

Abstract
Although stem cell therapy has tremendous therapeutic potential, clinical translation of stem cell therapy has yet to be fully realized. Recently, patient comorbidities and lifestyle choices have emerged to be important factors in the efficacy of stem cell therapy. Tobacco usage is an important risk factor for numerous diseases, and nicotine exposure specifically has become increasing more prevalent with the rising use of electronic cigarettes. This review describes the effects of nicotine exposure on the function of various stem cells. We place emphasis on the differential effects of nicotine exposure in vitro and as well as in preclinical models. Further research on the effects of nicotine on stem cells will deepen our understanding of how lifestyle choices can impact the outcome of stem cell therapies.
Keywords: : electronic cigarettes, nicotine, stem cell therapy, tobacco
As we enter a new decade in 2020, there are over 5000 clinical trials using stem cells to treat a variety of diseases. Among these, over 1000 clinical trials involve stem cell therapy to treat cardiovascular diseases such as peripheral arterial disease, atherosclerosis and congestive heart failure. Despite the popularity of stem cell-based clinical trials, the clinical efficacy of stem cell therapeutics has had mixed results. Some of the most well-described hurdles to clinical translation are related to the production and purification of stem cells and subsequent implantation and engraftment at the location of interest [1]. However, some aspects of translation that are often overlooked are the comorbidities and lifestyle choices of patients receiving the treatment. Among these, tobacco usage is the most prevalent contributor to preventable deaths worldwide and remains an important lifestyle consideration in the clinic [2]. In recent years, the increasing popularity of electronic cigarettes (e-cigarettes)/vaping has provided new ways for consumers to be exposed to tobacco substances. One constant among the many chemicals produced in tobacco-related products is nicotine, the addictive component. The effect of nicotine on stem cells in the context of their therapeutic efficacy is largely unknown, due to the limited amount research in this area. Here, we provide an overview of the current knowledge of nicotine and its effect on adult and pluripotent stem cells, with a focus on in vitro systems and preclinical models.
Clinical & biological impact of nicotine
Nicotine is the addictive component of tobacco-related products such as cigarettes, cigars and chewing tobacco. Nicotine is also the active ingredient in nicotine replacement therapies such as chewing gum or transdermal patches that are intended to aide in tobacco use cessation. These nicotine replacement therapy products are considered safe and have shown efficacy in tobacco use cessation [3]. More recently, with the rising popularity of e-cigarette use, e-cigarettes have become an additional route of nicotine exposure. Initially, e-cigarettes were touted as a less harmful alternative than tobacco and an effective nicotine replacement therapy for tobacco cessation [4], but some clinical data suggest that e-cigarette use reduces the chance of quitting tobacco use [5]. Furthermore, e-cigarette usage among youth has been concerning. The prevalence of e-cigarette usage in high school students in the USA progressively increased in recent years, from 1–2% in 2010–2011 to 10–20% in 2013–2017 and then to 27% in 2019 [6–8]. Data suggest that adolescence e-cigarette use leads to increased tobacco use later in life [9].
Although nicotine is generally perceived as safe, nicotine and its metabolites have been linked to adverse systemic effects. Such examples can be seen in the cardiovascular system, where nicotine’s actions can lead to increase incidence of atherosclerosis and angiogenesis [10,11]. Nicotine exposure reduces blood flow to the coronary vessels that leads to ischemia [12]. Over an extended period, the repetition of acute ischemia causes myocardial dysfunction. In addition, nicotine also has direct effects on vascular smooth muscle cells and endothelial cells by increasing the secretion of basic FGF. This results in intimal thickening and atherosclerotic plaque progression [13]. In other areas such as cancer, nicotine exposure has been shown to increase tumor angiogenesis, leading to accelerated tumor growth [11]. There is also evidence that nicotine regulates proliferation and apoptotic pathways to promote tumorigenesis [14].
The biological effects of nicotine are mediated by the nicotinic acetylcholine receptors (nAChR), which are composed of five subunits that form a central pore (Figure 1) [15]. There are 18 nAChR subunits, designated as α1–10, β1–5, γ, δ and ϵ [15]. The receptor is either heteromeric or homomeric, and is most commonly composed of (α4)2(β2)3 and (α7)5 respectively [15]. In the context of stem cells, expression of nAChR subunits have been confirmed [16]. Once the receptor is activated by acetylcholine or nicotine, the central pore opens to allow for sodium, potassium and calcium cations to enter the cell. The influx of these positive ions causes depolarization, which is then propagated by voltage gated ion channels in neuronal cells within the nervous system and neuromuscular junctions to control contraction. These neurogenic effects have been well documented due to the addictive properties of nicotine in the interference of dopamine pathways [17]. The relevant effect of nAChR activation in stem cells is the influx of calcium ions, triggering a number of intracellular signaling transduction pathways that affect cellular behaviors, including survival, proliferation and apoptosis (Figure 1) [18].
Figure 1. . Overview of nicotinic acetylcholine receptor signaling pathway.
