

Highlights
-
•
In the 2019 American College of Sports Medicine Wolffe lecture George Brooks described history Lactate Shuttle development.
-
•
Lactate is a fuel energy source, the main gluconeogenic precursor, and a signaling molecule with autocrine, paracrine, and endocrine functions.
-
•
Endurance training develops the capacities to produce, remove and utilize lactate.
-
•
Lactate is favored as a fuel by working red muscle, heart, liver, and brain. Clinical trials are under way to improve outcomes in traumatic brain injury patients using lactate supplementation.
-
•
The importance of Lactate Shuttle in normal physiology is emphasized by the realization that dysfunctional lactate shuttling is involved in cancer, metabolic inflexibility, and inflammation.
Keywords: Anaerobic metabolism; Exercise; Glycolysis; Oxidative metabolism, Warburg Effect
Abstract
Once thought to be a waste product of oxygen limited (anaerobic) metabolism, lactate is now known to form continuously under fully oxygenated (aerobic) conditions. Lactate shuttling between producer (driver) and consumer cells fulfills at least 3 purposes; lactate is: (1) a major energy source, (2) the major gluconeogenic precursor, and (3) a signaling molecule. The Lactate Shuttle theory is applicable to diverse fields such as sports nutrition and hydration, resuscitation from acidosis and Dengue, treatment of traumatic brain injury, maintenance of glycemia, reduction of inflammation, cardiac support in heart failure and following a myocardial infarction, and to improve cognition. Yet, dysregulated lactate shuttling disrupts metabolic flexibility, and worse, supports oncogenesis. Lactate production in cancer (the Warburg effect) is involved in all main sequela for carcinogenesis: angiogenesis, immune escape, cell migration, metastasis, and self-sufficient metabolism. The history of the tortuous path of discovery in lactate metabolism and shuttling was discussed in the 2019 American College of Sports Medicine Joseph B. Wolffe Lecture in Orlando, FL.
Graphical Abstract
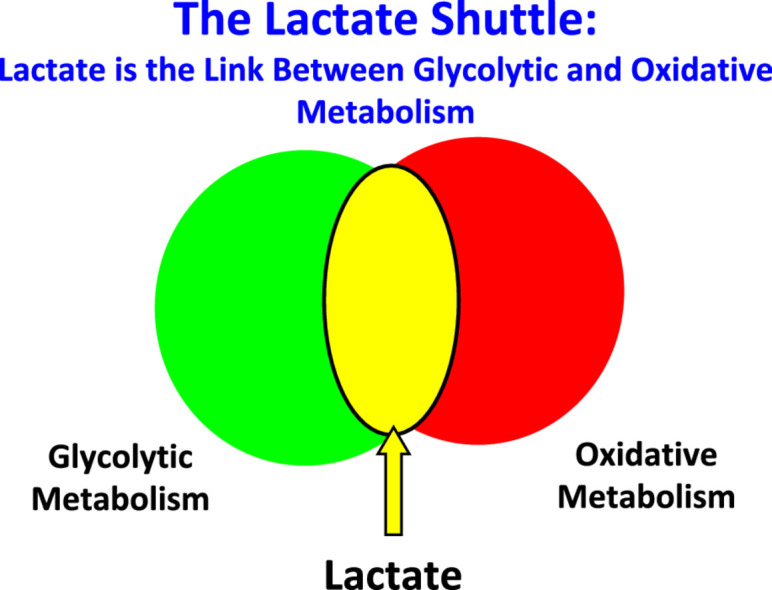
1. Introduction
1.1. Author's perspective
Fifty-five years ago, the author's understanding of the biochemistry and physiology was simple: the accumulation of lactic acid indicated O2 debt. Lactate was a dead-end metabolite, a metabolic waste that caused muscle fatigue, rigor, cramps, and soreness. Certainty of that understanding was assured because those ideas came from the founders of modern biochemistry (O. Meyerhof)1 and muscle physiology (A.V. Hill).2 However, from a teleological view, those ideas made no sense to the author, a 20-year-old at the time. Furthermore, the ideas seemed dated and inconsistent with emerging ideas in mitochondrial energetics,3,4 muscle metabolism and contraction.5 In contrfrom lactate shuttle theory (Fig. 1),6, 7, 8 today we know that lactate is continuously produced under fully aerobic conditions (Fig. 2) and is a major fuel for muscle,9 heart,10 and brain;11 the major gluconeogenic precursor;12,13 and a signaling molecule.14 Furthermore, lactate may serve to alleviate muscle fatigue due to extracellular K+ accumulation15 and, because it is the terminal step in glycolysis, catabolized by lactate dehydrogenase (LDH), produces lactate anion, not lactic acid.16, 17, 18
Fig. 1.

The lactate shuttle concept, depicting lactate as the vehicle linking glycolytic and oxidative metabolism. Linkages between lactate “producer” and “consumer” exist within and among cells, tissues, and organs. As the product of one metabolic pathway (glycolysis) and the substrate for a downstream pathway of disposal (mitochondrial respiration), lactate is the link between the glycolytic and aerobic pathways. Importantly, according to the lactate shuttle hypothesis, this linkage occurs continuously under fully aerobic conditions, can transcend compartment barriers and occur within and among cells, tissues and organs. Modified from Refs 6, 23, 194 with permission.
Fig. 2.

Lactate production occurs continuously under fully aerobic conditions in intact animals, mammalian tissue preparations, intact animals, and humans in vivo. In muscles and arterial blood of resting healthy humans, lactate concentration approximates 1.0 mmol/L, while pyruvate concentration approximates 0.1 mmol/L. The lactate/pyruvate (L/P) approximates 10, with net lactate production and release from resting muscle of healthy individuals occurring when arterial partial pressure of oxygen (PO2) approximates 100 Torr and intramuscular PO2 approximates 40 Torr, well above the critical mitochondrial PO2 for maximal mitochondrial respiration (1–2 Torr).195, 196, 197 During exercise at about 65% of maximal oxygen consumption (VO2max), lactate production and net lactate release from working muscle beds rise and the L/P rises more than an order of magnitude (to approximately 500).57 However, the intramuscular PO2 remains at 3–4 Torr, well above the critical mitochondrial O2 level. Hence, it is appropriate to conclude that in healthy humans, glycolysis proceeds to lactate under fully aerobic conditions. Importantly, most (75%–80%) lactate is disposed of immediately within the tissue or subsequent to release and reuptake by working muscle, with significant uptake and oxidation by heart or oxidation by liver for gluconeogenesis. From diverse sources9,45,57,144,197,198 with permission.
Given the early history, how paradoxical it is that we19 and others20,21 are evaluating the efficacy of using lactate-containing solutions to provide support in the setting of critical care medicine. Specifically, for treatment of traumatic brain injury (TBI), lactate formulations directly support neuronal metabolism when glucose uptake is limited following injury, achieve exquisite glycemic control by supplying the major gluconeogenic precursor for liver and kidneys, and limit postinjury cerebral swelling.19 In the last century, a very insightful man said, “der Alte würfelt nicht!” (“the old one (i.e., creator) does not play dice with the universe”).22 So it is that lactate production and use is now to be viewed as a basic biological response that is accelerated to mitigate a variety of metabolic stresses.
1.2. Physiological stress and lactate strain
In physiology, it is typical that lactate accumulates when there is a physiologic stress. The classical interpretation was that lactate accumulation was a stress (a fatigue agent, a dead-end metabolite, the result of O2 insufficiency, etc.);1 retrospectively, that was a biased and incorrect view. To the contrary, increased glycolysis in response to stress could have, and should have, been regarded as a compensatory strain response.23
That luminaries in biology and medicine were involved in O2Debt theory has historically been a problem for the field. For instance, in the unperfused and unoxygenated frog hemicorpus preparation of Meyerhof,1 electrical stimulation led to glycogen depletion, lactate accumulation and fatigue. This was an ideal setup to quantitatively relate glycogen depletion to lactate accumulation, but the experimental setup (Fig. 3) was nonphysiological and incapable of representing what happens in vivo. With the benefit of a century of research we now appreciate that in a closed, oxygen-limited environment, glycolysis was a major means to support adenosine triphosphate (ATP) flux. In concert, A.V. Hill, one of the founders of muscle and exercise physiology, measured postexercise O2 consumption, but not lactate kinetics in humans recovering from exercise,24 but he never measured either O2 consumption or lactate production in the isolated frog muscle preparations he studied.2 The evidence linking lactate production to muscle oxygen insufficiency was circumstantial, and, unfortunately, ill-considered. Regrettably, the Pavlovian and knee-jerk responses to lactate accumulation and attributing it to O2 insufficiency have persisted.25
Fig. 3.
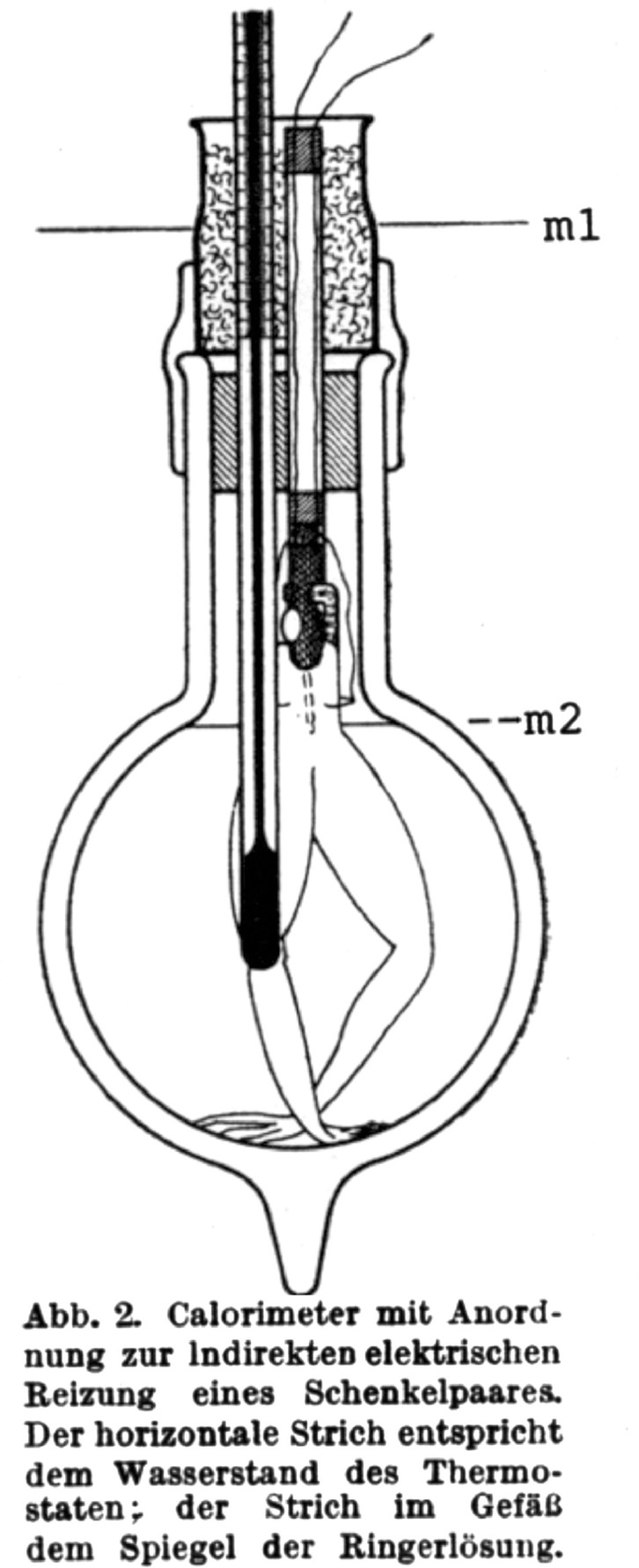
The Meyerhof calorimeter. m1 is the device for indirect electrical stimulation of frog hemicorpus thighs; the horizontal line (m2) is Ringers solution level. This device was used to demonstrate quantitative conversion of glycogen to lactate under nonperfused and nonoxygenated conditions. From Ref. 1 with permission.
1.3. Controversies within the Nobel tradition
Despite widespread interpretation of data by Nobel Prize winners1,2 that lactate production was the result of O2-limited metabolism, observations that lactate could be produced under fully aerobic conditions were made by other Nobelists.26,27 Glucose consumption leading to lactate production under fully aerobic conditions was revealed to be a hallmark of cancer.27 While originally ascribed to the presence of mitochondrial defects in cancer cells, subsequently the ability of mitochondria in cancer cells to respire with substrates, including lactate, was demonstrated.28 Importantly, the efficacious use of lactate produced in fully oxygenated muscles to support gluconeogenesis and maintain euglycemia was demonstrated by the Coris.26 However, while there was good reason to think that lactate could be efficacious in vivo,26 the general belief of lactate as a metabolic waste and stress metabolite persisted through much of the 20th century.24,29
1.4. Origin of the Lactate Shuttle theory
The inspiration leading to articulation of the Lactate Shuttle hypothesis came from several lines of evidence using lab rats as models of mammalian metabolism;6 evidence came from radiotracer studies of glucose and lactate kinetics and measurements of blood and muscle lactate concentrations in resting and exercising animals. At its essence, the hypothesis is that lactate shuttles among producer (driver) and consumer (recipient) tissues, cells, and cellular compartments.
