

Abstract
Quorum quenching (QQ), the obstruction of quorum sensing, is the most attractive way to break down the N-acyl-homoserine lactones (AHL) molecules. This work was focused at isolating AHL degrading bacteria from gastrointestinal tract of Oreochromis niloticus, with abilities appropriate for use as probiotic in aquaculture. The presence of an autoinducer inactivation (aiiA) homolog gene and AHL inactivation assay showed that Enterococcus faecium QQ12, which was one among the 20 isolates, could rapidly degrade synthetic C6-HSL in vitro and hampered violacein production by Chromobacterium violaceum. It had excellent biodegrading ability of natural N-AHL produced by Aeromonas hydrophila, suggesting that it can be used as a potential quencher bacterium for disrupting the virulence of A. hydrophila. It was susceptible to all the five antibiotics tried out. The isolate grew well at pH 3.0–7.0, was resistant to high level of bile salts (0–0.9%) and 0.5% of phenol. QQ12 also exhibited high degree of auto-aggregation and co-aggregation, confirming that it possessed good probiotic attributes. Goldfish fed diet incorporated with 108 and 1010 CFU g−1 of the QQ12 for 30 days showed 76.66–86.66% survival when challenged with A. hydrophila. The study indicates that Enterococcus faecium QQ12 could be used as a non-antibiotic feed additive in aquaculture to control bacterial diseases.
Keywords: Enterococcus faecium, N-acyl-homoserine lactones, Probiotic, Quorum sensing, Quorum quenching
Introduction
Aeromonas hydrophila, a Gram-negative, motile rod is regarded as one of the most important aquaculture pathogen causing motile aeromonas septicemia or ulcer disease in fish [1, 2]. This disease affects large number of tropical or ornamental fish. The unlimited use of antibiotics for the control of bacterial infection in aquaculture can lead to the development of antibiotic resistance in pathogenic bacteria. Alternative biocontrol strategies are therefore needed for sustainable development of the aquaculture industry. One such approach that is as equally functional as antibiotics is to impede the bacterial signaling pathways controlling the production of virulence factors.
Quorum sensing (QS) is a mechanism by which majority of microorganisms communicate with each other and make response collectively to a dynamic environment. For this purpose, they synthesize and secrete small chemical signal molecules such as acyl-homoserine lactones (AHLs) in Gram-negative bacteria, oligopeptides in Gram-positive bacteria, and autoinducer-2 (AI-2) in both Gram-negative and Gram-positive bacteria; the concentration of which could be recognized by bacterial specific detection system. This permits bacteria to understand about the surrounding cell density. When the critical threshold concentration of signal molecule is reached, it induces or represses the quorum sensing regulated gene expression. In bacteria, physiological functions like sporulation, antibiotic resistance, luminescence, biofilm formation, virulence, and motility are coordinated by QS systems [3–5]. In Aeromonas, expression of virulence factors such as adhesins, cytotoxins, hemolysins, lipases, and proteases is regulated through quorum sensing mechanism, which is mediated by signaling molecules N-acyl homoserine lactones. Disruption of these signaling molecules could be an effective strategy to control virulence in these bacteria.
Quorum quenching (QQ) refers to the process of interrupting QS by small molecule antagonists or signal degrading enzymes and has been regarded as an innovative biocontrol approach to attenuate bacterial pathogens [6] and is proven to be an alternative to conventional antibiotic control of infections in aquatic system [7, 8]. Quorum quenching enzymes, including lactonase, acylase, oxidoreductase, and paraoxonase, have been recognized in quorum sensing and non-quorum sensing microbes [9–11]. As a more sustainable alternative to antibiotic, the use of probiotics too is gaining acceptance for the control of bacterial pathogens in aquaculture. Probiotics eliminate pathogens by several mechanisms such as enhancement of the epithelial barrier, modulation of the immune system, increased adhesion to intestinal mucosa, and concomitant inhibition of pathogen adhesion, competitive exclusion of pathogenic bacteria by elimination of available bacterial receptor sites, production and secretion of antimicrobial substances and selective metabolites, and competitive depletion of essential nutrients. The health benefits associated with application of probiotics in fish are related to nutrient contribution, growth promotion, improvement of host immune system, and increased survival [12, 13]. These beneficial microorganisms have been discovered, characterized, and used in aquaculture during the last three decades. In this context, application of signal degrading (quorum quenching) bacteria that can at the same time act as probiotic would be a unique dual strategy to control antibiotic-resistant pathogens and to support the host in a positive manner. Recently, some research works have been reported in quorum quenching bacteria isolated from gastrointestinal tract of aquatic animals [8, 14–19]. It has also been shown that probiotic bacteria such as Enterococcus durans and Bacillus spp. inactivate the signal molecules of pathogenic bacteria by enzymatic action [20, 21].
Enterococcus faecium is a Gram-positive, alpha-hemolytic, or non-hemolytic lactic acid bacteria generally occurring as a commensal in the gastrointestinal tract of humans and animals. It has been recognized as a good probiotic for a long time and is widely used in the field of aquaculture. Though its probiotic activity and safe utilization has been well proven [22–24] in aquatic animals, currently there are no studies on the quorum quenching potential of this bacteria. In the present work, E. faecium strain was isolated from the gastrointestinal tract of healthy Nile tilapia (Oreochromis niloticus) and tested for both quorum quenching and probiotic potential against pathogenic A. hydrophila. Tolerance to acid, bile, and phenol, surface properties, and antibiotic resistance are considered as the major attributes that give a probiotic the potential to remain alive in the bile-rich and extremely acidic gastrointestinal tract of the animals [25, 26]. These essential features were screened in E. faecium in the present study. By analyzing both quorum quenching attributes and probiotic properties of E. faecium strain, we were able to develop an effective biocontrol strategy to protect goldfish against A. hydrophila infection.
Materials and methods
Bacterial strains and growth conditions
CV026, a mini-Tn5 mutant derived from Chromobacterium violaceum was used as a biosensor to find out the presence of exogenous AHLs (C6-HSL). It was purchased from Microbial Culture Collection (MCC), NCCS, Pune, India. CV026 cannot synthesize AHL, but it can detect and respond to exogenous AHLs with acyl chain of four to eight carbons, by production of the purple-colored violacein pigment. It was grown in Luria Bertani (LB) medium at 28 °C supplemented with 50 μg mL−1 of kanamycin. The target fish pathogen A. hydrophila used in this study was provided by the National Bureau of Fish Genetic Resources (ICAR, Kochi, India). It was grown in LB broth (pH 7.2 ± 0.2) at 30 °C overnight. Escherichia coli DH5α (Promega), also grown in LB medium at 37 °C, served as negative control in AHL inactivation assay. All media used for AHLs assay were buffered with 50 mmol L−1 3-[N-morpholino] propane sulfonic acid (MOPS) to pH 6.8, to prevent spontaneous degradation of AHLs.
