

Abstract
Over two decades, γ-secretase has been the target for extensive therapeutic development due to its pivotal role in pathogenesis of Alzheimer’s disease and cancer. However, it has proven to be a challenging task owing to its large set of substrates and our limited understanding of the enzyme’s structural and mechanistic features. The scientific community is taking bigger strides towards solving this puzzle with recent advancement in techniques like cryogenic electron microscopy (cryo-EM) and photo-affinity labelling (PAL). This review highlights the significance of the PAL technique with multiple examples of photo-probes developed from γ-secretase inhibitors and modulators. The binding of these probes into active and/or allosteric sites of the enzyme has provided crucial information on the γ-secretase complex and improved our mechanistic understanding of this protease. Combining the knowledge of function and regulation of γ-secretase will be a decisive factor in developing novel γ-secretase modulators and biological therapeutics.
Keywords: γ-Secretase, Photo affinity labeling, Inhibitor, Modulator, Mechanism
1. Alzheimer disease (AD) and γ-secretase
Alzheimer disease (AD) is a progressive and fatal neurodegenerative disease. Histopathologically, AD is characterized by the presence of extracellular plaques and intracellular neurofibrillary tangles in the brain. In accordance with the amyloid cascade hypothesis [1,2], β-amyloid peptide (Aβ) is the major constituent of plaques, and it’s believed to play a central role in the neuropathology of AD. Aβ is derived from the processing of the amyloid precursor protein (APP) by two proteases, β-secretase (BACE1) and γ-secretase, which generate the N and C termini of Aβ, respectively [3,4].
The γ-secretase complex is one of the most well studied aspartyl proteases belonging to a family of intramembrane proteases [5]. The enzyme complex is composed of four essential membrane proteins, namely presenilin (PS), nicastrin (Nct), anterior pharynx-defective 1 (Aph1), and presenilin enhancer 2 (Pen2) [6]. Although the presenilin (PS) subunit consists of catalytic site of the enzyme [7–10], the sequential assembly of all four subunits is known to be essential for an active γ-secretase complex [11–13].
The enzyme carries out proteolysis within the hydrophobic environment of the transmembrane domain leading to the release of an N-terminal fragment of the substrate into extracellular space and a C-terminal fragment into the cytoplasm [14]. To date, more than 90 substrates are reported to be processed by γ-secretase [15]. Two γ-secretase substrates – the amyloid precursor protein (APP) and Notch are extensively studied because of their implications in pathogenesis of Alzheimer’s disease and other human disorders. The involvement of γ-secretase in the disease pathogenesis has led to relentless efforts in drug development [4,16–18]. Nonetheless, γ-secretase has proven to be a rather elusive target [19] because of the large set of enzyme substrates, which lead to our limited knowledge of the structural features of the enzyme complex and undesired side-effects in clinical trials.
The advances in cryo-electron microscopy has recently led to two new structural reports of γ-secretase bound to APP and Notch substrates [20,21]. The substrate bound complex determined at 2.7 and 2.6 Å resolution provides great details about enzyme conformation and interactions with transmembrane domains (Fig.1). Long before these structural studies, chemical probes derived from γ-secretase inhibitors (GSIs) and modulators (GSMs) have been used to investigate γ-secretase and offered structural and mechanistic insights into this protease. Particularly, “Photo-affinity labelling” is a valuable and practical approach in such studies. Herein, we have reviewed recent advances in photo-affinity probes and their application in elucidating the function and regulation of γ-secretase.
Fig. 1.
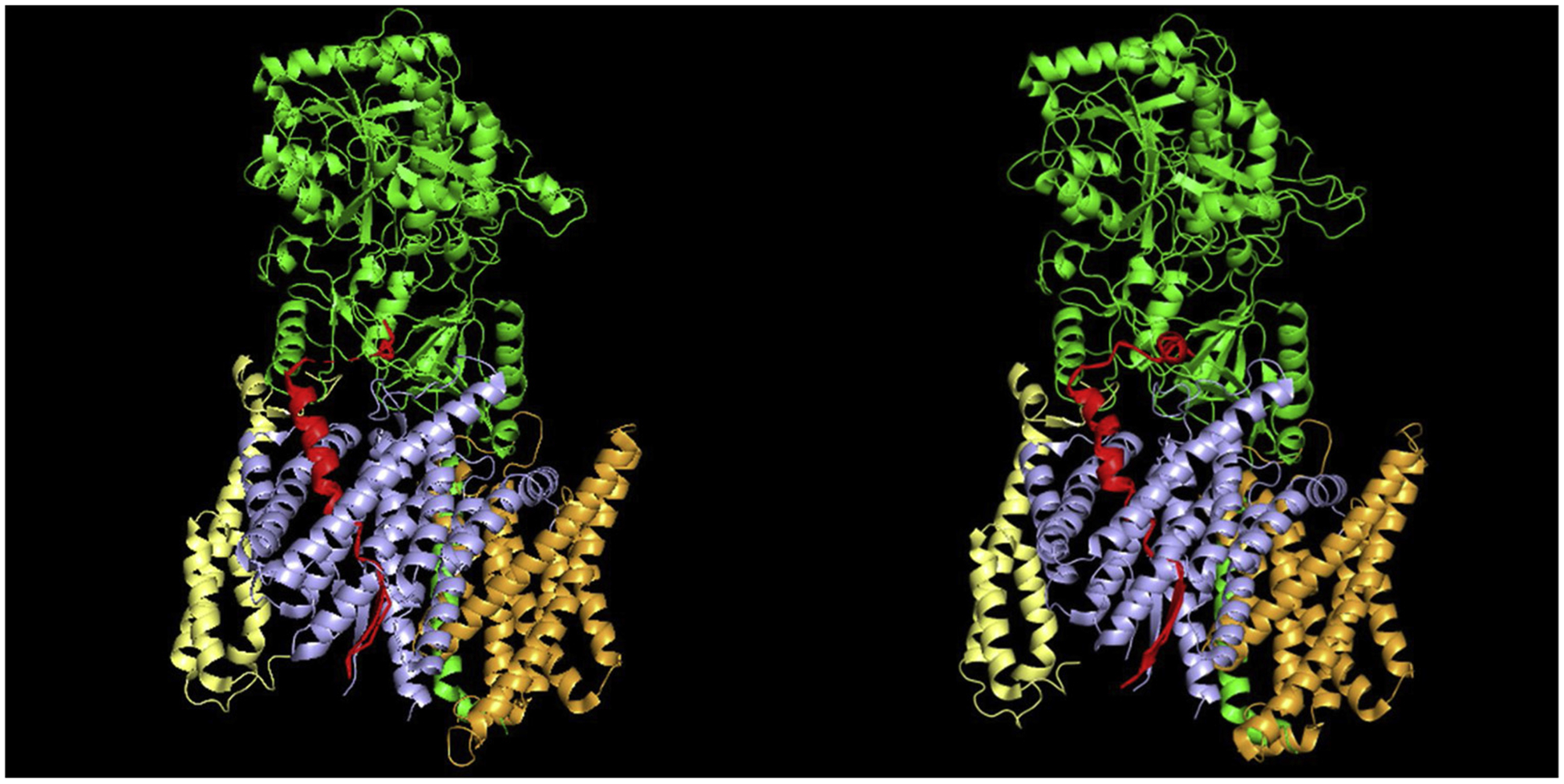
Cryo-EM structures of human γ-secretase in complex with APP-C83 (left, PDB bank ID 6IYC) and Notch-100 (right, PDB bank ID 6IDF). Color: APP-C83/Notch-100 in red, PS1 in light blue, Aph1 in bright orange, and Pen2 in pale yellow and Nicastrin in green.
2. Photo-affinity labeling (PAL) probes
Photo-affinity labeling has proven to be an effective tool in the field of chemical biology and pharmacology. The technique has come a long way since the first report of photo activated labelling of the enzyme chymotrypsin by Westheimer et al. [22] in 1962. At present, PAL is extensively used for target identification [23,24], structural elucidation of binding site [25], analysis of ligand-receptor interaction [26,27], and protein labelling and monitoring [28].
2.1. Photophores
The chemical entity responsible for PAL is called the “Photo-affinity probe”. A photo-affinity probe generally consists of three components, namely
A ligand, which could be a native substrate or a lead molecule. Most of the time these ligands are target specific with high affinity.
A photoactive group which upon irradiation leads to highly reactive chemical species that covalently reacts with the target and
A reporter or identification tag that can be used for monitoring and isolation of desired target.
The activation of a photoprobe should be mild such that the photoirradiation will cause minimal disruption of the biological system. The photoprobe should be stable in the dark or ambient light and a wide range of pHs. Modification of the ligand with these components should not affect the parent ligand’s activity. The use of a spacer between ligand and the other two components combined with the choice of components significantly affect the outcome. The synthetic ease with which these photoactive groups can be incorporated also dictates the selection of the photoactive group. The steric bulk of the photo-probe, modification site on the parent ligand, half-life of reactive intermediate, possible non-specific labelling, and physio-chemical properties of the probe are some of the factors that need to be considered while designing an optimal photo-affinity probe.
The process of optimizing a photo-probe resembles a hit to lead drug development. Several photo-affinity probes have been reported in the last 50 years with different synthetic design, photoactive groups and reporter tags. Nonetheless, it is a considerably difficult task to assemble all the features mentioned earlier in a single molecule. As a result, there is “No Ideal” photo-probe design to date which can be utilized efficiently with different pharmacophores for PAL.
The three most common photoactive groups are benzophenone, aryl azides, and diazirines (Fig. 2). Benzophenones are activated using a light of wavelength around 360 nm. The irradiation leads to a reactive diradical carbonyl species which can add to any XeH bond where X can be a C, N, or O atom. The stability of benzophenones in most organic solvents, their ease of preparation, orthogonality with other chemical transformations and mild photo-activation are some of the factors responsible for their extensive use in PAL. However, benzophenone activation requires longer irradiation time which could lead to non-specific interactions. Another disadvantage of benzophenones is their bulkiness which can sterically hinder the binding of a ligand with a desired target.
Fig. 2.