1.5. Key results from isotope tracer studies
In contrast to the one-fifth/four-fifths oxidation/glyconeogenesis theory of lactate disposal during exercise recovery 14C-lactate injected into lab rats resulted in label exhalation as 14CO2, with little incorporation into muscle glycogen.30,31 Second, technologies were developed to make metabolic measurements on resting, exercising, and recovering lab animals32 and then determine concentrations and specific activities of CO2, glucose, lactate, and related metabolites33 (Fig. 4A and 4B), respectively. Indeed, it is likely that none of these advances would have been possible without isotope tracer technologies enabling measurements of metabolite production and disposal rates. With those methods available, it became possible to simultaneously compare glucose34 and lactate35 flux (turnover, production, and disposal) rates as well as rates of lactate disposal via oxidation and gluconeogenesis in trained and untrained rats both at rest and when exercising. Hence, the presence of lactate shuttling in a mammalian (rat) system was revealed (Fig. 5). In resting rats, lactate turnover and oxidation rates were surprisingly high, but were typically less than corresponding glucose flux rates. However, during exercise, lactate flux and oxidation easily exceeded corresponding glucose flux rates. This was because of the contribution of glycogen to glycolytic flux during exercise. Moreover, during exercise, most glucose production came from lactate via gluconeogenesis.34 Training not only increased the capacity of gluconeogenesis, but also had dramatic effects on lactate the metabolic clearance rate (disposal rate/concentration). The classic effect of exercise training on lowering blood lactate concentration was observed in running rats. However, tracer data showed that while lactate production was high in running rats, lactate production was balanced by disposal. Hence, the lower circulating blood lactate concentration in trained rats during exercise was explained by greater clearance rates due to increased oxidation and gluconeogenesis.34,35 Subsequently, with the advent of stable, nonradioactive isotope tracers, metabolite flux studies could be conducted on human subjects. In short, the same effects of exercise and exercise training on glucose and lactate production and disposal rates as observed in rats were replicated in cross-sectional36, 37, 38, 39 and longitudinal training studies on humans.9,13,40
Fig. 4.

Devices use to support the presence of lactate shuttling in resting and exercising mammals in vivo. These include (A) a motorized treadmill with sensors to determine oxygen consumption (VO2), rate of elimination of carbon dioxide (VCO2), electrocardiogram (ECG), RER (= VCO2/VO2)32 and (B) blood-specific activities of glucose, lactate glucose (Gluc), fructose di-phosphate (FDP), and other metabolic intermediates using 2-dimensional paper chromatography, autoradiography and enzymatic analyses.33 See original papers32,33 for details on how lactate flux (production and disposal, oxidation and gluconeogenic) rates were determined in a mammalian model organism during physical exercise. From Ref. 23 with permission.
Fig. 5.

Depiction of the lactate shuttle as it fulfills 3 physiologic functions: (1) lactate as a major energy source, (2) lactate as the major gluconeogenic precursor, and (3) lactate as a signaling molecule with autocrine-, paracrine-, and endocrine-like effects (called a “lactormone”). “Cell–Cell” and “Intracellular Lactate Shuttle” concepts describe the roles of lactate in the delivery of oxidative and gluconeogenic substrates as well as in cell signaling. Examples of the Cell–Cell Lactate Shuttles include lactate exchanges between white glycolytic and red oxidative fibers within a working muscle bed and between working skeletal muscle and heart, brain, liver, and kidneys. Examples of Intracellular Lactate Shuttles include cytosol–mitochondrial and cytosol–peroxisome exchanges. Indeed, most, if not all, lactate shuttles are driven by a concentration or pH gradient or by redox state. FG = fast glycolytic; SO = slow oxidative. The figure is annotated because the original model did not anticipate cerebral lactate oxidation and hepatic and renal gluconeogenesis. Compiled from diverse sources6, 7, 8 and194 with permission.
1.6. Key results from tissue lactate concentration studies
The results of isotope tracer studies34,35 showing high rates of lactate disposal via tissue exchange and oxidation were complemented by results of carefully done rat muscle studies by Paul Molé, Kenneth Baldwin, and their associates.41,42 The Lactate Shuttle hypothesis was necessary to explain why in working red skeletal muscles, lactate concentrations were not only lower than those in white sections of the same muscle but also were lower than those in the blood perfusing them. Thus, the concept of a lactate shuttle6 was supported by independent lines of confirmatory data,6 which were subsequently described more fully (Figs. 1, 2, and 5).43,44
1.7. Key results from tissue net and isotope tracer balance studies
Results of whole body tracer and tissue lactate concentration studies were informative but provided minimal information on the tissue sites of lactate production and disposal in vivo. However, in the early 1980s, studies on tissue specificity of lactate metabolism were under way,9,45,46 and those efforts served as templates for further research.9,10,40 For example, in 1988, fueling of the heart during exercise with lactate released from working muscle beds was observed and was key to envisioning lactate shuttling in humans.10 In terms of Cell–Cell, or Tissue–Tissue lactate shuttling, it was subsequently recognized that lactate shuttled and carbon recycled between and among tissue beds, such as between skeletal muscle, heart and liver (Fig. 5). In terms of driver cells, fast white skeletal cells and tissues42,47, 48, 49 and the integument50 were identified. In terms of recipient cells and tissues, the beating heart takes up and oxidizes lactate,10,46,51 as do working skeletal muscle beds.9,45 However, what is the path between lactate uptake and the formation and release of CO2 within an organ, tissue or cell? Was there something else, perhaps an Intracellular Lactate Shuttle, to be discovered and elucidated?
1.8. The intracellular, cytosol-to-mitochondrial lactate shuttle
Observations of lactate exchange between cells, tissue, and organs in humans and other mammals52,53 provided impetus for discovery of intracellular lactate shuttling. Studies on humans38,39 and other mammals35,54,55 revealed that most lactate was disposed of via intramuscular oxidation, a result inverse of the one-fifth/four-fifths theory of O2Debt (vide supra). Lactate disposal via oxidation was particularly prominent when muscles and heart were engaged and blood flow, oxygen consumption, and metabolite flux rates were high.9,10,45,46 With that realization, issues arose about where within a working muscle fiber or beating heart cell lactate was oxidized. At the time, much thinking about this issue was that the first step in lactate oxidation to pyruvate occurred in the cytosol, but for several reasons that idea made no sense. Beating heart46 and working red muscle45,56 simultaneously consume glucose and take up and oxidize glucose and lactate. Additionally, measurements of the lactate (L) to pyruvate (P) ratio (L/P) were inconsistent with the one-fifth/four-fifths theory of lactate disposal. In a resting person, the L/P in leg muscle venous blood ranged from 10 to 20;29,57 however, when muscles contracted to achieve a moderate exercise power output, the L/P in venous effluent of working muscle rose more than an order of magnitude (i.e., L/P >500).57 Because of the presence LDH in erythrocytes and lung parenchyma,58,59 relative rise of the L/P in arterial blood of exercising individuals is significant but blunted.9,29,57 Given these data, the notion that lactate oxidation occurs in the cytosol was implausible. If not in the cytosol, where, then, does lactate oxidation commence? Where else, but in the mitochondrial reticulum!
2. The mitochondrial lactate oxidation complex (mLOC)
Controversy surrounds the idea of mitochondrial lactate oxidation; some investigators have produced mitochondrial preparations that oxidize lactate,60, 61, 62, 63, 64, 65 whereas some others66, 67, 68 have failed in the attempt. Conceptually the issue is resolved if one realizes that the cellular respiratory apparatus is not located in discrete, capsular-shaped organelles (i.e., mitochondria), but rather in an extensive network, the mitochondrial reticulum.69,70 Hence, attempts to isolate mitochondria for ex vivo respiratory studies inevitably results in disruption of the mitochondrial reticulum69,71 and lability and fragility of mitochondrial constituents such as cytochrome c and LDH. Still, it is possible to show lactate oxidation in mitochondrial preparations from mammalian muscle, including human skeletal muscle.72 Additionally, the presence of muscle mitochondrial lactate oxidation can be demonstrated by magnetic resonance spectroscopy.73,74 Moreover, studies of muscle cells and tissues using confocal laser scanning microscopy, immunoprecipitation and immunohistochemistry show the presence of LDH in the mLOC.75,76 Seemingly, then, the issue of mitochondrial lactate oxidation has been resolved. In retrospect, the inability of some investigators to produce mitochondrial preparations that do respire lactate is not unique. A similar problem with the ability of muscle mitochondrial preparations to oxidize long chain fatty acids has been ascribed to use of the proteolytic enzyme nagarse, which increases muscle mitochondrial protein yield, but the resulting preparations lose the ability to oxidize fatty acids or their carnitine derivatives.77 In this instance, the inability of mitochondria preparations to recapitulate what is known to happen in vivo can be ascribed to isolation artifact. It is the same with mitochondrial preparations which lose LDH during isolation and are unable to oxidize lactate. Losing or knocking out mLDH, an essential part of the mLOC, only proves that mLDH is essential for mitochondrial lactate oxidation.
In vivo tissue lactate production, uptake, and oxidation occur because glycolysis and mitochondrial respiration occur simultaneously. For the mitochondrial reticulum to oxidize lactate, the reticulum contain a transporter or transporters64,78 and LDH,62,75 both of which were found as soon as probed for using immunohistochemistry and immunocoprecipitation.75,76,79 LDH is now listed in mitochondrial constituent databases such as the MitoCarta80,81 (https://www.broadinstitute.org/scientific-community/science/programs/metabolic-disease-program/publications/mitocarta/mitocarta-in-0) and MitoMiner (http://mitominer.mrc-mbu.cam.ac.uk/release-4.0/begin.do).
The first clues to the structure of mLOC (Fig. 6) and mitochondrial ability to oxidize lactate as well as pyruvate was deduced based on the functionality of mitochondrial preparations and a single report in the literature on how to mitigate the effects of LDH-contaminated mitochondrial preparations.82 To oxidize lactate mitochondrial preparations requires all the usually understood components (pyruvate dehydrogenase, Krebs cycle enzymes, components of the electron transport chain), a lactate transporter, and LDH. Upon investigation, mitochondrial preparations from rat and human muscle were found to contain monocarboxylate transporter isoform 1 (MCT1), its membrane chaperone basigin (BSG or CD147), LDH and cytochrome oxidase.75 Subsequently, essential elements of the mLOC were found in mitochondrial preparations from liver,63,65 brain,76 and various model systems, such as brain slices,83 primary neuronal cultures,76,84 normal breast, and transformed breast cancer cells28 and tumors.85 Immunohistologic evidence of the presence of mLOC is presented in Fig. 7. At the time of discovery, it was known that, while lactate was preferred over pyruvate as an oxidizable substrate, monocarboxylate transporters (MCTs) could also transport pyruvate and β-hydroxybutyrate,86,87 but a unique mitochondrial pyruvate transporter had not been identified.
Fig. 6.

A schematic showing the putative mitochondrial lactate oxidation complex (mLOC). The lactate-pyruvate transporter (MCT1) is inserted into the mitochondrial inner membrane, strongly interacting with its chaperone protein CD147, and is also associated with cytochrome oxidase (COx) as well as mitochondrial lactate dehydrogenase (mLDH), which could be located at the outer side of the inner membrane. Lactate, which is always produced in the cytosol of muscle and other tissues because of the abundance, activity, and characteristics of cytosolic LDH, is oxidized to pyruvate via the lactate oxidation complex in mitochondria of the same cell. This endergonic lactate oxidation reaction is coupled to the exergonic redox change in COx during mitochondrial electron transport. ETC = electron transport chain; GP = glycerol phosphate; Mal-Asp = malate-aspartate; MCT = monocarboxylate (lactate) transporter; mPC = mitochondrial pyruvate carrier; TCA = tricarboxylic acid. Redrawn from Ref. 75 with permission.
Fig. 7.

Immunohistochemical images demonstrating some components of the mitochondrial lactate oxidation complex (mLOC) in L6 cells. The mLOC contains the inner mitochondrial membrane cytochrome oxidase (COX), the lactate-pyruvate transporter (MCT1), lactate dehydrogenase (LDH) and the MCT anchoring protein, CD147 (Basigin). MCT1 was detected at both sarcolemmal and intracellular domains (A-1). Mitochondrial reticulum (MR), identified by MitoTracker, was extensively elaborated in L6 cells (A-2). The merged images of MCT1 (green, A-1) and MR (red, A-2) showed intense yellow, indicating co-localization of MCT1 and components of the MR, particularly at perinuclear cell domains (A-3). (B) LDH (B-1) and COX (B-2) are imaged. Superposition of signals for LDH (red, B-1) and COX (green, B-2) shows co-localization of LDH in the MR (yellow) of muscle cells (B-3). Depth of field approximately 1 μm, scale bar = 10 μm. Similar data have been obtained on rat plantaris leg muscles in vivo. From Ref. 75 with permission.
2.1. Mitochondrial lactate and pyruvate carriers
Following reports of discovery of the mitochondrial pyruvate carrier (mPC),88,89 and with access to our own custom antibodies to MCT1 as well as commercially available antibodies to the putative mPC, we obtained images assessing co-localization of MCT1 and mPC in L6 cells. In those preliminary studies, co-localization analysis of mMCT1 and mPC1 in Imaris software showed an r2 of 0.8. It appears that both MCT1 and mPC are co-localized to the mitochondria (r2 = 0.8) (Fig. 8). However, at the light microscopic level it is impossible to know if mMCT and mPC interact physically and functionally. Immunocoprecipitation, x-ray crystallography, mass spectrometry, and protein deletion (knockout) studies are needed to definitively answer questions about mMCT and mPC co-localization and functionality and the role of the mPC in mitochondrial lactate oxidation.
Fig. 8.

