

ABSTRACT
In plants, macroautophagy/autophagy has been reported to function in various biotic and abiotic stress-response pathways, but few direct regulators linking stress and autophagy have yet been identified. Other than the conserved nutrient sensing kinase TOR (Target of Rapamycin), negative regulators that can directly modulate plant autophagy are unknown. We recently identified a mutant, termed cost1 (Constitutively Stressed 1), which has strong drought tolerance with constitutive induction of autophagy and broad expression of normally stress-responsive genes. The COST1 protein negatively regulates autophagy by direct interaction with the key autophagy adaptor ATG8E, thus directly linking autophagy and drought tolerance. Moreover, plant growth and development in a cost1 mutant is greatly retarded, suggesting that COST1 controls the tradeoff between growth and stress tolerance.
KEYWORDS: Arabidopsis, autophagy, COST1, drought, stress
In plants, macroautophagy/autophagy has been reported to function in various biotic and abiotic stress-response pathways, but few direct regulators linking stress and autophagy have yet been identified. Other than the conserved nutrient sensing kinase TOR (TARGET OF RAPAMYCIN), negative regulators that can directly modulate plant autophagy are unknown. We recently identified a mutant, termed cost1 (CONSTITUTIVELY STRESSED 1), which has strong drought tolerance with constitutive induction of autophagy and broad expression of normally stress-responsive genes. The COST1 protein negatively regulates autophagy by direct interaction with the key autophagy adaptor ATG8E, thus directly linking autophagy and drought tolerance. Moreover, plant growth and development in a cost1 mutant is greatly retarded, suggesting that COST1 controls the tradeoff between growth and stress tolerance.
COST1 is a new plant-unique attenuator of autophagy
Autophagy participates in an array of biotic and abiotic stress responses, including the response to drought. Disruption of autophagy-related (ATG) genes renders plants more susceptible to starvation, drought, and other environmental stresses. However, the signaling pathways leading from drought perception to autophagy induction remain unknown. Our recent study suggests that COST1 (ID: AT2G45260; Pfam: PF04859) is one missing piece of this puzzle [1], based on the following observations: (i) a cost1 mutant has constitutively activated autophagy; (ii) autophagy is dramatically repressed in COST1-overexpression plants, especially under dehydration; (iii) cost1 mutants are drought tolerant, and this tolerance requires a functional autophagy pathway; (iv) COST1 can colocalize with the autophagy receptor NBR1 and autophagosome marker ATG8E, and physically interact with ATG8E; (v) consistent with the knowledge that autophagy has diverse functions in senescence and various plant stress responses, COST1 participates in a broad array of stress responses beyond drought, including salt tolerance and senescence.
COST1 is primarily regulated post-transcriptionally, as COST1 transcript levels are not significantly changed by various abiotic stress treatments. The COST1 protein physically interacts with the autophagosome protein ATG8E and inhibits autophagy when overexpressed, indicating that COST1 is a direct negative regulator of autophagy. After drought treatment, a portion of the COST1 protein is ubiquitinated and degraded by the 26S-proteasome. In parallel, autophagy also degrades the COST1 protein. These two pathways work cooperatively to quickly reduce the overall amount of endogenous COST1, releasing its repression of autophagy, and thus conferring plant drought tolerance. A potential feedback loop between COST1 and autophagy is depicted in the model in Figure 1: under normal growth conditions, COST1 inhibits autophagy and promotes plant growth (Figure 1, green lines); during drought, COST1 can be degraded through the 26S-proteasome and autophagy pathways, releasing its inhibition of autophagy and conferring drought tolerance (Figure 1, black arrows). Notably, in addition to its key role in autophagy, COST1 regulates the level of stress-responsive gene transcripts, including genes that are enriched in ABA, osmotic, cold and jasmonic acid pathways.
Figure 1.
