

Abstract
IgD-Fc-Ig fusion protein, a new biological agent, is constructed by linking a segment of human IgD-Fc with a segment of human IgG1-Fc, which specifically blocks the IgD-IgDR pathway and selectively inhibits the abnormal proliferation, activation, and differentiation of T cells. In this study we investigated whether IgD-Fc-Ig exerted therapeutic effects in collagen-induced arthritis (CIA) rats. CIA rats were treated with IgD-Fc-Ig (1, 3, and 9 mg/kg) or injected with biological agents etanercept (3 mg/kg) once every 3 days for 40 days. In the PBMCs and spleen lymphocytes of CIA rats, both T and B cells exhibited abnormal proliferation; the percentages of CD3+ total T cells, CD3+CD4+ Th cells, CD3+CD4+CD25+-activated Th cells, Th1(CD4+IFN-γ+), and Th17(CD4+IL-17+) were significantly increased, whereas the Treg (CD4+CD25+Foxp3+) cell percentage was decreased. IgD-Fc-Ig administration dose-dependently decreased the indicators of arthritis; alleviated the histopathology of spleen and joint; reduced serum inflammatory cytokines levels; decreased the percentages of CD3+ total T cells, CD3+CD4+ Th cells, CD3+CD4+CD25+-activated Th cells, Th1 (CD4+IFN-γ+), and Th17(CD4+IL-17+); increased Treg (CD4+CD25+Foxp3+) cell percentage; and down-regulated the expression of key molecules in IgD-IgDR-Lck-NF-κB signaling (p-Lck, p-ZAP70, p-P38, p-NF-κB65). Treatment of normal T cells with IgD (9 μg/mL) in vitro promoted their proliferation. Co-treatment with IgD-Fc-Ig (0.1–10 μg/mL) dose-dependently decreased IgD-stimulated T cell subsets percentages and down-regulated the IgD-IgDR-Lck-NF-κB signaling. In summary, this study demonstrates that IgD-Fc-Ig alleviates CIA and regulates the functions of T cells through inhibiting IgD-IgDR-Lck-NF-κB signaling.
Keywords: IgD, IgDR, IgD-Fc-Ig fusion protein, CD4+ T cell, collagen-induced-arthritis, IgD-IgDR-Lck-NF-κB signaling, etanercept
Introduction
Rheumatoid arthritis (RA) is a chronic systemic autoimmune disease. Synovitis is the main pathological manifestation [1]. The pathogenesis of RA is related to the abnormal activation of T and B cells. Activated CD4+ T cells differentiate into special effector cells and play an important role in specific immune responses. A large number of activated CD4+ T cells infiltrate into synovial tissue and secrete inflammatory cytokines that cause inflammation in the RA synovium [2]. The abnormalities of CD4+ T cells mainly manifest as an imbalance in the T cell subsets. Th1 and Th2 cells mediate cellular immune and humoral immune responses, and Tregs mainly inhibit autoimmunity. Th17 cells mediate autoimmune and inflammatory reactions [3].
Currently, the main therapeutic drugs for RA are steroidal anti-inflammatory drugs, nonsteroidal anti-inflammatory drugs, disease-modifying drugs, and biological agents. However, these drugs have only partial effects on the relief of joint inflammation and result in many adverse reactions, such as gastrointestinal side effects, hepatotoxicity, and myelosuppression, which seriously hinder the therapeutic effects. Therefore, it is of great clinical significance to explore new therapeutic targets for RA and develop new therapeutic drugs for RA.
Immunoglobulin D (IgD) was discovered in 1965 and consists of two identical light chains and two heavy chains [4]. IgD includes secreted IgD (sIgD) and membrane-bound IgD (mIgD). The C-terminus of mIgD has more transmembrane and intracellular regions than that of sIgD. These structural features and molecular configurations are important for the specific biological functions of IgD. mIgD is a hallmark of mature B cells. Although IgD levels are low in the body, IgD exerts important immune functions [5, 6]. An abnormally elevated level of IgD can lead to inflammation and tissue damage by inducing the activation and infiltration of immune cells [7, 8]. Accumulating evidence has suggested that IgD may contribute to some autoimmune diseases, such as RA, systemic lupus erythematosus (SLE) and Sjogren’s syndrome (SS) [9]. Previous studies in our lab showed that the levels of sIgD and mIgD in patients with RA were higher than those in healthy controls, and after stimulation with IgD, the expression of IgDR on peripheral T and B cells was significantly increased [10]. In an IgD transgenic mouse model, skin ulcers and abnormal swelling of the liver, spleen and kidney were found, which may be related to the high expression of sIgD in serum [11].
In 1980, IgDR was discovered on T cells in human peripheral blood. sIgD exerts biological functions by binding to a specific membrane receptor (IgDR). The molecular weight of IgDR is approximately 70 kDa, but the IgDR gene is still unknown and has not yet been cloned [12–14]. Studies have shown that IgD can induce IgDR expression to increase by more than five-fold on the surfaces of T cells, and IgDR expression on the T cells of RA patients can increase by more than 12-fold [15]. Studies have shown that protein tyrosine kinase (PTK) is involved in the process of IgD-induced IgDR expression. Therefore, we have hypothesized that IgD may activate some key proteins downstream of the PTK pathway, but PTK inhibitors can completely block the induction by IgD on IgDR expression, suggesting that IgD may induce IgDR expression through one or more PTK activation pathways. The lymphocyte-specific protein tyrosine kinase (Lck) and zeta-associated protein 70 (ZAP70), which are involved in the PTK signaling pathway, play important roles in T cell activation. Animal experiments show that IgD activates the phosphorylation of Lck and phospholipase C-γ (PLC-γ) in T cell hybridomas [16]. Lck mainly exists in T cells and participates in signal transduction that is involved in T cell development, differentiation, and activation [17, 18]. The interaction of T cells with antigen-presenting cells (APC) leads to Lck activation and the phosphorylation of immunoreceptor tyrosine-based activation motifs (ITAMs). The activation of Lck kinase induces the production of the SH2 domain, which binds to ZAP70 kinase, thereby activating ZAP70 kinase. The activation of ZAP70 kinase further stimulates multiple signaling networks downstream, including calcium mobilization, Ras/MAPK and PI3K pathways [19]. Under normal conditions, the expression and activity of Lck are limited. When stimulated by certain external factors, the expression and activity of Lck can become abnormal, causing some autoimmune diseases, such as RA and SLE [20]. The above findings suggest that Lck may be a key downstream signaling molecule for IgD/IgDR. IgD may induce abnormal T cell activation through activation of the IgD-IgDR-Lck-NF-κB signaling pathway.
The development of new targeted drugs for RA has important clinical implications. Based on previous results, our research team successfully linked the human IgD-Fc segment to the human IgG1-Fc segment to form a fusion protein (IgD-Fc-Ig) targeting IgD/IgDR, and IgD-Fc-Ig has been patented in China. IgD-Fc-Ig competes with overexpressed IgD for binding to IgDR, which specifically blocks the IgD/IgDR-mediated signaling pathway and selectively reduces the abnormal activation of T and B cells. Etanercept is an anti-TNF-α agent that reduces the percentage of B cells in the peripheral blood of RA patients by neutralizing TNF-α, which is crucial for B cell proliferation. Etanercept inhibits memory B cell induction by interfering with germinal center formation and the follicular dendritic cell network [21, 22]. It has been unclear whether IgD-Fc-Ig has therapeutic effects on experimental arthritis. Does IgD-Fc-Ig affect the proliferation, activation, and differentiation of T cells in experimentally induced arthritis? Could IgD-Fc-Ig regulate the IgD-IgDR-Lck signaling pathway in T cells? To answer these questions, in this study, rats with collagen-induced arthritis (CIA) were used as subjects to observe the therapeutic effects of IgD-Fc-Ig on CIA and T cell proliferation, activation and differentiation in rats, and to elucidate the mechanisms by which IgD-Fc-Ig inhibits T cell proliferation, activation, and differentiation by blocking the IgD/IgD signaling pathway. This study provides an experimental basis for further understanding the pathogenesis of RA and elucidating the characteristics and mechanisms of the treatment of RA with IgD-Fc-Ig.
