

Abstract
La rage est encore aujourd’hui responsable d’environ 60 000 décès humains par an, principalement dans les populations démunies en Afrique et en Asie. Pourtant, depuis le développement du premier vaccin par Louis Pasteur, il y a 130 ans, les mesures prophylactiques se sont considérablement améliorées et allégées. Elles sont désormais composées du vaccin associé à des immunoglobulines antirabiques d’origine équine ou humaine purifiées. Mais, d’une manière générale, les protocoles de prophylaxie post-exposition sont longs et coûteux et les immunoglobulines, qui servent à la sérothérapie associée, sont chères et peu accessibles dans les pays en voie de développement. Enfin, la rage chez l’Homme reste incurable lorsque les symptômes apparaissent, voire même lorsque les patients sont pris en charge trop tardivement pendant la période d’incubation. Dans ces deux cas, des approches, qui laissent espérer un changement de paradigme au bénéfice des populations exposées, ont été développées. Enfin, les campagnes de vaccination antirabique de masse des chiens, qui constituent la mesure de prévention de la rage chez l’Homme la plus efficace et la moins coûteuse, se heurtent à des difficultés d’implémentation et présentent, parfois, une efficacité modérée. À ce titre, l’identification et l’analyse des moteurs épidémiologiques, conditionnant la circulation du virus dans les populations canines, permettent de mieux comprendre les points clés du contrôle qui doivent être associés à ces campagnes.
Mots clés: Rage, Prévention, Prophylaxie, Immunoglobulines
Abstract
Rabies still causes about 60,000 human deaths per year, mainly in poor populations in Africa and Asia. However, since Louis Pasteur developed the first vaccine 130 years ago, prophylactic measures have been considerably improved and simplified. They now consist of the vaccine combined with purified rabies immunoglobulins of equine or human origin. In general, however, post-exposure prophylaxis protocols are long and expensive. Furthermore, the immunoglobulins used for associated serotherapy are costly and not widely available in developing countries. Approaches have been developed to deal with these two issues that offer hope for a paradigm shift for the benefit of exposed populations. Finally, mass rabies vaccination in dogs, which are the most cost-effective measure for preventing rabies in humans, are difficult to implement and sometimes have moderate effectiveness. The identification and analysis of the epidemiological drivers conditioning the circulation of the virus in dog populations allow a better understanding of the key control points that need to be associated with these campaigns for a better efficacy.
Keywords: Rabies, Prevention, Prophylaxis, Immunoglobulines
Introduction et rappels sur la rage
La rage est une encéphalite virale aiguë, progressive et incurable. Le virus de la rage est transmis par effraction de la barrière cutanée et exposition à du matériel contaminé (la salive d’un animal infecté dans la plupart des cas), projection de virus sur des muqueuses et, de manière exceptionnelle, par transplantation d’organe à partir d’un donneur infecté [1]. Lors de l’exposition au virus, la transmission et le développement de la maladie peuvent être prévenus par la prophylaxie post-exposition (PPE), qui comprend lavage et antisepsie puis l’administration de plusieurs doses de vaccin antirabique, en combinaison avec l’injection d’immunoglobulines antirabiques (RIG) dans les cas les plus graves. Environ 30 millions de personnes reçoivent une prophylaxie post-exposition (PPE) chaque année. Malgré cela, des estimations récentes confirment au moins 59 000 décès humains par an dus à la rage [2]. Ceci est en grande partie dû au prix élevé et au manque de disponibilité locale des produits couplés à la complexité des protocoles de PPE, en particulier pour les populations rurales et/ou pauvres des pays endémiques, qui sont les plus à risque et au développement de produits contrefaits [3]. Enfin, l’efficacité de la PPE diminue à mesure que le laps de temps entre l’exposition et l’administration du vaccin augmente, jusqu’à devenir complètement inefficace à partir du développement des symptômes. Plusieurs tentatives ont été effectuées pour traiter la rage symptomatique [4], [5]. En 2004, un patient infecté du Wisconsin (États-Unis) a survécu grâce à une approche thérapeutique qui a été baptisée protocole de Milwaukee [6]. Depuis lors, des modifications ont été apportées à ce protocole mais il n’a pas fait preuve de son efficacité [7], [8].
