

ABSTRACT
Actomyosin networks are some of the most crucial force-generating components present in developing tissues. The contractile forces generated by these networks are harnessed during morphogenesis to drive various cell and tissue reshaping events. Recent studies of these processes have advanced rapidly, providing us with insights into how these networks are initiated, positioned and regulated, and how they act via individual contractile pulses and/or the formation of supracellular cables. Here, we review these studies and discuss the mechanisms that underlie the construction and turnover of such networks and structures. Furthermore, we provide an overview of how ratcheted processivity emerges from pulsed events, and how tissue-level mechanics are the coordinated output of many individual cellular behaviors.
KEY WORDS: Myosin, Actin, Morphogenesis, Pulsatility, Ratcheting
Summary: Actomyosin networks direct morphogenesis through contractile pulses or the formation of supracellular cables. This Review summarizes the mechanics of network assembly, and provides insights into cell ratcheting, force transmission and pulsatility.
Introduction
Any observer who has watched an animal embryo develop has seen that the tissues that make up the embryo show a remarkable dynamism; indeed, it is this dynamism that has drawn many researchers to developmental biology. As these tissues move and grow, it is obvious that there is a certain limited fluidity, or viscousness, to their movements. Also, as tissues collide, or as a tissue is mechanically probed, a resistance to deformation can be observed, and a ‘bouncing back’ of the tissue often occurs. It is this remarkable visco-elastic property of cellular life that underlies morphogenesis and is the ground state for all processes that seek to change tissue topologies. This essential quality of tissues is, of course, the result of the individual properties of the cells that form the tissue.
Given this materials backdrop, how then do cells exert forces that lead to the observed changes of morphogenesis? Although there are many ways that this can happen (for example, by using pushing forces generated by filament polymerization, or adhesive forces that are directed by protein engagement), one of the central mechanisms involves using the function of motor proteins to drive contractile force generation within the cellular cortex (Heer and Martin, 2017; Blanchard et al., 2018). These cortical cellular regions, which directly underlie and support the surface of the cell, are composed of filamentous actin networks that provide much of the surface-associated elasticity of the cell (Levayer and Lecuit, 2012; Salbreux et al., 2012). Non-muscle Myosin II motor proteins then act on these filamentous actin networks to drive filament sliding and contractile force generation. Together, these actin and myosin (actomyosin) networks provide the forces that can shape cells and tissues during morphogenesis. Indeed, across a huge variety of morphological processes, the tell-tale flash of tagged-Myosin fluorescence often precedes cell deformations.
Here, we summarize our current understanding of actomyosin networks and tissue-generated contractile forces based on recent work in the field. We first provide an overview of the two key types of actomyosin networks that function during development: pulsatile flows and supracellular cables. We then discuss how actomyosin assemblies are initiated and terminated, as well as the upstream regulatory networks that control the timing and localization of Myosin II activation. Finally, we review how progressive cell-shape changes are elicited out of cyclic force events and how these forces are actively transmitted across tissues.
Actomyosin network organization: pulsatile flows versus supracellular cables
Actomyosin networks are organized in a variety of structural archetypes, from the linear Formin-driven cytokinetic rings found in dividing cells to the highly regular sarcomeric arrays present in skeletal muscle cells. Two of the most relevant structural types to morphogenesis are: (1) pulsatile, highly-transient actomyosin populations; and (2) supracellular cables that are mechanically-linked across a number of cells (Fig. 1). Pulsatile populations are often (but not always) associated with apical cell surfaces and tend to be highly dynamic and relatively disorganized, whereas supracellular cables appear coordinated and to have higher stability (Rodal et al., 2015; Sutherland and Lesko, 2020). Pulsatile actomyosin networks may show remarkable degrees of ‘flow’, in which actomyosin arrays are physically displaced across cortical surfaces, although the degree of movement can vary greatly. Supracellular cables permit the coordinated application of tension at scales from many cells to entire tissues. For example, during wound healing, an actomyosin cable forms at the leading edge of the wound boundary and the applied mechanical tension coordinates the cell migration and remodeling of adhesive networks necessary for wound closure (Martin and Lewis, 1992; Brock et al., 1996; Kiehart et al., 2000; Davidson et al., 2002; Wood et al., 2002; Abreu-Blanco et al., 2012; Fernandez-Gonzalez and Zallen, 2013). Supracellular cables can also stiffen entire tissue geometries and direct resistance to mechanical stress (Duda et al., 2019), or provide a mechanical cue to orient cell divisions at compartmental boundaries (Scarpa et al., 2018). At a smaller scale, the local coordination of actomyosin cables can drive efficiencies in elongation movements through the formation of multicellular rosettes (Blankenship et al., 2006). By contrast, highly dynamic pulses of Myosin II can direct rapid oscillations in cell areas and, through ratcheting activities that direct unidirectional processivity, can mediate lasting changes in cell surfaces.
Fig. 1.

Pulsatile actomyosin network and supracellular actomyosin cables during morphogenesis. (A) During Drosophila gastrulation, intercalary processes use radial pulses from medial Myosin II networks as well as actomyosin cables that form on interfaces oriented along the dorsal-ventral axis to direct interface contraction. Polarized actomyosin cables also regulate the direction of oriented cell divisions. (B) During Drosophila dorsal closure, actomyosin cables at the leading edge of lateral epidermal cells coordinate the ratcheted dorsal advancement of the epithelium over pulsed contractions of the amnioserosal cells. (C) During Drosophila ventral furrow formation (mesoderm ingression), pulsatile apical-medial actomyosin networks direct the constriction of the apical cell surface. (D) During asymmetric cell division in the early C. elegans embryo, pulsatile myosin flows direct cytoplasmic flows and anterior-posterior polarity. (E) In models of wound healing, pulsed myosin flows contribute to the building of a supracellular actomyosin cable around the wound to promote wound closure. (F) In Xenopus, an apical-medial actomyosin network in bottle cells (BC) directs apical constriction and the invagination of the mesoderm. Black arrows indicate the movement of cells/tissues/actomyosin flows.
Interestingly, pulsatile actomyosin populations and supracellular cables are often found in the same tissues and even the same cells. Indeed, these populations are often inter-related. In wound healing in the Drosophila notum, rapid waves of actomyosin pulses flow towards the wound margin immediately after wounding and locally drive the apical constriction of cells (Antunes et al., 2013). These waves then appear to coalesce at the margin to form the coordinating actomyosin cable. Similarly, actomyosin pulses have been observed to flow from apical cell surfaces to E-cadherin-containing cell junctions during cell intercalation to form junctional cable-like arrays (Rauzi et al., 2010), although recent evidence has also implicated dedicated Myosin-activating signaling pathways (De Las Bayonas et al., 2019). Non-pulsatile flows can also contribute to the formation of an embryo-encompassing actomyosin subcellular ring during zebrafish epiboly (Behrndt et al., 2012). Interestingly, supracellular behaviors can also emerge out of seemingly pulsatile networks if there are high enough levels of connectivity (Yevick et al., 2019). In the following sections, we discuss recent advances in understanding the mechanisms that initiate and terminate changes in actomyosin behaviors as well as those that mechanically link force generation to produce enduring changes in cell shapes.
Actomyosin flows and the role of pulsatility
A key aspect of actomyosin function during development is that it is transient and often occurs in pulses that act over a time scale of several minutes (Munro et al., 2004; Martin et al., 2009; Solon et al., 2009; Rauzi et al., 2010; Blanchard et al., 2010; David et al., 2010; Fernandez-Gonzalez and Zallen, 2011; Sawyer et al., 2011; Xie and Martin, 2015; Miao et al., 2019). Actomyosin flows result from imbalances in either myosin activation or how the actomyosin network is anchored to the cell cortex or sites of cell-cell/cell-extracellular matrix adhesion. Actomyosin flows were first observed in Caenorhabditis elegans embryos at the one-cell stage, when cortical flows were shown to generate early embryo asymmetries that are essential for controlling the localization of partitioning defective (PAR) proteins that direct asymmetric cell fates (Munro et al., 2004). Pulsed actomyosin contractions were then observed to drive the apical constriction events that underlie mesoderm invagination (Martin et al., 2009) as well as cell intercalation and dorsal closure in Drosophila (Solon et al., 2009; Rauzi et al., 2010; Blanchard et al., 2010; David et al., 2010; Fernandez-Gonzalez and Zallen, 2011; Sawyer et al., 2011). Although these pulses often drive oscillations in apical cell areas, they can also occur in basal cell regions (e.g. during tissue elongation in the fly ovary; He et al., 2010), and can direct the coordinated invagination of large numbers of cells (e.g. during mesoderm invagination and dorsal closure; Martin et al., 2009; Solon et al., 2009; Blanchard et al., 2010; David et al., 2010) or the ingression of single cells (e.g. endodermal precursor cell internalization in C. elegans and neuroblast ingression in Drosophila; Roh-Johnson et al., 2012; Simões et al., 2017; An et al., 2017). Actomyosin pulsing also directs compaction of the early mouse embryo (Maître et al., 2015) as well as convergent extension movements in Xenopus (Kim and Davidson, 2011) and epithelial cell extrusion in Drosophila (Michel and Dahmann, 2020). In addition, although often thought of as driving the contraction of cell surfaces, depending on how cell geometries are arranged, actomyosin pulses can lead to the lengthening of associated cell interfaces (Collinet et al., 2015; Yu and Fernandez-Gonzalez, 2016) as well as the ratcheted sliding of tricellular vertices (Vanderleest et al., 2018).