Isolation and identification of quorum quenching bacteria from Oreochromis niloticus gut
O. niloticus were collected from aquaculture farms in Ernakulum district, Kerala, India. The fish were dissected aseptically (dissection was performed under tricaine methanesulfonate (Sigma) (100 mg L−1 anesthesia) and the gastrointestinal tract was removed as quickly as possible. A gut homogenate was prepared in sterile physiological saline ((pH 7.4) 0.85% NaCl). Samples were then enriched in minimal medium (KG medium) with AHL as the sole source of carbon and nitrogen. One hundred microliters of the homogenate was inoculated into 100 mL flask containing 10 mL of KG medium (pH 6.8) with 500 μg L−1 of C6-HSL, as previously described [27] and incubated at 28 °C. After 24 h, 1 mL of culture was transferred to fresh C6-HSL containing KG medium for enrichment culturing. At the third time enrichment cycle, a diluted suspension was plated onto LB agar. After 48 h of incubation, bacteria with different colony morphology were picked and streaked on LB agar. Pure colonies were obtained by repeated streaking on LB agar. The selected bacterium was identified following Bergey’s Manual of Systematic Bacteriology [28] in accordance with different biochemical and physiological characteristics. Species level identification was carried out by 16S rDNA sequencing (SciGenom Labs, India) using universal primers 27F and 1492R and analyzed using NCBI nucleotide database. Antibiotic susceptibility of the isolate was checked by disc diffusion method as stated by the guidelines of the Clinical and Laboratory Standard Institute [29]. The antibiotic discs used in this test included ampicillin (10 μg), erythromycin (15 μg), penicillin G (10 U), vancomycin (30 μg), and tetracycline (30 μg).
Screening of quorum quenching activity
PCR amplification of aiiA homolog gene
Initially, the quorum quenching activity of all isolates was checked by screening for the presence of aiiA (autoinducer inactivation homolog) gene by PCR. Total DNA was extracted using HiPurA bacterial genomic DNA purification Kit (Himedia, India). The forward and reverse primers used were aiiA F (5′-A T G G G A T C C A T G A C A G T A A A G A A G C T T T A T-3′) and aiiA R (5′-G T C G A A T T C C T C A A C A A G A T A C T C C T A A T G-3′) respectively. PCR amplification was performed in a thermal cycler (MJ MINI, Biorad, USA), in 0.2 mL reaction tube consisting of 25 μL total reaction volume containing 9 μL nuclease free water, 12.5 μL GoTaq® Colorless Master Mix 2X (Promega, USA), 1.25 μL (10 μM) of each primer, and 1 μL of template DNA (100 ng). The reaction consisted of an initial denaturation of 94 °C for 10 min, followed by 30 cycles of 94 °C for 30 s, 52 °C for 30 s, 72 °C for 1 min, and a final extension of 72 °C for 5 min. Samples electrophoresed in 1.5% agarose gel at 70 V were visualized using gel documentation system (Biorad, USA). The amplified product was sequenced (Agrigenome lab, India) and the sequence was analyzed using NCBI nucleotide database.
Whole-cell AHL inactivation assay
The whole-cell AHL inactivation assay was carried out as previously reported [30] with minor modifications. The isolates showing the presence of aiiA homolog gene grown overnight at 30 °C in MRS media were centrifuged at 2800g for 10 min at 4 °C. Cell pellets were washed two times in 100 mM PBS (pH 6.8) and resuspended in the same buffer to get OD 600 of 1.0 (BIOPHOTOMETER, Eppendorf, Germany). 10 μg μL−1 C6-HSL (a synthetic AHL, Sigma-Aldrich, India) in absolute ethanol was transferred to individual sterile microcentrifuge tube and dried by evaporation under aseptic condition. Each of the bacterial cell suspensions in PBS was added to rehydrate AHL to the final concentration of 0.1 μg μL−1. The mixtures were then incubated at 30 °C with gentle shaking for 12 h. C6-HSL inactivation was assessed at 3 h, 6 h, and 12 h using CV026 as biosensor. Heat-denatured reaction mixtures (10 μL) at abovementioned time periods were loaded into the wells of LB agar bioassay plate overlaid with the biosensor CV026 and incubated at 28 °C for 24 h. E. coli strain DH5α served as negative control. Absence of violacein (purple zone) shown by CV026 indicated AHL degradation. The assay was carried out in triplicate.
AHL degradation with culture supernatant
To find out whether the quorum quenching factor is released out of the cell or is bound to cell, an in vitro assay was carried out as previously described by Chu et al. [20] with minor modification. Selected quorum quenching isolate (QQ12, the isolate showing strong and rapid AHL degrading activity) grown overnight at 30 °C in MRS medium was centrifuged for 10 min at 5500g and the filter-sterilized supernatant of the overnight culture was taken for testing the AHL degrading activity. One hundred microliters of the supernatant was mixed with an equal volume of 100 mM PBS (pH 6.8) containing 0.2 μg μL−1 C6-HSL. Following that, the reaction mixture was incubated at 30 °C for 24 h with gentle shaking, followed by incubation at 95 °C for 5 min to stop the reaction. Ten microliters of the reaction mixture was loaded into the well of a LB agar plate seeded with the biosensor CV026 and incubated at 28 °C for 24 h. The assay was carried out in triplicate.
Degradation of N-AHL produced by Aeromonas hydrophila
Fish pathogen A. hydrophila was inoculated in 10 mL LB medium and incubated at 30 °C for 24 h. Bacterial cells were removed by centrifugation at 16200g for 5 min at 4 °C. Filter-sterilized cell-free culture supernatant was added to equal volume of fresh LB medium and QQ12 was inoculated in this medium. Bacterial culture was incubated at 30 °C for 48 h and AHL inactivation was assessed at 0 h and 48 h using CV026 as biosensor. The assay was carried out in triplicate.
Screening of probiotic activity
Bile salt and acid tolerance
The isolate QQ12 was tested for bile salt tolerance and survival in acidic condition. Bacterial strain was grown overnight in MRS media and 0.1 mL of culture suspension (absorbance was adjusted to 0.02) was inoculated into tubes containing 10 mL of autoclaved MRS media with 0%, 0.3%, 0.6%, and 0.9% bile salt (Himedia, India). The inoculated tubes were incubated at 30 °C for 18 h and the absorbance at 600 nm was measured to evaluate growth. To determine acidic tolerance of QQ12, 0.1 mL of actively grown overnight culture at 30 °C in MRS medium was transferred to autoclaved MRS broth adjusted to pH 1–7 with HCl (Sigma, India), which were then incubated at 30 °C for 18 h followed by measurement of absorbance at 600 nm. Both assays were carried out in triplicate.
Phenol tolerance assay
To check the phenol tolerance, actively growing overnight culture of QQ12 was inoculated into MRS media with concentration of 0.2% and 0.5% phenol or without phenol. Cell growth of the isolate was evaluated after 18 h of incubation at 30 °C at A600 nm. The assay was carried out in triplicate.
Auto-aggregation and co-aggregation assays
To evaluate the probiotic potential of QQ12, auto-aggregation and co-aggregation rate were measured according to Del Re et al. [31] with some modifications. Isolate was grown for 18 h at 30 °C in MRS media. The cells were harvested by centrifugation at 2800g for 15 min at 4 °C, washed twice with PBS (pH 7.2) and resuspended in the same buffer. Initial absorbance (A600 nm) was adjusted to 0.2. Cell suspension (5 mL) was mixed by vortexing for 10 s and the same suspension was left to rest for 5 h at room temperature without vortexing. Auto-aggregation of cell suspension was determined by taking 0.1 mL of the upper suspension at every 1 h interval to another tube with 4.9 mL of PBS and the absorbance of suspension at 600 nm was recorded. Cell auto-aggregation was measured by decrease in absorbance and auto-aggregation percentage is demonstrated as 1 − (At/A0) × 100, where At represents the absorbance at time t = 1, 2, 3, 4, or 5 h and A0 the absorbance at t = 0.
