

In rats as in humans, nonhelping bystanders suppress helping whereas naive bystanders boost helping above individual levels.
Abstract
To investigate whether the classic bystander effect is unique to humans, the effect of bystanders on rat helping was studied. In the presence of rats rendered incompetent to help through pharmacological treatment, rats were less likely to help due to a reduction in reinforcement rather than to a lack of initial interest. Only incompetent helpers of a strain familiar to the helper rat exerted a detrimental effect on helping; rats helped at near control levels in the presence of incompetent helpers from an unfamiliar strain. Duos and trios of potential helper rats helped at superadditive rates, demonstrating that rats act nonindependently with helping facilitated by the presence of competent-to-help bystanders. Furthermore, helping was facilitated in rats that had previously observed other rats’ helping and were then tested individually. In sum, the influence of bystanders on helping behavior in rats features characteristics that closely resemble those observed in humans.
INTRODUCTION
In 1964, on a cold March night, Catherine Genovese was murdered on a crowded residential street in Queens. The New York Times reported that more than 35 of Ms. Genovese’s neighbors heard or witnessed her murder and not a single one intervened or called the police (1). Although this account is now known to be nonfactual (2), the case inspired psychologists B. Latané and J. Darley (3) to investigate the phenomenon. They tested the behavior of human subjects in the presence of individuals who were part of the research team and had been instructed not to help in various contrived scenarios; the nonhelping accomplices are termed confederates in the psychology literature. Subjects were consistently less likely to help when tested with confederates than when tested alone. This phenomenon of less helping in the presence of nonhelping, confederate bystanders than when alone is enshrined in modern psychology as the bystander effect (4–7) and has been ascribed to a diffusion of responsibility, pluralistic ignorance, or the sheer fear of appearing foolish (8).
Here, we deliberately copied the design of psychology studies by testing how one or two nonhelping rats, analogous to human confederates, influence the behavior of a subject rat who could help. We hypothesized that, in parallel to humans, rats would help less in the presence of others who were not helping and that this effect would be stronger in the presence of two nonhelpers than one.
Bystanders may not always suppress helping. The main finding of a recent study of naturally occurring conflicts recorded on video by surveillance cameras was that people intervened in over 90% of the 219 conflicts recorded and analyzed (9). Of relevance to the present study, the likelihood of intervention increased by 10% with the addition of each additional observer (bystander). This result reveals that, although a group of individuals help less frequently than would be expected if the individuals acted independently (10), the overall chance of an individual in distress getting help increases, which is evidence that bystanders do not always suppress the occurrence of helping.
To examine how bystanders, either potential helpers or those incompetent to help, influence the helping behavior of rats, we used the trapped rat helping paradigm (11–12). In this paradigm, rats are placed in an arena with a centrally located restrainer containing a trapped rat. Most individually tested rats learn to open the restrainer door for a trapped rat of a familiar strain, doing so with increasing likelihood and at progressively shorter latencies over the course of 12 daily sessions (11, 13–14). In the current study, rats were tested with (i) one or two rats who were incompetent to help (incompetent helpers analogous to human confederates as above), (ii) one or two rats who could potentially help [potential helpers analogous to naïve bystanders as in the video study mentioned above (9)], or (iii) alone (Controls). All rats in a testing group were strangers to one another.
Rats were rendered incompetent to help through the administration of the benzodiazepine anxiolytic midazolam (4 mg/kg, ip). This dose of midazolam neither anesthetized nor severely sedated animals. Treated rats maintained a normal posture (see Fig. 1) and moved freely around the arena. They were even able to open the restrainer door, although this happened rarely (in 8 of 376 testing sessions that included either one or two incompetent helper rats).
Fig. 1. The presence of incompetent helper bystanders antagonizes rat helping by reducing reinforcement.

(A and B) One Sprague-Dawley potential helper rat (p)was tested with one (A) or two (B) Sprague-Dawley incompetent helpers (n) and a trapped rat (t). Photographs show the first door-opening (left) and a later day when the potential helper did not open the restrainer door (right). (C) Rats tested with incompetent helpers had fewer total and consecutive openings [median and interquartile range (IQR), 25 to 75 percentile] than did Control rats [total: Kruskal-Wallis rank sum χ2(2, N = 23) = 5.28, P = 0.07, η2 = 0.16; consecutive: χ2(2, N = 23) = 7.19, P = 0.03, η2 = 0.26; Dunn’s pairwise tests with Bonferroni corrections: two versus Control: P = 0.04; one versus Control: P = 0.12; two versus one: P = 1.00]. The dotted line shows perfect reinforcement. (D) The latency decreased from the first to third day of opening for Control rats [Wilcoxon signed rank V(N = 8) = 36, P = 0.007, r2 = 0.38] but not for rats tested with one [V(N = 8) = 16, P = 0.63] or two [V(N = 7) = 8, P = 0.85] incompetent helpers. (E) The number of rats in each group (Control, one, or two incompetent helpers) that opened three times or more, one or two times, or never opened is shown. Photo credits: M.C. Jacobi, University of Chicago.