First images assessing co-localization of the monocarboxylate (lactate, pyruvate, β-hydroxybutyrate) transporter (MCT1) and mitochondrial the pyruvate carrier (mPC) in L6 cells, which shows the localization of DAPI-positive nuclei (A), MCT1 (B), mPC1 (C), and Mito Tracker-positive MR (D) in L6 cells. The merged images are shown in E. Co-localization analysis of mPC1 (C) and mitochondria (D) showed a Pearson correlation coefficient (r2) value of 0.8. Co-localization analysis of MCT1 (B) and mPC1 (C) showed an r2 of 0.3, largely because MCT1 occupies sarcolemmal, mitochondrial, and peroxisomal compartments. A channel to represent the co-localization of MCT1 and mitochondria was created to image mMCT1; subsequent co-localization of mMCT1 with mPC1 resulted in an r2 of 0.8 (F). White dots indicate the co-localization of mMCT1 and mPC1 as observed in Image J software. Whole images were contrast enhanced in A, B, C, D, and E. Similar results were observed for mPC2. Scale bar = 20 μm. It appears that both MCT1 and the putative mPC are co-localized to the mitochondria (r2 = 0.8). However, at the light microscopic level, it is impossible to know if the 2 proteins interact physically and functionally. Also, with benefit of the Orbitrap liquid chromatography/mass spectrometry device, we would be able to determine fractional synthesis rates of mitochondrial lactate oxidation complex and mPC proteins.
Finally, with regard to the site of intramuscular lactate oxidation, Gladden and associates,18 in an attempt to integrate ideas about the intracellular lactate oxidation, modified Fig. 6 to include the hypothesis that mitochondrial lactate oxidation to pyruvate occurred in the cytosol or mitochondrial intermembrane space. However, that conclusion is inconsistent with what we28,76 and others63,65,84 have found concerning the location of mitochondrial LDH.
Future research efforts to better define mitochondrial lactate and pyruvate oxidation complexes, perhaps using Betzig-type super-resolution microscopy, electron microscopy, or hyperpolarized C studies,13 are eagerly anticipated. Moreover, all extant models may prove to be too simplistic. For instance, others have proposed90 that intracellular lactate shuttling is associated with the well-known malate–aspartate shuttle. Because redundancy in regulation is a physiologic principle, we shall not be surprised if intracellular lactate shuttling will eventually be shown to be accomplished by multiple mechanisms in vivo.
2.2. Other Lactate Shuttle and other roles of lactate
Because of the recognition of the presence of lactate shuttling within and among various cells, tissues and organs such as muscle, heart, and liver,6,7,43,44 we and others have extended the concept to include other cells, tissues, and organs such as the brain,90,91 lungs,58,59 sperm,92,93 adipose,94, 95, 96 and peroxisomes.97 Descriptions of these lactate shuttles are described elsewhere.23
3. Lactate signaling, lipolysis, fatty acid oxidation, glucose tolerance, and inflammation
An inverse relationship between blood and plasma free fatty acid (FFA) concentration and oxidation has long been recognized,98 but the associations are under-appreciated (Fig. 9). In the 1960s, Issekutz and colleagues99,100 noted the effect of lactacidemia on diminishing circulating FFA in dogs and humans during hard exercise, and lactate infusion into running dogs caused circulating FFA to decline.99,101,102 In their work, these investigators could clearly observe an effect of the lactate on circulating FFA, but whether the mechanism was an inhibition of lipolysis or a stimulation or re-esterification was not addressed.
Fig. 9.

Illustration of how lactatemia affects blood (glucose) and peripheral glucose uptake as well as the production, uptake and oxidation of FFA, giving rise to metabolic inflexibility in muscle. Lactate is the inevitable consequence of glycolysis,18 the minimal muscle lactate (L) to pyruvate (P) ratio (L/P) being 10 and rising to an L/P of >100 when glycolytic flux is high.57 Lactate is the favored oxidizable substrate and provides product inhibition of glucose and FFA oxidation. As the products of glycolysis, lactate and pyruvate provide negative feedback inhibition of glucose disposal (blue dashed lines). Also, as the predominant mitochondrial substrate, lactate gives rise to acetyl-coenzyme A (CoA), and in turn malonyl-CoA. Acetyl-CoA inhibits β-ketothiolase and, hence, β-oxidation, while malonyl-CoA inhibits mitochondrial FFA-derivative uptake via CPT1 (T).199 Moreover, lactate is the main gluconeogenic precursor raising glucose production and blood (glucose) (red lines). Via GPR81 binding, lactate inhibits lipolysis in WAT (T), depressing circulating FFA.96,104 This model explains the paradoxical presence of lactatemia in high-intensity exercise and insulin-resistant states with limited ability to oxidize fat (green lines). Modified from.76 CPT1 = carnitine palmitoyl transporter-1; FAT = fatty acid translocator comprised of CD36 and FABPc; FFA = free fatty acid; GLUT = glucose transporter; m = mitochondrial; Malonyl = CoA formed from exported TCA citrate controlled by the interactions of malonyl-CoA decarboxylase (MCD) and acetyl-CoA carboxylase (ACC); MCT = monocarboxylate transporter; mPC = mitochondrial pyruvate transporter; PDH = pyruvate dehydrogenase; s = sarcolemmal; T = inhibition; WAT = white adipose tissue. Not shown is fatty acyl-Co (FA-CoA) that will accumulate if FFAs are taken up by myocytes, but blocked from mitochondrial entry by the effect of malonyl-CoA on CPT1. Accumulated intracellular FA-CoA will give rise to intramyocellular triglyceride (IMTG) and the formulation of LC-FA, DAG, and ceramides via inhibition of PI3 Kinase (PI3-K) and reducing GLUT4 translocation; from Ref. 23 with permission.
The mechanism by which lactatemia suppresses circulating FFA is now known to be due to suppression of adipose lipolysis. Recently, several groups of investigators94, 95, 96,103 have shown that, independent of pH or sodium ions, lactate inhibits lipolysis in fat cells through activation of a previously identified orphan G-protein coupled receptor, now termed hydroxycarboxylic acid receptor 1 (HCAR-1). In mouse, rat, and human adipocytes, HCAR-1 appears to act as a lactate sensor with the inhibitory effect on lipolysis operating through cyclic adenosine monophosphate and cyclic adenosine monophosphate response element binding.104, 105, 106
Most recently, a large international group of investigators has expanded knowledge of the role of lactate signaling via transforming growth factor β (TGF-β2) secreted from adipose.107 TGF-β is a multifunctional cytokine belonging to the TGF superfamily that includes 3 different mammalian isoforms (TGF-β1 to TGF-β3). Because of its role in immune and stem cell regulation and differentiation, TGF-β2 is a highly researched cytokine in the fields of cancer, auto-immune and infectious diseases, disruption of the blood–brain barrier in epilepsy, aging, and TBI.108 While all TGF-β isoforms are known to be secreted by white blood cells, Takahashi et al.107 showed that after endurance training, adipose of mice secreted TGF-β2 in response to lactate signaling. In turn, TGF-β2 improved glucose tolerance in mice, leading the authors to conclude that exercise training improves systemic metabolism through an inter-organ (adipose to liver) communication via a “lactate–TGF-β2 signaling cycle”.
On first impression, one might think that the effects of lactate on HCAR-1 inhibiting lipolysis and mitochondrial lactate oxidation (Fig. 6)96 are contradictory with increasing glucose tolerance via TGF-β2.107 However, a reasonable alternative explanation is that the effects of lactate on HCAR-1 in adipose are acute (as occurs during hard exercise) and that the effects of TGF-β2 are long term (as occurs during recovery from exercise) when glucose tolerance and lipid oxidation are improved in men and women.109 While the purported effects of lactate signaling HCAR-1 and TGF-β2 observed in rodent models await validation in humans, for the present it is certain that lactate both inhibits lipolysis and mitochondrial FFA oxidation and stimulates mitochondrial biogenesis and glucose tolerance and lipid oxidation in humans in vivo.
Beyond a role in inhibiting lipolysis, lactate also plays an important role in limiting inflammation following injury. Consequently, lactate-containing solutions are being evaluated as anti-inflammatory resuscitation fluids for use in a variety of other therapies, including acute pancreatitis,104,110 hepatitis,104 and dengue fever.21 Compared to normal saline, lactate-containing resuscitation solutions offer the advantage of providing calories in addition to fluid and electrolytes. However, it was not until Hoque and colleagues104 found that lactate binding to HCAR-1 negatively regulates toll-like receptor induction of the pyrin-domain-containing protein 3 inflammasome and production of Interleukin (IL)-1β, via Arrestin β 2. It was then appreciated that lactate and HCAR-1 were involved in suppression of inflammation in patients with acute organ injury. For the treatment of brain swelling and elevated intracranial pressure following TBI, hypertonic sodium-L-lactate, a hypertonic solution, can help manage intracranial pressure by its osmotic effects.111, 112, 113
4. The dark side of lactate metabolism—When lactate production and accumulation may or may not be efficacious
Some table salt improves the taste of many foods, but too much salt can ruin a soup or entrée. Similarly, insulin and insulin action are keys to metabolic flexibility and health. However, too much exogenous insulin can be lethal. So it is with lactate accumulation, metabolism, and supplementation.
4.1. Glucose–lactate interactions and cancer metabolism
Warburg and Minami114 first described the metabolic phenotype characteristic of cancer cells. They noted high glucose uptake and excessive lactate formation in cancer cells even under fully oxygenated conditions. This discovery was subsequently named the Warburg Effect.115 While the high glucose uptake/lactate release phenotype remains a hallmark of cancer,116,117 today there is no consensus on the meaning of the Warburg Effect. The excessive lactate formation of cancer cells and tumors led Warburg to propose that cancer was an injury to the cellular respiratory apparatus. More recently, however, we28 and others118 observed that cancer cell mitochondria are capable of respiring with lactate. In a recent review, San-Millán and Brooks119 described many similarities between cancer and healthy exercise phenotypes. Consequently, we proposed that augmented lactate production (lactagenesis) initiated by gene mutations is the reason and purpose of the Warburg Effect and that dysregulated lactate metabolism and signaling are key elements in carcinogenesis.119 Specifically, we identified the following steps by which lactagenesis may support carcinogenesis: angiogenesis, immune escape, cell migration, metastasis, and self-sufficient metabolism. Justification for our hypothesis that dysregulated lactate metabolism and signaling are key elements in carcinogenesis are presented separately.23,119 In this regard, it is to be noted that we are not the only exercise physiologists and biochemists to have commented on the meaning of exaggerated lactate shuttling in cancer.120
4.2. Blocking lactate shuttles in cancer
Since Sonveaux et al.85 in the Feron lab identified lactate shuttling in tumors, and as commented on by Semenza,121 there have been serious attempts to repress tumorigenesis by blocking the release of lactate from glucose-consuming and highly glycolytic cells and cells respiring lactate. That cancer cells respire with lactate drawn from the tumor microenvironment is an important realization in itself.28,118 However, oxidative lactate disposal within tumors also sets up the concentration gradient necessary for lactate shuttling. Following the lead of Sonveaux et al.,85 the search is on to develop MCT1 and MCT4 inhibitors.85,122, 123, 124, 125 However, the lack of MCT specificity has been a problem, even for investigators in big pharma.126 Because a quest to find cancer-specific MCT blockers has as yet been unsuccessful, others are looking for alternative approaches to blocking lactate shuttling in tumors and cancer, such as by limiting the expression of CD147, the scaffold for MCT insertion into cell membranes (vide supra).127, 128, 129, 130
4.3. Exercise, lactatemia, and carcinogenesis
Endurance training and cancer phenotypes have a lot in common, including the presence of high glycolytic rates and lactate production and accumulation.119 Accordingly, it is of concern that elevated circulating lactate levels resulting from high-intensity interval exercise training can provoke cancer-prone cells to transform. Fortunately, epidemiologic studies support the idea that regular physical activity reduces the risk of many common cancers, including cancer of the breast, colon, bladder, uterus, esophagus, kidney, lung, and stomach.131 It is noteworthy that the organs of cancer have apparently little to do with regard to exercise itself, suggesting the systemic circulation of a protective cytokine, myokine, adipokine, or metabolite during exercise. Given this observation, a suggestion is that intermittent lactate release and circulation of lactate during physical activity improves lactate clearance and preconditions cells, tissues, and organs for reducing the chance that lactagenesis promotes carcinogenesis.119
4.4. Lactate insulin and hypoglycemia
Lactate–glucose interactions can be complex, and interference with lactate shuttling by glucose-insulin signaling can be disruptive. As recognized in the Lactate Shuttle6,43 and Cori Cycle,26 glucose and glycogen are the precursors to lactate formation,7,132 and lactate is the major gluconeogenic precursor.13,26,36,133,134 However, whereas the blood glucose level provides important feedback in the regulation of insulin and counter-regulatory hormones, lactate normally plays only a small role in the regulation of insulin secretion, and is, by nature's design, excluded from the regulatory processes.