Materials and methods
Animals
Wistar rats (male, 150 ± 20 g) were purchased from Beijing Vital River Laboratory Animal Technology Company (Certificate No. SCXK [Jing] 2016-0006). The rats were maintained under standard laboratory conditions. All studies adhered to the principles of laboratory animal care and were approved by the Ethics Review Committee for Animal Experimentation of the Institute of Clinical Pharmacology, Anhui Medical University.
Reagents
Roswell Park Memorial Institute (RPMI)-1640 medium and fetal bovine serum (FBS) were obtained from Gibco Co. (CA, USA). The CCK-8 kit was purchased from Tianjin Baiying Biological Technology Co., Ltd (China). The enzyme-linked immunosorbent assay (ELISA) kits for IL-4 and IL-17 were purchased from Research & Development Co., Ltd (Minneapolis, MN, USA). Chicken type II collagen (CII) was purchased from Chondrex, Inc. (Redmond, WA, USA). Freund’s complete adjuvant was purchased from Sigma. T/B lymphocyte separation fluid was purchased from Dakewe Biotechnology Co., Ltd. Human IgD protein was purchased from Abcam (Cambridge, MA, USA). The cell stimulation cocktail was purchased from eBioscience (San Diego, CA, USA). Anti-rat FITC-CD3 (559975), PE-CyTM7-CD4 (561578), PE-CD25 (554866), APC-IFN-γ (562213), PE-IL-4 (555082), PE-IL-17, and FITC-Foxp3 (4307321) antibodies were obtained from BD Pharmingen (San Diego, CA, USA). The β-actin antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies against Lck (YT2545) and phosphorylated-Lck (p-Lck) (SAB4300118) were obtained from Sigma-Aldrich (St. Louis, MO, USA). ZAP70 (ab32410), phosphorylated-ZAP70 (p-ZAP70), p38 mitogen-activated protein kinase (P38) (ab170099), phosphorylated P38 (p-P38) (4511T), NF-κB65 (8242T), and p-NF-κB65 (3033T) were obtained from Abcam (Cambridge, MA, USA). The rat protein antibody array was obtained from RayBiotech (Norcross, GA, USA).
Drugs
The IgD-Fc-Ig fusion protein was synthesized by our laboratory. Etanercept was provided by Shanghai Guojian Pharmaceutical Co., Ltd (China).
Induction and treatment of CIA
CII was dissolved in 0.1 mol/L acetic acid at a concentration of 4 mg/mL overnight at 4 °C under sterile conditions. The CII mixture was then emulsified with an equal volume of Freund’s complete adjuvant (6 mg/mL). The emulsion of CII (0.2 mL) was injected intradermally into the back, the base of the tail and the foot of rats on day 0 to induce CIA, followed by a booster injection on D7. After the onset of arthritis on D7, the number of rats was 10 in each group. The CIA rats were randomly divided into five groups and treated with IgD-Fc-Ig (1, 3, and 9 mg/kg) or etanercept (3 mg/kg) once every 3 days from D17 to D55. The normal rats and CIA model rats were given equal volumes of normal sodium, which were administered at the same time.
Clinical assessment of CIA
An evaluation of the severity of CIA was performed by two independent observers with no knowledge of the treatment protocol. Beginning on D10 after immunization, the rats were evaluated every 3 days using three clinical parameters: the arthritis index (AI), the swollen joint count (SJC), and the paw swelling volume. A scale from 0 to 4 was used to grade the swelling: 0 = no swelling or spiloplaxia; 1 = spiloplaxia and the slight swelling of the ankle; 2 = spiloplaxia and the slight swelling of the joints, including the ankle, metatarsal joints, and palm joints; 3 = spiloplaxia and the moderate swelling of the joints, including the ankle, metatarsal joints, and palm joints; 4 = spiloplaxia and the severe swelling of the joints, including the ankle and metatarsal joints. All feet were evaluated, and the maximum score value for each rat was 16. Each paw has five phalanx joints and one ankle or wrist joint, so the maximum SJC for each rat was 24 [23]. The weight and paw volumes of the CIA rats were also evaluated once every 3 days after immunization.
The measurement of the thymus index and the spleen index
The rats were sacrificed by cervical dislocation after ether anesthesia on D55 after immunization. The thymus and spleen were aseptically removed. The thymus and spleen indices were determined according to the ratio of the weight of the thymus or spleen and the weight of the rat.
Analysis of T and B lymphocyte proliferation
T lymphocytes from the thymus and B lymphocytes from the spleen were separated with lymphocyte separation fluid in each group, and the concentration of cells was adjusted to 1 × 106/mL. T or B lymphocyte suspensions (200 μL) in RPMI-1640 supplemented with 10% fetal calf serum were seeded into 96-well culture plates (1 × 105 cells per well). ConA and LPS were used to stimulate the T and B cells, which were cultured for an additional 48 h. Triplicate measurements were performed. Ten microliters of CCK-8 reagent was added to each well. The cells were incubated at 37 °C in 5% CO2 for 2 h. Then, the absorbance was measured at 450 nm.
Histopathological examination of the spleen and joints
Paraffin sections of the spleen and joints were stained with hematoxylin and eosin. The histological changes were evaluated by two independent observers. Five parameters were evaluated in the spleen: the periarteriolar lymphoid sheaths (PALs), lymphoid follicles, marginal zone, red pulp, and the total number of germinal centers (GCs). Five parameters were assessed in the joints: synovial hyperplasia, the pannus, cartilage erosion, and infiltrating inflammatory cells. The grading scheme consisted of ordinal categories ranging from 0 (no effect) to 4 (severe effect) [24, 25].
The percentages of the T cell subsets in peripheral blood mononuclear cells and splenic cells from CIA rats were analyzed by flow cytometry
Peripheral blood mononuclear cells (PBMCs) were obtained from the blood of rats from erythrocyte lysate. The lymphocytes in spleens were separated using lymphocyte separation fluid. A 100 μL aliquot of lymphocyte cell suspension was then transferred into a round-bottomed tube. Antibody combinations of CD3-FITC, CD4-PE-CyTM7, and CD25-PE, CD4-PE-CyTM7, and IFN-γ-APC, CD4-PE-CyTM7 and IL-4-PE or CD4-PE-CyTM7 and IL-17-PE were added into the flow tube. After 40 min of incubation at room temperature in the dark, the cells were washed and analyzed immediately by flow cytometry. However, the cell suspensions used for detecting the percentage of Th17 cells were cultured for 8 h with RPMI-1640 supplemented with 10% fetal calf serum and a cell-stimulating agent. When detecting the percentages of Th1, Th2, Th17, and Treg cells, the cells were first labeled with surface antibodies, and then intracellular antibodies were added after the disruption of the cell membrane. The cell-associated fluorescence was analyzed with a Cytoflex flow cytometer (Beckman Coulter, USA). In vitro, the same method as that described above was used to examine the effects of IgD-Fc-Ig on the differentiation of T cells.
