencing
RNA was isolated from parasite secreted products using TRIzol, and libraries were prepared using the Illumina TruSeq small RNA preparation kit. Sensitivity to RNase was again assessed by digesting some RNA samples with 4 µg ml−1 RNase A/10 U ml−1 RNase T1 at 37 °C for 1 h prior to library preparation. Sequencing was performed by the Medical Research Council London Institute of Medical Sciences High Throughput Sequencing Facility, UK, using a Hiseq 2000 system. For data analysis, adapters were removed using fastx-trimmer, and files were collapsed using fastx-collapser. First nucleotide and length for each sequence were extracted using a custom Perl script. To identify miRNAs, miRDeep2 annotation of T. spiralis performed previously (Sarkies et al., 2015) was used, and mature miRNA sequences were identified in each library by sequence matching using a custom Perl script. BLAST was used to examine sequence files for any matches to Y-RNA. Potential targets in mouse 3′ untranslated regions (UTRs) were assessed by searching for complementary sequences to the seed sequence (nt 2–8) within the 3′UTRs of mouse RNAs, downloaded from Ensembl. All figures were prepared in R.
3. Results
3.1. Characterisation of RNA protection in T. spiralis-secreted material
In order to characterise the RNA content of secreted material from T. spiralis, we isolated secreted material from adult worms and MSL. We treated secreted material with different concentrations of exonuclease to test whether the extracellular RNA was encapsulated in EVs or otherwise protected, then extracted RNA and analysed size and quantity using a bioanalyzer. We recovered abundant small RNAs from both adults and larvae. Adult small RNA was largely unaffected by RNase treatment, indicating that it was protected from exonuclease digestion. In contrast, the majority of small RNA from larvae was degraded by the same treatment (Fig. 1A–F). Quantification of the area under the bioanalyzer trace demonstrated that whilst the amount of RNA remaining was similar after treatment of adult-secreted material (mock/maximum RNase = 1.1), only approximately 50% remained after treatment of MSL-secreted material (mock/maximum RNase = 2). This suggests that adult T. spiralis secretes small RNA that is protected whilst MSL-secreted material is not protected. Previous characterisation of extracellular RNA has shown that some secreted RNA can be bound by proteins in the absence of vesicles (Arroyo et al., 2011, Turchinovich et al., 2011); however this affords protection to RNase at much lower concentrations than used here (7 nM as opposed to 1 μM) (Turchinovich et al., 2011). Therefore, in line with previous studies on parasitic helminths (Coakley et al., 2015), we considered whether EVs might be protecting adult small RNAs, whilst MSL-secreted small RNAs might not be encapsulated.
Fig. 1.

Profile of RNA in Trichinella spiralis secreted material. Bioanalyzer traces showing small RNA profiles from secreted material extracted from adult (A–C) and muscle stage larvae (MSL; D–F) after exposure to different concentrations of exonuclease as indicated above each plot. nt, nucleotides.
We used a standard protocol (see Section 2) to isolate EVs from MSL-secreted products by ultracentrifugation, and examined the pelleted material by Transmission Electron Microscopy (TEM), which revealed vesicle-like structures up to 150 nm in diameter (Fig. 2A). To confirm this, we used Nanoparticle Tracking Analysis (NTA) to profile secreted material from MSL. The results showed a disperse profile of particle size consistent with TEM images, confirming the presence of EVs with many in the size range typical of exosomes (Fig. 2B), with a similar profile observed by NTA of secreted material from adult worms (Fig. 2C). Taken together with the lack of exonuclease protection, this indicates that whilst MSL do secrete EVs, the majority of secreted small RNAs do not appear to be contained within these structures. Quantitation of the number of vesicles secreted by each life stage indicated that over the 72 h period of maintenance in vitro, adult T. spiralis secreted on average 3.15 × 105 vesicles per parasite per 24 h, in comparison to 1.26 × 105 vesicles per parasite per 24 h for MSL. Given that adult worms are estimated to be between 1.4 to 4× the length of larvae, this is indicative of broadly similar vesicle production relative to body mass. Taken together with the lack of protection of RNAs, this implies that adult T. spiralis secrete RNAs in vesicles whereas T. spiralis MSL secrete vesicles and RNA separately.
Fig. 2.

