

Abstract
We present a model-based method for inferring full-brain neural activity at millimeter-scale spatial resolutions and millisecond-scale temporal resolutions using standard human intracranial recordings. Our approach makes the simplifying assumptions that different people’s brains exhibit similar correlational structure, and that activity and correlation patterns vary smoothly over space. One can then ask, for an arbitrary individual’s brain: given recordings from a limited set of locations in that individual’s brain, along with the observed spatial correlations learned from other people’s recordings, how much can be inferred about ongoing activity at other locations throughout that individual’s brain? We show that our approach generalizes across people and tasks, thereby providing a person- and task-general means of inferring high spatiotemporal resolution full-brain neural dynamics from standard low-density intracranial recordings.
Keywords: electrocorticography (ECoG), intracranial electroencephalography (iEEG), local field potential (LFP), epilepsy, maximum likelihood estimation, Gaussian process regression
Introduction
Modern human brain recording techniques are fraught with compromize (Sejnowski et al. 2014). Commonly used approaches include functional magnetic resonance imaging (fMRI), scalp electroencephalography (EEG), and magnetoencephalography (MEG). For each of these techniques, neuroscientists and electrophysiologists must choose to optimize spatial resolution at the cost of temporal resolution (e.g., as in fMRI) or temporal resolution at the cost of spatial resolution (e.g., as in EEG and MEG). A less widely used approach (due to requiring work with neurosurgical patients) is to record from electrodes implanted directly onto the cortical surface (electrocorticography; ECoG) or into deep brain structures (intracranial EEG; iEEG). However, these intracranial approaches also require compromize: the high spatiotemporal resolution of intracranial recordings comes at the cost of substantially reduced brain coverage, since safety considerations limit the number of electrodes one may implant in a given patient’s brain. Furthermore, the locations of implanted electrodes are determined by clinical, rather than research, needs.
An increasingly popular approach is to improve the effective spatial resolution of MEG or scalp EEG data by using a geometric approach called “beamforming” to solve the biomagnetic or bioelectrical inverse problem (Sarvas 1987). This approach entails using detailed brain conductance models (often informed by high spatial resolution anatomical MRI images) along with the known sensor placements (localized precisely in 3D space) to reconstruct brain signals originating from theoretical point sources deep in the brain (and far from the sensors). Traditional beamforming approaches must overcome two obstacles. First, the inverse problem beamforming seeks to solve has infinitely many solutions. Researchers have made progress toward constraining the solution space by assuming that signal-generating sources are localized on a regularly spaced grid spanning the brain and that individual sources are small relative to their distances to the sensors (Snyder 1991; Baillet et al. 2001; Hillebrand et al. 2005). The second, and in some ways much more serious, obstacle is that the magnetic fields produced by external (noise) sources are substantially stronger than those produced by the neuronal changes being sought (i.e., at deep structures, as measured by sensors at the scalp). This means that obtaining adequate signal quality often requires averaging the measured responses over tens to hundreds of responses or trials (e.g., see review by Hillebrand et al. 2005).
Another approach to obtaining high spatiotemporal resolution neural data has been to collect fMRI and EEG data simultaneously. Simultaneous fMRI-EEG has the potential to balance the high spatial resolution of fMRI with the high temporal resolution of scalp EEG, thereby, in theory, providing the best of both worlds. In practice, however, the signal quality of both recordings suffers substantially when the two techniques are applied simultaneously (e.g., see review by Huster et al. 2012). In addition, the experimental designs that are ideally suited to each technique individually are somewhat at odds. For example, fMRI experiments often lock stimulus presentation events to the regularly spaced image acquisition time (TR), which maximizes the number of poststimulus samples. In contrast, EEG experiments typically employ jittered stimulus presentation times to maximize the experimentalist’s ability to distinguish electrical brain activity from external noise sources such as from 60 Hz alternating current power sources.
The current “gold standard” for precisely localizing signals and sampling at high temporal resolution is to take (ECoG or iEEG) recordings from implanted electrodes (but from a limited set of locations in any given brain). This begs the following question: what can we infer about the activity exhibited by the rest of a person’s brain, given what we learn from the limited intracranial recordings we have from their brain and additional recordings taken from other people’s brains? Here, we develop an approach, which we call “SuperEEG” (The term “SuperEEG” was coined by Robert J. Sawyer in his popular science fiction novel The Terminal Experiment (Sawyer 1995). SuperEEG is a fictional technology that measures ongoing neural activity throughout the entire living human brain at arbitrarily high spatiotemporal resolution.), based on Gaussian process regression (Rasmussen 2006). SuperEEG entails using data from multiple people to estimate activity patterns at arbitrary locations in each person’s brain (i.e., independent of their electrode placements). We test our SuperEEG approach using two large datasets of intracranial recordings (Sederberg et al. 2003, 2007a,b; Manning et al. 2011, 2012; Ezzyat et al. 2017, 2018; Horak et al. 2017; Kragel et al. 2017; Kucewicz et al. 2017, 2018; Lin et al. 2017; Solomon et al. 2018; Weidemann et al. 2019). We show that the SuperEEG algorithm recovers signals well from electrodes that were held out of the training dataset. We also examine the factors that influence how accurately activity may be estimated (recovered), which may have implications for electrode design and placement in neurosurgical applications.
Approach
The SuperEEG approach to inferring high temporal resolution full-brain activity patterns is outlined and summarized in Figure 1. We describe (in this section) and evaluate (in Results) our approach using two large previously collected datasets comprising multisession intracranial recordings. Dataset 1 comprises multisession recordings taken from 6876 electrodes implanted in the brains of 88 epilepsy patients (Sederberg et al. 2003, 2007a,b; Manning et al. 2011, 2012). Each recording session lasted from 0.2 to 3 h (total recording time: 0.3–14.2 h; Supplementary Fig. S6E). During each recording session, the patients participated in a free recall list learning task, which lasted for up to approximately 1 h. In addition, the recordings included “buffer” time (the length varied by patient) before and after each experimental session, during which the patients went about their regular hospital activities (confined to their hospital room, and primarily in bed). These additional activities included interactions with medical staff and family, watching television, reading, and other similar activities. For the purposes of the Dataset 1 analyses presented here, we aggregated all data across each recording session, including recordings taken during the main experimental task as well as during nonexperimental time. We used Dataset 1 to develop our main SuperEEG approach and to examine the extent to which SuperEEG might be able to generate task-general predictions. Dataset 2 comprised multisession recordings from 14 860 electrodes implanted in the brains of 131 epilepsy patients (Ezzyat et al. 2017, 2018; Horak et al. 2017; Kragel et al. 2017; Kucewicz et al. 2017, 2018; Lin et al. 2017; Solomon et al. 2018; Weidemann et al. 2019). Each recording session lasted from 0.4 to 2.2 h (total recording time: 0.4–6.6 h; Supplementary Fig. S6K). Whereas Dataset 1 included recordings taken as the patients participated in a variety of activities, Dataset 2 included recordings taken as each patient performed each of two specific experimental memory tasks: a random word list free recall task (Experiment 1) and a categorized word list free recall task (Experiment 2). We used Dataset 2 to further examine the ability of SuperEEG to generalize its predictions within versus across tasks. Supplementary Figure S6 provides additional information about both datasets.
Figure 1.

Methods overview. A. Electrode locations. Each dot reflects the location of a single electrode implanted in the brain of a Dataset 1 patient. A held-out recording location from one patient is indicated in red, and the patient’s remaining electrodes are indicated in black. The electrodes from the remaining patients are colored by k-means cluster (computed using the full-brain correlation model shown in Panel D). B. Radial basis function kernel. Each electrode contributed by the patient (black) weights on the full set of locations under consideration (all dots in Panel A, defined as 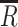 in the text). The weights fall off with positional distance (in MNI152 space) according to an RBF. C. Per-patient correlation matrices. After computing the pairwise correlations between the recordings from each patient’s electrodes, we use RBF-weighted averages to estimate correlations between all locations in
in the text). The weights fall off with positional distance (in MNI152 space) according to an RBF. C. Per-patient correlation matrices. After computing the pairwise correlations between the recordings from each patient’s electrodes, we use RBF-weighted averages to estimate correlations between all locations in  . We obtain an estimated full-brain correlation matrix using each patient’s data. D. Merged correlation model. We combine the per-patient correlation matrices (Panel C) to obtain a single full-brain correlation model that captures information contributed by every patient. Here, we have sorted the rows and columns to reflect k-means clustering labels (using k = 7; Yeo et al. 2011), whereby we grouped locations based on their correlations with the rest of the brain (i.e., rows of the matrix displayed in the panel). The boundaries denote the cluster groups. The rows and columns of Panel C have been sorted using the Panel D-derived cluster labels. E. Reconstructing activity throughout the brain. Given the observed recordings from the given patient (shown in black; held-out recording is shown in blue), along with a full-brain correlation model (Panel D), we use equation (12) to reconstruct the most probable activity at the held-out location (red).