MCTs are bidirectional symporters facilitating movement of protons and lactate anions down concentration gradients. While MCTs are ubiquitous and scaffolded in plasma membranes of most cells, including erythrocytes cells in the heart, muscle, and brain,8,135,136 via small interfering RNAs MCTs are excluded from insertion into pancreatic β-cell plasma membranes.137,138 In pancreatic β-cells, MCT expression is silenced to keep extracellular lactate from affecting intracellular redox and thereby interfering with glucose sensing and insulin secretion.139 The silencing of MCT1 in pancreatic β-cells is evolutionary proof of how lactate overrides glucose in regulating energy substrate partitioning in general and insulin secretion in particular when the dominant role of lactate must be suppressed. In this regard, it is noteworthy that, in persons with failed suppression of pancreatic β-cells, MCT expression becomes hypoglycemic during hard exercise. If MCT1 is not silenced, lactate will gain entry into pancreatic β-cells and affect cell redox, just as if blood glucose were elevated. Consequently, individuals in whom pancreatic β-cell MCT expression is not silenced during exercise experience lactatemia and become hypoglycemic due to hyperinsulinemia and increased glucose disposal through metabolism.140
4.5. The “Anaerobic Threshold”
The exercise intensity at which a rise in arterial blood lactate concentration commences is the “Anaerobic Threshold” (AT). At one time, the AT was reasonably associated with a limitation in working muscle oxygen delivery, which resulted in a Pasteur effect involving augmented glycolysis due to oxygen limitation.25,141 The idea was straightforward and at the time was based on the widely accepted O2Debt theory.24 However, because the AT concept assumed that lactatemia during exercise was due to increased lactate production rather than an imbalance between lactate production (appearance in the blood, Ra) and lactate disposal (disappearance from the blood, Rd),9,142 the AT concept has been severely challenged.43,143 Importantly, and regrettably, the AT concept failed because of the recognition that muscle produced and released lactate9,38,45,142 under fully aerobic conditions.144 Still, in the hands of knowledgeable and skilled clinicians, AT testing can be part of an armamentarium in dissecting changes in blood lactate Ra due to chronic obstructive pulmonary disease, pulmonary hypertension, cardiovascular disease, mitochondrial defects, catecholamine or other endocrine effects,145 pharmacologic toxicity,146 altered carbohydrate nutrition,147 environmental effects,148 or other effects.
4.6. Lactate–brain fuel after TBI
In comatose TBI patients, the role of lactate in brain fueling was found to be impressive because most (70%–80%) of circulating blood glucose was produced via gluconeogenesis from lactate.11,12 Also, net lactate uptake provided 12% of brain fuel. Hence, most (57%) brain fuel was from lactate, either directly (12%) or indirectly via gluconeogenesis (45%).
In healthy humans, plasma lactate levels during exercise can increase by an order of magnitude or more. For example, with a 10-fold rise in concentration, net lactate uptake provides 25% of total brain energy needed.149,150 Disregarding for a moment that cerebral disposal of glucose is accomplished by conversion to lactate, preference of cerebral lactate over glucose has been found in studies on rats151 and humans,150,152 and the provision of lactate to a healthy brain decreases glucose net uptake. This observation indicates an important cerebral lactate shuttling phenomenon. Alternatively, some might consider that lactate can serve as an alternative brain fuel. However, as emphasized by Schurr, lactate is the brain fuel because, regardless of whether extracellular glucose or lactate is taken up by brain, the path of glucose disposal is through conversion to lactate and a Cell-Cell Lactate Shuttle.153,154
4.7. Lactate and brain-derived neurotrophic factor
In addition to serving as a cerebral energy substrate, circulating lactate can signal secretion of cerebral brain-derived neurotrophic factor (BDNF). As discussed in part elsewhere in this work, circulating blood lactate can be raised during exercise by production in muscle, the integument and other epinephrine sensitive tissues. Also, circulating lactate level can be raised during rest or exercise by vascular L-lactate infusion into the systemic circulation. Indeed, infused sodium lactate raises circulating BDNF levels.155 BDNF is a member of the neurotrophic family of proteins and facilitates neurogenesis, neuroprotection, neuroregeneration and synaptic plasticity, as well as formation, retention, and memory recall.156 BDNF is produced both in the central nervous system and in other tissues, including the vascular endothelium. High levels of BDNF messenger RNA are found in the hippocampus and in the cerebral cortex, and in rodent models physical exercise is the strongest known stimulus to BDNF expression in the hippocampus.157 In contrast, attenuated expression of BDNF messenger RNA in the hippocampus may constitute a pathogenic factor common to Alzheimer's disease and major depression.158 Circulating BDNF is typically elevated in exercise, but levels are reduced in patients with depression and type 2 diabetes.159 Even though acute exercise increases BDNF production in the hippocampus and cerebral cortex, studying the effects of exercise on BDNF expression in the human brain is difficult. In what may become noted as a classic study in neurochemistry, Seifert et al.156 compared BDNF levels in arterial and internal jugular vein blood and demonstrated the effects of exercise and exercise training on cerebral BDNF net release.
While many posit a positive role for BDNF in terms of cognition, it is important to know also that lactate therapy has been shown to raise BDNF levels and improve cognitive function in rodent models following brain injury.160, 161, 162 Most recently, in an extraordinary set of experiments on healthy men who volunteered to exercise at high intensities with arterial and jugular bulb catheters in place, Hashimoto et al.163 showed that executive function was directly correlated to blood (lactate) and cerebral lactate uptake. Wang et al.164 also recently provided data showing that optogenetic activation of astrocytes in the anterior cingulate cortex triggers lactate release and improves decision making in rats with chronic visceral pain. As predicted in papers resulting from studies on rodents164,165 and healthy and injured humans,11 Hashimoto et al.163 found that brain fueling with lactate improved cerebral functioning. Additionally, with respect to cognition, it has been shown that long-term memory consolidation in rat hippocampus relies on lactate and the Astrocyte-Neuron Lactate Shuttle (ANLS).166,167
5. A role for lactate supplementation in sepsis
Although we have discussed numerous instances in which it may be clinically beneficial to achieve elevated blood lactate concentrations, it is noteworthy to mention that a blood lactate value of 4 mmol/L is a biomarker of severe sepsis.168,169 Interestingly, the lactate threshold in exercise frequently corresponds with 4 mmol/L blood lactate.170 What then is the significance of a slightly elevated blood lactate concentration (e.g., >2.0 mmol/L) in a febrile patient?171 Concern over the prospect of a blooming bacterial, viral, or fungal attack leading to sepsis needs to be appreciated from the context of the huge lactate clearance capacity in a healthy person.172 Also, lactatemia is unlikely to be due to oxygen insufficiency (hypoxia) in either exercise23,143 or sepsis.20,173,174 Alternatively, is the body readying for some challenge by raising the level of a key myocardial51 and whole body energy substrate,7 a gluconeogenic precursor13,36,44 or an anti-inflammatory moiety?104,110 In realizing the importance of lactate in supporting metabolism, some have proposed that lactated Ringers solution be used as a resuscitation fluid in sepsis,174 and, by extension of the same thinking, that the use of isotonic or hypertonic sodium-L-lactate and Sanguisal™ (a physiologically balanced mixture of Na+-, K+-, Mg2+-, and Ca2+-lactate plus phosphate buffer) also needs to be evaluated for resuscitation in sepsis.
6. Conclusion
Studies on humans and mammals such as rats and dogs during rest and exercise led to discovery of the Lactate Shuttle, a mechanism by which lactate production in rapidly glycolyzing driver cells provides energy substrate for recipient cells where lactate is a fuel energy source, gluconeogenic precursor, and signaling molecule. In heart and red skeletal muscle, lactate disposal is by mitochondrial respiration. In the liver and kidneys, mitochondria play roles in oxidizing lactate to pyruvate and then in the conversion to oxaloacetate and phosphoenolpyruvate via pyruvate carboxylase and phosphoenolpyruvate carboxykinase. In the liver and kidneys, the oxidation of lactate and other fuel sources also provides energy for completion of gluconeogenesis and glycogen synthesis. By changes in cell redox owing to the production and oxidation of lactate, as well as allosteric binding to HCAR-1 and TGF-β signaling, lactate affects numerous processes. Discoveries of intracellular, cell–cell, and organ–organ lactate exchanges has led to articulation of numerous lactate shuttles, including peroxisomal and astrocyte–neuron lactate shuttles, the basis of which are interactive effects between driver producer and recipient consumer cells.
In terms of the organization of energy substrate partitioning in humans and other mammals, lactate is at the fulcrum of intermediary metabolism. Lactate is the inevitable product of glycolysis. With rapid glycolysis, as occurs in hard muscle exercise, lactate limits lipid metabolism in 2 ways. First, lactate released into the blood reaches white adipose tissue, and by binding to HCAR-1, lactate acts to inhibit lipolysis via a cyclic adenosine monophosphate-dependent pathway (CREB), which limits subsequent release of fatty acids into blood. Second, lactate produces an abundance of mitochondrial acetyl-CoA that gives rise to malonyl-CoA, which in turn inhibits carnitine palmitoyl transporter-1 (CPT-1) and blocks mitochondrial FFA uptake.
Recognizing that lactate is produced under full aerobic conditions, as well as understanding the roles of lactate shuttling in biology, is providing new opportunities to treat the ill and injured. For example, exogenous L-lactate vascular infusion is being evaluated in the treatment of heart failure,175, 176, 177 TBI,19,113,178,179 pancreatitis,104,110 hepatitis,104 dengue fever,21 and sepsis.174
Recognizing that lactate, particularly rising blood lactate concentration, is a biomarker for an imbalance between lactate production and removal provides practitioners in diverse fields with important information on physiologic status of athletes and the ill and injured. The applications for this information include providing fluid, energy and electrolyte support to athletes and others,180 and apply to fields as diverse as pulmonary medicine,141 sports medicine,181,182 critical care medicine,173 and oncology.85 Indeed, the recognition that lactate shuttles among driver producer and recipient consumer cells in tumors offers the exciting possibility of reducing carcinogenesis and tumor size by blocking either or both producer and recipient arms of lactate shuttles within and among tumor cells. With growing appreciation of the importance and extent of lactate shuttling in vivo, it is to be anticipated that by controlling blood (lactate) via a closed loop, a continuous monitoring system can improve care of the ill, injured, and malnourished.183, 184, 185
In closing, profound thanks are offered to those students, postdoctoral fellows, visiting scientists and University of California at Berkeley colleagues who participated in the discovery of lactate shuttling; those contributors are identified in the references, typically as first authors. Also, it is important to fully recognize the many contributions of the American College of Sports Medicine (ACSM) and to recognize that our contributions, as described in this paper, occurred within the context of a greater whole. It is risky to limit the examples of how ACSM colleagues have advanced science and medicine, since other important contributions are easily and unintentionally omitted. Nevertheless, contributions of the following individuals to physiology, and sports medicine should not be overlooked: Jere Mitchell, in cardiovascular physiology and medicine;186 John Holloszy, in mitochondrial biogenesis;187 L. Bruce Gladden, in the regulation of intermediary metabolism;18,188 Jim Barnard, V. Reggie Edgerton, and Ken Baldwin, in muscle fiber heterogeneity;47,48 Scott Powers, in redox signaling and preconditioning of the heart to cardiovascular insults;189 Karl Wasserman and Jerry Dempsey, in pulmonary physiology and medicine;25,190 Tom Fahey, in translating the lessons of physiology and sports medicine to the literature for health care professionals and the lay public;191, 192, 193 and Tim White, in the administration of higher education (https://www2.calstate.edu/csu-system/chancellor/Pages/meet-the-chancellor.aspx). In the aggregate, our scientists, clinicians, and staff at the ACSM know well how the body works under stress, in illness, and after injury. Thus, collectively we can think of and accomplish things that few others could imagine. One such example is an idea from exercise physiology that has wide applicability to science and medicine—the Lactate Shuttle—in which the product of one process, glycolysis, is the substrate for another process, oxidative metabolism. The linkage between the two is lactate, the metabolite that can transcend cellular, tissue and organ compartment barriers to be a preferred fuel energy source, the major gluconeogenic precursor and a powerful signaling molecule.
Acknowledgments
Acknowledgments
This review is due to the combined efforts of all students, fellows, visiting scientists, colleagues, and human subjects associated with GAB. Appreciation is also expressed to the officers and staff of the American College of Sports Medicine (ACSM) for the privilege of delivering the 2019 J.B. Wolffe Memorial Lecture. Supported by NIH R01 AG059715-01 and Pac-12 Conference CAL-Brooks-17-02 and the University of California at Berkeley's Center for Research and Education on Aging (CREA).
Competing interests
The author declares that he has no competing interests.
Footnotes
This paper represents a text version of the Joseph B. Wolffe Lecture to the American College of Sports Medicine, May 28, 2019, in Orlando, FL. ICU stands for intensive care unit. Accompanying video adapted with permission of the American College of Sports Medicine © American College of Sports Medicine 2019. ICU stands for Intensive Care Unit.
Peer review under responsibility of Shanghai University of Sport.
Supplementary materials
Supplementary material associated with this article can be found in the online version at doi:10.1016/j.jshs.2020.02.006.