Characterisation of extracellular vesicles secreted by Trichinella spiralis parasites at different life stages. (A) Transmission electron microscopy images of EVs derived from muscle stage larvae. (B, C) Histograms showing size distributions of EVs derived from muscle stage larvae and adult parasites, respectively, measured by nanoparticle tracking analysis. Modal size was considered as the representative size of the vesicle population isolated from conditioned culture medium. Values were expressed as mean ± S.D., where n = 4 measurements per sample.
3.2. Specific small RNAs are secreted by T. spiralis adults and MSL
We next characterised parasite small RNAs by high-throughput sequencing. Material extracted from both adult and MSL somatic tissue showed a profile peaking at 23 nt in length with a substantial fraction of 24 and 25 nt small RNAs, mapping to genes and transposable elements (Fig. 3A, D, F). This is consistent with our previous demonstration that T. spiralis produces abundant endogenous short interfering RNAs (siRNAs) with 23–25 nt length that map to transposable elements (Supplementary Fig. S1; (Sarkies et al., 2015)). In contrast, secreted RNAs from both larvae and adult worms peaked at 22 nt in length (Fig. 3C and F), and indeed small RNAs derived from transposable elements are depleted from secreted material (Fig. 3G). We then assessed whether small RNAs in secreted material were protected from exonuclease digestion. The profile of secreted small RNAs from adult worms was similar with and without exonuclease treatment, however in larvae the majority of small RNAs were removed by exonuclease treatment under the same conditions, suggesting that most small RNAs in MSL-secreted material are unprotected (Fig. 3B, C, E and F). Consistent with this observation, small RNAs from purified EVs from MSL showed a flat profile, whilst small RNAs in the supernatant were enriched for small RNAs with a size of ∼22 nt (Fig. 3G and H). Of note, in contrast to other studies on parasitic nematode-secreted material (Buck et al., 2014), we did not detect Y-RNAs, using BLAST to test for divergent alignment to C. elegans or human Y-RNAs in the secreted material from T. spiralis or in the material isolated from whole worms. However, the Y-RNA family has yet to be described in T. spiralis, potentially due to the rapid divergence in sequence over evolutionary distance (Boria et al., 2010), and thus we cannot exclude the presence of Y-RNA in secreted material. Again in contrast to earlier studies on parasitic nematodes (Buck et al., 2014, Chow et al., 2019), 22G-RNAs were not found in secreted material, as these evolved in Chromadorea nematodes and are not found in T. spiralis (Sarkies et al., 2015).
Fig. 3.

Characterisation of small RNAs secreted by Trichinella spiralis by high-throughput sequencing. (A–C) The length in nucleotides (nt) and the first nucleotide (AGC or U) of small RNA sequences in adult-secreted material either with or without exonuclease treatment compared with total worm material. (D–E) The length and first nucleotide of small RNA sequences in muscle stage larvae-secreted material either with or without exonuclease treatment compared with total worm material. (G, H) Supernatant (unencapsulated) or pellet (extracellular vesicles) after ultracentrifugation of muscle stage larvae-secreted material. All plots show the number of distinct sequences found in the high-throughput small RNA sequencing data. (I) Fraction of reads mapping to different genomic features across datasets.
miRNAs are an important component of RNA secreted by parasitic nematodes (Buck et al., 2014, Quintana et al., 2019). We therefore mapped small RNAs from secreted material to our previous annotations of miRNAs in T. spiralis (Sarkies et al., 2015). Secreted products contained approximately 15% miRNA in both larvae and adults, approximately two-fold depleted relative to the whole worm. This fraction was similar for adult material treated with exonuclease, suggesting that most miRNAs secreted by adult worms are protected from exonuclease activity, in contrast to miRNAs in MSL-secreted products (Fig. 3I). We found several specific miRNAs at high levels in secreted products. Most miRNAs from MSL were completely removed by exonuclease treatment, whereas miRNAs secreted by adult parasites were protected from digestion. Many miRNAs showed differential abundance in secreted material relative to total worm tissue (Fig. 4). The majority of miRNAs were depleted in secreted material, whilst a small number were enriched (Fig. 5A), suggesting some selectivity in which miRNAs are secreted relative to those retained within cells. Notably, the correlation between enrichment in MSL and adult-secreted material was weak (Fig. 5B), suggesting either differences in specificity and abundance of miRNA expression, or mechanisms of secretion between the two stages.