The procedure for making the cell suspension for co-aggregation assay was the same as that for auto-aggregation assay. QQ12 prepared as mentioned above was mixed with equal volume (2 mL) of the culture of fish pathogen A. hydrophila and incubated at room temperature without agitation. In control tubes, 4 mL of each bacterial suspension alone was added. After 5 h of incubation, the absorbance (A) at 600 nm of the suspensions was measured. Co-aggregation percentage was calculated using the equation of Handley et al. [32]. Co-aggregation % = [(Apathog + AQQ)/2 − (Amix)/(Apathog + AQQ)/2] × 100, where Apathog and AQQ constitute the absorbance in the tubes containing solely the pathogen or the quorum quenching bacteria (control tubes), respectively, and Amix represents the absorbance of the mixture. Auto-aggregation and co-aggregation assays were performed in triplicate.
In vivo study
Maintenance of experimental fish
To confirm the probiotic activity of QQ12 in fish, uniform-sized fingerlings of goldfish C. auratus (Linnaeus, 1758) were initially acclimatized in tanks of 300 L capacity for 3 weeks before starting the experiment. The fish were healthy and exhibited no signs of disease (confirmed through the active swimming movement, examination of gills, fins, and skin). The pathogen-free status of the fish was confirmed by determination of total viable aerobic count by standard plate count method. During this period, the fish were fed twice daily with commercial feed. All tanks were provided with proper aeration (dissolved oxygen level was maintained at 5–7 ppm) and water temperature was maintained at 26 ± 1 °C. pH was recorded at regular intervals and maintained at 7.2 ± 0.1.
Determination of LD50
Before conducting the challenge study, the infectious dose of A. hydrophila was selected by 50% lethal dose (LD50) determination. Five groups of six gold fish, each in triplicate (3.34–4.32 g weight and 85.35–94.40 mm length) were introduced into tanks of 50 L capacity. Four groups received intraperitoneal injection of 0.1 mL of 1 × 102, 1 × 104, 1 × 106, and 1 × 108 cells of A. hydrophila, respectively, while the fifth group was maintained as control and injected with 0.1 mL of PBS alone. Fish were observed up to 7 days for abnormal clinical appearance. Dead fish were removed immediately for bacteriological examination.
Safety of the QQ12
The pathogenicity of the QQ12 was also ascertained before preparing probiotic feed. Two groups of six goldfish (3.34–4.32 g weight and 85.35–94.40 mm length), were challenged with 0.1 mL of PBS with 1.0 × 107 cells and 1.0 × 1010 cells of QQ12 respectively by intraperitoneal injection. Each group was maintained in triplicate. The control group fish received 0.1 mL of PBS alone. Fish were observed for mortality for 7 days. During this period, behavior of fish was recorded daily.
Preparation of probiotic feed and feeding trial
The probiotic feed was prepared by inoculating the QQ12 in MRS broth and incubated at 30 °C for 24 h. Bacterial culture with different cell densities (104 CFU mL−1, 106 CFU mL−1, 108 CFU mL−1, and 1010 CFU mL−1) were prepared and harvested at frequent time intervals by centrifugation at 1000g for 15 min at 4 °C. After centrifugation, bacteria were washed twice with PBS (pH 7.2) and resuspended in PBS, maintaining the cell densities. This suspension was added at the rate of 1 mL of culture g−1 of commercial feed to incorporate 104 cells g−1 feed, 106 cells g−1 feed, 108 cells g−1 feed, and 1010 cells g−1 feed respectively. A binder (Brand: Aqua one, Salem Microbes Private limited, India) was used 1 mL 10 g−1 feed. Binder alone was added in control feed. After proper mixing of the ingredients, the feeds were air dried and stored in screw capped glass bottles at room temperature until used. To ensure a required probiotic level in the supplemented feed, new probiotic diets were made on a weekly basis. Five groups of 10 goldfish each, C. auratus were introduced into glass tanks of 50 L capacity. Four groups were fed with 104 CFU g−1, 106 CFU g−1, 108 CFU g−1, and 1010 CFU g−1 of probiotic diet respectively, while the fifth group was maintained as control group. Feeding was done two times daily at the rate of 3% of the body weight of C. auratus for 30 days. The fish of all groups were fed with respective feed in triplicate. Continuous aeration and water flow were maintained in all glass tanks. During the study period, activity and behavior of the fish were monitored and recorded daily.
Bacterial challenge study
Control and probiotic fed fish were challenged (10 numbers in each group, maintained in triplicate) via intraperitoneal injection with 0.1 mL of 1 × 106 cells of A. hydrophila. External signs of infection, behavioral abnormalities, and mortalities, if any, were recorded for 2 weeks. Dead fish were removed immediately for bacteriological examination. Bacterial isolation was carried out from hemorrhagic and ulcerative lesions and visceral organs.
Study of QQ12 survival in goldfish intestine
Thirty five samples of pathogen-free goldfish were used in this experiment. Goldfish were fed for 7 days with feed containing QQ12 (104 cells g−1 per day). Feeding with probiotic diet was then discontinued, and the average concentration of QQ12 in the goldfish intestine was determined 0 (control), 1, 2, 3, 4, 5, 6, and 7 days after the probiotic feeding period (by standard plate count method).
Statistical analysis
All the experiments were performed in triplicate and the results were expressed as mean ± standard deviation (SD) of triplicates. Data were statistically processed by one way ANOVA using SPSS (Version 21.0). Statistically significant differences were defined at p < 0.01.
Results
Isolation and identification of quorum quenching bacteria
Twenty bacterial isolates in the KG medium containing C6-HSL were screened. Finally, one representative isolate showing strong AHL degrading activity was selected. It was characterized at the physiological, biochemical, and morphology levels. Based on biochemical properties, the strain showed close resemblance to Enterococcus spp. To further identify the strain, 16S rDNA sequencing was carried out. Results showed the QQ12 shared 100% homology with Enterococcus faecium species (GenBank accession number KM257657). The isolate QQ12 was susceptible to five tested antibiotics (ampicillin, erythromycin, penicillin G, tetracycline, and vancomycin) (Table 1).
Table 1.
Antibiotic resistances of Enterococcus faecium QQ12
| Zone of growth inhibition diameter (mm) | ||||
| AM | E | P | TE | VA |
| 29 ± 0.22 | 23 ± 0.29 | 17 ± 0.03 | 26 ± 0.05 | 25 ± 0.48 |
| Interpretative criteria (CLSI) zone inhibition diameter (mm) | ||||
| AM | E | P | TE | VA |
|
S-≥ 17 I-Nil R-≤ 16 |
S-≥ 23 I-14–22 R-≤ 13 |
S-≥ 15 I-Nil R-≤ 14 |
S-≥ 19 I-15–18 R-≤ 14 |
S-≥ 17 I-15–16 R-≤ 14 |
AM ampicillin, E erythromycin, P penicillin G, TE tetracycline, VA vancomycin, S sensitive; I intermediate, R resistant
The mean of three values of zone of growth inhibition of each antibiotic are presented along with ± SD
Detection of aiiA homolog gene
Among the 20 bacterial isolates that were initially screened for presence of aiiA homolog gene by PCR, six isolates with aiiA homolog gene were observed. The expected amplicon size of approximately 800 base pairs was detected (Fig. 1). The sequence of the amplified product (Accession No.: BankIt2290444 BSeq#1MN803161) showed 100% homology with aiiA gene from other bacterial isolates.
Fig. 1.