. We obtain an estimated full-brain correlation matrix using each patient’s data. D. Merged correlation model. We combine the per-patient correlation matrices (Panel C) to obtain a single full-brain correlation model that captures information contributed by every patient. Here, we have sorted the rows and columns to reflect k-means clustering labels (using k = 7; Yeo et al. 2011), whereby we grouped locations based on their correlations with the rest of the brain (i.e., rows of the matrix displayed in the panel). The boundaries denote the cluster groups. The rows and columns of Panel C have been sorted using the Panel D-derived cluster labels. E. Reconstructing activity throughout the brain. Given the observed recordings from the given patient (shown in black; held-out recording is shown in blue), along with a full-brain correlation model (Panel D), we use equation (12) to reconstruct the most probable activity at the held-out location (red).
We first applied fourth-order Butterworth notch filters to remove 60 Hz (± 0.5 Hz) line noise from every recording (from every electrode). Next, we downsampled the recordings (regardless of the original samplerate) to 250 Hz. This downsampling step served to both normalize for differences in sampling rates across patients and to ease the computational burden of our subsequent analyses. We then excluded any electrodes that showed putative epileptiform activity. Specifically, we excluded, from further analysis, any electrode that exhibited a maximum kurtosis of 10 or greater across all of that patient’s recording sessions. We also excluded any patients with fewer than two electrodes that passed this criterion, as the SuperEEG algorithm requires measuring correlations between two or more electrodes from each patient. For Dataset 1, this yielded clean recordings from 4168 electrodes implanted throughout the brains of 67 patients (Fig. 1A, colored dots); for Dataset 2, this yielded clean recordings from 5023 electrodes implanted in 78 patients. Each individual patient contributed electrodes from a limited set of brain locations, which localized in a common space (MNI152; Grabner et al. 2006); an example, Dataset 1 patient’s 54 electrodes that survived the kurtosis thresholding procedure are highlighted in black and red (Fig. 1A).
The recording from a given electrode is maximally informative about the activity of the neural tissue immediately surrounding its recording surface. However, brain regions that are distant from the recording surface of the electrode also contribute to the recording, albeit (ceteris paribus) to a much lesser extent. One mechanism underlying these contributions is volume conduction. The precise rate of falloff due to volume conduction (i.e., how much a small volume of brain tissue at location x contributes to the recording from an electrode at location η) depends on the size of the recording surface, the electrode’s impedance, and the conductance profile of the volume of brain between x and η. As an approximation of this intuition, we place a Gaussian radial basis function (RBF) at the location η of each electrode’s recording surface (Fig. 1B). We use the values of the RBF at any brain location x as a rough estimate of how much structures around x contributed to the recording from location η
 |
(1) |
where the width variable λ is a parameter of the algorithm (which may in principle be set according to location-specific tissue conductance profiles) that governs the level of spatial smoothing. In choosing λ for the analyses presented here, we sought to maximize spatial resolution (which implies a small value of λ) while also maximizing the algorithm’s ability to generalize to any location throughout the brain, including those without dense electrode coverage (which implies a large value of λ). Here, we set λ = 20, guided in part by our prior related work (Manning et al. 2014, 2018) and in part by examining the brain coverage with non-zero weights achieved by placing RBFs at each electrode location in Dataset 1 and taking the sum (across all electrodes) at each voxel in a 4-mm3 MNI brain. (We then held λ fixed for our analyses of Dataset 2.) We note that this value could in theory be further optimized, e.g., using cross validation or a formal model (e.g., Manning et al. 2018).
A second mechanism whereby a given region x can contribute to the recording at η is through (direct or indirect) anatomical connections between structures near x and η. Although anatomical and functional correlations can differ markedly (e.g., Honey et al. 2009; Adachi et al. 2012; Goñi et al. 2014), we use temporal correlations in the data to estimate these anatomical connections (Becker et al. 2018). Let 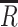 be the set of locations at which we wish to estimate local field potentials (LFPs), and let
be the set of locations at which we wish to estimate local field potentials (LFPs), and let 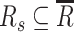 be the set of locations at which we observe LFPs from patient s (excluding the electrodes that did not pass the kurtosis test described above). In the analyses below, we define
be the set of locations at which we observe LFPs from patient s (excluding the electrodes that did not pass the kurtosis test described above). In the analyses below, we define  . We can calculate the expected interelectrode correlation matrix for patient s, where
. We can calculate the expected interelectrode correlation matrix for patient s, where  is the correlation between the time series of voltages for electrodes i and j from subject s during session k, using:
is the correlation between the time series of voltages for electrodes i and j from subject s during session k, using:
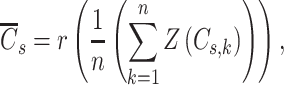 |
(2) |
where
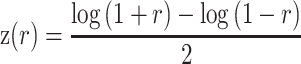 |
(3) |
is the Fisher z-transformation and
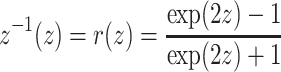 |
(4) |
is its inverse.
Next, we use equation (1) to construct a number of to-be-estimated locations by the number of patient electrode locations weight matrix, Ws. Specifically, Ws approximates how informative the recordings at each location in Rs is in reconstructing activity at each location in 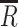 , where the contributions fall off with an RBF according to the distances between the corresponding locations:
, where the contributions fall off with an RBF according to the distances between the corresponding locations:
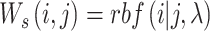 |
(5) |
Given this weight matrix, Ws, and the observed interelectrode correlation matrix for patient s,  , we can estimate the correlation matrix for all locations in
, we can estimate the correlation matrix for all locations in 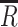 (
( ; Fig. 1C) using:
; Fig. 1C) using:
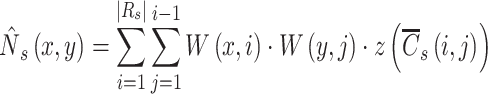 |
(6) |
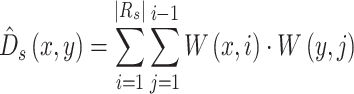 |
(7) |
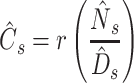 |
(8) |
After estimating the numerator ( ) and denominator (
) and denominator (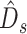 ) placeholders for each
) placeholders for each 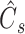 , we aggregate these estimates across the S patients to obtain a single expected full-brain correlation matrix (
, we aggregate these estimates across the S patients to obtain a single expected full-brain correlation matrix (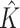 ; Fig. 1D)
; Fig. 1D)
 |
(9) |
.
Intuitively, the numerators capture the general structures of the patient-specific estimates of full-brain correlations, and the denominators account for which locations were near the implanted electrodes in each patient. To obtain 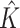 , we compute a weighted average across the estimated patient-specific full-brain correlation matrices, where patients with observed electrodes near a particular set of locations in
, we compute a weighted average across the estimated patient-specific full-brain correlation matrices, where patients with observed electrodes near a particular set of locations in 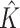 contribute more to the estimate.
contribute more to the estimate.
Having used the multipatient data to estimate a full-brain correlation matrix at the set of locations in 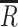 that we wish to know about, we next use
that we wish to know about, we next use  to estimate activity patterns everywhere in
to estimate activity patterns everywhere in 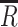 , given observations at only a subset of locations in
, given observations at only a subset of locations in 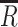 (Fig. 1E).
(Fig. 1E).
Let αs be the set of indices of patient s’s electrode locations in 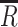 (i.e., the locations in Rs), and let βs be the set of indices of all other locations in
(i.e., the locations in Rs), and let βs be the set of indices of all other locations in 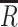 . In other words, βs reflects the locations in
. In other words, βs reflects the locations in 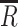 where we did not observe a recording for patient s (these are the recording locations we will want to fill in using SuperEEG). We can subdivide
where we did not observe a recording for patient s (these are the recording locations we will want to fill in using SuperEEG). We can subdivide 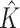 as follows:
as follows:
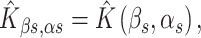 |
(10) |
and
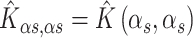 |
(11) |
Here, 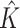 βs,αs represents the correlations between the “unknown” activity at the locations indexed by βs and the observed activity at the locations indexed by αs, and
βs,αs represents the correlations between the “unknown” activity at the locations indexed by βs and the observed activity at the locations indexed by αs, and 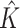 αs,αs represents the correlations between the observed recordings (at the locations indexed by αs).
αs,αs represents the correlations between the observed recordings (at the locations indexed by αs).