La rage est due à des virus neurotropes appartenant à la famille Rhabdoviridae, genre Lyssavirus. À ce jour, 17 espèces de lyssavirus ont été identifiées (https://talk.ictvonline.org/taxonomy/). Les chauves-souris hébergent 15 de ces 17 espèces de lyssavirus et elles sont considérées comme les hôtes ancestraux de ces virus. Les lyssavirus sont des virus à ARN simple brin, de sens négatif, dont le génome, d’une taille d’environ 12 kb, code pour cinq protéines : la nucléoprotéine (N), la phosphoprotéine (P), la protéine de matrice (M), la protéine d’enveloppe ou glycoprotéine (G) et l’ARN polymérase (L) [1]. Les anticorps neutralisants, qui constituent le corrélat le plus proche de la protection, sont essentiellement dirigés contre la glycoprotéine. Les lyssavirus se dissimulent aux réponses de l’hôte. Ceci découle de la nature multifonctionnelle de plusieurs de leurs protéines qui, hormis leurs rôles dans la réplication, l’assemblage et l’attachement viral, sont aussi impliquées dans l’évasion vis-à-vis des réponses immunitaires innées de l’hôte par la régulation de la signalisation cellulaire [9].
Cette anthropozoonose affecte de nombreuses espèces animales différentes qui agissent en tant qu’hôte primaire du virus, par exemple les chiens, les renards, les ratons laveurs, les mouffettes, les mangoustes et de très nombreuses espèces de chauves-souris hématophages, insectivores et frugivores. La rage présente aussi une épidémiologie qui évolue au cours du temps. Comme les autres virus à ARN, les lyssavirus présentent des taux de mutation élevés en raison d’un manque d’activité de correction de la protéine L lors de la réplication du génome. Ceci leur confère une capacité d’adaptation à des changements environnementaux et une propension à franchir la barrière d’espèce et à se propager chez de nouvelles espèces animales [10]. Ainsi, de nouveaux cycles épidémiologiques émergent régulièrement. Cependant, la rage du chien constitue aujourd’hui le principal problème de santé publique. Plus de 98 % des décès, dans le monde, sont liés à une origine canine du virus et l’OMS estime que 3 milliards de personnes sont exposées au risque de contracter la rage par la morsure de chiens infectés, principalement en Asie et en Afrique, où la moitié des victimes sont des enfants âgés de moins de 15 ans. Une seule espèce de lyssavirus, le virus de la rage (RABV), est présente sur la plupart des continents et circule dans un ensemble diversifié d’hôtes primaires, dont le chien [10].
Cette revue présente les avancées récentes concernant trois axes majeurs de développement de la recherche en matière de contrôle et d’élimination de la rage canine chez l’Homme [11]. La vaccination antirabique des populations canines est aujourd’hui considérée comme la mesure de choix pour le contrôle à bas coût et à long terme de la rage chez l’animal mais aussi chez l’Homme. Notre premier point concernera donc la compréhension de la dynamique d’infection chez le chien. Elle devient essentielle afin de rationaliser et d’améliorer l’efficacité des campagnes de vaccination antirabique de masse chez le chien. Il est aussi nécessaire d’apporter des solutions thérapeutiques aux populations défavorisées qui sont le plus à risque et qui n’ont pas ou peu accès à la PPE. Dans notre deuxième point, nous aborderons donc les progrès récents en matière de raccourcissement des protocoles de PPE et de développement de produits de substitutions aux produits coûteux qui sont actuellement sur le marché. Enfin, nous finirons par la recherche d’un changement de paradigme en matière de PPE et de traitement de la rage déclarée chez l’Homme.
Dynamique de propagation du virus de la rage et identification des critères épidémiologiques impliqués dans la persistance virale chez le chien
La présence à long terme de la rage dépend de la source de transmission zoonotique. Les populations de chiens domestiques restent la principale cible de prévention et d’intervention, car elles sont en cause dans près de 99 % des décès chez l’Homme. Il est donc essentiel de comprendre les facteurs qui influencent la transmission et le maintien de la rage dans les populations canines et l’efficacité des campagnes de vaccination. Ces aspects ont été longuement négligés et une approche dogmatique, visant à essayer d’atteindre un pourcentage de couverture vaccinale théorique de 70 %, a été favorisée sans succès notoire à long terme. Aujourd’hui, une analyse fine de la dynamique et de l’écologie des populations canines permet d’entrevoir les raisons de certains de ces échecs et de mieux appréhender l’organisation des campagnes de vaccination. L’apport de la modélisation et des études phylodynamiques couplées à des données épidémiologiques, obtenues grâce la technologie du système mondial de géolocalisation (GPS), améliorent notre capacité à mieux comprendre les processus qui influent sur la persistance et la diffusion des virus [12], [13].