Appendix. Supplementary materials
References
- 1.Meyerhof O. Uber's energy conversions in muscle II. The € Fate of the milk sour in the recovery period of the muscle (Über die Energieumwandlungen im Muskel II. Das Schicksal der Milchsaure in der Erholungsperiode des Muskels) Pflügers Archiv ges Physiol Mensch Tiere. 1920;182:284–317. [in German] [Google Scholar]
- 2.Hill A.V. Muscular activity and carbohydrate metabolism. Science. 1924;60:505–514. doi: 10.1126/science.60.1562.505. [DOI] [PubMed] [Google Scholar]
- 3.Mitchell P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 1961;191:144–148. doi: 10.1038/191144a0. [DOI] [PubMed] [Google Scholar]
- 4.Chance B., Williams G.R. Respiratory enzymes in oxidative phosphorylation. VI. The effects of adenosine diphosphate on azide-treated mitochondria. J Biol Chem. 1956;221:477–489. [PubMed] [Google Scholar]
- 5.Huxley H.E. Recent x-ray diffraction and electron microscope studies of striated muscle. J Gen Physiol. 1967;50(Suppl.):S71–83. doi: 10.1085/jgp.50.6.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brooks G.A. Circulation, respiration, and metabolism: current comparative approaches. Springer; Berlin, Heidelberg: 1984. Glycolytic end product and oxidative substrate during sustained exercise in mammals–the “lactate shuttle; pp. 208–218. [Google Scholar]
- 7.Brooks G.A. Lactate shuttles in nature. Biochem Soc Trans. 2002;30:258–264. doi: 10.1042/. [DOI] [PubMed] [Google Scholar]
- 8.Brooks G.A. Cell-cell and intracellular lactate shuttles. J Physiol. 2009;587:5591–5600. doi: 10.1113/jphysiol.2009.178350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bergman B.C., Wolfel E.E., Butterfield G.E., Lopaschuk G.D., Casazza G.A., Horning M.A. Active muscle and whole body lactate kinetics after endurance training in men. J Appl Physiol (1985) 1999;87:1684–1696. doi: 10.1152/jappl.1999.87.5.1684. [DOI] [PubMed] [Google Scholar]
- 10.Gertz E.W., Wisneski J.A., Stanley W.C., Neese R.A. Myocardial substrate utilization during exercise in humans. Dual carbon-labeled carbohydrate isotope experiments. J Clin Invest. 1988;82:2017–2025. doi: 10.1172/JCI113822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Glenn T.C., Martin N.A., Horning M.A., McArthur D.L., Hovda D.A., Vespa P. Lactate: brain fuel in human traumatic brain injury: a comparison to normal healthy control subjects. J Neurotrauma. 2015;32:820–832. doi: 10.1089/neu.2014.3483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Glenn T.C., Martin N.A., McArthur D.L., Hovda D.A., Vespa P., Johnson M.L. Endogenous nutritive support following traumatic brain injury: peripheral lactate production for glucose supply via gluconeogenesis. J Neurotrauma. 2015;32:811–819. doi: 10.1089/neu.2014.3482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bergman B.C., Horning M.A., Casazza G.A., Wolfel E.E., Butterfield G.E., Brooks G.A. Endurance training increases gluconeogenesis during rest and exercise in men. Am J Physiol Endocrinol Metab. 2000;278:E244–E251. doi: 10.1152/ajpendo.2000.278.2.E244. [DOI] [PubMed] [Google Scholar]
- 14.Hashimoto T., Hussien R., Oommen S., Gohil K., Brooks G.A. Lactate sensitive transcription factor network in L6 cells: activation of MCT1 and mitochondrial biogenesis. FASEB J. 2007;21:2602–2612. doi: 10.1096/fj.07-8174com. [DOI] [PubMed] [Google Scholar]
- 15.de Paoli F.V., Overgaard K., Pedersen T.H., Nielsen O.B. Additive protective effects of the addition of lactic acid and adrenaline on excitability and force in isolated rat skeletal muscle depressed by elevated extracellular K+ J Physiol. 2007;581:829–839. doi: 10.1113/jphysiol.2007.129049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Robergs R.A., Ghiasvand F., Parker D. Biochemistry of exercise-induced metabolic acidosis. Am J Physiol Regul Integr Comp Physiol. 2004;287:R502–R516. doi: 10.1152/ajpregu.00114.2004. [DOI] [PubMed] [Google Scholar]
- 17.Robergs R.A. Invited review: quantifying proton exchange from chemical reactions - implications for the biochemistry of metabolic acidosis. Comp Biochem Physiol A Mol Integr Physiol. 2019;235:29–45. doi: 10.1016/j.cbpa.2019.04.024. [DOI] [PubMed] [Google Scholar]
- 18.Rogatzki M.J., Ferguson B.S., Goodwin M.L., Gladden L.B. Lactate is always the end product of glycolysis. Front Neurosci. 2015;9:22. doi: 10.3389/fnins.2015.00022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brooks G.A., Martin N.A. Cerebral metabolism following traumatic brain injury: new discoveries with implications for treatment. Front Neurosci. 2015;8:408. doi: 10.3389/fnins.2014.00408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Garcia-Alvarez M., Marik P., Bellomo R. Stress hyperlactataemia: present understanding and controversy. Lancet Diabetes Endocrinol. 2014;2:339–347. doi: 10.1016/S2213-8587(13)70154-2. [DOI] [PubMed] [Google Scholar]
- 21.Somasetia D.H., Setiati T.E., Sjahrodji A.M., Idjradinata P.S., Setiabudi D., Roth H. Early resuscitation of dengue shock syndrome in children with hyperosmolar sodium-lactate: a randomized single-blind clinical trial of efficacy and safety. Crit Care. 2014;18:466. doi: 10.1186/s13054-014-0466-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Einstein A. Letter to Max Born. In: Born I., editor. The Born-Einstein letters. 1st ed. Walker and Company; New York, NY: 1926. [Google Scholar]
- 23.Brooks G.A. The science and translation of lactate shuttle theory. Cell Metab. 2018;27:757–785. doi: 10.1016/j.cmet.2018.03.008. [DOI] [PubMed] [Google Scholar]
- 24.Hill A.V., Lupton H. Muscular exercise, lactic acid and the supply and utilization of oxygen. Q J Med. 1923;16:135–171. [Google Scholar]
- 25.Wasserman K., McIlroy M.B. Detecting the threshold of anaerobic metabolism in cardiac patients during exercise. Am J Cardiol. 1964;14:844–852. doi: 10.1016/0002-9149(64)90012-8. [DOI] [PubMed] [Google Scholar]
- 26.Cori C.F., Cori G.T. Carbohydrate metabolism. Annu Rev Biochem. 1946;15:193–218. doi: 10.1146/annurev.bi.15.070146.001205. [DOI] [PubMed] [Google Scholar]
- 27.Warburg O., Wind F., Negelein E. The metabolism of tumors in the body. J Gen Physiol. 1927;8:519–530. doi: 10.1085/jgp.8.6.519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hussien R., Brooks G.A. Mitochondrial and plasma membrane lactate transporter and lactate dehydrogenase isoform expression in breast cancer cell lines. Physiol Genomics. 2011;43:255–264. doi: 10.1152/physiolgenomics.00177.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wasserman K., Beaver W.L., Davis J.A., Pu J.Z., Heber D., Whipp B.J. Lactate, pyruvate, and lactate-to-pyruvate ratio during exercise and recovery. J Appl Physiol (1985) 1985;59:935–940. doi: 10.1152/jappl.1985.59.3.935. [DOI] [PubMed] [Google Scholar]
- 30.Brooks G.A., Brauner K.E., Cassens R.G. Glycogen synthesis and metabolism of lactic acid after exercise. Am J Physiol. 1973;224:1162–1166. doi: 10.1152/ajplegacy.1973.224.5.1162. [DOI] [PubMed] [Google Scholar]
- 31.Brooks G.A., Gaesser G.A. End points of lactate and glucose metabolism after exhausting exercise. J Appl Physiol Respir Environ Exerc Physiol. 1980;49:1057–1069. doi: 10.1152/jappl.1980.49.6.1057. [DOI] [PubMed] [Google Scholar]
- 32.Brooks G.A., White T.P. Determination of metabolic and heart rate responses of rats to treadmill exercise. J Appl Physiol Respir Environ Exerc Physiol. 1978;45:1009–1015. doi: 10.1152/jappl.1978.45.6.1009. [DOI] [PubMed] [Google Scholar]
- 33.Brooks G.A., Bissell M.J., Bassham J.A. Ion-retardation desalting of blood and other animal tissues for separation of soluble metabolites by two-dimensional chromatography. Anal Biochem. 1977;83:580–588. doi: 10.1016/0003-2697(77)90061-6. [DOI] [PubMed] [Google Scholar]
- 34.Brooks G.A., Donovan C.M. Effect of endurance training on glucose kinetics during exercise. Am J Physiol. 1983;244:E505–E512. doi: 10.1152/ajpendo.1983.244.5.E505. [DOI] [PubMed] [Google Scholar]
- 35.Donovan C.M., Brooks G.A. Endurance training affects lactate clearance, not lactate production. Am J Physiol. 1983;244:E83–E92. doi: 10.1152/ajpendo.1983.244.1.E83. [DOI] [PubMed] [Google Scholar]
- 36.Emhoff C.A., Messonnier L.A., Horning M.A., Fattor J.A., Carlson T.J., Brooks G.A. Gluconeogenesis and hepatic glycogenolysis during exercise at the lactate threshold. J Appl Physiol (1985) 2013;114:297–306. doi: 10.1152/japplphysiol.01202.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Emhoff C.A., Messonnier L.A., Horning M.A., Fattor J.A., Carlson T.J., Brooks G.A. Direct and indirect lactate oxidation in trained and untrained men. J Appl Physiol (1985) 2013;115:829–838. doi: 10.1152/japplphysiol.00538.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mazzeo R.S., Brooks G.A., Schoeller D.A., Budinger T.F. Disposal of blood [1-13C]lactate in humans during rest and exercise. J Appl Physiol (1985) 1986;60:232–241. doi: 10.1152/jappl.1986.60.1.232. [DOI] [PubMed] [Google Scholar]
- 39.Stanley W.C., Gertz E.W., Wisneski J.A., Morris D.L., Neese R.A., Brooks G.A. Systemic lactate kinetics during graded exercise in man. Am J Physiol. 1985;249:E595–E602. doi: 10.1152/ajpendo.1985.249.6.E595. [DOI] [PubMed] [Google Scholar]
- 40.Bergman B.C., Butterfield G.E., Wolfel E.E., Lopaschuk G.D., Casazza G.A., Horning M.A. Muscle net glucose uptake and glucose kinetics after endurance training in men. Am J Physiol. 1999;277:E81–E92. doi: 10.1152/ajpendo.1999.277.1.E81. [DOI] [PubMed] [Google Scholar]
- 41.Hooker A.M., Baldwin K.M. Substrate oxidation specificity in different types of mammalian muscle. Am J Physiol. 1979;236:C66–C69. doi: 10.1152/ajpcell.1979.236.1.C66. [DOI] [PubMed] [Google Scholar]
- 42.Molé P.A., Baldwin K.M., Terjung R.L., Holloszy J.O. Enzymatic pathways of pyruvate metabolism in skeletal muscle: adaptations to exercise. Am J Physiol. 1973;224:50–54. doi: 10.1152/ajplegacy.1973.224.1.50. [DOI] [PubMed] [Google Scholar]
- 43.Brooks G.A. Anaerobic threshold: review of the concept and directions for future research. Med Sci Sports Exerc. 1985;17:22–34. [PubMed] [Google Scholar]
- 44.Brooks G.A. Lactate production under fully aerobic conditions: the lactate shuttle during rest and exercise. Fed Proc. 1986;45:2924–2929. [PubMed] [Google Scholar]
- 45.Stanley W.C., Gertz E.W., Wisneski J.A., Neese R.A., Morris D.L., Brooks G.A. Lactate extraction during net lactate release in legs of humans during exercise. J Appl Physiol (1985) 1986;60:1116–1120. doi: 10.1152/jappl.1986.60.4.1116. [DOI] [PubMed] [Google Scholar]
- 46.Gertz E.W., Wisneski J.A., Neese R., Bristow J.D., Searle G.L., Hanlon J.T. Myocardial lactate metabolism: evidence of lactate release during net chemical extraction in man. Circulation. 1981;63:1273–1279. doi: 10.1161/01.cir.63.6.1273. [DOI] [PubMed] [Google Scholar]
- 47.Baldwin K.M., Campbell P.J., Cooke D.A. Glycogen, lactate, and alanine changes in muscle fiber types during graded exercise. J Appl Physiol. 1977;43:288–291. doi: 10.1152/jappl.1977.43.2.288. [DOI] [PubMed] [Google Scholar]
- 48.Barnard R.J., Edgerton V.R., Furukawa T., Peter J.B. Histochemical, biochemical, and contractile properties of red, white, and intermediate fibers. Am J Physiol. 1971;220:410–414. doi: 10.1152/ajplegacy.1971.220.2.410. [DOI] [PubMed] [Google Scholar]
- 49.Peter J.B., Sawaki S., Barnard R.J., Edgerton V.R., Gillespie C.A. Lactate dehydrogenase isoenzymes: distribution in fast-twitch red, fast-twitch white, and slow-twitch intermediate fibers of guinea pig skeletal muscle. Arch Biochem Biophys. 1971;144:304–307. doi: 10.1016/0003-9861(71)90482-6. [DOI] [PubMed] [Google Scholar]
- 50.Johnson J.A., Fusaro R.M. The role of the skin in carbohydrate metabolism. Adv Metab Disord. 1972;60:1–55. doi: 10.1016/b978-0-12-027306-5.50006-1. [DOI] [PubMed] [Google Scholar]
- 51.Bergman B.C., Tsvetkova T., Lowes B., Wolfel E.E. Myocardial glucose and lactate metabolism during rest and atrial pacing in humans. J Physiol. 2009;587:2087–2099. doi: 10.1113/jphysiol.2008.168286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Depocas F., Minaire Y., Chatonnet J. Rates of formation and oxidation of lactic acid in dogs at rest and during moderate exercise. Can J Physiol Pharmacol. 1969;47:603–610. doi: 10.1139/y69-106. [DOI] [PubMed] [Google Scholar]
