

Abstract
Background:
Hurricane Maria struck Puerto Rico on September 20th, 2017 causing catastrophic devastation. Prolonged shortage of food had been a substantial challenge to the residents after Maria. Experiencing food insecurity in utero has been associated with negative health outcomes later in life. We aim to examine whether there is any alteration in the infant gut microbiome that is associated with prenatal food insecurity.
Methods:
We established a cohort of infants aged 2–6 months who were exposed in utero to Hurricane Maria near San Juan, Puerto Rico and examined the gut microbiota (n=29) using 16S rRNA gene sequencing.
Results:
Among the enrolled infants, 30% of their mothers experienced “post-Maria poor access to food” for at least one month during pregnancy. The relative abundance of gut Veillonella spp. is significantly decreased among infants who experienced prenatal food insecurity, compared to those who did not (adjusted p=0.025). There is no significant difference observed by prenatal food insecurity at the microbial community level in this cohort.
Conclusion:
Our finding indicated that infants who experienced prenatal food insecurity post hurricane harbor microbial alternations of specific bacterial taxa, which may further influence the microbial maturation and place the individual at a high-risk health trajectory.
Introduction
The deadly Category 4 tropical cyclone, Hurricane Maria, struck Puerto Rico on September 20th, 2017. The whole island experienced catastrophic devastation and sustained a humanitarian crisis with an estimated death toll ranging from 2975 to 4645 and over $90 billion in damages (1, 2). In the aftermath of Maria, widespread loss of electricity and communication, the collapse of the healthcare system, and the prolonged shortage of food and other critical resources, combined with delayed disaster response and aid relief, lead to substantial physical and psychological challenges to the residents of Puerto Rico (3). Previous epidemiological studies have demonstrated that prenatal exposure to a devastating hurricane is associated with negative birth outcomes and increased disease risks later in life (4–7). However, due to the extremely complex mix of exposures during a hurricane crisis, the impact of each major exposure on the unborn has not been evaluated. To investigate specific exposures in the context of hurricane is a critical step to start understanding biological pathways from prenatal exposure to health outcomes.
Prenatal maternal food insecurity has been identified as a major factor that directly affects the nutrients available for the growing fetus and profoundly influences disease risks later in life (8–10). With growing knowledge of the human microbiome, we have recognized that maternal diet during pregnancy plays an important role in shaping infant gut microbiota, a major stimulus for immune and metabolic function development (11, 12). The prenatal maternal diet can potentially exert its influence on the infant gut microbiota both before and after birth. In mice, prenatally, maternal nutrients and bacterial metabolites shape the infant intestinal mucosal immune cell composition, regulating the intestinal transcription pattern, which applies select pressure on the pioneer bacterial communities at birth and in early infancy (13). Postnatally, maternal bacterial strains can directly transfer to the infant gut and have better ecological adaptation than those acquired from other sources. (14). This mother-to-infant microbial transmission has been observed for a variety of bacterial species and from multiple body sites of the mothers (14–16). Additionally, food insecurity is among the top problems that are associated with stress, anxiety or depression after a hurricane (17). These effects on maternal mental health have also exhibited strong associations with alterations of the infant gut microbiome (18, 19). Thus, food insecurity is an important exposure for the unborn, due to its potential impact on the developing infant gut microbiota and may be the mechanism underlying prenatal disaster exposure leading to long-term health consequences.
To this end, we conducted a study, named Hurricane as the Origin of Later Alterations in microbiome (HOLA), with infants who were exposed in utero to Hurricane Maria in the San Juan greater metropolitan area. We investigated their major prenatal exposures due to Hurricane Maria and evaluated the gut microbiome by prenatal maternal food security state.
Methods
Subject recruitment, sample collection and DNA extraction
HOLA study was reviewed and approved by the Institutional Review Boards of San Juan City Hospital (ID: EPDC-Microbiome) and deemed exempt by Washington University Human Research Protection Office as all data was de-identified. All parents signed a written informed consent for infant-mother pair participation in the study. Between April and August 2018, we recruited infants through San Juan City Hospital Research Unit. Study inclusion criteria were 1) child’s mother resided in Puerto Rico throughout the pregnancy; 2) aged more than two months and less than six; 3) born at term via vaginal delivery. The exclusion criteria include 1) delivery by cesarean section; 2) premature birth (defined as less than 37 weeks of gestational age at the time of birth); 3) admission to Neonatal Intensive Care Unit; 4) acute illness at the time of acquiring the samples; 5) airway or pulmonary malformations; 6) identified chromosomal or genetic abnormalities.
The infants’ fresh stool samples were collected at clinic or during a home visit by a trained health professional, taken directly from the diaper liner using a sterile swab. The swab was then immediately placed into the DNA/RNA Shield collection solution (Zymo Resesrch, Irvine, CA), and shipped to Washington University for bacterial DNA extraction. The DNA/RNA Shield collection tube contains DNA and RNA stabilization solution, which preserves the integrity of nucleic acids of the samples. Therefore, the storage and transportation of collection tubes were at ambient temperatures. Thirty infants were recruited and 29 stool samples were used for final analysis since one subject was under antibiotic treatment when the stool sample was collected. At the time of sample collection, we conducted a questionnaire-based interview in Spanish with the mother of each subject. The questionnaire was based off various sources, including personal experience with disaster of some of the members of the research teams, particularly those who lived or were actively involved in disaster relief immediately after the Hurricane. The questions assessing hurricane exposures are listed in Supplementary file 2. We additionally administered the Edinburgh Postnatal Depression Scale (EPDS), a standard psychological instrument for detection of depressive symptoms (20). Two researchers independently extracted data from the questionnaire and calculated EPDS score. Any discrepancy in the data input was carefully discussed to resolve.
DNA extraction, PCR Amplification and Sequencing of Bacterial 16S rRNA Genes
Total genomic DNA was extracted from the swab collection using Zymo Fecal/Soil Microbe Miniprep Kit (Zymo Research, Irvine, CA). Fourteen PCR amplicons, representing all 9 16S variable regions, were produced using the Fluidigm Access Array System. 5ng/ul of DNA were input into each reaction. The reaction mix consisted of 1X High Fidelity FastStart Reaction Buffer without MgCl2 (Roche), 4.5nM MgCl2 (Roche), 5% DMSO (Roche), 200uM PCR Grade Nucleotide Mix (Roche), 0.05 U/μL 5 U/μL FastStart High Fidelity Enzyme Blend (Roche), 1X Access Array Loading Reagent (Fluidigm), 1ul DNA, and water. Two hundred nM each of the forward and reverse primers and 1X Access Array Loading Reagent were added to the reaction mix. PCR amplification was performed on the BioMark HD system from Fluidigm. The reaction products were harvested and indexed using unique 10 base pair sequences with 14 rounds of PCR to incorporate each index sequence. All samples were pooled into 48 sample libraries and cleaned using bead purification. Libraries were sequenced using an Illumina MiSeq sequencer (2×150 base pair kit). Amplification and sequencing was performed at Genome Technology Access Center at Washington University.