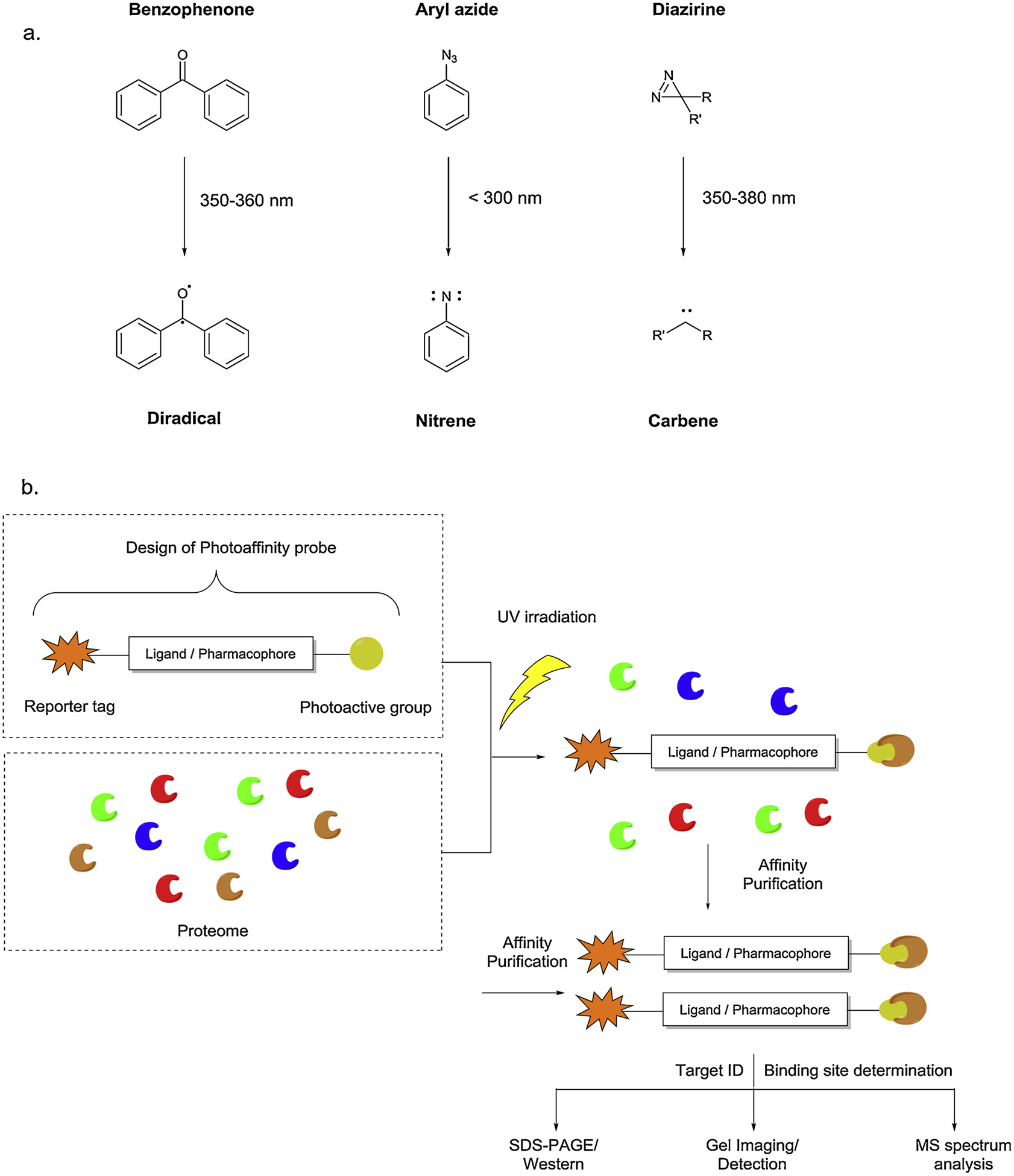
a. Activation mechanism of photoactive groups and b. schematic illustration of PAL.
Aryl azides are activated using shorter wavelength of light (< 300 nm) which leads to reactive singlet nitrene species for binding. The aryl azides can be easily synthesized and incorporated into a probe design. The smaller size of the photo-active group is the major advantage over benzophenone-based probes. But the shorter wavelength of activation can cause damage to the other biomolecules. The nitrenes are also reported to form undesired side products such as benzazirine and ketenimines [29] resulting in the lower labelling efficiency. Substituted aryl azides such as tetrafluorophenylazide are developed to improve the stability of these probes. Tetrafluorophenylazides have shown to prevent the formation of undesired ketenimines [30]. It is also observed that ortho-substituted phenylazides should be avoided since they are prone to intra-molecular cyclization leading to benzazirine intermediate followed by a ring expansion [31].
Aliphatic diazirines are most frequently used photo-active group among the three because of their smaller size. Both aliphatic and aryl diazirines have been used in PAL which are photo irradiated using higher wavelength of light around 360 nm. The three membered ring system containing one carbon atom and two nitrogen atoms linked with a double bond upon irradiation results in highly reactive carbene species with the release of N2 gas. The carbenes are more reactive and can undergo a range of reactions with C–H or XeH bonds where X is any heteroatom. The shorter half-life of reactive carbene intermediates reduces the non-specific interactions but also effects the overall labeling yields. The in situ generated carbenes are also quenched by water reducing the labeling yields further. The final major disadvantage of diazirines is the comparatively difficult synthetic route to afford the desired probe. The early methods of synthesizing aliphatic and substituted aromatic diazirines along with the recent advances in the synthetic strategies are reviewed by Robertson et al. [32].
2.2. Reporter/Identification tag
The final component is the reporter tag, which is required to successfully monitor and isolate the photo-labelled target. Radioactive isotopes such as 19F, 125I and 3H and variety of fluorophores like rhodamine, BODIPY, and fluorescein are a common choice of reporter tags to measure and monitor the desired target [33,34]. However, with the advent of click chemistry “alkyne handles” have become the primary choice of reporter system for isolation of photo-labeled targets [35,36]. The alkyne handle is reacted with an azide terminated affinity tag such as biotin using Cu-catalyzed azide-alkyne cycloaddition. The biotin linked targets can be isolated using avidin beads which have a high affinity to biotin.
3. γ-Secretase inhibitor (GSIs)
3.1. GSIs in AD clinical trials
In order to suppress the γ-secretase activity for AD and cancer therapy, a variety of GSIs have been developed [4,37–40] and later advanced to Phase II and III clinical trials. Among them are Semagacestat and Avagacestat (BMS-708,163), the clinical studies of which have been reported [41,42]. The most surprising outcome of the clinical trial of Semagacestat was the worsening of memory in patients [41]. The other major adverse effect of the trial was the increased risk of a skin cancer, which likely resulted from inhibition of Notch signaling [43,44]. While the Notch-associated side effects are somewhat understood, the mechanism of cognitive decline is elusive. It may be informative to compare clinical studies of Avagacestat [42]. The major adverse effects included predominantly gastrointestinal and dermatologic complications resulting from probable Notch inhibition. The 100 mg and 125 mg dose arms also led to a trend for cognitive worsening [42] suggesting that both compounds may share a common mechanism of toxicity. While Avagacestat was reported as a Notch sparing inhibitor at the first place [45], we and others were unable to reproduce this result [46,47]. Therefore, these studies, from compound selections to clinical design, have been questioned [19,46,47]. Moreover, non-selective inhibition of γ-secretase drastically affects the processing and metabolism of APP proteins, which have been shown to regulate various neuronal and synaptic functions conferred by distinct APP domains [48,49]. The accumulation of APP βCTF that results from γ-secretase inhibition has been implicated in neurotoxicity [50]. Also, it has been shown that low concentrations of GSIs can cause Aβ elevation and withdrawing of GSIs leads to a rebound of Aβ production in the plasma [51]. In addition, an increase in βCTF can augment the Aβ42/Aβ40 ratio [52]. Together the data suggest that total inhibition of APP processing could actually aggravate AD pathology and lead to worsening cognition. Tagami et al. [53] recently reported a novel mechanism indicating that non-transition state analog GSIs like Semagacestat are pseudo-inhibitors, which inhibited secretion of intramembranous γ-byproducts to the soluble space and increased peptide byproducts and long Aβ species in the cells. These accumulated intracellular Aβ species could be toxic and cause the aggravation of cognitive decline observed in the clinical trials.
Although none of the GSIs achieved clinically viable therapy so far, utilization of these molecules as chemical tool probes facilitates our understanding of the regulatory mechanism of γ-secretase. Several representative categories of photo affinity probes with the emphasis on their chemical design and biological application are summarized here.
3.2. GSI-based probes
3.2.1. Active site directed transition state probes
The earliest generation of GSIs were transition state inhibitors that targeted the active site of the aspartic protease, such as L-685,458 (1, Fig. 3) and III-31-C (8, Fig. 4). These analogues utilize a hydroxyethylene dipeptide isostere (Figs. 3 & 4, labeled in blue color) that directs the compound into the active site without hydrolyzation. These active site-directed GSIs only bind to the active form of aspartyl protease, but not the inactive one. The discovery of L-685,458 as a γ-secretase inhibitor [54] and development of L-685,458 based photo-probes (L-852,505 and L852,646) [8] (2–3, Fig. 3) have provided a critical insight into the identity of γ-secretase. Mutagenesis of the two conserved aspartate residues in the transmembrane regions of PS1 significantly reduces γ-secretase activity [7], suggesting that PS1 is either an essential diaspartyl cofactor or is itself γ-secretase. Direct labeling of PS1 by L-852,505 and L-852,646 has provided compelling evidence that the catalytic active site of γ-secretase is located at the interface of two PS1 subunits. In a more recent study [55], Crump et al. reported two clickable photoaffinity probes based on both L-852,505 and L-852,646, which allowed conjugation with any types of azide-linked reporters for affinity pulldown and fluorescent visualization. By this approach, the probes efficiently labeled PS1-NTF in protease complex, as well as a related aspartyl intramembrane protease signal peptide peptidase (SPP).
Fig. 3.
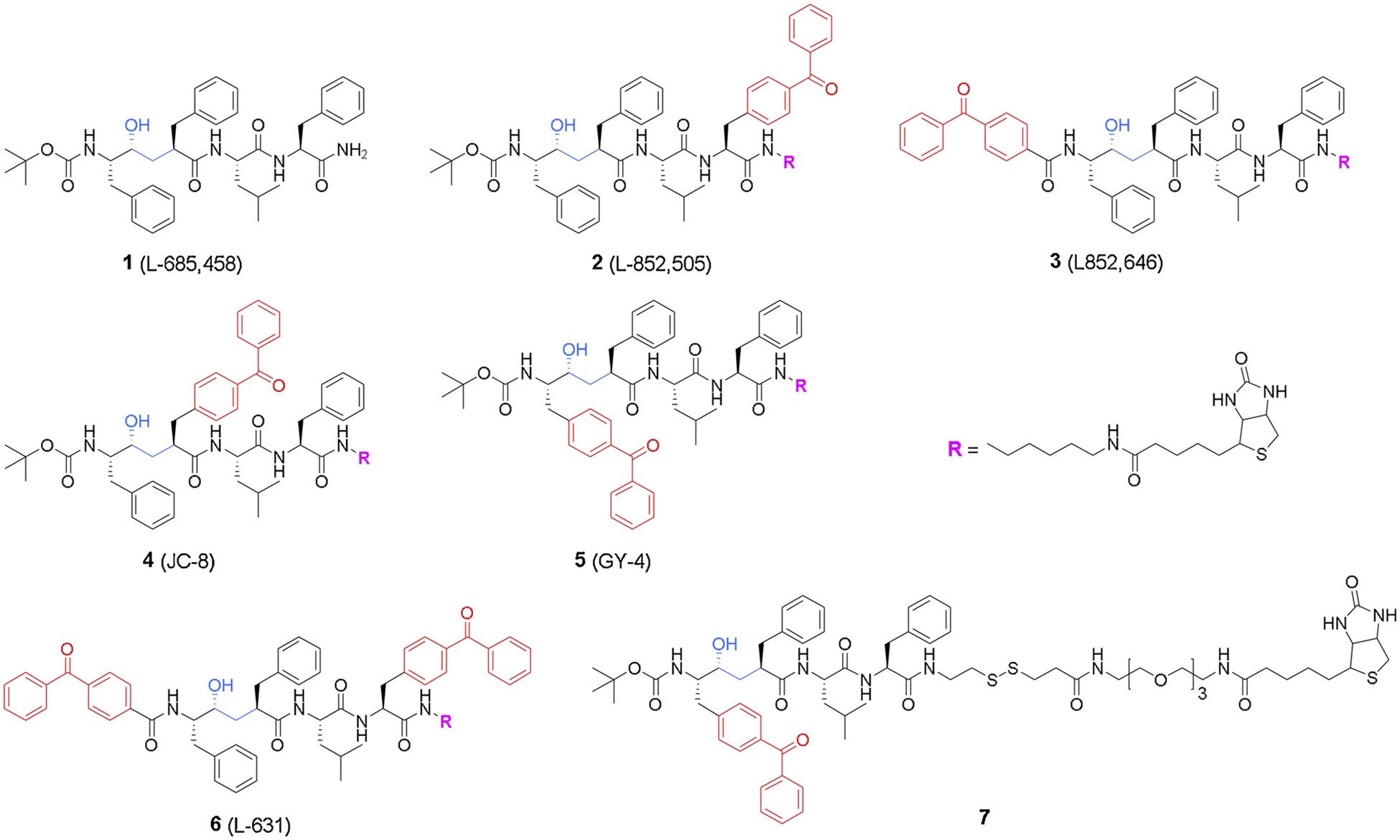
L-685,458 based photo affinity probes.
Fig. 4.