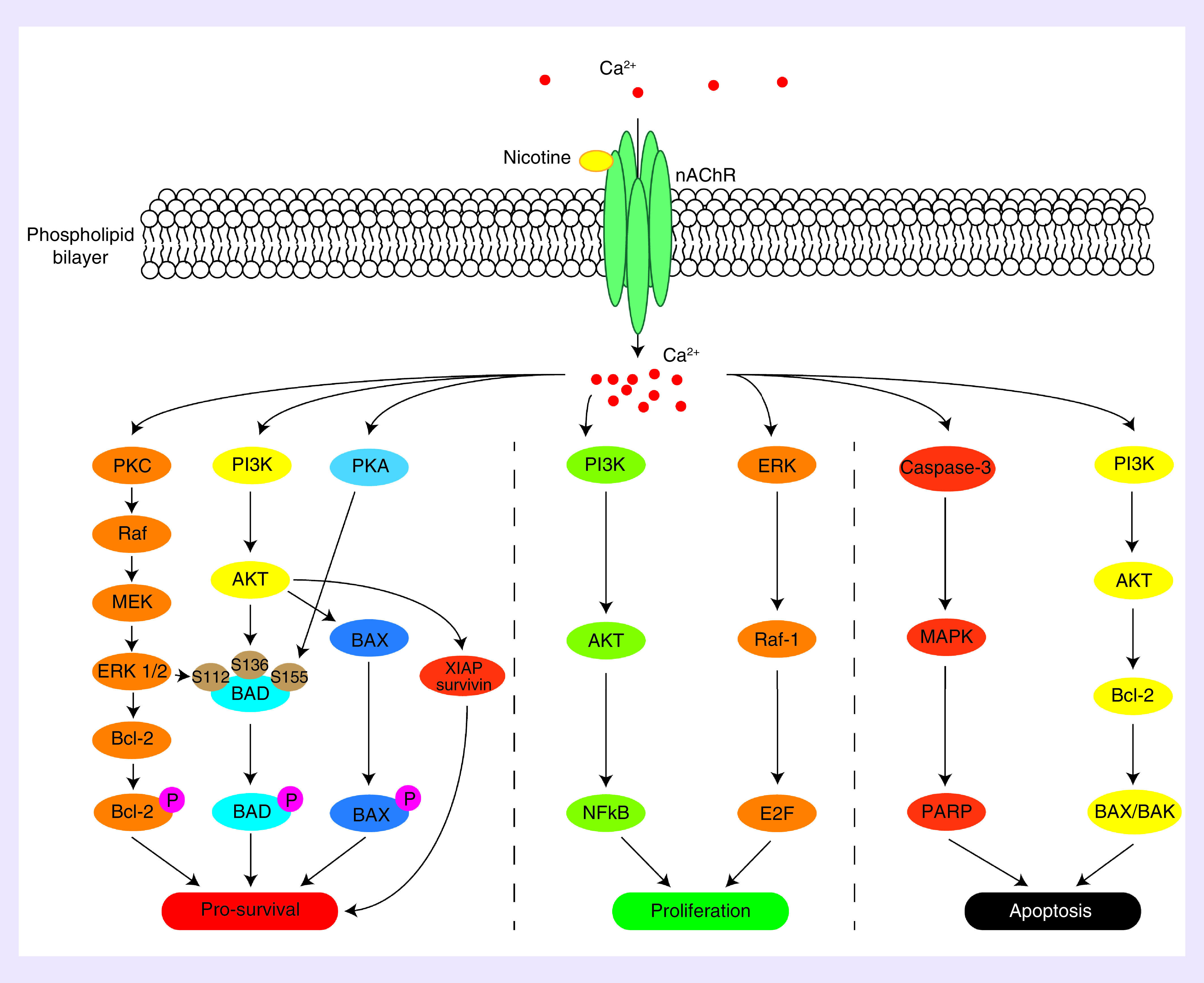
Reproduced with permission from [18].
Effects of nicotine on stem cells
Whereas the effects of nicotine in disease have been well examined, less is known about the effects of nicotine on therapeutic stem cells. The following section describes the current knowledge of nicotine exposure’s effect on various stem cell types (Figure 2), including circulating progenitor and stem cells, mesenchymal stem cells (MSCs), embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs).
Figure 2. . Schematic depiction of stem cell types for studying or modeling nicotine exposure.
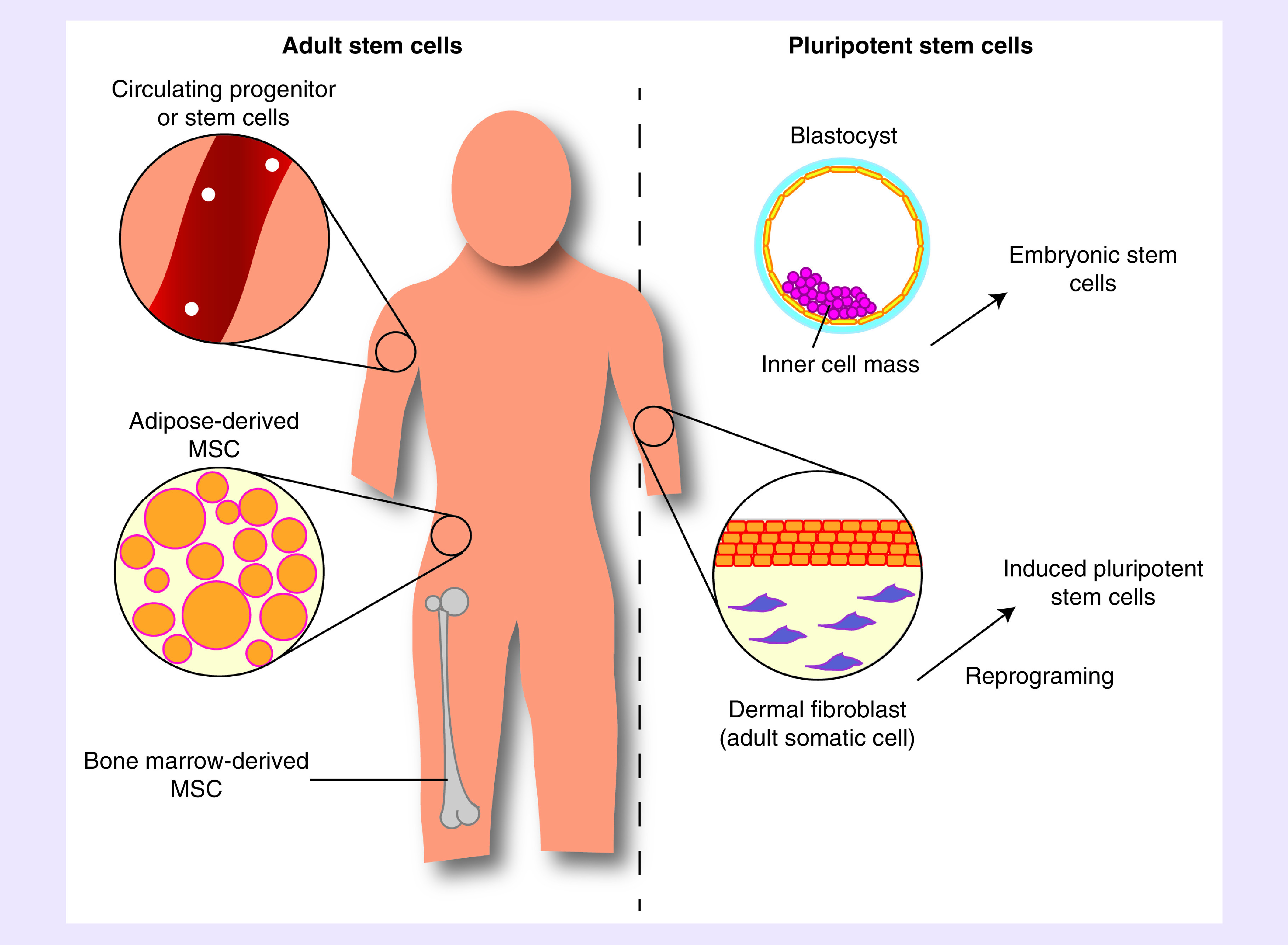
MSC: Mesenchymal stem cells.
