

Abstract
Rheumatoid arthritis (RA) is one of the most common autoimmune diseases caused by abnormal immune activation and immune tolerance. Immunomodulatory cells (ICs) play a critical role in the maintenance and homeostasis of normal immune function and in the pathogenesis of RA. The human gastrointestinal tract is inhabited by trillions of commensal microbiota on the mucosal surface that play a fundamental role in the induction, maintenance, and function of the host immune system. Gut microbiota dysbiosis can impact both the local and systemic immune systems and further contribute to various diseases, such as RA. The neighbouring intestinal ICs located in distinct intestinal mucosa may be the most likely intermediary by which the gut microbiota can affect the occurrence and development of RA. However, the reciprocal interaction between the components of the gut microbiota and their microbial metabolites with distinct ICs and how this interaction may impact the development of RA are not well studied. Therefore, a better understanding of the gut microbiota, ICs, and their interactions might improve our knowledge of the mechanisms by which the gut microbiota contribute to RA and facilitate the further development of novel therapeutic approaches. In this review, we have summarized the roles of the gut microbiota in the immunopathogenesis of RA, especially the interactions between the gut microbiota and ICs, and further discussed the strategies for treating RA by targeting/regulating the gut microbiota.
1. Introduction to RA
Rheumatoid arthritis (RA) is a chronic autoimmune disease that affects approximately 1% of the population worldwide, caused by abnormal immune activation and immune tolerance and characterized by synovial inflammation, cartilage damage, and bone destruction [1]. Previous studies have suggested that immune abnormalities, such as immunomodulatory cell (IC) activation or inhibition, that take place at a local and, subsequently, at a systemic level are present in patients who are at risk of developing RA [2]. The frequency and function of T and B lymphocyte subsets are associated with the pathogenesis of RA [3, 4]. The pathologically increased production of CXCL13 and interleukin- (IL-) 21 by the peripheral T helper (TPH) cell phenotype can recruit T follicular (Tfh) helper cells and B cells, and the TPH cell phenotype can expand the spectrum of B cell-helper T cells within the inflamed synovium of RA patients [5]. T helper 17 (Th17) cells, which can produce IL-17, contribute to the onset of RA [6], while regulatory T (Treg) cells, which secrete the anti-inflammatory cytokine IL-10 and transforming growth factor- (TGF-) β1, are pivotal players in the control of RA [7]. Many approaches for the treatment of RA may promote Treg generation/expansion while inhibiting Th17 cell differentiation and consequently restoring the balance of the Th17/Treg cell ratio [8–11]. Some drugs can even decrease the frequency of Th1 cells but increase that of Th2 cells [12]. Autoreactive B cells are also crucial in the aetiology of RA and producing anticitrullinated protein antibodies (ACPAs) and rheumatoid factors (RFs) [3]. In the lymphoid tissue of RA patients, increased T cell activation is related to the persistence of switched memory B cells [13]. A memory B cell subset that expresses the immunoglobulin A (IgA) receptor, termed Fc receptor-like 4 (FcRL4), is a component of the local autoimmune response that can contribute to the joint destruction in RA patients via receptor activation of nuclear factor-κB ligand (RANKL) expression [14]. Numerous immune-related abnormalities in RA notably occur at the mucosal level. During the development of RA, the intestinal mucosal immune response is excessively exaggerated, antigen-presenting cells (APCs) are abnormally activated, and immune tolerance is disturbed. The intestinal mucosal immune system, especially Peyer's patches, is important for inducing immunity tolerance. The higher numbers of CD8+ cells and CD4+ cells are observed in Peyer's patches of rats with collagen-induced arthritis (CIA) than those of control rats [15]. The CIA animal model shares many clinical, histopathological, and immunological features with clinical human RA, and these similarities are commonly exploited in the use of the CIA model for studying RA [16]. One study showed that in rats with CIA, the concentrations of secretory IgA (sIgA) in the small intestine and interferon-γ (IFN-γ) in small intestinal tissue homogenates were upregulated, the ratios of CD4+/CD8+ in both the epithelium and lamina propria of the small intestine were increased, and the expression of CD80, CD86, IL-6, and IL-12 mRNA in the small intestine was also significantly increased compared with those in the control group [17]. Evans-Marin et al. showed that CD4+ T cells in the lamina propria were activated before the onset of arthritis in mice with CIA, following the significant upregulation of IL-17A, tumor necrosis factor- (TNF-) α, and granulocyte-macrophage colony-stimulating factor (GM-CSF), and the severity of arthritis was significantly reduced if Th17 cells were lacking [18]. Serum analysis of recent-onset RA patients who received antirheumatic therapy showed that the ACPA, sIgA, and IgM levels rapidly decreased, which were related to the decreased disease activity. The decreased mucosal immunity to citrullinated proteins/peptides and the recruitment of new B cells are crucial features of antirheumatic therapy responses in early RA patients [19].
Since genetic factors cannot completely account for the risk of this disease [20], the substantive role of other risk factors needs to be explored. Recently, it has been proposed that environmental factors are potentially involved in RA progression [21]. Additional evidence has shown that the gut microbiota might be an important experimental factor in the occurrence and development of RA [22]. Diverse ICs, especially those located in the intestinal mucosa, are the most likely intermediary by which the gut microbiota can impact the occurrence and development of RA. As a result, there is a possible interaction among the gut microbiota, ICs, and initiation and propagation of RA, which has recently attracted more interest. In this review, we summarize the roles of the gut microbiota in the pathogenesis and treatment of RA, focusing on the interactions between the gut microbiota and intestinal ICs.
2. General Introduction to the Gut Microbiota
The gastrointestinal tract of mammals is abundantly colonized by trillions of different prokaryotic microorganisms, including bacteria, archaea, fungi, and viruses, collectively termed the gut microbiota, which have a mutual relationship with their host. The gut microbiota contains 1000-5000 different species of microorganisms that numerically exceed host cells by approximately 10-fold, with 99% belonging to the phyla Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria [23–25]. The microbiota composition of the host in adulthood is shaped during the early neonatal period [26]. The microbiota consists of “beneficial” symbiotic microbes that maintain host homeostasis in a cooperative and nonpathogenic manner, “sensitive” microbes that become dysregulated due to diseases, “pathogenic” microbes that can cause disease, and “therapeutic” microbes that can help reverse alterations [27]. The beneficial microbes not only can help to digest and absorb food but also have a protective function that can prevent the adherence of pathogenic microbes to the mucosal layer [28]. Furthermore, the host can provide a niche for the survival of the beneficial microbes. The gut microbiota can be influenced by diet, probiotics, prebiotics, antibiotics, exogenous enzymes, faecal microbiota transplantation (FMT), and other environmental factors [29]. The distribution of the gut microbiota is also affected according to the local regional environment in the gastrointestinal tract [30, 31]. The gut microbiota is necessary for host health, especially for immune homeostasis and function. The components of the gut microbiota play a profound role in modulating the innate and adaptive immunity of the host. The host immune system has the ability to induce immune tolerance towards the existence of beneficial microbes and to prevent the uncontrolled growth of opportunistic pathogens [32]. Commensal microorganisms are required for the development and differentiation of the local and systemic immune systems and nonimmune components [33, 34]. The intestinal barrier plays a critical role in maintaining immune homeostasis via these microorganisms. The gut microbiota participates in immune responses presumably by altering intestinal barrier permeability, modifying self-antigen integrity, mimicking epitopes, and modulating cell apoptosis mechanisms [35–37]. The tolerance to the gut microbiota must be maintained in order for the host to benefit from their coexistence; on the contrary, colonization with specific pathogenic microbes may be detrimental to the host, leading to diseases [33]. The gut microbiota has been found to play important roles in the pathogenesis of many intestinal and extraintestinal diseases [38, 39]. In addition, gut microbiota dysbiosis, which closely interacts with the intestinal mucosal immune system, has been associated with autoimmune diseases such as RA; moreover, the diversity, various taxa from the phylum to genus levels, and associated functions of the gut microbiota are already altered in patients in the early stages of RA [40]. However, the underlying cellular and molecular mechanisms are undefined.
3. Interactions of the Gut Microbiota and Metabolites with ICs
The immune system has coevolved with the microbial community inhabiting not only the body surface but also the mucosal barriers [41]. Mucosal sites, such as the oral and intestinal mucosa, may be the sites where autoimmunity is initiated. Although the gut microbiota is spatially confined in the intestinal lumen, these microbes and their related metabolites can shape the function of the host immune system. Some microbial clusters may be harmful and others may be helpful in maintaining the balance of the immune network. The normal immune system is maintained by a balance of protective immunity against pathogens and immune tolerance to self-antigens, and the disruption of this balance may cause the abnormal activation and proliferation of lymphocytes. Recent evidence has highlighted the immunomodulatory effect of the gut microbiota and their metabolites on ICs [42, 43].
3.1. Gut Microbiota and ICs
The interaction between the host and gut microbes is partially mediated by intestinal ICs, which sustain a complex balance. Gut microbiota dysbiosis leads to changes in the functionality of intestinal IC subsets [36, 44]. The interplay between the gut microbiota, intestinal epithelium, and innate and adaptive immune cells at homeostasis prevents immune-mediated disease [45, 46]. Gut microbiota may affect intestinal immunity by regulating T cell-mediated mucosal immunity. One of the most prominent functions of the gut microbiota and their metabolites is the regulation of the balance between proinflammatory Th1 and Th17 cells and protective Treg cells at mucosal surfaces and systemically. A recent study demonstrated that Lactobacillus helveticus SBT2171 (LH2171) directly inhibited the proliferation of lipopolysaccharide- (LPS-) stimulated mouse T and B cells and the cell cycle progression of human lymphoma cell lines (BJAB) in vitro by suppressing the c-Jun N-terminal kinase (JNK) signalling pathway [47]. Segmented filamentous bacteria (SFB) can induce Th17 cell activation [48]. The modulation of cytokine production by APCs may be critical for the anti-inflammatory effects of the gut microbiota. A previous study suggested that L. helveticus SBT2171 (LH2171) could induce the expression of A20 (a negative regulator of NF-κB/MAPK signalling) by Toll-like receptor 2 (TLR2) signalling, thus inhibiting IL-6 and IL-1β production by APCs [49]. When autoimmune-prone Dark Agouti (DA) rats were intraperitoneally injected with their own colonic E. coli or Enterococcus for two days, the proportion of resident and anti-inflammatory macrophages was diminished, the proportion of activated neutrophils was increased, and the inflammatory polarization of peritoneal cells was induced [50]. Tanoue et al. reported that a consortium of 11 bacterial strains isolated from healthy human donor faeces could induce IFN-γ-producing CD8+ T cells without causing inflammation in the intestine [51].