Let Ys,k,αs be the number-of-timepoints (T) by 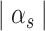 matrix of (observed) voltages from the electrodes in αs during session k from patient s. Then, we can estimate the voltage from patient s’s kth session at the locations in βs as follows (Rasmussen 2006):
matrix of (observed) voltages from the electrodes in αs during session k from patient s. Then, we can estimate the voltage from patient s’s kth session at the locations in βs as follows (Rasmussen 2006):
 |
(12) |
This equation is the foundation of the SuperEEG algorithm. Whereas we observe recordings only at the locations indexed by αs, equation (12) allows us to estimate the recordings at all locations indexed by βs, which can define a priori to include any locations we wish, throughout the brain. This yields the estimates of the time-varying voltages at every location in 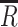 provided that we define
provided that we define 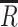 in advance to include the union of all of the locations in Rs and all of the locations at which we wish to estimate recordings (i.e., a timeseries of voltages).
in advance to include the union of all of the locations in Rs and all of the locations at which we wish to estimate recordings (i.e., a timeseries of voltages).
We designed our approach to be agnostic to electrode impedances, as electrodes that do not exist do not have impedances. Therefore, our algorithm recovers voltages in standard deviation (z-scored) units rather than attempting to recover absolute voltages. (This property reflects the fact that 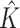 βs,αs and
βs,αs and 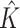 αs,αs are correlation matrices rather than covariance matrices.) Also, we note that equation (12) requires computing a T by T matrix, which can become computationally expensive when T is very large (e.g., for the Dataset 1 patient with the longest recording time, T = 12 786 750; also see Supplementary Fig. S6E and K). However, because equation (12) is time invariant, we may compute Ys,k,βs in a piecewise manner by filling in Ys,k,βs one row at a time (using the corresponding samples from Ys,k,αs).
αs,αs are correlation matrices rather than covariance matrices.) Also, we note that equation (12) requires computing a T by T matrix, which can become computationally expensive when T is very large (e.g., for the Dataset 1 patient with the longest recording time, T = 12 786 750; also see Supplementary Fig. S6E and K). However, because equation (12) is time invariant, we may compute Ys,k,βs in a piecewise manner by filling in Ys,k,βs one row at a time (using the corresponding samples from Ys,k,αs).
The SuperEEG algorithm described above and in Figure 1 allows us to estimate, up to a constant scaling factor, LFPs for each patient at all arbitrarily chosen locations in the set 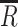 , even if we did not record that patient’s brain at all of those locations. We next turn to an evaluation of the accuracy of those estimates.
, even if we did not record that patient’s brain at all of those locations. We next turn to an evaluation of the accuracy of those estimates.
Results
We used a cross-validation approach to test the accuracy with which the SuperEEG algorithm reconstructs activity throughout the brain. For each patient in turn, we estimated full-brain correlation matrices (eq. 9) using data from all of the other patients. This step ensured that the data we were reconstructing could not also be used to estimate the between-location correlations that drove the reconstructions via equation (12) (otherwise the analysis would be circular). For that held-out patient, we held out each electrode in turn. We used equation (12) to reconstruct activity at the held-out electrode location, using the correlation matrix learned from all other patients’ data as 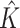 and using activity recorded from the other electrodes from the held-out patient as Ys,k,αs. (For analyses examining the stability of our estimates of
and using activity recorded from the other electrodes from the held-out patient as Ys,k,αs. (For analyses examining the stability of our estimates of  across time and patients, respectively, see Supplementary Figs S7 and S8). We then asked: how closely did each of the SuperEEG-estimated recordings at those electrodes match the observed recordings from those electrodes (i.e., how closely did the estimated
across time and patients, respectively, see Supplementary Figs S7 and S8). We then asked: how closely did each of the SuperEEG-estimated recordings at those electrodes match the observed recordings from those electrodes (i.e., how closely did the estimated 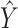 s,k,βs match the observed Ys,k,βs)?
s,k,βs match the observed Ys,k,βs)?
We used this general approach to quantify the algorithm’s performance across the full dataset. For each held-out electrode, from each held-out patient in turn, we computed the average correlation (across recording sessions) between the SuperEEG-reconstructed voltage traces and the observed voltage traces from that electrode. For this analysis, we set  to be the union of all electrode locations across all patients. This yielded a single correlation coefficient for each electrode location in
to be the union of all electrode locations across all patients. This yielded a single correlation coefficient for each electrode location in  , reflecting how well the SuperEEG algorithm was able to recover the recording at that location by incorporating data across patients (black histogram in Fig. 2A, map in Fig. 2C). The observed distribution of correlations was centered well above zero (mean: r = 0.51; t-test comparing mean of distribution of z-transformed average patient correlation coefficients to 0: t(66) = 23.55, P < 10−10), indicating that the SuperEEG algorithm recovers held-out activity patterns substantially better than random guessing.
, reflecting how well the SuperEEG algorithm was able to recover the recording at that location by incorporating data across patients (black histogram in Fig. 2A, map in Fig. 2C). The observed distribution of correlations was centered well above zero (mean: r = 0.51; t-test comparing mean of distribution of z-transformed average patient correlation coefficients to 0: t(66) = 23.55, P < 10−10), indicating that the SuperEEG algorithm recovers held-out activity patterns substantially better than random guessing.
Figure 2.
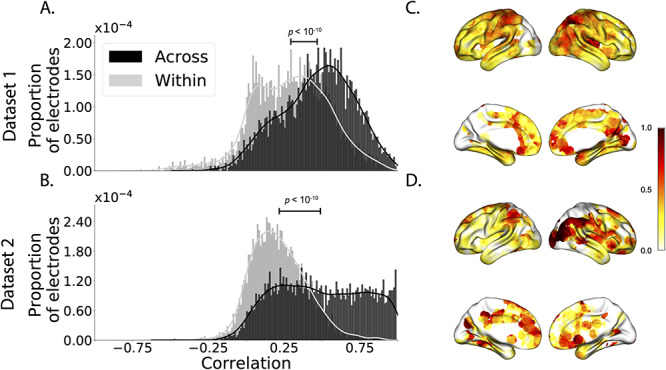
Reconstruction accuracy across all electrodes in two ECoG datasets. A. Distributions of correlations between observed versus reconstructed activity by electrode, for Dataset 1. The across-patient distribution (black) reflects reconstruction accuracy (correlation) using a correlation model learned from all but one patient’s data, and then applied to that held-out patient’s data. The within-patient distribution (gray) reflects performance using a correlation model learned from the same patient who contributed the to-be-reconstructed electrode. B. Distributions of correlations for Dataset 2. This panel is in the same format as Panel A but reflects results obtained from Dataset 2. The histograms aggregate data across both Dataset 2 experiments; for results broken down by experiment see Supplementary Figures S2 and S3. C, D. Reconstruction accuracy by location. The colors denote the average across-session correlations, using the across-patient correlation model, between the observed and reconstructed activity at the given electrode location projected to the cortical surface (Combrisson et al. 2019). Panel C displays the map for Dataset 1 and Panel D displays the map for Dataset 2.
Figure 3.
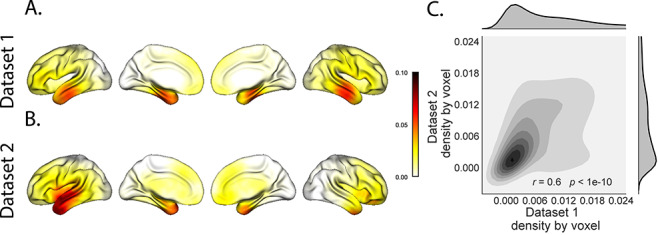
Electrode sampling density by location. A. Electrode sampling density by voxel location in Dataset 1. Each voxel is colored by the proportion of total electrodes in the dataset that are located within 20 MNI units of the given voxel. B. Electrode sampling density by voxel location in Dataset 2. This panel displays the sampling density map for Dataset 2, in the same format as Panel A. C. Correspondence in sampling density by voxel location across Datasets 1 and 2. The two-dimensional histogram displays the per-voxel sampling densities in the two Datasets, and the one-dimensional histograms display the proportions of voxels in each dataset with the given density value. The correlation reported in the panel is across voxels in the 4-mm3 MNI152 brain.