But why is there this commonality in pulsing in so many different systems? In part, this is likely because of the common biochemical elements that regulate, and potentially feedback on, actomyosin function (as discussed in the sections below). The ‘deeper’ question of what this pulsing achieves for developing cells and tissues is also now beginning to emerge. The answer may lay in part in the amount of contraction disordered actomyosin networks can achieve as well as the timescales of the various processes that are coordinated with mechanical deformations to drive lasting cell-shape changes (see ‘Ratcheting mechanisms that drive progressive changes in cell shapes’ section). It has been shown that the endocytic processes responsible for remodeling cell surface adhesion networks coordinate with actomyosin-driven contractions and typically occur on a minutes time scale (Rappoport and Simon, 2003; Ehrlich et al., 2004; Loerke et al., 2009; Levayer et al., 2011; Jewett et al., 2017; Miao et al., 2019). In addition, filamentous actin remodeling often needs time to occur to decrease the elastic response present in the cell cortex (Clément et al., 2017). Interestingly, recent work using an optogenetic system in cultured Caco-2 epithelial cells to artificially regulate myosin II pulse lengths demonstrated a saturation in the amount of irreversible contraction a single pulse can achieve (Cavanaugh et al., 2020). Pulses that varied from 10 min to 40 min produced a similar ∼20% irreversible reduction in interface length after pulse termination. This same study also implicated endocytic and filamentous actin mechanisms in providing the ratchet-like activity that shortens individual cell interfaces and suggested that a distinct central region of cellular interfaces is the primary region capable of directing ratcheting. The observed saturation of contractile lengths poses an important possible explanation for why discretizing contractions into pulsed regimes is an effective cellular approach. Finally, pulsing may permit an essential absence of synchronicity between individual cell contractions. This may allow the staggered deformation of cells within a larger tissue context to keep strain rates at a level that maintain tissue and cytoskeletal integrity. Indeed, in circumstances where pulsatility, but not contractility, is compromised, tears and separations in cytoskeletal networks and their connections to adhesive nodes are observed (Jodoin et al., 2015; Mason et al., 2016). Thus, pulsatility appears crucial over a variety of different scales, from permitting cortical and endocytic-dependent remodeling to preventing excess tissue strain rates.
The control of Myosin II activity
The myosin superfamily is composed of distinct classes of proteins, each containing head domains, which can interact with actin and hydrolyze ATP, and a diverse tail domain. The conventional two-headed myosins, myosin II class members, consist of a pair of heavy chains, a pair of light chains (ELCs) and a pair of regulatory light chains (RLCs) (Hartman and Spudich, 2012). Myosin II network assembly, activity and function are regulated by phosphorylation and dephosphorylation (Fig. 2). Electron microscopy has shown that the two heads of dephosphorylated Myosin interact with each other, which blocks actin binding and ATPase activity (Wendt et al., 2001; Jung et al., 2008). Additional electrostatic interactions between a blocked head and the first section of the tail may also contribute to the stability of inactivated Myosin; this concept is supported by biochemical data and crystal structure analysis of a portion of the Myosin tail (Woodhead et al., 2005; Blankenfeldt et al., 2006). The inhibitory interactions between head domains are then disrupted when Myosin is activated (Jung et al., 2008).
Fig. 2.

Non-muscle Myosin II structure and activation. Myosin II can be phosphorylated by a number of different kinases (ROCK, MRCK, MLCK and Citron kinase). This phosphorylation causes Myosin II activation and unfolding and its assembly into bipolar filaments. Phosphorylated Myosin II is inactivated by Myosin phosphatase.
Myosin activation is primarily regulated by phosphorylation of the RLCs (Bresnick, 1999; Heissler and Manstein, 2013). At the amino acid level, highly conserved residues (serine-19 and threonine-18 in mammals, serine-21 and threonine-20 in Drosophila) act as the primary and secondary phosphorylation sites, respectively. In vitro experiments suggest that phosphorylation at these sites induces conformational changes in the head and tail domains, allowing Myosin to unfold and form bipolar filaments, bind filamentous actin and promote actin-activated ATPase activity (Scholey et al., 1980; Craig et al., 1983). The kinases primarily responsible for RLC phosphorylation, and consequently myosin II activation, are rho-associated coiled coil-containing kinase (ROCK) and myosin light chain kinase (MLCK; also known as Mylk) (Ikebe and Hartshorne, 1985; Amano et al., 1996; Totsukawa et al., 2000). ROCK activates myosin II through phosphorylation, but can also phosphorylate the myosin binding subunit (MBS) of myosin phosphatase to render it inactive (Kimura et al., 1996; Hartshorne et al., 1998). Other kinases have also been found to possess similar activating abilities, such as myotonic dystrophy related Cdc42-binding kinase (MRCK; also known as CDC42BPA) and citron kinase (CIT; Tan et al., 2008; Yamashiro et al., 2003). MRCK can preferentially phosphorylate myosin RLC at serine 19 and acts during cell protrusion and neurite outgrowth (Tan et al., 2008; Chen et al., 1999). Citron kinase, like ROCK, is an effector of activated Rho and can regulate myosin II activation through phosphorylation of both serine-19 and threonine-18, and is required for cytokinetic ring function (Di Cunto et al., 1998; Yamashiro et al., 2003). However, in vitro experiments suggest that, unlike ROCK, MBS cannot be phosphorylated by citron kinase (Yamashiro et al., 2003).
Myosin phosphatases are equally important in the regulation of Myosin II activity. For example, basal Myosin II recruitment and area oscillations in basal cellular regions are increased by loss of function of the myosin light chain phosphatase subunit gene flapwing during egg formation in the Drosophila ovary (Valencia-Expósito et al., 2016). Myosin phosphatase (MBS/Mypt-75D) function is also required for pulsing during cell intercalation and apical construction during Drosophila gastrulation (Vasquez et al., 2014; Munjal et al., 2015). Thus, Myosin II activity is most proximally controlled by the kinases and phosphatases that regulate RLC phosphorylation states.
Upstream activation of Myosin II networks
As discussed above, Myosin II is activated through phosphorylation by the conserved action of kinases in a diverse array of morphogenetic processes, such as apical constriction, cell intercalation and cytokinesis. But how is this activation matched to the particular process that Myosin II function is used in? Below, we discuss three different signaling pathways underlying the regulation of Myosin II pulsatility, flow and activation (Fig. 3).
Fig. 3.
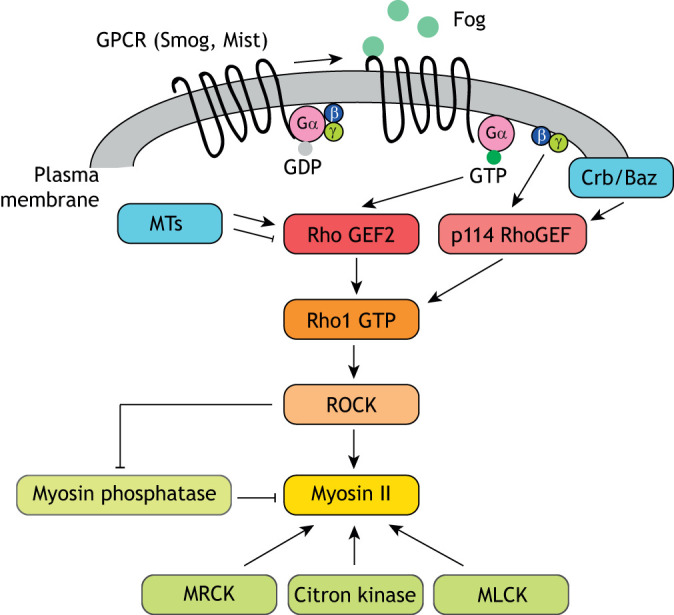
Signaling pathways leading to Myosin II activation. Myosin II can be activated by the ROCK, MRCK, Citron kinase and MLCK kinases and is inactivated by Myosin phosphatase. The activation of ROCK is controlled by GPCRs and their downstream G proteins, as well as apical polarity proteins Crumbs (Crb) and Bazooka (Baz), and microtubule networks (MTs). In Drosophila, the ligand Fog binds to GPCRs to activate heterotrimeric G proteins, which in turn bind to different RhoGEFs to activate the Rho1/ROCK/Myosin signaling pathway.