Various kinds of ICs and their secreted cytokines are necessary to maintain immunological homeostasis and oral tolerance to dietary antigens in the gastrointestinal tract mucosa [52]. The functional actions of ICs require the regulation of gut microbiota, thus establishing a detrimental vicious cycle. The ICs in specific compartments are also regulated in a regional specialization-dependent manner by microorganisms [53]. Colonic epithelial cells consist of different cellular subtypes, including colonocytes, gradients of progenitor cells, and goblet cells within intestinal crypts. The colonic epithelium facilitates interactions between the host and microbiota to form a mucus barrier, control mucosal immunity, and coordinate nutrient recycling through the specific contribution of each epithelial cell subtype [54]. In the small intestine, Zhou et al. showed that group-3 innate lymphoid cells (ILC3s) were the dominant cellular source of IL-2, which was required to maintain Treg; moreover, IL-2 was selectively induced by the IL-1β secreted in the small intestine, and activating IL-1β production by macrophages required MYD88- and NOD2-dependent sensing of the microbiota [55]. As major innate and adaptive lymphocyte populations, subsets of innate lymphoid cells (ILCs) and conventional T cells sequentially shape the mature commensal gut microbiota and help maintain tissue metabolic homeostasis [56].
3.2. Microbial Metabolites and ICs
The gut microbiota produces dozens of metabolites, which can participate in various physiological processes, including the modulation of immune cell function in the host gut mucosa [57]. As one of the active microbial metabolites, short-chain fatty acids (SCFAs) can induce metabolic alterations in T cells by activating the mTOR complex and modulating glucose metabolism [43]. Butyrate, a functional SCFA produced by the anaerobic gut microbiota, has been shown to prevent CIA in mice; butyrate might mediate the differentiation of CD4+ T cells towards Treg cells in the spleen, increase systemic Treg cells and decrease systemic Th17 cells, and enhance the polarization of Treg cells but not that of Th17 cells [58]. Shen et al. showed that antibiotic treatment could reduce the abundance of the microbiota in the colon, which led to decreased proportions of Treg cells and SCFAs in IL-10-deficient mice with colitis [59]. The bacterial metabolites pyruvic acid and lactic acid, which are produced in a bacteria-dependent manner, can lead to enhanced immune responses by GPR31-mediated induction of dendrite protrusions in small intestinal mononuclear cells that express CX3CR1 (CX3CR1+ cells) [60]. The aryl hydrocarbon receptor (AhR), which is a ligand-dependent transcription factor, can recognize not only tryptophan metabolites but also endogenous microbiota-derived factors and dietary components [61]. AhR can mediate crosstalk between ILCs and other immune cells in host tissues, especially in the intestinal mucosal surface [62], which may be the bridge between the gut microbiota and ICs. Li et al. demonstrated that human umbilical mesenchymal stem cells played a therapeutic role in rats with CIA by modulating the interactions between the immune status of Treg, Th17 cells, and B cells and the gut microbiota in the ileum via AhR [63].
4. Gut Microbiota and Metabolites in RA
Because of the interactions between the gut microbiota and host homeostasis, the former is believed to trigger RA through the regulation of ICs, which are both near to and distant from the site where they occur to induce. Recent advances have shown that the complex interaction between the genetic and environmental factors can contribute to the aetiology of RA [64]. However, the mechanisms underlying the outcomes still need to be elucidated. Lu et al. reported that mice deficient in TYRO3/AXL/MER (TAM) receptors spontaneously developed clinical characteristics of RA at the age of 6 months [65], but a subsequent study showed that TAM triple knockout (TKO) mice did not spontaneously develop any macroscopic arthritis-like symptoms until 52 weeks of age [66]; this difference might be due to an interplay between the genetic and environmental factors. A previous study found that the human leukocyte antigen (HLA) alleles HLA-B27 and HLA-DRB1 could favour a more inflammatory gut microbiome and/or aberrant immunologic responses to bacteria, which drive the immunopathogenesis of RA [67]. The gut microbiota and microbial metabolites strongly interact with many RA-related genes and affect RA-associated immune pathways and immunological phenotypes [68]. Observations in patient studies and in animal model experiments about RA have revealed that alterations in the host gut microbiota can influence susceptibility to onset and subsequent progression of RA (Table 1).
Table 1.
The alteration of the gut microbiota in RA patients and animal models.
| Study objects | Sample type | Technology | Increased | Decreased | References |
|---|---|---|---|---|---|
| Human, compared RA patients with nonarthritic controls | Stool | Metagenomic shotgun sequencing | Clostridiaceae | [80] | |
| Human, compared RA patients with osteoarthritis patients | Stool | 16S ribosome (r)RNA sequencing | Lactobacilli, Prevotella | Bacteroides, Bifidobacterium, Bacteroidetes/Firmicutes | [81] |
| Human, compared RA patients with healthy controls | Stool | 16S rRNA sequencing | Bacteroides, Escherichia-Shigella | Lactobacillus, Alloprevotella, Enterobacter, Odoribacter | [78] |
| Human, compared RA patients with healthy controls | Stool | Whole-genome shotgun sequencing | Prevotella | [73] | |
| Human, compared RA patients with healthy controls | Stool | 16S rRNA sequencing | Verrucomicrobiae, Akkermansia | [75] | |
| Human, compared preclinical RA patients with first-degree relatives (FDR) of RA patients | Stool | 16S rRNA sequencing | Prevotellaceae, Prevotella spp. | [70] | |
| Human, compared FDR of RA patients with healthy controls | Stool | 16S rRNA sequencing | Collinsella | Actinobacteria | [69] |
| Human, compared RA patients with healthy controls | Stool | Metagenomic shotgun sequencing | Lactobacillus salivarius | Haemophilus spp. | [77] |
| Human, compared RA patients with healthy controls | Stool | qPCR | Bacteroides, Prevotella | Clostridium leptum | [97] |
| Mouse, compared mice with CIA at the initial peak and relapse of arthritis with healthy controls | Stool | 16S rRNA gene sequencing | Bacteroides, Bacteroidales | Firmicutes | [86] |
| Mouse, compared mice with CIA with healthy controls | Stool | 16S rRNA gene sequencing | Clostridiales, Deferribacterales, Mucispirillum | Enterobacteriales | [94] |
RA: rheumatoid arthritis; CIA: collagen-induced arthritis.
4.1. Gut Microbiota Dysbiosis Triggers Inflammatory Arthritis in Human RA Patients
A certain number of bacterial species and metabolite profiles may characterize some type of RA and predict RA progression. Chen et al. showed that the abundance of Collinsella segregated with RA was correlated with high levels of α-aminoadipic acid and asparagine and with the secretion of IL-17A. The Collinsella increased gut permeability by reducing the expression of tight junction proteins in the human epithelial cell line CACO-2 in vitro, which suggested that the expansion of Collinsella enhances proinflammatory conditions through a loss of gut epithelial integrity [69]. A previous study showed an enrichment of the bacterial family Prevotellaceae, particularly Prevotella spp., in the “preclinical RA” group compared with that in the first-degree relative (FDR) (asymptomatic patients without autoantibodies) control group [70]. The abundance of Prevotella in some early RA patients is higher compared with that in the control group [71]. Pianta et al. revealed that subgroups of RA patients had differential IgG or IgA immune reactivity against P. copri, which was associated with Th17 cytokine responses and frequent ACPAs, so they suggested that P. copri is immunologically relevant to RA pathogenesis [72]. The presence of multiple Prevotella spp. in the gut microbiota, in addition to P. copri, was associated with RA aetiology by metagenome-wide shotgun sequencing [73]. Studies showed that compared to that of healthy controls, the faecal microbiota of RA patients contained more Lactobacillus communities, according to the higher richness, Shannon-Wiener, and evenness measures results [74], and had more members of the phylum Verrucomicrobiae and the genus Akkermansia. Interestingly, a higher abundance of Enterobacteriaceae and Klebsiella and a lower abundance of Bifidobacterium were detected in RA patients who had high serum levels of TNF-α or IL-17A [75]. In RA patients, the phylum Euryarchaeota was directly associated with the disease activity score on 28 joints (DAS-28) and emerged as an independent risk factor, and patients treated with etanercept (ETN) presented a partial restoration of the gut microbiota (Cyanobacteria, the class Nostocophycideae, and the order Nostocales increased, while the class Deltaproteobacteria and the family Clostridiaceae decreased) [76]. Haemophilus spp. were decreased in RA patients and negatively correlated with the levels of serum autoantibodies [77]. A recent study found that there was an increase in Bacteroides and Escherichia-Shigella and a decrease in Lactobacillus, Alloprevotella, Enterobacter, and Odoribacter in RA patients. Furthermore, using the Spearman correlation analysis, Dorea and Ruminococcus were positively correlated with RF-IgA and anti-CCP antibodies, and Alloprevotella was positively correlated with numerous rheumatoid factors, such as RF-IgM, RF-IgA, and RF-IgG, and with inflammatory biomarkers, including the erythrocyte sedimentation rate and C-reactive protein [78]. By assessing the freshly collected faecal samples from RA patients, Ebrahimi et al. reported that the serum levels of RF, ESR, CRP, anticyclic citrullinated peptide (anti-CCP), and antimutated citrullinated vimentin (anti-MCV) were significantly upregulated in Helicobacter pylori- (H. pylori-) positive patients compared with those in H. pylori-negative patients [79].