Fig. 4.

Characterisation of micro RNAs (miRNAs) in Trichinella spiralis secreted material. Heatmap indicating the normalised abundances of miRNAs. miRNAs are sorted in descending order of abundance in muscle stage larvae-secreted material.
Fig. 5.

Selective secretion of micro RNAs (miRNAs) by Trichinella spiralis muscle stage larvae. (A) Scatter plot indicating the relationship between abundance in total worms and abundance in secreted material. The black line indicates equal abundance, the green line indicates a ∼1.4-fold enrichment (log2 0.5) and purple indicates. a 1.4-fold depletion. (B) Scatter plot showing correlation between enrichment in adult-secreted material (X-axis) and enrichment in larval-secreted material (Y-axis). The line of best fit according to a linear model is shown, together with the correlation coefficient.
3.3. Potential roles of secreted miRNAs in T. spiralis infection
To investigate the possible consequences of miRNA secretion by T. spiralis larvae, we identified miRNAs >1.4-fold enriched in the secreted material (Fig. 5A; Table 1). Interestingly, these included a homologue of mammalian miR-31, previously annotated as a homologue of C. elegans miR-72 (Fig. 5, Table 1). This was notable because miR-31 has a well-documented role in muscle development by regulating translation of the myogenic determination gene Myf5. miR-31 binds to the 3′UTR of Myf5 through a perfect match to the seed sequence, which is conserved in primates, rodents and canines (Supplementary Fig. S2). The miR-31 seed sequence is shared between mammalian miR-31 and the T. spiralis homologoue, thus the T. spiralis homologue is also predicted to target Myf5. miR-31 is transcribed at high levels in quiescent satellite cells, and is sequestered together with Myf5 RNA in messenger ribonucleoprotein (mRNP) granules. Following activation of satellite cells during development or after injury, mRNP granules are dissociated, miR-31 is reduced, and Myf5 mRNA is released from repression, allowing myogenesis to proceed (Crist et al., 2012). Some enriched miRNAs showed no homology to mammalian miRNAs but nevertheless were predicted to target sites in mammalian 3′UTRs (Table 1), indicating that exposure to these miRNAs may have effects on gene expression in infected cells.
Table 1.
MicroRNAs (miRNAs) found to be enriched in Trichinella spiralis-secreted material. The sequence, potential homology to annotated Caehorhabditis elegans and human miRNAs, and the number of potential targets in mammalian 3′ untranslated regions are shown.
| Name | Sequence | Caenorhabditis elegans homologue | Mammalian homologue | Number of target UTRs (mouse) | Enrichment secreted material (MSL) | Enrichment secreted material (adult) |
|---|---|---|---|---|---|---|
| Tsp miR_68 | TGAGATCACCGTGAAAGCCT | miR-80 | n/a | 3430 | 16.4 | 2.6 |
| Tsp miR 15 | TGAGGTAGTAGGTTGTATAGTT | let7 | let7 | 3815 | 13.5 | 0.25 |
| Tsp miR 41 | TCACCGGTTACTAAAACATGCAT | n/a | n/a | 606 | 10.3 | 0.0035 |
| Tsp miR-65 | AACCCGTAGATCCGAACTTGT | miR-51 | miR-100 | 483 | 8.5 | 34.7 |
| Tspi miR-19 | GTGAGCAAAGTTTCAGGTGTGT | miR-87 | n/a | 4476 | 7.6 | 0.0067 |
| Tsp miR-72 | TCACCGGTCCATTTTATCTTCT | n/a | n/a | 606 | 3.7 | 0.80 |
| Tsp miR-8 | CGGATAGCACTTTTGGTAGCT | n/a | n/a | 2624 | 3.6 | 0.94 |
| Tsp miR-60 | AGGCAAGATGTTGGCATAGCT | miR-72 | miR-31 | 4568 | 1.8 | 1.5 |
| Tsp miR-78 | TAACCGTTTCCCTCTTTCAGTGC | n/a | n/a | 738 | 1.8 | 2.8 |
| Tsp miR-85 | TCACCGGATCATTTTATCTT | n/a | n/a | 824 | 1.7 | 0.088 |
MSL, muscle stage larvae.