Prévention primaire de la rage canine par la vaccination
La prévention de la rage canine par la vaccination est essentielle pour limiter ou éliminer la rage humaine dans les pays où les chiens sont la principale source d’infection [14], [15]. Ceci a été largement démontré dans les pays développés qui ont éliminé la rage humaine par le contrôle et la vaccination de leurs populations canines. Cette approche a aussi été mise en place dans de nombreux pays en développement [16], [17], et son intérêt a été largement validé par des travaux qui montrent que l’élimination de la rage canine est à la fois épidémiologiquement et opérationnellement faisable, et pourrait être réalisée dans un large éventail de contextes en Afrique et en Asie [18]. Cette approche est maintenant favorisée par l’association tripartite de l’OMS de la FAO et de l’OIE qui envisage l’objectif de l’élimination de la rage humaine transmise par le chien d’ici 2030 [19]. Cependant, ces programmes de vaccination et leur implémentation à long terme restent un défi pour la plupart des pays en développement qui souvent échouent dans l’atteinte de l’objectif d’une élimination durable, voire même d’un contrôle efficace et à large échelle. Tous les travaux récents concluent sur l’importance des études sur l’écologie et la démographie canine dans les pays où la rage canine est fortement endémique [20], [21] afin d’améliorer la mise en œuvre et l’efficacité de ces campagnes.
Analyse et modélisation phylogéographiques comme outils d’étude de la dynamique épidémiologique du RABV canin
Les travaux, visant à étudier l’écologie et la démographie canine et leurs rôles dans la persistance et la diffusion de la rage, reposent sur deux approches qui restent pour le moment très souvent distinctes (leur intégration demeurant actuellement un réel défi scientifique). La première d’entre elle vise à modéliser les données épidémiologiques récoltées et simuler le rôle de différents facteurs et d’analyser les conséquences de certaines mesures d’intervention et, en particulier, la vaccination de masse des chiens [22].
L’ensemble des travaux montrent que la capacité du virus rabique à se transmettre de chien à chien est très faible comparée à d’autres maladies infectieuses. Les estimations du nombre de reproduction de base (R0) pour la rage dans différents contextes montrent que ce taux est la plupart du temps en dessous de 2 et proche de 1, voire inférieur dans certains contextes [22]. Le taux de reproduction a même été suivi de manière détaillée au cours du temps dans certaines grandes villes par des approches épidémiologique et génomique combinées [23], [24], [25]. Il s’avère que si le virus de la rage canine semble être endémique dans ces grandes villes africaines, son épidémiologie est en fait façonnée par l’extinction régulière des chaînes de transmission locale couplée à l’introduction continue de nouveaux virus en provenance de l’extérieur, générant des vagues successives de propagation [24], [26]. Il est à noter que le nombre effectif de reproduction au cours de chaque vague était rarement supérieur à la valeur critique de 1, de sorte que la rage ne s’auto-entretient pas dans ces grandes villes.
La couverture vaccinale est un facteur clé pour le succès de la prévention et du contrôle de la rage. Considérant le faible niveau du R0 observé, le seuil de vaccination critique (immunité de troupeau), calculé à partir du R0, devrait donc être de l’ordre de 10 à 50 % de la population canine [22]. Or, le taux de couverture vaccinale efficace observé, de manière empirique, chez le chien est dans la plupart des régions d’au moins 70 % [27]. Il y a donc un hiatus entre le fait qu’il faille 70 % ou plus des chiens efficacement vaccinés, à un moment donné, pour prévenir la survenue d’épidémies et permettre une élimination locale et les observations locales et nationales concernant la capacité du virus à se transmettre et aboutissant au calcul du R0. En fait, de nombreux autres facteurs jouent un rôle dans l’entretien de l’enzootie et l’efficacité des interventions et la couverture vaccinale. Ainsi, le taux de natalité des chiens, la densité des chiens [22] et l’hétérogénéité spatiale de la couverture vaccinale [28] ont été identifiés comme des facteurs cruciaux dans le succès de ces campagnes. Compte tenu des différents niveaux d’endémicité de la rage dans chaque pays, la prise en compte de ces facteurs est donc cruciale pour la faisabilité et l’optimisation de la mise en œuvre des interventions de lutte contre la rage. Enfin, de nombreux travaux, sur les mouvements de chiens, indiquent que les réintroductions continuelles de chiens enragés par le biais de mouvements médiés par l’Homme sont à l’origine du maintien de la maladie dans un pays ou une région donnée, en dépit de niveaux de couverture vaccinale atteints qui frôlent 80 % [23], [25], [29], [30]. Les déplacements en liberté des chiens domestiques sont, en effet, limités à de très courtes distances de quelques kilomètres par jour ou moins pour les chiens enragés [31]. Les données disponibles, provenant d’enquêtes auprès des ménages, indiquent la pratique assez courante consistant à transporter des chiens avec des mouvements fréquents, même sur de grandes distances [28], [32]. Plus récemment, un modèle de méta-population spatialement explicite pour la diffusion de la rage en République Centrafricaine a montré que la fragmentation spatiale des chiens, les mouvements induits par l’Homme et les fluctuations de la période d’incubation chez les chiens sont des éléments déterminants dans la persistance de la rage dans le pays [33].