The comparison of patients with RA and comorbidities mediated by the gut microbiota has also been studied. A potential microbial link for inflammatory arthritis may exist in patients with RA and inflammatory bowel disease- (IBD-) associated arthropathy because these patients share a higher abundance of Clostridiaceae than nonarthritic controls [80]. The gut microbiota is also different between RA and osteoarthritis (OA) patients. Lee et al. demonstrated that RA patients had a lower relative abundance of Bacteroides and Bifidobacterium and a lower Bacteroidetes : Firmicutes ratio than OA patients and that the abundance of certain bacterial species, including Fusicatenibacter saccharivorans, Dialister invisus, Clostridium leptum, Ruthenibacterium lactatiformans, Anaerotruncus colihominis, Bacteroides faecichinchillae, Harryflintia acetispora, Bacteroides acidifaciens, and Christensenella minuta [81], was significantly lower in RA patients.
4.2. Gut Microbiota Dysbiosis Triggers Inflammatory Arthritis in CIA
The CIA animal model is a common experimental animal model of RA that is established by immunization of animals with type II collagen, and this model has clinical features similar to those of human RA. The interplay between the gut microbiota and inflammatory arthritis in animal models has been demonstrated in vivo [82]. Previous studies reported that the alteration of the gut microbiota composition marked the preclinical phase of murine CIA and preceded the development of disease [83, 84]. Compared with nontreated mice, CIA-susceptible HLA-DQ8 mice treated with C. aerofaciens increased the incidence and severity of arthritis [69]. Liu et al. showed that the microbial richness and diversity were different between mice that were susceptible and resistant to CIA before the initiation of arthritis. With the progression of CIA, the abundance of the operational taxonomic units (OTUs) affiliated with the families Bacteroidaceae, Lachnospiraceae, and S24-7 was markedly upregulated in CIA-susceptible mice; if germ-free mice were administered the microbiota from either CIA-susceptible or CIA-resistant mice, the former group presented a higher incidence of arthritis with increased IL-17 levels and CD8+ T cell and Th17 lymphocyte proportions but decreased dendritic cells (DCs), B cells, and Treg cells in the spleen than the latter group [85]. The immune response and the gut microbiota profiles occur at different stages of CIA. Nemoto et al. reported that the percentage of Foxp3+CD4+ T cells was only increased in the mesenteric lymph nodes (MLNs) in the relapse stage of CIA. The percentage of RORγ+CD4+ T cells was increased in the MLNs at the initial peak and was decreased in the relapse stage of CIA, but the opposite changes were observed in the spleen. The concentration of IgA in the faeces increased with the progression of arthritis and showed positive correlations with Bacteroidales in the CIA group [86]. Gut commensal bacteria can regulate gut immunity. Balakrishnan et al. showed that RA-associated bacteria (Eggerthella lenta or Collinsella aerofaciens) enhanced gut permeability in DQ8 mice. The splenocytes from naive DQ8 mice gavaged with E. lenta produced the proinflammatory cytokines IL-6, IL-21, and IL-23. When compared with the controls, DQ8 mice gavaged with non-RA-associated bacteria (Prevotella histicola or Bifidobacterium sp.) exhibited decreased numbers of inflammatory monocytes and CD11c+Ly6c+ cells and reduced levels of proinflammatory monocyte chemotactic protein- (MCP-) 1 and MCP-3 [87]. In a recent metabolomic study, altered metabolites contained citric acid and l-isoleucine both in the serum and in the faeces were identified in rats with CIA compared with the control rats [88]. Jubair et al. found that in the CIA model, the absence of a dominant microbiota resulted in an approximately 40% reduction in disease severity through the regulation of mucosal and systemic cytokines and autoantibodies, and gut microbial dysbiosis was associated with the mucosal Th17 immune response, stimulating mucosal lymphoid tissue-producing autoantibodies and regulating autoantibody effector functions in the preclinical phase of CIA [89]. Maeda et al. suggested that T cells from germ-free, arthritis-prone SKG mice were activated in the intestine by dysregulated microbiota from RA patients, which caused joint inflammation [90]. Certain antibiotics can induce sustained changes in gut immunity by increasing the ratios of Th1 and Th17 cells in the MLNs, which might be responsible for the aggravation of CIA and attributed to the disruption dysregulation of microbes [91].
Whether the dysregulation of the gut microbiota precedes the onset of RA or is a consequence of RA still needs to be investigated. Previous research showed that the gut microbiota could be changed with the aggravation of CIA by orally administering Porphyromonas gingivalis [92]. Hablot et al. suggested that concomitant experimental colitis in mice with CIA could slightly delay arthritis onset and reduce arthritis severity, which was associated with changes in the gut microbial composition [93]. The difference in the faecal microbial composition was correlated with disease severity in CIA mice [94]. The partial depletion of the natural gut microbiota, which was observed in the colonic content, could aggravate CIA symptoms and increase in vitro Th1/Th17 cytokine production by axillary lymph node cells (ALNCs) from arthritic mice treated with antibiotics compared to the positive control [95]. However, in contrast to these findings, Wing et al. showed that the increased CIA susceptibility of germ-free, reactive oxygen species- (ROS-) deficient Ncf1 mutant mice was not dependent on commensal bacteria when they were reared in specific pathogen-free (SPF) conditions [96]. These differences may be attributed to the distinct local bacteria in the respective SPF facilities where the different experiments were conducted.
4.3. Gut Microbiota in Distinct Compartments of the Gastrointestinal Tract Plays Region-Specific Roles in RA (and CIA)
Gut microbiota dysbiosis could contribute to the onset and development of RA, and one of the mechanisms may be through the disruption of the proportions and activation status of local ICs along the intestine, including the dysregulation of effector and regulatory ICs. Microbial analyses of stool samples have been accepted as methods to explore the relationship of the gut microbiota with RA and CIA [98]. However, functional heterogeneity of distinct gastrointestinal tract segments gives rise to regional differences in the gut microbiota [30], with its components that closely interact with their neighbouring ICs, and both display regional specificity within the same individual [99]. The lower gastrointestinal tract comprises a variety of different microbiota along the small intestine, caecum, and colon. The small intestine contains lower microbial diversity than the caecum and colon [100]. Asquith et al. showed that the microbiota composition was strikingly different in stool samples compared to that in mucosal samples, and there was also a marked difference in the ileal site compared with the colonic site [67]. In addition, a less diverse bacterial population was found in the ileum (mucosa and lumen) than that in the caecum of broiler chickens, by terminal restriction fragment length polymorphism (T-RFLP) analysis and sequence analyses of 16S rRNA genes [101]. Another study reported that early antibiotic exposure in suckling piglets changed the abundance of the gut microbiota and its related metabolites in the ileal digesta, which were different from those in the caecal digesta [102]. In aged rats, Lee et al. found that the β-diversity of the microbiota was higher in the ileum than in the caecum, but the α-diversity of microbiota composition was higher in the caecum than in the ileum. The family Lactobacillaceae was more enriched in the ileum than in the caecum, while Ruminococcaceae and Lachnospiraceae were more abundant in the caecum [31]. Doonan et al. demonstrated that the gut microbiota showed different dysregulations between ileal and colonic contents of mice with CIA; the former exhibited outgrowths of Firmicutes and Proteobacteria in the ileum and decreased Firmicutes with a compensatory increase in Bacteroidetes in the colon, and treatment with ES-62 increased the overall species diversity in the ileum but not in the colon of mice with CIA [103]. Thus, it may not be sufficient to measure faecal samples in order to explore the mechanisms of RA that are mediated by the numerous intestinal microbes in the host.
The large intestine and the small intestine have different functions, in which the functions of the large intestine are mainly the extraction of water and salt from solid wastes, while the main functions of the small intestine are the absorption of nutrients and minerals contained in the diet. In addition, the distinct regions of the intestinal tract not only exhibit different functions but also possess different properties of immune cells with distinct roles in immune modulation [53]. The human colon and ileum contain DCs with distinct roles in mucosal immunity; thus, they should be regarded as separate entities. Mann et al. indicated that a lower proportion of colonic DCs produced TNF-α and IL-1β when compared with their (paired) ileal counterparts and that colonic DCs exhibited an enhanced ability to induce the expansion/development of CD4+FoxP3+IL-10+ T cells (Treg). It also demonstrated that colonic and ileal DCs have different abilities to imprint homing properties on T cells [104]. RORγ+ Treg, as a unique microbe-responsive cell type, were detected in lower proportions in the small intestine compared with those in the colon [105]. A recent analysis of T cells from iFoxp3 mice after 5 weeks of tamoxifen pulse labelling demonstrated a significant reduction in the frequency and number of CD4+Foxp3+ T cells in most of the analysed compartments (including MLNs, small intestine epithelium, and lamina propria), except the large intestine, but found an accumulation of intraepithelial CD4+ in the small intestine. The plasticity of the Treg cells in the epithelium is microbiota dependent, and these cells exhibit intratissue specialization, which is shaped by discrete niches in the intestine [106]. The follicle-associated epithelium (FAE) of the human ileum, which includes Peyer's patches, is functionally distinct from the regular villus epithelium (VE) because the former is more prone to bacterial-epithelial cell interaction and antigen delivery to the mucosal immune system [107]. The partial elimination of the gut microbiota during established CIA modulates the mucosal T helper cell balance, which mainly presents differently in distinct tracts of the intestine. Broad-spectrum antibiotic (ABX) treatment of mice with CIA significantly reduced the expression of IL-17 mRNA in the terminal ileum, which is a main site for the microbiota-induced T cell modulation, while ABX treatment did not affect the expression of IL-17 in the colon. In addition, the expression of the Treg-related transcription factor FoxP3 was significantly upregulated in the colon tissue of ABX-treated mice, but it was not affected in the small intestine [83]. A proteome analysis study found that the colon mucosa could trigger the production of ACPAs, which is known to contribute to the onset of RA [108]. Therefore, the interplay of the gut microbiota and intestinal ICs may participate in RA in a special tissue-dependent manner that is shaped by discrete compartments in the intestine. Observations of the compartmentalization of the ICs and gut microbial species along specific regions of the intestine at a steady state in RA may provide a platform for understanding the pathogenesis of this disease. Therefore, characterizing the microbial alterations in distinct intestinal tracts during established arthritis may be suggested.