Next, we compared the quality of these across-participant reconstructions (i.e., computed using a correlation model learned from other patients’ data) to reconstructions generated using a correlation model trained using the in-patient’s data. In other words, for this within-patient benchmark analysis, we estimated 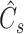 (eq. 8) for each patient in turn, using recordings from all of that patient’s electrodes except at the location we were reconstructing. These within-patient reconstructions serve as an estimate of how well data from all of the other electrodes from that single patient may be used to estimate held-out data from the same patient. This allows us to ask how much information about the activity at a given electrode might be inferred through 1) volume conductance or other sources of “leakage” from activity patterns measured from the patient’s other electrodes and 2) across-electrode correlations learned from that single patient. As shown in Figure 2A (gray histogram), the distribution of within-patient correlations was centered well above zero (mean: r = 0.32; t-test comparing mean of distribution of z- transformed average patient correlation coefficients to 0: t(66) = 15.16, P < 10−10). However, the across-patient correlations were substantially higher (t-test comparing average z-transformed within versus across patient electrode correlations: t(66) = 9.17, P < 10−10). This is an especially conservative test, given that the across-patient SuperEEG reconstructions exclude (from the correlation matrix estimates) all data from the patient whose data are being reconstructed. We repeated each of these analyses on a second independent dataset and found similar results (Fig. 2B, D; within vs across reconstruction accuracy: t(77) = 11.25, P < 10−10). We also replicated this result separately for each of the two experiments from Dataset 2 (Supplementary Fig. S3). This overall finding, which reconstructions of held-out data using correlation models learned from other patient’s data yield higher reconstruction accuracy than correlation models learned from the patient whose data are being reconstructed, has two important implications. First, it implies that distant electrodes provide additional predictive power to the data reconstructions beyond the information contained solely in nearby electrodes. This follows from the fact that each patient’s grid, strip and depth electrodes are implanted in a unique set of locations, so for any given electrode, the closest electrodes in the full dataset tends to come from the same patient. Second, it implies that the spatial correlations learned using the SuperEEG algorithm are, to some extent, similar across people.
(eq. 8) for each patient in turn, using recordings from all of that patient’s electrodes except at the location we were reconstructing. These within-patient reconstructions serve as an estimate of how well data from all of the other electrodes from that single patient may be used to estimate held-out data from the same patient. This allows us to ask how much information about the activity at a given electrode might be inferred through 1) volume conductance or other sources of “leakage” from activity patterns measured from the patient’s other electrodes and 2) across-electrode correlations learned from that single patient. As shown in Figure 2A (gray histogram), the distribution of within-patient correlations was centered well above zero (mean: r = 0.32; t-test comparing mean of distribution of z- transformed average patient correlation coefficients to 0: t(66) = 15.16, P < 10−10). However, the across-patient correlations were substantially higher (t-test comparing average z-transformed within versus across patient electrode correlations: t(66) = 9.17, P < 10−10). This is an especially conservative test, given that the across-patient SuperEEG reconstructions exclude (from the correlation matrix estimates) all data from the patient whose data are being reconstructed. We repeated each of these analyses on a second independent dataset and found similar results (Fig. 2B, D; within vs across reconstruction accuracy: t(77) = 11.25, P < 10−10). We also replicated this result separately for each of the two experiments from Dataset 2 (Supplementary Fig. S3). This overall finding, which reconstructions of held-out data using correlation models learned from other patient’s data yield higher reconstruction accuracy than correlation models learned from the patient whose data are being reconstructed, has two important implications. First, it implies that distant electrodes provide additional predictive power to the data reconstructions beyond the information contained solely in nearby electrodes. This follows from the fact that each patient’s grid, strip and depth electrodes are implanted in a unique set of locations, so for any given electrode, the closest electrodes in the full dataset tends to come from the same patient. Second, it implies that the spatial correlations learned using the SuperEEG algorithm are, to some extent, similar across people.
The recordings we analyzed from Dataset 1 comprised data collected as the patients performed a variety of (largely idiosyncratic) tasks throughout each day’s recording session. That we observed reliable reconstructions across patients suggests that the spatial correlations derived from the SuperEEG algorithm are, to some extent, similar across tasks. We tested this finding more directly using Dataset 2. In Dataset 2, the recordings were limited to times when each patient was participating in one of two experiments. Experiment 1 is a random-word list free recall task; Experiment 2 is a categorized list free recall task (24 patients participated in both). We wondered whether a correlation model learned from data from one experiment might yield good predictions of data from the other experiment. Furthermore, we wondered about the extent to which it might be beneficial or harmful to combine data across tasks.
To test the task specificity of the SuperEEG-derived correlation models, we restricted the dataset to the 24 patients that participated in both experiments and repeated the above within- and across-patient cross-validation procedures separately for Experiment 1 and Experiment 2 data from Dataset 2. We then compared the reconstruction accuracies for held-out electrodes, for models trained within versus across the two experiments, or combining across both experiments (Supplementary Fig. S1). In every case, we found that across-patient models trained using data from all other patients out-performed within-patient models trained on data only from the subject contributing the given electrode (ts(23) > 6.50, Ps < 10−5). All reconstruction accuracies also reliably exceeded chance performance (ts(23) > 8.00, Ps < 10−8). Average reconstruction accuracy was highest for the across-patient models limited to data from the same experiment (mean accuracy: r = 0.68); next-highest for the models that combined data across both experiments (mean accuracy: r = 0.61); and lowest for models trained across tasks (mean accuracy: r = 0.47). This pattern of results also held for each of the Dataset 2 experiments individually (Supplementary Fig. S2). Taken together, these results indicate that there are reliable commonalities in the spatial correlations of full-brain activity across tasks, but that there are also reliable differences in these spatial correlations across tasks. Whereas reconstruction accuracy benefits from incorporating data from other patients, reconstruction accuracy is highest when constrained to within-task data, or data that includes a variety of tasks (e.g., Dataset 1, or combining across the two Dataset 2 experiments).
Although both datasets we examined provide good full-brain coverage (when considering data from every patient), electrodes were not sampled uniformly throughout the brain. For example, in our patient population, electrodes are more likely to be implanted in regions such as the medial temporal lobe (MTL) and are rarely implanted in occipital cortex (Fig. S3A, B). Separately for each dataset, for each voxel in the 4-mm3 voxel MNI152 brain, we computed the proportion of electrodes in the dataset that were contained within a 20 MNI unit radius sphere centered on that voxel. We defined the “density” at that location as this proportion. Across Datasets 1 and 2, the electrode placement densities were similar (correlation by voxel: r = 0.6, P < 10−10). We wondered whether regions with good coverage might be associated with better reconstruction accuracy. For example, Figure 2C, D indicate that some electrodes in the MTL (which tends to be relatively densely sampled) have relatively high reconstruction accuracy, and occipital electrodes (which tend to be relatively sparsely sampled) tend to have relatively low reconstruction accuracy. To test whether this held more generally across the entire brain, for each dataset, we computed the electrode placement density for each electrode from each patient (using the proportion of other patients’ electrodes within 20 MNI units of the given electrode). We then correlated these density values with the across-patient reconstruction accuracies for each electrode. We found no reliable correlation between reconstruction accuracy and density for Dataset 1 (r = 0.05, P = 0.70) and a reliable negative correlation for Dataset 2 (r = −0.21, P = 0.05). This suggests that the reconstruction accuracies we observed are not driven solely by sampling density but rather may also reflect higher order properties of neural dynamics such as functional correlations between distant voxels (Betzel et al. 2017).
Prior work in humans and animals has shown that the spatial profile of the LFP differs by frequency band (e.g., with respect to volume conductance properties and contribution to the LFP; Fries et al. 2007; Crone et al. 2011; Buzsaki et al. 2012). For example, lower frequency components of the LFP tend to have higher power and extend further in space than high-frequency components (e.g., Miller et al. 2007; Manning et al. 2009). We wondered whether the reconstructions we observed might be differently weighting or considering the contributions of activity at different frequency bands. We therefore examined a range of frequency bands (δ: 2–4 Hz; θ: 4–8 Hz; α: 8–12 Hz; β: 12–30 Hz; γL: 30–60 Hz; and γH: 60–100 Hz), along with a measure of broadband (BB) power. We used second-order Butterworth bandpass filters to compute the activity patterns within each narrow frequency band. We defined BB power as the mean height of a linear robust regression fit in log-log space to the fourth-order Morelet wavelet-computed power spectrum at 50 log-spaced frequencies from 2 to 100 Hz (Manning et al. 2009). We then repeated our within-subject and across-subject cross-validated reconstruction accuracy tests (analogous to Fig. 2) separately for each frequency band (Fig. 4). (We also carried out a similar analysis on the Hilbert transform-computed spectral power within each narrow band; see Supplementary Fig. S4) Across both datasets, we found that our approach is best at reconstructing patterns of BB activity (right-most bars in Fig. 4A, D), a correlate of population firing rate (Manning et al. 2009). We also achieved good reconstruction accuracy within each narrow frequency band (Fig. 4 and Supplementary Fig. S4). Activity at lower frequencies (δ, θ, α, and β) tended to be reconstructed better than high-frequency patterns (γL and γH), with reconstruction accuracy peaking in the θ band. Overall, these results indicate that our approach is able to accurately recover information within the 2–100 Hz range.
Figure 4.