4. Discussion
Small RNAs encapsulated in EVs are secreted from many cell types, and there is growing evidence that this is a common feature of all the major classes of helminth parasites (Tritten and Geary, 2018). Small RNAs secreted by the parasitic nematodes Heligmosomoides polygyrus and Litomosoides sigmodontis are resistant to degradation by RNase, but sensitive in the presence of Triton X-100, indicative of encapsulation in membrane-bound vesicles (Buck et al., 2014, Quintana et al., 2019). The number of exosome-sized particles released by L. sigmodontis as determined by NTA was very low in comparison to H. polygyrus, and sensitivity to RNase was also conferred by exposure to proteinase K, suggesting that extracellular RNAs might also be stabilised via interaction with proteins (Quintana et al., 2019). Analysis of small RNAs secreted by larval Schistosoma mansoni identified a comparable abundance of miRNAs and tRNA derived small (ts) RNAs in EV-enriched and EV-depleted fractions, again suggesting extracellular existence outside vesicles and possible stabilisation by proteins (Nowacki et al., 2015).
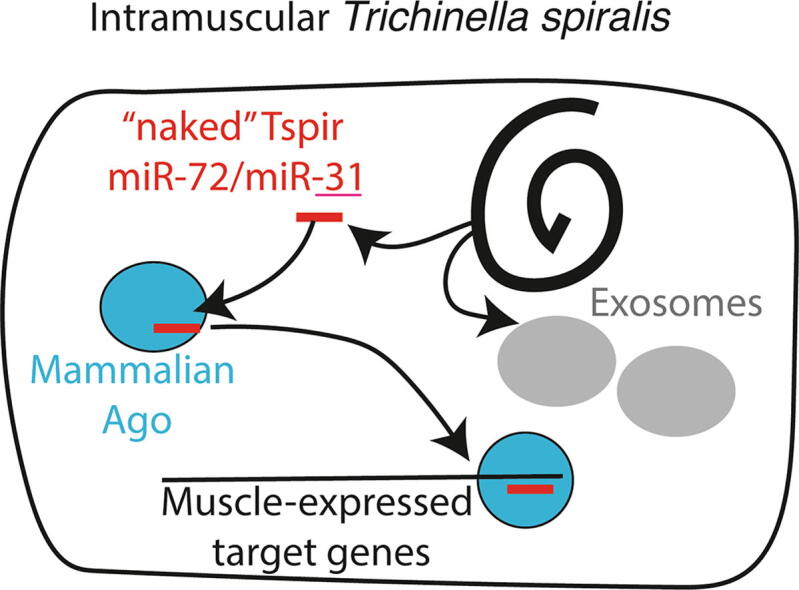
Our results indicate that adult T. spiralis secrete small RNAs that are resistant to exonuclease digestion, but that those secreted by MSL are largely unprotected. It is possible that this dichotomy is caused by differences in protein binding, in particular the presence of an RNA-binding protein in adult secreted material but not secreted from MSL. There is some evidence that miRNAs bound to Ago2 can be secreted by human cells (Arroyo et al., 2011, Turchinovich et al., 2011), and that this offers some protection against nucleases (albeit tested only to 100-fold lower concentrations than we tested here). However, a more prevalent view is that encapsulation in EVs is responsible for protection of extracellular small RNAs (Coakley et al., 2015, Turchinovich et al., 2016). Therefore we suggest that small RNAs secreted by adults are encapsulated within vesicles but small RNAs in MSL-secreted material are not. The authors of the study on S. mansoni suggested that small RNAs might be secreted in vesicles which were highly labile, lysing during in vitro culture to release their contents (Nowacki et al., 2015). Whilst that is possible, our current data were obtained via culture of two different stages of T. spiralis and subsequent processing of material under identical conditions, and intrinsic lability of vesicles from MSL alone appears unlikely. There is also no major difference in the viability of both stages under the culture conditions employed, assessed by Adenosine Triphosphate (ATP) assay. In addition, the few miRNAs that are resistant to exonuclease treatment in larval-secreted material do not show similar abundance to those that are sensitive (Fig. 4, Fig. 5B). We suggest that T. spiralis larvae secrete small RNAs via an alternative mechanism, potentially involving direct release or in complex with a chaperone (Fig. 6). This model is highly speculative and remains a question for future work. Recent studies have discovered a secreted Argonaute, part of the worm-specific WAGO clade that binds to siRNAs, although not miRNAs, in secreted material from parasitic nematodes within EVs (Chow et al., 2019). We have previously characterised the T. spiralis Argonautes, finding that they have homologues both of miRNA-binding Agos and WAGOs (Sarkies et al., 2015). It is therefore possible that a secreted Ago is involved in T. spiralis small RNA stabilisation, either in EVs or outside, but this still remains to be determined. Interestingly, not all miRNAs found in secreted material from other helminth parasites appear to be encapsulated within vesicles (Buck et al., 2014, Nowacki et al., 2015), suggesting that secretion of miRNAs by T. spiralis MSL might be an extreme example of processes found in other species.