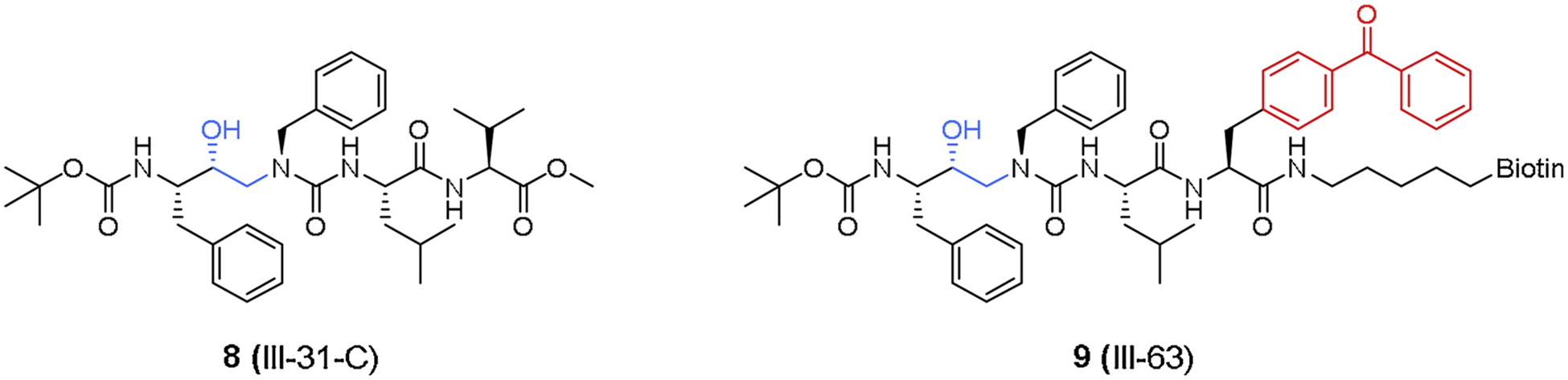
III-31-C based photo affinity probes.
Since then, many scaffolds of transition state inhibitors have been rapidly studied and developed as molecular probes due to their high potency and specificity. Benzophenone as a photoaffinity group is generally thought to be bulky with large steric hindrance, but it has been remarkably successful in case of the transition state inhibitor analogues.
Wolfe and colleagues also developed a similar photoaffinity probe III-63 (9, Fig. 4) [56–58] based on transition state inhibitor III-31-C (8, Fig. 4), which was used to characterize numerous structurally diverse GSIs by competitive labeling studies [57]. As a result, the GSIs were classified into different mechanistic categories – directly binding to active site, altering active site by allosteric binding or blocking protease activity without affecting active site. The III-63 was later used in a subsequent study [58] to identify an initial substrate-binding site (docking site) of the γ-secretase, which recognizes the peptide substrate before it enters the active site.
Transition state probes are not only utilized to characterize presenilin, but also represent valuable tools to study γ-secretase interacting proteins [59,60]. Studies using the activity-based photo probe confirmed that only a small portion (~14 %) of PS1 is engaged in active γ-secretase complex [61]. Meanwhile, other studies also proved that only a small fraction of γ-secretase complexes are catalytically active [59,62,63]. An early example showed that γ-secretase subunits Pen2, Nct, and Aph1 could be captured in company with presenilin by active site directed probes (7, Fig. 3), which were further used to characterize stoichiometry and dynamics of γ-secretase [59]. For instance, over-expression of Pen2 or PS1 mutants modulates the equilibrium of PS1 and PS2 in γ-secretase complex, resulting in an elevation in the Aβ42:Aβ40 ratio. Using the same probe (7, Fig. 3), a more recent study successfully pulled down hypoxia-inducible factor-1α (Hif-1α) together with γ-secretase complex [60], which proved the direct association of Hif-1α with the active enzyme complex and thus demonstrated a novel regulatory mechanism of γ-secretase under hypoxia condition. Notably, these studies also suggested that a minimal linker length (> 34 Å) for the affinity probe is required to capture the native γ-secretase complex emphasizing the crucial role of the linker during the probe design.
3.2.2. DAPT-derived and sulfonamide-based probes
As transition state inhibitors are generally peptidomimetics with poor pharmacokinetics, a lot of efforts have been taken to develop new classes of GSIs for clinical use [38,39]. N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) scaffold (10, Fig. 5) was identified [64] with an allosteric binding site that is distinct from the active site. Morohashi et al. reported the first DAPT-based probe DAPBpB [65] (11, Fig. 5), which consists a benzophenone moiety connecting DAPT and biotin group for affinity-based pulldown. The probe helped to define a novel function domain on PS-CTF as a DAPT binding site, which is distinct from either the active site or the initial substrate binding site. DAPT derivatives were therefore believed to block the substrate migration from the substrate binding site to the catalytic active site. A follow up study from the same group reported conversion of two DAPT derivatives, Compound E (12, Fig. 5) and LY411,575 analogue (DBZ, 15, Fig. 5), into photoreactive probes through divergent synthesis [66]. Different types of photoreactive groups and linkers were incorporated into the probes (13–14, 16–17, Fig. 5) to compare their labeling efficiency. Interestingly, both Compound E and DBZ based probes cross-linked to PS-NTF, and not CTF like DAPT. This result suggests that the binding site of DAPT derivatives is at the interface of the heterodimer. The labeling selectivity likely depends on distance or orientation of the probe to a specific residue around the binding site. The results also showed that only benzophenone containing probes labeled PS, and hydrophobic linkers are preferred compared with hydrophilic linkers.
Fig. 5.
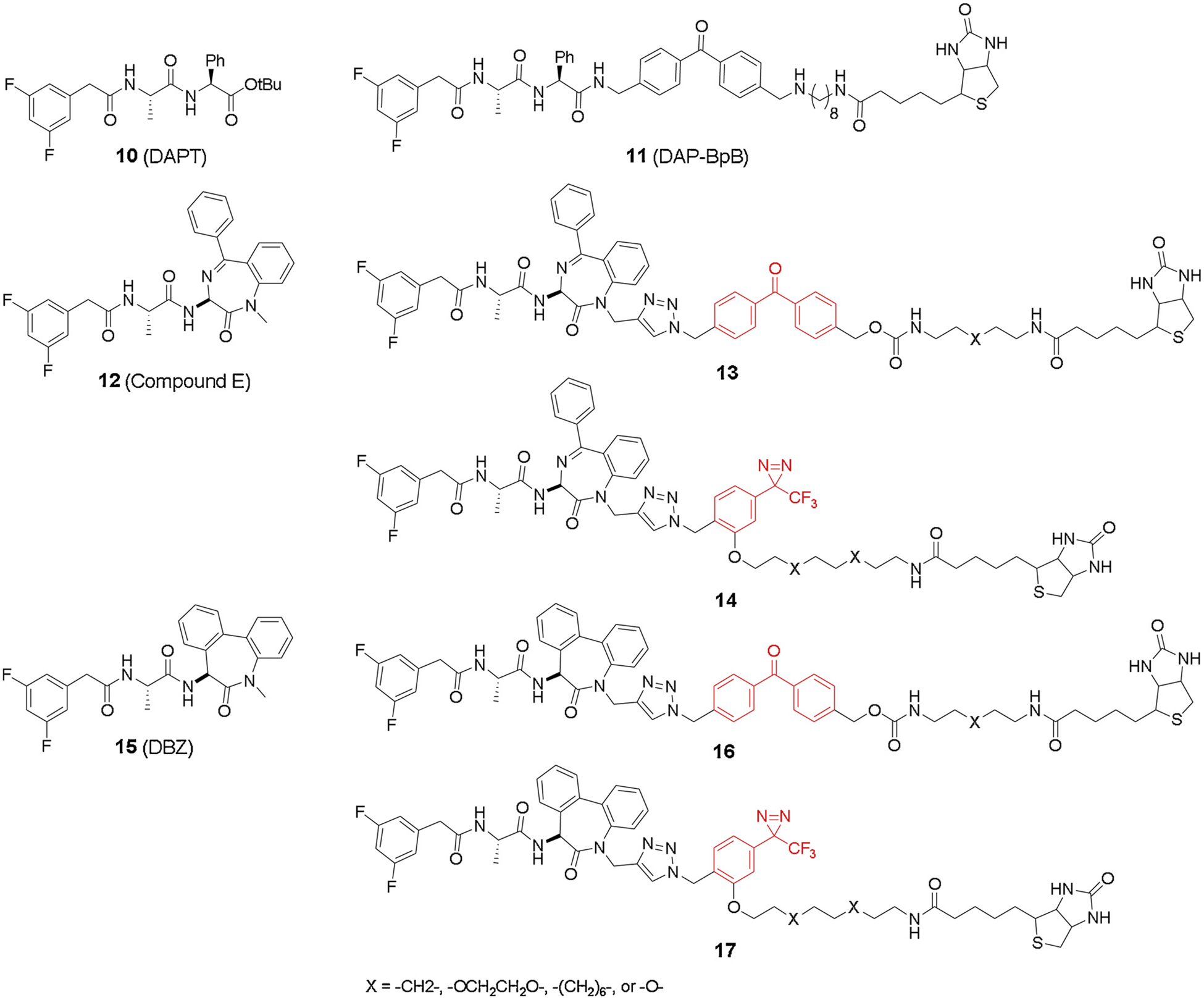
DAPT analogues and photo affinity probes.
BMS-708,163 (Avagacestat) initially gained quite an interest because of its reported selectivity to APP inhibition over Notch [45]. However, this reported Notch-sparing inhibitor was later challenged by a series of photo affinity labeling studies [46,47,67]. 163-BPyne (19, Fig. 6) was synthesized based on BMS-708,163 with a benzophenone moiety and a clickable terminal alkyne [46]. 163-BPyne was found to bind PS1-NTF with high specificity. Subsequent competition studies showed that the binding can be competed with pan-GSIs but not GSMs.
Fig. 6.
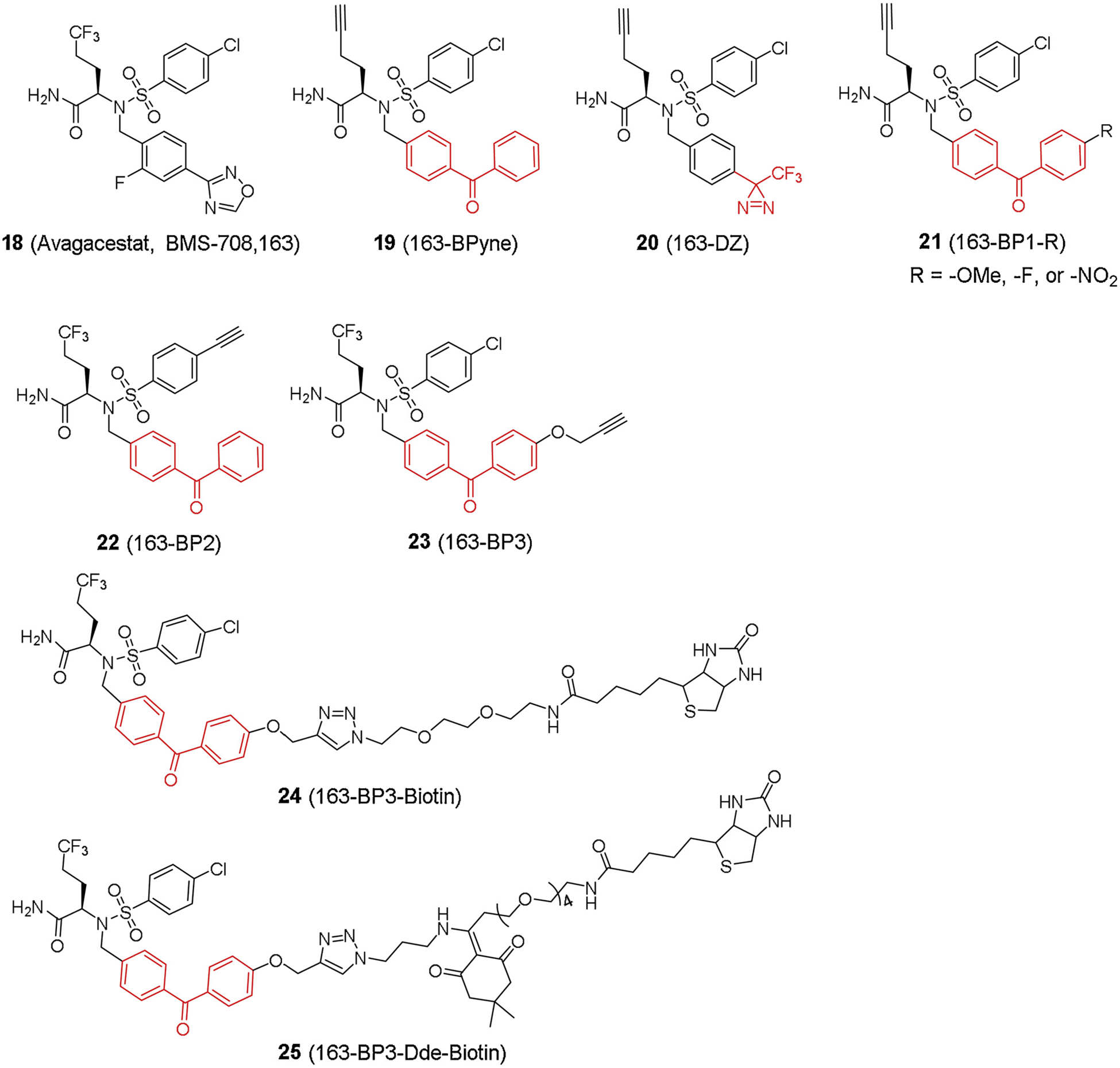
BMS-708,163 based photo affinity probes.