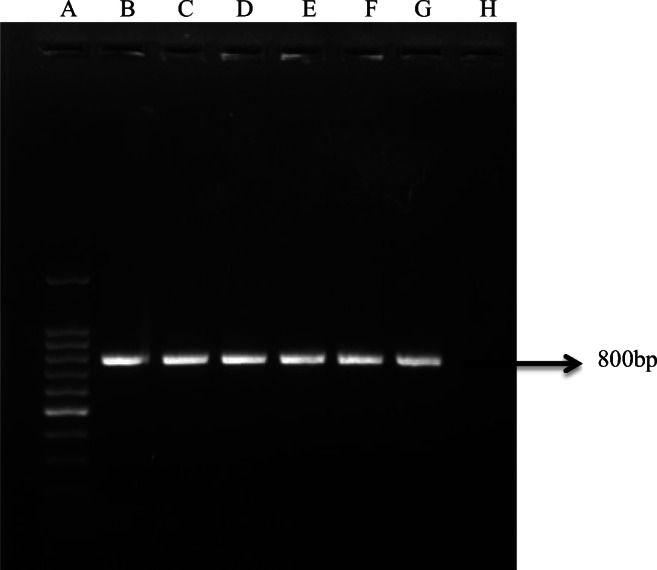
PCR detection of aiiA homolog gene. Lane A, 100 bp DNA ladder (Promega); lane B, Enterococcus faecium QQ12; lane C–G, other five isolates (QQ8, QQ10, QQ15, QQ17, and QQ20); lane H, negative control. Arrow shows the expected amplicon size of approximately 800 bp
Whole-cell AHL inactivation assay
Whole-cell AHL inactivation assay was performed to evaluate the AHL degrading efficiency of six QQ isolates (QQ8, QQ10, QQ12, QQ15, QQ17, and QQ20) that harbored aiiA homolog gene. The results (given in Table 2) showed that only QQ12 (identified as Enterococcus faecium QQ12) out of the six isolates tested was able to degrade C6-HSL almost completely within 6 h of incubation showing rapid quorum quenching activity (Fig. 2, A). Figure 2 B showed the AHL degradation of E. faecium QQ12 after 12 h of incubation. Only leftover C6-HSL was detected by CV026 when the reaction was ceased after incubation for 3 h (Fig. 2, D). No visible AHL degradation was noticed in DH5α that served as negative control (Fig. 2, C). Results showed that there was no visible AHL degradation observed in other four isolates (QQ8, QQ10, QQ15, and QQ17) within 12 h of incubation (Table 2). One isolate (QQ20) showed the potential to degrade C6-HSL partially within 12 h of incubation (Table 2). Finally, Enterococcus faecium QQ12 which showed the potential to degrade C6-HSL rapidly and completely within 6 h of incubation was selected for further studies. The supernatant of QQ12 had no AHL inactivating activity, and the diameter of the purple-pigmented zone had no remarkable difference with that of negative control DH5α well (data not shown). In order to confirm AHL degrading activity of QQ12, crude cell–free culture supernatant of A. hydrophila as natural N-AHL was used instead of synthetic C6-HSL. Complete degradation of natural N-AHL after 48 h incubation with QQ12 was observed (data not shown). No AHL degradation was observed and presence of violacein (purple zone) was shown by CV026 at 0 h incubation.
Table 2.
AHL degrading activity of QQ isolates
| Isolates | CV026 (C6-HSL) | ||
|---|---|---|---|
| 3 h | 6 h | 12 h | |
| QQ8 | N | N | N |
| QQ10 | N | N | N |
| QQ12 | P | C | C |
| QQ15 | N | N | N |
| QQ17 | N | N | N |
| QQ20 | N | N | P |
N no AHL degradation, P partial AHL degradation, C complete AHL degradation
Only one isolate (QQ12) showed complete C6-HSL degradation within 6 h of incubation
Fig. 2.
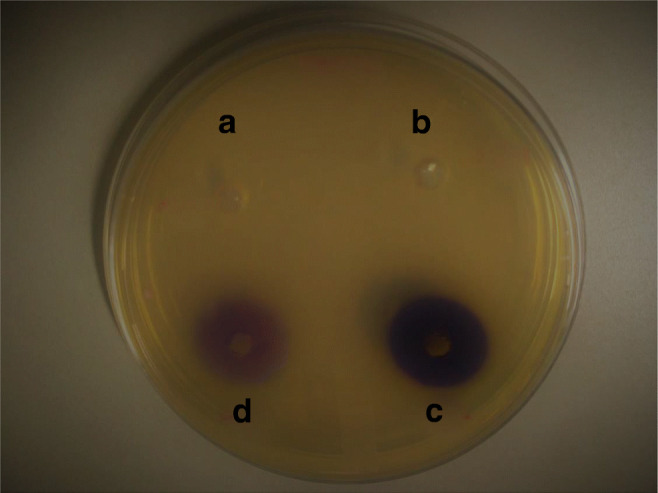
AHL degrading activity of Enterococcus faecium QQ12. QQ 12 was incubated with C6-HSL for 3 h (D), 6 h (A), and 12 h (B). Escherichia coli DH5α (C) (negative control). Pigment formation indicates the presence of C6-HSL; degradation of C6-HSL is evident by loss of pigment formation on the biosensor lawn
Bile salt, pH, and phenol tolerance of E. faecium QQ12
E. faecium QQ12 was checked for bile salt, pH, and phenol tolerance. E. faecium QQ12 grown successfully in all tested concentrations of bile (0–0.9%). There were no significant differences in optical density between control (without addition of bile salt) and E. faecium with different concentrations of bile salt after 18 h of incubation. This data suggests that E. faecium QQ12 is resistant to high bile salt concentration (Fig. 3 a). pH tolerance studies showed that E. faecium QQ12 grew at pH 3 or above but did not grow in conditions less than pH 3 (Fig. 3 b). The isolate grown well at 0–0.5% of phenol in MRS media (Fig. 3 c).
Fig. 3.
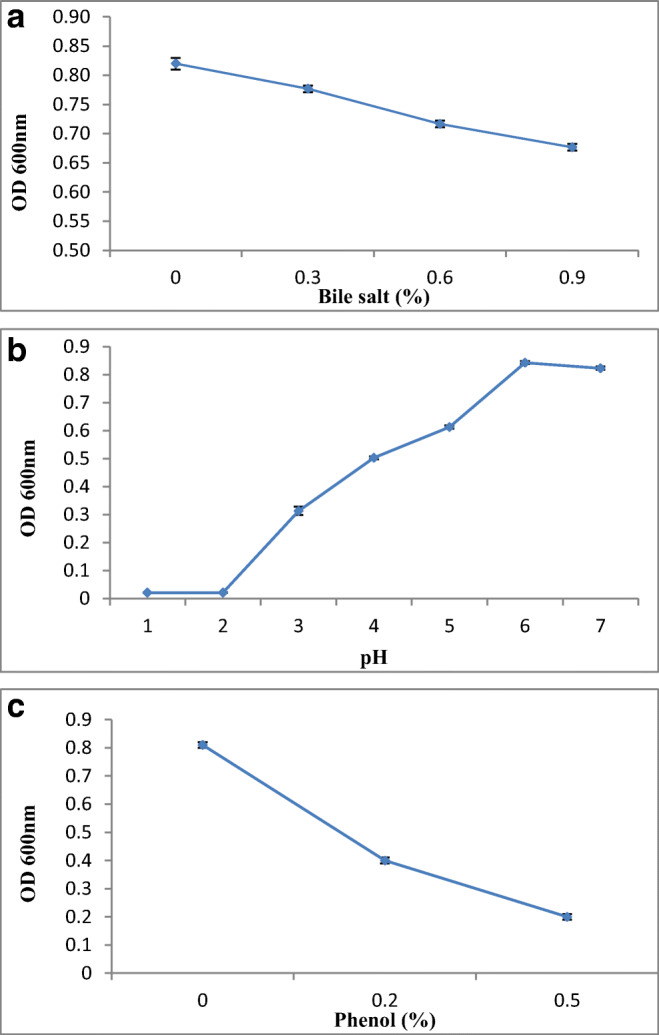
Effect of bile salt (a), pH (b), and phenol (c) on the survival of Enterococcus faecium QQ12 at 30 °C. To check bile salt, pH, and phenol resistance, the growth of E. faecium QQ12 in MRS medium for 18 h was determined by measuring OD at 600 nm after adjusting the culture media to the specific pH, salt, and phenol concentration. Values are mean ± SD of three different observations
Auto-aggregation and co-aggregation assays
The capability of E. faecium QQ12 to adhere to the intestinal cell wall was evaluated by auto-aggregation and co-aggregation assays. The result showed that E. faecium QQ12 had excellent auto-aggregation property (72.91 ± 0.06%) and aggregation values increased with time (Fig. 4). E. faecium QQ12 also exhibited very good co-aggregation ability after 5 h of incubation with A. hydrophila, during which 50.6 ± 0.04% of QQ12 co-aggregated with A. hydrophlila (data not shown).