Much recent work has focused on G protein-coupled receptors (GPCRs) and their downstream heterotrimeric G proteins. In the Drosophila embryo, the GPCRs Smog and Mist can be bound by the secreted ligand Fog (Manning et al., 2013; Kerridge et al., 2016). Upon binding, the guanine nucleotide exchange activity of these receptors catalyzes and separates heterotrimeric G proteins. After Gα12/13 [Concertina (Cta) in Drosophila] dissociates from the Gβ13F/Gγ1 subunits, the active Gα12/13-GTP binds to RhoGEF2, which promotes GTP exchange on the small GTPase Rho1 and in turn activates Rho Kinase and Myosin II at the apical surface (Kerridge et al., 2016). The transmembrane protein T48 also contributes to apical constriction and RhoGEF2 activation in a Fog-independent manner (Kölsch et al., 2007). Interestingly, the heterotrimeric Gβ13F/Gγ1 subunits act in a separate pathway in which they can bind to and activate a different RhoGEF, p114RhoGEF/Cysts. The activation of p114RhoGEF/Cysts turns on junctional, but not apical-medial, Rho1/ROCK/Myosin II signaling in the ectoderm (De Las Bayonas et al., 2019). p114RhoGEF/Cysts also represents an intriguing potential convergence point between GPCR signaling and apical identity proteins; it physically interacts with Crumbs and Bazooka, which are key apical polarity proteins, and disrupted Crumbs function phenocopies the Myosin II junctional depletion observed after p114RhoGEF/Cysts disruption in later stage embryos (Silver et al., 2019). This suggests that apical positioning cues and GPCR-based activation may cooperate to localize actomyosin function in the developing epithelium. In addition, the above evidence indicates a multi-pronged effect of GPCR-based signaling pathways on Myosin II activities and the cell-shape changes that underlie gastrulation movements.
The main factors that regulate these GPCR signaling pathways and the levels of activated Myosin are ligand availability and endocytic-based control of GPCR function. During Drosophila gastrulation, high levels of apical-medial Myosin II activation in the invaginating mesoderm are a result of high levels of Fog expression in this tissue, whereas the low apical Myosin II activation typical of ectodermal cells corresponds to low Fog expression in the ectoderm. This potentially indicates that ligand availability controls the level of GPCR signaling and, therefore, Myosin concentration (Kerridge et al., 2016). Indeed, increasing Fog expression increases medial Myosin levels and can induce apical flattening in the ectoderm (Morize et al., 1998; Dawes-Hoang et al., 2005). Interestingly, overexpression of Fog induces the partitioning of Smog GPCRs into tubular plasma membrane clusters, potentially indicative of GPCR-Rho1 signaling hubs (Jha et al., 2018). Clathrin-mediated endocytosis, mediated by G protein-coupled receptor (GRKs) and β-arrestin function, results in the removal of activated GPCRs from the surface as well as reducing the availability of receptors for ligand activation (Jha et al., 2018).
In a second pathway, ion-dependent signaling is coupled to Myosin II contractility. This is observed, for example, during zebrafish primordial germ cell migration. In this context, calcium levels have been observed to locally rise at sites where cell protrusions form, and an artificial reduction in Ca2+ levels causes aberrant cell mobility and migration, processes in which Myosin II-mediated force generation is required (Blaser et al., 2006). But how does calcium affect Myosin II? In vitro work has indicated that RLC is not phosphorylated and cannot bind to actin in the absence of calcium (Scholey et al., 1980). Moreover, in scratch-wounded cell culture chambers, inhibition of calcium fluxes with BAPTA leads to failure of RLC phosphorylation and Myosin II recruitment, indicating that calcium might be a primary determinant of phosphorylation (Betapudi et al., 2010). However, negative feedback pathways have also been shown to depend on calcium levels. The activation by MLCK phosphorylation of Myosin is inhibited by increased calcium levels (Stull et al., 1990). Furthermore, a continuous increase in inhibitory MLCK phosphorylation leads to decreased MLCK kinase activity and force generation, which provides negative feedback to control the amplitude of contractile forces (Tansey et al., 1994).
Finally, Myosin II contractility is regulated by factors that provide apical-basal spatial information in epithelial cells. Both microtubule networks and cell polarity proteins are important regulators of actomyosin localization and function. There are several potential mechanisms through which microtubules can impact actomyosin function. One such mechanism involves microtubule depolymerization, which is thought to promote actomyosin-based contractility in various systems. For example, actomyosin-based cortical flows in Xenopus oocytes were observed to speed up when microtubules are depolymerized with nocodazole (Canman and Bement, 1997). More specifically, RhoGEF2 has been found to bind microtubule ‘comets’ at the plus end in an EB1-dependent manner in Drosophila S2 cells and embryos (Rogers et al., 2004; De Las Bayonas et al., 2019). When constitutively active Gα12/13/Cta is expressed in the fly germband epithelium, RhoGEF2 accumulates at high levels in medial pools and is depleted from EB1+ comets. In addition, microtubule depolymerization leads to an apical, but not junctional, increase in Myosin II, suggesting that microtubule ends act to sequester RhoGEF2 from medial/apical surfaces (De Las Bayonas et al., 2019). However, microtubules can also positively regulate Myosin II recruitment and actomyosin contractility via direct physical interactions and transport. In the early Drosophila syncytium, actomyosin networks colocalize with microtubules, and inhibiting microtubule function in this context leads to a loss of cortical actomyosin function (Foe et al., 2000). In addition, dual color imaging in laser-wounded Xenopus oocytes confirms that there is colocalization and cotransport between actomyosin networks and microtubules at the wound border (Mandato and Bement, 2003). Perturbing microtubules causes a failure to properly recruit Myosin II and the Arp2/3 complex at the wound border. Intriguingly, after wounding, actomyosin flows also drive the recruitment of microtubules near the wound, demonstrating a codependency between the cytoskeletal networks.
As mentioned briefly above, apical polarity proteins such as the Par and Crumbs complexes also provide important spatiotemporal information that guides actomyosin assembly and function, although the effects of disrupting these complexes vary greatly. For example, Baz/Par-6/aPKC complex function is essential for the anchorage and accumulation of apical myosin during Drosophila oogenesis (Wang and Riechmann, 2007), whereas Par-3 (Bazooka in Drosophila) acts antagonistically during germband extension to constrain Myosin II function to anterior-posterior (AP), and not dorsal-ventral (DV), interfaces (Simões et al., 2010). Interestingly, different PAR complex components have been implicated in distinct roles in regulating contractile pulse properties during the apical constriction of amnioserosa cells: Par-3 acts to elongate the period of contractile steps, whereas Par-6 and aPKC promote the rest duration between pulses (David et al., 2010). Similarly, the Crumbs complex can both promote and inhibit actomyosin function in different tissue contexts. During the formation of the Drosophila salivary gland, Crumbs and aPKC coordinately control formation of a supracellular actomyosin cable by limiting Rok recruitment to regions of low Crumbs activity (Röper, 2012). Negative regulation of actomyosin networks by Crumbs has also been observed during dorsal closure (Flores-Benitez and Knust, 2015). However, as discussed previously, Crumbs and Par-3/Bazooka can interact with p114RhoGEF/Cysts, a Drosophila RhoGEF, to promote junctional myosin accumulation through Rho1 (Silver et al., 2019). Thus, the proteins that provide an internal cellular compass that enables epithelia to maintain distinct apical and basolateral identities are also used to direct the asymmetric accumulation of contractile actomyosin networks.
Taken together, it is not surprising that, given the variety of tissue processes Myosin II is necessary for, there are a large number of regulatory mechanisms by which Myosin II activity is controlled.
Actomyosin pulsatility: from initiation to cycling
As we have highlighted above, actomyosin activity is regulated by Rho1/RhoA activity. But how do pulsatile regimes emerge? It is interesting to note that pulsing has many characteristics of a stochastic process, and lacks tightly-controlled periodicities in many systems (Rauzi et al., 2010; Sokolow et al., 2012; Xie and Martin, 2015; Miao et al., 2019). Indeed, individual actomyosin pulses have highly variable amplitudes and periods. Recent work has indicated that pulsing requires a combination of positive and negative inputs on the activation signal (Rho1/RhoA), as well as Myosin dephosphorylation and the turnover of filamentous actin networks (Levayer and Lecuit, 2012; Coravos and Martin, 2016; Blanchard et al., 2018). The localization of Rho kinase shows pulse dynamics that are similar to those exhibited by Myosin II, and phosphomimetic Myosin II mutants fail to localize and pulse properly, suggesting that upstream cycles of activation/deactivation are essential for determining pulse dynamics, although spatiotemporal limitations of phosphomimetic approaches can complicate potential interpretations (Munjal et al., 2015; Vasquez et al., 2014; Kasza et al., 2014). Interestingly, medial and junctional Myosin II pools have been observed to have regulatory RhoGEFs that specialize in promoting their populations, with RhoGEF2 driving medial Rho activity and Myosin II accumulation, and p114RhoGEF/Cysts driving junctional Myosin II pools (Mason et al., 2016; De Las Bayonas et al., 2019). Other work has observed medial Myosin II flows that appear to precede and join with junctional or vertex Myosin populations (Rauzi et al., 2010; Vanderleest et al., 2018). Negative regulation of Rho1 activity by GTPase-activating proteins (GAPs) is also essential for both the proper localization and cycling of Rho (Mason et al., 2016; Michaux et al., 2018). For example, in the invaginating Drosophila mesoderm, the Rho GAP Cumberland-GAP (C-GAP) restricts RhoA pathway activity spatially to a central position in the apical cortex, and actomyosin pulsing is lost in the absence of C-GAP activity (Mason et al., 2016).