- 53.Hochachka P.W. Cross-species studies of glycolytic function. Adv Exp Med Biol. 1999;474:219–229. doi: 10.1007/978-1-4615-4711-2_18. [DOI] [PubMed] [Google Scholar]
- 54.Depocas F., Minaire Y., Chatonnet J. Rates of formation and oxidation of lactic acid in dogs at rest and during moderate exercise. Can J Physiol Pharmacol. 1969;47:603–610. doi: 10.1139/y69-106. [DOI] [PubMed] [Google Scholar]
- 55.Freminet A., Bursaux E., Poyart C.F. Effect of elevated lactataemia on the rates of lactate turnover and oxidation in rats. Pflugers Arch. 1974;346:75–86. doi: 10.1007/BF00592652. [DOI] [PubMed] [Google Scholar]
- 56.Zinker B.A., Lacy D.B., Bracy D., Jacobs J., Wasserman D.H. Regulation of glucose uptake and metabolism by working muscle. An in vivo analysis. Diabetes. 1993;42:956–965. doi: 10.2337/diab.42.7.956. [DOI] [PubMed] [Google Scholar]
- 57.Henderson G.C., Horning M.A., Wallis G.A., Brooks G.A. Pyruvate metabolism in working human skeletal muscle. Am J Physiol Endocrinol Metab. 2007;292:E366. doi: 10.1152/ajpendo.00363.2006. [DOI] [PubMed] [Google Scholar]
- 58.Johnson M.L., Emhoff C.A., Horning M.A., Brooks G.A. Transpulmonary lactate shuttle. Am J Physiol Regul Integr Comp Physiol. 2012;302:R143–R149. doi: 10.1152/ajpregu.00402.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Johnson M.L., Hussien R., Horning M.A., Brooks G.A. Transpulmonary pyruvate kinetics. Am J Physiol Regul Integr Comp Physiol. 2011;301:R769–R774. doi: 10.1152/ajpregu.00206.2011. [DOI] [PubMed] [Google Scholar]
- 60.Brandt R.B., Laux J.E., Spainhour S.E., Kline E.S. Lactate dehydrogenase in rat mitochondria. Arch Biochem Biophys. 1987;259:412–422. doi: 10.1016/0003-9861(87)90507-8. [DOI] [PubMed] [Google Scholar]
- 61.Kline E.S., Brandt R.B., Laux J.E., Spainhour S.E., Higgins E.S., Rogers K.S. Localization of L-lactate dehydrogenase in mitochondria. Arch Biochem Biophys. 1986;246:673–680. doi: 10.1016/0003-9861(86)90323-1. [DOI] [PubMed] [Google Scholar]
- 62.Baba N., Sharma H.M. Histochemistry of lactic dehydrogenase in heart and pectoralis muscles of rat. J Cell Biol. 1971;51:621–635. doi: 10.1083/jcb.51.3.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.De Bari L., Atlante A., Valenti D., Passarella S. Partial reconstruction of in vitro gluconeogenesis arising from mitochondrial l-lactate uptake/metabolism and oxaloacetate export via novel L-lactate translocators. Biochem J. 2004;380:231–242. doi: 10.1042/BJ20031981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Brooks G.A., Brown M.A., Butz C.E., Sicurello J.P., Dubouchaud H. Cardiac and skeletal muscle mitochondria have a monocarboxylate transporter MCT1. J Appl Physiol (1985) 1999;87:1713–1718. doi: 10.1152/jappl.1999.87.5.1713. [DOI] [PubMed] [Google Scholar]
- 65.Passarella S., Paventi G., Pizzuto R. The mitochondrial L-lactate dehydrogenase affair. Front Neurosci. 2014;8:407. doi: 10.3389/fnins.2014.00407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Yoshida Y., Holloway G.P., Ljubicic V., Hatta H., Spriet L.L., Hood D.A. Negligible direct lactate oxidation in subsarcolemmal and intermyofibrillar mitochondria obtained from red and white rat skeletal muscle. J Physiol. 2007;582:1317–1335. doi: 10.1113/jphysiol.2007.135095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Rasmussen H.N., van Hall G., Rasmussen U.F. Lactate dehydrogenase is not a mitochondrial enzyme in human and mouse vastus lateralis muscle. J Physiol. 2002;541:575–580. doi: 10.1113/jphysiol.2002.019216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sahlin K., Fernstrom M., Svensson M., Tonkonogi M. No evidence of an intracellular lactate shuttle in rat skeletal muscle. J Physiol. 2002;541:569–574. doi: 10.1113/jphysiol.2002.016683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kirkwood S.P., Munn E.A., Brooks G.A. Mitochondrial reticulum in limb skeletal muscle. Am J Physiol. 1986;251:C395–C402. doi: 10.1152/ajpcell.1986.251.3.C395. [DOI] [PubMed] [Google Scholar]
- 70.Kirkwood S.P., Packer L., Brooks G.A. Effects of endurance training on a mitochondrial reticulum in limb skeletal muscle. Arch Biochem Biophys. 1987;255:80–88. doi: 10.1016/0003-9861(87)90296-7. [DOI] [PubMed] [Google Scholar]
- 71.Glancy B., Hartnell L.M., Combs C.A., Femnou A., Sun J., Murphy E. Power grid protection of the muscle mitochondrial reticulum. Cell Rep. 2018;23:2832. doi: 10.1016/j.celrep.2018.05.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Jacobs R.A., Meinild A.K., Nordsborg N.B., Lundby C. Lactate oxidation in human skeletal muscle mitochondria. Am J Physiol Endocrinol Metab. 2013;304:E686–E694. doi: 10.1152/ajpendo.00476.2012. [DOI] [PubMed] [Google Scholar]
- 73.Chen Y.J., Mahieu N.G., Huang X., Singh M., Crawford P.A., Johnson S.L. Lactate metabolism is associated with mammalian mitochondria. Nat Chem Biol. 2016;12:937–943. doi: 10.1038/nchembio.2172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Park J.M., Josan S., Mayer D., Hurd R.E., Chung Y., Bendahan D. Hyperpolarized 13C NMR observation of lactate kinetics in skeletal muscle. J Exp Biol. 2015;218:3308–3318. doi: 10.1242/jeb.123141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hashimoto T., Hussien R., Brooks G.A. Colocalization of MCT1, CD147, and LDH in mitochondrial inner membrane of L6 muscle cells: evidence of a mitochondrial lactate oxidation complex. Am J Physiol Endocrinol Metab. 2006;290:1237–1244. doi: 10.1152/ajpendo.00594.2005. [DOI] [PubMed] [Google Scholar]
- 76.Hashimoto T., Hussien R., Cho H.S., Kaufer D., Brooks G.A. Evidence for the mitochondrial lactate oxidation complex in rat neurons: demonstration of an essential component of brain lactate shuttles. PloS one. 2008;3:e2915. doi: 10.1371/journal.pone.0002915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Pande S.V., Blanchaer M.C. Preferential loss of ATP-dependent long-chain fatty acid activating enzyme in mitochondria prepared using Nagarse. Biochim Biophys Acta. 1970;202:43–48. doi: 10.1016/0005-2760(70)90216-x. [DOI] [PubMed] [Google Scholar]
- 78.Butz C.E., McClelland G.B., Brooks G.A. MCT1 confirmed in rat striated muscle mitochondria. J Appl Physiol (1985) 2004;97:1059–1066. doi: 10.1152/japplphysiol.00009.2004. [DOI] [PubMed] [Google Scholar]
- 79.Hashimoto T., Masuda S., Taguchi S., Brooks G.A. Immunohistochemical analysis of MCT1, MCT2 and MCT4 expression in rat plantaris muscle. J Physiol. 2005;567:121–129. doi: 10.1113/jphysiol.2005.087411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pagliarini D.J., Calvo S.E., Chang B., Sheth S.A., Vafai S.B., Ong S.E. A mitochondrial protein compendium elucidates complex I disease biology. Cell. 2008;134:112–123. doi: 10.1016/j.cell.2008.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Calvo S.E., Clauser K.R., Mootha V.K. MitoCarta2.0: an updated inventory of mammalian mitochondrial proteins. Nucleic Acids Res. 2016;44:D1251–D1257. doi: 10.1093/nar/gkv1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chretien D., Pourrier M., Bourgeron T., Séné M., Rötig A., Munnich A. An improved spectrophotometric assay of pyruvate dehydrogenase in lactate dehydrogenase contaminated mitochondrial preparations from human skeletal muscle. Clin Chim Acta. 1995;240:129–136. doi: 10.1016/0009-8981(95)06145-6. [DOI] [PubMed] [Google Scholar]
- 83.Schurr A. Lactate: a major and crucial player in normal function of both muscle and brain. J Physiol. 2008;586:2665–2666. doi: 10.1113/jphysiol.2008.155416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Atlante A., de Bari L., Bobba A., Marra E., Passarella S. Transport and metabolism of L-lactate occur in mitochondria from cerebellar granule cells and are modified in cells undergoing low potassium dependent apoptosis. Biochim Biophys Acta. 2007;1767:1285–1299. doi: 10.1016/j.bbabio.2007.08.003. [DOI] [PubMed] [Google Scholar]
- 85.Sonveaux P., Végran F., Schroeder T., Wergin M.C., Verrax J., Rabbani Z.N. Targeting lactate-fueled respiration selectively kills hypoxic tumor cells in mice. J Clin Invest. 2008;118:3930–3942. doi: 10.1172/JCI36843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Roth D.A., Brooks G.A. Lactate transport is mediated by a membrane-bound carrier in rat skeletal muscle sarcolemmal vesicles. Arch Biochem Biophys. 1990;279:377–385. doi: 10.1016/0003-9861(90)90505-s. [DOI] [PubMed] [Google Scholar]
- 87.Roth D.A., Brooks G.A. Lactate and pyruvate transport is dominated by a pH gradient-sensitive carrier in rat skeletal muscle sarcolemmal vesicles. Arch Biochem Biophys. 1990;279:386–394. doi: 10.1016/0003-9861(90)90506-t. [DOI] [PubMed] [Google Scholar]
- 88.Herzig S., Raemy E., Montessuit S., Veuthey J.L., Zamboni N., Westermann B. Identification and functional expression of the mitochondrial pyruvate carrier. Science. 2012;337:93–96. doi: 10.1126/science.1218530. [DOI] [PubMed] [Google Scholar]
- 89.Bricker D.K., Taylor E.B., Schell J.C., Orsak T., Boutron A., Chen Y.C. A mitochondrial pyruvate carrier required for pyruvate uptake in yeast, Drosophila, and humans. Science. 2012;337:96–100. doi: 10.1126/science.1218099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Pellerin L., Pellegri G., Bittar P.G., Charnay Y., Bouras C., Martin J.L. Evidence supporting the existence of an activity-dependent astrocyte-neuron lactate shuttle. Dev Neurosci. 1998;20:291–299. doi: 10.1159/000017324. [DOI] [PubMed] [Google Scholar]
- 91.Liu L., MacKenzie K.R., Putluri N., Maletic-Savatic M., Bellen H.J. The glia-neuron lactate shuttle and elevated ROS promote lipid synthesis in neurons and lipid droplet accumulation in glia via APOE/D. Cell Metabolism. 2017;27:719–737. doi: 10.1016/j.cmet.2017.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Boussouar F., Benahmed M. Lactate and energy metabolism in male germ cells. Trends Endocrinol Metab. 2004;15:345–350. doi: 10.1016/j.tem.2004.07.003. [DOI] [PubMed] [Google Scholar]
- 93.Storey B.T., Kayne F.J. Energy metabolism of spermatozoa. VI. Direct intramitochondrial lactate oxidation by rabbit sperm mitochondria. Biol Reprod. 1977;16:549–556. [PubMed] [Google Scholar]
- 94.Ahmed K., Tunaru S., Tang C., Müller M., Gille A., Sassmann A. An autocrine lactate loop mediates insulin-dependent inhibition of lipolysis through GPR81. Cell Metab. 2010;11:311–319. doi: 10.1016/j.cmet.2010.02.012. [DOI] [PubMed] [Google Scholar]
- 95.Cai T.Q., Ren N., Jin L., Cheng K., Kash S., Chen R. Role of GPR81 in lactate-mediated reduction of adipose lipolysis. Biochem Biophys Res Commun. 2008;377:987–991. doi: 10.1016/j.bbrc.2008.10.088. [DOI] [PubMed] [Google Scholar]
- 96.Liu C., Wu J., Zhu J., Kuei C., Yu J., Shelton J. Lactate inhibits lipolysis in fat cells through activation of an orphan G-protein-coupled receptor, GPR81. J Biol Chem. 2009;284:2811–2822. doi: 10.1074/jbc.M806409200. [DOI] [PubMed] [Google Scholar]
- 97.McClelland G.B., Khanna S., González G.F., Butz C.E., Brooks G.A. Peroxisomal membrane monocarboxylate transporters: evidence for a redox shuttle system? Biochem Biophys Res Commun. 2003;304:130–135. doi: 10.1016/s0006-291x(03)00550-3. [DOI] [PubMed] [Google Scholar]
- 98.Brooks G.A., Mercier J. Balance of carbohydrate and lipid utilization during exercise: the “crossover” concept. J Appl Physiol (1985) 1994;76:2253–2261. doi: 10.1152/jappl.1994.76.6.2253. [DOI] [PubMed] [Google Scholar]
- 99.Issekutz B., Jr, Miller H. Plasma free fatty acids during exercise and the effect of lactic acid. Proc Soc Exp Biol Med. 1962;110:237–239. [Google Scholar]
- 100.Rodahl K., Miller H.I., Issekutz B., Jr Plasma free fatty acids in exercise. J Appl Physiol. 1964;19:489–492. doi: 10.1152/jappl.1964.19.3.489. [DOI] [PubMed] [Google Scholar]
- 101.Gold M., Miller H.I., Issekutz B., Jr., Spitzer J.J. Effect of exercise and lactic acid infusion on individual free fatty acids of plasma. Am J Physiol. 1963;205:902–904. doi: 10.1152/ajplegacy.1963.205.5.902. [DOI] [PubMed] [Google Scholar]