16S rRNA gene sequence processing
Of the 14 PCR amplicons sequenced, reads from one amplicon which covers the 16S V4 variable regions were first examined using the QIIME pipeline. Reads from the other 13 amplicons were either discarded or used for the MVRSION taxonomic analysis described below. The paired end reads were joined into single sequences using fastq-join with the default parameters where the minimum length of the overlap is 6 and the maximum hamming distance considered is 8% of the overlap length. Only the joined V4 reads where used for analysis using the QIIME pipeline version 1.9.0 (21). Open-reference operational taxonomic units (OTU) were called using the Greengenes May 2013 release as the reference database (22). Reads were clustered into OTUs by QIIME using UCLUST at a threshold of 97% similarity (23). Representative sequences for each OTU were classified taxonomically with the UCLUST consensus taxonomy assigner in QIIME using a sequence similarity of 0.9. The resultant OTU table was rarefied to the lowest individual total read count of 5600.
To optimize taxonomic assignment, we then used the MVRSION pipeline which uses sequencing reads from 7 of the 14 amplicons to produce a list of microbial species along with the number of reads assigned to each species for each sample (24). MVRSION was run with default parameters except for the novoalign –tval parameter of 180. The data was run on a filtered version of the SILVA 16S database (25). MVRSION produces a taxon read count table where the counts are the number of reads classified for each species averaged across all variable regions. This taxon read count table was rarefied to a depth of 2390 reads.
Statistical analysis
Alpha (e.g. Shannon index) and beta diversity (e.g., weighted UniFrac distance) calculations were performed from rarefied reads in the V4 region. Anosim comparisons of beta diversity distance matrices by prenatal malnutrition status were performed in QIIME. A random forest machine-learning model was used to rank feature importance of the bacteria genera in predicting prenatal exposure to food insecurity. The prediction performance was evaluated using the relative abundance matrices at genus level. We applied Leave-One-Out as the dataset splitting method in which each sample is used once as a test set (singleton) while the remaining samples for the training set. Default parameters were chosen on the Scikit-learn python package v. 0.19.1 (26). The algorithm was repeated 999 times to generate a ranked list in the order of feature importance. R package DESeq2 was used to detect differential bacteria genera using a generalized linear regression model with a logarithmic link and following a negative binomial distribution (27). Resulting p-values were adjusted for multiple testing according to the Benjamini–Hochberg procedure. We used R v. 3.4.2 and Python3 for statistical analyses and ‘ggplot2’ for visualization (28). α values < 0.05 were considered statistically significant.
Results
Prevalence of prenatal maternal food insecurity in HOLA and the gut microbiome at the community level
We enrolled 30 infants aged 2 to 6 months in the HOLA study. All of our subjects resided in the same metropolitan area as defined by the US Census Bureau. The municipalities included are interconnected by main highways and on an average day can be reached from San Juan proper within an hour’s drive time. In the aftermath of Hurricane Maria, limited access to fuel, 100% power outage and loss of local crops affected all areas equally. The prevalence of major hurricane consequences for their mothers during pregnancy is summarized in Figure 1. All the participants’ mothers experienced at least one month of power outage due to the hurricane and were exposed to molds, fumes, or debris. 30% of the mothers experienced “poor access to food due to Hurricane Maria” for at least one month. One participant was under antibiotic treatment for diarrhea when we collected stool samples, so we excluded this subject for microbiome analysis. The demographics of the 29 subjects were described in Table 1. We compared infants whose mothers answered that the household had poor access to “food” due to hurricanes Maria and Irma (occurred two weeks prior to Maria) during gestation (n=8) and infants whose mothers answered that the household did not have poor access to food during gestation (n=21).
Figure 1. Prevalence of self-reported experience due to Hurricane Maria among the mothers of 30 participating infants.
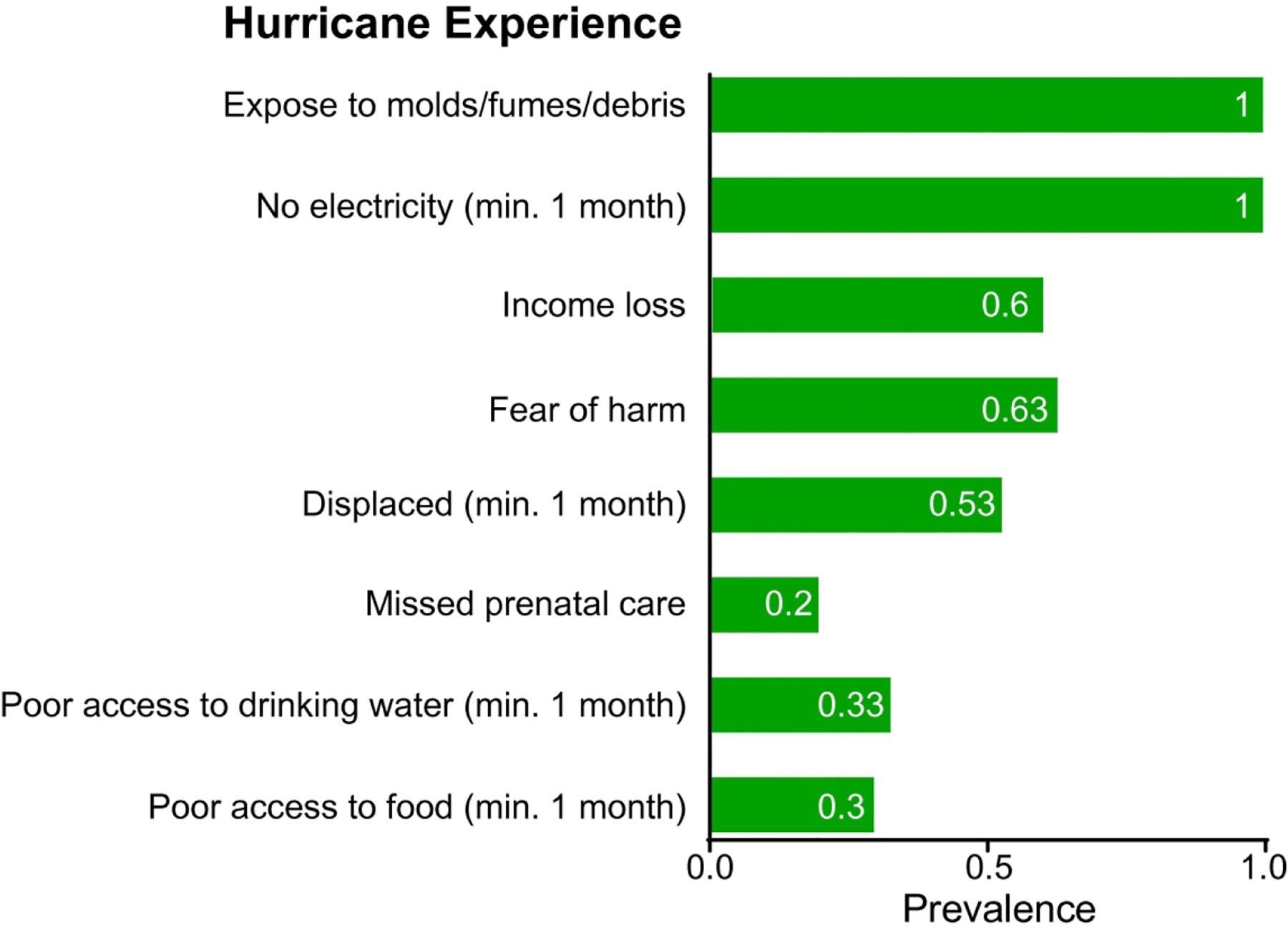
Table 1.