Reconstruction accuracy across all electrodes in two ECoG datasets for each frequency band. A. Distributions of correlations between observed versus reconstructed activity by electrode for each frequency band in Dataset 1. Each color denotes a different frequency band. Within each color group, the darker dots and bar on the left display the distribution (and mean) across-patient reconstruction accuracies (analogous to the black histograms in Fig. 2). The lighter dots and bar on the right display the distribution (and mean) within-patient reconstruction accuracies (analogous to the gray histograms in Fig. 2). Each dot indicates the reconstruction accuracy for one electrode in the dataset. To facilitate visual comparison with the frequency-specific results, the leftmost bars (gray) re-plot the histograms in Figure 2A. B. Statistical summary of across-patient reconstruction accuracy by electrode for each frequency band in Dataset 1. In the upper triangles of each map, warmer colors (positive t-values) indicate that the reconstruction accuracy for the frequency band in the given row was greater (via a two-tailed paired-sample t-test) than for the frequency band in the given column. Cooler colors (negative t-values) indicate that reconstruction accuracy for the frequency band in the given row was lower than for the frequency band in the given column. The lower triangles of each map denote the corresponding P-values for the t-tests. The diagonal entries display the average reconstruction accuracy within each frequency band. C. Statistical summary of within-patient reconstruction accuracy by electrode for each frequency band in Dataset 1. This panel displays the within-patient statistical summary, in the same format as Panel B. D. Distributions of correlations between observed versus reconstructed activity by electrode, for each frequency band in Dataset 2. This panel displays reconstruction accuracy distributions for each frequency band for Dataset 2. E, F. Statistical summaries of across-patient and within-patient reconstruction accuracy by electrode for each frequency band in Dataset 2. These panels are in the same format as Panels B and C, but display results from Dataset 2.
A basic assumption of our approach (and of most prior ECoG work) is that electrode recordings are most informative about the neural activity near the recording surface of the electrode. But if we consider that activity patterns throughout the brain are meaningfully correlated, are there particular implantation locations that, if recorded from a given patient’s brain, yield especially high reconstruction accuracies throughout the rest of their brain? For example, one might hypothesize that brain structures that are heavily interconnected with many other structures could be more informative about full-brain activity patterns than comparatively isolated structures. To test this hypothesis, we computed the average reconstruction accuracy across all of each patient’s electrodes (using our across-patients cross validation test; black histograms in Fig. 2A, B). We first labeled each patient’s electrodes, in each dataset, with the average reconstruction accuracy for that patient. In other words, we assigned every electrode from each patient the same value, reflecting how well the activity patterns for that patient were reconstructed. Next, for each voxel in the 4-mm3 MNI brain, we computed the average value across any electrode (from any patient) that came within 20 MNI units of that voxel’s center. This yielded an information score for each voxel, reflecting the (weighted) average reconstruction accuracy across any patients with electrodes near each voxel, where the averages were weighted to reflect patients who had more electrodes implanted near that location. We created a single map of these information scores for each dataset, highlighting regions that are especially informative about activity in other brain areas (Fig. 5A, B). Despite task and patient differences across the two datasets, we nonetheless found that the information score maps from both datasets were correlated (voxelwise correlation between information scores across the two datasets: r = 0.18, P < 10−10). Our finding that there were some commonalities between the two datasets’ information score maps lends support to the notion that different brain areas are (reliably) differently informative about full-brain activity patterns. We also examined the intersection between the top 10% most informative voxels across the two datasets (gray areas in Fig. 5C, networks shown in Fig. 6A, top row). Supporting the notion that structures that are highly interconnected with the rest of the brain are most informative about full-brain activity patterns, the intersecting set of voxels with the highest information scores included major portions of the dorsal attention network (e.g., inferior parietal lobule, precuneus, inferior temporal gyrus, thalamus, and striatum) as well as some portions of the default mode network (e.g., angular gyrus) that are highly interconnected with a large proportion of the brain’s gray matter (e.g., Tomasi and Volkow 2011).
Figure 5.

Most informative recording locations. A. Dataset 1 information scores by voxel. The voxel colors reflect the weighted average reconstruction accuracy across all electrodes from any patients with at least one electrode within 20 MNI units of the given voxel. B. Dataset 2 information scores by voxel. This panel is in the same format as Panel A. C. Intersection. Gray areas indicate the intersections between the top 10% most informative voxels in each map and projected onto the cortical surface (Combrisson et al. 2019). D. Correspondence in information scores by voxel across Datasets 1 and 2. The correlation reported in the Panel is between the per-voxel information scores across Datasets 1 and 2.
Figure 6.

Most informative recording locations by frequency band. A. Intersections between information score maps by frequency band. The regions indicated in each row depict the intersection between the top 10% most informative locations across Datasets 1 and 2. B. Network memberships of the most informative brain regions. The pie charts display the proportions of voxels in each region that belong to the seven networks identified by Yeo et al. (2011). The relative sizes of the charts for each frequency band reflect the average across-subject reconstruction accuracies (Fig. 4A, D). The voxels in Panel A are colored according to the same network memberships. C. Neurosynth terms associated with the most informative brain regions, by frequency band. The lists in each row display the top five neurosynth terms (Rubin et al. 2017) decoded for each region. D. Network parcellation map and legend. The parcellation defined by Yeo et al. (2011) is displayed on the inflated brain maps. The colors and network labels serve as a legend for Panels A and B. E. Combined map of the most informative brain regions. The map displays the union of the most informative maps in Panel A, colored by frequency band. The labels also serve as a legend for Panel C.
We also wondered whether the map of information scores might vary as a function of the spectral components of the activity patterns under consideration. We computed analogous maps of information scores for each individual frequency band. Across Datasets 1 and 2 (with the exception of α-band activity), we observed reliable positive correlations between the voxelwise maps of information scores (δ: r = 0.09, P < 10−57; θ: r = 0.24, P < 10−60; α: r = −0.03, P < 0.001; β: r = 0.02, P = 0.0011; γL: r = 0.1, P < 10−67; γH: r = 0.03, P < 10−7; BB: r = 0.21, P < 10−297).
To gain additional insight into which regions were most informative about full-brain activity patterns at different frequency bands, we next computed (for each frequency band) the intersection of the top 10% highest information scores across the maps for Datasets 1 and 2 (analogous to our approach in Fig. 5C). This yielded a single map of the (reliably) most informative locations, for each frequency band we examined. We then carried out post hoc analyses on each of these maps to characterize the underlying structural and functional properties of each set of regions we identified as being particularly informative about one or more types of neural pattern (Fig. 6 and Supplementary Fig. S5).
A growing body of neuroscientific research is concerned with characterizing the parcellations of anatomical and functional brain networks (for review see Zalesky et al. 2010; Arslan et al. 2018). The dominant approaches entail obtaining a full-brain connectivity matrix using either diffusion tensor imaging (DTI) to identify the brain’s network of white matter connections, or functional connectivity (typically applied to resting state data) to correlate the patterns of activity exhibited by different brain structures. One can then apply graph theoretic approaches to assign each brain structure (typically a single fMRI voxel) to one or more networks (for review see Bullmore and Sporns 2009). The result is a set of distinct (or partially overlapping) brain “networks” that may be further examined to elucidate their potential functional role. We over-laid a well-cited seven-network parcellation map identified by Yeo et al. (2011) onto the maps of brain locations that were most informative about each type of neural pattern. For each of these information maps, we computed the proportion of voxels in the most informative brain regions that belonged to each of the seven networks identified by Yeo et al. (2011); Figure 6D. We found that the regions we identified as being most informative about different neural patterns varied markedly with respect to which functional networks they belonged to (Fig. 6A, B).
The variability we observed in the frequency-specific information score maps is consistent with the notion that there is no “universal” brain region that reflects all types of activity patterns throughout the rest of the brain. Rather, each region’s activity patterns appear to be characterized by different spectral profiles, and the ability to infer full-brain activity patterns at a particular frequency band depends on the structural and functional connectome specific to that frequency band (Fig. 6E). We wondered how the maps we found might fit in with prior work. To this end, in addition to examining the anatomical profiles of each map, we used neurosynth (Rubin et al. 2017) to identify (using meta analyses of the neuroimaging literature) the top five most common terms associated with each frequency-specific map (Fig. 6C). We found that δ patterns across the brain were best predicted by regions of ventromedial prefrontal cortex, striatum, and thalamus (yellow). These regions are also implicated in modulating δ oscillations during sleep and are heavily interconnected with cortex (e.g., Amzica and Steriade 1998). The brain areas most informative about full-brain θ patterns were occipital and parietal regions associated with visual processing and visual attention (light green). Prior work has implicated θ oscillations in these areas in periodic sampling of visual attention (e.g., Busch and VanRullen 2010). We found that full-brain α patterns were best predicted by motor areas (dark green), which also exhibit α band changes during voluntary movements (e.g., Jurkiewicz et al. 2006). Striatum and thalamus (teal) were most informative about full-brain β patterns. Prior work has implicated striatal β activity in sensory and motor processing (Feingold et al. 2015), and thalamic β activity has been implicated in modulating widespread β patterns across neocortex (Sherman et al. 2016). Somatosensory areas (dark blue) were most informative about full-brain γL patterns. Prior work has implicated somatosensory γL in somatosensory processing and motor planning (Ihara et al. 2003). Occipital cortex (purple) was most informative about full-brain γH patterns. Occipital γH has also been linked with visual processing and reading (Wu et al. 2011) and the transmission of visual representations from low-order to high-order visual areas (Matsumoto et al. 2013). Full-brain BB patterns were best predicted by inferior parietal cortex precuneus (maroon). Functional neuroimaging BOLD responses (Simony et al. 2016) and BB ECoG patterns (Honey et al. 2012) in these default-mode hubs have been implicated in processing context-dependent representations that unfold over long timescales.