In addition to the above signaling components, it should be noted that the mechanical effect of contraction may, in turn, regulate pulsing. For example, contraction-based advection may lead to concentration of actomyosin filaments and their regulators, and the potential mechanosensitivity of Myosin II may favor greater activity under load-based conditions (Fernandez-Gonzalez et al., 2009; Hundt et al., 2016), whereas dispersion and F-actin turnover may limit pulse length (Dierkes et al., 2014; Munjal et al., 2015). F-actin turnover is also essential for maintaining the connection between contractile actomyosin networks and their anchorage to cell-cell E-cadherin adherens junctions (Jodoin et al., 2015). In addition, Myosin II phosphatase function is required for pulsing (Munjal et al., 2015; Vasquez et al., 2014). It is interesting that plasma membrane-associated compartmental localization of Rab35 and its associated GEF Sbf appears to precede Myosin II accumulation by ∼60 s during both apical constriction and cell intercalation, suggesting that a higher-order coordination between actomyosin and trafficking networks may exist (Jewett et al., 2017; Miao et al., 2019). Indeed, disrupting Rab35 or Sbf leads to compromised Myosin II activity, although the mechanism coordinating these activities remains to be identified.
Ratcheting mechanisms that drive progressive changes in cell shapes
If actomyosin networks are continually being remodeled, with cycles of activation followed by disassembly, how does such a cyclic system give rise to progressive changes in cell shape or behavior? To obtain lasting changes from such systems, there is a requirement for a ratcheting mechanism that extracts unidirectional changes from contractile cycles. This has been an area of intense focus over the last few years, and several intriguing themes and mechanisms have emerged (Fig. 4). These involve harnessing the continual remodeling of cortical elastic forces of the cell, as well as adhesive engagement forces, to permit topological reshaping under certain compatible time frames.
Fig. 4.

Ratcheting mechanisms that direct cell-shape changes. (A) Remodeling of the actin cortex enforces new cell-shape changes after Myosin II-driven cell-shape changes. During this process, primary actin filaments (red) are disassembled and then re-form (blue) to take on the shape of the new cell topology. (B) Endocytic pathways can direct the uptake of contracted cell surfaces and adhesion proteins. During cell oscillations (arrows), Formin and Myosin II function mediate the clustering of E-cadherin complexes. Tubulated Rab35 compartments then form to take up plasma membrane and E-cadherin complexes, causing the shortening of cell interfaces. Myosin II and Arp2/3 function aid Clathrin-mediated endocytosis to direct the termination of Rab35 compartments and deliver excess plasma membrane to endosomes, leading to processive interface contraction. (C) Ratcheted vertex sliding is responsible for interface contraction. During cell oscillations, tricellular vertices experience asymmetric forces, leading to their sliding along the cell periphery, resulting in the shortening of cell interfaces.
For example, one study has found that pulses during Drosophila cell intercalation need to have a certain duration for cell-shape changes to become irreversible during a pulse (Clément et al., 2017). These longer pulses are congruent with the time frame for actin remodeling; this remodeling allows the short-term cortical elasticity to be overcome and a longer-term viscous response is generated in which the cell can relax into the new cell shape. This relaxation is enforced by the assembly of cortical actin that conforms to the new cellular configuration (Fig. 4A). Indeed, artificially stabilizing actin filaments with small molecule treatments leads to a greater elastic response and a higher degree of reversible cell-shape changes, whereas using an optical laser trap to exert pulling forces for longer periods of time enhances ratcheting and irreversibility (Clément et al., 2017). Myosin II activity has been found to aid the fluidization of actin filaments (Le Goff et al., 2002; Humphrey et al., 2002), and thus Myosin activity may also directly aid the mobilization and turnover of pre-existing actin filaments that is required for this elastic relaxation. Interestingly, a recent study in C. elegans also found that the ability to enforce a newly remodeled actin stiffness through the function of a bundling F-actin cross-linker (α-spectrin SPC-1) is essential to permit tissue elongation (Lardennois et al., 2019). The optogenetic modulation of myosin activation dynamics, mentioned above, also indicated that there is a minimum time (∼5 min for Caco-2 cells) that is required to obtain irreversible interface length changes (Cavanaugh et al., 2020). Combined, these results suggest that the cortical elasticity of a starting cellular configuration must be disassembled before a new supporting framework can be built in the shape of a motor-driven cell deformation.
Other work has examined how direct remodeling of the cell surface and, presumably, the adhesive proteins embedded in the plasma membrane, can provide a cellular ratcheting activity. For example, oscillations in cell area as well as interface lengths produce cycles of high and low tension. These are periods during which trafficking networks can use these areas of slackened cell surface to internalize plasma membrane and integral proteins (Fig. 4B). This in turn would decrease adhesive complexes on particular cell surfaces and thereby prevent interfaces from rebounding in length when contractile force generation terminates. Such an increase in endocytic rates has been observed at contractile surfaces during cell intercalation (Levayer et al., 2011; Jewett et al., 2017), and it has also been shown that efficient endocytosis is a requirement for apical constriction in both Xenopus and Drosophila (Lee and Harland, 2010; Sumi et al., 2018; Miao et al., 2019). Specific filamentous actin networks have been implicated in directing individual aspects of this internalization. Whereas the Arp2/3-activating protein Scar is required to mediate increased endocytosis at contractile surfaces, the formin Diaphanous functions to cluster E-cadherin adhesive complexes for subsequent internalization (Levayer et al., 2011). In addition, when endocytosis through highly-tubulated Rab35 compartments is disrupted, interfaces show a lack of cell ratcheting and exhibit a ‘wobble’ phenotype in which cell interfaces have rapid reversing failures and a loss of processive changes in cell shape (Jewett et al., 2017). Disrupting early endocytic events through the expression of dominant negative Rab5 also produces ruffled cell interfaces in the amnioserosa during dorsal closure, indicative of aberrant domains of lower plasma membrane tension (Sumi et al., 2018). These results suggest that cell surfaces can be destabilized through the joint function of actomyosin activity and endocytic uptake of adhesion proteins and/or surface areas. A further interesting feature of these studies was the absence of paired endocytic events at juxtaposed plasma membranes (Jewett et al., 2017). This demonstrates that adhesion protein uptake likely occurs from only one cell surface, and is consistent with the generation of interfacial shear forces at contracting surfaces (Jewett et al., 2017; Kale et al., 2018).
Cell-shape changes can also be driven by a ratcheted sliding of tricellular vertices (Curran et al., 2017; Vanderleest et al., 2018). In these sliding events, radially directed pulses pull on cell vertices to cause their displacement along the cell periphery (Fig. 4C). These medial Myosin II pulses then drive the enrichment of E-cadherin adhesive complexes at tricellular vertices by flowing from cell apices into cell vertices. The vertex enrichment of E-cadherin, in turn, inhibits the back-sliding of vertices after pulse termination and causes a ratcheted change in cell dimensions. Vertex E-cadherin levels are particularly sensitive to changes in endocytic rates (Vanderleest et al., 2018), and the force balance at tricellular junctions is dependent on the summed local tensions that are influenced by the actin, adhesion and endocytic pathways discussed above. However, it will be important to explore how cell vertex function integrates and scales with the various medial and interfacial force pathways that act in an epithelial sheet. Finally, ratcheting can also emerge through the combinatorial activity of supracellular cables and actomyosin pulses. During the zippering up of the dorsal embryonic surface during Drosophila dorsal closure, the amnioserosa is surrounded by an encroaching epithelium that has a substantial purse-string cable that begins to constrict inwards (Franke et al., 2005). As the amnioserosa undergoes cyclic pulses of actomyosin contraction, the surrounding purse-string ratchets inwards, preventing amnioserosa cells from expanding out to their original size once a myosin pulse terminates (Solon et al., 2009). In this manner, the supracellular cable helps guide a processive advancement of the epithelium to eventually close over the apoptotic amnioserosa. On a smaller scale, the enrichment of Myosin II at cell junctions was found to aid the stabilization of pulsed apically-driven deformations during cell intercalation (Munjal et al., 2015).
Taken together, the above findings suggest that cell ratcheting in pulsatile systems involves a combination of cortical remodeling, endocytic regulation and coordination between distinct actomyosin populations and cell geometries.
Force transmission within cells and across tissues
There are two primary conditions for force transmission within tissues: (1) a cell must be able to generate a force that can propagate through a cell without being lost through viscous dissipation effects; and (2) cells within a tissue must possess a connectivity that permits the coupling of generated forces by adhesive contacts that transmit forces through adjacent cytoskeletal networks. As mentioned above, when the ability of actin networks to remodel is compromised, cytoskeletal connectivity can be lost (Jodoin et al., 2015; Mason et al., 2016). This is interesting, and would at first appear paradoxical, as it is the disruption of pathways that lead to actin turnover that cause breaks in microfilament networks. However, this illustrates that the ability to continually tear down and then rebuild actin structures that match the evolving changes in intracellular, as well as cell-adjacent, tensions is a crucial property that leads to tissue connectivity. It also appears that dedicated actin networks may be involved in building the cortical actin required for tissue force transmission. Recent work has discovered a formin – Frl (Fmnl) – that acts to nucleate a persistent actin network: if Frl function is compromised, cells become more deformable but do not show efficient transmission across entire cell areas and into adjacent cells (Dehapiot et al., 2019). Intriguingly, disruption of the persistent actin network leads to an upregulation of medial pulsatile networks, suggesting an antagonistic interplay between these entangled cortical pools. In addition, the function of adhesion complexes is essential for coupling actin cytoskeletons across cells and tissues. Indeed, disrupting the function of cadherins, their associated α-catenins/β-catenins or potential actin-connecting proteins such as Afadin, deeply disrupts tissue-level transmission of deforming forces (Larue et al., 1994; Tepass et al., 1996; Costa et al., 1998; Pokutta et al., 2002; Sawyer et al., 2009, 2011; Bazellières et al., 2015; Chanet et al., 2017).