Characterization of the microbiome study cohorta
| Maternal access to food | |||
|---|---|---|---|
| Poor (n=8) | Normal (n=21) | p-value | |
| Ethnicity (Latino) | 8 (100%) | 21 (100%) | |
| Vaginally delivered | 8 (100%) | 21 (100%) | |
| Gestational age in weeks | 39±0.84 | 39±1.07 | 0.73 |
| Trimester on the day of Hurricane Maria | 0.46 | ||
| 1st | 3 (37.5%) | 5 (23.8%) | |
| 2ndb | 5 (62.5%) | 16 (76.2%) | |
| Gender (male) | 2 (25%) | 8 (38.1%) | 0.51 |
| Infant age at sample collection in weeks | 18±5.9 | 15±5.2 | 0.18 |
| Feeding type | 0.43 | ||
| Exclusive breast milk | 2 (25.0%) | 6 (28.6%) | |
| Mixed | 5 (62.5%) | 8 (38.1%) | |
| Exclusive formula | 1 (12.5%) | 7 (33.3%) | |
| Maternal EPDS score | 6.75±7.09 | 4.90±3.78 | 0.50 |
Statistical significance was determined by t-test or chi-square test. SD: standard deviation. EPDS: The Edinburgh Postnatal Depression Scale. Values are presented as number (percentage) or mean ± SD.
totally 29 infants were included in the gut microbiome study. One participant was under antibiotic treatment for diarrhea when we collected stool samples, so we excluded this subject for microbiome analysis.
one of the subjects was 30-week pregnant.
In QIIME, from a total of 226771 reads, we observed 263 operational taxonomic units (OTUs). The OTU table is in Supplementary file 3. The most abundant phyla in the gut microbial community were Firmicutes (37%), Actinobacteria (27%), Bacteroidetes (20%), and Proteobacteria (15%). To evaluate general features of the infant microbiome, we compared the alpha diversity (Shannon’s index) by infant feeding types and age, influential factors on the infant gut microbial community. As shown in Figure 2A, feeding type was significantly associated with the gut microbial community composition, with exclusive breast milk feeding resulting in significantly lower α-diversity compared to exclusive formula feeding (p=0.02), which is consistent with previous studies (29). In Figure 2B, the overall α-diversity shows an increasing trend with infant age, but likely since we captured a narrow age window (2–6 months old), the increase was not significant. In Supplemental Figure S1 A–C (online), the Principal Coordinate Analysis (PCoA) of weighted UniFrac distance shaded for the relative abundance of three major families, Bifidobacteriaceae, Bacteroidaceae, and Enterobacteriaceae, highlights three distinct clusters. Supplemental Figure S1 D–F (online) further shows that the composition of the three major bacterial families is influenced by feeding type with Bifidobacteriaceae significantly correlated with breastfeeding vs. formula feeding (p = 0.03). Notably, there is no significant difference in the α- or β-diversity measured by Shannon’s index (Wilcoxon, p=0.83) or weighted UniFrac distances (ANOSIM, p=0.69) by prenatal food security state (Figure 2C and 2D).