Discussion
Are our brain’s networks static or dynamic? And to what extent are the network properties of our brains stable across people and tasks? One body of work suggests that our brain’s functional networks are dynamic (e.g., Manning et al. 2018; Owen et al. 2019), person-specific (e.g., Finn et al. 2015), and task-specific (e.g., Turk-Browne 2013). In contrast, although the gross anatomical structure of our brains changes meaningfully over the course of years as our brains develop, on the timescales of typical neuroimaging experiments (i.e., hours to days), our anatomical networks are largely stable (e.g., Casey et al. 2000). Furthermore, many aspects of brain anatomy, including white matter structure, are largely preserved across people (e.g., Talairach and Tournoux 1988; Mori et al. 2008; Jahanshad et al. 2013). There are several possible means of reconciling this apparent inconsistency between dynamic person- and task-specific functional networks versus stable anatomical networks. For example, relatively small magnitude anatomical differences across people may be reflected in reliable functional connectivity differences. Along these lines, one recent study found that DTI structural data are similar across people but may be used to predict person-specific resting state functional connectivity data (Becker et al. 2018). Similarly, other work indicates that task-specific functional activations may be predicted by resting state functional connectivity data (Cole et al. 2016; Tavor et al. 2016). Another (potentially complementary) possibility is that our functional networks are constrained by anatomy but, nevertheless, exhibit (potentially rapid) task-dependent changes (e.g., Sporns and Betzel 2016). This prior work differs from ours in a number of ways. For example, fMRI data have substantially higher spatial resolution than (raw) ECoG data, and fMRI data have nearly complete spatial overlap across participants, whereas ECoG data have minimal spatial overlap across participants. Nevertheless, our work draws inspiration from those studies in that we also attempt to estimate held-out activity patterns across people and tasks.
Here, we have taken a model-based approach to studying whether high spatiotemporal resolution activity patterns throughout the human brain may be explained by a static connectome model that is shared across people and tasks. Specifically, we trained a model to take in recordings from a subset of brain locations and then predicted activity patterns during the same interval, but at other locations that were held out from the model. Our model, based on Gaussian process regression, was built on three general hypotheses about the nature of the correlational structure of neural activity (each of which we tested). First, we hypothesized that functional correlations are stable over time and across tasks. We found that, although aspects of the patients’ functional correlations were stable across tasks, we achieved better reconstruction accuracy when we trained the model on within-task data. This suggests that our general approach could be extended to better model across-task changes, e.g., following Cole et al. (2016); Tavor et al. (2016); and others. Second, we hypothesized that some of the correlational structure of people’s brain activity is similar across individuals. Consistent with this hypothesis, our model explained each patient’s data best when trained using data from other patients–even when compared models trained within-patient. Third, we resolved ambiguities in the data by hypothesizing that neural activity from nearby sources tends to be similar, all else being equal. This hypothesis was supported through our finding that all of the models we trained that incorporated this spatial smoothness assumption predicted held-out data well above chance.
Another important finding is that SuperEEG-based reconstructions accurately recover activity patterns at a broad range of frequencies (as well as BB patterns). However, brain networks differed in how informative they were about activity within each frequency band. Prior work has largely treated region-specific narrowband and BB activity as an indicator that activity at those frequency ranges reflects that the given region is representing or supporting a particular function. Our work suggests a complementary interpretation that when we observe a particular neural pattern in a particular brain region, it may instead (or in addition) reflect how that region is transmitting information to the rest of the brain via signaling at the given frequency range.
One potential limitation of our approach is that it does not provide a natural means of estimating the precise timing of single-neuron action potentials. Prior work has shown that gamma band and BB activity in the LFP may be used to estimate the firing rates of neurons that underlie the population contributing to the LFP (Miller et al. 2008; Manning et al. 2009; Jacobs et al. 2010; Crone et al. 2011). Because SuperEEG reconstructs LFPs throughout the brain, one could in principle use BB power in the reconstructed signals to estimate the corresponding firing rates (though not the timings of individual action potentials). We found that we were able to reconstruct full-brain patterns of BB power well (Fig. 4).
A second potential limitation of our approach is that it relies on ECoG data from epilepsy patients. Recent work comparing functional correlations in epilepsy patients (measured using ECoG) and healthy individuals (measured using fMRI) suggests that there are gross similarities between these populations (e.g., Kucyi et al. 2018; Reddy et al. 2018). Nevertheless, because all of the patients we examined have drug-resistant epilepsy, it remains uncertain how generally the findings reported here might apply more broadly to the population at large (e.g., nonclinical populations).
Beyond providing a means of estimating ongoing activity throughout the brain using already-implanted electrodes, our work also has implications how to optimize electrode placements in neurosurgical evaluations. Electrodes are typically implanted to maximize coverage of suspected epileptogenic tissue. However, our findings suggest that this approach might be improved upon. Specifically, one could leverage not only the noninvasive recordings taken during an initial monitoring period (as is currently done routinely) but also recordings collected from other patients. We could then ask: given what we learn from other patients’ data (and potentially from the scalp EEG recordings of this new patient), where should we place a fixed number of electrodes to maximize our ability to map seizure foci? As shown in Figures 5 and 6, and Supplementary Figure S5, recordings from different regions vary with respect to how informative they are about different narrowband and BB full-brain activity patterns.
By providing a means of reconstructing full-brain activity patterns, the SuperEEG approach maps ECoG recordings from different patients into a common neural space, despite those different patients’ electrodes were implanted in different locations. This feature of our approach enables across-patient ECoG studies, analogous to across-subject fMRI studies (e.g., Haxby et al. 2001, 2011; Norman et al. 2006). Whereas the focus of this manuscript is to specifically evaluate which aspects of neural activity patterns SuperEEG recovers well (or poorly), in parallel work, we are training across-patient classifiers by leveraging the common neural spaces obtained by applying SuperEEG to multipatient ECoG data. For example, we have shown that SuperEEG-derived activity patterns may be used to accurately predict psychiatric conditions such as depression (Scangos et al. 2020). Analogous approaches could in principle be used to develop improved brain–computer interfaces and/or to carry out other analyses that would benefit from high spatiotemporal resolution full-brain data in individuals, projected into a common ECoG space across people.
Concluding remarks
Over the past several decades, neuroscientists have begun to leverage the strikingly profound mathematical structure underlying the brain’s complexity to infer how our brains carry out computations to support our thoughts, actions, and physiological processes. Whereas traditional beamforming techniques rely on geometric source-localization of signals measured at the scalp, here, we propose an alternative approach that leverages the rich correlational structure of two large datasets of human intracranial recordings. In doing so, we are one step closer to observing, and perhaps someday understanding, the full spatiotemporal structure of human neural activity.
Code availability
We have published an open-source toolbox implementing the SuperEEG algorithm. It may be downloaded here. Additionally, we have provided code for all analyses and figures reported in the current manuscript, available here.
Data availability
The datasets analyzed in this study were generously shared by Michael J. Kahana. A portion of Dataset 1 may be downloaded here. Dataset 2 may be downloaded here.
Notes
We are grateful for useful discussions with Luke J. Chang, Uri Hasson, Josh Jacobs, Michael J. Kahana, and Matthijs van der Meer. We are also grateful to Michael J. Kahana for generously sharing the ECoG data we analyzed in our paper, which was collected under NIMH grant MH55687 and DARPA RAM Cooperative Agreement N66001-14-2-4-032, both to M.J.K. Our work was also supported in part by NSF EPSCoR Award Number 1632738 and by a sub-award of DARPA RAM Cooperative Agreement N66001-14-2-4-032 to J.R.M. The content is solely the responsibility of the authors and does not necessarily represent the official views of our supporting organizations.