5. Targeting the Gut Microbiota for the Treatment of RA
Recent studies have proposed that the commensal microbiota is one of the important environmental triggers of RA due to its interactions with the host immune system [2]. Therefore, immunosuppressive agents that can restore the gut microbial composition and immunologic balance may act as therapeutic drugs for inflammatory arthritis (Table 2).
Table 2.
The partial restoration of the gut microbiota in the treatment of arthritis.
| Objects and therapy | Samples | Technology | Increased | Decreased | References |
|---|---|---|---|---|---|
| Human, etanercept group vs. naive patient group | Stool | Metagenomic sequencing | Cyanobacteria, Nostocophycideae, Nostocales | Deltaproteobacteria, Clostridiaceae | [76] |
| Mice, kaempferol group vs. CIA group | Stool | 16S rRNA sequencing | Bacteroidales_S24-7_group, Prevotellaceae, Erysipelotrichaceae, Alcaligenaceae | Lachnospiraceae, Staphylococcaceae | [115] |
| Mice, ES-62 group vs. CIA group | Ileal and colonic content | Metagenomic shotgun sequencing | Clostridaceae, Lachnospiraceae (in the ileum) | Helicobacter, Escherichia (in the colon) | [103] |
| Mice, PSE group vs. CIA group | Stool | 16S rRNA sequencing | Bacteriodetes, S24-7, Rikenella | Desulfovibrio, Rikenellaceae_RC9, Mucispirillum, Helicobacter, Lachnospiraceae | [110] |
| Rats, berberine group vs. CIA group | Caecal content | 16S rRNA sequencing | Blautia, Butyricicoccus, Parabacteroides | Prevotella, Paraprevotella, Coprococcus | [109] |
| Rats, TGP group vs. CIA group | Stool | 16S rRNA sequencing | Tenericutes, Mollicutes, Mollicutes RF9, Christensenellaceae, Unclassified_Erysipelotrichaceae, Anaerovorax | [111] | |
| Rats, QLT group vs. AA group | Stool | 16S rRNA gene sequencing | Ruminococcus_1, Clostridium_sensu_stricto_1, Atopostipes, Turicibacter, Ruminococcaceae_UCG-013, Roseburia | Anaerofustis, Blautia, Parasutterella, Leuconostoc | [114] |
| CIA rats, ZT group vs. CIA group | Stool | 16S rRNA gene sequencing | Coriobacteriaceae | Bacteroidaceae, Porphyromonadaceae | [88] |
| AA rats, L. casei-treated group vs. AA group | Stool | Metagenomic sequencing | Acinetobacter unclassified, Corynebacterium casei, L. acidophilus | Corynebacterium urealyticum, Desulfovibrio desulfuricans, Erysipelotrichaceae | [116] |
CIA: collagen-induced arthritis; AA: adjuvant-induced arthritis; PSE: Paederia scandens extract; TGP: total glucosides of paeony; QLT: Qingluo Tongbi decoction; ZT: Zushima tablet.
5.1. Gut Microbiota Mediate the Effect of Drugs on RA
The gut microbial composition is quite different between RA patients who received disease-modifying antirheumatic drugs (DMARDs) and healthy controls. Rodrigues et al. found that relative expression units (REU) of Bacteroides and Prevotella species were increased and REU of Clostridium leptum in the faecal samples from RA patients receiving DMARDs were decreased compared with those in samples from healthy controls [97]. The administration of clindamycin for 4 weeks increased the incidence and severity of CIA in mice, and the abundance of anaerobic bacteria was significantly decreased compared to that observed after vancomycin administration [91]. Yue et al. demonstrated that oral administration of berberine can ameliorate symptoms in rats with CIA by downregulating the diversity and richness of gut bacteria, including the abundance of Prevotella, and upregulating the abundance of butyrate-producing bacteria to increase the generation of butyrate and stabilize intestinal hypoxia and nitrate supply [109]. Doonan et al. indicated that the subcutaneous administration of ES-62 (an immunomodulator secreted by tissue-dwelling Acanthocheilonema) could protect against joint disease in mice with CIA, which was associated with the normalization of the gut microbiota and the prevention of intestinal barrier integrity loss [103]. Paederia scandens extract (PSE) effectively inhibited paw swelling, tissue fibrosis, and inflammatory cell infiltration and decreased the serum levels of TNF-α, IL-1β, IL-6, IL-7, and IL-23 in mice with CIA. Moreover, PSE treatment restored the gut microbial ecosystem of mice with CIA by decreasing the relative abundance of inflammation-related microorganisms, including Desulfovibrio, Mucispirillum, Helicobacter, and Lachnospiraceae [110]. A long-term experiment indicated that the gut microbiota might play a profound role in mediating the therapeutic effects of total glucosides of paeony (TGP) in rats with CIA because of its capability to significantly reverse the taxonomic changes in this animal model, to increase the relative abundance of beneficial symbiotic bacteria, and to inhibit the levels of intestinal cytokines, sIgA and IFN-γ [111]. The oral administration of the gut microbial metabolites SCFAs ameliorated the severity of CIA, which was associated with inhibiting Th1 cells but promoting Treg cells [112]. Guo et al. reported that total clematis triterpenoid saponins (CTSs) in CIA rats could improve arthritis symptoms and significantly downregulate the total SCFA concentration, and by LEfSe and DESeq2 analyses, the CTSs could restore the most significantly increased Gram-negative (G(-)) and decreased Gram-positive (G(+)) genera [113].
Certain formulas of traditional Chinese medicine (TCM) also have an effect on RA by impacting the gut microbiota. Qingluo Tongbi decoction (QLT) had a beneficial effect on altered bacterial genera and families (Lachnospiraceae, Eubacteriaceae, and Leuconostocaceae) in rats with adjuvant-induced arthritis (AA) and significantly decreased the expression levels of cadherin-11, IL-17α, TLR2, and TLR4 in synovial tissues, which negatively correlated with the abundance of Staphylococcus and Candidatus_Saccharimonas [114]. The families of bacteria in the faeces and the metabolites in both the serum and faeces were altered in rats with CIA, and the Zushima tablet (ZT) restored most of these metabolites (including l-isoleucine, l-aspartic acid, pyruvic acid, cholic acid, and hypoxanthine) and bacteria (such as Coriobacteriaceae, Bacteroidaceae, and Porphyromonadaceae) [88]. The route of administration may also influence the effect of drugs mediated by the gut microbiota and their metabolites on animals with CIA. It has been reported that kaempferol treatment administered intragastrically rather than intraperitoneally could restore the gut microbiota composition and regulate the microbial metabolism (energy production and tryptophan, fatty acid, and secondary bile acid) in CIA mice and might be responsible for the antiarthritis effects [115].
5.2. Probiotics Would Be a Potential Therapy for RA
The gut microbiota might serve as a therapeutic target for many kinds of diseases. Restoring the aberrant gut microbiota to the healthy state is a potential therapeutic approach for preventing RA. At present, resetting gut microbial dysbiosis through probiotics, prebiotics, or FMT is emerging as a potential approach for the prevention and treatment of RA. Probiotics are nonpathogenic microorganisms that can interact with the gut microbiota and provide benefits for the host. The treatment of rats with AA with Lactobacillus casei (L. casei, ATCC334) could inhibit joint swelling, decrease arthritis scores, improve bone destruction, restore some Lactobacillus strains to normal, and decrease the expression of the proinflammatory cytokines IFN-γ, TNF-α, IL-1β, IL-17, and IL-6 [116]. The oral administration of L. casei had an antiarthritic effect and inhibited cyclooxygenase- (COX-) 2 by decreasing proinflammatory cytokines in a model of CIA [117]. Liu et al. demonstrated that the administration of L. salivarius UCC118 and L. plantarum WCFS1 isolated from RA patients could alleviate arthritis in mice with CIA, decrease Th17 cells, and increase Treg, and L. salivarius-treated mice with CIA also showed a significant increase in the anti-inflammatory IL-10 serum levels [118]. Esvaran et al. reported that the administration of L. fermentum PC1 could markedly reduce paw inflammation and synovial infiltration, attenuate cartilage damage, decrease the proinflammatory cytokine IL-12, and increase the anti-inflammatory cytokines IL-4 and IL-10 in DBA/1 mice with CIA [119]. Yamashita et al. suggested that both the oral administration and intraperitoneal injection of L. helveticus SBT2171 could prevent the CIA symptoms of mice and decrease the subsequent production of bovine type II collagen- (bCII-) specific antibodies; in addition, intraperitoneal injection of L. helveticus SBT2171 could also reduce the numbers of immune cells, including the total B cells, germinal centre B cells, and CD4+ T cells, in the draining lymph nodes and the serum level of IL-6 [120]. Oral administration of L. delbrueckii subsp. bulgaricus OLL 1073R-1 prevented CIA in DBA/1J mice, inhibited the secretion of proinflammatory cytokine IFN-γ by lymph node cells in response to bCII, and reduced the IL-6, TNF-α, and MCP-1 produced by accessory cells [121, 122]. Moreover, probiotic administration in RA may reduce organ damage to a greater extent than nonsteroidal anti-inflammatory drugs (NSAIDs). In Wistar rats with CIA, L. acidophilus decreased arthritis scores and maintained the normal histology of reproductive organs and the oxidative stress parameters in ovaries and testes [123]. Hosoya et al. reported that the intraperitoneal administration of L. helveticus SBT2171 (LH2171) alleviated CIA symptoms in DBA/1J mice, and the mechanisms might be the inhibition of excessive lymphocyte proliferation and the generation of immunosuppressive effects in vivo [47]. The oral administration of L. casei suppressed arthritis severity in rats with CIA, which was associated with the more effective inhibition of CII-reactive Th1-type IgG isotypes (IgG2a and IgG2b), the more effective promotion of IL-10 levels, and the reduction in proinflammatory molecules (such as IL-1β, IL-2, IL-6, IL-12, IL-17, IFN-γ, TNF-α, and COX-2) by CD4+ T cells [124]. The systemic administration of exopolysaccharide (EPS), derived from L. rhamnosus KL37, could ameliorate active CIA induced by the systemic injection of collagen and lipopolysaccharide via the inhibition of arthritogenic CII-specific antibody production [125]. In addition, dietary interventions targeting the microbiota may become a potential therapy for RA. A recent study proved that after 28 days of intervention with a high-fibre diet, RA patients had increased circulating Treg cell numbers, favourable Th1/Th17 ratios, and improved symptoms, which might be due to the regulation of the gut microbiota and microbial metabolites [126].