BMS-708,163 might not be a selective inhibitor, but its sub-nanomolar potency and high synthetic feasibility make it an ideal scaffold to explore the molecular determinants for effective photoprobe design. Crump et al. reported a structure-photoreactivity relationship study [67] to investigate how photoreactive groups, alkyne handle position, and substituents affected the labeling efficiency of PS1 subunit. Not surprisingly, phenyl diazirine probe failed to label PS1-NTF, which was consistent with the previous results in both transition state probes and DAPT derivatives. Meanwhile, the data also indicated that benzophenone is the most frequently used photoreactive group for γ-secretase. Most of the substituents on benzophenone group (21, Fig. 6) are well tolerated for photo affinity labeling except the p-nitro substitution. Moreover, the position of the alkyne group was evaluated by directly comparing the labeling efficiencies of the probes. Although their inhibitory potency was almost the same, the location of the alkyne on these probes had a significant impact on labeling efficiency. The mechanism of this dramatic difference was not clear until a recent study revealed the binding model of BMS-708,163 [68]. In this work, four BMS-708,163 based probes with distinct cleavable linkers were made. Afterwards, PS1 was labeled with these probes and eluted under different cleavage conditions. The most efficient probe (25, Figs. 6 and 7) was selected to enrich enough PS1 for proteomics and molecular dynamics (MD) simulation. A PS1-NTF fragment sequence (NETLFPALIYSST, 279–291) was identified as the probe labeling site with photo-insertion on the residue L282 by MS proteomics. Then MD simulations could be utilized to reveal the binding model of BMS-708,163, which showed that the alkyne moiety on the probe was facing towards solvent and easily accessible for azide reporter moiety. This work provided a working model for BMS-708,163 analogues and discovered a new path for exploring binding sites of other clinically relevant GSIs and GSMs.
Fig. 7.
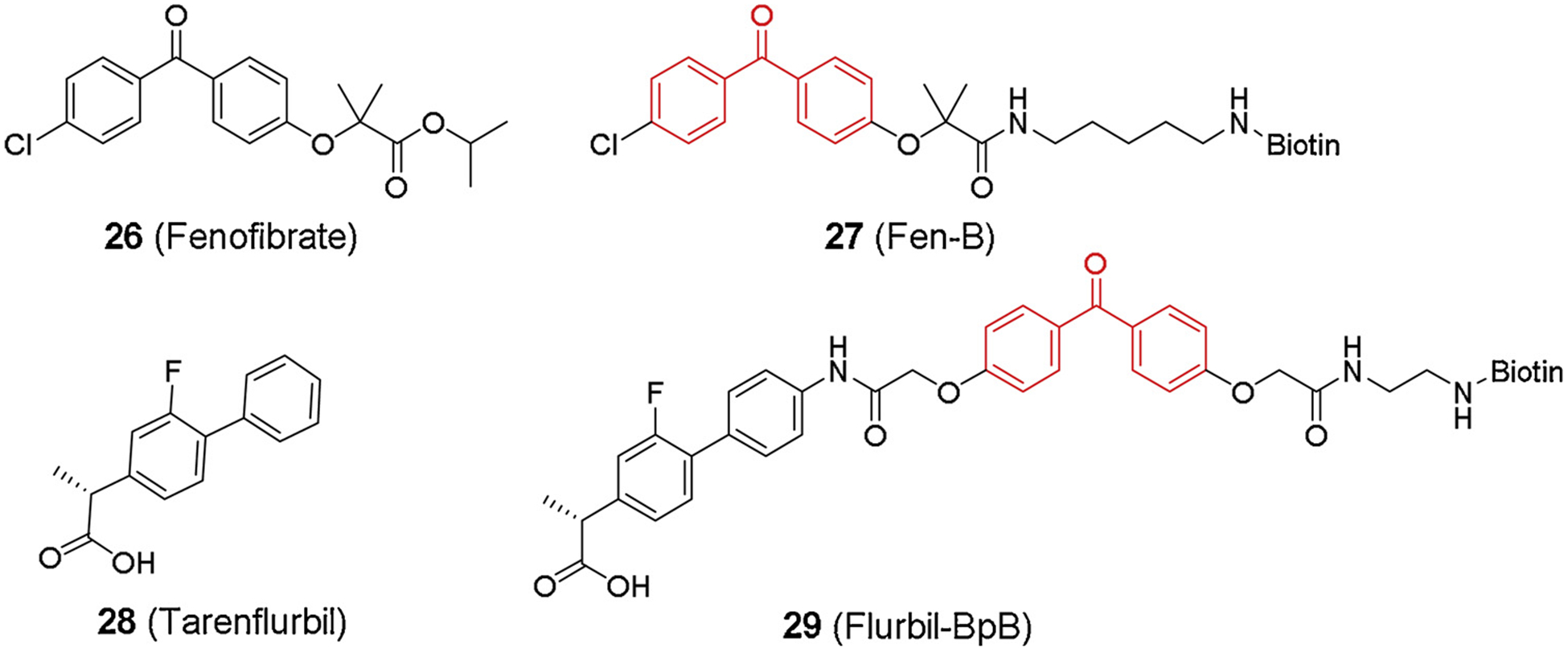
NSAID modulator-based probes.
4. γ-Secretase modulators (GSMs)
Lessons from the clinical studies point to the need for the development of disease modifying agents that reduce γ-secretase activity for the production of Aβ42 without affecting the overall processing of APP and function of other γ-secretase substrates [4,69].
4.1. Development of first generation GSMs
The concept of γ-secretase modulation was discovered when a subset of NSAIDs, such as ibuprofen, indomethacin and sulindac sulfide, were found to selectively lower the formation of Aβ42 in favor of Aβ38 without inhibiting the Notch1 cleavage [69]. Furthermore, the effect of these NSAIDs on Aβ modulation was dissociated from their COX activity [69]. GSMs have many unique characteristics, which include: 1) reducing Aβ42 production; 2) promoting production of shorter forms of Aβ species (Aβ38 or Aβ37); 3) having no significant effect on the total amount of Aβ produced or accumulation of βCTF; and 4) lacking inhibitory effect on Notch cleavage and the cleavage of other γ-secretase substrates. These ideal properties have inspired the development of GSMs as potential disease modifying agents for AD treatment. However, first generation GSMs (NSAIDs derivatives) exhibited low potency in modulation of Aβ42 production and lacked efficacy in clinical studies [70–73].
4.2. Development of second-generation GSMs
A key goal in the development of second-generation GSMs has been to improve the potency and brain availability [4,40,74–77]. Structurally, second generation GSMs can be divided into three categories: acid derived GSMs (acid GSMs), imidazole-based GSMs (imidazole GSMs) and natural product-derived GSMs.
The first two classes have been extensively investigated in cellular and animal models from mouse/rat to guinea pig [4]. Acid GSMs are exemplified by GSM-1 (30, Fig. 8) and close analogs GSM-2 and GSM-10 h. GSM-1 significantly decreased Aβ42 and increased Aβ38 in cells expressing either WT PS1 or WT PS2 [78]. GSM-10 h has been demonstrated to have excellent bioavailability and good CNS penetration [79]. Acute and sub-chronic administration of GSM-10 h to rats decreased Aβ42 in plasma, CSF and brain [80]. Furthermore, GSM-10 h did not cause Aβ rebound in rat plasma nor did it accumulate β-CTF [81].
Fig. 8.
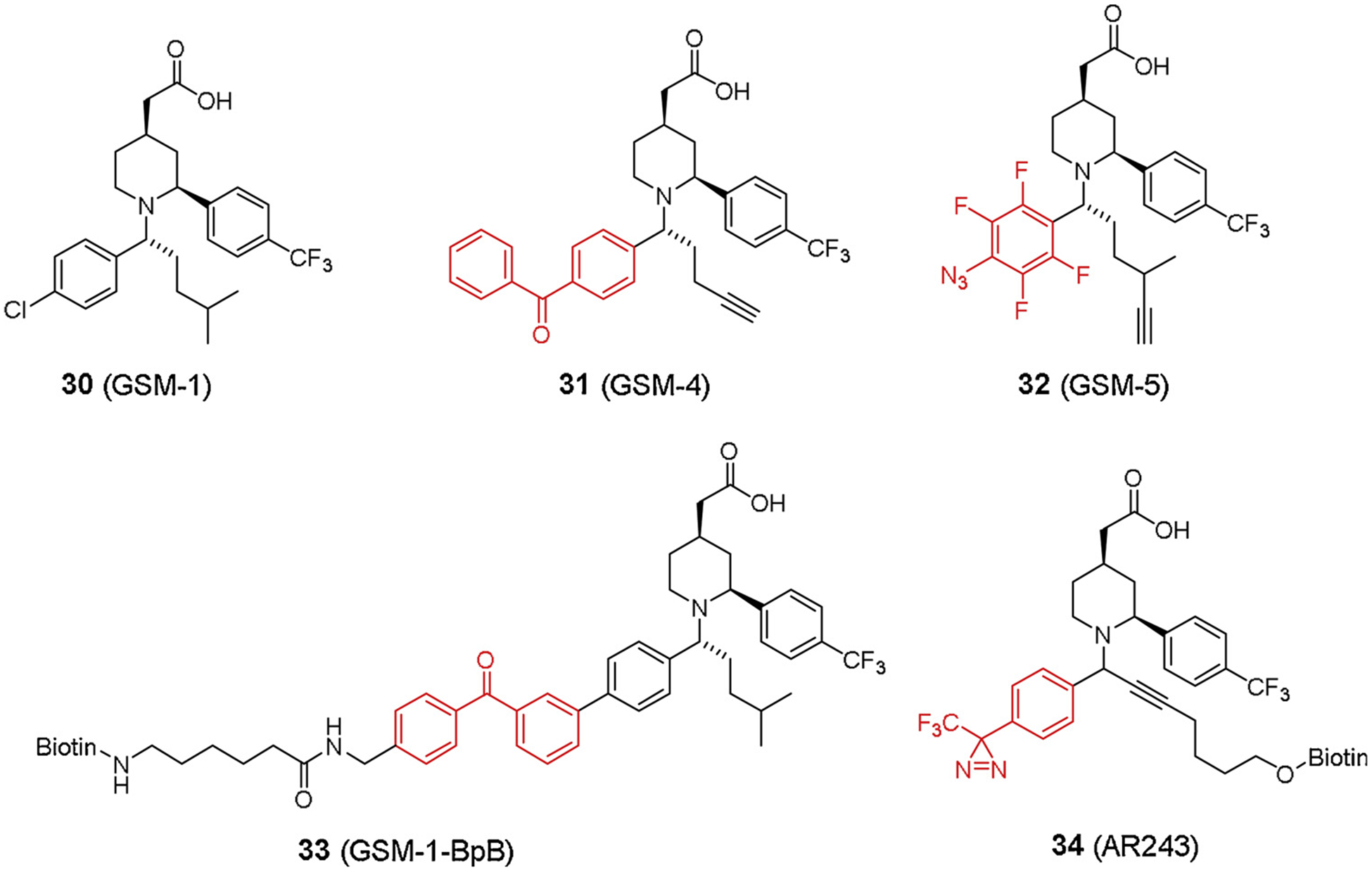
GSM-1 based probes.