RESULTS
The influence of incompetent helper bystanders on helping
We first tested rats with either one or two incompetent helper rats. Most rats tested with incompetent helpers (14 of 16) opened the door at least once during the first 6 days of testing as did all Control rats. Yet, unlike Control rats, rats tested with incompetent helpers failed to consistently open on subsequent days after their first opening. Therefore, rats tested with incompetent helpers had fewer consecutive door-openings (Fig. 1C).
For most Control rats, successive door-openings occurred on successive days so that the interval between the first and third door-openings was 2 days, the minimum possible (Fig. 1D). In contrast, the interval between first and third door-openings was typically more than 3 or 5 days in rats tested with one or two incompetent helpers, respectively. Furthermore, whereas the latency on the third day of opening was less than that on the first day of opening for Control rats, this was not the case for rats tested with incompetent helpers (Fig. 1D).
Rat helping is socially selective with rats only helping rats from familiar strains, regardless of their individual familiarity (13). To determine whether the bystander effect is similarly socially selective, we tested whether rats would be influenced by the presence of an incompetent helper from an unfamiliar strain (Fig. 2A). Two weeks of cohousing is sufficient to confer strain familiarity. Therefore, albino Sprague-Dawley rats were either cohoused with another Sprague-Dawley rat or with a black-caped Long-Evans rat, rendering them unfamiliar or familiar with Long-Evans rats, respectively. Sprague-Dawley rats from both of these conditions were tested with a trapped Sprague-Dawley rat in the presence of one Long-Evans incompetent helper rat (Fig. 2A). Thus, the protocol was identical for the two conditions with the only difference being whether the potential helper Sprague-Dawley was familiar with the Long-Evans strain. As above, rats in the testing groupings were strangers to each other; specifically, Sprague-Dawley rats in the familiar group were tested with a stranger Long-Evans rat that was not the Long-Evans cagemate.
Fig. 2. Strain familiarity is required for the antagonism of helping produced by incompetent helper bystanders.

(A) A Sprague-Dawley potential helper rat was tested with a trapped Sprague-Dawley rat and a Long-Evans incompetent helper. (B) Rats familiar with Long-Evans rats opened less frequently and accrued fewer consecutive openings than rats unfamiliar with Long-Evans rats who opened similarly to control rats [total: χ2(2, N = 64) = 8.81, P = 0.01, η2 = 0.11; familiar versus control: P = 0.009, unfamiliar versus control: P = 1.00, unfamiliar versus familiar: P = 0.15; consecutive χ2(2, N = 50) = 12.78, P = 0.002, η2 = 0.23; familiar versus control: P = 0.001, unfamiliar versus control: P = 0.81, unfamiliar versus familiar: P = 0.11]. (C) Although the first day of opening was similar [χ2(2, N = 50) = 0.38, P = 0.83], the third opening differed across the conditions [χ2(2, N = 50) = 11.12, P = 0.004, η2 = 0.19]. The latency decreased from first to third openings for control [V(N = 38) = 36, P < 0.0001, r2 = 0.87] and unfamiliar [V(N = 7) = 0, p = 0.009, r2 = 0.40] rats but not for rats familiar with the Long-Evans strain [V(N = 7) = 1, P = 0.13]. Latencies are graphed individually for the two rats that opened in the familiar condition (dashed lines). (D) Graph has the same format as Fig. 1E. The right ordinate is for the control rats (N = 48). Photo credits: R.R. Kukreja, University of Chicago.
Sprague-Dawley rats familiar with the Long-Evans strain and tested with a Long-Evans incompetent helper rat resembled rats tested with one or two incompetent Sprague-Dawley helpers in that they opened less frequently than did an expanded group of 48 control rats gleaned from contemporaneously run experiments (n = 48; Fig. 2B). [Note that the expanded group of 48 control rats is denoted by the lowercase control in contrast to the eight (uppercase) Control rats tested specifically during this study. The 48 control rats are inclusive of the eight Control rats.] In contrast to the Sprague-Dawley rats familiar with Long-Evans rats, Sprague-Dawley rats unfamiliar with Long-Evans rats behaved similarly to control rats (Fig. 2B). These findings suggest that clues from rats of familiar strains influence helping behavior, whereas clues from rats of unfamiliar strains are largely ignored.
Most control rats (30 of 38 rats that opened at least once) were perfectly reinforced for the first three openings, with the first and third days of opening occurring 2 days apart (Fig. 2C). This was also true for five of the seven rats unfamiliar with the Long-Evans strain who opened at least once. In contrast, none of the rats familiar with Long-Evans rats were perfectly reinforced. Furthermore, the opening latency decreased from the first to the third opening both in control rats and rats unfamiliar with Long-Evans rats. In contrast, only two rats familiar with Long-Evans rats opened at least three times; in both of these cases, the latency of the third opening was greater than that of the first opening (Fig. 2, C and D).