Circulating progenitor & stem cells
Endogenous circulating progenitor and stem cells are cells that originate in the bone marrow and circulate throughout the body, and have been shown to participate in numerous regenerative processes in the body. In the context of cardiovascular regenerative medicine, endothelial progenitor cells (EPCs) have gained considerable interest due to their ability to form mature endothelial cells and contribute to angiogenesis in response to ischemic injury [19]. With the increased prevalence of ischemic diseases such as coronary heart disease and peripheral artery diseases, EPCs remain an important option in stem cell therapy.
EPCs have generally been characterized by expression of cell surface markers such as CD31, CD144 and VEGFR. However, due to overlaps in cell surface expression with endothelial cells, there is currently no unique EPC marker [20]. Recently, there has been a shift toward a functional definition of EPCs through the assessment of self-renewal and potency. EPCs are thereby defined by their ability to form colonies in vitro and induce angiogenic capacity in vivo.
In vitro experiments that assessed the effects of nicotine exposure on late EPCs from human umbilical cord blood showed that nicotine increases proliferation, migration and tubule formation [21]. These effects were evident at a concentration of 10 nM for 36 h, and the positive effects were abrogated when cells were treated with mecamylamine, a nAChR inhibitor. These observations were also seen in bone marrow mononuclear cells-derived EPCs with a shorter nicotine exposure duration of 32 h [22]. In contrast, a study using EPCs from peripheral blood showed that nicotine did not change proliferation and migration capabilities after 48 h of nicotine exposure (20 nM–20 μM) in vitro, although an anti-apoptotic effect was observed [23]. Interestingly, one study showed that even only a 30-min exposure to tobacco secondhand smoke caused an increase in EPC numbers, although the migratory response of EPCs was also impaired [24].
When examining the effects of nicotine on the functionality of EPCs in vivo, investigators observed that systemic nicotine exposure increased the recruitment and angiogenic actions of EPCs in a mouse model of hindlimb ischemia (HLI) [21]. This study also employed a mouse parabiosis model, which links the circulation of a wild-type mouse with that of a tie2-LacZ transgenic mouse, to show improved mobilization and recruitment of EPCs. Nicotine-treated mice exhibited greater EPC numbers and showed increased EPC migration ex vivo. Similarly, Sugimoto et al. showed increases in the efficacy of EPCs delivered venously in a mouse model of HLI [25]. In this study, mice were exposed to nicotine via their drinking water one week before the HLI and throughout 3-week experiment.
While the aforementioned studies show convincing evidence for improvement in the number of EPCs in vivo and improved functions with short-term exposure to nicotine, other studies investigating the long-term effects of nicotine exposure showed the opposite. In one such study, the mice were exposed to nicotine systemically in their drinking water for up to 6 months [26], which was notably longer in duration compared with the 3-week exposure in Sugimoto et al. After 1 month of exposure to nicotine, EPC number and activity were consistent with earlier short-term exposure. However, after 3 and 6 months of nicotine exposure, the EPC number significantly decreased, and cell proliferation and migration functions were impaired compared with control EPCs without nicotine treatment. The authors also investigated a potential biochemical mechanism governing this switch in cell behavior. They found that nAChR subunit expression was significantly decreased after 6 months of nicotine exposure, thereby limiting the activation of cholinergic angiogenic pathways. Telomerase expression and activity were also decreased after 6 months of nicotine exposure. These studies suggest that chronic nicotine exposure adversely affects the number and function of EPCs, which may lead to a decrease in therapeutic capacity (Table 1).
Table 1. . Effect of nicotine exposure on multipotent stem cells.