Individual cellular behaviors are also affected by the cumulative transmission of forces within a tissue. For example, during apical constriction in the Drosophila mesoderm, cells constrict their apical surfaces anisotropically, and this is also observable at the level of the actomyosin networks (Chanet et al., 2017). However, this asymmetric constriction is a result of the rectangular array of cells that is oriented along the AP axis and thus produces a greater AP-directed tension. If tissue dimensions are altered to produce a more isotropic tissue, then apical constriction, as well as the underlying actomyosin network, also becomes isotropic (Chanet et al., 2017). Interestingly, optogenetic manipulation of RhoGEF2 function has shown that almost any AP or DV region in the early fly embryo is capable of forming an ingressing furrow, and that this is simply dependent on the dimensions of Rho activation (Izquierdo et al., 2018). Tissue-level tension, as applied by microaspiration or tissue stretching, can also lead to enhanced recruitment of Myosin II activity (Fernandez-Gonzalez et al., 2009; Duda et al., 2019). Furthermore, tissue movements can be harnessed to drive the propagation of biomechanical properties. For example, during invagination of the Drosophila posterior midgut, an anterior-propagating wave of apical constriction and Myosin II activity occurs (Bailles et al., 2019). Remarkably, this wave is induced owing to the pushing of cells into the eggshell membrane caused by the movement of the invaginating tissue. As cells are pushed into the eggshell, an activation of integrin-mediated adhesion leads to Myosin II recruitment (Bailles et al., 2019; Münster et al., 2019). Thus, the transmission of individual cellular actomyosin forces leads to tissue-level effects that can, in turn, feedback on cortical actomyosin networks.
Conclusions
The last decade has been an exceptional time for advancing our understanding of actomyosin function, but key questions still remain to be answered. For example, how do actin meshworks of randomly oriented fibers direct contractile force generation? Part of the answer to this may lie in a non-apparent underlying organization. Indeed, recent studies of apical constriction have demonstrated a sarcomere-like organization in which barbed actin ends are enriched near junctions and Myosin II is present in the central/medial region of cell apices (Coravos and Martin, 2016). However, similar organizations of actin have not (yet) been found in other contexts or during other morphogenetic processes. Other questions revolve around the multitude of lipid compartments and topological complexities that are present at the plasma membrane – how do these influence the construction and physical linkages of productive actomyosin networks? Recent work has observed that the apical surface becomes deeply convoluted when membrane trafficking networks are disrupted, and this produces heterogeneities in actomyosin network construction/organization (Miao et al., 2019). Indeed, mild blebbing can be observed even in wild-type cells (Leptin and Grunewald, 1990; Sweeton et al., 1991), suggesting that the pathways that remodel the apical cell surface must actively deal with how contractile forces deform the plasma membrane. If this is the case, how are these processes coordinated? The plasma membrane often shows constrained or corralled diffusion of particular components, so could this provide crucial landmarks for efficient network construction? As noted above, GPCRs cluster in small invaginations in the plasma membrane, which already suggests higher-order organization of signaling complexes (Jha et al., 2018). Could this organization translate into the anisotropic activities that are required for polarized processes, such as cell intercalation and apical constriction? If this is the case, there should be some informational relationship between the Toll receptors that mediate planar polarity and GPCR function (Paré et al., 2014). Interestingly, a recent report indicates that the aGPCR family member Cirl can bind to Toll-8 (TLR8) and direct Myosin II polarization to cell interfaces (Lavalou et al., 2020 preprint).
New technologies may also aid in advancing our understanding of the dynamics of actomyosin assembly, pulsing and ratcheting. Lattice light sheet microscopy should permit deeper tissue penetration and a reduction in noise/background, whereas total internal reflection fluorescence-based single-molecule imaging would allow the true cortical and plasma membrane diffusion and recruitment rates of cytoskeletal and regulatory components to be determined. Regardless, actomyosin networks will clearly remain front and center in our attempts to understand the force-generating events that drive developmental dynamics and the mechanisms of morphogenesis.
Acknowledgements
We thank members of the Blankenship labs for critical reading and constructive comments on the manuscript. We apologize to colleagues whose relevant work was not cited here because of space limitations and decisions on manuscript organization and scope.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Funding
This work was supported by grants from the National Institute of General Medical Sciences (National Institutes of Health; R01GM127447 and R15GM126422 to J.T.B.). Deposited in PMC for release after 12 months.
References
- Abreu-Blanco M. T., Verboon J. M., Liu R., Watts J. J. and Parkhurst S. M. (2012). Drosophila embryos close epithelial wounds using a combination of cellular protrusions and an actomyosin purse string. J. Cell Sci. 125, 5984-5997. 10.1242/jcs.109066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amano M., Ito M., Kimura K., Fukata Y., Chihara K., Nakano T., Matsuura Y. and Kaibuchi K. (1996). Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase). J. Biol. Chem. 271, 20246-20249. 10.1074/jbc.271.34.20246 [DOI] [PubMed] [Google Scholar]
- An Y., Xue G., Shaobo Y., Mingxi D., Zhou X., Yu W., Ishibashi T., Zhang L. and Yan Y. (2017). Apical constriction is driven by a pulsatile apical myosin network in delaminating Drosophila neuroblasts. Development 144, 2153-2164. 10.1242/dev.150763 [DOI] [PubMed] [Google Scholar]
- Antunes M., Pereira T., Cordeiro J. V., Almeida L. and Jacinto A. (2013). Coordinated waves of actomyosin flow and apical cell constriction immediately after wounding. J. Cell Biol. 202, 365-379. 10.1083/jcb.201211039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailles A., Collinet C., Philippe J.-M., Lenne P.-F., Munro E. and Lecuit T. (2019). Genetic induction and mechanochemical propagation of a morphogenetic wave. Nature 572, 467-473. 10.1038/s41586-019-1492-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bazellières E., Conte V., Elosegui-Artola A., Serra-Picamal X., Bintanel-Morcillo M., Roca-Cusachs P., Muñoz J. J., Sales-Pardo M., Guimerà R. and Trepat X. (2015). Control of cell–cell forces and collective cell dynamics by the intercellular adhesome. Nat. Cell Biol. 17, 409-420. 10.1038/ncb3135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrndt M., Salbreux G., Campinho P., Hauschild R., Oswald F., Roensch J., Grill S. W. and Heisenberg C.-P. (2012). Forces driving epithelial spreading in zebrafish gastrulation. Science 338, 257-260. 10.1126/science.1224143 [DOI] [PubMed] [Google Scholar]
- Betapudi V., Rai V., Beach J. R. and Egelhoff T. (2010). Novel regulation and dynamics of myosin II activation during epidermal wound responses. Exp. Cell Res. 316, 980-991. 10.1016/j.yexcr.2010.01.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanchard G. B., Murugesu S., Adams R. J., Martinez-Arias A. and Gorfinkiel N. (2010). Cytoskeletal dynamics and supracellular organisation of cell shape fluctuations during dorsal closure. Development 137, 2743-2752. 10.1242/dev.045872 [DOI] [PubMed] [Google Scholar]
- Blanchard G. B., Étienne J. and Gorfinkiel N. (2018). From pulsatile apicomedial contractility to effective epithelial mechanics. Curr. Opin. Genet. Dev. 51, 78-87. 10.1016/j.gde.2018.07.004 [DOI] [PubMed] [Google Scholar]
- Blankenfeldt W., Thomä N. H., Wray J. S., Gautel M. and Schlichting I. (2006). Crystal structures of human cardiac β-myosin II S2-Δ provide insight into the functional role of the S2 subfragment. Proc. Natl Acad. Sci. USA 103, 17713-17717. 10.1073/pnas.0606741103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blankenship J. T., Backovic S. T., Sanny J. S. P., Weitz O. and Zallen J. A. (2006). Multicellular rosette formation links planar cell polarity to tissue morphogenesis. Dev. Cell 11, 459-470. 10.1016/j.devcel.2006.09.007 [DOI] [PubMed] [Google Scholar]
- Blaser H., Reichman-Fried M., Castanon I., Dumstrei K., Marlow F. L., Kawakami K., Solnica-Krezel L., Heisenberg C.-P. and Raz E. (2006). Migration of zebrafish primordial germ cells: a role for myosin contraction and cytoplasmic flow. Dev. Cell 11, 613-627. 10.1016/j.devcel.2006.09.023 [DOI] [PubMed] [Google Scholar]