Administration of NGP-555 (35, Fig. 9) to 8-month old Tg2576 mice for 7 months caused significant reduction in plaque density and amyloid deposition [82,83]. E2012 (38, Fig. 9) lowered Aβ42, Aβ40 and Aβ39 and raised Aβ37 and, to a lesser extent, Aβ38 [84–86]. E2012 entered phase I trials in 2006, and it represents the first non-NSAID GSM to enter clinical development. Following the observation of lenticular opacity in a 13-week rat safety study, clinical development of E2012 was halted [87]. Eisai announced that E2012 was not developed further in favor of an improved compound, E2212. Clinical studies (doses ranging from 10 mg to 250 mg) showed that E2212 has acceptable safety profiles and promising pharmacodynamic responses, supporting further clinical development of E2212 [88].
Fig. 9.
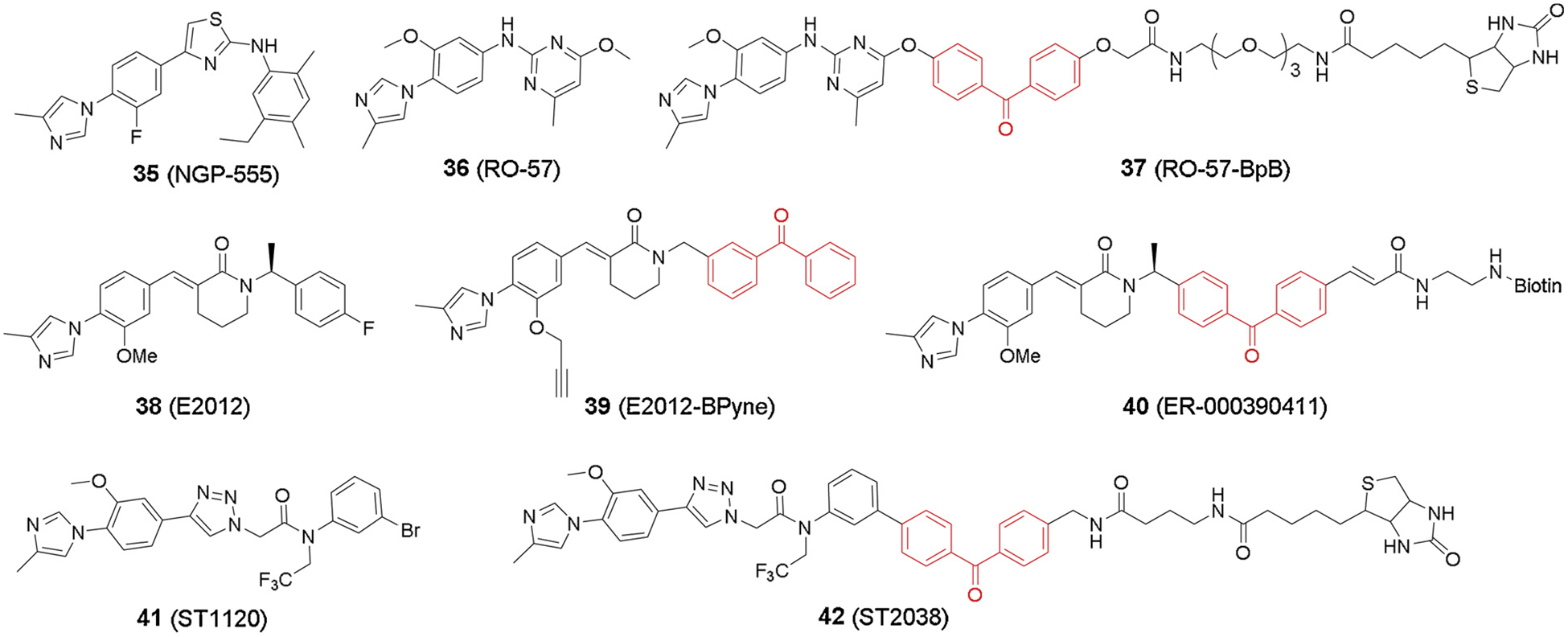
Imidazole-based GSM probes.
Importantly, some studies indicate that GSMs offer advantages over GSIs for development of AD treatment [50,89,90]. A study was conducted comparing the efficacy of two GSIs (Semagacestat and Avagacestat) and GSM-2 [50]. These compounds were administered to wild-type and 5.5 month-old Tg2576 mice for 8 days and Y-maze tests were conducted to evaluate spatial working memory. Only GSM-2 ameliorated the cognitive deficit in Tg2576 mice. While all three drugs reduced hippocampal Aβ42 levels, the β-CTF levels only increased with the two GSIs. Subchronic treatment with Semagacestat impaired normal cognitive function in WT mice, while treatment with GSM-2 had no effect. These findings suggest that the cognitive impairment associated with GSI treatment could be due to, at least in part, β-CTF elevation [50]. Weissmiller et al. found that GSI treatment induced defects in BDNF axonal trafficking and signaling, but GSM treatment did not [89]. D’Avanzo et al. reported that GSMs decreased the production of Aβ42 in human neural progenitor cells and had no adverse effect on neuronal differentiation that is particular to GSI treatment [90]. The data suggests that GSM treatment may not only circumvent the gastrointestinal and cancer toxicities associated with Notch signaling inhibition that is characteristic of GSIs, but may also avoid the β-CTF accumulation that can lead to cognitive decline.
4.3. GSM based probes
4.3.1. NSAID-derived carboxylic acid GSM probe
The earliest GSM PAL probes were designed based on two NSAID compounds, Aβ42-raising fenofibrate and Aβ42-lowering tarenflurbil [91] (Fig. 7). Like many other GSI probes, a benzophenone moiety and a biotin tag were incorporated into the compounds. Interestingly, both type of probes failed to label subunits of γ-secretase complex but instead labeled amyloid precursor protein (APP). Meanwhile, Fen-B labeled APP more efficiently than Notch, which therefore explained the selectivity of GSMs to APP inhibition over Notch. This data revealed a new mechanism of Aβ inhibition through substrate-targeting. To overcome the low potency and poor brain penetration of NSAID GSMs, GSM-1 has been developed selectively inhibiting Aβ42 (IC50 120–348 nM) and increasing Aβ38 [78,92,93]. Soon later, photo affinity labeling studies were concurrently reported to elucidate the molecular mechanism of this compound [94–97] (Fig. 8). Ohki et al. [94] developed a benzophenone-based probe GSM-1-BpB (33, Fig. 8) with a biotin terminus. GSM-1-BpB directly labeled transmembrane domain 1 (TMD1) of PS1, thereby causing a conformational change in the catalytic active site that favored Aβ38 production over Aβ42. Meanwhile, Crump et al. [97] also reported a few clickable GSM-1 based probes (31–32, Fig. 8). Both benzophenone and perfluorophenyl azide were used respectively to substitute chlorophenyl group on GSM-1, and two types of probes directly labeled PS1-NTF of γ-secretase as well, which was consistent with the previous study [94].
Other researches [95,96] reported two continuous studies on design and evaluation of GSM-1 probes (34, Fig. 8) for γ-secretase labeling. Different from previous probes, photoreactive diazirine moiety was used in these studies, which also confirmed PS1-NTF as molecular target of GSM-1.
Based on our previous results, photo group localization on a probe is directly associated with labeling specificity. The idea that utilizes location of benzophenone to label distinct subsites within the active site has been further developed into a multiple photo-affinity approach, referred as photophore walking [98–100]. In brief, the photophore benzophenone group was separately incorporated into different side chains along L-685,458 to target different subsites within the active site (Fig. 3), and thus the conformational change in the active site will also lead to an alteration of the labeling efficiency for the corresponding probe. The photophore walking approach was firstly applied to detect conformational changes of the active site upon small molecule inhibitor or modulator binding [98]. For instance, an enhancement of GY4 labeling was observed in the present of modulator GSM-1 [97], which indicates a possible mechanism that GSM1 allosterically changes the S1 subsite of γ-secretase and therefore leads to selective inhibition of Aβ42. With the same strategy, studies have also shown PS1 FAD mutations like M146 L and E280A result in deeper and distinct S2 subsites, which prefer the substrate with larger residues like Ile in both Aβ42 and Aβ48 and thus lead to an increase in Aβ42 production [99]. Furthermore, a recent study explored the conformational change of γ-secretase induced by γ-secretase activating protein (GSAP) using an L-685,458 derived probe L631 with two benzophenone moieties [101]. Both PS1-NTF and PS1-CTF could be crosslinked together by L631 when GSAP is present in the cells, whereas the L631 cross-linking is absent in GSAP KO cells. Although the role of GSAP in γ-secretase regulation has been questioned [102] because of limited mechanistic evidence, these results strongly support the mechanism that GSAP directly regulates activity and specificity of γ-secretase by altering the catalytic active site.
4.3.2. Non-NSAID imidazole-based GSM probes
Ebke et al. [103] developed GSM RO-57-BpB (37, Fig. 9) that specifically labeled both PS1-NTF and PS2-NTF, but none of the other γ-secretase components, supporting that this class of GSMs target the catalytic subunit of γ-secretase. Pozdnyakov et al. [104] developed the non-NSAID heterocyclic GSM probe E2012-BPyne (39, Fig. 9), which also labeled the PS1-NTF site in membrane or live cells. Unlike acid GSM probes, E2012-BPyne labeling of PS1-NTF was significantly enhanced in the presence of transition state inhibitor L-685,458, suggesting that an E2012 favoring conformation change was induced upon L-685,458 binding. A later study by Takeo et al. [105] proposed a more detailed model for mechanism of action using the phenylimidazole-type GSM probe ST2038 (42, Fig. 9). The labeling efficiency of ST2038 on PS was evaluated both by alanine scanning and by the substituted cysteine accessibility method thereby helping to locate the extracellular pocket formed by HL1/TM2/TM5 for phenylimidazole GSM binding. The subsequent study from the same group [106] further explored this modulation mechanism by another E2012 derived photo probe (40, Fig. 9). The N-terminal region of the HL1 on PS1-NTF was then determined as E2012 binding site, and the following piston movement of the transmembrane domain 1 led to selectivity in Aβ production. Together, all this information helped elevate our understanding of the working mechanism of phenylimidazole GSMs.
5. Conclusion and future perspective
The amyloid cascade hypothesis has greatly influenced both academic and pharmaceutical research for the last thirty years. Under the hypothesis, γ-secretase was regarded as one of therapeutic targets and corresponding GSIs and GSMs have been developed for clinical applications. Unfortunately, none of them has reached the efficacy endpoints in clinical trials, which put the Aβ hypothesis under close scrutiny. Encouragingly, Biogen recently announced their phase III trial results for the Aβ clearance antibody Aducanumab. In one of the two cohorts, early onset AD patients treated with high dose antibody (10 mg/kg) showed a significant reduction of clinical decline after 18 months. It’s still hard to make a conclusion how much benefits patients could get from Aducanumab at this moment, but the new data is a great breakthrough in the field and provides support for Aβ hypothesis.