| Cell type | In vivo/in vitro | Nicotine dosage | Exposure duration | Outcome | Ref. |
|---|---|---|---|---|---|
| Endogenous mice EPCs | In vivo, mouse parabiosis model with HLI model | 100 μg/ml in drinking water or intramuscular injection (0.03 μg/kg) | 30 days | • Systemic nicotine exposure induced greater angiogenesis; local delivery also increased angiogenesis but not to the same extent • Systemic nicotine exposure increased number of EPCs in circulation and chemotaxis migration |
[21] |
| Human EPCs from healthy nonsmokers | In vitro, media | 2 μM | 48 h | • No effect on proliferation and migration • Improved survival |
[25] |
| Mouse EPCs | Ex vivo | 100 ng/ml in drinking water | 3 weeks | • Increased EPC activity | [25] |
| Mouse EPCs | In vivo (mice HLI model) | 100 ng/ml in drinking water | 5 weeks | • Increased perfusion in HLI | [25] |
| Human EPCs from umbilical cord blood | In vitro | 10 nM | 36 h | • Increased proliferation | [23] |
| Mouse EPCs | Ex vivo | 100 ng/ml in drinking water | 1, 3 and 6 months | • Increased number of EPCs after 1 month, but decreased over subsequent time • Same trends with migration and proliferation |
[26] |
| Human MSCs | In vitro | 0.1, 1 and 10 μM | 24 h | • Increase proliferation | [27] |
| Human MSCs | In vivo | 0.01 μM, iv. | 24 h | • Migration to bone marrow and spleen was decreased, lung was not affected. | [27] |
| Rat BM MSCs | In vitro | 25, 50 and 100 μM | 4 weeks | • No effect on cell viability • Decreased efficacy of differentiation |
[28] |
| Human MSCs | In vitro | 0.1, 0.5 and 1 μM | 48 h | • Cytoplasm and nucleus became stiffer in a concentration dependent manner | [29] |
| Human MSCs and periodontal ligament derived stem cells | In vitro | 1 μM | Up to 7 days | • Proliferation and migration impaired • Osteogenic differentiation inhibited |
[30] |
| Human umbilical cord derived MSCs | In vitro | 0.1, 0.5, 1 and 1.5 mg/ml | 24 h | • Apoptosis significantly increased at high concentrations of nicotine • Proteomic analysis showed differences in cell structure and motility |
[31] |
BM: Bone marrow; EPC: Endothelial progenitor cell; HLI: Hindlimb ischemia; iv.: Intravenously; MSC: Mesenchymal stem cell.
Mesenchymal stem cells
MSCs are multipotent adult stem cells capable of differentiating into mesodermal, ectodermal and endodermal lineages. MSCs were first isolated from bone marrow, but they have since been isolated from various other tissues such as adipose tissue, dental tissue, skin, umbilical cord, placental membrane and synovial fluid. In general, MSCs are characterized by the expression of cell surface markers (such as CD73, CD90 and CD105) and lack of CD14, CD34, CD45 and human leucocyte antigen-DR. MSCs are the most investigated stem cell type for clinical stem cell therapeutics, with a rapid increase in clinical trials from the mid-2000s [32]. The advantages of MSCs include their ability to be easily isolated and purified from bone marrow and adipose tissue. Additionally, MSCs obviate potential ethical concerns associated with ESCs and have not been shown to give rise to teratomas. Given the many ongoing clinical trials with MSCs, it is important to consider the effects of lifestyle choice such as smoking and nicotine exposure on their therapeutic efficacy.
The in vitro effects of nicotine on MSCs have been widely studied, but the dosage and duration of nicotine have varied among the studies. The common consensus is that high doses of nicotine at the micromolar level cause apoptosis in MSCs and impair proliferation. Schraufstatter et al. found that only a short 24-h exposure of nicotine (1 μM) significantly increased apoptosis in human MSCs, as demonstrated by propidium iodide staining [27]. The same study also showed that chemotactic migration was inhibited, and that this effect could be reversed by pharmacological inhibition of the nAChR subunit α7. These results were replicated in a study comparing human MSCs with periodontal ligament-derived stem cells. Both cell types were exposed to nicotine (1 μM) over 5 and 7 days which then resulted in impaired proliferation and migration as early as 1 day after nicotine treatment in MSCs [30]. Another study showed increased apoptosis with 6.2 mM nicotine exposure and changes in cell morphology compared with nontreated cells, namely increased vacuole formation in the cytoplasm and pyknotic nuclei [31]. Proteomic analysis showed differential expression of cell structure and movement markers, which may help explain the observed impairments in migratory functions after nicotine exposure [31]. This concept was also explored in an investigation of nicotine’s effects on the mechanical properties of MSCs. It was found that after a 48-h exposure regime, cells were significantly stiffer than nontreated controls [29], which is an indicator for cell motility. Another important consideration is the efficacy of differentiation. Findings using nicotine alone were similar to data generated with cigarette smoke. Cigarette smoke-infused media decreased the proliferation and migration of human adipose derived MSCs [33]. The authors also observed decreases in the secretion of inflammatory makers IL6 and IL8, which are important for chemotaxis in wound-healing processes. In vitro chondrogenic differentiation was significantly impaired when MSCs were exposed to nicotine in a concentration dependent manner [28,30].
In vivo studies of MSCs in the context of nicotine exposure have shown that homing of intravenously delivered MSCs to the bone marrow and spleen was impaired in mice, whereas the lung was not affected [27]. This data corroborates other in vitro results showing decreased migratory function of MSCs after nicotine exposure. In a more recent study, Zhao et al. isolated alveolar bone marrow MSCs from healthy nonsmokers and smokers to assess their efficacy in osteogenic function [34]. Using a subcutaneous murine model, polylactic acid scaffolds seeded with MSCs were assessed for bone matrix after 8 weeks. MSCs from smokers produced significantly less bone matrix compared with nonsmokers. These scaffolds were transplanted autologously in the patients, and nonsmokers exhibited significantly higher implant stability after 4 weeks. In a direct comparison of adipose derived MSCs from smokers or nonsmokers, Barwinska et al. utilized a mouse HLI model to show that MSCs derived from smokers had significantly dampened angiogenesis, compared with nonsmokers [35]. In vitro experiments from the same study suggest a possible molecular mechanism involving activin A, whereby activin A diminishes secretion of HGF and stromal-derived factor-1 (Table 1).
Embryonic stem cells
ESCs are a kind of pluripotent stem cells that are capable of unlimited expansion. They are derived from the inner cell mass of an embryo and express cell surface markers such as stage-specific embryonic antigen-4, Tra-1-60 and Tra-1-81. Pluripotent markers such as Oct3/4 are also expressed in ESCs, as well as high levels of telomerase. Human and mouse ESCs have been shown to expand in feeder cells and feeder-free systems, while maintaining a high level of consistency in karyotype and phenotype with extended expansion. Due to the pluripotency of ESCs, they have been applied in many disease settings and preclinical studies such as spinal injury, cardiovascular disease and degenerative neurological diseases [36,37]. However, as a result of the ethical concerns over the destruction of viable embryos, human ESCs have largely fallen out of favor for use in regenerative medicine [38].