- Bresnick A. R. (1999). Molecular mechanisms of nonmuscle myosin-II regulation. Curr. Opin. Cell Biol. 11, 26-33. 10.1016/S0955-0674(99)80004-0 [DOI] [PubMed] [Google Scholar]
- Brock J., Midwinter K., Lewis J. and Martin P. (1996). Healing of incisional wounds in the embryonic chick wing bud: characterization of the actin purse-string and demonstration of a requirement for Rho activation. J. Cell Biol. 135, 1097-1107. 10.1083/jcb.135.4.1097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canman J. C. and Bement W. M. (1997). Microtubules suppress actomyosin-based cortical flow in Xenopus oocytes. J. Cell Sci. 110, 1907-1917. [DOI] [PubMed] [Google Scholar]
- Cavanaugh K. E., Staddon M. F., Munro E., Banerjee S. and Gardel M. L. (2020). RhoA mediates epithelial cell shape changes via mechanosensitive endocytosis. Dev. Cell 52, 152-166.e5. 10.1016/j.devcel.2019.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chanet S., Miller C. J., Vaishnav E. D., Ermentrout B., Davidson L. A. and Martin A. C. (2017). Actomyosin meshwork mechanosensing enables tissue shape to orient cell force. Nat. Commun. 8, 15014 10.1038/ncomms15014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X.-Q., Tan I., Leung T. and Lim L. (1999). The myotonic dystrophy kinase-related Cdc42-binding kinase is involved in the regulation of neurite outgrowth in PC12 cells. J. Biol. Chem. 274, 19901-19905. 10.1074/jbc.274.28.19901 [DOI] [PubMed] [Google Scholar]
- Clément R., Dehapiot B., Collinet C., Lecuit T. and Lenne P.-F. (2017). Viscoelastic dissipation stabilizes cell shape changes during tissue morphogenesis. Curr. Biol. 27, 3132-3142.e4. 10.1016/j.cub.2017.09.005 [DOI] [PubMed] [Google Scholar]
- Collinet C., Rauzi M., Lenne P.-F. and Lecuit T. (2015). Local and tissue-scale forces drive oriented junction growth during tissue extension. Nat. Cell Biol. 17, 1247-1258. 10.1038/ncb3226 [DOI] [PubMed] [Google Scholar]
- Coravos J. S. and Martin A. C. (2016). Apical sarcomere-like actomyosin contracts nonmuscle Drosophila epithelial cells. Dev. Cell 39, 346-358. 10.1016/j.devcel.2016.09.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costa M., Raich W., Agbunag C., Leung B., Hardin J. and Priess J. R. (1998). A putative catenin–cadherin system mediates morphogenesis of the Caenorhabditis elegans embryo. J. Cell Biol. 141, 297-308. 10.1083/jcb.141.1.297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig R., Smith R. and Kendrick-Jones J. (1983). Light-chain phosphorylation controls the conformation of vertebrate non-muscle and smooth muscle myosin molecules. Nature 302, 436-439. 10.1038/302436a0 [DOI] [PubMed] [Google Scholar]
- Curran S., Strandkvist C., Bathmann J., de Gennes M., Kabla A., Salbreux G. and Baum B. (2017). Myosin II controls junction fluctuations to guide epithelial tissue ordering. Dev. Cell 43, 480-492.e6. 10.1016/j.devcel.2017.09.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- David D. J. V., Tishkina A. and Harris T. J. C. (2010). The PAR complex regulates pulsed actomyosin contractions during amnioserosa apical constriction in Drosophila. Development 137, 1645-1655. 10.1242/dev.044107 [DOI] [PubMed] [Google Scholar]
- Davidson L. A., Ezin A. M. and Keller R. (2002). Embryonic wound healing by apical contraction and ingression in Xenopus laevis. Cell Motil. Cytoskelet. 53, 163-176. 10.1002/cm.10070 [DOI] [PubMed] [Google Scholar]
- Dawes-Hoang R. E., Parmar K. M., Christiansen A. E., Phelps C. B., Brand A. H. and Wieschaus E. F. (2005). Folded gastrulation, cell shape change and the control of myosin localization. Development 132, 4165-4178. 10.1242/dev.01938 [DOI] [PubMed] [Google Scholar]
- De Las Bayonas A. G., Philippe J.-M., Lellouch A. C. and Lecuit T. (2019). Distinct RhoGEFs activate apical and junctional contractility under control of G proteins during epithelial morphogenesis. Curr. Biol. 29, 3370-3385.e7. 10.1016/j.cub.2019.08.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dehapiot B., Clément R., Alégot H., Gazsó-Gerhát G., Philippe J.-M. and Lecuit T. (2019). Assembly of a persistent apical actin network by the formin Frl/Fmnl tunes epithelial cell deformability. Nat. Cell Biol. 22, 791-802. 10.1038/s41556-020-0524-x [DOI] [PubMed] [Google Scholar]
- Di Cunto F., Calautti E., Hsiao J., Ong L., Topley G., Turco E. and Dotto G. P. (1998). Citron Rho-interacting kinase, a novel tissue-specific ser/thr kinase encompassing the Rho-Rac-binding protein Citron. J. Biol. Chem. 273, 29706-29711. 10.1074/jbc.273.45.29706 [DOI] [PubMed] [Google Scholar]
- Dierkes K., Sumi A., Solon J. and Salbreux G. (2014). Spontaneous oscillations of elastic contractile materials with turnover. Phys. Rev. Lett. 113, 148102 10.1103/PhysRevLett.113.148102 [DOI] [PubMed] [Google Scholar]
- Duda M., Kirkland N. J., Khalilgharibi N., Tozluoglu M., Yuen A. C., Carpi N., Bove A., Piel M., Charras G., Baum B. et al. (2019). Polarization of myosin II refines tissue material properties to buffer mechanical stress. Dev. Cell 48, 245-260.e7. 10.1016/j.devcel.2018.12.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrlich M., Boll W., Van Oijen A., Hariharan R., Chandran K., Nibert M. L. and Kirchhausen T. (2004). Endocytosis by random initiation and stabilization of clathrin-coated pits. Cell 118, 591-605. 10.1016/j.cell.2004.08.017 [DOI] [PubMed] [Google Scholar]
- Fernandez-Gonzalez R. and Zallen J. A. (2011). Oscillatory behaviors and hierarchical assembly of contractile structures in intercalating cells. Phys. Biol. 8, 045005 10.1088/1478-3975/8/4/045005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Gonzalez R. and Zallen J. A. (2013). Wounded cells drive rapid epidermal repair in the early Drosophila embryo. Mol. Biol. Cell 24, 3227-3237. 10.1091/mbc.e13-05-0228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Gonzalez R., Simoes S. D. M., Röper J.-C., Eaton S. and Zallen J. A. (2009). Myosin II dynamics are regulated by tension in intercalating cells. Dev. Cell 17, 736-743. 10.1016/j.devcel.2009.09.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flores-Benitez D. and Knust E. (2015). Crumbs is an essential regulator of cytoskeletal dynamics and cell-cell adhesion during dorsal closure in Drosophila. eLife 4, e07398 10.7554/eLife.07398 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foe V. E., Field C. M. and Odell G. M. (2000). Microtubules and mitotic cycle phase modulate spatiotemporal distributions of F-actin and myosin II in Drosophila syncytial blastoderm embryos. Development 127, 1767-1787. [DOI] [PubMed] [Google Scholar]
- Franke J. D., Montague R. A. and Kiehart D. P. (2005). Nonmuscle myosin II generates forces that transmit tension and drive contraction in multiple tissues during dorsal closure. Curr. Biol. 15, 2208-2221. 10.1016/j.cub.2005.11.064 [DOI] [PubMed] [Google Scholar]
- Hartman M. A. and Spudich J. A. (2012). The myosin superfamily at a glance. J. Cell Sci. 125, 1627-1632. 10.1242/jcs.094300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartshorne D. J., Ito M. and Erdo F. (1998). Myosin light chain phosphatase: subunit composition, interactions and regulation. J. Muscle Res. Cell Motil. 19, 325 10.1023/A:1005385302064 [DOI] [PubMed] [Google Scholar]
- He L., Wang X., Tang H. L. and Montell D. J. (2010). Tissue elongation requires oscillating contractions of a basal actomyosin network. Nat. Cell Biol. 12, 1133-1142. 10.1038/ncb2124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heer N. C. and Martin A. C. (2017). Tension, contraction and tissue morphogenesis. Development 144, 4249-4260. 10.1242/dev.151282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heissler S. M. and Manstein D. J. (2013). Nonmuscle myosin-2: mix and match. Cell. Mol. Life Sci. 70, 1-21. 10.1007/s00018-012-1002-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphrey D., Duggan C., Saha D., Smith D. and Käs J. (2002). Active fluidization of polymer networks through molecular motors. Nature 416, 413-416. 10.1038/416413a [DOI] [PubMed] [Google Scholar]
- Hundt N., Steffen W., Pathan-Chhatbar S., Taft M. H. and Manstein D. J. (2016). Load-dependent modulation of non-muscle myosin-2A function by tropomyosin 4.2. Sci. Rep. 6, 20554 10.1038/srep20554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikebe M. and Hartshorne D. J. (1985). Phosphorylation of smooth muscle myosin at two distinct sites by myosin light chain kinase. J. Biol. Chem. 260, 10027-10031. [PubMed] [Google Scholar]