Figure 2. The gut microbial community evaluated by alpha- and beta- diversity.
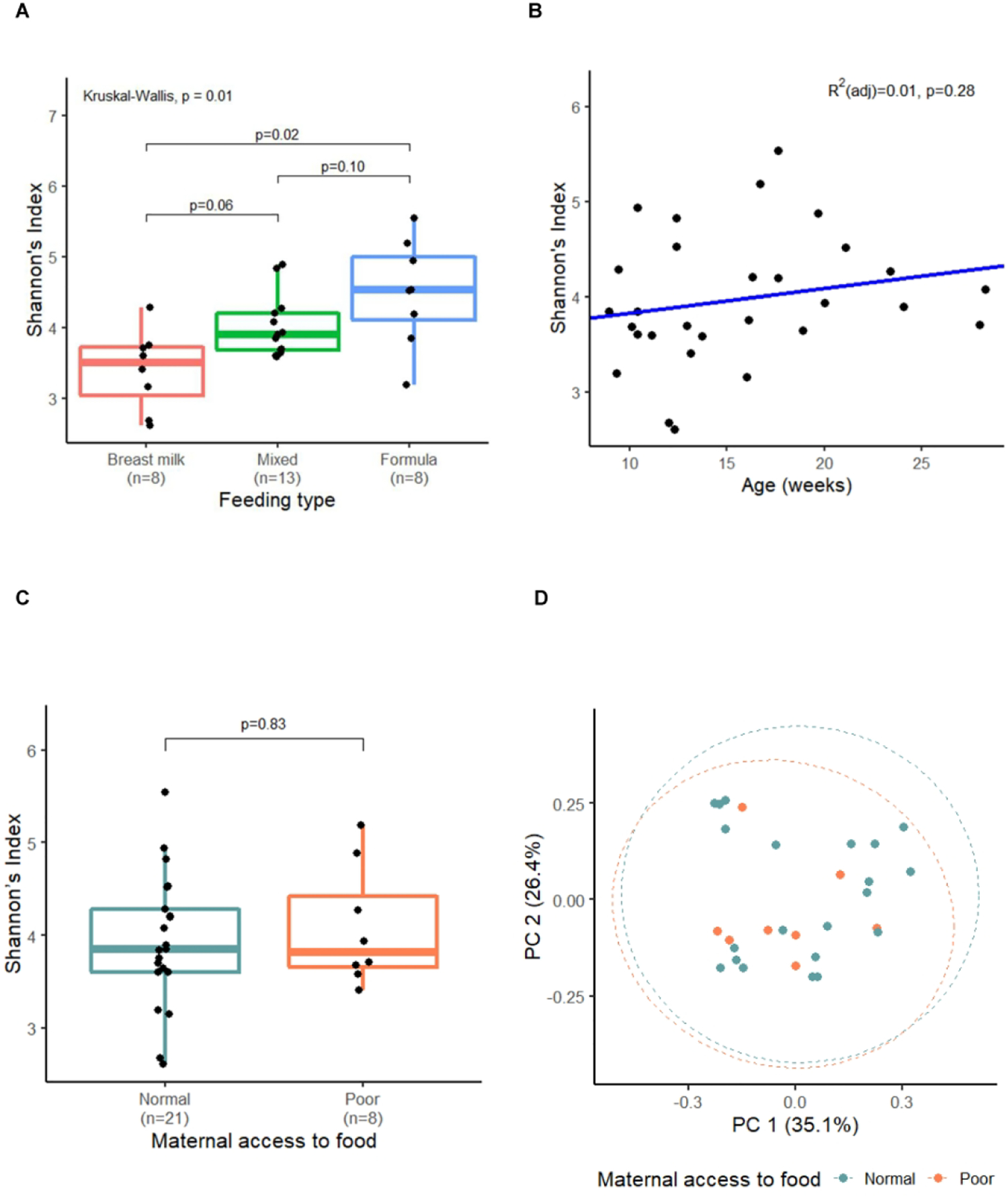
A. Shannon’s index by feeding types. B. Shannon’s index by infant age. The blue line represents the best fit line in the linear regression model. C. Shannon’s index by prenatal maternal food security status. Statistical significance in A-C was determined using Kruskal-Wallis test, Wilcoxon rank sum test, or linear regression model. D. Principal Coordinate Analysis (PCoA) plot based on the weighted UniFrac distance, colored by prenatal maternal food security status (ANOSIM p=0.69). PC1, principal coordinate axis 1. PC2, principal coordinate axis 2.
Decreased relative abundance of Veillonella spp. is associated with prenatal maternal food insecurity.
The relative abundance of most prevalent genera (present in >20% of subjects, 38 in total), grouped by prenatal maternal food security state are shown in Figure 3. To evaluate the potential impact of prenatal food insecurity on the infant microbiome, we conducted a random forest machine-learning model to rank the 38 most prevalent genera by their feature importance on predicting the prenatal food security status. With the cross-validation accuracy of 0.724 in the model, genus Veillonella is ranked as the most important feature with the importance score of 0.13, nearly doubled the importance score of the second-best feature, genus Erwinia with the importance score of 0.07 as shown in Figure 4A and Supplemental Table S1 (online). We then applied a generalized linear regression model using DESeq2 including all 206 genera to identify bacterial genera with significant differential abundance by prenatal food security. After adjusting for multiple comparisons, Veillonella is significantly decreased in infants who experienced food insecurity during gestation, compared to infants who did not (means of relative abundance: 2.3% vs. 7.6%, adjusted p=0.025, Figure 4B and Supplemental Table S2 (online)).
Figure 3. The relative abundance of genera, grouped by food access state, the infant feeding type, and the infant age.
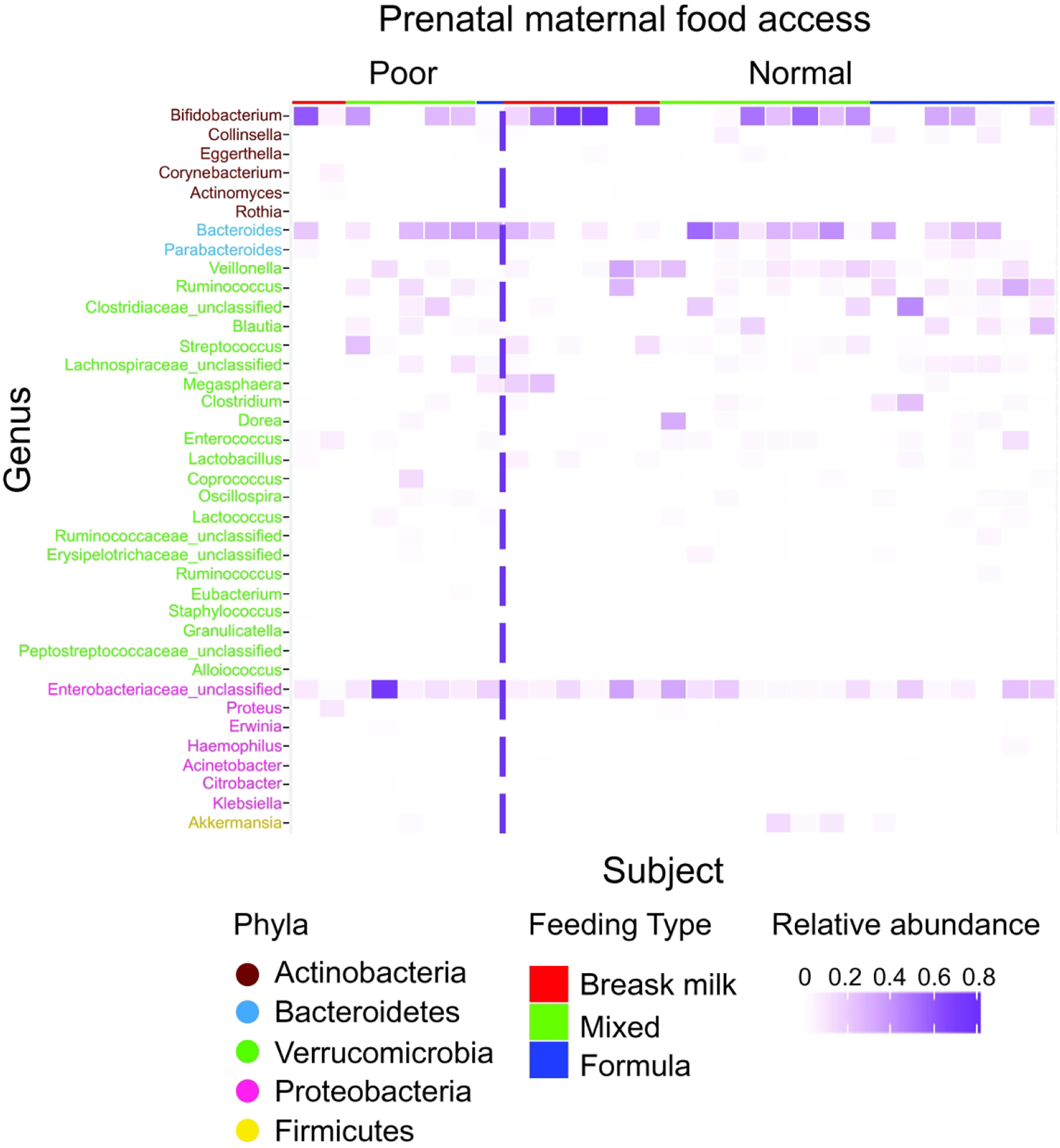
Each column represents one infant. The most prevalent genera (present in at >20% subjects) are included, sorted vertically by phylum and total abundance of all subjects.