Fig. 4.
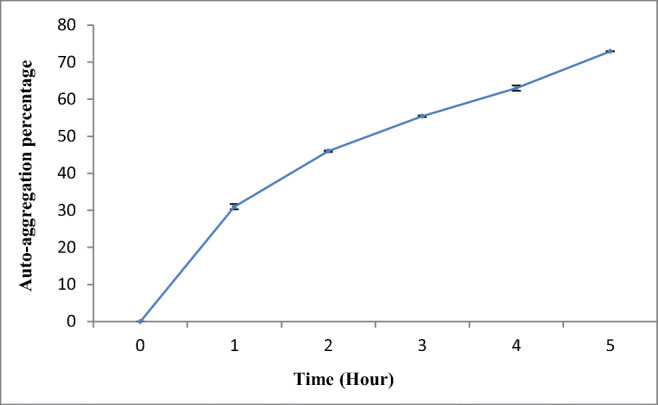
Auto-aggregation rate of quorum quenching isolate Enterococcus faecium QQ12. Aggregation percentage increased every 1 h and highest rate was observed at 5 h. Values are mean ± SD of three different observations
Determination of LD50 for bacterial challenge
The administration of A. hydrophila at 1 × 102 cells per fish did not show any potential adverse effect on fish. At higher doses, abnormal movement and loss of balance was observed 24 h after the injection. 50% mortality was observed by the administration of 1 × 106 cells per fish within 2 days, while in 1 × 108 cells per fish, 100% mortality was observed. A. hydrophila could be isolated from dead fish. Therefore, 1 × 106 cells per fish was selected as the dose for bacterial challenge.
Safety of the E. faecium QQ12
The administration of E. faecium QQ12 even at the concentration of 1 × 1010 cells per fish did not result in any unfavorable effect on fish activity. All fish were clinically healthy and behaved no different from the control group, suggesting that the isolate E. faecium QQ12 is not pathogenic to fish.
Experimental challenge with A. hydrophila
The administration of QQ12 diet (E. faecium) afforded effective protection against experimental A. hydrophila infection. In control group, following challenge with A. hydrophila, all fish showed severe skin lesions and 50% mortality was observed in 2 days. One fish each died in 104 CFU g−1 feed and 106 CFU g−1 feed in 2 days and some of the fish in both these treatments showed mild skin lesions and hemorrhages. In contrast, during the same time, there was no mortality in the two groups fed with QQ12 diet of 108 CFU g−1 feed and 1010 CFU g−1 feed. At the end of 2 weeks, the highest survival rate was noticed in groups of fish fed with 108 CFU g−1 (76.66%) and 1010 CFU g−1 (86.66%) probiotic diet. ANOVA showed that there was significant difference (p ≤ 0.01) in the survival rates among different concentrations. Post hoc analysis using Duncan’s Multiple Range Test grouped the concentrations into three homogenous groups viz.: (1) control (had only 13.33% survival), (2) 104 CFU g−1 and 106 CFU g−1 probiotic feed (had 43.33% survival), and (3) groups fed with 108 CFU g−1 (76.66% survival) and 1010 CFU g−1 (86.66% survival) (Table 3). A. hydrophila was isolated from hemorrhagic lesions of both dead and survived fish.
Table 3.
Survival percentages of Carassius auratus (fed with different concentrations of probiotic diet) 2 weeks after the experimental infection with Aeromonas hydrophila by intraperitoneal injection
| Probiotic concentration | Survival percent |
|---|---|
| Control | 13.33 ± 5.8 |
| 1 × 104 CFU | 43.33 ± 5.8 |
| 1 × 106 CFU | 43.33 ± 5.8 |
| 1 × 108 CFU | 76.66 ± 5.8 |
| 1 × 1010 CFU | 86.66 ± 5.8 |
Values are mean ± SD of triplicate observations
Study of E. faecium QQ12 survival in goldfish intestine
Goldfish were fed with feed containing E. faecium QQ12 for 7 days; bacterial feeding was then discontinued. The number of E. faecium QQ12 in goldfish intestine 1 day after discontinuation of probiotic feeding was 7.44 ± 1.02 × 104 CFU per individual, which was not significantly (p > 0.05) different from day 0 (control) (Table 4). However, the number of E. faecium QQ12 in goldfish intestine decreased by about 40.55% (4.76 ± 2.05 × 104 CFU per individual) from the control by day 2, by 62% (3.05 ± 3.25 × 104 CFU per individual) by day 3, by 76.50% (1.87 ± 1.25 × 104 CFU per individual) by day 4, by 86% (1.11 ± 3.22 × 104 CFU per individual) by day 5, by 88.2% (9.5 ± 1.78 × 103 CFU per individual) by day 6, and by 90.0% (8.02 ± 2.87 × 103 CFU per individual) by day 7 (Table 4).
Table 4.