The levels of IL-4 and IL-17 were measured by ELISA
The levels of serum IL-4 and IL-17 were measured. Each serum sample was assayed in triplicate. First, assay diluent (80 μL) was added to each well of a 96-well plate. The standard or sample (40 μL) was added to each well and incubated for 2 h at room temperature. After washing, rat antibodies against IL-4 and IL-17 were added to each well and incubated for 2 h. Then, the substrate solution was added to each well and incubated for 30 min. The optical density was measured at 450 nm.
The inflammatory cytokine levels were measured by protein chip array systems
The levels of serum IFN-γ, IL-1β, IL-6, TNF-α, leptin, monocyte chemoattractant protein-1 (MCP-1), and tissue inhibitor of metalloproteinase-1 (TIMP-1) were analyzed. The steps of the procedure were as follows. (1) The diluent (100 μL) was added to each well and incubated for 30 min at room temperature for blocking. After discarding the buffer, the samples (100 μL) were added to each well and incubated for 1–2 h at room temperature. (2) A cocktail of detection antibodies (80 μL) was added to each well, and the glass chip was incubated for 1–2 h. (3) The Cy3 equivalent dye streptavidin (80 μL) was injected into each well. The device was covered with aluminum foil and incubated at room temperature for 1 h. (4) Fluorescence detection was performed using a laser scanner, such as the Axon GenePix, at the appropriate wavelength for Cy3.
T cell magnetic separation
T cells in spleens were isolated using magnetic cell separation (MACS) by positive selection. The lymphocytes (108 cells) in the thymus and spleen were isolated by lymphocyte separation fluid. PBMCs were incubated with anti-CD3-FITC in MACS buffer for 20 min, followed by incubation with anti-PE beads for an additional 20 min. The stained cell suspension was applied to an LS column (Miltenyi Biotec, Germany). The labeled T cells were collected after washing with degassed buffer three times.
The expression levels of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 were analyzed by Western blotting
The T cells isolated by MACS were lysed in cell lysis buffer with 1 mM PMSF at 4 °C for 30 min, followed by centrifugation at 14,000 × g for 10 min at 4 °C. The proteins were isolated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. The membranes were blocked with blocking buffer for 2 h at room temperature and then incubated with primary monoclonal antibodies against rat Lck, p-Lck, ZAP70, p-ZAP70, P38, p-P38, NF-κB65, and p-NF-κB65 and rat monoclonal anti-β-actin antibody at 4 °C overnight.
Effects of IgD-Fc-Ig on T cell activation stimulated by IgD in vitro
Lymphocytes from spleens were separated by lymphocyte separation fluid, and then the T cells were isolated using MACS with positive selection. The final concentration of the cells was adjusted to 1 × 106/mL. IgD (9 μg/mL), IgD-Fc-Ig (0.1, 1, and 10 μg/mL), and etanercept (2 μg/mL) were incubated with T cells for 48 h (37 °C). In vitro, carboxyfluorescein succinimidylamino ester (CFSE) was used to detect the effect of IgD-Fc-Ig on T cell proliferation. The percentage of proliferating CD3+ T cells was measured by flow cytometry according to the CFSE instructions. The same method as that described above was used to examine the effects of IgD-Fc-Ig on T cell proliferation, the percentages of the T cell subsets, and the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 on IgD-stimulated T cells.
Statistical analysis
SPSS 16.0 was used for the statistical analysis, and the two groups were compared using the t-test. The differences between groups were compared with one-way analysis of variance (ANOVA). Data were expressed as the mean ± standard deviation (SD). P values less than 0.05 were considered to be significant.
Results
IgD-Fc-Ig relieved paw swelling, decreased the paw volume, AI and SJC, and reduced weight loss in CIA rats
The results showed that the onset of inflammation appeared on approximately D17 after primary immunization. The forefeet and hind feet appeared to suffer from erythema and swelling in sequence, and nodules were evident on the tail. The swelling of rat paws during different stages of inflammation was observed. Compared with the untreated CIA rats, IgD-Fc-Ig was found to relieve paw swelling in treated CIA rats (Fig. 1a). The peak paw volume in the untreated CIA rats appeared on D27 after primary immunization (Fig. 1b). Paw swelling in the CIA rats in the IgD-Fc-Ig (9 mg/kg) group gradually decreased after D31. IgD-Fc-Ig decreased the AI and SJC and reduced weight loss in CIA rats. The AI in the IgD-Fc-Ig (9 mg/kg) group was significantly reduced on D48 (Fig. 1c). The SJC in the etanercept group was reduced on D41 (Fig. 1d). Compared with those of the untreated CIA rats, the weights of the IgD-Fc-Ig (9 mg/kg) group rats were significantly higher on D34 (Fig. 1e). Etanercept decreased AI on D34.
Fig. 1. The effects of IgD-Fc-Ig on paw swelling, AI, SJC, weight, paw volume, thymus/spleen indices, and T/B cell proliferation in CIA rats.

a The effects of IgD-Fc-Ig on paw swelling in CIA rats during different inflammation stages. b The paw swelling volume in CIA rats and the effects of IgD-Fc-Ig was observed. c The AI of CIA rats was assessed, and the effects of IgD-Fc-Ig were assessed. d The SJC of CIA rats was observed, and the effects of IgD-Fc-Ig were observed. e The weight of CIA rats was observed, and the effects of IgD-Fc-Ig were observed. f The effects of IgD-Fc-Ig on the thymus and spleen indices in CIA rats. g The effects of IgD-Fc-Ig on the proliferation of T and B cells in CIA rats. *P < 0.05, **P < 0.01 vs Normal; #P < 0.05 vs CIA; $P < 0.05 vs IgD-Fc-Ig (9 mg/kg) (n = 10).
IgD-Fc-Ig decreased the thymus and spleen indices and inhibited the proliferation of thymus T cells and spleen B cells
Compared with those in normal rats, the thymus and spleen indices were obviously increased in the treated rats. IgD-Fc-Ig (9 mg/kg) and etanercept decreased the thymus and spleen indices. The effects of etanercept on the thymus and spleen indices were stronger than those of IgD-Fc-Ig (9 mg/kg) (Fig. 1f). The proliferation of T/B cells was measured by CCK-8 assays. The proliferation of T and B cells in CIA rats was significantly higher than that in normal rats. IgD-Fc-Ig (9 mg/kg) and etanercept clearly inhibited the abnormal proliferation of T and B cells (Fig. 1g).
IgD-Fc-Ig alleviated spleen and joint histopathological changes
Histopathological examinations of the spleen and joints were performed by H&E staining. An increased number of GCs and infiltrating inflammatory cells and increased lymphoid follicular hyperplasia were observed in CIA rats. Compared with normal rats, an increase in PALs, the marginal zone, and red pulp were also found in CIA rats. Compared with CIA rats, IgD-Fc-Ig (9 mg/kg) and etanercept alleviated these abnormalities. Red pulp congestion was alleviated, and the number of GCs was reduced in the IgD-Fc-Ig (9 mg/kg) and etanercept groups (Fig. 2a, b). There were marked differences among the IgD-Fc-Ig (9 mg/kg), etanercept, and CIA groups. Synovial hyperplasia and pannus formation were observed in CIA rats, as well as the destruction of cartilage, the infiltration of inflammatory cells into the synovium, and the elevation of inflammation scores. IgD-Fc-Ig (9 mg/kg) and etanercept significantly improved the pathological changes caused by synovial cell hyperplasia, inflammatory cell infiltration, and pannus formation (Fig. 2c, d).