Fig. 6.

Model showing secretion of small RNAs by muscle stage larvae Trichinella spiralis or adult T. spiralis, and how these might play roles in host gene expression regulation by the parasite.
Differences between secreted RNAs in adult and larval T. spiralis might also relate to the unusual life cycle of this nematode. In the intestinal phase, development of infective larvae to adult worms requires invasion of epithelia (ManWarren et al., 1997, Gagliardo, 2002). Although described as occupying a multicellular niche, worms migrate through epithelial cell monolayers in vitro, leaving a trail of dead cells (ManWarren et al., 1997), and have been observed migrating in and out of the epithelial cell layer in vivo (Wright, 1979). This is therefore not an intracellular parasite in the conventional sense in terms of permanent enclosure and development within a single cell.
In contrast, invasion of myofibres by L1s and subsequent development is entirely intracellular, forming an interaction which can remain stable for years. The early stages of remodelling of skeletal muscle by T. spiralis has gross similarities to repair of muscle following injury with respect to recruitment, activation and proliferation of satellite cells, presumably in response to the damage caused by parasite invasion (Wu et al., 2008). The subsequent processes diverge, leading to repair and regeneration of a contractile myofibre following injury, and remodelling into a nurse cell by T. spiralis. In this respect, secretion of a homologue of miR-31 by T. spiralis larvae is interesting, given its key role in repression of the myogenic programme (Crist et al., 2012). Moreover, miR-31 expression is associated with Duchenne Muscular Dystrophy (DMD): it is found at higher levels in human DMD biopsies, and its persistent upregulation in mouse models of the disease (mdx mice) is linked to delay of the muscle differentiation programme, reduced fibre maturation and intensive regeneration (Greco et al., 2009, Cacchiarelli et al., 2011). The intracellular location of larval T. spiralis within skeletal muscle would mean that unencapsulated small RNAs might have a higher chance of engaging with mammalian pathways of gene regulation. Differentiation and repair of skeletal muscle is highly complex, and the cellular remodelling effected by T. spiralis infection is quite unique, thus in order to test the plausibility of this model further, confirmation of miR-31/72 and other parasite-secreted miRNAs within muscle cells in vivo is required.
Acknowledgements
This study was supported by a UK Biotechnology and Biological Sciences Research Council (BBSRC) studentship to PJT (BB/M0111788/1), BBSRC grants to MES (BB/S001085/1) and MB (BB/L023091/1) and a grant from the UK Medical Research Council, to PS (Transgenerational Epigenetic Inheritance and Evolution).
Footnotes
Supplementary data to this article can be found online at https://doi.org/10.1016/j.ijpara.2020.05.008.
Contributor Information
Murray E. Selkirk, Email: m.selkirk@imperial.ac.uk.
Peter Sarkies, Email: p.sarkies@lms.mrc.ac.uk.
Appendix A. Supplementary data
The following are the Supplementary data to this article:
Supplementary Fig. S1.

Small RNAs aligning to (A) microRNAS (miRNAs) and (B) transposable elements using adult total Trichinella spiralis material as an example. X-axis shows length in nucleotides (nt) and colour shows the first nucleotide (AGC or U). Note the increased length of TE-mapping small RNAs relative to miRNA-derived reads.
Supplementary Fig. S2.

Conservation of binding site for the seed sequence of microRNA miR-31 in the 3′ untranslated region (UTR) of various mammalian myf5 mRNAs. mRNA sequences were taken from Ensembl (release 100, April 2020). The seed sequence is highlighted. Numbers corresponding to the human myf5 UTR downstream of the stop codon are shown.