Figure 4. Veillonella spp. is significantly associated with prenatal food insecurity.
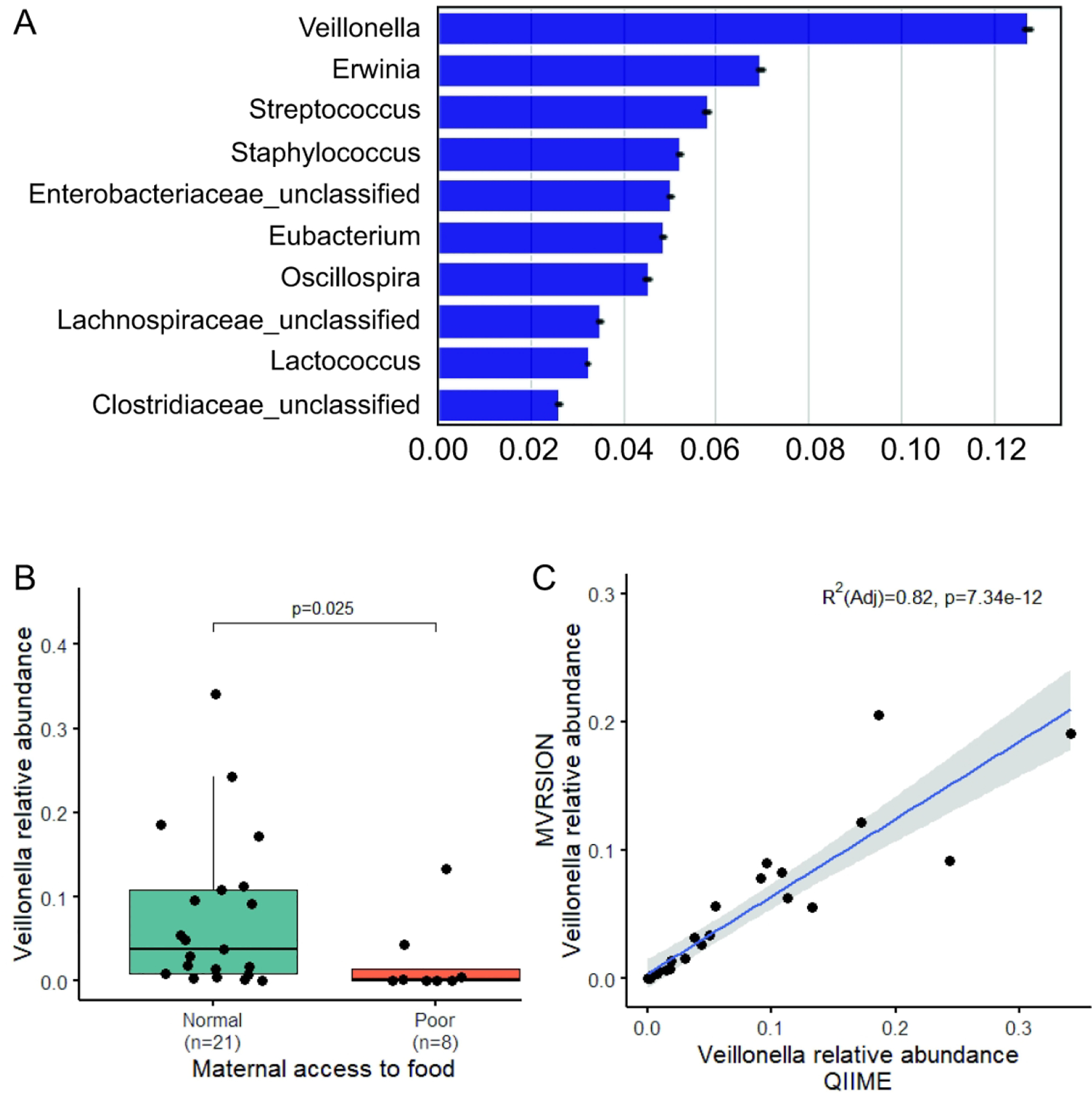
A. The top 10 most important genera in predicting maternal prenatal food security status. The most prevalent 38 genera (present in at >20% subjects, list in A) were included in the random Forest model (999 replicates). The x-axis shows the importance of the genus to the accuracy. The error bar represents the standard error. B. The relative abundance of Veillonella spp. identified in QIIME by prenatal maternal food security status. Statistical significance was determined using Wilcoxon rank sum test. C. Correlation of Veillonella spp. relative abundance calculated by QIIME and MVRSION in the linear regression model. The blue line represents the best-fit line and the shaded gray area represents the 95% confidence interval.
We then performed MVRSION, a novel 16S sequencing and analysis pipeline to validate this observed association in QIIME. We compared the resulting relative abundance of Veillonella spp. between the traditional QIIME pipeline and MVRSION pipeline. As shown in Figure 3D, the abundance of Veillonella spp. are consistent from the two methods (R2=0.82, p=7.34×10−12). MVRSION results show the same significant difference between food insecurity and security groups regarding the relative abundance of Veillonella (Supplemental Figure S2 A (online)). Furthermore, MVRSION identified seven species in genus Veillonella in our samples, including V. parvula, V. dispar, V. ratti, V. magna, V. cricetid, V. atypica, and V. tobetsuensis. As shown in Supplemental Figure S2 B (online), with an increase in the relative abundance of Veillonella spp, more species were present, regardless of the food security status, which indicated a correlation between the relative abundance and species diversity of Veillonella spp (R2=0.55, p=2.19×10−6).
The identified association with Veillonella spp. is not confounded by feeding types, or infant age. Also, this association is not with hurricane-related drinking water insecurity or displacement.