Fig. 2. Histopathological evaluation of the spleen and joints in CIA rats and the effects of IgD-Fc-Ig.

a The effects of IgD-Fc-Ig on histopathological changes in the spleen. In normal rats, → shows red pulp and ↓ shows white pulp. In CIA rats, → shows germinal centers, ↓ shows lymphoid follicular hyperplasia, ↙ shows periarteriolar lymphoid sheaths, and ↘ shows red medullary congestion. b Comparison of the pathological changes in the spleen in different groups. c The effects of IgD-Fc-Ig on histopathological changes in the joints. In normal rats, ↓ shows the synovial membrane. In CIA rats, ↑ shows synovial hyperplasia, ↓ shows inflammatory cell infiltration, ← shows vascular pannus, and ↖ shows cartilage damage. d Comparison of the pathological changes in joints in different groups. #P < 0.05, ##P < 0.01 vs CIA (n = 6).
The percentages of T cell subsets among PBMCs from CIA rats and the effect of IgD-Fc-Ig on D55
The percentages of the T cell subsets in PBMCs were analyzed by flow cytometry. Compared with those in normal rats, the percentages of total CD3+, CD3+CD4+, and CD3+CD4+CD25+ T cells were increased in CIA rats. Compared with the CIA rats, the IgD-Fc-Ig (9 mg/kg) and etanercept groups showed obviously decreased percentages of total CD3+, CD3+CD4+, and CD3+CD4+CD25+ T cells (Fig. 3a–c).
Fig. 3. The effects of IgD-Fc-Ig on CD3+CD4+CD25+ T cell subsets in PBMCs on D55 and in the spleens of CIA rats.
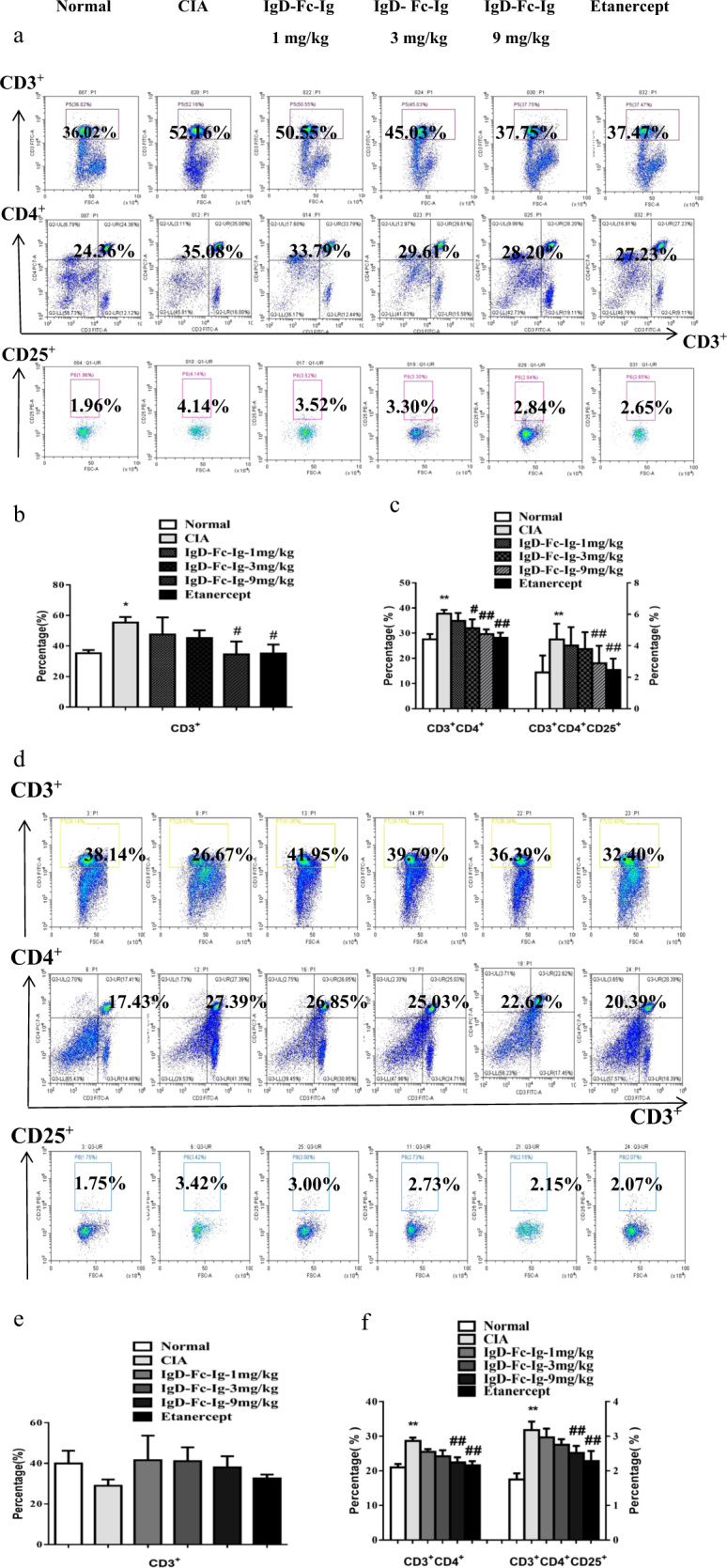
a The percentages of the T cell subsets in PBMCs were observed by flow cytometry. b Bar graphs show the percentage of total CD3+ T cells in CIA rats on D55 and the effects of IgD-Fc-Ig. c Bar graphs show the percentages of CD3+CD4+ and CD3+CD4+CD25+ T cells in CIA rats on D55 and the effects of IgD-Fc-Ig. d The percentages of T cell subsets in the spleen were observed by flow cytometry. e Bar graphs show the percentage of total CD3+ T cells and the effects of IgD-Fc-Ig. f Bar graphs show the percentages of CD3+CD4+ and CD3+CD4+CD25+ T cells in CIA rats and the effects of IgD-Fc-Ig. *P < 0.05, **P < 0.01 vs Normal, #P < 0.05, ##P < 0.01 vs CIA (n = 10).
IgD-Fc-Ig regulated the differentiation of T cell subsets
Compared with those in normal rats, the CD3+CD4+ and CD3+CD4+CD25+ T cell percentages were increased in CIA rats. IgD-Fc-Ig (9 mg/kg) and etanercept decreased the percentages of CD3+CD4+ and CD3+CD4+CD25+ T cells, but IgD-Fc-Ig had no effect on the total CD3+ T cells (Fig. 3d–f). The percentages of CD4+IFN-γ+ Th1 and CD4+IL-17+ Th17 cells were increased, and the percentage of CD4+CD25+Foxp3+ Tregs was decreased in CIA rats. IgD-Fc-Ig (9 mg/kg) and etanercept decreased the percentages of CD4+ IFN-γ+ Th1 and CD4+ IL-17+ Th17 cells and increased the percentage of CD4+CD25+Foxp3+ Tregs, but IgD-Fc-Ig had no effect on CD4+IL-4+ Th2 cells (Fig. 4a–d).
Fig. 4. The effects of IgD-Fc-Ig on T cell subsets in the spleens of CIA rats.