References
- Arden S.R., Smith A.M., Booth M.J., Tweedie S., Gounaris K., Selkirk M.E. Identification of serine/threonine protein kinases secreted by Trichinella spiralis infective larvae. Mol. Biochem. Parasitol. 1997;90:111–119. doi: 10.1016/s0166-6851(97)00145-x. [DOI] [PubMed] [Google Scholar]
- Arroyo J.D., Chevillet J.R., Kroh E.M., Ruf I.K., Pritchard C.C., Gibson D.F., Mitchell P.S., Bennett C.F., Pogosova-Agadjanyan E.L., Stirewalt D.L., Tait J.F., Terawi M. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. U. S. A. 2011;108:5003–5008. doi: 10.1073/pnas.1019055108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartel D.P. Metazoan MicroRNAs. Cell. 2018;173:20–51. doi: 10.1016/j.cell.2018.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boria I., Gruber A.R., Tanzer A., Bernhart S.H., Lorenz R., Mueller M.M., Hofacker I.L., Stadler P.F. Nematode sbRNAs: Homologs of vertebrate y RNAs. J. Mol. Evol. 2010;70:346–358. doi: 10.1007/s00239-010-9332-4. [DOI] [PubMed] [Google Scholar]
- Buck A.H., Coakley G., Simbari F., McSorley H.J., Quintana J.F., Le Bihan T., Kumar S., Abreu-Goodger C., Lear M., Harcus Y., Ceroni A., Babayan S., Blaxter M., Ivens A., Maizels R.M. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat. Commun. 2014;5:5488. doi: 10.1038/ncomms6488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cacchiarelli D., Incitti T., Martone J., Cesana M., Cazzella V., Santini T., Sthandier O., Bozzoni I. miR-31 modulates dystrophin expression: new implications for Duchenne muscular dystrophy therapy. EMBO Rep. 2011;12:136–141. doi: 10.1038/embor.2010.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chow F.-W.-N., Koutsovoulos G., Ovando-Vázquez C., Neophytou K., Bermúdez-Barrientos J.R., Laetsch D.R., Robertson E., Kumar S., Claycomb J.M., Blaxter M., Abreu-Goodger C., Buck A.H. Secretion of an Argonaute protein by a parasitic nematode and the evolution of its siRNA guides. Nucleic Acids Res. 2019;47:3594–3606. doi: 10.1093/nar/gkz142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coakley G., Maizels R.M., Buck A.H. Exosomes and other extracellular vesicles: the new communicators in parasite infections. Trends Parasitol. 2015;31:477–489. doi: 10.1016/j.pt.2015.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coakley G., Buck A.H., Maizels R.M. Host parasite communications—Messages from helminths for the immune system: parasite communication and cell-cell interactions. Mol. Biochem. Parasitol. 2016;208:33–40. doi: 10.1016/j.molbiopara.2016.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coakley G., McCaskill J.L., Borger J.G., Simbari F., Robertson E., Millar M., Harcus Y., McSorley H.J., Maizels R.M., Buck A.H. Extracellular vesicles from a helminth parasite suppress macrophage activation and constitute an effective vaccine for protective immunity. Cell Rep. 2017;19:1545–1557. doi: 10.1016/j.celrep.2017.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crist C.G., Montarras D., Buckingham M. Muscle satellite cells are primed for myogenesis but maintain quiescence with sequestration of Myf5 mRNA targeted by microRNA-31 in mRNP granules. Cell Stem Cell. 2012;11:118–126. doi: 10.1016/j.stem.2012.03.011. [DOI] [PubMed] [Google Scholar]
- Faruqu F.N., Xu L., Al-Jamal K.T. Preparation of exosomes for siRNA delivery to cancer cells. J. Vis. Exp. 2018;2018 doi: 10.3791/58814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gagliardo L.F. Molting, ecdysis, and reproduction of Trichinella spiralis are supported in vitro by intestinal epithelial cells. Infect. Immun. 2002;70:1853–1859. doi: 10.1128/IAI.70.4.1853-1859.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gebert L.F.R., MacRae I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019;20:21–37. doi: 10.1038/s41580-018-0045-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbings D.J., Ciaudo C., Erhardt M., Voinnet O. Multivesicular bodies associate with components of miRNA effector complexes and modulate miRNA activity. Nat. Cell Biol. 2009;11:1143–1149. doi: 10.1038/ncb1929. [DOI] [PubMed] [Google Scholar]