The number of Enterococcus faecium QQ12 in Carassius auratus intestine after 7 days of feeding and after 1, 2, 3, 4, 5, 6, and 7 days following discontinuation of probiotic feeding
| Time after discontinuation of Enterococcus faecium QQ12 feeding (day) | Number of Enterococcus faecium QQ12 in Carassius auratus (CFU/individual) | Percent decrease |
|---|---|---|
| 0 | 8.0 ± 3.07 × 104 | 0 |
| 1 | 7.44 ± 1.02 × 104 | 7.75 |
| 2 | 4.76 ± 2.05 × 104 | 40.55 |
| 3 | 3.05 ± 3.25 × 104 | 62 |
| 4 | 1.87 ± 1.25 × 104 | 76.5 |
| 5 | 1.11 ± 3.22 × 104 | 86 |
| 6 | 9.5 ± 1.78 × 103 | 88.2 |
| 7 | 8.02 ± 2.87 × 103 | 90 |
Values are mean ± SD of triplicate observations
Discussion
The present study focused on fish gut bacteria Enterococcus faecium QQ12 that exhibited both probiotic and quorum quenching ability. To the best of our knowledge, there are hardly any reports demonstrating the quorum quenching ability of probiotic E. faecium. In this study, synthetic N-hexanoyl-L-homoserine lactone (C6-HSL) was used as a test compound. The AHL degrading ability of isolated bacteria was initially screened by PCR amplification of aiiA homolog gene, encoding the AHL inactivation enzyme. Previous studies by Dong et al. [6] revealed that the aiiA gene is responsible for AHL degradation in Bacillus sp. and is conserved between Gram-positive and Gram-negative bacteria. As the presence of aiiA homolog gene can only predict but does not confirm the AHL degrading function, the whole-cell inactivation assay was also carried out. Enterococcus faecium QQ12, which synthesize AHL degrading enzyme, was selected based on its ability to stop AHL-dependent violacein production by the bio indicator CV026. In whole-cell in vitro AHL inactivation assay, nearly all synthetic C6-HSL was degenerated after incubating with E. faecium QQ12 for 6 h, indicating rapid and strong QQ activity. Similar result was observed in a study by Chu et al. [20] in which the isolate QSI-1 (Bacillus spp.) degraded C6-HSL completely within 6 h in whole-cell AHL inactivation assay. The supernatant of E. faecium QQ12 could not inactivate C6-HSL, indicating that the degrading enzyme is not discharged out of the cell that agrees with the reports by Molina et al. [33], Martinez et al. [34], and Chu et al. [20] suggesting that the signaling molecules diffuse into the quorum quenching bacterial cells where molecule inactivation takes place. The efficacy evaluation of the E. faecium QQ12 for degradation of natural N-AHL produced by A. hydrophila resulted in the complete inactivation of N-AHL within 48 h of incubation. C4-HSL and C6-HSL are the major autoinducers produced by A. hydrophila [35] and can be detected by CV026 in AHL inactivation assay. This result suggests that E. faecium QQ12 can be used as potential quencher bacterium in aquatic environment very effectively for inhibiting the virulence of A. hydrophila.
The implementation of quorum quenching property to choose a new probiotic gives an alternative to antibiotics to control pathogenic bacteria in aquaculture [7]. The results of the present study showed that, in addition to possessing excellent quorum quenching properties, E. faecium isolated from tilapia gut has very good probiotic properties such as bile salt, acid, and phenol resistance, auto-aggregation, co-aggregation, and antibiotic sensitivity. In the present study, E. faecium QQ12 tested for bile salt tolerance exhibited survival even in 0.9% bile salt at 18 h of incubation and there were no significant differences in optical density between control and different concentrations of bile salt treatment suggesting that it has the capacity to withstand the conditions in fish gut. Many reports are found to describe the bile salt tolerance of E. faecium [36–39]. When a bacterium prefers a pH above 5.5, but able to grow at pH 5.5 or below, it can be classified as acid tolerant. Fish gastrointestinal pH shows great variation among species with a range of 1.47 to 5.12 and the lowest value observed was 1.18 [40]. However, such extreme low pH is transient. The pH value raises to 3 and above in the presence of food [25]. In the present study, we found that E. faecium QQ12 grew at pH 3 or above. This result indicates that the QQ isolate E. faecium given as a probiotic diet will be able to survive the harsh conditions of the gut environment and colonize the intestinal tract, thereby will be capable of imparting their benefits. In this study, the isolate could also persist well at 0.5% of phenol in MRS media. Phenol may be synthesized in the intestine by bacterial deamination of various aromatic amino acids obtained from dietary or endogenously derived protein [41]. Studies on different animal models reveal that phenol has a bacteriostatic effect against gut bacteria [42]. Since probiotics should withstand the harsh gut environment, tolerance to phenol is considered as a mandatory probiotic property.
Auto-aggregation and co-aggregation properties are considered as major characteristics of probiotic bacteria. Assessment of auto-aggregation and potential to co-aggregate with harmful intestinal pathogens can be used for initial evaluation and selection of the best probiotic strain. Bao et al. [43] stated that auto-aggregation property of probiotic bacteria is responsible for the bacterial adhesion on to the intestinal cell wall; an essential feature for colonization of probiotic strains in the gastrointestinal tract. Jelena and Natasha [44] tested the auto-aggregation property of 12 lactic acid bacteria and among them, E. faecium and L. paracasei strains had the highest degree of auto-aggregation (44% and 50%, respectively). In the present study, the E. faecium QQ12 exhibited high degree of auto-aggregation (72.91 ± 0.06%). Co-aggregation ability, also called “barrier effect” of probiotics, might become an obstacle that prevents colonization of pathogenic bacteria in the gastrointestinal tract [45]. In order to measure interbacterial adherence, Blagoeva et al. [46] examined the co-aggregation of 34 LAB strains with two pathogenic strains (Salmonella sp. NBIMCC 1425 and Listeria monocytogenes NBIMCC 8669). Among them, Lactobacillus fermentum and Enterococcus faecium showed highest rate of co-aggregation (27.31% and 24.03% respectively) with Salmonella sp. NBIMCC 1425. In the present study, E. faecium QQ12 demonstrated excellent co-aggregation ability (50.6 ± 0.04%) with A. hydrophila. This ability of E. faecium QQ12 might be a reason for the prevention of colonization of A. hydrophila in the goldfish gastrointestinal tract.
The antibiotic sensitivity of bacteria is considered as one of the important strategy to develop safe probiotic products for aquaculture applications. Antibiotic resistance in probiotic bacteria may result in active transfer of antibiotic-resistant genes from probiotics to other intestinal microflora and finally to opportunistic pathogens that reside in the same harsh environment. The bacteria which are used as probiotics should be free from acquired antibiotic resistance genes to prevent lateral spread of resistance [47]. In the present study, E. faecium QQ12 exhibited susceptibility to all five antibiotics tested. This result supports the possibility of the E. faecium QQ12 to be developed as probiotic.
The goldfish, C. auratus, has high vulnerability to A. hydrophila. The QQ isolate E. faecium QQ12 isolated in the present study had no harmful effect on goldfish and the probiotic diet supplemented with 108 CFU and 1010 CFU for 30 days protected the fish when challenged with A. hydrophila. Lowest (13.33%) survival was observed in the control (fed basal diet) compared with probiotic fed groups. Highest survival of fish was recorded in the group fed with probiotic diet of 1010 CFU g−1 feed and 108 CFU g−1 feed (86.66% and 76.66% respectively). Statistical analysis showed that there was no significant difference in the survival rate between these two groups (post hoc analysis) suggesting that 108 CFU g−1 may be sufficient to afford protection to the fish against A. hydrophila infection. These results were comparable to findings by Chang and Liu [48] where they used E. faecium at a dose of 5 × 108 cells g−1 feed as feed additive to control infection caused by Edwardsiella tarda in European eels (Anguilla anguilla L.). Survival rate (73%) was significantly higher in fish fed with E. faecium than in the control group and the untreated control experienced mortality of 75–100% within 2 weeks when challenged with E. tarda. However, only a few E. faecium QQ12 cells remained in the goldfish intestine at the end of 7 days after probiotic feeding was discontinued. One day after the discontinuation of probiotic feed, no significant reduction in E. faecium QQ12 cell number was observed in goldfish intestine. Therefore, E. faecium QQ12 is recommended for every alternate day use to prevent A. hydrophila infection during goldfish culture.
The role of E. faecium as a quorum quenching bacteria has not received much attention. This is the first study to isolate and characterize AHL degrading E. faecium from gastrointestinal tract of tilapia and to provide evidence that it is quorum quenching as well as probiotic strain. The results of the present study clearly suggest that the higher survival rate of goldfish after challenging with A. hydrophila is due to the combined effect of quorum quenching ability of E. faecium QQ12 together with its probiotic activity. Production of AHL degrading enzyme might have inhibited the pathogenicity of A. hydrophila, while, the probiotic potential of the QQ12 might have simultaneously helped to out-compete A. hydrophila for nutrients and space and exclude the pathogenic bacteria through antagonistic activity.