a The percentages of Th17 and Treg cells in the spleen were observed by flow cytometry. b Bar graphs show the percentages of CD4+IL-17+ Th17 and CD4+CD25+Foxp3+ Treg cells. c The percentages of Th1 and Th2 cells in the spleen were observed by flow cytometry. d Bar graphs show the percentages of CD4+IFN-γ+ Th1 and CD4+IL-4+ Th2 cells. **P < 0.01 vs Normal; #P < 0.05, ##P < 0.01 vs CIA (n = 10).
IgD-Fc-Ig regulated cytokine levels in the serum of CIA rats
The levels of serum IL-4 and IL-17 were detected by ELISA. IL-17 levels were significantly elevated in CIA rats. IgD-Fc-Ig (9 mg/kg) and etanercept decreased IL-17 levels, but IgD-Fc-Ig had no effect on IL-4 in CIA rats (Fig. 5a). Compared with those in normal rats, the serum levels of ICAM-1, IFN-γ, IL-1β, IL-6, TNF-α, leptin, and MCP-1 were significantly increased in CIA rats. TIMP-1 levels were significantly decreased in CIA rat serum. IgD-Fc-Ig (9 mg/kg) and etanercept decreased the levels of ICAM-1, IFN-γ, IL-1β, IL-6, TNF-α, leptin, and MCP-1 and increased TIMP-1 levels (Fig. 5b–e).
Fig. 5. The levels of serum IL-4, IL-17, IFN-γ, IL-1β, IL-6, TNF-α, leptin, MCP-1, and TIMP-1 in CIA rats and the effects of IgD-Fc-Ig on the expression of p-Lck, p-ZAP-70, p-P38, and p-NF-κB65.

a The levels of serum IL-4 and IL-17 in CIA rats and the effects of IgD-Fc-Ig. b The level of serum IL-1β in CIA rats and the effects of IgD-Fc-Ig. c The levels of serum IL-6 and IFN-γ in CIA rats and the effects of IgD-Fc-Ig. d The levels of serum MCP-1 and leptin in CIA rats and the effects of IgD-Fc-Ig. e The levels of serum TNF-α and TIMP-1 in CIA rats and the effects of IgD-Fc-Ig. f The expression levels of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 in T cells in the spleens of CIA rats were analyzed by Western blotting. g Comparison of protein expression in different groups. The effects of IgD-Fc-Ig on the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65. *P < 0.05, **P < 0.01 vs Normal, ##P < 0.01 vs CIA (n = 10).
IgD-Fc-Ig inhibited the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 in CIA rats
The expression levels of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 were analyzed by Western blotting. Compared with those in normal rats, the expression levels of p-Lck (tyr394), p-ZAP70 (Tyr319), p-P38, and p-NF-κB65 were obviously increased in CIA rats. IgD-Fc-Ig (9 mg/kg) and etanercept decreased the elevation of the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 in the spleens of CIA rats. IgD-Fc-Ig (9 mg/kg) and etanercept reduced p-Lck expression to less than normal levels (Fig. 5f, g).
IgD-Fc-Ig inhibited IgD-induced T cell proliferation and activation and the differentiation of T cell subsets in vitro
Cell proliferation experiments showed that IgD (9 μg/mL) significantly promoted the proliferation of normal T cells (Fig. 6a). IgD (9 μg/mL), IgD-Fc-Ig (0.1, 1, and 10 μg/mL), and etanercept (2 μg/mL) were incubated with T cells for 48 h (37 °C). IgD-Fc-Ig (10 μg/mL) and etanercept (2 μg/mL) inhibited T cell proliferation stimulated by IgD (Fig. 6b, c). Compared with those in the control group, the CD3+CD4+ and CD3+CD4+CD25+ T cell percentages were obviously increased in the IgD (9 μg/mL) group. IgD-Fc-Ig (10 μg/mL) and etanercept (2 μg/mL) decreased the percentages of CD3+CD4+ and CD3+CD4+CD25+ T cells, but IgD-Fc-Ig had no effect on total CD3+ T cells (Fig. 6d–f). The percentages of CD4+IFN-γ+ Th1 and CD4+IL-17+ Th17 cells were increased, and the percentage of CD4+CD25+Foxp3+ Tregs was decreased in the IgD (9 μg/mL) group. IgD-Fc-Ig (10 μg/mL) and etanercept (2 μg/mL) decreased the percentages of CD4+IFN-γ+ Th1 and CD4+IL-17+ Th17 cells and increased the percentage of CD4+CD25+Foxp3+ Tregs, but IgD-Fc-Ig had no effect on CD4+IL-4+ Th2 cells (Fig. 7a–d).
Fig. 6. The effects of IgD-Fc-Ig on the function of T cells stimulated by IgD.
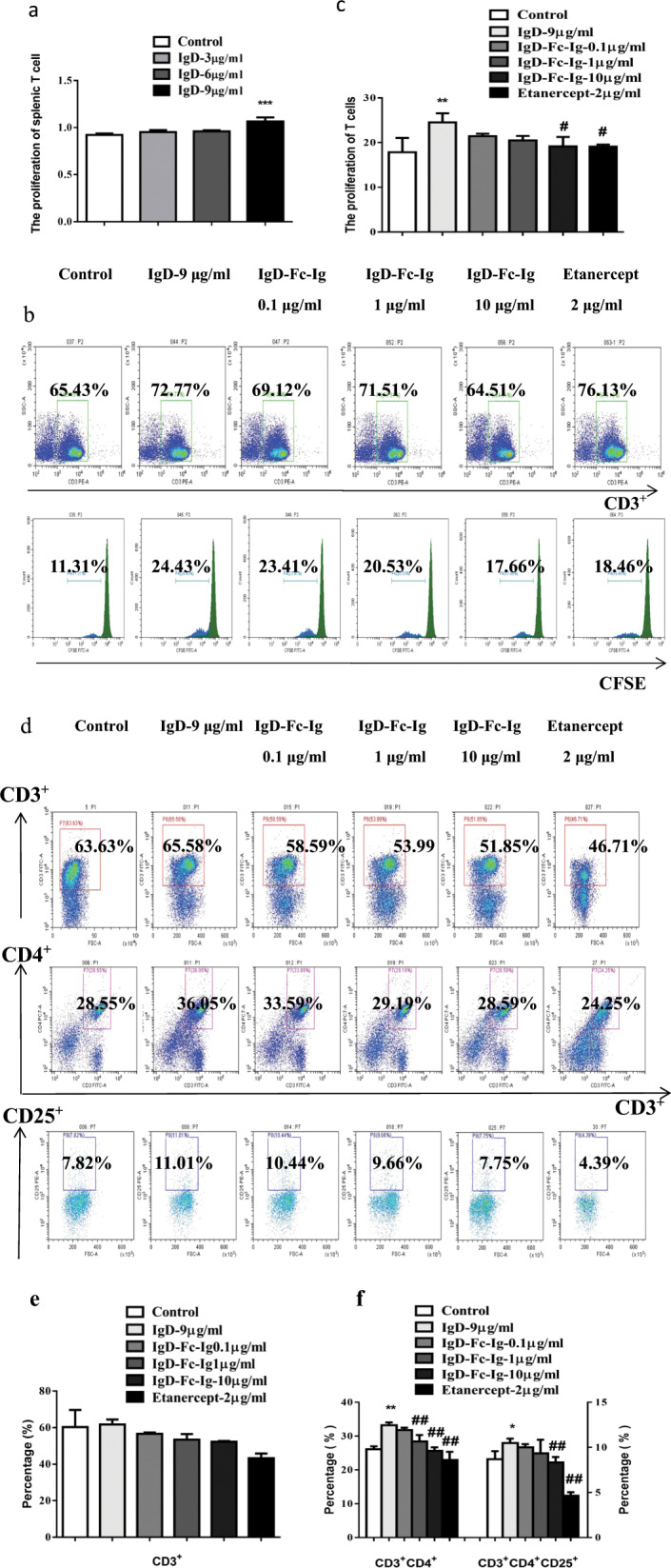
a The effect of IgD at different concentrations on the proliferation of T cells. b The proliferation of total CD3+ T cells was observed with CFSE. c Bar graphs show the proliferation of total CD3+ T cells. d The percentages of T cell subsets stimulated by IgD were observed by flow cytometry. e Bar graphs show the percentage of total CD3+ T cells. f Bar graphs show the percentages of CD3+CD4+ and CD3+CD4+CD25+ T cells. *P < 0.05, **P < 0.01 vs Control; #P < 0.05, ##P < 0.01 vs IgD 9 μg/mL (n = 10).