6. Perspective and Conclusion
The interactions between the gut microbiota and ICs, especially intestinal ICs, may hold the keys for developing novel biomarkers and treatment strategies as well as for understanding the pathophysiology of RA (Figure 1). However, the mechanisms of the alteration of specific gut microbiota clusters in RA need to be further investigated. Studies are also required to explore whether some specific gut microbial dysbiosis is either an active driver of RA or is only an epiphenomenon. Furthermore, the potential molecular mechanisms of the therapeutic actions of probiotics for RA remain unclear. Additional studies need to be conducted to explore the involvement of the gut microbiota in distinct intestinal tracts in immune modulation during RA pathogenesis.
Figure 1.
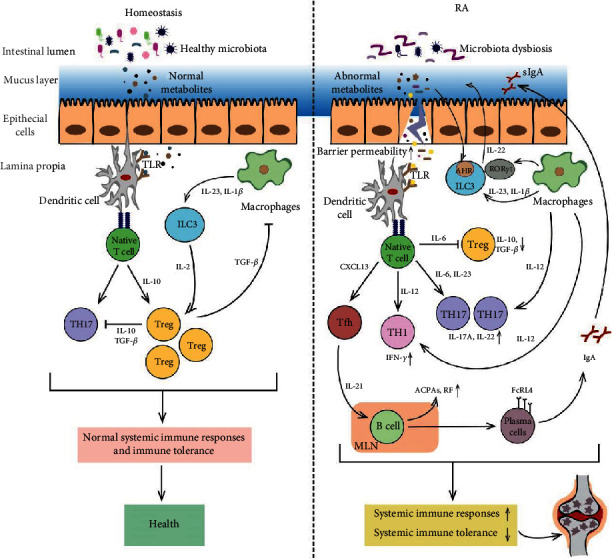
Gut microbiota dysbiosis contributes to the pathogenesis of RA, and a variety of ICs act as a bridge linking the gut microbiota and RA via multiple molecular mechanisms. The healthy gut microbiota and their normal metabolites maintain the integrity of the intestinal epithelial cell layer and the homeostasis of gut immunity.
Acknowledgments
This research was funded by grants from the National Natural Science Foundation of China (81673844, 81873223 to CX and 81573845 to HYZ) and the International Cooperation Project of the Ministry of Science and Technology (No. 2014DFA31490 to CX).
Contributor Information
Cheng Xiao, Email: xc2002812@126.com.
Qingdong Guan, Email: qguan@hsc.mb.ca.
Conflicts of Interest
The authors declare no conflict of interest regarding this manuscript.
Authors' Contributions
CX and QG proposed the manuscript and formulated the concept; HHX and HYZ wrote the manuscript; JFC and YX searched and arranged documents; HHX and DPF made the figures; MJL, DHJ, CX, and QG revised the manuscript. All the authors have approved the final manuscript. HHX and HYZ contributed equally to this work.
References
- 1.Reina-Bueno M., Gonzalez-Lopez J. R., Lopez-Lopez D., et al. Development and validation of the overall foot health questionnaire for patients with rheumatoid arthritis: a cross-sectional descriptive analysis. Medicina. 2019;55(6):p. 290. doi: 10.3390/medicina55060290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lucchino B., Spinelli F. R., Iannuccelli C., Guzzo M. P., Conti F., Di Franco M. Mucosa-environment interactions in the pathogenesis of rheumatoid arthritis. Cells. 2019;8:p. 700. doi: 10.3390/cells8070700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lu D. R., McDavid A. N., Kongpachith S., et al. T cell-dependent affinity maturation and innate immune pathways differentially drive autoreactive B cell responses in rheumatoid arthritis. Arthritis & Rhematology. 2018;70(11):1732–1744. doi: 10.1002/art.40578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Picchianti Diamanti A., Rosado M. M., Scarsella M., et al. Abatacept (cytotoxic T lymphocyte antigen 4-immunoglobulin) improves B cell function and regulatory T cell inhibitory capacity in rheumatoid arthritis patients non-responding to anti-tumour necrosis factor-α agents. Clinical and Experimental Immunology. 2014;177(3):630–640. doi: 10.1111/cei.12367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rao D. A., Gurish M. F., Marshall J. L., et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature. 2017;542(7639):110–114. doi: 10.1038/nature20810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kuwabara T., Ishikawa F., Kondo M., Kakiuchi T. The role of IL-17 and related cytokines in inflammatory autoimmune diseases. Mediators of Inflammation. 2017;2017:11. doi: 10.1155/2017/3908061.3908061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Xu H., Zhao H., Lu C., et al. Triptolide inhibits osteoclast differentiation and bone ResorptionIn Vitrovia enhancing the production of IL-10 and TGF-β1 by regulatory T cells. Mediators of Inflammation. 2016;2016:10. doi: 10.1155/2016/8048170.8048170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yang P., Qian F., Zhang M., et al. Zishen Tongluo formula ameliorates collagen-induced arthritis in mice by modulation of Th17/Treg balance. Journal of Ethnopharmacology. 2020;250:p. 112428. doi: 10.1016/j.jep.2019.112428. [DOI] [PubMed] [Google Scholar]
- 9.Niu H. Q., Li Z. H., Zhao W. P., et al. Sirolimus selectively increases circulating Treg cell numbers and restores the Th17/Treg balance in rheumatoid arthritis patients with low disease activity or in DAS28 remission who previously received conventional disease-modifying anti-rheumatic drugs. Clinical and Experimental Rheumatology. 2020;38(1):58–66. [PubMed] [Google Scholar]
- 10.Vasilev G., Ivanova M., Ivanova-Todorova E., et al. Secretory factors produced by adipose mesenchymal stem cells downregulate Th17 and increase Treg cells in peripheral blood mononuclear cells from rheumatoid arthritis patients. Rheumatology International. 2019;39(5):819–826. doi: 10.1007/s00296-019-04296-7. [DOI] [PubMed] [Google Scholar]
- 11.Li C., Zhang J., Wang W., Wang H., Zhang Y., Zhang Z. Arsenic trioxide improves Treg and Th17 balance by modulating STAT3 in treatment-naive rheumatoid arthritis patients. International Immunopharmacology. 2019;73:539–551. doi: 10.1016/j.intimp.2019.05.001. [DOI] [PubMed] [Google Scholar]
- 12.Li C., Zhang J., Wang W., Wang H., Zhang Y., Zhang Z. Data on arsenic trioxide modulates Treg/Th17/Th1/Th2 cells in treatment-naïve rheumatoid arthritis patients and collagen-induced arthritis model mice. Data in Brief. 2019;27:p. 104615. doi: 10.1016/j.dib.2019.104615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ramwadhdoebe T. H., van Baarsen L. G. M., Boumans M. J. H., et al. Effect of rituximab treatment on T and B cell subsets in lymph node biopsies of patients with rheumatoid arthritis. Rheumatology. 2019;58(6):1075–1085. doi: 10.1093/rheumatology/key428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Amara K., Clay E., Yeo L., et al. B cells expressing the IgA receptor FcRL4 participate in the autoimmune response in patients with rheumatoid arthritis. Journal of Autoimmunity. 2017;81:34–43. doi: 10.1016/j.jaut.2017.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Xiao C., Zhao L., Liu Z., et al. The effect of triptolide on CD4+ and CD8+ cells in the Peyer’s patch of DA rats with collagen induced arthritis. Natural Product Research. 2009;23(18):1699–1706. doi: 10.1080/14786410802187783. [DOI] [PubMed] [Google Scholar]
- 16.Bessis N., Decker P., Assier E., Semerano L., Boissier M. C. Arthritis models: usefulness and interpretation. Seminars in Immunopathology. 2017;39(4):469–486. doi: 10.1007/s00281-017-0622-4. [DOI] [PubMed] [Google Scholar]
- 17.Wang T., Wei Z. F., Dou Y. N., Xia Y. F., Dai Y. Effect of madecassoside on intestinal mucosal immunity in collagen-induced arthritis rats. Zhong Yao Cai. 2015;38(2):333–338. [PubMed] [Google Scholar]
- 18.Evans‐Marin H., Rogier R., Koralov S. B., et al. Microbiota-dependent involvement of Th17 cells in murine models of inflammatory arthritis. Arthritis & Rhematology. 2018;70(12):1971–1983. doi: 10.1002/art.40657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kastbom A., Roos Ljungberg K., Ziegelasch M., Wettero J., Skogh T., Martinsson K. Changes in anti-citrullinated protein antibody isotype levels in relation to disease activity and response to treatment in early rheumatoid arthritis. Clinical and Experimental Immunology. 2018;194(3):391–399. doi: 10.1111/cei.13206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wells P. M., Williams F. M. K., Matey-Hernandez M. L., Menni C., Steves C. J. RA and the microbiome: do host genetic factors provide the link? Journal of Autoimmunity. 2019;99:104–115. doi: 10.1016/j.jaut.2019.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Heydari-Kamjani M., Beckler M. D., Kesselman M. M. Reconsidering the use of minocycline in the preliminary treatment regime of rheumatoid arthritis. Cureus. 2019;11, article e5351 doi: 10.7759/cureus.5351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Maeda Y., Takeda K. Host-microbiota interactions in rheumatoid arthritis. Experimental & Molecular Medicine. 2019;51(12):1–6. doi: 10.1038/s12276-019-0283-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ribaldone D. G., Caviglia G. P., Abdulle A., et al. Adalimumab therapy improves intestinal dysbiosis in Crohn’s disease. Journal of Clinical Medicine. 2019;8(10):p. 1646. doi: 10.3390/jcm8101646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Masoodi I., Alshanqeeti A. S., Ahmad S., et al. Microbial dysbiosis in inflammatory bowel diseases: results of a metagenomic study in Saudi Arabia. Minerva Gastroenterologica e Dietologica. 2019;65(3):177–186. doi: 10.23736/S1121-421X.19.02576-5. [DOI] [PubMed] [Google Scholar]