Acknowledgments
The authors thank the authorities of the Kerala University of Fisheries and Ocean Studies (KUFOS), Kochi, Kerala for the facilities extended for carrying out this work at the Centre for Aquatic Animal Health Management, KUFOS. We thank Dr. Mathew Sebastian, KUFOS for the statistical analysis of results.
Compliance with ethical standards
Conflict of interest
The authors declare that there is no conflict of interest.
Statement of ethical approval
The present research involving fish was carried out in strict accordance with the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA). The protocol was reviewed and approved by the institutional animal ethics committee of Kerala University of Fisheries and Ocean Studies, India. All dissection was performed under tricaine methanesulfonate (Sigma) (100 mg L−1) anesthesia and all efforts were made to minimize suffering.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Angka SL, Lam TJ, Sin YM. Some virulence characteristics of Aeromonas hydrophila in walking catfish (Clarias gariepinus) Aquaculture. 1995;130:103–112. doi: 10.1016/0044-8486(94)00216-B. [DOI] [Google Scholar]
- 2.Rahman MH, Suzuki S, Kawai K. The effect of temperature on Aeromonas hydrophila infection in goldfish, Carassius auratus. J Appl Ichthyol. 2001;17:282–285. doi: 10.1046/j.1439-0426.2001.00291.x. [DOI] [Google Scholar]
- 3.Truchado P, Lopez GF, Gil MI, Tomas BFA, Allende A. Quorum sensing inhibitory and antimicrobial activities of honeys and the relationship with individual phenolics. Food Chem. 2009;115:1337–1344. doi: 10.1016/j.foodchem.2009.01.065. [DOI] [Google Scholar]
- 4.Mangwani N, Dash H, Chauhan A, Das S. Bacterial quorum sensing: functional features and potential applications in biotechnology. J Mol Microbiol Biotechnol. 2012;22:202–208. doi: 10.1159/000341847. [DOI] [PubMed] [Google Scholar]
- 5.Abudoleh S, Mahasneh A. Anti-quorum sensing activity of substances isolated from wild berry associated bacteria. Avicenna J Med bBiotechnol. 2017;9:32–38. [PMC free article] [PubMed] [Google Scholar]
- 6.Dong YH, Xu JL, Li XZ, Zhang LH. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum sensing signal and attenuates the virulence of Erwinia carotovora. PNAS USA. 2000;97:3526–3531. doi: 10.1073/pnas.060023897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Defoirdt T, Boon N, Sorgeloos P, Verstraete W, Bossier P. Alternatives to antibiotics to control bacterial infections: luminescent vibriosis in aquaculture as an example. Trends Biotechnol. 2007;25:472–479. doi: 10.1016/j.tibtech.2007.08.001. [DOI] [PubMed] [Google Scholar]
- 8.Cam DTV, Nhan DT, Ceuppens S, Hao NV, Dierckens K, Wille M, Sorgeloos P, Bossier P. Effect of N–acyl homoserine lactone–degrading enrichment cultures on Macrobrachium rosenbergii larviculture. Aquaculture. 2009;294:5–13. doi: 10.1016/j.aquaculture.2009.05.015. [DOI] [Google Scholar]
- 9.Dong YH, Wang LH, Xu JL, Zhang HB, Zhang XF, Zhang LH. Quenching quorum sensing dependent bacterial infection by N-acyl homoserine lactonase. Nature. 2001;411:813–817. doi: 10.1038/35081101. [DOI] [PubMed] [Google Scholar]
- 10.Zhang HB, Wang LH, Zhang LH. Genetic control of quorum-sensing signal turnover in Agrobacterium tumefaciens. Proc Natl Acad Sci U S A. 2002;99:4638–4643. doi: 10.1073/pnas.022056699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lin YH, Xu JL, Hu J, Wang SL, Ong JR. Acyl-homoserine lactone acylase from ralstonia strain XJ12B represent a novel and potent class of quorum quenching enzymes. Mol Microbiol. 2003;47:849–860. doi: 10.1046/j.1365-2958.2003.03351.x. [DOI] [PubMed] [Google Scholar]
- 12.Akhter N, Wu B, Memon AM, Mohsin M. Probiotics and prebiotics associated with aquaculture: a review. Fish Shellfish Immun. 2015;45:733–741. doi: 10.1016/j.fsi.2015.05.038. [DOI] [PubMed] [Google Scholar]
- 13.Irianto A, Austin B. Probiotics in aquaculture. J FishDis. 2002;25:633–642. [Google Scholar]
- 14.Nhan DT, Cam DTV, Wille M, Defoirdt T, Bossier P, Sorgeloos P. Quorum quenching bacteria protect Macrobrachium rosenbergii larvae from Vibrio harveyi infection. J Appl Microbiol. 2010;109:1007–1016. doi: 10.1111/j.1365-2672.2010.04728.x. [DOI] [PubMed] [Google Scholar]
- 15.Chu W, Zhou S, Zhu W, Zhuang X. Quorum quenching bacteria Bacillus sp. QSI-1 protect zebrafish (Danio rerio) from Aeromonas hydrophila infection. Sci Rep. 2014;4:1–6. doi: 10.1038/srep05446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vinoj G, Vaseeharan B, Thomas S, Spiers AJ, Shanthi S. Quorum quenching activity of the AHL-lactonase from Bacillus licheniformis DAHB1 inhibits vibrio biofilm formation in vitro and reduces shrimp intestinal colonization and mortality. Mar Biotechnol. 2014;16:707–715. doi: 10.1007/s10126-014-9585-9. [DOI] [PubMed] [Google Scholar]
- 17.Tinh NTN, Gunasekara RAYSA, Boon N, Dierckens K, Sorgeloos P, Bossier P. N-acyl homoserine lactone-degrading microbial enrichment cultures isolated from Penaeus vannamei shrimp gut and their probiotic properties in Brachionus plicatilis cultures. FEMS Microbiol Ecol. 2007;62:45–53. doi: 10.1111/j.1574-6941.2007.00378.x. [DOI] [PubMed] [Google Scholar]
- 18.Tinh NTN, Yen VHN, Dierckens K, Sorgeloos P, Bossier P. An acyl homsoerine lactone-degrading microbial community improves the survival of first feeding turbot larvae (Scophthalmus maximus L.) Aquaculture. 2008;285:56–62. doi: 10.1016/j.aquaculture.2008.08.018. [DOI] [Google Scholar]
- 19.Defoirdt T, Loan DT, Delsen BA, Schryver PD, Sorgeloos P, Boon N, Bossier P. N-acylhomoserine lactone-degrading Bacillus strains isolated from aquaculture animals. Aquaculture. 2011;311:258–260. doi: 10.1016/j.aquaculture.2010.11.046. [DOI] [Google Scholar]
- 20.Chu W, Lu F, Zhu W, Kang C. Isolation and characterization of new potential probiotic bacteria based on quorum-sensing system. J Appl Microbiol. 2010;110:202–208. doi: 10.1111/j.1365-2672.2010.04872.x. [DOI] [PubMed] [Google Scholar]
- 21.Boopathi S, Selvakumar N, Sivakumar G. Quorum quenching potentials of probiotic Enterococcus durans Lab38 against methicillin resistant Staphylococcus aureus. Asian J Pharm Clin Res. 2017;10:445–450. doi: 10.22159/ajpcr.2017.v10i4.17039. [DOI] [Google Scholar]