To summarize the behavior of rats tested with incompetent helpers, we compared door-opening streaks for rats that opened at least once. Rats unfamiliar with the strain of the incompetent helper/s behaved similarly to control rats who were tested solo, with both groups consistently opening for many consecutive days upon the first opening (Fig. 3A). However, rats familiar with the strain of the incompetent helper/s failed to open on the days following their first opening. This latter finding held regardless of whether the incompetent helper rat was from the same (Sprague-Dawley) or a different (Long-Evans) strain as the potential helper rat. Thus, as is the case for helping, bystander influence depends on strain familiarity rather than on strain identity (13).
Fig. 3. Incompetent helper bystanders of a familiar strain antagonize helping by reducing reinforcement.

(A) The length of the initial streak of door-opening, the streak inclusive of and following upon a rat’s first opening, is graphed by the first day of door-opening for conditions involving one or two incompetent helpers and for controls. The dotted line represents perfect reinforcement, the streak length when an animal opens on every day following his first opening. Jitter (0.05 to 0.1) was introduced to separate overlapping symbols (control versus unfamiliar; two incompetent helpers versus familiar). (B) The proportion of all rats that opened at least once and showed perfect reinforcement thereafter is greatest for control rats and least for rats tested with one or two incompetent helpers from a familiar strain. Rats tested with an incompetent helper from an unfamiliar strain showed an intermediate level of reinforcement. (C) After opening for the first time, the proportion of days that rats opened thereafter is graphed. Symbols are shown in the lower panel. The N in the lower panel refers to the number of animals in each group that opened at least once.
Perfect reinforcement after the first opening was observed in more than 75% of control rats and nearly half of rats in the unfamiliar condition but was nearly absent in rats tested with incompetent helpers from familiar strains (Fig. 3B). Last, the proportion of days on which rats helped, after opening once, was greatest for rats tested alone (control) or with an incompetent helper from an unfamiliar strain and least for rats tested with two incompetent helpers of the same strain or with one incompetent helper of a foreign but familiar strain (Fig. 3C). Reinforcement was intermediate for rats tested with one incompetent helper of the same strain.
The influence of potential helper bystanders on helping
To examine the consequences of additional untreated and competent-to-open bystanders, we compared the opening behavior of Controls with the behavior of groups of two (duos) or three (trios) rats, all of whom were potential openers (Fig. 4, A and B). Collectively, duos and trios are termed the plus conditions. All rats in plus conditions were naïve, untreated, and therefore potential helpers. Rats in the trio condition opened the door consistently and at short latency by day 3 and were therefore tested as a group for only 6 days. Thus, analysis of duos and trios is restricted to the initial six testing days.
Fig. 4. Additional potential helper rats facilitate opening relative to rats tested alone.
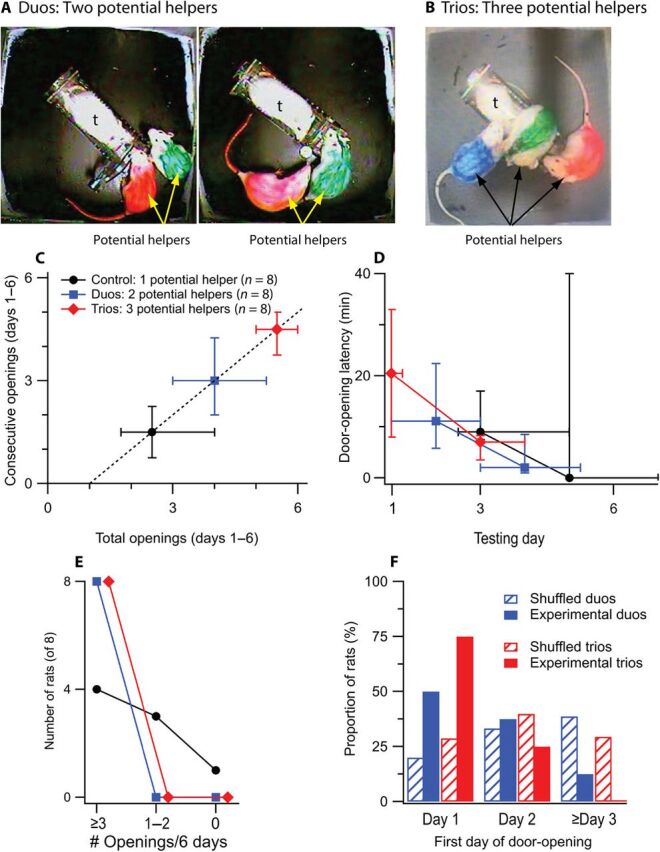
(A and B) Photographs of duos (A) and trios (B) of rats on the first (left) and a subsequent (right) door-opening. (C) Trios exhibited a greater frequency of openings than Control rats [total: Kruskal-Wallis rank sum χ2(2, N = 24) = 9.42, P = 0.009, η2 = 0.35; Dunn’s pairwise tests with Bonferroni corrections: duos versus Control, P = 0.27; trios versus Control, P = 0.007; Consecutive: χ2(2, N = 23) = 6.63, P = 0.04, η2 = 0.23; duos versus Control, P = 0.41; trios versus Control, P = 0.03]. (D) Trio rats opened earlier than Control and duo rats [first day: χ2(2, N = 23) = 10.15, P = 0.006, η2 = 0.61; duos versus control, P = 0.19; trios versus control, P = 0.004; third day: χ2(2, N = 23) = 8.04, P = 0.02, η2 = 0.30; duos versus control, P = 0.19; trios versus control, P = 0.004]. Duos and trios decreased their latency from first to third day of opening but Control rats did not [Controls: Wilcoxon signed rank V(N = 7) = 10, P = 0.77; duos: V(N = 8) = 34, P = 0.01; trios: V(N = 8) = 34.5, P = 0.01]. (E) Graph has the same format as Fig. 1E. (F) Greater proportions of rats in the experimental duos and trios opened on the first day of testing than did rats in the shuffled groups [trios versus shuffled trios χ2(df = 2) = 8.60, P = 0.01; duos versus shuffled duos χ2(df = 2) = 5.91, P = 0.05]. Photo credits: J.L. Havlik, University of Chicago.