To investigate whether feeding type is confounding the association of Veillonella spp with prenatal food insecurity, we divided the subjects into breast milk or mixed fed group and exclusively formula fed group. As shown in Figure 5A, regardless of feeding type, the trends are similar: infants experiencing prenatal food insecurity have less abundant Veillonella spp. when compared to the infants whose mother did not experience food insecurity. In the breast milk/mixed group, there is a significant decrease of Veillonella spp. in the prenatal food insecure group (p=0.04). With smaller numbers of subjects in the formula group, we did not observe significant difference but can still see the same trend. Infant age is not correlated with Veillonella spp. (p=0.64, data not shown) and the age distributions are not different between these two groups (p=0.18, Table 1). However, the two groups displayed opposite trends regarding the relative abundance of Veillonella with age. As shown in Figure 5B, relative abundance of Veillonella spp. increases with age in the control group. However, the converse is true in the prenatal food insecurity group.
Figure 5. Subgroup analysis for the association between Veillonella and prenatal food insecurity.
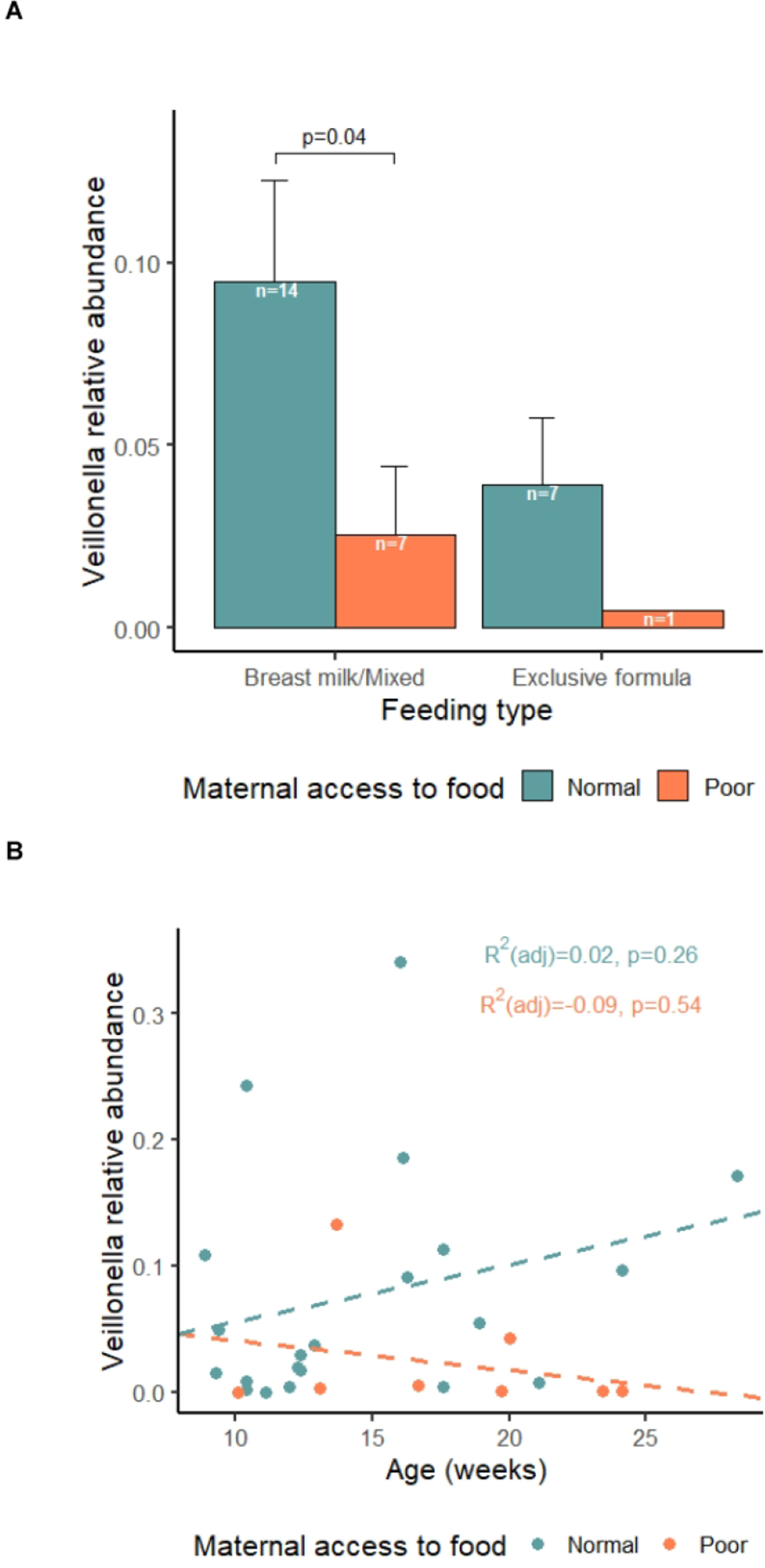
A. the relative abundance of Veillonella spp. by feeding types and prenatal maternal food security status. Values are means ± SEM. Statistical significance was determined using Wilcoxon rank sum test. B. the relative abundance of Veillonella spp. versus age by prenatal maternal food security status. Statistical significance was determined using linear model. The lines are the best fit line in the corresponding linear regression model by maternal food security status respectively.
We then examined the association of Veillonella spp. with other top hurricane-related stressors, including drinking water insecurity and displacement (17, 30). As shown in Supplemental Figure 3 A (online), the relative abundance of Veillonella spp. is not associated with prenatal drinking water security. The mean relative abundance of Veillonella in drinking water secure group vs. drinking water insecure group is 6.04% vs. 6.34% (p=0.68). Similarly, in Supplemental Figure 3 B (online), the relative abundance of Veillonella spp. is not associated with displacement. The mean relative abundance of not-displaced group vs. displaced group is 5.27% vs. 9.52% (p=0.29). These results did not show association of Veillonella abundance with two other hurricane-stressors, prenatal drinking water insecurity or displacement.
Discussion
This is the first human study to examine the impact of maternal prenatal food insecurity after a devastating hurricane on one specific aspect of infant outcomes, the early in life gut microbiome. We identified that the relative abundance and species diversity of gut Veillonella spp. is significantly decreased among infants who had prenatal maternal food insecurity, compared to those who did not. Our findings indicate that infants who experienced prenatal food insecurity post hurricane may harbor alterations of specific bacterial taxa in their gut microbiota, which may further influence the individual’s microbial maturation, immune development and long-term health. There are two possible routes through which maternal food insecurity affects offspring gut microbiota: 1) the maternal nutritional availability is associated with an altered in utero environment that influences fetal programming, resulting in changes to the fetal gut niche that then imposes selective pressures on microbiota early-in-life; 2) an altered maternal microbiota due to events surrounding hurricane Maria can be vertically transmitted to the spring prenatally and/or postnatally. Further studies are needed to investigate these two potential routes.