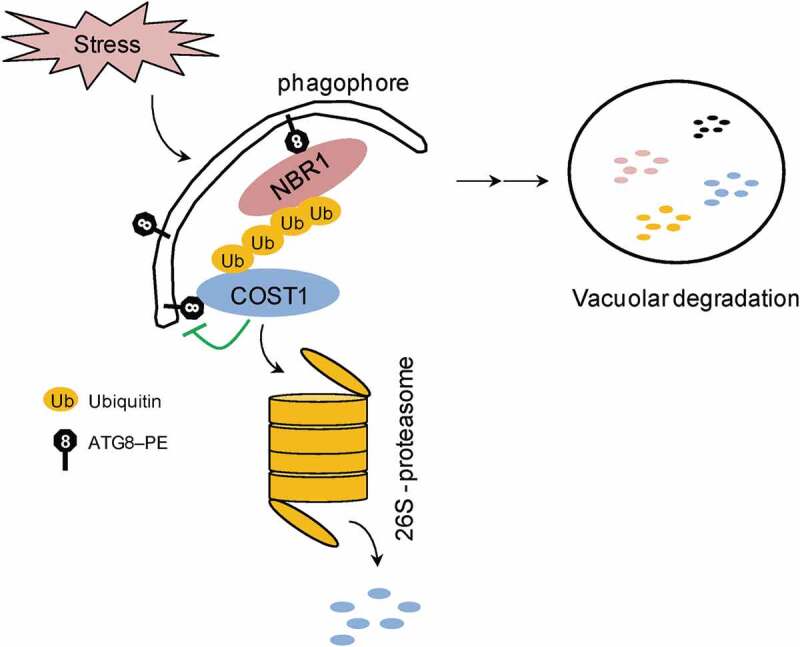
A proposed working model of COST1 function. Under normal growth conditions, COST1 inhibits autophagy and promotes plant growth (green lines). During drought stress, COST1 is degraded by both the 26S-proteasome and autophagy pathways, releasing its inhibition of autophagy and conferring stress tolerance (black arrows).
Conservation of both COST proteins and the autophagy pathway in all vascular plants suggests that the interplay between autophagy and COST1 is universal in the plant kingdom. COST1 tends to be retained as a single copy in land plant species, suggesting that this gene has been subjected to high selection pressure during evolution. By contrast, there are typically multiple copies of genes encoding the COST1-interacting partner ATG8 in plant genomes. Future detailed studies of the interactions between COST1 and ATG8 isoforms, along with narrowing down the specific domain/residues that mediate the interaction between these two proteins, will increase our understanding of the mechanism of COST1-mediated autophagy regulation.
Beyond stress responses, COST1 functions in plant growth and development
In addition to its critical role in stress tolerance, loss of COST1 dramatically affects plant growth and development. Plant size, including height and leaf area, is greatly reduced in the cost1 mutant, due to decreased cell expansion. Compared with WT plants, the cost1 mutant has increased chlorophyll per fresh weight and delayed leaf senescence. Intriguingly, while the cost1 stress tolerant and delayed leaf senescence phenotypes are restored by introduction into an autophagy-deficient mutant background, the cost1 defects in growth and development are not recovered, suggesting that COST1-controlled plant growth is autophagy-independent. Thus, there are as yet-to-be-identified endogenous signaling components controlling COST1-mediated plant growth, uncoupled from stress response pathways. Engineered versions of COST1 that restore the growth defect of the cost1 mutant but still retain cost1-like stress resistance would provide an ideal solution for generating more stress-tolerant crops without a yield penalty.
Funding Statement
This work has been supported by grant no. IOS 1353867 to DCB from the US National Science Foundation
Disclosure statement
No potential conflict of interest was reported by the authors.
Reference
- [1].Bao Y, Song WM, Wang P, Yu X, Li B, Jiang C, Shiu SH, Zhang H, and Bassham DC.. COST1 regulates autophagy to control plant drought tolerance. Proc Natl Acad Sci U S A. 2020. doi: 10.1073/pnas.1918539117 [DOI] [PMC free article] [PubMed] [Google Scholar]