The use of ESCs for modeling embryonic development in the context of tobacco and nicotine exposure has become an important tool in developmental biology. Studies using mouse and human ESCs have shown the negative effects of cigarette smoke exposure. Specific studies comparing regular cigarettes with harm reduction cigarettes showed that 24 h exposure of main-stream smoke or side-stream smoke (secondhand smoke) all produced higher levels of cytotoxicity and impaired adhesion in both mouse and human ESCs, relative to nontreated controls [39,40], with side-stream smoke being more cytotoxic to ESCs. Experiments using only nicotine showed an interesting trend in which low concentrations (0.01 and 0.1 μM) of nicotine promoted proliferation of mouse ESCs, whereas high concentrations (1 and 10 μM) impaired proliferation after 48 h of exposure [41]. The same study investigated the pathways affected by nicotine exposure and found that Wnt signaling was dysregulated. These findings were supported by in vitro findings, in which cigarette smoke and e-cigarette smoke showed impaired cardiac differentiation of human ESCs [42]. The results suggest that maturation of cardiomyocytes was impaired because a lower expression of contractile proteins and junction proteins was observed. Mechanistic investigation also indicated Wnt signaling pathway dysregulation in this study. A further study investigated the effects of 48-h cigarette smoke condensate exposure on mouse ESCs from different genetic backgrounds and maternal ages [43]. The results showed that cigarette smoke condensate exposure had different effects depending on the strain and maternal age. For example, apoptotic indicator Bax/Bcl2 ratio was significantly increased with low concentrations of cigarette smoke condensate in C57BL/6 mice at 2 months maternal age. However, it was decreased in the mice of the same strain at 6 months maternal age and also decreased in a Naval Medical Research Institute outbred strain at 2 months maternal age. Similar trends were seen in expression of pluripotent markers. A study investigating human embryoid body differentiation under nicotine (10 μM) exposure for 21 days showed that nicotine has different effects on specific cell type lineages, with muscle cell lineages exhibiting the greatest differences in gene expression [44]. For example, in ESC-derived cardiomyocytes, nicotine exposure during differentiation causes dysregulation of intracellular calcium handling genes, resulting in arrhythmia (Table 2).
Table 2. . Effects of nicotine exposure on pluripotent stem cells.
| Cell type | In vivo/in vitro | Nicotine dosage | Exposure duration | Outcome | Ref. |
|---|---|---|---|---|---|
| Embryonic stem cell derived endothelial cells | In vivo (mice LAD model) | 10 nM in vitro treatment 100 ng/ml in drinking water |
16 h in vitro treatment 3 weeks in vivo |
• Tubule formation negatively affected under hypoxic conditions • Improved survival of injected cells and increased number of blood vessels |
[45] |
| Mouse embryonic stem cells | In vitro | 0.01–10 μM | 48 h | • Decreased proliferation at concentrations of 1 and 10 μM • Wnt signaling dysregulated |
[41] |
| Human embryoid bodies | In vitro | 10 μM | 21 days | • Nicotine has different gene expression effects in different cell types, with muscle cells exhibiting the greatest differences compared with control | [44] |
| Mouse iPSCs | In vitro | 300 nM | 4 days | • Increased proliferation • Neural differentiation not affected |
[46] |
| iPSC derived dopaminergic neurons | In vitro | 0.1, 1 and 10 μM | 72 h | • Increased nAChR α6 subunit activity • Induced structural plasticity in neurons |
[47] |
| iPSC derived endothelial cells | In vitro | Up to 18 mg/ml e-liquids diluted in media up to 1% | 48 h | • Cell viability, ROS generation and apoptosis all negatively affected | [48] |
iPSC: Induced pluripotent stem cell; LAD: Left anterior descending artery; nAChR: Nicotinic acetylcholine receptor; ROS: Reactive oxygen species.
Human ESC-derived endothelial cells were treated with nicotine (10 nM) for 24 h and then injected intramyocardially following left anterior descending artery ligation in a murine model to assess the effects of nicotine on the regenerative potential of ESC-derived ECs [45]. Results showed that nicotine improved ESC-derived endothelial cell survival at the site of injection and increased in the number of blood vessels. In vitro experiments from the same study also showed that high concentrations of nicotine negatively affect tube formation under hypoxic conditions, and similar trends were seen in apoptosis [45]. Gene expression investigations showed increases in basic FGF and VEGF under hypoxia, which is consistent with paracrine effects of stem cell therapy. Other in vivo assessment in zebrafish comparing the effect of cigarette smoke and e-cigarette smoke on embryonic development showed both smokes increased the incidence of cardiac defects after 72 h of exposure [42]. Interestingly, a control of nicotine alone did not increase cardiac defects in zebrafish embryos. Understanding the mechanisms of nicotine exposure in ESCs may lead to a better understanding of autologous stem cell therapies.
Induced pluripotent stem cells
Although the effects of nicotine have been studied in ESCs, more recently iPSCs have become the pluripotent cell type of choice, because of the ability to generate these cells autologously for regenerative medicine applications. Human iPSCs are generated from adult cells, most commonly from dermal fibroblasts or mononuclear cells from blood. These adult cells are reverted to a pluripotent state through a reprogramming process in which a set of transcriptional factors (c-Myc, Oct3/4, Sox2 and Klf4) are induced [49]. Initial studies utilized viral vectors for induction of transcriptional factors, but the field is progressively developing small molecules that can induce pluripotency [50]. With the substantial therapuetic potential, there is much interest in the effect of nicotine on iPSC proliferation and differentiation.