Fig. 7. The effects of IgD-Fc-Ig on T cell subsets and the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 in T cells stimulated by IgD.

a The percentages of Th17 and Treg cells were observed by flow cytometry. b Bar graphs show the percentages of CD4+IL-17+ Th17 cells and CD4+CD25+Foxp3+ Treg cells. c The percentages of Th1 and Th2 cells were observed by flow cytometry. d Bar graphs show the percentages of CD4+IFN-γ+ Th1 and CD4+IL-4+ Th2 cells. e The expression levels of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 were analyzed by Western blotting. f The effects of IgD-Fc-Ig on the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65. **P < 0.01 vs Control; #P < 0.05, ##P < 0.01 vs IgD 9 μg/mL (n = 10).
IgD-Fc-Ig inhibited the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 in normal T cells stimulated by IgD in vitro
The expression levels of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 were analyzed by Western blotting. Compared with those in the control group, the expression levels of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 were obviously increased in normal T cells stimulated by IgD. IgD-Fc-Ig (10 μg/mL) and etanercept (2 μg/mL) decreased the elevation of the expression of p-Lck, p-ZAP70, p-P38, and p-NF-κB65 in normal T cells stimulated by IgD (Fig. 7e, f).
Discussion
RA is a chronic autoimmune disease characterized by synovial hyperplasia and the progressive destruction of articular cartilage [26–28]. CIA rats are a classic animal model used for studies of the pathological mechanisms and therapeutic drugs of RA [29, 30]. In this study, the paw volume, AI, and SJC of CIA rats were significantly increased, while the weights of CIA rats were obviously decreased, and the proliferation of T and B cells in CIA rats was significantly higher than that in normal rats. IgD-Fc-Ig and etanercept could clearly reduce these abnormalities. At the same time, synovial hyperplasia, pannus formation, and destroyed cartilage were observed in CIA rats, and IgD-Fc-Ig and etanercept markedly alleviated these pathological changes. These results suggest that IgD-Fc-Ig has a good therapeutic effect on CIA rats.
CD4+ T cells play an important role in coordinating protective immune responses to various pathogens, but CD4+ T cells also contribute to the development of allergies, inflammation, and immune responses in autoimmune diseases. CD4+ T cells can differentiate into T cell subsets that perform different functions [31], including Th1, Th2, and Th17 cells and Tregs. Th1 and Th2 cells mediate cellular and humoral immune responses, respectively [32]. Tregs mainly suppress autoimmunity. Th17 cells mainly participate in autoimmune and inflammatory reactions. Therefore, disorders of T cell subsets may be closely related to the occurrence and development of autoimmune diseases. During the differentiation of Th1 cell subsets, Th1 cells can secrete cytokines and induce the development of inflammatory processes. Th2 cells mainly secrete IL-4, which mediates the humoral immune response. Studies have shown that Th1/Th2 imbalances play a role in RA and SLE. These studies have indicated that the imbalance of Th1/Th2 is closely related to the pathogenesis of autoimmune diseases [33–35]. The IL-17 level is high in the peripheral blood of RA patients, and this abnormality indicates that Th17 cells play an important role in RA. IL-17 is the major effector cytokine of Th17 cells. IL-17 can induce the production of TNF-α, and TNF-α is the main pathogenic cytokine of RA. In addition, IL-17 can cooperate with TNF-α and IL-1 to promote the chronic inflammatory progression of RA. Tregs are characterized by the nuclear expression of Foxp3. Tregs secrete inhibitory cytokines to exert anti-inflammatory and immune tolerance effects [36]. In this study, the percentages of CD3+CD4+ T cells, CD3+CD4+CD25+ T cells, CD4+IFN-γ+ Th1 cells, and CD4+IL-17+ Th17 cells were elevated, and the percentage of CD4+CD25+Foxp3+ Tregs was decreased in CIA rats. IgD-Fc-Ig and etanercept significantly decreased the percentages of abnormally increased T cell subsets and increased the percentage of Tregs. These results indicate that IgD-Fc-Ig could inhibit the activation and differentiation of T cells, suggesting that IgD-Fc-Ig exerts a therapeutic effect on CIA rats by regulating the function of T cells.
T cell subsets secrete many inflammatory cytokines during the immune response. Cytokines are extracellular signaling molecules that mediate cell-to-cell communication and play critical roles in many biological processes, such as cellular growth and differentiation, immunity and inflammation [37]. In this study, the levels of IL-17, ICAM-1, IFN-γ, IL-1β, IL-6, TNF-α, leptin, and MCP-1 were significantly increased, and TIMP-1 levels were significantly decreased in CIA rat serum. IgD-Fc-Ig and etanercept could decrease the high levels of these cytokines and increase the level of TIMP-1. These results suggest that IgD-Fc-Ig inhibits the secretion of inflammatory cytokines by regulating the functions of T cells.
IgDR is expressed on the surfaces of T cells; human CD4+ T cells and CD8+ T cells express IgDR, and IgD binds to IgDR and mediates the activation of T cells and the interaction of T and B cells [38]. Previous studies have also found that IgD can significantly increase the levels of the inflammatory cytokines IL-1α, IL-1β, IL-6, IL-8, and TNF-α in culture supernatants of CD4+ T cells [39]. A high level of sIgD in RA serum was positively correlated with the levels of rheumatoid factor (RF), C-reactive protein (CRP), and soluble nuclear factor kappa B receptor activator ligand (sRANKL). These findings suggest that IgD may participate in the abnormal activation of T cells in patients with RA through IgDR. Lck mainly exists in T cells and participates in signal transduction involved in T cell development, differentiation, and activation. The phosphorylation of Lck can induce the activation of ZAP70. Subsequently, Lck and ZAP70 can further stimulate multiple signaling networks [40]. In this study, the results showed that IgD-Fc-Ig inhibited T cell proliferation stimulated by IgD, and IgD-Fc-Ig alone had no significant effect on T cell proliferation (data not shown), which further confirmed the hypothesis that IgD-Fc-Ig is a pseudoligand of IgDR. The results showed that p-Lck, p-ZAP70, p-P38, and p-NF-κB65 levels were obviously increased in CIA rats and in T cells stimulated by IgD. IgD-Fc-Ig and etanercept could obviously decrease the elevation of the expression of p-Lck, p-ZAP70, p-P38 and p-NF-κB65. The above results suggest that Lck may be a key signaling molecule involved in IgD activation of downstream signaling pathways. IgD-Fc-Ig selectively inhibits the activation and proliferation of T cells stimulated by IgD, and this may be related to its inhibition of the IgD-IgDR-Lck-NF-κB signaling pathway (Fig. 8).