- 102.Miller H.I., Issekutz B., Jr., Rodahl K., Paul P. Effect of lactic acid on plasma free fatty acids in pancreatectomized dogs. Am J Physiol. 1964;207:1226–1230. doi: 10.1152/ajplegacy.1964.207.6.1226. [DOI] [PubMed] [Google Scholar]
- 103.Ge H., Weiszmann J., Reagan J.D., Gupte J., Baribault H., Gyuris T. Elucidation of signaling and functional activities of an orphan GPCR, GPR81. J Lipid Res. 2008;49:797–803. doi: 10.1194/jlr.M700513-JLR200. [DOI] [PubMed] [Google Scholar]
- 104.Hoque R., Farooq A., Ghani A., Gorelick F., Mehal W.Z. Lactate reduces liver and pancreatic injury in Toll-like receptor- and inflammasome-mediated inflammation via GPR81-mediated suppression of innate immunity. Gastroenterology. 2014;146:1763–1774. doi: 10.1053/j.gastro.2014.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Bergersen L.H. Lactate transport and signaling in the brain: potential therapeutic targets and roles in body-brain interaction. J Cereb Blood Flow Metab. 2015;35:176–185. doi: 10.1038/jcbfm.2014.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lauritzen K.H., Morland C., Puchades M., Holm-Hansen S., Hagelin E.M., Lauritzen F. Lactate receptor sites link neurotransmission, neurovascular coupling, and brain energy metabolism. Cereb Cortex. 2014;24:2784–2795. doi: 10.1093/cercor/bht136. [DOI] [PubMed] [Google Scholar]
- 107.Takahashi H., Alves C.R.R., Stanford K.I., Middelbeek R.J.W., Nigro Pasquale, Ryan R.E. TGF-β2 is an exercise-induced adipokine that regulates glucose and fatty acid metabolism. Nat Metab. 2019;1:291–303. doi: 10.1038/s42255-018-0030-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Friedman A., Kaufer D., Heinemann U. Blood-brain barrier breakdown-inducing astrocytic transformation: novel targets for the prevention of epilepsy. Epilepsy Res. 2009;85:142–149. doi: 10.1016/j.eplepsyres.2009.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Henderson G.C., Fattor J.A., Horning M.A., Faghihnia N., Johnson M.L., Mau T.L. Lipolysis and fatty acid metabolism in men and women during the postexercise recovery period. J Physiol. 2007;584:963–981. doi: 10.1113/jphysiol.2007.137331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Wu B.U., Hwang J.Q., Gardner T.H., Repas K., Delee R., Yu S. Lactated Ringer's solution reduces systemic inflammation compared with saline in patients with acute pancreatitis. Clin Gastroenterol Hepatol. 2011;9:710. doi: 10.1016/j.cgh.2011.04.026. [DOI] [PubMed] [Google Scholar]
- 111.Carteron L., Solari D., Patet C., Quintard H., Miroz J.P., Bloch J. Hypertonic lactate to improve cerebral perfusion and glucose availability after acute brain injury. Crit Care Med. 2018;46:1649–1655. doi: 10.1097/CCM.0000000000003274. [DOI] [PubMed] [Google Scholar]
- 112.Oddo M., Vespa P., Menon D.K. Boosting the injured brain with supplemental energy fuels. Intensive Care Med. 2019;45:872–875. doi: 10.1007/s00134-018-05517-6. [DOI] [PubMed] [Google Scholar]
- 113.Patet C., Suys T., Carteron L., Oddo M. Cerebral lactate metabolism after traumatic brain injury. Curr Neurol Neurosci Rep. 2016;16:31. doi: 10.1007/s11910-016-0638-5. [DOI] [PubMed] [Google Scholar]
- 114.Warburg O., Minami S. Tests on over-living carcinoma tissue (Versuche an Überlebendem Carcinom-gewebe) Klinische Wochenschrift. 1923;2:776–777. [in German] [Google Scholar]
- 115.Racker E. Bioenergetics and the problem of tumor growth. Am Sci. 1972;60:56–63. [PubMed] [Google Scholar]
- 116.Hanahan D., Weinberg R.A. The hallmarks of cancer. Cell. 2000;100:57–70. doi: 10.1016/s0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 117.Koppenol W.H., Bounds P.L., Dang C.V. Otto Warburg's contributions to current concepts of cancer metabolism. Nat Rev Cancer. 2011;11:325–337. doi: 10.1038/nrc3038. [DOI] [PubMed] [Google Scholar]
- 118.Hensley C.T., Faubert B., Yuan Q., Lev-Cohain N., Jin E., Kim J. Metabolic heterogeneity in human lung tumors. Cell. 2016;164:681–694. doi: 10.1016/j.cell.2015.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.San-Millan I., Brooks G.A. Reexamining cancer metabolism: lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis. 2017;38:119–133. doi: 10.1093/carcin/bgw127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Goodwin M.L., Gladden L.B., Nijsten M.W., Jones K.B. Lactate and cancer: revisiting the Warburg effect in an era of lactate shuttling. Front Nutr. 2014;1:27. doi: 10.3389/fnut.2014.00027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Semenza G.L. Tumor metabolism: cancer cells give and take lactate. J Clin Invest. 2008;118:3835–3837. doi: 10.1172/JCI37373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Doherty J.R., Yang C., Scott K.E., Cameron M.D., Fallahi M., Li W. Blocking lactate export by inhibiting the Myc target MCT1 disables glycolysis and glutathione synthesis. Cancer Res. 2014;74:908–920. doi: 10.1158/0008-5472.CAN-13-2034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Draoui N., Feron O. Lactate shuttles at a glance: from physiological paradigms to anti-cancer treatments. Dis Model Mech. 2011;4:727–732. doi: 10.1242/dmm.007724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Draoui N., Schicke O., Seront E., Bouzin C., Sonveaux P., Riant O. Antitumor activity of 7-aminocarboxycoumarin derivatives, a new class of potent inhibitors of lactate influx but not efflux. Mol Cancer Ther. 2014;13:1410–1418. doi: 10.1158/1535-7163.MCT-13-0653. [DOI] [PubMed] [Google Scholar]
- 125.Doherty J.R., Cleveland J.L. Targeting lactate metabolism for cancer therapeutics. J Clin Invest. 2013;123:3685–3692. doi: 10.1172/JCI69741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Ovens M.J., Davies A.J., Wilson M.C., Murray C.M., Halestrap A.P. AR-C155858 is a potent inhibitor of monocarboxylate transporters MCT1 and MCT2 that binds to an intracellular site involving transmembrane helices 7–10. Biochem J. 2010;425:523–530. doi: 10.1042/BJ20091515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Baba M., Inoue M., Itoh K., Nishizawa Y. Blocking CD147 induces cell death in cancer cells through impairment of glycolytic energy metabolism. Biochem Biophys Res Commun. 2008;374:111–116. doi: 10.1016/j.bbrc.2008.06.122. [DOI] [PubMed] [Google Scholar]
- 128.Schneiderhan W., Scheler M., Holzmann K.H., Marx M., Gschwend J.E., Bucholz M. CD147 silencing inhibits lactate transport and reduces malignant potential of pancreatic cancer cells in in vivo and in vitro models. Gut. 2009;58:1391–1398. doi: 10.1136/gut.2009.181412. [DOI] [PubMed] [Google Scholar]
- 129.Su J., Chen X., Kanekura T. A CD147-targeting siRNA inhibits the proliferation, invasiveness, and VEGF production of human malignant melanoma cells by down-regulating glycolysis. Cancer Lett. 2009;273:140–147. doi: 10.1016/j.canlet.2008.07.034. [DOI] [PubMed] [Google Scholar]
- 130.Zou W., Yang H., Hou X., Zhang W., Chen B., Xin X. Inhibition of CD147 gene expression via RNA interference reduces tumor cell invasion, tumorigenicity and increases chemosensitivity to paclitaxel in HO-8910pm cells. Cancer Lett. 2007;248:211–218. doi: 10.1016/j.canlet.2006.07.005. [DOI] [PubMed] [Google Scholar]
- 131.Powell K.E., King A.C., Buchner D.M., Campbell W.W., DiPietro L., Erickson K.I. The scientific foundation for the physical activity guidelines for Americans, 2nd ed. J Phys Act Health. 2018;16:1–11. doi: 10.1123/jpah.2018-0618. [DOI] [PubMed] [Google Scholar]
- 132.Hultman E.A. Physiological role of muscle glycogen in man. Physiol Muscular Exerc. 1967:I99–I112. [Google Scholar]
- 133.Meyer C., Dostou J.M., Welle S.L., Gerich J.E. Role of human liver, kidney, and skeletal muscle in postprandial glucose homeostasis. Am J Physiol Endocrinol Metab. 2002;282:E419–E427. doi: 10.1152/ajpendo.00032.2001. [DOI] [PubMed] [Google Scholar]
- 134.Meyer C., Stumvoll M., Dostou J., Welle S., Haymond M., Gerich J. Renal substrate exchange and gluconeogenesis in normal postabsorptive humans. Am J Physiol Endocrinol Metab. 2002;282:E428–E434. doi: 10.1152/ajpendo.00116.2001. [DOI] [PubMed] [Google Scholar]
- 135.Garcia C.K., Goldstein J.L., Pathak R.K., Anderson R.G., Brown M.S. Molecular characterization of a membrane transporter for lactate, pyruvate, and other monocarboxylates: implications for the Cori cycle. Cell. 1994;76:865–873. doi: 10.1016/0092-8674(94)90361-1. [DOI] [PubMed] [Google Scholar]
- 136.Price N.T., Jackson V.N., Halestrap A.P. Cloning and sequencing of four new mammalian monocarboxylate transporter (MCT) homologues confirms the existence of a transporter family with an ancient past. Biochem J. 1998;329:321–328. doi: 10.1042/bj3290321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Pullen T.J., Khan A.M., Barton G., Butcher S.A., Sun G., Rutter G.A. Identification of genes selectively disallowed in the pancreatic islet. Islets. 2010;2:89–95. doi: 10.4161/isl.2.2.11025. [DOI] [PubMed] [Google Scholar]
- 138.Rutter G.A., Pullen T.J., Hodson D.J., Martinez-Sanchez A. Pancreatic β-cell identity, glucose sensing and the control of insulin secretion. Biochem J. 2015;466:203–218. doi: 10.1042/BJ20141384. [DOI] [PubMed] [Google Scholar]
- 139.Bender K., Newsholme P., Brennan L., Maechler P. The importance of redox shuttles to pancreatic beta-cell energy metabolism and function. Biochem Soc Trans. 2006;34:811–814. doi: 10.1042/BST0340811. [DOI] [PubMed] [Google Scholar]
- 140.Otonkoski T., Jiao H., Kaminen-Ahola N., Tapia-Paez I., Ullah M.S., Parton L.E. Physical exercise-induced hypoglycemia caused by failed silencing of monocarboxylate transporter 1 in pancreatic β cells. Am J Hum Genet. 2007;81:467–474. doi: 10.1086/520960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Casaburi R., Barstow T.J., Robinson T., Wasserman K. Influence of work rate on ventilatory and gas exchange kinetics. J Appl Physiol (1985) 1989;67:547–555. doi: 10.1152/jappl.1989.67.2.547. [DOI] [PubMed] [Google Scholar]
- 142.Messonnier A.L., Emhoff C.A., Fattor J.A., Horning M.A., J. T., Brooks G.A. Lactate kinetics at the lactate threshold in trained and untrained men. J Appl Physiol (1985) 2013;114:1593–1602. doi: 10.1152/japplphysiol.00043.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ferguson B.S., Rogatzki M.J., Goodwin M.L., Kane D.A., Rightmire Z., Gladden L.B. Lactate metabolism: historical context, prior misinterpretations, and current understanding. Eur J Appl Physiol. 2018;118:691–728. doi: 10.1007/s00421-017-3795-6. [DOI] [PubMed] [Google Scholar]
- 144.Richardson R.S., Noyszewski E.A., Leigh J.S., Wagner P.D. Lactate efflux from exercising human skeletal muscle: role of intracellular PO2. J Appl Physiol (1985) 1998;85:627–634. doi: 10.1152/jappl.1998.85.2.627. [DOI] [PubMed] [Google Scholar]
- 145.Mazzeo R.S., Brooks G.A., Butterfield G.E., Podolin D.A., Wolfel E.E., Reeves J.T. Acclimatization to high altitude increase muscle sympathetic activity both at rest and during exercise. Am J Physiol. 1995;269:R201–R207. doi: 10.1152/ajpregu.1995.269.1.R201. [DOI] [PubMed] [Google Scholar]
- 146.Mercier J.G., Hokanson J.F., Brooks G.A. Effects of cyclosporine A on skeletal muscle mitochondrial respiration and endurance time in rats. Am J Respir Crit Care Med. 1995;151:1532–1536. doi: 10.1164/ajrccm.151.5.7735611. [DOI] [PubMed] [Google Scholar]
- 147.Hughes E.F., Turner S.C., Brooks G.A. Effects of glycogen depletion and pedaling speed on “anaerobic threshold”. J Appl Physiol Respir Environ Exerc Physiol. 1982;52:1598–1607. doi: 10.1152/jappl.1982.52.6.1598. [DOI] [PubMed] [Google Scholar]
- 148.Brooks G.A., Butterfield G.E., Wolfe R.R., Groves B.M., Mazzeo R.S., Sutton J.R. Decreased reliance on lactate during exercise after acclimatization to 4,300 m. J Appl Physiol (1985) 1991;71:333–341. doi: 10.1152/jappl.1991.71.1.333. [DOI] [PubMed] [Google Scholar]
- 149.Gallagher C.N., Carpenter K.L., Grice P., Howe D.J., Mason A., Timofeev I. The human brain utilizes lactate via the tricarboxylic acid cycle: a 13C-labelled microdialysis and high-resolution nuclear magnetic resonance study. Brain. 2009;132:2839–2849. doi: 10.1093/brain/awp202. [DOI] [PubMed] [Google Scholar]