- 25.Nishida A., Inoue R., Inatomi O., Bamba S., Naito Y., Andoh A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clinical Journal of Gastroenterology. 2018;11(1):1–10. doi: 10.1007/s12328-017-0813-5. [DOI] [PubMed] [Google Scholar]
- 26.Fulde M., Sommer F., Chassaing B., et al. Neonatal selection by Toll-like receptor 5 influences long-term gut microbiota composition. Nature. 2018;560(7719):489–493. doi: 10.1038/s41586-018-0395-5. [DOI] [PubMed] [Google Scholar]
- 27.Yacoub R., Jacob A., Wlaschin J., McGregor M., Quigg R. J., Alexander J. J. Lupus: the microbiome angle. Immunobiology. 2018;223(6-7):460–465. doi: 10.1016/j.imbio.2017.11.004. [DOI] [PubMed] [Google Scholar]
- 28.Qi X. Z., Tu X., Zha J. W., Huang A. G., Wang G. X., Ling F. Immunosuppression-induced alterations in fish gut microbiota may increase the susceptibility to pathogens. Fish & Shellfish Immunology. 2019;88:540–545. doi: 10.1016/j.fsi.2019.03.035. [DOI] [PubMed] [Google Scholar]
- 29.Guan Q. A comprehensive review and update on the pathogenesis of inflammatory bowel disease. Journal of Immunology Research. 2019;2019:16. doi: 10.1155/2019/7247238.7247238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Martinez-Guryn K., Leone V., Chang E. B. Regional diversity of the gastrointestinal microbiome. Cell Host & Microbe. 2019;26(3):314–324. doi: 10.1016/j.chom.2019.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee S. M., Kim N., Park J. H., Nam R. H., Yoon K., Lee D. H. Comparative analysis of ileal and cecal microbiota in aged rats. Journal of Cancer Prevention. 2018;23(2):70–76. doi: 10.15430/JCP.2018.23.2.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tanoue T., Umesaki Y., Honda K. Immune responses to gut microbiota-commensals and pathogens. Gut Microbes. 2014;1(4):224–233. doi: 10.4161/gmic.1.4.12613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nell S., Suerbaum S., Josenhans C. The impact of the microbiota on the pathogenesis of IBD: lessons from mouse infection models. Nature Reviews. Microbiology. 2010;8(8):564–577. doi: 10.1038/nrmicro2403. [DOI] [PubMed] [Google Scholar]
- 34.Saleh M., Elson C. O. Experimental inflammatory bowel disease: insights into the host-microbiota dialog. Immunity. 2011;34(3):293–302. doi: 10.1016/j.immuni.2011.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Caviglia G. P., Rosso C., Ribaldone D. G., et al. Physiopathology of intestinal barrier and the role of zonulin. Minerva Biotecnologica. 2019;31(3):83–92. doi: 10.23736/s1120-4826.19.02554-0. [DOI] [Google Scholar]
- 36.Kalinkovich A., Livshits G. A cross talk between dysbiosis and gut-associated immune system governs the development of inflammatory arthropathies. Seminars in Arthritis and Rheumatism. 2019;49(3):474–484. doi: 10.1016/j.semarthrit.2019.05.007. [DOI] [PubMed] [Google Scholar]
- 37.Stewart L., Edgar J. D. M., Blakely G., Patrick S. Antigenic mimicry of ubiquitin by the gut bacterium Bacteroides fragilis: a potential link with autoimmune disease. Clinical and Experimental Immunology. 2018;194(2):153–165. doi: 10.1111/cei.13195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fredricks D. N. The gut microbiota and graft-versus-host disease. The Journal of Clinical Investigation. 2019;129:1808–1817. doi: 10.1172/jci125797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Durazzo M., Ferro A., Gruden G. Gastrointestinal microbiota and type 1 diabetes mellitus: the state of art. Journal of Clinical Medicine. 2019;8(11):p. 1843. doi: 10.3390/jcm8111843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jeong Y., Kim J. W., You H. J., et al. Gut microbial composition and function are altered in patients with early rheumatoid arthritis. Journal of Clinical Medicine. 2019;8(5):p. 693. doi: 10.3390/jcm8050693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Oh J., Unutmaz D. Immune cells for microbiota surveillance. Science. 2019;366(6464):419–420. doi: 10.1126/science.aaz4014. [DOI] [PubMed] [Google Scholar]
- 42.Konig M. F. The microbiome in autoimmune rheumatic disease. Best Practice & Research. Clinical Rheumatology. 2020;34(1, article 101473) doi: 10.1016/j.berh.2019.101473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Luu M., Visekruna A. Short-chain fatty acids: bacterial messengers modulating the immunometabolism of T cells. European Journal of Immunology. 2019;49(6):842–848. doi: 10.1002/eji.201848009. [DOI] [PubMed] [Google Scholar]
- 44.Ding T., Niu H., Zhao X., Gao C., Li X., Wang C. T-follicular regulatory cells: potential therapeutic targets in rheumatoid arthritis. Frontiers in Immunology. 2019;10:p. 2709. doi: 10.3389/fimmu.2019.02709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Honda K., Littman D. R. The microbiota in adaptive immune homeostasis and disease. Nature. 2016;535(7610):75–84. doi: 10.1038/nature18848. [DOI] [PubMed] [Google Scholar]
- 46.Maynard C. L., Elson C. O., Hatton R. D., Weaver C. T. Reciprocal interactions of the intestinal microbiota and immune system. Nature. 2012;489(7415):231–241. doi: 10.1038/nature11551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hosoya T., Sakai F., Yamashita M., et al. Lactobacillus helveticus SBT2171 inhibits lymphocyte proliferation by regulation of the JNK signaling pathway. PLoS One. 2014;9(9, article e108360) doi: 10.1371/journal.pone.0108360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yang Y., Torchinsky M. B., Gobert M., et al. Focused specificity of intestinal TH17 cells towards commensal bacterial antigens. Nature. 2014;510(7503):152–156. doi: 10.1038/nature13279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kawano M., Miyoshi M., Miyazaki T. Lactobacillus helveticus SBT2171 induces A20 expression via toll-like receptor 2 signaling and inhibits the lipopolysaccharide-induced activation of nuclear factor-kappa B and mitogen-activated protein kinases in peritoneal macrophages. Frontiers in Immunology. 2019;10:p. 845. doi: 10.3389/fimmu.2019.00845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Blagojevic V., Kovacevic-Jovanovic V., Curuvija I., et al. Rat strain differences in peritoneal immune cell response to selected gut microbiota: a crossroad between tolerance and autoimmunity? Life Sciences. 2018;197:147–157. doi: 10.1016/j.lfs.2018.02.011. [DOI] [PubMed] [Google Scholar]
- 51.Tanoue T., Morita S., Plichta D. R., et al. A defined commensal consortium elicits CD8 T cells and anti-cancer immunity. Nature. 2019;565(7741):600–605. doi: 10.1038/s41586-019-0878-z. [DOI] [PubMed] [Google Scholar]
- 52.Ahluwalia B., Magnusson M. K., Ohman L. Mucosal immune system of the gastrointestinal tract: maintaining balance between the good and the bad. Scandinavian Journal of Gastroenterology. 2017;52(11):1185–1193. doi: 10.1080/00365521.2017.1349173. [DOI] [PubMed] [Google Scholar]
- 53.Morhardt T. L., Hayashi A., Kao J. Y., Kamada N. Regional control of regulatory immune cells in the intestine. Current Pathobiology Reports. 2018;6(1):29–34. doi: 10.1007/s40139-018-0156-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Parikh K., Antanaviciute A., Fawkner-Corbett D., et al. Colonic epithelial cell diversity in health and inflammatory bowel disease. Nature. 2019;567(7746):49–55. doi: 10.1038/s41586-019-0992-y. [DOI] [PubMed] [Google Scholar]
- 55.Zhou L., Chu C., Teng F., et al. Innate lymphoid cells support regulatory T cells in the intestine through interleukin-2. Nature. 2019;568(7752):405–409. doi: 10.1038/s41586-019-1082-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mao K., Baptista A. P., Tamoutounour S., et al. Innate and adaptive lymphocytes sequentially shape the gut microbiota and lipid metabolism. Nature. 2018;554(7691):255–259. doi: 10.1038/nature25437. [DOI] [PubMed] [Google Scholar]
- 57.Li Z., Quan G., Jiang X., et al. Effects of metabolites derived from gut microbiota and hosts on pathogens. Frontiers in Cellular and Infection Microbiology. 2018;8:p. 314. doi: 10.3389/fcimb.2018.00314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hui W., Yu D., Cao Z., Zhao X. Butyrate inhibit collagen-induced arthritis via Treg/IL-10/Th17 axis. International Immunopharmacology. 2019;68:226–233. doi: 10.1016/j.intimp.2019.01.018. [DOI] [PubMed] [Google Scholar]
- 59.Shen B., Hu J., Song H., et al. Antibiotics exacerbated colitis by affecting the microbiota, Treg cells and SCFAs in IL10-deficient mice. Biomedicine & Pharmacotherapy. 2019;114, article 108849 doi: 10.1016/j.biopha.2019.108849. [DOI] [PubMed] [Google Scholar]
- 60.Morita N., Umemoto E., Fujita S., et al. GPR31-dependent dendrite protrusion of intestinal CX3CR1(+) cells by bacterial metabolites. Nature. 2019;566(7742):110–114. doi: 10.1038/s41586-019-0884-1. [DOI] [PubMed] [Google Scholar]