In addition to the antibody therapy, GSM is another competitive approach in therapeutic discovery. It has been widely found that the complete inhibition of the protease activity by clinical GSIs led to adverse side effects. Therefore, GSMs that selectively lower Aβ42 are much more promising than GSIs for potential clinical use in the future. Moreover, different categories of GSMs have distinct Aβ profiles, indicating existence of multiple modulation mechanism of GSMs. While understandings of the molecular basis of GSMs have rapidly advanced for the last few years based on techniques like cryo-EM and photo affinity labeling, there is much more progress to be made to facilitate clinical development of GSM. Recently, pharmacological studies shown that two GSMs, BPN-15060 [107] and PF-06648671 [108] effectively reduce toxic Aβ peptides at non-toxic doses in preclinical and clinal investigation, showing the promise to develop GSMs as treatment of AD.
Finally, only a small fraction of γ-secretase is catalytically active, which puts a formidable challenge in the field of γ-secretase. The function and activation of the inactive complex is unknown. It is likely that the inactive complex functions as a pool, permitting quick modulation of γ-secretase activity through conversion to active complexes to adapt to cellular and chemical environmental changes. How GSIs and GSMs affect the dynamics of the active and inactive complex in vivo deserves further investigations. Clearly, to be therapeutically efficacious, GSIs and GSMs must block activated γ-secretase.
Acknowledgments
We thank Dr. Grady Nelson for proofreading the manuscript. This work is supported by NIH grant R01NS096275 (YML), RF1AG057593 (YML), R01AG061350 (YML), the JPB Foundation (YML), the MetLife Foundation (YML), Cure Alzheimer’s Fund (YML), The Edward and Della L. Thome Memorial Foundation (YML) and Coins for the Alzheimer’s Research Trust (YML). Authors also acknowledge the MSK Cancer Center Support Grant/Core Grant (Grant P30 CA008748), Mr. William H. Goodwin and Mrs. Alice Goodwin and the Commonwealth Foundation for Cancer Research, the Experimental Therapeutics Center of MSKCC, and the William Randolph Hearst Fund in Experimental Therapeutics
Footnotes
Declaration of Competing Interest
LYM is co-inventor of intellectual property (assay for gamma secretase activity and screening method for gamma secretase inhibitors) licensed to Jiangsu Continental Medical Development.
References
- [1].Hardy JA, Higgins GA, Alzheimer’s disease: the amyloid cascade hypothesis, Science 256 (5054) (1992) 184–185. [DOI] [PubMed] [Google Scholar]
- [2].Hardy J, Selkoe DJ, The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics, Science 297 (5580) (2002) 353–356. [DOI] [PubMed] [Google Scholar]
- [3].O’Brien RJ, Wong PC, Amyloid precursor protein processing and Alzheimer’s disease, Annu. Rev. Neurosci 34 (2011) 185–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Crump CJ, Johnson DS, Li Y-M, Development and mechanism of γ-Secretase modulators for alzheimer’s disease, Biochemistry 52 (19) (2013) 3197–3216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Brown MS, Ye J, Rawson RB, Goldstein JL, Regulated intramembrane proteolysis: a control mechanism conserved from bacteria to humans, Cell 100 (4) (2000) 391–398. [DOI] [PubMed] [Google Scholar]
- [6].De Strooper B, Aph-1, Pen-2, and Nicastrin with Presenilin generate an active gamma-Secretase complex, Neuron 38 (1) (2003) 9–12. [DOI] [PubMed] [Google Scholar]
- [7].Wolfe MS, Xia W, Ostaszewski BL, Diehl TS, Kimberly WT, Selkoe DJ, Two transmembrane aspartates in presenilin-1 required for presenilin endoproteolysis and gamma-secretase activity, Nature 398 (6727) (1999) 513–517. [DOI] [PubMed] [Google Scholar]
- [8].Li YM, Xu M, Lai MT, Huang Q, Castro JL, DiMuzio-Mower J, Harrison T, Lellis C, Nadin A, Neduvelil JG, Register RB, Sardana MK, Shearman MS, Smith AL, Shi XP, Yin KC, Shafer JA, Gardell SJ, Photoactivated gamma-secretase inhibitors directed to the active site covalently label presenilin 1, Nature 405 (6787) (2000) 689–694. [DOI] [PubMed] [Google Scholar]
- [9].Esler WP, Kimberly WT, Ostaszewski BL, Diehl TS, Moore CL, Tsai JY, Rahmati T, Xia W, Selkoe DJ, Wolfe MS, Transition-state analogue inhibitors of gamma-secretase bind directly to presenilin-1, Nat. Cell Biol 2 (7) (2000) 428–434. [DOI] [PubMed] [Google Scholar]
- [10].Ahn K, Shelton CC, Tian Y, Zhang X, Gilchrist ML, Sisodia SS, Li YM, Activation and intrinsic gamma-secretase activity of presenilin 1, Proc Natl Acad Sci U S A 107 (50) (2010) 21435–21440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Takasugi N, Tomita T, Hayashi I, Tsuruoka M, Niimura M, Takahashi Y, Thinakaran G, Iwatsubo T, The role of presenilin cofactors in the gamma-secretase complex, Nature 422 (6930) (2003) 438–441. [DOI] [PubMed] [Google Scholar]
- [12].Edbauer D, Winkler E, Regula JT, Pesold B, Steiner H, Haass C, Reconstitution of gamma-secretase activity, Nat. Cell Biol 5 (5) (2003) 486–488. [DOI] [PubMed] [Google Scholar]
- [13].Kimberly WT, LaVoie MJ, Ostaszewski BL, Ye W, Wolfe MS, Selkoe DJ, Gamma-secretase is a membrane protein complex comprised of presenilin, nicastrin, Aph-1, and Pen-2, Proc Natl Acad Sci 100 (11) (2003) 6382–6387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Kopan R, Ilagan MX, Gamma-secretase: proteasome of the membrane? Nat. Rev. Mol. Cell Biol 5 (6) (2004) 499–504. [DOI] [PubMed] [Google Scholar]
- [15].Haapasalo A, Kovacs DM, The many substrates of presenilin/gamma-secretase, J. Alzheimers Dis 25 (1) (2011) 3–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Kumar D, Ganeshpurkar A, Kumar D, Modi G, Gupta SK, Singh SK, Secretase inhibitors for the treatment of Alzheimer’s disease: long road ahead, Eur. J. Med. Chem 148 (2018) 436–452. [DOI] [PubMed] [Google Scholar]
- [17].Olsauskas-Kuprys R, Zlobin A, Osipo C, Gamma secretase inhibitors of notch signaling, Onco. Ther 6 (2013) 943–955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Ran Y, Hossain F, Pannuti A, Lessard CB, Ladd GZ, Jung JI, Minter LM, Osborne BA, Miele L, Golde TE, Gamma-Secretase inhibitors in cancer clinical trials are pharmacologically and functionally distinct, EMBO Mol. Med 9 (7) (2017) 950–966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].De Strooper B, Lessons from a failed gamma-secretase alzheimer trial, Cell 159 (4) (2014) 721–726. [DOI] [PubMed] [Google Scholar]
- [20].Yang G, Zhou R, Zhou Q, Guo X, Yan C, Ke M, Lei J, Shi Y, Structural basis of notch recognition by human gamma-secretase, Nature 565 (7738) (2019) 192–197. [DOI] [PubMed] [Google Scholar]
- [21].Zhou R, Yang G, Guo X, Zhou Q, Lei J, Shi Y, Recognition of the amyloid precursor protein by human gamma-secretase, Science 363 (6428) (2019) eaaw0930. [DOI] [PubMed] [Google Scholar]
- [22].Singh A, Thornton ER, Westheimer FH, The photolysis of diazoacetylchymotrypsin, J. Biol. Chem 237 (9) (1962) 3006–3008. [PubMed] [Google Scholar]
- [23].Smith E, Collins I, Photoaffinity labeling in target- and binding-site identification, Future Med. Chem 7 (2) (2015) 159–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Sumranjit J, Chung SJ, Recent advances in target characterization and identification by photoaffinity probes, Molecules 18 (9) (2013) 10425–10451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Vodovozova EL, Photoaffinity labeling and its application in structural biology, Biochemistry Mosc. 72 (1) (2007) 1–20. [DOI] [PubMed] [Google Scholar]
- [26].Murale DP, Hong SC, Haque MM, Lee JS, Photo-affinity labeling (PAL) in chemical proteomics: a handy tool to investigate protein-protein interactions (PPIs), Proteome Sci. 15 (1) (2016) 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Lapinsky DJ, Johnson DS, Recent developments and applications of clickable photoprobes in medicinal chemistry and chemical biology, Future Med. Chem 7 (16) (2015) 2143–2171. [DOI] [PubMed] [Google Scholar]
- [28].Geurink PP, Prely LM, van der Marel GA, Bischoff R, Overkleeft HS, Photoaffinity labeling in activity-based protein profiling, Top. Curr. Chem 324 (2011) 85–113. [DOI] [PubMed] [Google Scholar]
- [29].Inui H, Sawada K, Oishi S, Ushida K, McMahon RJ, Aryl nitrene rearrangements: spectroscopic observation of a benzazirine and its ring expansion to a ketenimine by heavy-atom tunneling, J. Am. Chem. Soc 135 (28) (2013) 10246–10249. [DOI] [PubMed] [Google Scholar]
- [30].Leyva E, Platz MS, Persy G, Wirz J, Photochemistry of phenyl azide - the role of singlet and Triplet Phenylnitrene as transient intermediates, J. Am. Chem. Soc 108 (13) (1986) 3783–3790. [Google Scholar]
- [31].Ghosh B, Jones LH, Target validation using in-cell small molecule clickable imaging probes, J MedChemComm 5 (3) (2014) 247–254. [Google Scholar]
- [32].Hill JR, Robertson AAB, Fishing for drug targets: a focus on diazirine photo-affinity probe synthesis, J. Med. Chem 61 (16) (2018) 6945–6963. [DOI] [PubMed] [Google Scholar]
- [33].Johnsson N, Johnsson K, Chemical tools for biomolecular imaging, ACS Chem. Biol 2 (1) (2007) 31–38. [DOI] [PubMed] [Google Scholar]
- [34].Liu DS, Tangpeerachaikul A, Selvaraj R, Taylor MT, Fox JM, Ting AY, Diels–Alder cycloaddition for fluorophore targeting to specific proteins inside living cells, J. Am. Chem. Soc 134 (2) (2012) 792–795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Thirumurugan P, Matosiuk D, Jozwiak K, Click chemistry for drug development and diverse chemical–biology applications, Chem. Rev 113 (7) (2013) 4905–4979. [DOI] [PubMed] [Google Scholar]
- [36].Wright M, Sieber S, Chemical proteomics approaches for identifying the cellular targets of natural products, Nat. Prod. Rep 33 (5) (2016) 681–708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Imbimbo BP, Therapeutic potential of gamma-secretase inhibitors and modulators, Curr. Top. Med. Chem 8 (1) (2008) 54–61. [DOI] [PubMed] [Google Scholar]
- [38].Josien H, Recent advances in the development of gamma-secretase inhibitors, Curr. Opin. Drug Discov. Devel 5 (4) (2002) 513–525. [PubMed] [Google Scholar]
- [39].D’Onofrio G, Panza F, Frisardi V, Solfrizzi V, Imbimbo BP, Paroni G, Cascavilla L, Seripa D, Pilotto A, Advances in the identification of gamma-secretase inhibitors for the treatment of Alzheimer’s disease, Expert Opin. Drug Discov 7 (1) (2012) 19–37. [DOI] [PubMed] [Google Scholar]
- [40].Oehlrich D, Berthelot DJ-C, Gijsen HJ, γ-Secretase modulators as potential disease modifying anti-Alzheimer’s drugs, J. Med. Chem 54 (3) (2010) 669–698. [DOI] [PubMed] [Google Scholar]
- [41].Doody RS, Raman R, Farlow M, Iwatsubo T, Vellas B, Joffe S, Kieburtz K, He F, Sun X, Thomas RG, Aisen PS, Alzheimer’s C Disease Cooperative Study Steering, E. Siemers, G. Sethuraman, R. Mohs, G. Semagacestat Study, A phase 3 trial of semagacestat for treatment of Alzheimer’s disease, N. Engl. J. Med 369 (4) (2013) 341–350. [DOI] [PubMed] [Google Scholar]
- [42].Coric V, van Dyck CH, Salloway S, Andreasen N, Brody M, Richter RW, Soininen H, Thein S, Shiovitz T, Pilcher G, Colby S, Rollin L, Dockens R, Pachai C, Portelius E, Andreasson U, Blennow K, Soares H, Albright C, Feldman HH, Berman RM, Safety and tolerability of the gamma-secretase inhibitor avagacestat in a phase 2 study of mild to moderate alzheimer disease, Arch. Neurol 69 (11) (2012) 1430–1440. [DOI] [PubMed] [Google Scholar]