On the first testing day, no rats in the Control condition freed the trapped rat, whereas 75% of the trios and half of the duos did so. Thereafter, all groups in the plus conditions opened at least three times in 6 days, whereas only half of the Control rats did (Fig. 4, C to E). Furthermore, groups in the plus conditions opened the restrainer door at lower latencies on the third day of opening than on the first day, as was true for Control rats (Fig. 4D). Breaks in opening were rare, with a total of only two to three recorded from all rats in each of the three conditions in 6 days. In sum, rats in the plus conditions initiated opening early and achieved consistent, day-after-day opening at short latency within 6 days.
We next examined whether the actions of groups of rats are simply the sum of the actions of individual rats. To this end, we constructed all possible combinations of two and three rats from our cohort of 48 control rats, resulting in the construction of shuffled duos (n = 1128) and trios (n = 17,296), respectively. Rats in the experimental duos and trios started opening earlier than did shuffled duos and trios (Fig. 4F). These data are evidence that groups of rats interact to augment door-opening more frequently and earlier in the testing schedule than would be expected if their individual contributions to door-opening were independent (10).
Observational learning
As mentioned above, trios opened consistently almost immediately, rendering further group testing of little value. Therefore, after 6 days of group testing, rats in the trios were tested individually for an additional 6 days, an approach that afforded analyses of how group testing influenced subsequently expressed individual helping behavior.
While trios opened on nearly every opportunity, one rat within the trio was the opener on any given day. To examine the effect of one rat’s opening on the other two rats who observed this opening, we looked at the subsequent opening behavior of rats who did not open on the final day of group testing (day 6). Most rats (13 of 17) who were not the trio’s opener during group testing on day 6 opened on day 7 when tested solo (Fig. 5A). This is markedly different from control rats among whom only 2 of 19 opened on day 7 after not opening on day 6 (Fig. 5A). Four of the trio rats that opened on day 7 when tested individually had never opened before; nonetheless, these four rats opened at latencies ranging from 0 to 7 s [median, 1.5 s; interquartile range (IQR), 0.0 to 4.0 s]. In contrast, the median latency on the first day of opening for controls was 8.0 s (IQR, 6.0 to 15.0). These data suggest that trio rats learned from observing the openings of others in their group and that this learning enabled and quickened their subsequent openings. Moreover, opening latencies decreased at the same rate when rats in trios observed others in their trio opening on the previous (base) day as when control rats opened on sequential days, demonstrating that rats learned from the actions of others as well as they did from their own actions (Fig. 5B).
Fig. 5. The experience of being in a trio influences future opening.