Author Contributions
J.R.M conceived and initiated the project. L.L.W.O., T.A.M., and A.C.H. performed the analyses using software packages that all authors contributed to. J.R.M. and L.L.W.O. wrote the manuscript with input from all other authors.
Supplementary Material
References
- Adachi Y, Osada T, Sporns O, Watanabe T, Matsui T, Miyamoto K, Miyashita Y. 2012. Functional connectivity between anatomically unconnected areas is shaped by collective network-level effects in the macaque cortex. Cereb Cortex. 22:1586–1592. [DOI] [PubMed] [Google Scholar]
- Amzica F, Steriade M. 1998. Electrophysiological correlates of sleep delta waves. Electroencephalogr Clin Neurophysiol. 107:69–83. [DOI] [PubMed] [Google Scholar]
- Arslan S, Ktena SI, Makropoulos A, Robinson EC, Rueckert D, Parisot S. 2018. Human brain mapping: a systematic comparison of parcellation methods for the human cerebral cortex. NeuroImage. 170:5–30. [DOI] [PubMed] [Google Scholar]
- Baillet S, Mosher JC, Leahy RM. 2001. Electromagnetic brain mapping. IEEE Signal Process Mag. 18:14–30. [Google Scholar]
- Becker CO, Pequito S, Pappas GJ, Miller MB, Bassett DS, Grafton ST, Preciado VM. 2018. Spectral mapping of brain functional connectivity from diffusion imaging. Sci Rep. 8. doi: 10.1038/s41598-017-18769-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betzel RF, Medaglia JD, Kahn AE, Soffer J, Schonhaut DR, Bassett DS. 2017. Inter-regional ECoG correlations predicted by communication dynamics, geometry, and correlated gene expression. arXiv 170606088. [Google Scholar]
- Bullmore E, Sporns O. 2009. Complex brain networks: graph theoretical analysis of structural and functional systems. Nat Rev Neurosci. 10:186–198. [DOI] [PubMed] [Google Scholar]
- Busch NA, VanRullen R. 2010. Spontaneous EEG oscillations reveal periodic sampling of visual attention. Proc Nat Acad Sci USA. 107:16048–16053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buzsaki G, Anastassiou C, Koch C. 2012. The origin of extracellular fields and currents - eeg, ecog, lfp and spikes. Nat Rev Neurosci. 13:407–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Giedd JN, Thomas KM. 2000. Structural and functional brain development and its relation to cognitive development. Biol Psychol. 54:241–257. [DOI] [PubMed] [Google Scholar]
- Cole MW, Ito T, Bassett DS, Schultz DH. 2016. Activity flow over resting-state networks shapes cognitive task activations. Nat Neurosci. 19:1718–1726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Combrisson E, Vallat R, O’Reilly C, Jas M, Pascarella A, Saive A, Thiery T, Meunier D, Altukhov D, Lajnef T et al. 2019. Visbrain: a multi-purpose GPU-accelerated open- source suite for multimodal brain data visualization. Front Neuroinf. 13:1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crone NE, Korzeniewska A, Franaszczuk PJ. 2011. Cortical gamma responses: searching high and low. Int J Psychophysiol. 79:9–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ezzyat Y, Kragel JE, Burke JF, Levy DF, Lyalenko A, Wanda P, O’Sullivan L, Hurley KB, Busygin S, Pedisich I et al. 2017. Direct brain stimulation modulates encoding states and memory performance in humans. Curr Biol. 27:1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ezzyat Y, Wanda PA, Levy DF, Kadel A, Aka A, Pedisich I, Sperling MR, Sharan AD, Lega BC, Burks A et al. 2018. Closed-loop stimulation of temporal cortex rescues functional networks and improves memory. Nat Commun. 9. doi: 10.1038/s41467-017-02753-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feingold J, Gibson DJ, DePasquale B, Graybiel AM. 2015. Bursts of beta oscillation differntiate postperformance activity in the striatum and motor cortex of monkeys performing movement tasks. Proc Nat Acad Sci USA. 112:13687–13692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finn ES, Shen X, Scheinost D, Rosenberg MD, Huang J, Chun MM, Papademetris X, Constable RT. 2015. Functional connectome fingerprinting: identifying individuals using patterns of brain connectivity. Nat Neurosci. 18:1664–1671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fries P, Nikolić D, Singer W. 2007. The gamma cycle. Trends Neurosci. 30:309–316. [DOI] [PubMed] [Google Scholar]
- Goñi J, Heuvel MP, Avena-Koenigsberger A, Mendizabal NV, Betzel RF, Griffa A, Hagmann P, Corominas-Murtra B, Thiran J-P, Sporns O. 2014. Resting-brain functional connectivity predicted by analytic measures of network communication. Proc Natl Acad Sci. 111:833–838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabner G, Janke AL, Budge MM, Smith D, Pruessner J, Collins DL. 2006. Symmetric atlasing and model based segmentation: an application to the hippocampus in older adults. Med Image Comput Comput-Assistend Intervention. 9:58–66. [DOI] [PubMed] [Google Scholar]
- Haxby JV, Gobbini MI, Furey ML, Ishai A, Schouten JL, Pietrini P. 2001. Distributed and overlapping representations of faces and objects in ventral temporal cortex. Science. 293:2425–2430. [DOI] [PubMed] [Google Scholar]
- Haxby JV, Guntupalli JS, Connolly AC, Halchenko YO, Conroy BR, Gobbini MI, Hanke M, Ramadge PJ. 2011. A common, high-dimensional model of the representational space in human ventral temporal cortex. Neuron. 72:404–416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillebrand A, Singh KD, Holliday IE, Furlong PL, Barnes GR. 2005. A new approach to neuroimaging with magnetoencephalography. Hum Brain Mapp. 25:199–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honey CJ, Sporns O, Cammoun L, Gigandet X, Thiran JP, Meuli R, Hagmann P. 2009. Predicting human resting-state functional connectivity from structural connectivity. Proc Nat Acad Sci USA. 106:2035–2040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honey CJ, Thesen T, Donner TH, Silbert LJ, Carlson CE, Devinsky O, Doyle JC, Rubin N, Heeger DJ, Hasson U. 2012. Slow cortical dynamics and the accumulation of information over long timescales. Neuron. 76:423–434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horak PC, Meisenhelter S, Song Y, Testorf ME, Kahana MJ, Viles WD, Bujarski KA, Connolly AC, Robbins AA, Sperling MR et al. 2017. Interictal epileptiform discharges impair word recall in multiple brain areas. Epilepsia. 58:373–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huster RJ, Debener S, Eichele T, Herrmann CS. 2012. Methods for simultaneous EEG-fMRI: an introductory review. J Neurosci. 32:6053–6060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ihara A, Hirata M, Yanagihara K, Ninomiya H, Imai K, Ishii R, Osaki Y, Sakihara K, Izumi H, Imaoka H et al. 2003. Neuromagnetic gamma-band activity in the primary and secondary somatosensory areas. NeuroReport. 14:273–277. [DOI] [PubMed] [Google Scholar]
- Jacobs J, Manning JR, Kahana MJ. 2010. Response to Miller: “broadband” vs. “high gamma” electrocorticographic signals. J Neurosci. 30. [Google Scholar]
- Jahanshad N, Kochunov PV, Sprooten E, Mandl RC, Nichols TE, Almasy L, Blangero J, Brouwer RM, Curran JE, Zubicaray GI et al. 2013. Multi-site genetic analysis of diffusion images and voxelwise heritability analysis: a pilot project of the enigma–dti working group. NeuroImage. 81:455–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jurkiewicz MT, Gaetz WC, Bostan AC, Cheyne D. 2006. Post-movement beta rebound is generated in motor cortex: evidence from neuromagnetic recordings. NeuroImage. 32:1281–1289. [DOI] [PubMed] [Google Scholar]
- Kragel JE, Ezzyat Y, Sperling MR, Gorniak R, Worrell GA, Berry BM, Inman C, Lin J-J, Davis KA, Das SR et al. 2017. Similar patterns of neural activity predict memory formation during encoding and retrieval. NeuroImage. 155:70–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucewicz MT, Berry BM, Kremen V, Brinkmann BH, Sperling MR, Jobst BC, Gross RE, Lega B, Sheth SA, Stein JM et al. 2017. Dissecting gamma frequency actiivty during human memory processing. Brain. 140:1337–1350. [DOI] [PubMed] [Google Scholar]
- Kucewicz MT, Berry BM, Miller LR, Khadjevand F, Ezzyat Y, Stein JM, Kremen V, Brinkmann BH, Wanda P, Sperling MR et al. 2018. Evidence for verbal memory enhancement with electrical brain stimulation in the lateral temporal cortex. Brain. 141:971–978. [DOI] [PubMed] [Google Scholar]