- 150.van Hall G., Str ø mstad M., Rasmussen P., Jans O., Zaar M., Gam C. Blood lactate is an important energy source for the human brain. J Cereb Blood Flow Metab. 2009;29:1121–1129. doi: 10.1038/jcbfm.2009.35. [DOI] [PubMed] [Google Scholar]
- 151.Smith D., Pernet A., Hallett W.A., Bingham E., Marsden P.K., Amiel S.A. Lactate: a preferred fuel for human brain metabolism in vivo. J Cereb Blood Flow Metab. 2003;23:658–664. doi: 10.1097/01.WCB.0000063991.19746.11. [DOI] [PubMed] [Google Scholar]
- 152.Quistorff B., Secher N.H., Van Lieshout J.J. Lactate fuels the human brain during exercise. FASEB J. 2008;22:3443–3449. doi: 10.1096/fj.08-106104. [DOI] [PubMed] [Google Scholar]
- 153.Schurr A. Lactate: the ultimate cerebral oxidative energy substrate? J Cereb Blood Flow Metab. 2006;26:142–152. doi: 10.1038/sj.jcbfm.9600174. [DOI] [PubMed] [Google Scholar]
- 154.Schurr A., Payne R.S. Lactate, not pyruvate, is neuronal aerobic glycolysis end product: an in vitro electrophysiological study. Neuroscience. 2007;147:613–619. doi: 10.1016/j.neuroscience.2007.05.002. [DOI] [PubMed] [Google Scholar]
- 155.Coco M., Caggia S., Musumeci G., Perciavalle V., Graziano A.C., Pannuzzo G. Sodium L-lactate differently affects brain-derived neurothrophic factor, inducible nitric oxide synthase, and heat shock protein 70 kDa production in human astrocytes and SH-SY5Y cultures. J Neurosci Res. 2013;91:313–320. doi: 10.1002/jnr.23154. [DOI] [PubMed] [Google Scholar]
- 156.Seifert T., Brassard P., Wissenberg M., Rasmussen P., Nordby P., Stallknecht B. Endurance training enhances BDNF release from the human brain. Am J Physiol Regul Integr Comp Physiol. 2010;298:R372–R377. doi: 10.1152/ajpregu.00525.2009. [DOI] [PubMed] [Google Scholar]
- 157.Cotman C.W., Berchtold N.C. Exercise: a behavioral intervention to enhance brain health and plasticity. Trends Neurosci. 2002;25:295–301. doi: 10.1016/s0166-2236(02)02143-4. [DOI] [PubMed] [Google Scholar]
- 158.Tsai S.J. Brain-derived neurotrophic factor: a bridge between major depression and Alzheimer's disease? Med Hypotheses. 2003;61:110–113. doi: 10.1016/s0306-9877(03)00141-5. [DOI] [PubMed] [Google Scholar]
- 159.Krabbe K.S., Nielsen A.R., Krogh-Madsen R., Plomgaard P., Rasmussen P., Erikstrup C. Brain-derived neurotrophic factor (BDNF) and type 2 diabetes. Diabetologia. 2007;50:431–438. doi: 10.1007/s00125-006-0537-4. [DOI] [PubMed] [Google Scholar]
- 160.Holloway R., Zhou Z., Harvey H.B., Levasseur J.E., Rice A.C., Sun D. Effect of lactate therapy upon cognitive deficits after traumatic brain injury in the rat. Acta Neurochirurgica (Wien) 2007;149:919–927. doi: 10.1007/s00701-007-1241-y. discussion 927. [DOI] [PubMed] [Google Scholar]
- 161.Rice A.C., Zsoldos R., Chen T., Wilson M.S., Alessandri B., Hamm R.J. Lactate administration attenuates cognitive deficits following traumatic brain injury. Brain Res. 2002;928:156–159. doi: 10.1016/s0006-8993(01)03299-1. [DOI] [PubMed] [Google Scholar]
- 162.Bisri T., Utomo B.A., Fuadi I. Exogenous lactate infusion improved neurocognitive function of patients with mild traumatic brain injury. Asian J Neurosurg. 2016;11:151–159. doi: 10.4103/1793-5482.145375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Hashimoto T., Tsukamoto H., Takenaka S., Olesen N.D., Petersen L.G., Sørensen H. Exercise improves brain executive function related to cerebral lactate metabolism in men. FASEB J. 2018;32:1417–1427. doi: 10.1096/fj.201700381RR. [DOI] [PubMed] [Google Scholar]
- 164.Wang J., Tu J., Cao B., Mu L., Yang X., Cong M. Astrocytic l-lactate signaling facilitates amygdala-anterior cingulate cortex synchrony and decision making in rats. Cell Rep. 2017;21:2407–2418. doi: 10.1016/j.celrep.2017.11.012. [DOI] [PubMed] [Google Scholar]
- 165.Schurr A., Gozal E. Aerobic production and utilization of lactate satisfy increased energy demands upon neuronal activation in hippocampal slices and provide neuroprotection against oxidative stress. Front Pharmacol. 2012;2:96. doi: 10.3389/fphar.2011.00096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Suzuki A., Stern S.A., Bozdagi O., Huntley G.W., Walker R.H., Magistretti P.J. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell. 2011;144:810–823. doi: 10.1016/j.cell.2011.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Steinman M.Q., Gao V., Alberini C.M. The role of lactate-mediated metabolic coupling between astrocytes and neurons in long-term memory formation. Front Integr Neurosci. 2016;10:10. doi: 10.3389/fnint.2016.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Shapiro N.I., Howell M.D., Talmor D., Nathanson L.A., Lisbon A., Wolfe R.E. Serum lactate as a predictor of mortality in emergency department patients with infection. Ann Emerg Med. 2005;45:524–528. doi: 10.1016/j.annemergmed.2004.12.006. [DOI] [PubMed] [Google Scholar]
- 169.Zhang Z., Xu X. Lactate clearance is a useful biomarker for the prediction of all-cause mortality in critically ill patients: a systematic review and meta-analysis*. Crit Care Med. 2014;42:2118–2125. doi: 10.1097/CCM.0000000000000405. [DOI] [PubMed] [Google Scholar]
- 170.Messonnier L.A., Emhoff C.A., Fattor J.A., Horning M.A., Carlson T.J., Brooks G.A. Lactate kinetics at the lactate threshold in trained and untrained men. J Appl Physiol (1985) 2013;114:1593–1602. doi: 10.1152/japplphysiol.00043.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Rivers E., Nguyen B., Havstad S., Ressler J., Muzzin A., Knoblich B. Early goal-directed therapy in the treatment of severe sepsis and septic shock. N Engl J Med. 2001;345:1368–1377. doi: 10.1056/NEJMoa010307. [DOI] [PubMed] [Google Scholar]
- 172.Miller B.F., Fattor J.A., Jacobs K.A., Horning M.A., Suh S.H., Navazio F. Metabolic and cardiorespiratory responses to “the lactate clamp”. Am J Physiol Endocrinol Metab. 2002;283:E889–E898. doi: 10.1152/ajpendo.00266.2002. [DOI] [PubMed] [Google Scholar]
- 173.Marik P., Bellomo R. Lactate clearance as a target of therapy in sepsis: a flawed paradigm. OA Critical Care. 2013;1:3–5. [Google Scholar]
- 174.Marik P., Bellomo R. A rational approach to fluid therapy in sepsis. Br J Anaesth. 2016;116:339–349. doi: 10.1093/bja/aev349. [DOI] [PubMed] [Google Scholar]
- 175.Nalos M., Leverve X., Huang S., Weisbrodt L., Parkin R., Seppelt I. Half-molar sodium lactate infusion improves cardiac performance in acute heart failure: a pilot randomised controlled clinical trial. Crit Care. 2014;18:R48. doi: 10.1186/cc13793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Sharma A.B., Barlow M.A., Yang S.H., Simpkins J.W., Mallet R.T. Pyruvate enhances neurological recovery following cardiopulmonary arrest and resuscitation. Resuscitation. 2008;76:108–119. doi: 10.1016/j.resuscitation.2007.04.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Sharma A.B., Knott E.M., Bi J., Martinez R.R., Sun J., Mallet R.T. Pyruvate improves cardiac electromechanical and metabolic recovery from cardiopulmonary arrest and resuscitation. Resuscitation. 2005;66:71–81. doi: 10.1016/j.resuscitation.2004.12.016. [DOI] [PubMed] [Google Scholar]
- 178.Oddo M., Levine J.M., Frangos S., Maloney-Wilensky E., Carrera E., Daniel R.T. Brain lactate metabolism in humans with subarachnoid hemorrhage. Stroke. 2012;43:1418–1421. doi: 10.1161/STROKEAHA.111.648568. [DOI] [PubMed] [Google Scholar]
- 179.Wolahan S.M., Mao H.C., Real C., Vespa P.M., Glenn T.C. Lactate supplementation in severe traumatic brain injured adults by primed constant infusion of sodium L-lactate. J Neurosci Res. 2018;96:688–695. doi: 10.1002/jnr.24085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Azevedo J.L., Tietz E., Two-Feathers T., Paull J., Chapman K. Lactate, fructose and glucose oxidation profiles in sports drinks and the effect on exercise performance. PloS One. 2007;2:e927. doi: 10.1371/journal.pone.0000927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Hofmann P., Pokan R. Value of the application of the heart rate performance curve in sports. Int J Sports Physiol Perform. 2010;5:437–447. doi: 10.1123/ijspp.5.4.437. [DOI] [PubMed] [Google Scholar]
- 182.San-Millán I., Brooks G.A. Assessment of metabolic flexibility by means of measuring blood lactate, fat, and carbohydrate oxidation responses to exercise in professional endurance athletes and less-fit individuals. Sports Med. 2018;48:467–479. doi: 10.1007/s40279-017-0751-x. [DOI] [PubMed] [Google Scholar]
- 183.Horning MA, Brooks GA, Inventors. Formulation and methods to provide nutrition to human and other patients. US patent 20140120238. May 1, 2014.
- 184.Horning MA, Brooks GA, Inventors. Formulations containing labels for medical diagnostics. US patent 09500657. Nov. 22, 2016.
- 185.Horning MA, Brooks GA, Inventors. Systems and methods to estimate nutritional needs of human and other patients. US patent 8927490. Jan. 6, 2015.
- 186.Mitchell J.H., Sproule B.J., Chapman C.B. The physiological meaning of the maximal oxygen intake test. J Clin Invest. 1958;37:538–547. doi: 10.1172/JCI103636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Holloszy J.O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J Biol Chem. 1967;242:2278–2282. [PubMed] [Google Scholar]
- 188.Gladden L.B. Lactate metabolism: a new paradigm for the third millennium. J Physiol. 2004;558:5–30. doi: 10.1113/jphysiol.2003.058701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Smuder A.J., Morton A.B., Hall S.E., Wiggs M.P., Ahn B., Wawrzyniak N.R. Effects of exercise preconditioning and HSP72 on diaphragm muscle function during mechanical ventilation. J Cachexia Sarcopenia Muscle. 2019;10:767–781. doi: 10.1002/jcsm.12427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Dempsey J.A., Rankin J. Physiologic adaptations of gas transport systems to muscular work in health and disease. Am J Phys Med. 1967;46:582–647. [PubMed] [Google Scholar]
- 191.Fahey T.D., Del Valle-Zuris A., Oehlsen G., Trieb M., Seymour J. Pubertal stage differences in hormonal and hematological responses to maximal exercise in males. J Appl Physiol Respir Environ Exerc Physiol. 1979;46:823–827. doi: 10.1152/jappl.1979.46.4.823. [DOI] [PubMed] [Google Scholar]
- 192.Rivera M.A., Fahey T.D. Association between aquaporin-1 and endurance performance: a systematic review. Sports Med Open. 2019;5:40. doi: 10.1186/s40798-019-0213-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Fahey T.D., Insel P.M., Roth W.T. 13th ed. McGraw-Hill Higher Education; Mountain View, CA: 2019. Fit and well: core concepts and labs in physical fitness and wellness. [Google Scholar]
- 194.Brooks G.A., Fahey T.D., Baldwin K.M. 5th ed. 1. Kindle Direct Publishing; Lexington, KY: 2019. (Exercise physiology: human bioenergetics and its applications). [Google Scholar]
- 195.Rumsey W.L., Schlosser C., Nuutinen E.M., Robiolio M., Wilson D.F. Cellular energetics and the oxygen dependence of respiration in cardiac myocytes isolated from adult rat. J Biol Chem. 1990;265:15392–15402. [PubMed] [Google Scholar]
- 196.Bendahan D., Chatel B., Jue T. Comparative NMR and NIRS analysis of oxygen dependent metabolism in exercising finger flexor muscles. Am J Physiol Regul Integr Comp Physiol. 2017;313:R740–R753. doi: 10.1152/ajpregu.00203.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Connett R.J., Honig C.R., Gayeski T.E., Brooks G.A. Defining hypoxia: a systems view of VO2, glycolysis, energetics, and intracellular PO2. J Appl Physiol (1985) 1990;68:833–842. doi: 10.1152/jappl.1990.68.3.833. [DOI] [PubMed] [Google Scholar]
- 198.Brooks G.A., Fahey T.D., Baldwin K.M. 5th ed. 2. Kindle Direct Publishing; San Bernardino, CA: 2019. (Exercise physiology: human bioenergetics and its applications). [Google Scholar]
- 199.Saddik M., Gamble J., Witters L.A., Lopaschuk G.D. Acetyl-CoA carboxylase regulation of fatty acid oxidation in the heart. J Biol Chem. 1993;268:25836–25845. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.