The microbial alteration among prenatal food insecure infants may be attributable to nutrition deficiency as well as maternal mental distress related to food insecurity. Previous studies on Hurricane Katrina have found that lack of basic necessities, such as food, drinking water, or displacement is strongly associated with posttraumatic stress, anxiety, and depression (17). While mothers in the prenatal food insecure and secure groups had no significant difference in terms of postpartum depression, according to the EPDS tests shown in Table 1, we lacked evaluation of the maternal distress during gestation to delineate the potential impact of food insecurity on the prenatal stress. We examined whether Veillonella was associated with other hurricane-stressors. We found that there was no significant difference in the relative abundance of Veillonella spp. by drinking water security, or displacement experience. However, considering all the mothers experienced a complex mix of chronic stress during gestation, their distress level may not be well reflected by a certain adverse experience in the aftermath of Maria. Therefore, we could not tease out the potentially critical role maternal distress may play on the association.
As shown by our analyses of alpha- and beta- diversity, the microbial community structure of our cohort was influenced by feeding types, which has also been seen in previous studies of other infant cohorts (29). This indicated that prenatal food insecurity may not disturb the microbial community structure of the gut of infants aged 2 to 6 months. However, a previous study has revealed that Veillonella spp. is a dominant taxa in defining ecological structures at age 6 months (31). Therefore, it is possible that the decrease of Veillonella spp.early in life has the potential to initiate structural disturbance which would not be seen until later in life. Follow-up studies are needed to assess long-term effects of prenatal maternal food insecurity on the offspring gut microbiome at both community and taxa levels.
Veillonella spp. are gram-negative anaerobic flora commonly present in multiple body sites, such as mouth, gastrointestinal tract, vagina, and breast milk (32–35). Veillonella spp. is one of the major producers of short-chain fatty acid (SCFA) (36, 37). Low levels of SCFA have been associated with diseases in which inflammatory processes are involved (38, 39). In the Canadian Healthy Infant Longitudinal Development (CHILD) study, Veillonella was among the four genera which were significantly reduced in the early infancy among children at high risk of asthma (39).
Several limitations should be acknowledged in the current study. First, because the cohort size is relatively small, we may not have enough power to detect more subtle changes in the prenatal malnutrition-associated bacterial taxa. Potentially larger effects of prenatal food insecurity on the infant gut microbiome may exist. Therefore, repeated and longitudinal evaluations of the infant gut microbiome in a large-scale cohort post disaster will be necessary to confirm and extend our findings and further examine the influence of prenatal food insecurity after a disaster. Second, although we evaluated potential confounding factors, some important information may still be missing in the current study, such as socioeconomic status (SES), sanitation and pathogen burden in the residing area. These factors can affect food accessibility after Hurricane Maria or directly disturb the gut microbiota. Third, our questionnaire provided us with objective and categorical information to define the state of food access in the household due to Hurricane Maria. Detailed questions regarding the access to different food categories and the major reasons causing food insecurity in a similar cohort would provide a more broad and in-depth view of the severe and prolonged food insecurity problem post Hurricane Maria. Fourth, although our findings focused on the food insecurity issue in the aftermath of Hurricane Maria, the disaster caused widespread destruction and devastating losses to the whole island. More research is warrantied to investigate the link between in-utero exposure to this deadly disaster and biological markers related to the health outcomes.
In summary, HOLA is the first study to examine the influence of prenatal maternal food insecurity after a devastating hurricane on the offspring microbiota. We identified a significant association between limited prenatal access to nourishment and decrease in infant Veillonella spp. While not causal, our findings begin to provide evidence of an underlying mechanism linking in utero exposure to a disaster and increased disease risk later in life. Future human cohort and animal studies will help to validate our findings and determine the mechanisms by which lack of nourishment during pregnancy in the context of a substantial hurricane influences the relative abundance of certain gut microbial taxa in the offspring in early infancy.
Supplementary Material
Impact Statement:
We identified that in-utero exposure to food insecurity post Hurricane Maria is associated with decreased abundance of Veillonella in the infant gut.
Our findings indicated that infants who experienced prenatal food insecurity post hurricane may harbor alterations of specific bacterial taxa in their gut microbiota.
This study showed the association between prenatal adverse exposure and alterations of gut microbiome early in life in the context of an extreme event.
This study provided insights into the mechanisms underlying prenatal adverse exposure and increased disease risks later in life.
Our findings will potentially raise awareness of the negative impact of extreme climate events on the unborn.
Acknowledgements
We express our deepest gratitude to the participating children and their families for their commitment and contribution. We appreciate the aid of Dr. Edna Acosta, under the tutelage of Dr. Duarte (Columbia University) and Dr. Glorisa Canino (University of Puerto Rico) in helping us formulate the disaster-related questions to the HOLA questionnaire. We appreciate the scientific advice from Dr. Gregory Storch (Washington University), Dr. Lora Iannotti (Washington University), Dr. Andrew DeWan (Yale University), and Dr. Gautam Dantas (Washington University).
Statement of Financial Support:
Dr. Leyao Wang is supported by grant 1R21AI139649-01A1 from the NIH/National Institute of Allergy and Infectious Diseases (NIAID). This project was partially supported by the Washington University Institute of Clinical and Translational Sciences which is, in part, supported by the NIH/National Center for Advancing Translational Sciences (NCATS), CTSA grant UL1TR002345.
Footnotes
Availability of sequencing data
The 16S rRNA sequencing files have been deposited into the European Nucleotide Archive (ENA). The accession code is PRJEB32915.
Patient Consent:
All the parents of the participating infants signed a written informed consent.
Disclosure Statement:
The authors declare no competing interests.