- Kucyi A, Schrouff J, Bickel S, Foster BL, Shine JM, Parvizi J. 2018. Intracranial electrophysiology reveals reproducible intrinsic functional connectivity with human brain networks. J Neurosci. 38:4230–4242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin J-J, Rugg MD, Das S, Stein J, Rizzuto DS, Kahana MJ, Lega BC. 2017. Theta band power increases in the posterior hippocampus predict successful episodic memory encoding in humans. Hippocampus. 27:1040–1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning JR, Jacobs J, Fried I, Kahana MJ. 2009. Broadband shifts in LFP power spectra are correlated with single-neuron spiking in humans. J Neurosci. 29:13613–13620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning JR, Polyn SM, Baltuch G, Litt B, Kahana MJ. 2011. Oscillatory patterns in tem- poral lobe reveal context reinstatement during memory search. Proc Nat Acad Sci USA. 108:12893–12897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning JR, Ranganath R, Norman KA, Blei DM. 2014. Topographic factor analysis: a Bayesian model for inferring brain networks from neural data. PLoS One. 9:e94914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning JR, Sperling MR, Sharan A, Rosenberg EA, Kahana MJ. 2012. Spontaneously reactivated patterns in frontal and temporal lobe predict semantic clustering during memory search. J Neurosci. 32:8871–8878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning JR, Zhu X, Willke TL, Ranganath R, Stachenfeld K, Hasson U, Blei DM, Norman KA. 2018. A probabilistic approach to discovering dynamic full-brain functional connectivity patterns. NeuroImage. 180:243–252. [DOI] [PubMed] [Google Scholar]
- Matsumoto Joseph Y, Matt S, Kucewicz Michal T, Matsumoto Andrew J, Peters Pierce A, Brinkmann Benjamin H, Danstrom Jane C, Goerss Stephan J, Richard MW, Meyer Fred B et al. 2013. Network oscillations modulate interictal epileptiform spike rate during human memory. Brain. 136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller KJ, Leuthardt EC, Schalk G, Rao RPN, Anderson NR, Moran DW, Miller JW, Ojemann JG. 2007. Spectral changes in cortical surface potentials during motor movement. J Neurosci. 27:2424–2432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller KJ, Shenoy P, Nijs M, Sorensen LB, Rao RPN, Ojemann JG. 2008. Beyond the gamma band: the role of high-frequency features in movement classification. IEEE Trans Biomed Eng. 55:1634–1637. [DOI] [PubMed] [Google Scholar]
- Mori S, Oishi K, Jiang H, Jiang L, Li X, Akhter K, Hua K, Faria AV, Mahmood A, Woods R et al. 2008. Stereotaxic white matter atlas based on diffusion tensor imaging in an icbm template. NeuroImage. 40:570–582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norman KA, Newman E, Detre G, Polyn SM. 2006. How inhibitory oscillations can train neural networks and punish competitors. Neural Comput. 18:1577–1610. [DOI] [PubMed] [Google Scholar]
- Owen LLW, Chang TH, Manning JR. 2019. High-level cognition during story listen- ing is reflected in high-order dynamic correlations in neural activity patterns. bioRxiv. doi: 10.1101/763821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasmussen CE. 2006. Gaussian processes for machine learning. MIT Press. [Google Scholar]
- Reddy PG, Betzel RF, Khambhati AN, Shah P, Kini L, Litt B, Lucas TH, Davis KA, Bassett DS. 2018. Genetic and neuroanatomical support for functional brain network dynamics in epilepsy authors genetic and neuroanatomical support for functional brain network dynamics in epilepsy . arXiv. 1809.03934.
- Rubin TN, Kyoejo O, Gorgolewski KJ, Jones MN, Poldrack RA, Yarkoni T. 2017. Decoding brain activity using a large-scale probabilistic functional-anatomical atlas of human cognition. PLoS Comput Biol. 13:e1005649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarvas J. 1987. Basic mathematical and electromagnetic concepts of the biomagnetic inverse problem. Phys Med Biol. 32:11–22. [DOI] [PubMed] [Google Scholar]
- Sawyer Robert J. 1995. The Terminal Experiment, HarperPrism.
- Scangos KW, Khambhati AN, Daly PM, Owen LLW, Manning JR, Ambrose JB, Austin E, Dawes HE, Krystal AD, Chang EF. 2020. Biomarkers of depression symptoms defined by direct intracranial neurophysiology. bioRxiv. doi: 10.1101/2020.02.14.943118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sederberg PB, Kahana MJ, Howard MW, Donner EJ, Madsen JR. 2003. Theta and gamma oscillations during encoding predict subsequent recall. J Neurosci. 23:10809–10814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sederberg PB, Schulze-Bonhage A, Madsen JR, Bromfield EB, Litt B, Brandt A, Kahana MJ. 2007a. Gamma oscillations distinguish true from false memories. Psychol Sci. 18:927–932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sederberg PB, Schulze-Bonhage A, Madsen JR, Bromfield EB, McCarthy DC, Brandt A, Tully MS, Kahana MJ. 2007b. Hippocampal and neocortical gamma oscillations predict memory formation in humans. Cereb Cortex. 17:1190–1196. [DOI] [PubMed] [Google Scholar]
- Sejnowski TJ, Churchland PS, Movshon JA. 2014. Putting big data to good use in neuroscience. Nat Neurosci. 17:1440–1441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sherman MA, Lee S, Law R, Haegens S, Thorn CA, Hämäläinen MS, Moore CI, Jones SR. 2016. Neural mechanisms of transient neocortical beta rhythms: converging evidence from humans, computational modeling, monkeys, and mice. Proc Nat Acad Sci USA. 113:E4885–E4894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simony E, Honey CJ, Chen J, Hasson U. 2016. Uncovering stimulus-locked network dynamics during narrative comprehension. Nat Commun. 7:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snyder AZ. 1991. Dipole source localization in the study of EP generators: a critique. Electroencephalogr Clin Neurophysiol. 80:321–325. [DOI] [PubMed] [Google Scholar]
- Solomon EA, Gross R, Lega B, Sperling MR, Worrell G, Sheth SA, Zaghloul KA, Jobst BC, Stein JM, Das S et al. 2018. Mtl functional connectivity predicts stimulation-induced theta power. Nature Communications In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sporns O, Betzel RF. 2016. Modular brain networks. Annu Rev Psychol. 67:613–640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. 1988. Co-planar stereotaxic atlas of the human brain. Stuttgart: Verlag. [Google Scholar]
- Tavor I, Parker JO, Mars RB, Smith SM, Behrens TE, Jbabdi S. 2016. Task-free MRI predicts individual differences in brain activity during task performance. Science. 352:216–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomasi D, Volkow ND. 2011. Association between functional connectivity hubs and brain networks. Cereb Cortex. 21:2003–2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turk-Browne NB. 2013. Functional interactions as big data in the human brain. Science. 342:580–584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weidemann CT, Kragel JE, Lega BC, Worrell GA, Sperling MR, Sharan AD, Jobst BC, Khadjevand F, Davis KA, Wanda PA et al. 2019. Neural activity reveals interactions between episodic and semantic memory systems during retrieval. J Exp Psychol Gen. 148:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu HC, Nagasawa T, Brown EC, Juhasz C, Rothermel R, Hoechstetter K, Shah A, Mittal S, Fuerst D, Sood S et al. 2011. Gamma-oscillations modulated by picture naming and word reading: intracranial recording in epileptic patients. Clin Neurophysiol. 122:1929–1942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeo BTT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, Hollinshead M, Roffman JL, Smoller JW, Zollei L, Polimieni JR et al. 2011. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 106:1125–1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zalesky A, Fornito A, Harding IH, Cocchi L, Yücel M, Pantelis C, Bullmore ET. 2010. Whole- brain anatomical networks: does the choice of nodes matter? NeuroImage. 50:970–983. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets analyzed in this study were generously shared by Michael J. Kahana. A portion of Dataset 1 may be downloaded here. Dataset 2 may be downloaded here.
Notes
We are grateful for useful discussions with Luke J. Chang, Uri Hasson, Josh Jacobs, Michael J. Kahana, and Matthijs van der Meer. We are also grateful to Michael J. Kahana for generously sharing the ECoG data we analyzed in our paper, which was collected under NIMH grant MH55687 and DARPA RAM Cooperative Agreement N66001-14-2-4-032, both to M.J.K. Our work was also supported in part by NSF EPSCoR Award Number 1632738 and by a sub-award of DARPA RAM Cooperative Agreement N66001-14-2-4-032 to J.R.M. The content is solely the responsibility of the authors and does not necessarily represent the official views of our supporting organizations.