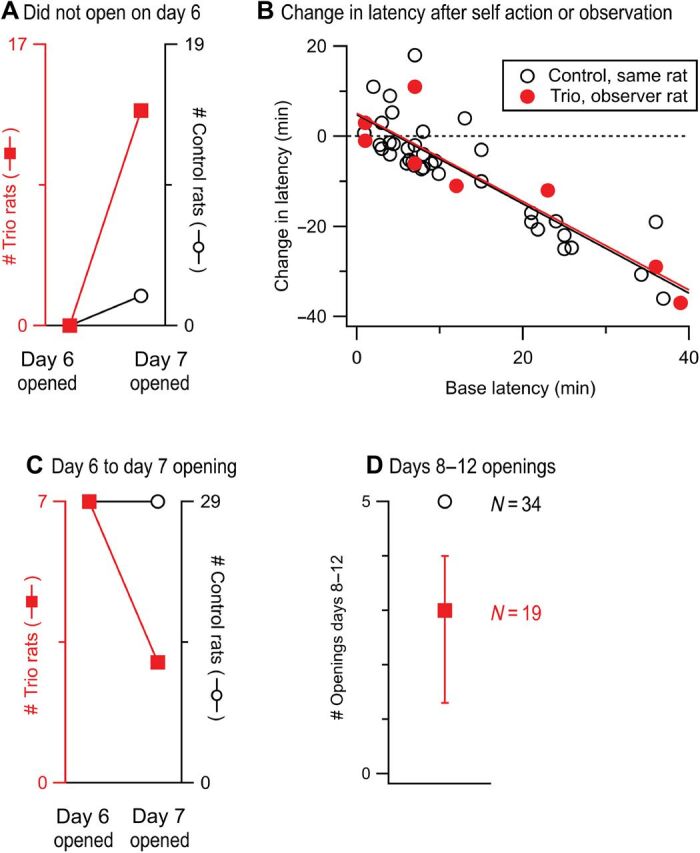
(A) Of the trio rats (left ordinate) that did not open on day 6, most opened on day 7 whereas only 2 of 19 control rats (right ordinate) did so. (B) When two sequential openings first occurred, the change in opening latency was inversely correlated to the latency on the base day (ΔLatency ~ LatencyN-1 * Condition; LatencyN-1 P < 0.0001). This correlation held regardless of whether the same rat opened on both days (black line) or two different rats from the trio condition opened (red line; Condition, P = 0.95). (C) Of the trio rats that opened on day 6 (tested as a group), less than half opened on day 7 (tested individually), whereas all control rats (right ordinate) who opened on day 6 also opened on day 7. (D) Control rats (empty black circle, IQR is within the symbol) who opened at least once during the first 6 days of testing (34 of 48) opened on all five of the final days of testing (days 8 to 12) but fewer than 20% of the trio rats who opened at least once during days 1 to 6 (19 of 24) opened on all 5 days.
The effect of trio experience on subsequent opening behavior
We next examined how the presence of two onlookers who witnessed a rat’s opening affected that rat’s subsequent opening behavior. We found that most trio rats (four of seven) who opened for their trio on day 6 failed to open when tested individually on day 7 (Fig. 5C). In stark contrast, all 29 control rats that opened on day 6 also opened on day 7. Thus, the loss of onlookers to witness a rat’s opening appears detrimental to future opening, suggestive of a facilitating audience effect. In support of this idea, individual trio rats who opened at least once during group testing (n = 19) opened a median of 3 (of 5) times during days 8 to 12 when tested solo (Fig. 5D). This stands in stark contrast to the 85% of control rats (who opened at least once during the first 6 days of testing) who opened every day during the final 5 days of testing. These data show that door-opening is not as reinforcing in rats tested alone after being tested as part of a group.
DISCUSSION
Here, we demonstrate bidirectional effects of bystanders on rat helping. The effect of rat bystanders depends on their capacity to help, with helping antagonized by incompetent helpers and facilitated by additional potential helpers. Recent evidence that naïve human bystanders facilitate or, at the very least, have no negative effect upon helping (9, 15) suggests that human helping may also be bidirectional.
Two incompetent helper rats antagonized helping more than did one. Similarly, helping is progressively more suppressed as the number of human bystanders increases (3). Another similarity between the rat and human versions of the bystander effect is that subjects are only influenced by bystanders of the same in-group (16). These similarities raise the possibility that similar circuits support the classic bystander effect in rats and humans. If this is the case, then either rats have greater cognitive and cultural capacities than currently appreciated or human helping operates independently of rational reasoning and cultural influences. Evidence that the building blocks for helping are fundamental circuits involved in parental care and affiliative interactions are shared across mammals (17) supports the latter possibility.
Incompetent helpers did not prevent or retard the initiation of helping. The first day of opening was the same for rats tested with one additional rat and for control rats and was even earlier for rats tested with two additional rats. It is likely that additional rats, regardless of their competency to help, hasten the initiation of opening by socially buffering potential helpers. Social buffering refers to the improved recovery from stressors afforded by the presence of conspecifics (18, 19). Of particular relevance here, conspecifics reduce the autonomic and behavioral expression of anxiety in response to novel environments (18). In the trapped rat paradigm, social buffering likely reduces the stress of the testing conditions, facilitating proximity to the centrally located restrainer door and thereby increasing opportunities for door-opening. After the first door-opening, however, social buffering can no longer account for the disparate effects on opening elicited by the presence of additional rats of different types. Additional rats that are incompetent to help exert a negative effect on reinforcement, whereas those that are potential helpers appear to facilitate reinforcement, as revealed by the reduction in helping shown by rats tested alone following group testing.