Fig. 8. Schematic diagram of the IgD-IgDR-Lck signaling pathway and the regulation by IgD-Fc-Ig of the IgD-IgDR-Lck signaling pathway.

IgD binds to IgDR, which may be upregulated by Lck activity by binding to Lck and phosphorylating the Tyr394 site of Lck. Phosphorylated-Lck activates ZAP70 kinase and phosphorylates the Tyr319 site of ZAP70, further stimulating the activation of the NF-κB signaling pathway. IgD-Fc-Ig can competitively bind to IgDR, inhibit the expression of Lck and ZAP70, and inhibit the activation of the NF-ĸB signaling pathway even further, thereby regulating the abnormal proliferation, differentiation, and activation of T cells.
Conclusion
In conclusion, IgD-Fc-Ig has therapeutic effects on CIA rats by decreasing the paw volume, AI, and SJC and inhibiting T and B cell proliferation. IgD-Fc-Ig inhibited activated T cell function by downregulating the percentages of T cell subsets, decreasing inflammatory cytokine levels, and inhibiting the activation of the IgD-IgDR-Lck-NF-κB signaling pathway. The results of this study indicate that IgD-IgDR plays an important role in the occurrence and development of RA. Excessive sIgD activates the signaling pathway and promotes the activation of T cells by binding to IgDR on T cells. These findings suggest that IgD-Fc-Ig might be a potential immunomodulatory drug with anti-inflammatory effects. This provides a theoretical basis for further elucidating the occurrence and development of RA and provides experimental evidence for the development of IgD-Fc-Ig as a new targeted therapeutic drug for highly selective therapy of RA.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (U1803129, 81973332, and 81673444).
Author contributions
LH and XZZ. performed the study, wrote the paper, and analyzed the data. CW, XYT, YZ, XYC, YJW, JLS., Q.-t.W., J.-y.C., Y.C., and H.-x.W. contributed to the study. L.-l.Z. and W.W. designed the study, analyzed the data and wrote the paper.
Competing interests
The authors declare no competing interests.
Footnotes
These authors contributed equally: Le Han, Xian-zheng Zhang
Contributor Information
Ling-ling Zhang, Email: ll-zhang@hotmail.com.
Wei Wei, Email: wwei@ahmu.edu.cn.
References
- 1.Zhao W, Li D, Su Y, Zhao H, Pang W, Sun Y, et al. MicroRNA-147 negatively regulates expression of toll-like receptor-7 in rat macrophages and attenuates pristane induced rheumatoid arthritis in rats. Am J Transl Res. 2019;11:2219–31. [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 2.Klammt C, Novotna L, Li DT, Wolf M, Blount A, Zhang K, et al. T cell receptor dwell times control the kinase activity of Zap70. Nat Immunol. 2015;16:961–9. doi: 10.1038/ni.3231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gizinski AM, Fox DA. T cell subsets and their role in the pathogenesis of rheumatic disease. Curr Opin Rheumatol. 2014;26:204–10. doi: 10.1097/BOR.0000000000000036. [DOI] [PubMed] [Google Scholar]
- 4.Chen K, Xu W, Wilson M, He B, Miller NW, Bengten E, et al. Immunoglobulin D enhances immune surveillance by activating antimicrobial, proinflammatory and B cell-stimulating programs in basophils. Nat Immunol. 2009;10:889–98. doi: 10.1038/ni.1748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nguyen TG, Little CB, Yenson VM, Jackson CJ, McCracken SA, Warning J, et al. Anti-IgD antibody attenuates collagen-induced arthritis by selectively depleting mature B-cells and promoting immune tolerance. J Autoimmun. 2010;35:86–97. doi: 10.1016/j.jaut.2010.03.003. [DOI] [PubMed] [Google Scholar]
- 6.Choi JH, Wang KW, Zhang D, Zhan X, Wang T, Bu CH, et al. IgD class switching is initiated by microbiota and limited to mucosa-associated lymphoid tissue in mice. Proc Natl Acad Sci USA. 2017;114:E1196–204. doi: 10.1073/pnas.1621258114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wu YJ, Chen WS, Chen HS, Dai X, Dong J, Wang Y, et al. The immunoglobulin D Fc receptor expressed on fibroblast-like synoviocytes from patients with rheumatoid arthritis contributes to the cell activation. Acta Pharmacol Sin. 2017;38:1466–74. doi: 10.1038/aps.2017.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang P, Wei Z, Yan B, Huang T, Gou K, Dai Y, et al. Establishment of a transgenic mouse model with liver-specific expression of secretory immunoglobulin D. Sci China Life Sci. 2012;55:219–27. doi: 10.1007/s11427-012-4301-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kumar SP, Kashyap A, Silakari O. Exploration of the therapeutic aspects of Lck: a kinase target in inflammatory mediated pathological conditions. Biomed Pharmacother. 2018;108:1565–71. doi: 10.1016/j.biopha.2018.10.002. [DOI] [PubMed] [Google Scholar]
- 10.Feng S, Cheng X, Zhang L, Lu X, Chaudhary S, Teng R, et al. Myeloid-derived suppressor cells inhibit T cell activation through nitrating LCK in mouse cancers. Proc Natl Acad Sci USA. 2018;115:10094–99. doi: 10.1073/pnas.1800695115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lo WL, Shah NH, Ahsan N, Horkova V, Stepanek O, Salomon AR, et al. Lck promotes Zap70-dependent LAT phosphorylation by bridging Zap70 to LAT. Nat Immunol. 2018;19:733–41. doi: 10.1038/s41590-018-0131-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wu Y, Chen W, Chen H, Zhang L, Chang Y, Yan S, et al. The elevated secreted immunoglobulin D enhanced the activation of peripheral blood mononuclear cells in rheumatoid arthritis. PLoS ONE. 2016;11:e147788. doi: 10.1371/journal.pone.0147788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Setz CS, Khadour A, Renna V, Iype J, Gentner E, He X, et al. Pten controls B-cell responsiveness and germinal center reaction by regulating the expression of IgD BCR. EMBO J. 2019;38: e100249. doi: 10.15252/embj.2018100249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Xu B, Wang J, Zhang M, Wang P, Wei Z, Sun Y, et al. Expressional analysis of immunoglobulin D in cattle (Bos taurus), a large domesticated ungulate. PLoS ONE. 2012;7:e44719. doi: 10.1371/journal.pone.0044719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Vahedi S, Chueh FY, Chandran B, Yu CL. Lymphocyte-specific protein tyrosine kinase (Lck) interacts with CR6-interacting factor 1 (CRIF1) in mitochondria to repress oxidative phosphorylation. BMC Cancer. 2015;15:551. doi: 10.1186/s12885-015-1520-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhu Qingwen, Wang Jing, Zhang Lingli, Bian Wenjun, Lin Mengsi, Xu Xiaoning, Zhou Xiang. LCK rs10914542-G allele associates with type 1 diabetes in children via T cell hyporesponsiveness. Pediatric Research. 2019;86(3):311–315. doi: 10.1038/s41390-019-0436-2. [DOI] [PubMed] [Google Scholar]