Category of Study:
Clinical Research
References
- 1.Santos-Burgoa C, et al. 2018. Differential and persistent risk of excess mortality from Hurricane Maria in Puerto Rico: a time-series analysis. Lancet Planet Health 2:e478–e488. [DOI] [PubMed] [Google Scholar]
- 2.Verma AA, Murray J, Mamdani MM 2018. Mortality in Puerto Rico after Hurricane Maria. N Engl J Med 379:e30. [DOI] [PubMed] [Google Scholar]
- 3.Willison CE, Singer PM, Creary MS, Greer SL 2019. Quantifying inequities in US federal response to hurricane disaster in Texas and Florida compared with Puerto Rico. BMJ Glob Health 4:e001191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Antipova A, Curtis A 2015. The post-disaster negative health legacy: pregnancy outcomes in Louisiana after Hurricane Andrew. Disasters 39:665–686. [DOI] [PubMed] [Google Scholar]
- 5.Zahran S, et al. 2014. Maternal exposure to hurricane destruction and fetal mortality. J Epidemiol Community Health 68:760–766. [DOI] [PubMed] [Google Scholar]
- 6.Sotomayor O 2013. Fetal and infant origins of diabetes and ill health: evidence from Puerto Rico’s 1928 and 1932 hurricanes. Econ Hum Biol 11:281–293. [DOI] [PubMed] [Google Scholar]
- 7.Harville EW, Giarratano G, Savage J, Barcelona de Mendoza V, Zotkiewicz T 2015. Birth Outcomes in a Disaster Recovery Environment: New Orleans Women After Katrina. Matern Child Health J 19:2512–2522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Li J, et al. 2017. Prenatal exposure to famine and the development of hyperglycemia and type 2 diabetes in adulthood across consecutive generations: a population-based cohort study of families in Suihua, China. Am J Clin Nutr 105:221–227. [DOI] [PubMed] [Google Scholar]
- 9.Bahwere P 2014. Severe acute malnutrition during emergencies: burden management, and gaps. Food Nutr Bull 35:S47–51. [DOI] [PubMed] [Google Scholar]
- 10.Hoffman DJ, Reynolds RM, Hardy DB 2017. Developmental origins of health and disease: current knowledge and potential mechanisms. Nutr Rev 75:951–970. [DOI] [PubMed] [Google Scholar]
- 11.Lundgren SN, et al. 2018. Maternal diet during pregnancy is related with the infant stool microbiome in a delivery mode-dependent manner. Microbiome 6:109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chu DM, et al. 2016. The early infant gut microbiome varies in association with a maternal high-fat diet. Genome Med 8:77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gomez de Aguero M, et al. 2016. The maternal microbiota drives early postnatal innate immune development. Science 351:1296–1302. [DOI] [PubMed] [Google Scholar]
- 14.Ferretti P, et al. 2018. Mother-to-Infant Microbial Transmission from Different Body Sites Shapes the Developing Infant Gut Microbiome. Cell Host Microbe 24:133–145 e135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Makino H 2018. Bifidobacterial strains in the intestines of newborns originate from their mothers. Biosci Microbiota Food Health 37:79–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yassour M, et al. 2018. Strain-Level Analysis of Mother-to-Child Bacterial Transmission during the First Few Months of Life. Cell Host Microbe 24:146–154 e144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chan CS, Rhodes JE 2014. Measuring exposure in Hurricane Katrina: a meta-analysis and an integrative data analysis. PLoS One 9:e92899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Glover V, O’Donnell KJ, O’Connor TG, Fisher J 2018. Prenatal maternal stress, fetal programming, and mechanisms underlying later psychopathology-A global perspective. Dev Psychopathol 30:843–854. [DOI] [PubMed] [Google Scholar]
- 19.Hartman S, Sayler K, Belsky J 2019. Prenatal stress enhances postnatal plasticity: The role of microbiota. Dev Psychobiol 61:729–738. [DOI] [PubMed] [Google Scholar]
- 20.Garcia-Esteve L, Ascaso C, Ojuel J, Navarro P 2003. Validation of the Edinburgh Postnatal Depression Scale (EPDS) in Spanish mothers. J Affect Disord 75:71–76. [DOI] [PubMed] [Google Scholar]
- 21.Caporaso JG, et al. 2010. QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7:335–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.McDonald D, et al. 2012. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J 6:610–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Edgar RC 2010. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26:2460–2461. [DOI] [PubMed] [Google Scholar]
- 24.Schriefer AE, et al. 2018. A multi-amplicon 16S rRNA sequencing and analysis method for improved taxonomic profiling of bacterial communities. J Microbiol Methods 154:6–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Quast C, et al. 2013. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41:D590–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pedregosa F, 2011. Scikit-learn: Machine Learning in Python. http://www.jmlr.org/papers/volume12/pedregosa11a/pedregosa11a.pdf. Journal of Machine Learning Research 12 (2011) 2825–2830. [Google Scholar]
- 27.Love MI, Huber W, Anders S 2014. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15:550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wickham H 2016. ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New York. [Google Scholar]
- 29.Savage JH, et al. 2018. Diet during Pregnancy and Infancy and the Infant Intestinal Microbiome. J Pediatr 203:47–54 e44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Taioli E, et al. 2018. Mental health challenges and experiences in displaced populations following Hurricane Sandy and Hurricane Harvey: the need for more comprehensive interventions in temporary shelters. J Epidemiol Community Health 72:867–870. [DOI] [PubMed] [Google Scholar]
- 31.Lee MJ, et al. 2018. Perturbations of gut microbiome genes in infants with atopic dermatitis according to feeding type. J Allergy Clin Immunol 141:1310–1319. [DOI] [PubMed] [Google Scholar]
- 32.Yamashita Y, Takeshita T 2017. The oral microbiome and human health. J Oral Sci 59:201–206. [DOI] [PubMed] [Google Scholar]
- 33.Simonyte Sjodin K, Vidman L, Ryden P, West CE 2016. Emerging evidence of the role of gut microbiota in the development of allergic diseases. Curr Opin Allergy Clin Immunol 16:390–395. [DOI] [PubMed] [Google Scholar]
- 34.Nelson TM, et al. 2015. Vaginal biogenic amines: biomarkers of bacterial vaginosis or precursors to vaginal dysbiosis? Front Physiol 6:253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jost T, Lacroix C, Braegger CP, Rochat F, Chassard C 2014. Vertical mother-neonate transfer of maternal gut bacteria via breastfeeding. Environ Microbiol 16:2891–2904. [DOI] [PubMed] [Google Scholar]
- 36.Counotte GH, Prins RA, Janssen RH, Debie MJ 1981. Role of Megasphaera elsdenii in the Fermentation of dl-[2-C]lactate in the Rumen of Dairy Cattle. Appl Environ Microbiol 42:649–655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Flint HJ, Duncan SH, Scott KP, Louis P 2015. Links between diet, gut microbiota composition and gut metabolism. Proc Nutr Soc 74:13–22. [DOI] [PubMed] [Google Scholar]
- 38.Machiels K, et al. 2014. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 63:1275–1283. [DOI] [PubMed] [Google Scholar]
- 39.Arrieta MC, et al. 2015. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci Transl Med 7:307ra152. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.