The show of indifference by incompetent helper rats is distinct from, and far more detrimental to, motivating helping than the absence of additional rats in control conditions. In groups, bystanders influence a rat’s interpretation of his own behavior, whereas, when solo, a rat’s own internal assessment of his actions rules. For a rat that has only experienced solo testing, this assessment is enough. Yet, when solo testing follows group testing, the self-reward pales in comparison to the recalled group reinforcement. It is possible then that just as rats tested solo after having the group experience failed to reinforce their behavior, rats tested solo following testing with incompetent helpers may show a positive rebound in reinforcement and helping.
The influence of bystanders promotes conformity, the matching of one’s behavior to that of a group. Conformity need not be limited to helping. Rats who did not eat an unpalatable food when alone ate it in the presence of a “demonstrator rat” who was eating the unpalatable food (20). Thus, the effect of bystanders may be more inclusively imagined as one that promotes conformity in all manner of observable behaviors.
MATERIALS AND METHODS
Subjects
Sprague-Dawley and Long-Evans male rats (Charles River, Portage, MI) were used for all studies. Rats were 8 to 11 weeks old at the start of the experiment. Rats were housed in pairs with ad libitum access to chow and water in a 12:12 light-dark cycle, between 06:00 and 18:00. Animals were allowed 2 weeks to acclimate to the housing environment. All procedures were approved by the University of Chicago Institutional Animal Care and Use Committee.
Setup
Above every arena (50 cm by 50 cm, 32 to 60 cm high), a charge-coupled device color camera (KT&C Co, Seoul, South Korea) was mounted. The cameras were connected to a video card (Geovision, Irvine, CA) in a dedicated PC. An acrylic rodent restrainer (25 cm by 8.75 cm by 7.5 cm; Harvard Apparatus, Holliston, MA) was placed diagonally in the center of the arena. The body of the restrainer had several small slits and the back end had a large slit, allowing for visual, auditory, olfactory, and tactile communication between free and trapped rats. A customized door, designed to be opened only from the outside, was located at the other end. Two weights (25 g each) were placed on one side of the door to facilitate opening.
Conditions
This experiment tested seven conditions, five involving Sprague-Dawley rats exclusively and two conditions using a mix of Sprague-Dawley and Long-Evans rats.
Exclusively Sprague-Dawley rat conditions
In the Control condition (n = 8), one potential helper rat was placed in an arena with one trapped rat. In the duo and trio conditions (n = 8 for each), two or three potential helper rats, respectively, were placed in an arena with one trapped rat. In the two conditions with incompetent helpers (n = 8 for each), one potential helper rat was placed in an arena with one or two incompetent helpers and one trapped rat.
Mixed Sprague-Dawley and Long-Evans rat conditions
To evaluate the role of strain familiarity in accepting cues from incompetent helpers, two conditions (n = 8 each) used one potential Sprague-Dawley helper, with one incompetent Long-Evans helper, and a Sprague-Dawley trapped rat. In the unfamiliar condition, the free rat had never seen or interacted with a Long-Evans rat. In the familiar condition, the free Sprague-Dawley rat had been living with a Long-Evans rat starting 2 weeks before the experiment’s start date. The incompetent Long-Evans helper was always an unknown individual to the Sprague-Dawley free rat, never his Long-Evans cagemate.
Control versus control cohorts
For selected analyses, we used a larger cohort of control rats than the eight in the Control condition. Specifically, data from 40 additional control rats from two experiments performed contemporaneously were added to the data from the eight Control rats studied here. These 40 rats were tested in the standard trapped rat helping paradigm without manipulation. References to the 48 control rats are in lowercase (“control”), whereas the eight Control rats tested in this study are referred to with an uppercase “Control.” No data from the 40 extra control rats have been published to date.
Shuffled data
To determine whether the facilitated opening in plus conditions was simply due to the additive contributions of multiple rats, we created groups of two or three rats from the control (n = 48) rats. All possible shuffles of two (n = 48!/46!*2!; 1,128) and three [n = (48!)/(45!*3!); 17,296] rats were constructed. For each shuffled group, an “opening” occurred if any one of the rats opened the restrainer door before 40 min. The shortest latency within the shuffled group was used as the opening latency. If more than one rat shared this latency, both rats were given credit for the opening. From these data, we calculated expectations for opening if control rats were assembled into groups of independently acting individuals.
Handling
Animals were habituated to transportation to the testing room, to the experimenters, and to the arenas for 10 days before being tested. On habituation day 1, rats were transported to the testing room and left undisturbed for an hour in their home cages. On subsequent days, rats’ tails were marked for identification, and rats were handled for 5 min by each experimenter. Rats were weighed three times per week. Cagemate pairs of rats were habituated to the testing arena for 30 min on each of days 3 to 8. Free rats were placed in the same arena for both habituation and testing when possible. Rats were colored with permanent markers to facilitate computerized tracking.
On days 6 to 8, rats destined to be incompetent helpers were injected with 0.15 ml of an isotonic saline solution. On day 9, rats were placed individually in arenas for 10 min and their behavior was recorded (open field). On day 10 of habituation, rats were placed on an elevated plus maze for 5 min. After each habituation session, rats were returned to their home cages and to the housing room. All sessions were run during the rats’ light cycle.