- Greco S., De Simone M., Colussi C., Zaccagnini G., Fasanaro P., Pescatori M., Cardani R., Perbellini R., Isaia E., Sale P., Meola G., Capogrossi M.C., Gaetano C., Martelli F. Common micro-RNA signature in skeletal muscle damage and regeneration induced by Duchenne muscular dystrophy and acute ischemia. FASEB J. 2009;23:3335–3346. doi: 10.1096/fj.08-128579. [DOI] [PubMed] [Google Scholar]
- Jasmer D.P. Trichinella spiralis infected skeletal muscle cells arrest in G2/M and cease muscle gene expression. J. Cell Biol. 1993;121:785–793. doi: 10.1083/jcb.121.4.785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T., Zhang Q., Zhang J., Li C., Miao Y.R., Lei Q., Li Q., Guo A.Y. EVmiRNA: a database of miRNA profiling in extracellular vesicles. Nucleic Acids Res. 2019;47:D89–D93. doi: 10.1093/nar/gky985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ManWarren T., Gagliardo L., Geyer J., McVay C., Pearce-Kelling S., Appleton J. Invasion of intestinal epithelia in vitro by the parasitic nematode Trichinella spiralis. Infect. Immun. 1997;65:4806–4812. doi: 10.1128/iai.65.11.4806-4812.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowacki F.C., Swain M.T., Klychnikov O.I., Niazi U., Ivens A., Quintana J.F., Hensbergen P.J., Hokke C.H., Buck A.H., Hoffmann K.F. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J. Extracell. Vesicles. 2015;4:28665. doi: 10.3402/jev.v4.28665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quintana J.F., Kumar S., Ivens A., Chow F.W.N., Hoy A.M., Fulton A., Dickinson P., Martin C., Taylor M., Babayan S.A., Buck A.H. Comparative analysis of small RNAs released by the filarial nematode Litomosoides sigmodontis in vitro and in vivo. PLoS Negl. Trop. Dis. 2019;13 doi: 10.1371/journal.pntd.0007811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkies P., Miska E.A. Small RNAs break out: the molecular cell biology of mobile small RNAs. Nat. Rev. Mol. Cell Biol. 2014;15:525–535. doi: 10.1038/nrm3840. [DOI] [PubMed] [Google Scholar]
- Sarkies P., Selkirk M.E., Jones J.T., Blok V., Boothby T., Goldstein B., Hanelt B., Ardila-Garcia A., Fast N.M., Schiffer P.M., Kraus C., Taylor M.J., Koutsovoulos G., Blaxter M.J., Miska E.A. Ancient and novel small RNA pathways compensate for the loss of piRNAs in multiple independent nematode lineages. PLoS Biol. 2015;13 doi: 10.1371/journal.pbio.1002061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tritten L., Geary T.G. Helminth extracellular vesicles in host–parasite interactions. Curr. Opin. Microbiol. 2018;46:73–79. doi: 10.1016/j.mib.2018.08.002. [DOI] [PubMed] [Google Scholar]
- Turchinovich A., Weiz L., Langheinz A., Burwinkel B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011;39:7223–7233. doi: 10.1093/nar/gkr254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turchinovich A., Tonevitsky A.G., Burwinkel B. Extracellular miRNA: a collision of two paradigms. Trends Biochem. Sci. 2016;41:883–892. doi: 10.1016/j.tibs.2016.08.004. [DOI] [PubMed] [Google Scholar]
- Wright K.A. Trichinella spiralis: an intracellular parasite in the intestinal phase. J. Parasitol. 1979;65:441–445. [PubMed] [Google Scholar]
- Wu Z., Sofronic-Milosavljevic L., Nagano I., Takahashi Y. Trichinella spiralis: nurse cell formation with emphasis on analogy to muscle cell repair. Parasites Vectors. 2008;1:27. doi: 10.1186/1756-3305-1-27. [DOI] [PMC free article] [PubMed] [Google Scholar]