- Izquierdo E., Quinkler T. and De Renzis S. (2018). Guided morphogenesis through optogenetic activation of Rho signalling during early Drosophila embryogenesis. Nat. Commun. 9, 2366 10.1038/s41467-018-04754-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jewett C. E., Vanderleest T. E., Miao H., Xie Y., Madhu R., Loerke D. and Blankenship J. T. (2017). Planar polarized Rab35 functions as an oscillatory ratchet during cell intercalation in the Drosophila epithelium. Nat. Commun. 8, 476 10.1038/s41467-017-00553-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jha A., van Zanten T. S., Philippe J.-M., Mayor S. and Lecuit T. (2018). Quantitative control of GPCR organization and signaling by endocytosis in epithelial morphogenesis. Curr. Biol. 28, 1570-1584.e6. 10.1016/j.cub.2018.03.068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jodoin J. N., Coravos J. S., Chanet S., Vasquez C. G., Tworoger M., Kingston E. R., Perkins L. A., Perrimon N. and Martin A. C. (2015). Stable force balance between epithelial cells arises from F-actin turnover. Dev. Cell 35, 685-697. 10.1016/j.devcel.2015.11.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung H. S., Komatsu S., Ikebe M. and Craig R. (2008). Head–head and head–tail interaction: a general mechanism for switching off myosin II activity in cells. Mol. Biol. Cell 19, 3234-3242. 10.1091/mbc.e08-02-0206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kale G. R., Yang X., Philippe J. M., Mani M., Lenne P. F. and Lecuit T. (2018). Distinct contributions of tensile and shear stress on E-cadherin levels during morphogenesis. Nat. Commun. 9, 1-16. 10.1038/s41467-018-07448-8 [DOI] [PMC free article] [PubMed]
- Kasza K. E., Farrell D. L. and Zallen J. A. (2014). Spatiotemporal control of epithelial remodeling by regulated myosin phosphorylation. Proc. Natl Acad. Sci. USA111, 11732-11737. 10.1073/pnas.1400520111 [DOI]
- Kerridge S., Munjal A., Philippe J.-M., Jha A., De Las Bayonas A. G., Saurin A. J. and Lecuit T. (2016). Modular activation of Rho1 by GPCR signalling imparts polarized myosin II activation during morphogenesis. Nat. Cell Biol. 18, 261-270. 10.1038/ncb3302 [DOI] [PubMed] [Google Scholar]
- Kiehart D. P., Galbraith C. G., Edwards K. A., Rickoll W. L. and Montague R. A. (2000). Multiple forces contribute to cell sheet morphogenesis for dorsal closure in Drosophila. J. Cell Biol. 149, 471-490. 10.1083/jcb.149.2.471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H. Y. and Davidson L. A. (2011). Punctuated actin contractions during convergent extension and their permissive regulation by the non-canonical Wnt-signaling pathway. J. Cell Sci. 124, 635-646. 10.1242/jcs.067579 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura K., Ito M., Amano M., Chihara K., Fukata Y., Nakafuku M., Yamamori B., Feng J., Nakano T., Okawa K. et al. (1996). Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho-kinase). Science 273, 245-248. 10.1126/science.273.5272.245 [DOI] [PubMed] [Google Scholar]
- Kölsch V., Seher T., Fernandez-Ballester G. J., Serrano L. and Leptin M. (2007). Control of Drosophila gastrulation by apical localization of adherens junctions and RhoGEF2. Science 315, 384-386. 10.1126/science.1134833 [DOI] [PubMed] [Google Scholar]
- Lardennois A., Pásti G., Ferraro T., Llense F., Mahou P., Pontabry J., Rodriguez D., Kim S., Ono S., Beaurepaire E. et al. (2019). An actin-based viscoplastic lock ensures progressive body-axis elongation. Nature 573, 266-270. 10.1038/s41586-019-1509-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larue L., Ohsugi M., Hirchenhain J. and Kemler R. (1994). E-cadherin null mutant embryos fail to form a trophectoderm epithelium. Proc. Natl Acad. Sci. USA 91, 8263-8267. 10.1073/pnas.91.17.8263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavalou J., Mao Q., Harmansa S., Kerridge S., Lellouch A. C., Philippe J.-M., Audebert S., Camoin L. and Lecuit T. (2020). Formation of mechanical interfaces by self-organized Toll-8/Cirl GPCR asymmetry. bioRxiv. 10.1101/2020.03.16.993758 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Goff L., Hallatschek O., Frey E. and Amblard F. (2002). Tracer studies on f-actin fluctuations. Phys. Rev. Lett. 89, 258101 10.1103/physrevlett.89.258101 [DOI] [PubMed] [Google Scholar]
- Lee J.-Y. and Harland R. M. (2010). Endocytosis is required for efficient apical constriction during Xenopus gastrulation. Curr. Biol. 20, 253-258. 10.1016/j.cub.2009.12.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leptin M. and Grunewald B. (1990). Cell shape changes during gastrulation in Drosophila. Development 110, 73-84. [DOI] [PubMed] [Google Scholar]
- Levayer R. and Lecuit T. (2012). Biomechanical regulation of contractility: spatial control and dynamics. Trends Cell Biol. 22, 61-81. 10.1016/j.tcb.2011.10.001 [DOI] [PubMed] [Google Scholar]
- Levayer R., Pelissier-Monier A. and Lecuit T. (2011). Spatial regulation of Dia and Myosin-II by RhoGEF2 controls initiation of E-cadherin endocytosis during epithelial morphogenesis. Nat. Cell Biol. 13, 529-540. 10.1038/ncb2224 [DOI] [PubMed] [Google Scholar]
- Loerke D., Mettlen M., Yarar D., Jaqaman K., Jaqaman H., Danuser G. and Schmid S. L. (2009). Cargo and dynamin regulate clathrin-coated pit maturation. PLoS Biol. 7, e57 10.1371/journal.pbio.1000057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maître J.-L., Niwayama R., Turlier H., Nédélec F. and Hiiragi T. (2015). Pulsatile cell-autonomous contractility drives compaction in the mouse embryo. Nat. Cell Biol. 17, 849-855. 10.1038/ncb3185 [DOI] [PubMed] [Google Scholar]
- Mandato C. A. and Bement W. M. (2003). Actomyosin transports microtubules and microtubules control actomyosin recruitment during Xenopus oocyte wound healing. Curr. Biol. 13, 1096-1105. 10.1016/S0960-9822(03)00420-2 [DOI] [PubMed] [Google Scholar]
- Manning A. J., Peters K. A., Peifer M. and Rogers S. L. (2013). Regulation of epithelial morphogenesis by the G protein–coupled receptor mist and its ligand Fog. Sci. Signal. 6, ra98 10.1126/scisignal.2004427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin P. and Lewis J. (1992). Actin cables and epidermal movement in embryonic wound healing. Nature 360, 179-183. 10.1038/360179a0 [DOI] [PubMed] [Google Scholar]
- Martin A. C., Kaschube M. and Wieschaus E. F. (2009). Pulsed contractions of an actin–myosin network drive apical constriction. Nature 457, 495-499. 10.1038/nature07522 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason F. M., Xie S., Vasquez C. G., Tworoger M. and Martin A. C. (2016). RhoA GTPase inhibition organizes contraction during epithelial morphogenesis. J. Cell Biol. 214, 603-617. 10.1083/jcb.201603077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miao H., Vanderleest T. E., Jewett C. E., Loerke D. and Blankenship J. T. (2019). Cell ratcheting through the Sbf RabGEF directs force balancing and stepped apical constriction. J. Cell Biol. 218, 3845-3860. 10.1083/jcb.201905082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michaux J. B., Robin F. B., McFadden W. M. and Munro E. M. (2018). Excitable RhoA dynamics drive pulsed contractions in the early C. elegans embryo. J. Cell Biol. 217, 4230-4252. 10.1083/jcb.201806161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michel M. and Dahmann C. (2020). Tissue mechanical properties modulate cell extrusion in the Drosophila abdominal epidermis. Development 147, dev179606 10.1242/dev.179606 [DOI] [PubMed] [Google Scholar]
- Morize P., Christiansen A. E., Costa M., Parks S. and Wieschaus E. (1998). Hyperactivation of the folded gastrulation pathway induces specific cell shape changes. Development 125, 589-597. [DOI] [PubMed] [Google Scholar]
- Munjal A., Philippe J.-M., Munro E. and Lecuit T. (2015). A self-organized biomechanical network drives shape changes during tissue morphogenesis. Nature 524, 351-355. 10.1038/nature14603 [DOI] [PubMed] [Google Scholar]
- Munro E., Nance J. and Priess J. R. (2004). Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo. Dev. Cell 7, 413-424. 10.1016/j.devcel.2004.08.001 [DOI] [PubMed] [Google Scholar]
- Münster S., Jain A., Mietke A., Pavlopoulos A., Grill S. W. and Tomancak P. (2019). Attachment of the blastoderm to the vitelline envelope affects gastrulation of insects. Nature 568, 395-399. 10.1038/s41586-019-1044-3 [DOI] [PubMed] [Google Scholar]
- Paré A. C., Vichas A., Fincher C. T., Mirman Z., Farrell D. L., Mainieri A. and Zallen J. A. (2014). A positional Toll receptor code directs convergent extension in Drosophila. Nature515, 523-527. 10.1038/nature13953 [DOI]
- Pokutta S., Drees F., Takai Y., Nelson W. J. and Weis W. I. (2002). Biochemical and structural definition of the l-afadin-and actin-binding sites of α-catenin. J. Biol. Chem. 277, 18868-18874. 10.1074/jbc.M201463200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rappoport J. Z. and Simon S. M. (2003). Real-time analysis of clathrin-mediated endocytosis during cell migration. J. Cell Sci. 116, 847-855. 10.1242/jcs.00289 [DOI] [PubMed] [Google Scholar]