- [43].Xia X, Qian S, Soriano S, Wu Y, Fletcher AM, Wang XJ, Koo EH, Wu X, Zheng H, Loss of presenilin 1 is associated with enhanced beta-catenin signaling and skin tumorigenesis, Proc Natl Acad Sci U S A 98 (19) (2001) 10863–10868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Nicolas M, Wolfer A, Raj K, Kummer JA, Mill P, Van Noort M, Hui CC, Clevers H, Dotto GP, Radtke F, Notch1 functions as a tumor suppressor in mouse skin, Nat. Genet 33 (3) (2003) 416–421. [DOI] [PubMed] [Google Scholar]
- [45].Gillman KW, Starrett JE Jr., Parker MF, Xie K, Bronson JJ, Marcin LR, McElhone KE, Bergstrom CP, Mate RA, Williams R, Meredith JE Jr., Burton CR, Barten DM, Toyn JH, Roberts SB, Lentz KA, Houston JG, Zaczek R, Albright CF, Decicco CP, Macor JE, Olson RE, Discovery and evaluation of BMS-708163, a potent, selective and orally bioavailable gamma-secretase inhibitor, ACS Med. Chem. Lett 1 (3) (2010) 120–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Crump CJ, Castro SV, Wang F, Pozdnyakov N, Ballard TE, Sisodia SS, Bales KR, Johnson DS, Li YM, BMS-708,163 targets presenilin and lacks notch-sparing activity, Biochemistry 51 (37) (2012) 7209–7211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Chavez-Gutierrez L, Bammens L, Benilova I, Vandersteen A, Benurwar M, Borgers M, Lismont S, Zhou L, Van Cleynenbreugel S, Esselmann H, Wiltfang J, Serneels L, Karran E, Gijsen H, Schymkowitz J, Rousseau F, Broersen K, De Strooper B, The mechanism of gamma-Secretase dysfunction in familial alzheimer disease, EMBO J. 31 (10) (2012) 2261–2274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Zheng H, Koo EH, Biology and pathophysiology of the amyloid precursor protein, Mol. Neurodegener 6 (1) (2011) 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Zheng H, Koo EH, The amyloid precursor protein: beyond amyloid, Mol. Neurodegener 1 (2006) 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Mitani Y, Yarimizu J, Saita K, Uchino H, Akashiba H, Shitaka Y, Ni K, Matsuoka N, Differential effects between gamma-secretase inhibitors and modulators on cognitive function in amyloid precursor protein-transgenic and nontransgenic mice, J. Neurosci 32 (6) (2012) 2037–2050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Lanz TA, Karmilowicz MJ, Wood KM, Pozdnyakov N, Du P, Piotrowski MA, Brown TM, Nolan CE, Richter KE, Finley JE, Fei Q, Ebbinghaus CF, Chen YL, Spracklin DK, Tate B, Geoghegan KF, Lau LF, Auperin DD, Schachter JB, Concentration-dependent modulation of amyloid-beta in vivo and in vitro using the gamma-secretase inhibitor, LY-450139, J. Pharmacol. Exp. Ther 319 (2) (2006) 924–933. [DOI] [PubMed] [Google Scholar]
- [52].Yin YI, Bassit B, Zhu L, Yang X, Wang C, Li YM, {gamma}-Secretase substrate concentration modulates the Abeta42/Abeta40 ratio: implications for alzheimer disease, J. Biol. Chem 282 (32) (2007) 23639–23644. [DOI] [PubMed] [Google Scholar]
- [53].Tagami S, Yanagida K, Kodama TS, Takami M, Mizuta N, Oyama H, Nishitomi K, Chiu YW, Okamoto T, Ikeuchi T, Sakaguchi G, Kudo T, Matsuura Y, Fukumori A, Takeda M, Ihara Y, Okochi M, Semagacestat is a pseudo-inhibitor of gamma-secretase, Cell Rep. 21 (1) (2017) 259–273. [DOI] [PubMed] [Google Scholar]
- [54].Shearman MS, Beher D, Clarke EE, Lewis HD, Harrison T, Hunt P, Nadin A, Smith AL, Stevenson G, Castro JL, L-685,458, an aspartyl protease transition state mimic, is a potent inhibitor of amyloid beta-protein precursor gamma-secretase activity, Biochemistry 39 (30) (2000) 8698–8704. [DOI] [PubMed] [Google Scholar]
- [55].Crump CJ, am Ende CW, Ballard TE, Pozdnyakov N, Pettersson M, Chau DM, Bales KR, Li YM, Johnson DS, Development of clickable active site-directed photoaffinity probes for gamma-secretase, Bioorg. Med. Chem. Lett 22 (8) (2012) 2997–3000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Micchelli CA, Esler WP, Kimberly WT, Jack C, Berezovska O, Kornilova A, Hyman BT, Perrimon N, Wolfe MS, γ-Secretase/presenilin inhibitors for Alzheimer’s disease phenocopy notch mutations in drosophila, Faseb J. 17 (1) (2003) 79–81. [DOI] [PubMed] [Google Scholar]
- [57].Kornilova AY, Das C, Wolfe MS, Differential effects of inhibitors on the gamma-secretase complex. Mechanistic implications, J. Biol. Chem 278 (19) (2003) 16470–16473. [DOI] [PubMed] [Google Scholar]
- [58].Kornilova AY, Bihel F, Das C, Wolfe MS, The initial substrate-binding site of gamma-secretase is located on presenilin near the active site, Proc Natl Acad Sci U S A 102 (9) (2005) 3230–3235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Placanica L, Tarassishin L, Yang G, Peethumnongsin E, Kim SH, Zheng H, Sisodia SS, Li YM, Pen2 and presenilin-1 modulate the dynamic equilibrium of presenilin-1 and presenilin-2 gamma-secretase complexes, J. Biol. Chem 284 (5) (2009) 2967–2977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Villa JC, Chiu D, Brandes AH, Escorcia FE, Villa CH, Maguire WF, Hu CJ, de Stanchina E, Simon MC, Sisodia SS, Scheinberg DA, Li YM, Nontranscriptional role of Hif-1alpha in activation of gamma-secretase and notch signaling in breast cancer, Cell Rep. 8 (4) (2014) 1077–1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Lai MT, Chen E, Crouthamel MC, DiMuzio-Mower J, Xu M, Huang Q, Price E, Register RB, Shi XP, Donoviel DB, Bernstein A, Hazuda D, Gardell SJ, Li YM, Presenilin-1 and Presenilin-2 exhibit distinct yet overlapping {gamma}-Secretase activities, J. Biol. Chem 278 (25) (2003) 22475–22481. [DOI] [PubMed] [Google Scholar]
- [62].Gu Y, Sanjo N, Chen F, Hasegawa H, Petit A, Ruan X, Li W, Shier C, Kawarai T, Schmitt-Ulms G, Westaway D, St George-Hyslop P, Fraser PE, The presenilin proteins are components of multiple membrane-bound complexes that have different biological activities, J. Biol. Chem 279 (30) (2004) 31329–31336. [DOI] [PubMed] [Google Scholar]
- [63].Beher D, Fricker M, Nadin A, Clarke EE, Wrigley JD, Li YM, Culvenor JG, Masters CL, Harrison T, Shearman MS, In vitro characterization of the presenilin-dependent gamma-secretase complex using a novel affinity ligand, Biochemistry 42 (27) (2003) 8133–8142. [DOI] [PubMed] [Google Scholar]
- [64].Dovey HF, John V, Anderson JP, Chen LZ, de Saint Andrieu P, Fang LY, Freedman SB, Folmer B, Goldbach E, Holsztynska EJ, Hu KL, Johnson-Wood KL, Kennedy SL, Kholodenko D, Knops JE, Latimer LH, Lee M, Liao Z, Lieberburg IM, Motter RN, Mutter LC, Nietz J, Quinn KP, Sacchi KL, Seubert PA, Shopp GM, Thorsett ED, Tung JS, Wu J, Yang S, Yin CT, Schenk DB, May PC, Altstiel LD, Bender MH, Boggs LN, Britton TC, Clemens JC, Czilli DL, Dieckman-McGinty DK, Droste JJ, Fuson KS, Gitter BD, Hyslop PA, Johnstone EM, Li WY, Little SP, Mabry TE, Miller FD, Audia JE, Functional gamma-secretase inhibitors reduce beta-amyloid peptide levels in brain, J. Neurochem 76 (1) (2001) 173–181. [DOI] [PubMed] [Google Scholar]
- [65].Morohashi Y, Kan T, Tominari Y, Fuwa H, Okamura Y, Watanabe N, Sato C, Natsugari H, Fukuyama T, Iwatsubo T, Tomita T, C-terminal fragment of presenilin is the molecular target of a dipeptidic gamma-secretase-specific inhibitor DAPT (N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester), J. Biol. Chem 281 (21) (2006) 14670–14676. [DOI] [PubMed] [Google Scholar]
- [66].Fuwa H, Takahashi Y, Konno Y, Watanabe N, Miyashita H, Sasaki M, Natsugari H, Kan T, Fukuyama T, Tomita T, Iwatsubo T, Divergent synthesis of multifunctional molecular probes to elucidate the enzyme specificity of dipeptidic gamma-secretase inhibitors, ACS Chem. Biol 2 (6) (2007) 408–418. [DOI] [PubMed] [Google Scholar]
- [67].Crump CJ, Murrey HE, Ballard TE, Am Ende CW, Wu X, Gertsik N, Johnson DS, Li YM, Development of sulfonamide photoaffinity inhibitors for probing cellular gamma-secretase, ACS Chem. Neurosci 7 (8) (2016) 1166–1173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Gertsik N, Am Ende CW, Geoghegan KF, Nguyen C, Mukherjee P, Mente S, Seneviratne U, Johnson DS, Li YM, Mapping the binding site of BMS-708163 on gamma-secretase with cleavable photoprobes, Cell Chem. Biol 24 (1) (2017) 3–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Weggen S, Eriksen JL, Das P, Sagi SA, Wang R, Pietrzik CU, Findlay KA, Smith TE, Murphy MP, Bulter T, Kang DE, Marquez-Sterling N, Golde TE, Koo EH, A subset of NSAIDs lower amyloidogenic Abeta42 independently of cyclooxygenase activity, Nature 414 (6860) (2001) 212–216. [DOI] [PubMed] [Google Scholar]
- [70].Eriksen JL, Sagi SA, Smith TE, Weggen S, Das P, McLendon DC, Ozols VV, Jessing KW, Zavitz KH, Koo EH, Golde TE, NSAIDs and enantiomers of flurbiprofen target gamma-secretase and lower Abeta 42 in vivo, J. Clin. Invest 112 (3) (2003) 440–449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Wilcock GK, Black SE, Hendrix SB, Zavitz KH, Swabb EA, Laughlin MA, I.I.S.i. Tarenflurbil Phase, Efficacy and safety of tarenflurbil in mild to moderate Alzheimer’s disease: a randomised phase II trial, Lancet Neurol. 7 (6) (2008) 483–493. [DOI] [PubMed] [Google Scholar]
- [72].Green RC, Schneider LS, Amato DA, Beelen AP, Wilcock G, Swabb EA, Zavitz KH, Tarenflurbil Phase G 3 Study, Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial, JAMA J. Am. Med. Assoc 302 (23) (2009) 2557–2564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Imbimbo BP, Why did tarenflurbil fail in Alzheimer’s disease? J. Alzheimers Dis 17 (4) (2009) 757–760. [DOI] [PubMed] [Google Scholar]
- [74].Pettersson M, Kauffman GW, am Ende CW, Patel NC, Stiff C, Tran TP, Johnson DS, Novel gamma-secretase modulators: a review of patents from 2008 to 2010, Expert Opin. Ther. Pat 21 (2) (2011) 205–226. [DOI] [PubMed] [Google Scholar]
- [75].Pettersson M, Stepan AF, Kauffman GW, Johnson DS, Novel gamma-secretase modulators for the treatment of alzheimer’s disease: a review focusing on patents from 2010 to 2012, Expert Opin. Ther. Pat 23 (10) (2013) 1349–1366. [DOI] [PubMed] [Google Scholar]
- [76].Wagner SL, Tanzi RE, Mobley WC, Galasko D, Potential use of gamma-secretase modulators in the treatment of alzheimer disease, Arch. Neurol (2012) 1–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Hall A, Patel TR, Gamma-Secretase modulators: current status and future directions, Prog. Med. Chem 53 (2014) 101–145. [DOI] [PubMed] [Google Scholar]
- [78].Page RM, Baumann K, Tomioka M, Perez-Revuelta BI, Fukumori A, Jacobsen H, Flohr A, Luebbers T, Ozmen L, Steiner H, Haass C, Generation of Abeta38 and Abeta42 is independently and differentially affected by familial Alzheimer disease-associated presenilin mutations and gamma-secretase modulation, J. Biol. Chem 283 (2) (2008) 677–683. [DOI] [PubMed] [Google Scholar]
- [79].Hall A, Elliott RL, Giblin GM, Hussain I, Musgrave J, Naylor A, Sasse R, Smith B, Piperidine-derived gamma-secretase modulators, Bioorg. Med. Chem. Lett 20 (3) (2010) 1306–1311. [DOI] [PubMed] [Google Scholar]
- [80].Hawkins J, Harrison DC, Ahmed S, Davis RP, Chapman T, Marshall I, Smith B, Mead TL, Medhurst A, Giblin GM, Hall A, Gonzalez MI, Richardson J, Hussain I, Dynamics of Abeta42 reduction in plasma, CSF and brain of rats treated with the gamma-secretase modulator, GSM-10h, Neurodegener. Dis 8 (6) (2011) 455–464. [DOI] [PubMed] [Google Scholar]
- [81].Li T, Huang Y, Jin S, Ye L, Rong N, Yang X, Ding Y, Cheng Z, Zhang J, Wan Z, Harrison DC, Hussain I, Hall A, Lee DH, Lau LF, Matsuoka Y, Gamma-secretase modulators do not induce Abeta-rebound and accumulation of beta-C-terminal fragment, J. Neurochem 121 (2) (2012) 277–286. [DOI] [PubMed] [Google Scholar]
- [82].(http://www.neurogeneticpharmaceuticals.com/about-ngp.html).