- 61.Schiering C., Wincent E., Metidji A., et al. Feedback control of AHR signalling regulates intestinal immunity. Nature. 2017;542(7640):242–245. doi: 10.1038/nature21080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Li S., Bostick J. W., Zhou L. Regulation of innate lymphoid cells by aryl hydrocarbon receptor. Frontiers in Immunology. 2018;2017:p. 8. doi: 10.3389/fimmu.2017.01909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Li X., Lu C., Fan D., et al. Human umbilical mesenchymal stem cells display therapeutic potential in rheumatoid arthritis by regulating interactions between immunity and gut microbiota via the aryl hydrocarbon receptor. Frontiers in Cell and Development Biology. 2020;8:p. 131. doi: 10.3389/fcell.2020.00131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Gomez A., Luckey D., Yeoman C. J., et al. Loss of sex and age driven differences in the gut microbiome characterize arthritis-susceptible 0401 mice but not arthritis-resistant 0402 mice. PLoS One. 2012;7(4, article e36095) doi: 10.1371/journal.pone.0036095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lu Q., Lemke G. Homeostatic regulation of the immune system by receptor tyrosine kinases of the Tyro 3 family. Science. 2001;293(5528):306–311. doi: 10.1126/science.1061663. [DOI] [PubMed] [Google Scholar]
- 66.Waterborg C. E., Koenders M. I., van Lent P. L., van der Kraan P. M., van de Loo F. A. Tyro3/Axl/Mertk-deficient mice develop bone marrow edema which is an early pathological marker in rheumatoid arthritis. PLoS One. 2018;13(10, article e0205902) doi: 10.1371/journal.pone.0205902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Asquith M., Sternes P. R., Costello M.-E., et al. HLA alleles associated with risk of ankylosing spondylitis and rheumatoid arthritis influence the gut microbiome. Arthritis & Rhematology. 2019;71(10):1642–1650. doi: 10.1002/art.40917. [DOI] [PubMed] [Google Scholar]
- 68.Wang Q., Xu R. Data-driven multiple-level analysis of gut-microbiome-immune-joint interactions in rheumatoid arthritis. BMC Genomics. 2019;20(1):p. 124. doi: 10.1186/s12864-019-5510-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chen J., Wright K., Davis J. M., et al. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Medicine. 2016;8(1):p. 43. doi: 10.1186/s13073-016-0299-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Alpizar-Rodriguez D., Lesker T. R., Gronow A., et al. Prevotella copriin individuals at risk for rheumatoid arthritis. Annals of the Rheumatic Diseases. 2019;78(5):590–593. doi: 10.1136/annrheumdis-2018-214514. [DOI] [PubMed] [Google Scholar]
- 71.Maeda Y., Takeda K. Role of gut microbiota in rheumatoid arthritis. Journal of Clinical Medicine. 2017;6(6):p. 60. doi: 10.3390/jcm6060060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pianta A., Arvikar S., Strle K., et al. Evidence of the immune relevance of Prevotella copri, a gut microbe, in patients with rheumatoid arthritis. Arthritis & Rhematology. 2017;69(5):964–975. doi: 10.1002/art.40003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kishikawa T., Maeda Y., Nii T., et al. Metagenome-wide association study of gut microbiome revealed novel aetiology of rheumatoid arthritis in the Japanese population. Annals of the Rheumatic Diseases. 2019;79(1):103–111. doi: 10.1136/annrheumdis-2019-215743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Liu X., Zou Q., Zeng B., Fang Y., Wei H. Analysis of fecal Lactobacillus community structure in patients with early rheumatoid arthritis. Current Microbiology. 2013;67(2):170–176. doi: 10.1007/s00284-013-0338-1. [DOI] [PubMed] [Google Scholar]
- 75.Chiang H. I., Li J. R., Liu C. C., et al. An Association of gut microbiota with different phenotypes in Chinese patients with rheumatoid arthritis. Journal of Clinical Medicine. 2019;8(11):p. 1770. doi: 10.3390/jcm8111770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Picchianti-Diamanti A., Panebianco C., Salemi S., et al. Analysis of gut microbiota in rheumatoid arthritis patients: disease-related dysbiosis and modifications induced by etanercept. International Journal of Molecular Sciences. 2018;19(10):p. 2938. doi: 10.3390/ijms19102938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Zhang X., Zhang D., Jia H., et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nature Medicine. 2015;21(8):895–905. doi: 10.1038/nm.3914. [DOI] [PubMed] [Google Scholar]
- 78.Sun Y., Chen Q., Lin P., et al. Characteristics of gut microbiota in patients with rheumatoid arthritis in Shanghai, China. Frontiers in Cellular and Infection Microbiology. 2019;9:p. 369. doi: 10.3389/fcimb.2019.00369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ebrahimi A., Soofizadeh B., Ebrahimi F., et al. Relationship between Helicobacter pylori cytotoxin-associated gene A protein with clinical outcomes in patients with rheumatoid arthritis. Immunology Letters. 2019;211:49–52. doi: 10.1016/j.imlet.2019.05.014. [DOI] [PubMed] [Google Scholar]
- 80.Muñiz Pedrogo D. A., Chen J., Hillmann B., et al. An increased abundance of Clostridiaceae characterizes arthritis in inflammatory bowel disease and rheumatoid arthritis: a cross-sectional study. Inflammatory Bowel Diseases. 2019;25(5):902–913. doi: 10.1093/ibd/izy318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lee J. Y., Mannaa M., Kim Y., Kim J., Kim G. T., Seo Y. S. Comparative analysis of fecal microbiota composition between rheumatoid arthritis and osteoarthritis patients. Genes. 2019;10(10):p. 748. doi: 10.3390/genes10100748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhou X., Devescovi V., Liu Y., Dent J. E., Nardini C. Host-microbiome synergistic control on sphingolipid metabolism by mechanotransduction in model arthritis. Biomolecules. 2019;9(4):p. 144. doi: 10.3390/biom9040144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Rogier R., Evans-Marin H., Manasson J., et al. Alteration of the intestinal microbiome characterizes preclinical inflammatory arthritis in mice and its modulation attenuates established arthritis. Scientific Reports. 2017;7(1):p. 15613. doi: 10.1038/s41598-017-15802-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Taneja V. Arthritis susceptibility and the gut microbiome. FEBS Letters. 2014;588(22):4244–4249. doi: 10.1016/j.febslet.2014.05.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Liu X., Zeng B., Zhang J., et al. Role of the gut microbiome in modulating arthritis progression in mice. Scientific Reports. 2016;6(1):p. 30594. doi: 10.1038/srep30594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Nemoto N., Takeda Y., Nara H., et al. Analysis of intestinal immunity and flora in a collagen-induced mouse arthritis model: differences during arthritis progression. International Immunology. 2020;32(1):49–56. doi: 10.1093/intimm/dxz058. [DOI] [PubMed] [Google Scholar]
- 87.Balakrishnan B., Luckey D., Taneja V. Autoimmunity-associated gut commensals modulate gut permeability and immunity in humanized mice. Military Medicine. 2019;184(Supplement_1):529–536. doi: 10.1093/milmed/usy309. [DOI] [PubMed] [Google Scholar]
- 88.Shan J., Peng L., Qian W., et al. Integrated serum and fecal metabolomics study of collagen-induced arthritis rats and the therapeutic effects of the Zushima tablet. Frontiers in Pharmacology. 2018;9:p. 891. doi: 10.3389/fphar.2018.00891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jubair W. K., Hendrickson J. D., Severs E. L., et al. Modulation of inflammatory arthritis in mice by gut microbiota through mucosal inflammation and autoantibody generation. Arthritis & Rhematology. 2018;70(8):1220–1233. doi: 10.1002/art.40490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Maeda Y., Kurakawa T., Umemoto E., et al. Dysbiosis contributes to arthritis development via activation of autoreactive T cells in the intestine. Arthritis & Rhematology. 2016;68(11):2646–2661. doi: 10.1002/art.39783. [DOI] [PubMed] [Google Scholar]
- 91.Yang S., Chen H., Wei B., et al. Clindamycin administration increases the incidence of collagen-induced arthritis in mice through the prolonged impact of gut immunity. Inflammation. 2018;41:1900–1911. doi: 10.1007/s10753-018-0833-4. [DOI] [PubMed] [Google Scholar]
- 92.Sato K., Takahashi N., Kato T., et al. Aggravation of collagen-induced arthritis by orally administered Porphyromonas gingivalis through modulation of the gut microbiota and gut immune system. Scientific Reports. 2017;7(1):p. 6955. doi: 10.1038/s41598-017-07196-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hablot J., Peyrin-Biroulet L., Kokten T., et al. Experimental colitis delays and reduces the severity of collagen-induced arthritis in mice. PLoS One. 2017;12(9, article e0184624) doi: 10.1371/journal.pone.0184624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ben-Amram H., Bashi T., Werbner N., et al. Tuftsin-phosphorylcholine maintains normal gut microbiota in collagen induced arthritic mice. Frontiers in Microbiology. 2017;8:p. 1222. doi: 10.3389/fmicb.2017.01222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Dorozynska I., Majewska-Szczepanik M., Marcinska K., Szczepanik M. Partial depletion of natural gut flora by antibiotic aggravates collagen induced arthritis (CIA) in mice. Pharmacological Reports. 2014;66(2):250–255. doi: 10.1016/j.pharep.2013.09.007. [DOI] [PubMed] [Google Scholar]