- 22.Wang YB, Tian ZQ, Yao JT, Li WF. Effect of probiotics, Enterococcus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture. 2008;277:203–207. doi: 10.1016/j.aquaculture.2008.03.007. [DOI] [Google Scholar]
- 23.Krummenauer D, Abreu PC, Lara G, Poersch L, Encarnacao P, Wasielesky W., Jr . The effect of probiotic in Litopenaeus vannamei biofloc technology culture system contaminated with Vibrio parahaemolyticus. Mexico: World Aquaculture Conference; 2009. [Google Scholar]
- 24.Sun YZ, Yang HL, Ma RL, Song K, Li JS. Effect of Lactococcus lactis and Enterococcus faecium on growth performance, digestive enzymes and immune response of grouper Epinephelus coioides. Aquac Nutr. 2012;18:281–289. doi: 10.1111/j.1365-2095.2011.00894.x. [DOI] [Google Scholar]
- 25.Erkkila S, Petaja E. Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci. 2000;55:297–300. doi: 10.1016/S0309-1740(99)00156-4. [DOI] [PubMed] [Google Scholar]
- 26.Hyronimus B, Marrec C, Sassi AH, Deschamps A. Acid and bile tolerance of spore-forming lactic acid bacteria. Int J Food Microbiol. 2000;61:193–197. doi: 10.1016/S0168-1605(00)00366-4. [DOI] [PubMed] [Google Scholar]
- 27.Chan KG, Yin WF, Sam CK, Koh CL. A novel medium for the isolation of N-acylhomoserine lactone-degrading bacteria. J Ind Microbiol Biotechnol. 2009;36:247–251. doi: 10.1007/s10295-008-0491-x. [DOI] [PubMed] [Google Scholar]
- 28.Ludwig W, Schleifer KH, Whitman WB (2009) Bergey’s manual of systematic bacteriology, 2nd edn, Vol. 3, 594–624
- 29.Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals; approved standards. NCCLS document M100-S23, CLSI, Wayne, 2013
- 30.Chan KG, Tiew SZ, Ng CC. Rapid isolation method of soil bacilli and screening of their quorum quenching activity. AsPac J Mol Biol Biotechnol. 2007;15:153–156. [Google Scholar]
- 31.Del Re B, Sgorbati B, Miglioli M, Palenzona D. Adhesion, auto aggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett Appl Microbiol. 2000;31:438–442. doi: 10.1046/j.1365-2672.2000.00845.x. [DOI] [PubMed] [Google Scholar]
- 32.Handley PS, Harty DWS, Wyatt JE, Brown CR, Doran JP, Gibbs ACC. A comparison of the adhesion, co aggregation and cell-surface hydrophobicity properties of fibrillar and fimbriate strains of Streptococcus salivarius. J Gen Microbiol. 1987;133:3207–3217. doi: 10.1099/00221287-133-11-3207. [DOI] [PubMed] [Google Scholar]
- 33.Molina L, Constantinescu F, Michel L, Reimmann C, Duly B, De Fago G. Degradation of pathogen quorum-sensing molecules by soil bacteria: a preventive and curative biological control mechanism. FEMS Microbiol Ecol. 2003;45:71–81. doi: 10.1016/S0168-6496(03)00125-9. [DOI] [PubMed] [Google Scholar]
- 34.Martinez MS, Uyttendaele M, Rajkovic A, Nadal P, Debevere J. Degradation of N-acyl-L-homoserine lactones by Bacillus cereus in culture media and pork extract. J Appl Environ Microbiol. 2007;73:2329–2332. doi: 10.1128/AEM.01993-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Swift S, Karlyshev AV, Fish L, Durant EL, Winson MK, Chhabra SR, Williams P, Macintyre S, Stewart GS. Quorum sensing in Aeromonas hydrophila and Aeromonas salmonicida: identification of the LuxRI homologs AhyRI and AsaRI and their cognate N-acylhomoserine lactone signal molecules. J Bacteriol. 1997;179:5271–5281. doi: 10.1128/jb.179.17.5271-5281.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Galaz AG, Morales RP, Cinco MD, Felix EA. Resistance of Enterococcus strains isolated from pigs to gastrointestinal tract and antagonistic effect against Escherichia coli K88. Rev Latinoam Microbiol. 2004;46:5–11. [PubMed] [Google Scholar]
- 37.Marciakova M, Simonova M, Laukova A. Probiotic properties of Enterococcus faecium EF9296 strain isolated from silage. Acta Vet Brno. 2004;73:513–519. doi: 10.2754/avb200473040513. [DOI] [Google Scholar]
- 38.Saelim K, Sohsomboon N, Kaewsuwan S, Maneerat S. Probiotic properties of Enterococcus faecium CE5-1 producing a bacteriocin-like substance and its antagonistic effect against antibiotic-resistant enterococci in vitro. Czech J Anim Sci. 2012;57:529–539. doi: 10.17221/6386-CJAS. [DOI] [Google Scholar]
- 39.Salma H, Abu Hafsa A, Mendonca B, Brehm-Stecher AA, Hassan SA, Ibrahim K. Probiotic potential and antimicrobial activity of Enterococcus faecium isolated from chicken caecal and fecal samples. IJMRHS. 2015;9:378–382. [Google Scholar]
- 40.Welliton GF, Tania CP, Fabricio MD, Frank L, Eduardo L, Cupertino B, Leandro P. Gastrointestinal tract pH measurement in juveniles Pacu Piaractus mesopotamicus (Characiformes: Characidae) Pan-Am J Aquat Sci. 2017;12:254–258. [Google Scholar]
- 41.Suskovic J, Brkic B, Matosic S, Maric V. Lactobacillus acidophilus M92 as potential probiotic strain. Milchwissenschaft. 1997;52:430–435. [Google Scholar]
- 42.Hoier E. In The 25th Annual Convention. Sydney: Australian Institute of Food Science Technology; 1992. Use of probiotic starter cultures in dairy products. [Google Scholar]
- 43.Bao Y, Zhang Y, Zhang Y, Liu Y, Wang S, Dong X, Wang Y, Zhang H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control. 2010;21:695–701. doi: 10.5713/ajas.15.0849. [DOI] [Google Scholar]
- 44.Jelena R, Natasa J. Probiotic properties and safety assessment of lactic acid bacteria isolated from kajmak. Biol Nyssana. 2015;6:81–89. [Google Scholar]
- 45.Garcia CT, Korany AM, Bustos I, de CLP G, Requena T, Pelaez C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res Int. 2014;57:44–50. doi: 10.1016/j.foodres2014.01.010. [DOI] [Google Scholar]
- 46.Blagoeva G, Gotcheva V, Angelov A (2014) Aggregation and co-aggregation abilities of potentially probiotic amylolytic lactic acid bacteria strains. In Proceedings of the ENGIHR Conference 2014 by the ENGIHR project of the European Science Foundation
- 47.Peres CM, Alves M, Hernandez-Mendoza A, Moreira L, Silva S, Bronze MR. Novel isolates of lactobacilli from fermented Portuguese olive as potential probiotics. LWT. 2014;59:234–246. doi: 10.1016/j.lwt.2014.03.003. [DOI] [Google Scholar]
- 48.Chang CI, Liu WY. An evaluation of two probiotic bacterial strains, Enterococcus faecium SF68 and Bacillus toyoi, for reducing edwardsiellosis in cultured European eel, Anguilla anguilla L. J Fish Dis. 2002;25:311–315. doi: 10.1046/j.1365-2761.2002.00365.x. [DOI] [Google Scholar]