- Rauzi M., Lenne P.-F. and Lecuit T. (2010). Planar polarized actomyosin contractile flows control epithelial junction remodelling. Nature 468, 1110-1114. 10.1038/nature09566 [DOI] [PubMed] [Google Scholar]
- Rodal A. A., Del Signore S. J. and Martin A. C. (2015). Drosophila comes of age as a model system for understanding the function of cytoskeletal proteins in cells, tissues, and organisms. Cytoskeleton 72, 207-224. 10.1002/cm.21228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers S. L., Wiedemann U., Häcker U., Turck C. and Vale R. D. (2004). Drosophila RhoGEF2 associates with microtubule plus ends in an EB1-dependent manner. Curr. Biol. 14, 1827-1833. 10.1016/j.cub.2004.09.078 [DOI] [PubMed] [Google Scholar]
- Roh-Johnson M., Shemer G., Higgins C. D., McClellan J. H., Werts A. D., Tulu U. S., Gao L., Betzig E., Kiehart D. P. and Goldstein B. (2012). Triggering a cell shape change by exploiting preexisting actomyosin contractions. Science 335, 1232-1235. 10.1126/science.1217869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Röper K. (2012). Anisotropy of Crumbs and aPKC drives myosin cable assembly during tube formation. Dev. Cell 23, 939-953. 10.1016/j.devcel.2012.09.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salbreux G., Charras G. and Paluch E. (2012). Actin cortex mechanics and cellular morphogenesis. Trends Cell Biol. 22, 536-545. 10.1016/j.tcb.2012.07.001 [DOI] [PubMed] [Google Scholar]
- Sawyer J. K., Harris N. J., Slep K. C., Gaul U. and Peifer M. (2009). The Drosophila afadin homologue Canoe regulates linkage of the actin cytoskeleton to adherens junctions during apical constriction. J. Cell Biol. 186, 57-73. 10.1083/jcb.200904001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawyer J. K., Choi W., Jung K.-C., He L., Harris N. J. and Peifer M. (2011). A contractile actomyosin network linked to adherens junctions by Canoe/afadin helps drive convergent extension. Mol. Biol. Cell 22, 2491-2508. 10.1091/mbc.e11-05-0411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scarpa E., Finet C., Blanchard G. B. and Sanson B. (2018). Actomyosin-driven tension at compartmental boundaries orients cell division independently of cell geometry in vivo. Dev. Cell 47, 727-740.e6. 10.1016/j.devcel.2018.10.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholey J. M., Taylor K. A. and Kendrick-Jones J. (1980). Regulation of non-muscle myosin assembly by calmodulin-dependent light chain kinase. Nature 287, 233-235. 10.1038/287233a0 [DOI] [PubMed] [Google Scholar]
- Silver J. T., Wirtz-Peitz F., Simões S., Pellikka M., Yan D., Binari R., Nishimura T., Li Y., Harris T. J. C., Perrimon N. et al. (2019). Apical polarity proteins recruit the RhoGEF Cysts to promote junctional myosin assembly. J. Cell Biol. 218, 3397-3414. 10.1083/jcb.201807106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simões S. D. M., Blankenship J. T., Weitz O., Farrell D. L., Tamada M., Fernandez-Gonzalez R. and Zallen J. A. (2010). Rho-kinase directs Bazooka/Par-3 planar polarity during Drosophila axis elongation. Dev. Cell 19, 377-388. 10.1016/j.devcel.2010.08.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simões S., Oh Y., Wang M. F. Z., Fernandez-Gonzalez R. and Tepass U. (2017). Myosin II promotes the anisotropic loss of the apical domain during Drosophila neuroblast ingression. J. Cell Biol. 216, 1387-1404. 10.1083/jcb.201608038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokolow A., Toyama Y., Kiehart D. P. and Edwards G. S. (2012). Cell ingression and apical shape oscillations during dorsal closure in Drosophila. Biophys. J. 102, 969-979. 10.1016/j.bpj.2012.01.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solon J., Kaya-Çopur A., Colombelli J. and Brunner D. (2009). Pulsed forces timed by a ratchet-like mechanism drive directed tissue movement during dorsal closure. Cell 137, 1331-1342. 10.1016/j.cell.2009.03.050 [DOI] [PubMed] [Google Scholar]
- Stull J. T., Hsu L. C., Tansey M. G. and Kamm K. E. (1990). Myosin light chain kinase phosphorylation in tracheal smooth muscle. J. Biol. Chem. 265, 16683-16690. [PubMed] [Google Scholar]
- Sumi A., Hayes P., D'Angelo A., Colombelli J., Salbreux G., Dierkes K. and Solon J. (2018). Adherens junction length during tissue contraction is controlled by the mechanosensitive activity of actomyosin and junctional recycling. Dev. Cell 47, 453-463.e3. 10.1016/j.devcel.2018.10.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutherland A. and Lesko A. (2020). Pulsed actomyosin contractions in morphogenesis. F1000Res. 9, 142 10.12688/f1000research.20874.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweeton D., Parks S., Costa M. and Wieschaus E. (1991). Gastrulation in Drosophila: the formation of the ventral furrow and posterior midgut invaginations. Development 112, 775-789. [DOI] [PubMed] [Google Scholar]
- Tan I., Yong J., Dong J. M., Lim L. and Leung T. (2008). A tripartite complex containing MRCK modulates lamellar actomyosin retrograde flow. Cell 135, 123-136. 10.1016/j.cell.2008.09.018 [DOI] [PubMed] [Google Scholar]
- Tansey M. G., Luby-Phelps K., Kamm K. E. and Stull J. T. (1994). Ca (2+)-dependent phosphorylation of myosin light chain kinase decreases the Ca2+ sensitivity of light chain phosphorylation within smooth muscle cells. J. Biol. Chem. 269, 9912-9920. [PubMed] [Google Scholar]
- Tepass U., Gruszynski-DeFeo E., Haag T. A., Omatyar L., Török T. and Hartenstein V. (1996). shotgun encodes Drosophila E-cadherin and is preferentially required during cell rearrangement in the neurectoderm and other morphogenetically active epithelia. Genes Dev. 10, 672-685. 10.1101/gad.10.6.672 [DOI] [PubMed] [Google Scholar]
- Totsukawa G., Yamakita Y., Yamashiro S., Hartshorne D. J., Sasaki Y. and Matsumura F. (2000). Distinct roles of ROCK (Rho-kinase) and MLCK in spatial regulation of MLC phosphorylation for assembly of stress fibers and focal adhesions in 3T3 fibroblasts. J. Cell Biol. 150, 797-806. 10.1083/jcb.150.4.797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valencia-Expósito A., Grosheva I., Míguez D. G., González-Reyes A. and Martín-Bermudo M. D. (2016). Myosin light-chain phosphatase regulates basal actomyosin oscillations during morphogenesis. Nat. Commun. 7, 10746 10.1038/ncomms10746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderleest T. E., Smits C. M., Xie Y., Jewett C. E., Blankenship J. T. and Loerke D. (2018). Vertex sliding drives intercalation by radial coupling of adhesion and actomyosin networks during Drosophila germband extension. eLife 7, e34586 10.7554/eLife.34586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasquez C. G., Tworoger M. and Martin A. C. (2014). Dynamic myosin phosphorylation regulates contractile pulses and tissue integrity during epithelial morphogenesis. J. Cell Biol. 206, 435-450. 10.1083/jcb.201402004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y. and Riechmann V. (2007). The role of the actomyosin cytoskeleton in coordination of tissue growth during Drosophila oogenesis. Curr. Biol. 17, 1349-1355. 10.1016/j.cub.2007.06.067 [DOI] [PubMed] [Google Scholar]
- Wendt T., Taylor D., Trybus K. M. and Taylor K. (2001). Three-dimensional image reconstruction of dephosphorylated smooth muscle heavy meromyosin reveals asymmetry in the interaction between myosin heads and placement of subfragment 2. Proc. Natl Acad. Sci. USA 98, 4361-4366. 10.1073/pnas.071051098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood W., Jacinto A., Grose R., Woolner S., Gale J., Wilson C. and Martin P. (2002). Wound healing recapitulates morphogenesis in Drosophila embryos. Nat. Cell Biol. 4, 907-912. 10.1038/ncb875 [DOI] [PubMed] [Google Scholar]
- Woodhead J. L., Zhao F.-Q., Craig R., Egelman E. H., Alamo L. and Padrón R. (2005). Atomic model of a myosin filament in the relaxed state. Nature 436, 1195-1199. 10.1038/nature03920 [DOI] [PubMed] [Google Scholar]
- Xie S. and Martin A. C. (2015). Intracellular signalling and intercellular coupling coordinate heterogeneous contractile events to facilitate tissue folding. Nat. Commun. 6, 7161 10.1038/ncomms8161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamashiro S., Totsukawa G., Yamakita Y., Sasaki Y., Madaule P., Ishizaki T., Narumiya S. and Matsumura F. (2003). Citron kinase, a Rho-dependent kinase, induces di-phosphorylation of regulatory light chain of myosin II. Mol. Biol. Cell 14, 1745-1756. 10.1091/mbc.e02-07-0427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yevick H. G., Miller P. W., Dunkel J. and Martin A. C. (2019). Structural redundancy in supracellular actomyosin networks enables robust tissue folding. Dev. Cell 50, 586-598. 10.1016/j.devcel.2019.06.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J. C. and Fernandez-Gonzalez R. (2016). Local mechanical forces promote polarized junctional assembly and axis elongation in Drosophila. eLife 5, e10757 10.7554/eLife.10757 [DOI] [PMC free article] [PubMed] [Google Scholar]