- [83].Kounnas MZ, Danks AM, Cheng S, Tyree C, Ackerman E, Zhang X, Ahn K, Nguyen P, Comer D, Mao L, Yu C, Pleynet D, Digregorio PJ, Velicelebi G, Stauderman KA, Comer WT, Mobley WC, Li Y-M, Sisodia SS, Tanzi RE, Wagner SL, Modulation of γ-Secretase reduces β-Amyloid deposition in a transgenic mouse model of alzheimer’s disease, Neuron 67 (5) (2010) 769–780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Portelius E, Van Broeck B, Andreasson U, Gustavsson MK, Mercken M, Zetterberg H, Borghys H, Blennow K, Acute effect on the Abeta isoform pattern in CSF in response to gamma-secretase modulator and inhibitor treatment in dogs, J. Alzheimers Dis 21 (3) (2010) 1005–1012. [DOI] [PubMed] [Google Scholar]
- [85].Borgegard T, Jureus A, Olsson F, Rosqvist S, Sabirsh A, Rotticci D, Paulsen K, Klintenberg R, Yan H, Waldman M, Stromberg K, Nord J, Johansson J, Regner A, Parpal S, Malinowsky D, Radesater AC, Li T, Singh R, Eriksson H, Lundkvist J, First and second generation gamma-secretase modulators (GSMs) modulate amyloid-beta (Abeta) peptide production through different mechanisms, J. Biol. Chem 287 (15) (2012) 11810–11819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Hashimoto T, Ishibashi A, Hagiwara H, Murata Y, Takenaka O, Miyagawa T, E2012: a novel gamma-secretase modulator - pharmacology, Alzheimers Dement. 6 (Suppl) (2010) S242. [Google Scholar]
- [87].Nagy C, Schuck E, Ishibashi A, Nakatani Y, Rege B, Logovinsky V, E2012, a novel gamma-secretase modulator, decreases plasma amyloid-beta (Aβ) levels in humans, Alzheimers Dement 6 (Suppl) (2010) S574. [Google Scholar]
- [88].Yu Y, Logovinsky V, Schuck E, Kaplow J, Chang MK, Miyagawa T, Wong N, Ferry J, Safety, tolerability, pharmacokinetics, and pharmacodynamics of the novel gamma-secretase modulator, E2212, in healthy human subjects, J. Clin. Pharmacol 54 (5) (2014) 528–536. [DOI] [PubMed] [Google Scholar]
- [89].Weissmiller AM, Natera-Naranjo O, Reyna SM, Pearn ML, Zhao X, Nguyen P, Cheng S, Goldstein LS, Tanzi RE, Wagner SL, Mobley WC, Wu C, A gamma-secretase inhibitor, but not a gamma-secretase modulator, induced defects in BDNF axonal trafficking and signaling: evidence for a role for APP, PLoS One 10 (2) (2015) e0118379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].D’Avanzo C, Sliwinski C, Wagner SL, Tanzi RE, Kim DY, Kovacs DM, Gamma-Secretase modulators reduce endogenous amyloid beta42 levels in human neural progenitor cells without altering neuronal differentiation, FASEB J. 29 (8) (2015) 3335–3341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Kukar TL, Ladd TB, Bann MA, Fraering PC, Narlawar R, Maharvi GM, Healy B, Chapman R, Welzel AT, Price RW, Moore B, Rangachari V, Cusack B, Eriksen J, Jansen-West K, Verbeeck C, Yager D, Eckman C, Ye W, Sagi S, Cottrell BA, Torpey J, Rosenberry TL, Fauq A, Wolfe MS, Schmidt B, Walsh DM, Koo EH, Golde TE, Substrate-targeting gamma-secretase modulators, Nature 453 (7197) (2008) 925–929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Hannam JC, Hartmann S, Madin A, Ridgill MP, Piperidines and Related Compounds for the Treatment of Dementia, (2007). [Google Scholar]
- [93].Madin A, Ridgill MP, Kulagowski JJ, Piperidine Derivatives for Treatment of Alzheimer’s Disease, (2007). [Google Scholar]
- [94].Ohki Y, Higo T, Uemura K, Shimada N, Osawa S, Berezovska O, Yokoshima S, Fukuyama T, Tomita T, Iwatsubo T, Phenylpiperidine-type gamma-secretase modulators target the transmembrane domain 1 of presenilin 1, EMBO J. 30 (23) (2011) 4815–4824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Rennhack A, Jumpertz T, Ness J, Baches S, Pietrzik CU, Weggen S, Bulic B, Synthesis of a potent photoreactive acidic gamma-secretase modulator for target identification in cells, Bioorg. Med. Chem 20 (21) (2012) 6523–6532. [DOI] [PubMed] [Google Scholar]
- [96].Jumpertz T, Rennhack A, Ness J, Baches S, Pietrzik CU, Bulic B, Weggen S, Presenilin is the molecular target of acidic γ-secretase modulators in living cells, PLoS One 7 (1) (2012) e30484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Crump CJ, Fish BA, Castro SV, Chau DM, Gertsik N, Ahn K, Stiff C, Pozdnyakov N, Bales KR, Johnson DS, Li YM, Piperidine acetic acid based gamma-secretase modulators directly bind to Presenilin-1, ACS Chem. Neurosci 2 (12) (2011) 705–710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Shelton CC, Zhu L, Chau D, Yang L, Wang R, Djaballah H, Zheng H, Li YM, Modulation of gamma-secretase specificity using small molecule allosteric inhibitors, Proc. Natl. Acad. Sci. U S A 106 (48) (2009) 20228–20233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Chau DM, Crump CJ, Villa JC, Scheinberg DA, Li YM, Familial Alzheimer disease presenilin-1 mutations alter the active site conformation of gamma-secretase, J. Biol. Chem 287 (21) (2012) 17288–17296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Gertsik N, Chau DM, Li YM, Gamma-secretase inhibitors and modulators induce distinct conformational changes in the active sites of gamma-secretase and signal peptide peptidase, ACS Chem. Biol 10 (8) (2015) 1925–1931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Wong E, Liao GP, Chang JC, Xu P, Li YM, Greengard P, GSAP modulates gamma-secretase specificity by inducing conformational change in PS1, Proc. Natl. Acad. Sci. U S A 116 (13) (2019) 6385–6390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Hussain I, Fabregue J, Anderes L, Ousson S, Borlat F, Eligert V, Berger S, Dimitrov M, Alattia JR, Fraering PC, Beher D, The role of gamma-secretase activating protein (GSAP) and imatinib in the regulation of gamma-secretase activity and amyloid-beta generation, J. Biol. Chem 288 (4) (2013) 2521–2531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Ebke A, Luebbers T, Fukumori A, Shirotani K, Haass C, Baumann K, Steiner H, Novel {gamma}-secretase modulators directly target presenilin, J. Biol. Chem (2011), 10.1074/jbc.C111.276972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Pozdnyakov N, Murrey HE, Crump CJ, Pettersson M, Ballard TE, Am Ende CW, Ahn K, Li YM, Bales KR, Johnson DS, Gamma-Secretase modulator (GSM) photoaffinity probes reveal distinct allosteric binding sites on presenilin, J. Biol. Chem 288 (14) (2013) 9710–9720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Takeo K, Tanimura S, Shinoda T, Osawa S, Zahariev IK, Takegami N, Ishizuka-Katsura Y, Shinya N, Takagi-Niidome S, Tominaga A, Ohsawa N, Kimura-Someya T, Shirouzu M, Yokoshima S, Yokoyama S, Fukuyama T, Tomita T, Iwatsubo T, Allosteric regulation of gamma-secretase activity by a phenylimidazole-type gamma-secretase modulator, Proc. Natl. Acad. Sci. U S A 111 (29) (2014) 10544–10549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Cai T, Yonaga M, Tomita T, Activation of gamma-secretase trimming activity by topological changes of transmembrane domain 1 of presenilin 1, J. Neurosci 37 (50) (2017) 12272–12280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Wagner SL, Rynearson KD, Duddy SK, Zhang C, Nguyen PD, Becker A, Vo U, Masliah D, Monte L, Klee JB, Echmalian CM, Xia WM, Quinti L, Johnson G, Lin JH, Kim DY, Mobley WC, Rissman RA, Tanzi RE, Pharmacological and toxicological properties of the potent oral gamma-secretase modulator BPN-15606, J. Pharmacol. Exp. Ther 362 (1) (2017) 31–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Ahn JE, Carrieri C, Dela Cruz F, Fullerton T, Hajos-Korcsok E, He P, Kantaridis C, Leurent C, Liu RC, Mancuso J, da Costa LM, Qiu RL, Pharmacokinetic and pharmacodynamic effects of a gamma-secretase modulator, PF-06648671, on CSF amyloid-beta peptides in randomized phase I studies, Clin. Pharmacol. Ther (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]