Midazolam dosing
To parallel human psychology experiments, we produced incompetent helper rats by administration of the benzodiazepine midazolam. On the first day of the experiment with two incompetent helpers, rats were given a dose of 2 mg/kg ip 20 min before the testing session. However, this dose was ineffective in completely blocking door-opening. Therefore, the dose was increased to 4 mg/kg for all remaining experiments and days. Midazolam-treated rats displayed little interest in helping and rarely opened the restrainer door (two openings in eight sessions at the 2 mg/kg dose, eight openings in 376 sessions at the 4 mg/kg dose). In the rare instances when an incompetent helper opened the door, the door remained open and the trial was not included in the data analysis.
Testing procedures
At the start of each session, the trapped rat was placed in the restrainer, the door closed, and the restrainer placed in the arena center. The free rat or rats were then placed in the arena. If the free rats did not open the restrainer door within 40 min, then the investigator opened the restrainer door halfway, to a 45° angle, facilitating door-opening by either a free rat or the trapped rat; this prevents the development of learned helplessness. Note that only openings that occurred before the 40-min mark were considered as door-openings. Regardless of whether the door was opened before or after the 40-min mark, rats always remained in the arena for the hour-long session. After each session, the arena and restrainer were washed with 1% acetic acid followed by isopropyl alcohol. Rats were tested once per day for 12 days.
If a trapped rat succeeded in opening the door from inside the restrainer, the trapped rat was placed immediately back in the restrainer with an acrylic blocker, which prevented his access to the door. If the free rat subsequently opened the door, the blocker was removed, allowing the trapped rat to exit the restrainer. The blocker was then used for that trapped rat on all following test days. In two instances, the trapped rat was the one to open the door but this was not realized at the time. Therefore, the trapped rat remained free and these two trials were not included in analyses.
Trios developed consistent, low-latency door-opening rapidly, and the group’s performance was maximal by day 4. In other words, no additional information could be gleaned by persisting with the same protocol beyond the first several days. Thus, rats were tested in the trios for only 6 days. Thereafter, they were tested individually with a cutoff of 30 min rather than the standard 40 min. Note that among the 48 control rats, the door-opening latency was between 30 and 40 min on days 7 to 12 on only two occasions of 208 openings (<1%). It is therefore unlikely that shortening these sessions to 30 min had a substantial impact on the number of openings among the trios in the second half of the testing sessions.
Video analysis
EthoVision tracking software (Noldus Information Technology Inc., Leesburg, VA) was used to track the rats’ movements at a rate of 15 frames per second in the arena under the +1 and −1 conditions. Rats were colored to allow for individual tracking. These data were used to calculate latencies.
Door-opening latencies
Time to door-opening was calculated as the latency, in minutes or seconds, until the restrainer door was opened. For rats that never opened, a cutoff time of 40 min was assigned. The 40-min cutoff was also used for trio rats on days 7 to 12, although the rats were only tested for 30 min. As explained above, latencies between 30 and 40 min were rare (2% of all 324 openings in the control rats and <1% of days 7 to 12 openings as above). Thus, using a 40-min latency for the trio rats tested individually allowed us to compare these rat sessions to those from other conditions with minimal impact.
The first day of opening was the first day on which the rat opened. A consecutive opening was defined as openings on two sequential days. Days when either an incompetent helper or trapped rat opened (n = 10 for confederate openings, n = 2 for trapped openings) were not included. Thus, if a rat opened for the first time on day 2 after an incompetent helper opening occurred on day 1, then the first day of opening would be assigned as day 1. Similarly, two openings interrupted by an incompetent helper or trapped opening were considered consecutive.
To analyze the latencies and days of first and third door-openings, only rats that opened at least once were included. For rats that opened once or twice but not three times, a latency of 40 was assigned to the third opening. In these cases, the day of the third opening was calculated upon which the rat could have opened for the third time, if testing had continued. Thus, for a rat that opened twice and was tested for 12 days, the third day of opening was given a value of 13.
Statistical analysis
Door-opening metrics were calculated per rat and then for each condition. All data are presented and illustrated as median and IQR (25th to 75th percentile). Signed rank sum test was used on paired observations to determine decrease in latencies. Kruskal-Wallis rank sum test was used to determine differences between groups in the number or latency of door-openings. Post hoc analyses were performed using Dunn’s test of multiple comparisons with Bonferroni correction. Effect size estimates were calculated following (21). All statistics were calculated in R, and post hoc analyses were performed using the PMCMR (Pairwise Multiple Comparison of Mean Rank sums) package.