Previous reports have shown that iPSCs express nAChR subunits α4, α7 and α6, which are required for the activation of acetylcholine pathways [46,47]. Similar to the other cell types described above, in vitro experiments showed that short-term nicotine exposure increases the proliferation of iPSCs. Assessment of differentiation into neural cells showed that efficacy of differentiation was not affected by nicotine exposure. The authors suggested there is potential to exploit the nAChR pathway to aid in the expansion of iPSCs, while maintaining differentiation efficacy [46].
With advent of e-cigarettes and their rise in popularity, the toxicity of e-liquids has also become a concern. Lee et al. utilized iPSCs as a screening model to assess the toxicity of popular e-liquids on the market [48]. Human iPSC proliferation was significantly decreased after exposure to two of the e-liquids at concentrations of 0.1% and greater. The different levels of nicotine (0, ∼37 and ∼111 mM) within the e-liquids were assessed in parallel. The data showed that the differences in nicotine concentration caused minimal differences in proliferation, migration and tubule formation, but found that higher nicotine concentrations tended to cause higher reactive oxygen species generation and apoptosis. In the same study, iPSC-derived endothelial cells were exposed to serum from nonsmokers, cigarettes smokers and e-cigarette users. E-cigarette usage showed similar effects on endothelial dysfunction and reactive oxygen species generation to cigarette usage. This study highlights the importance of nicotine across various forms of exposure (Table 2).
A comparison of iPSC-derived MSCs and bone marrow-derived MSCs in the treatment of cigarette smoke induced cardiac injury showed that iPSC-derived MSCs performed better than bone marrow-derived MSCs in cardiac functional outcomes. They exhibited increased left ventricular ejection fraction and a higher percentage fractional shortening [51]. The authors also showed significant improvements in oxidative stress, lipid dysfunction, inflammatory cytokine secretion and fibrotic response. While the mechanisms underlying these effects are unknown, the results suggest that iPSC-derived MSCs were more resistant to cigarette smoke exposure and the associated physiological environment.
Conclusion & future perspective
As the field of stem cell therapy has progressed over the last decade, the knowledge gap in patient comorbidities and lifestyles have become increasingly apparent in clinical translation. In this review, we focused on one component of tobacco-related product, namely nicotine. Nicotine exposure showed significant impacts on stem cells and their potential efficacy when delivered in vivo. As the field progresses, there remain many unanswered questions. First, since many reported studies involving nicotine were conducted in vitro, an important question lies in how these findings transfer to in vivo disease settings. Furthermore, since the majority of these studies only assessed nicotine’s effect in acute settings, more studies are necessary to assess the effects of chronic (>1 month) nicotine exposure in disease settings (Tables 1 & 2). Side-by-side comparisons of various cell types using similar nicotine exposure conditions may enable a better understanding of cell-specific effects of nicotine. In the clinical context, it is reasonable to assume that chronic nicotine exposure is more relevant, as patients with prolonged nicotine exposure are likely to be patients who would benefit from stem cell therapeutics. This also leads to concerns of whether autologous stem cell therapy would be as effective when the cells are derived from tobacco users, given that current data suggests negative effects of nicotine on stem cell proliferation and function. It is also unknown whether comorbidities combined with nicotine exposure could have synergistic effects on the regenerative compacity of stem cells. Further investigations into this matter could promote the development of allogenic stem cell banks. Additionally, the differential effects of nicotine exposure caused by direct tobacco use, compared with secondhand smoke, requires further investigation. Despite these concerns, recent technological advances in organ-on-a-chip and high-throughput screening platforms may expedite experimental studies and help yield a greater understanding of nicotine’s effects on cardiovascular lineages. Such in vitro systems allow organoids to be exposed to nicotine under disease-mimicking conditions and provide quantitative analysis via high-throughput transcriptomic, proteomic and metabolomic screens. The answers to these questions may have important implications in the design of stem cell therapeutics for the treatment of cardiovascular diseases. More broadly, a better understanding of how nicotine exposure influences physiology will be important for evaluating the safety of nicotine replacement therapies that aid cigarette smoking cessation.
Executive summary.
An often overlooked barrier to the translation of stem cell therapies is lifestyle choices, especially tobacco usage.
Nicotine, the addictive component of tobacco, is increasingly common in new delivery routes such as electronic cigarettes, which are increasing in use among the youth.
Adult stem cells
Effects on endothelial progenitor cells varied with the duration of nicotine exposure: acute exposure increases proliferation and migration of endothelial progenitor cells, whereas chronic exposure has the opposite effect.
Mesenchymal stem cells exhibit increased apoptosis, decreased migration and impaired differentiation compared with other lineages with acute exposure.
Pluripotent stem cells
Embryonic stem cells have been an important tool in assessing the effects of tobacco smoke and nicotine exposure during development, with limited studies on translational applications.
Less established induced pluripotent stem cells are increasingly more prevalent in the field, with in vitro modeling being used to assess harmful effects of e-cigarette liquids.
Conclusion
Future research on the in vivo effects of nicotine exposure are needed to corroborate in vitro data, as well as further investigations into the effects of chronic exposure to nicotine.
Acknowledgments
We acknowledge A Muller from the Stanford Cardiovascular Institute for language editing assistance.
Footnotes
Financial & competing interests disclosure
This work was supported in part by grants to NFH from the US NIH (R01 HL127113 and R01 HL142718), and the US Department of Veterans Affairs (1I01BX002310 and 1I01BX004259). AHPC was supported by a postdoctoral fellowship from the Tobacco Related Disease Research Program (T30FT0860). This publication was made possible by a grant from the US FDA, which supports the UCSF-Stanford Center of Excellence in Regulatory Sciences and Innovation (U01FD005978). The contents of this article are solely the responsibility of the authors and do not necessarily represent the official views of the HHS or FDA. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.