- 17.Barrera-Vargas A, Gomez-Martin D, Alcocer-Varela J. T cell receptor-associated protein tyrosine kinases: the dynamics of tolerance regulation by phosphorylation and its role in systemic lupus erythematosus. Hum Immunol. 2014;75:945–52. doi: 10.1016/j.humimm.2014.08.207. [DOI] [PubMed] [Google Scholar]
- 18.Conigliaro P, Triggianese P, Perricone C, Chimenti MS, Di Muzio G, Ballanti E, et al. Restoration of peripheral blood natural killer and B cell levels in patients affected by rheumatoid and psoriatic arthritis during etanercept treatment. Clin Exp Immunol. 2014;177:234–43. doi: 10.1111/cei.12335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Daridon C, Burmester GR, Dorner T. Anticytokine therapy impacting on B cells in autoimmune diseases. Curr Opin Rheumatol. 2009;21:205–10. doi: 10.1097/BOR.0b013e32832a0760. [DOI] [PubMed] [Google Scholar]
- 20.Kobezda T, Ghassemi-Nejad S, Mikecz K, Glant TT, Szekanecz Z. Of mice and men: how animal models advance our understanding of T-cell function in RA. Nat Rev Rheumatol. 2014;10:160–70. doi: 10.1038/nrrheum.2013.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tanaka Y. Next stage of RA treatment: is TNF inhibitor-free remission a possible treatment goal? Ann Rheum Dis. 2013;72:i124–27. doi: 10.1136/annrheumdis-2012-202350. [DOI] [PubMed] [Google Scholar]
- 22.Wang QT, Wu YJ, Huang B, Ma YK, Song SS, Zhang LL, et al. Etanercept attenuates collagen-induced arthritis by modulating the association between BAFFR expression and the production of splenic memory B cells. Pharmacol Res. 2013;68:38–45. doi: 10.1016/j.phrs.2012.11.003. [DOI] [PubMed] [Google Scholar]
- 23.Wang D, Chang Y, Wu Y, Zhang L, Yan S, Xie G, et al. Therapeutic effects of TACI-Ig on rat with adjuvant arthritis. Clin Exp Immunol. 2011;163:225–34. doi: 10.1111/j.1365-2249.2010.04293.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang QT, Zhang LL, Wu HX, Wei W. The expression change of beta-arrestins in fibroblast-like synoviocytes from rats with collagen-induced arthritis and the effect of total glucosides of paeony. J Ethnopharmacol. 2011;133:511–6. doi: 10.1016/j.jep.2010.10.022. [DOI] [PubMed] [Google Scholar]
- 25.Zhang L, Li P, Song S, Liu Y, Wang Q, Chang Y, et al. Comparative efficacy of TACI-Ig with TNF-alpha inhibitor and methotrexate in DBA/1 mice with collagen-induced arthritis. Eur J Pharmacol. 2013;708:113–23. doi: 10.1016/j.ejphar.2013.02.025. [DOI] [PubMed] [Google Scholar]
- 26.Wang Q, Ma Y, Liu D, Zhang L, Wei W. The roles of B cells and their interactions with fibroblast-like synoviocytes in the pathogenesis of rheumatoid arthritis. Int Arch Allergy Immunol. 2011;155:205–11. doi: 10.1159/000321185. [DOI] [PubMed] [Google Scholar]
- 27.Shu J, Zhang F, Zhang L, Wei W. G protein coupled receptors signaling pathways implicate in inflammatory and immune response of rheumatoid arthritis. Inflamm Res. 2017;66:379–87. doi: 10.1007/s00011-016-1011-5. [DOI] [PubMed] [Google Scholar]
- 28.Chatzidionysiou K, Emamikia S, Nam J, Ramiro S, Smolen J, van der Heijde D, et al. Efficacy of glucocorticoids, conventional and targeted synthetic disease-modifying antirheumatic drugs: a systematic literature review informing the 2016 update of the EULAR recommendations for the management of rheumatoid arthritis. Ann Rheum Dis. 2017;76:1102–7. doi: 10.1136/annrheumdis-2016-210711. [DOI] [PubMed] [Google Scholar]
- 29.Jia XY, Chang Y, Sun XJ, Wu HX, Wang C, Xu HM, et al. Total glucosides of paeony inhibit the proliferation of fibroblast-like synoviocytes through the regulation of G proteins in rats with collagen-induced arthritis. Int Immunopharmacol. 2014;18:1–6. doi: 10.1016/j.intimp.2013.09.007. [DOI] [PubMed] [Google Scholar]
- 30.Zhu L, Wei W, Zheng YQ, Jia XY. Effects and mechanisms of total glucosides of paeony on joint damage in rat collagen-induced arthritis. Inflamm Res. 2005;54:211–20. doi: 10.1007/s00011-005-1345-x. [DOI] [PubMed] [Google Scholar]
- 31.Yu D, Linterman M. The temporospatial control of Tfh cells. Immunol Cell Biol. 2014;92:20–21. doi: 10.1038/icb.2013.86. [DOI] [PubMed] [Google Scholar]
- 32.Samuel RO, Ervolino E, de Azevedo QI, Azuma MM, Ferreira GT, Cintra L. Th1/Th2/Th17/Treg balance in apical periodontitis of normoglycemic and diabetic rats. J Endod. 2019;45:1009–15. doi: 10.1016/j.joen.2019.05.003. [DOI] [PubMed] [Google Scholar]
- 33.Jury EC, Kabouridis PS, Abba A, Mageed RA, Isenberg DA. Increased ubiquitination and reduced expression of LCK in T lymphocytes from patients with systemic lupus erythematosus. Arthritis Rheum. 2003;48:1343–54. doi: 10.1002/art.10978. [DOI] [PubMed] [Google Scholar]
- 34.Krishnan S, Nambiar MP, Warke VG, Fisher CU, Mitchell J, Delaney N, et al. Alterations in lipid raft composition and dynamics contribute to abnormal T cell responses in systemic lupus erythematosus. J Immunol. 2004;172:7821–31. doi: 10.4049/jimmunol.172.12.7821. [DOI] [PubMed] [Google Scholar]
- 35.Jelley-Gibbs DM, Strutt TM, McKinstry KK, Swain SL. Influencing the fates of CD4 T cells on the path to memory: lessons from influenza. Immunol Cell Biol. 2008;86:343–52. doi: 10.1038/icb.2008.13. [DOI] [PubMed] [Google Scholar]
- 36.Bowen H, Kelly A, Lee T, Lavender P. Control of cytokine gene transcription in Th1 and Th2 cells. Clin Exp Allergy. 2008;38:1422–31. doi: 10.1111/j.1365-2222.2008.03067.x. [DOI] [PubMed] [Google Scholar]
- 37.Monserrat J, Bohorquez C, Gomez LA, Movasat A, Perez A, Ruiz L, et al. The abnormal CD4(+)T lymphocyte subset distribution and Vbeta repertoire in new-onset rheumatoid arthritis can be modulated by methotrexate treament. Cells-Basel. 2019;8:871. doi: 10.3390/cells8080871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bucasas KL, Franco LM, Shaw CA, Bray MS, Wells JM, Nino D, et al. Early patterns of gene expression correlate with the humoral immune response to influenza vaccination in humans. J Infect Dis. 2011;203:921–29. doi: 10.1093/infdis/jiq156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wu YJ, Chen HS, Chen WS, Dong J, Dong XJ, Dai X, et al. CP-25 Attenuates the activation of CD4(+) T cells stimulated with immunoglobulin D in human. Front Pharmacol. 2018;9:4. doi: 10.3389/fphar.2018.00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bommhardt U, Schraven B, Simeoni L. Beyond TCR signaling: emerging functions of Lck in cancer and immunotherapy. Int J Mol Sci. 2019;20:3500. doi: 10.3390/ijms20143500. [DOI] [PMC free article] [PubMed] [Google Scholar]