Supplementary Material
Acknowledgments
We thank N. Molasky, M. Sol Bernardez Sarria, W. Tong, M. Nakamura, M. Barajas, and J. Woodruff Jr. for assisting in data collection and helpful discussions. Funding: M.C.J. was supported by The University of Chicago Pritzker School of Medicine Summer Research Program; J.L.H. and R.R.K. were supported by UChicago Neuroscience Metcalf Fellowships and J.L.H. was also supported by the Neuroscience Honors program. Additional support from UChicago Department of Neurobiology is gratefully acknowledged. Author contributions: J.L.H. and P.M. conceived the study; R.R.K. and P.M. conceived and designed the foreign confederate study; J.L.H., M.C.J., and R.R.K. ran the experiments and collected the data; J.L.H., M.C.J., Y.Y.V.S., and P.M. analyzed the data; J.H.C.J. and Y.Y.V.S. coded the shuffles; J.L.H., M.C.J., Y.Y.V.S., and P.M. wrote the manuscript; P.M. supervised the project. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data and R code are included in the Supplementary Materials. Additional data related to this paper may be requested from the authors.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/6/28/eabb4205/DC1
REFERENCES AND NOTES
- 1.M. Gansberg, 37 Who Saw Murder Didn’t Call the Police, The New York Times (27 March 1964).
- 2.J. Rasenberger, Kitty, 40 Years Later, The New York Times (8 Feb 2014).
- 3.B. Latané, J. M. Darley, The Unresponsive Bystander: Why Doesn't He Help? (Appleton-Century Crofts, 1970). [Google Scholar]
- 4.J. T. Cacioppo, L. Freberg, Discovering Psychology (Belmont, CA: Wadsworth Cengage Wadsworth, 2013), p. 668. [Google Scholar]
- 5.M. S. Gazzaniga, T. F. Heatherton, D. F. Halpern, Psychological Science (W. W. Norton & Co., 2010), pp. 556–558. [Google Scholar]
- 6.H. Gleitman, J. J. Gross, D. Reisberg, Psychology (W. W. Norton & Co., 2011), pp. 532–533. [Google Scholar]
- 7.D. Westen, Psychology: Brain, Behavior & Culture (John Wiley, 2002) pp. 637–638. [Google Scholar]
- 8.Lilienfeld S. O., Sauvigné K. C., Lynn S. J., Cautin R. L., Latzman R. D., Waldman I. D., Fifty psychological and psychiatric terms to avoid: A list of inaccurate, misleading, misused, ambiguous, and logically confused words and phrases. Front. Psychol. 6, 1100 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Philpot R., Liebst L. S., Levine M., Bernasco W., Lindegaard M. R., Would I be helped? Cross-national CCTV footage shows that intervention is the norm in public conflicts. Am. Psychol. 75, 66–75 (2020). [DOI] [PubMed] [Google Scholar]
- 10.Mason P., Shan H., A valence-free definition of sociality as any violation of inter-individual independence. Proc. Biol. Soc. 284, 20170948 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ben-Ami Bartal I., Decety J., Mason P., Empathy and pro-social behavior in rats. Science 334, 1427–1430 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Materials and methods are available as supporting material on Science Online.
- 13.Ben-Ami Bartal I., Rodgers D. A., Bernardez Sarria M. S., Decety J., Mason P., Pro-social behavior in rats is modulated by social experience. eLife 3, e01385 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Carvalheiro J., Seara-Cardoso A., Mesquita A. R., de Sousa L., Oliveira P., Summavielle T., Magalhães A., Helping behavior in rats (Rattus norvegicus) when an escape alternative is present. J. Comp. Psychol. 133, 452–462 (2019). [DOI] [PubMed] [Google Scholar]
- 15.Fischer P., Greitemeyer T., The positive bystander effect: Passive bystanders increase helping in situations with high expected negative consequences for the helper. J. Soc. Psychol. 153, 1–5 (2013). [DOI] [PubMed] [Google Scholar]
- 16.Levine M., Cassidy C., Brazier G., Reicher S., Self-categorization and bystander non-intervention: Two experimental studies. J. Appl. Soc. Psych. 32, 1452–1463 (2002). [Google Scholar]
- 17.Decety J., Bartal I. B., Uzefovsky F., Knafo-Noam A., Empathy as a driver of pro-social behavior: Highly conserved neurobehavioral mechanisms across species. Philos. Trans. R Soc. Lond. Biol. Sci. 371, 20150077 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kikusui T., Winslow J. T., Mori Y., Social buffering: Relief from stress and anxiety. Philos. Trans. R Soc. Lond. B Biol. Sci. 361, 2215–2228 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kiyokawa Y., Hennessy M. B., Comparative studies of social buffering: A consideration of approaches, terminology, and pitfalls. Neurosci. Biobehav. Rev. 86, 131–141 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Galef B. G., Whiskin E. E., 'Conformity' in Norway rats? Animal Behav. 75, 2035–2039 (2008). [Google Scholar]
- 21.Tomczak M., Tomczak E., The need to report effect size estimates revisited. An overview of some recommended measures of effect size. Trends Sport Sci. 21, 19–25 (2014). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/6/28/eabb4205/DC1