References
Papers of special note have been highlighted as: • of interest
- 1.Huang NF, Serpooshan V, Morris VB. et al. Big bottlenecks in cardiovascular tissue engineering. Commun. Biol. 1, 199 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Onor IO, Stirling DL, Williams SR. et al. Clinical effects of cigarette smoking: epidemiologic impact and review of pharmacotherapy options. Int. J. Environ. Res. Public Health 14(10), 1147 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Stead LF, Perera R, Bullen C. et al. Nicotine replacement therapy for smoking cessation. Cochrane Database Syst. Rev. 11, CD000146 (2012). [DOI] [PubMed] [Google Scholar]
- 4.Bullen C, Howe C, Laugesen M. et al. Electronic cigarettes for smoking cessation: a randomised controlled trial. Lancet 382(9905), 1629–1637 (2013). [DOI] [PubMed] [Google Scholar]
- 5.Kalkhoran S, Glantz SA. E-cigarettes and smoking cessation in real-world and clinical settings: a systematic review and meta-analysis. Lancet Respir. Med. 4(2), 116–128 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Singh T, Arrazola RA, Corey CG. et al. Tobacco use among middle and high school students--United States, 2011–2015. MMWR Morb. Mortal. Wkly. Rep. 65(14), 361–367 (2016). [DOI] [PubMed] [Google Scholar]
- 7.Anic GM, Sawdey MD, Jamal A, Trivers KF. Frequency of use among middle and high school student tobacco product users – United States, 2015–2017. MMWR Morb. Mortal. Wkly. Rep. 67(49), 1353–1357 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cullen KA, Gentzke AS, Sawdey MD. et al. e-Cigarette use among youth in the United States, 2019. JAMA 322(21), 2095–2103 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bold KW, Kong G, Camenga DR. et al. Trajectories of e-cigarette and conventional cigarette use among youth. Pediatrics 141(1), e20171832 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Santanam N, Thornhill BA, Lau JK. et al. Nicotinic acetylcholine receptor signaling in atherogenesis. Atherosclerosis 225(2), 264–273 (2012). [DOI] [PubMed] [Google Scholar]
- 11.Heeschen C, Jang JJ, Weis M. et al. Nicotine stimulates angiogenesis and promotes tumor growth and atherosclerosis. Nat. Med. 7(7), 833–839 (2001). [DOI] [PubMed] [Google Scholar]; • Describes the nicotine’s effect on the vascular system.
- 12.Kaijser L, Berglund B. Effect of nicotine on coronary blood-flow in man. Clin. Physiol. 5(6), 541–552 (1985). [DOI] [PubMed] [Google Scholar]
- 13.Lee J, Cooke JP. The role of nicotine in the pathogenesis of atherosclerosis. Atherosclerosis 215(2), 281–283 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mai H, May WS, Gao F, Jin Z, Deng X. A functional role for nicotine in Bcl2 phosphorylation and suppression of apoptosis. J. Biol. Chem. 278(3), 1886–1891 (2003). [DOI] [PubMed] [Google Scholar]
- 15.Albuquerque EX, Pereira EFR, Alkondon M, Rogers SW. Mammalian nicotinic acetylcholine receptors: from structure to function. Physiol. Rev. 89(1), 73–120 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Carballosa CM, Greenberg JM, Cheung HS. Expression and function of nicotinic acetylcholine receptors in stem cells. AIMS Bioeng. 3(3), 245–263 (2016). [Google Scholar]
- 17.Dani JA, Bertrand D. Nicotinic acetylcholine receptors and nicotinic cholinergic mechanisms of the central nervous system. Annu. Rev. Pharmacol. Toxicol. 47, 699–729 (2007). [DOI] [PubMed] [Google Scholar]
- 18.Resende RR, Adhikari A. Cholinergic receptor pathways involved in apoptosis, cell proliferation and neuronal differentiation. Cell Commun. Signal 7, 20 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]; • Reviews nicotinic acetylcholine receptors in stems cells and the associate pathways.
- 19.Takahashi T, Kalka C, Masuda H. et al. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat. Med. 5(4), 434–438 (1999). [DOI] [PubMed] [Google Scholar]
- 20.Fadini GP, Losordo D, Dimmeler S. Critical reevaluation of endothelial progenitor cell phenotypes for therapeutic and diagnostic use. Circ. Res. 110(4), 624–637 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Heeschen C, Chang E, Aicher A, Cooke JP. Endothelial progenitor cells participate in nicotine-mediated angiogenesis. J. Am. Coll. Cardiol. 48(12), 2553–2560 (2006). [DOI] [PubMed] [Google Scholar]
- 22.Wang X, Zhu J, Chen J, Shang Y. Effects of nicotine on the number and activity of circulating endothelial progenitor cells. J. Clin. Pharmacol. 44(8), 881–889 (2004). [DOI] [PubMed] [Google Scholar]
- 23.Yu M, Liu Q, Sun J, Yi K, Wu L, Tan X. Nicotine improves the functional activity of late endothelial progenitor cells via nicotinic acetylcholine receptors. Biochem. Cell Biol. 89(4), 405–410 (2011). [DOI] [PubMed] [Google Scholar]
- 24.Heiss C, Amabile N, Lee AC. et al. Brief secondhand smoke exposure depresses endothelial progenitor cells activity and endothelial function: sustained vascular injury and blunted nitric oxide production. J. Am. Coll. Cardiol. 51(18), 1760–1771 (2008). [DOI] [PubMed] [Google Scholar]
- 25.Sugimoto A, Masuda H, Eguchi M, Iwaguro H, Tanabe T, Asahara T. Nicotine enlivenment of blood flow recovery following endothelial progenitor cell transplantation into ischemic hindlimb. Stem Cells Dev. 16(4), 649–656 (2007). [DOI] [PubMed] [Google Scholar]
- 26.Li W, Du DY, Liu Y, Jiang F, Zhang P, Li YT. Long-term nicotine exposure induces dysfunction of mouse endothelial progenitor cells. Exp. Ther. Med. 13(1), 85–90 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]; • Extends nicotine exposure to 6 months in mice.