- 96.Wing K., Klocke K., Samuelsson A., Holmdahl R. Germ-free mice deficient of reactive oxygen species have increased arthritis susceptibility. European Journal of Immunology. 2015;45(5):1348–1353. doi: 10.1002/eji.201445020. [DOI] [PubMed] [Google Scholar]
- 97.Rodrigues G. S. P., Cayres L. C. F., Goncalves F. P., et al. Detection of increased relative expression units of Bacteroides and Prevotella, and decreased Clostridium leptum in stool samples from Brazilian rheumatoid arthritis patients: a pilot study. Microorganisms. 2019;7(10):p. 413. doi: 10.3390/microorganisms7100413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Vaahtovuo J., Munukka E., Korkeamaki M., Luukkainen R., Toivanen P. Fecal microbiota in early rheumatoid arthritis. The Journal of Rheumatology. 2008;35(8):1500–1505. [PubMed] [Google Scholar]
- 99.James K. R., Gomes T., Elmentaite R., et al. Distinct microbial and immune niches of the human colon. Nature Immunology. 2020;21(3):343–353. doi: 10.1038/s41590-020-0602-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Donaldson G. P., Lee S. M., Mazmanian S. K. Gut biogeography of the bacterial microbiota. Nature Reviews. Microbiology. 2016;14(1):20–32. doi: 10.1038/nrmicro3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Gong J., Forster R. J., Yu H., et al. Molecular analysis of bacterial populations in the ileum of broiler chickens and comparison with bacteria in the cecum. FEMS Microbiology Ecology. 2002;41(3):171–179. doi: 10.1111/j.1574-6941.2002.tb00978.x. [DOI] [PubMed] [Google Scholar]
- 102.Yu M., Mu C., Zhang C., Yang Y., Su Y., Zhu W. Marked response in microbial community and metabolism in the ileum and cecum of suckling piglets after early antibiotics exposure. Frontiers in Microbiology. 2018;9:p. 1166. doi: 10.3389/fmicb.2018.01166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Doonan J., Tarafdar A., Pineda M. A., et al. The parasitic worm product ES-62 normalises the gut microbiota bone marrow axis in inflammatory arthritis. Nature Communications. 2019;10(1):p. 1554. doi: 10.1038/s41467-019-09361-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Mann E. R., Bernardo D., English N. R., et al. Compartment-specific immunity in the human gut: properties and functions of dendritic cells in the colon versus the ileum. Gut. 2016;65(2):256–270. doi: 10.1136/gutjnl-2014-307916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Sefik E., Geva-Zatorsky N., Oh S., et al. Individual intestinal symbionts induce a distinct population of ROR + regulatory T cells. Science. 2015;349(6251):993–997. doi: 10.1126/science.aaa9420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Sujino T., London M., van Konijnenburg D. P. H., et al. Tissue adaptation of regulatory and intraepithelial CD4(+) T cells controls gut inflammation. Science. 2016;352(6293):1581–1586. doi: 10.1126/science.aaf3892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Keita A. V., Gullberg E., Ericson A. C., et al. Characterization of antigen and bacterial transport in the follicle-associated epithelium of human ileum. Laboratory Investigation. 2006;86(5):504–516. doi: 10.1038/labinvest.3700397. [DOI] [PubMed] [Google Scholar]
- 108.Bennike T. B., Ellingsen T., Glerup H., et al. Proteome analysis of rheumatoid arthritis gut mucosa. Journal of Proteome Research. 2016;16(1):346–354. doi: 10.1021/acs.jproteome.6b00598. [DOI] [PubMed] [Google Scholar]
- 109.Yue M., Tao Y., Fang Y., et al. The gut microbiota modulator berberine ameliorates collagen-induced arthritis in rats by facilitating the generation of butyrate and adjusting the intestinal hypoxia and nitrate supply. The FASEB Journal. 2019;33(11):12311–12323. doi: 10.1096/fj.201900425RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Xiao M., Fu X., Ni Y., et al. Protective effects of Paederia scandens extract on rheumatoid arthritis mouse model by modulating gut microbiota. Journal of Ethnopharmacology. 2018;226:97–104. doi: 10.1016/j.jep.2018.08.012. [DOI] [PubMed] [Google Scholar]
- 111.Peng J., Lu X., Xie K., et al. Dynamic alterations in the gut microbiota of collagen-induced arthritis rats following the prolonged administration of total glucosides of paeony. Frontiers in Cellular and Infection Microbiology. 2019;9:p. 204. doi: 10.3389/fcimb.2019.00204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Mizuno M., Noto D., Kaga N., Chiba A., Miyake S. The dual role of short fatty acid chains in the pathogenesis of autoimmune disease models. PLoS One. 2017;12(2, article e0173032) doi: 10.1371/journal.pone.0173032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Guo L. X., Wang H. Y., Liu X. D., et al. Saponins from Clematis mandshurica Rupr. regulates gut microbiota and its metabolites during alleviation of collagen-induced arthritis in rats. Pharmacological Research. 2019;149:p. 104459. doi: 10.1016/j.phrs.2019.104459. [DOI] [PubMed] [Google Scholar]
- 114.Huang Y., Li M., Zhou L., et al. Effects of Qingluo Tongbi decoction on gut flora of rats with adjuvant-induced arthritis and the underlying mechanism. Evidence-based Complementary and Alternative Medicine. 2019;2019:11. doi: 10.1155/2019/6308021.6308021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Aa L. X., Fei F., Qi Q., et al. Rebalancing of the gut flora and microbial metabolism is responsible for the anti-arthritis effect of kaempferol. Acta Pharmacologica Sinica. 2020;41(1):73–81. doi: 10.1038/s41401-019-0279-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Pan H., Guo R., Ju Y., et al. A single bacterium restores the microbiome dysbiosis to protect bones from destruction in a rat model of rheumatoid arthritis. Microbiome. 2019;7(1):p. 107. doi: 10.1186/s40168-019-0719-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Amdekar S., Singh V., Singh R., Sharma P., Keshav P., Kumar A. Lactobacillus casei reduces the inflammatory joint damage associated with collagen-induced arthritis (CIA) by reducing the pro-inflammatory cytokines: Lactobacillus casei: COX-2 inhibitor. Journal of Clinical Immunology. 2011;31(2):147–154. doi: 10.1007/s10875-010-9457-7. [DOI] [PubMed] [Google Scholar]
- 118.Liu X., Zhang J., Zou Q., et al. Lactobacillus salivarius isolated from patients with rheumatoid arthritis suppresses collagen-induced arthritis and increases Treg frequency in mice. Journal of Interferon & Cytokine Research. 2016;36(12):706–712. doi: 10.1089/jir.2016.0057. [DOI] [PubMed] [Google Scholar]
- 119.Esvaran M., Conway P. Lactobacillus fermentum PC1 has the capacity to attenuate joint inflammation in collagen-induced arthritis in DBA/1 mice. Nutrients. 2019;11(4):p. 785. doi: 10.3390/nu11040785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Yamashita M., Matsumoto K., Endo T., et al. Preventive effect of Lactobacillus helveticus SBT2171 on collagen-induced arthritis in mice. Frontiers in Microbiology. 2017;8:p. 1159. doi: 10.3389/fmicb.2017.01159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kano H., Mogami O., Uchida M. Oral administration of milk fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 to DBA/1 mice inhibits secretion of proinflammatory cytokines. Cytotechnology. 2002;40(1/3):67–73. doi: 10.1023/A:1023966121989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Kano H., Kaneko T., Kaminogawa S. Oral intake of Lactobacillus delbrueckii subsp. bulgaricus OLL1073R-1 prevents collagen-induced arthritis in mice. Journal of Food Protection. 2002;65(1):153–160. doi: 10.4315/0362-028X-65.1.153. [DOI] [PubMed] [Google Scholar]
- 123.Amdekar S., Singh V. Lactobacillus acidophilus maintained oxidative stress from reproductive organs in collagen-induced arthritic rats. Journal of Human Reproductive Sciences. 2016;9(1):41–46. doi: 10.4103/0974-1208.178638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.So J. S., Kwon H. K., Lee C. G., et al. Lactobacillus casei suppresses experimental arthritis by down-regulating T helper 1 effector functions. Molecular Immunology. 2008;45(9):2690–2699. doi: 10.1016/j.molimm.2007.12.010. [DOI] [PubMed] [Google Scholar]
- 125.Nowak B., Ciszek-Lenda M., Srottek M., et al. Lactobacillus rhamnosus exopolysaccharide ameliorates arthritis induced by the systemic injection of collagen and lipopolysaccharide in DBA/1 mice. Archivum Immunologiae et Therapiae Experimentalis (Warsz) 2012;60(3):211–220. doi: 10.1007/s00005-012-0170-5. [DOI] [PubMed] [Google Scholar]
- 126.Hager J., Bang H., Hagen M., et al. The role of dietary fiber in rheumatoid arthritis patients: a feasibility study. Nutrients. 2019;11(10):p. 2392. doi: 10.3390/nu11102392. [DOI] [PMC free article] [PubMed] [Google Scholar]