- 27.Schraufstatter IU, Discipio RG, Khaldoyanidi SK. Alpha 7 subunit of nAChR regulates migration of human mesenchymal stem cells. J. Stem Cells 4(4), 203–216 (2010). [PMC free article] [PubMed] [Google Scholar]
- 28.Deng Y, Li TQ, Yan YE, Magdalou J, Wang H, Chen LB. Effect of nicotine on chondrogenic differentiation of rat bone marrow mesenchymal stem cells in alginate bead culture. Biomed. Mater. Eng. 22(1–3), 81–87 (2012). [DOI] [PubMed] [Google Scholar]
- 29.Ruiz JP, Pelaez D, Dias J, Ziebarth NM, Cheung HS. The effect of nicotine on the mechanical properties of mesenchymal stem cells. Cell Health Cytoskelet. 4, 29–35 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ng TK, Carballosa CM, Pelaez D. et al. Nicotine alters microRNA expression and hinders human adult stem cell regenerative potential. Stem Cells Dev. 22(5), 781–790 (2013). [DOI] [PubMed] [Google Scholar]
- 31.Zeng HL, Qin YL, Chen HZ. et al. Effects of nicotine on proliferation and survival in human umbilical cord mesenchymal stem cells. J. Biochem. Mol. Toxicol. 28(4), 181–189 (2014). [DOI] [PubMed] [Google Scholar]
- 32.Olsen TR, Ng KS, Lock LT, Ahsan T, Rowley JA. Peak MSC: are we there yet? Front. Med. (Lausanne) 5, 178 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wahl EA, Schenck TL, Machens HG, Egana JT. Acute stimulation of mesenchymal stem cells with cigarette smoke extract affects their migration, differentiation, and paracrine potential. Sci. Rep. 6, 22957 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhao X, Zhu B, Duan Y, Wang X, Li D. The effect of smoking behavior on alveolar bone marrow mesenchymal stem cells of clinical implant patient. BioMed Res. Int. 2018, 7672695 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Barwinska D, Traktuev DO, Merfeld-Clauss S. et al. Cigarette smoking impairs adipose stromal cell vasculogenic activity and abrogates potency to ameliorate ischemia. Stem Cells 36(6), 856–867 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shroff G, Gupta R. Human embryonic stem cells in the treatment of patients with spinal cord injury. Ann. Neurosci. 22(4), 208–216 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tomaskovic-Crook E, Crook JM. Human embryonic stem cell therapies for neurodegenerative diseases. CNS Neurol. Disord. Drug Targets 10(4), 440–448 (2011). [DOI] [PubMed] [Google Scholar]
- 38.Hyun I. The bioethics of stem cell research and therapy. J. Clin. Invest. 120(1), 71–75 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lin S, Tran V, Talbot P. Comparison of toxicity of smoke from traditional and harm-reduction cigarettes using mouse embryonic stem cells as a novel model for preimplantation development. Hum. Reprod. 24(2), 386–397 (2009). [DOI] [PubMed] [Google Scholar]
- 40.Lin S, Fonteno S, Weng JH, Talbot P. Comparison of the toxicity of smoke from conventional and harm reduction cigarettes using human embryonic stem cells. Toxicol. Sci. 118(1), 202–212 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Qu Q, Zhang F, Zhang X, Yin W. Bidirectional regulation of mouse embryonic stem cell proliferation by nicotine is mediated through Wnt signaling pathway. Dose Response 15(4), 1559325817739760 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Palpant NJ, Hofsteen P, Pabon L, Reinecke H, Murry CE. Cardiac development in zebrafish and human embryonic stem cells is inhibited by exposure to tobacco cigarettes and e-cigarettes. PLoS ONE 10(5), e0126259 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Assadollahi V, Mohammadi E, Fathi F. et al. Effects of cigarette smoke condensate on proliferation and pluripotency gene expression in mouse embryonic stem cells. J. Cell. Biochem. 120(3), 4071–4080 (2019). [DOI] [PubMed] [Google Scholar]
- 44.Guo H, Tian L, Zhang JZ. et al. Single-cell RNA sequencing of human embryonic stem cell differentiation delineates adverse effects of nicotine on embryonic development. Stem Cell Rep. 12(4), 772–786 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yu J, Huang NF, Wilson KD. et al. nAChRs mediate human embryonic stem cell-derived endothelial cells: proliferation, apoptosis, and angiogenesis. PLoS ONE 4(9), e7040 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ishizuka T, Goshima H, Ozawa A, Watanabe Y. Effect of nicotine on the proliferation and differentiation of mouse induced pluripotent stem cells. Curr. Med. Chem. 19(30), 5164–5169 (2012). [DOI] [PubMed] [Google Scholar]
- 47.Collo G, Cavalleri L, Zoli M, Maskos U, Ratti E, Merlo Pich E. Alpha6-containing nicotinic acetylcholine receptors mediate nicotine-induced structural plasticity in mouse and human iPSC-derived dopaminergic neurons. Front. Pharmacol. 9, 572 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lee WH, Ong SG, Zhou Y. et al. Modeling cardiovascular risks of e-cigarettes with human-induced pluripotent stem cell-derived endothelial cells. J. Am. Coll. Cardiol. 73(21), 2722–2737 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]; • Tests e-liquids on human induced pluripotent stem cell-derived endothelial cells.
- 49.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126(4), 663–676 (2006). [DOI] [PubMed] [Google Scholar]
- 50.Hou P, Li Y, Zhang X. et al. Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds. Science 341(6146), 651–654 (2013). [DOI] [PubMed] [Google Scholar]
- 51.Liang Y, Li X, Zhang Y. et al. Induced pluripotent stem cells-derived mesenchymal stem cells attenuate cigarette smoke-induced cardiac remodeling and dysfunction. Front. Pharmacol. 8, 501–501 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]