En conclusion, ces approches épidémiologiques et génomiques permettent d’élaborer un cadre global pour comprendre la persistance de la maladie et informer les mesures de lutte. Tous ces travaux suggèrent que la rage à l’échelle géographique locale est due à la dispersion du virus rabique par l’Homme dans des zones périurbaines et rurales peu reliées entre elles (structure des populations canines en forme de méta-population), par opposition à une dispersion de proche en proche dans une population canine urbaine homogène relativement importante.
Prévention secondaire : développement de nouvelles approches concernant la prophylaxie post-exposition antirabique chez l’Homme
Raccourcissement des protocoles de prophylaxie post-exposition
La première des 13 doses de vaccin antirabique administrées sur 11 jours en PPE a été injectée par Jacques-Joseph Grancher en présence de Louis Pasteur à Joseph Meister, le 6 juillet 1885 [34] (Fig. 1 ). Dès ce moment, la durée des protocoles (et le nombre de doses administrées) de vaccins développés sur tissus nerveux a varié, allant jusqu’à un mois dans des cas considérés à risque élevé de transmission du virus de la rage [35].
Figure 1.

Notes de Louis Pasteur sur le premier protocole PPE administré à Joseph Meister, décrivant les 13 injections sur une période de 11 jours (©Institut Pasteur – Musée Pasteur).
Le développement dans les années 1960 de vaccins produit sur cultures cellulaires plus immunogènes a permis, vers le milieu des années 1970, puis des années 1980, le raccourcissement des schémas vaccinaux intramusculaires – protocoles Essen [36] puis Zagreb [37], recommandés par l’OMS à partir de 1992. L’antigénicité élevée de ces vaccins a également permis d’envisager l’administration intradermique (ID) de doses de volume réduit, mise à l’épreuve au laboratoire [38] ou sur le plan de la réponse immunitaire seulement [39], [40] puis mise en pratique clinique quotidienne au centre de la Croix-Rouge thaïlandaise (TRC) de Bangkok. À la fin des années 1990, l’OMS a recommandé, pour la première fois, un protocole PPE par la voie ID avec un schéma à 5 sessions sur 3 mois extrêmement efficace et à moindre coût. Des défauts d’approvisionnement mondial en vaccin antirabique ont amené le US-CDC à revoir les recommandations vaccinales, en 2008 [36], entérinées par l’OMS en 2009.
Sur la base de l’expérience, de données expérimentales, cliniques et biologiques, l’ACIP (Advisory Committee on Immunization Practices) a ainsi recommandé de réduire le schéma vaccinal à 4 sessions, en abandonnant le rappel à 3 mois. La protection clinique conférée par une dose intramusculaire ou deux doses ID étant équivalentes [41], le protocole TRC a donc lui aussi été réduit à 4 sessions ID sur un mois (j0, j3, j7, j28).
Révision des directives de gestion de l’OMS
Réouvert en 1995, le centre de prévention de la rage de l’Institut Pasteur du Cambodge (IPC) est un des plus actifs au Monde, avec plus de 20 000 patients ayant reçu le protocole ID développé par la TRC en 2010.
En avril 2011 est malheureusement survenu le décès par rage d’un jeune Cambodgien, attaqué 21 jours plus tôt par un chien dans un quartier de Phnom Penh. Ce petit garçon, sévèrement mordu avec décollement du scalp, avait dû être suturé avant de recevoir la PPE par voie ID à l’IPC le lendemain. Les résultats positifs de l’examen virologique de la tête du chien mordeur à l’IPC avaient conduit à l’infiltration un jour plus tard d’immunoglobulines antirabiques dans le pourtour de la plaie. Ce décès, survenu malgré la PPE, y compris l’injection des immunoglobulines en temps considéré encore utile [42], bien qu’elles aient été administrées après que la suture ait eu lieu, est un évènement très rare.
Après vérification de la continuité de la chaîne du froid et de la qualité du lot de vaccin, la question s’est posée de ce qu’était réellement le devenir des patients venus à l’IPC et ayant reçu une PPE. Une rapide étude par contact téléphonique avec des patients mordus par des chiens confirmés enragés au cours des mois précédents a confirmé l’efficacité de la PPE. Il a alors été constaté que la survie dans cette petite étude était de 100 % chez les patients qui n’étaient pas revenus pour leur 4e session de vaccin, à 1 mois.
Grâce à un financement de l’Institut Pasteur et de son réseau, l’équipe de l’unité d’épidémiologie et de santé publique de l’IPC a mené une étude clinico-épidémiologique en contactant tous les patients PPE ayant été mordus par un chien confirmé enragé ou non testé mais d’aspect malade sur la période 2003–2014 [43]. Malgré les effectifs importants, le nombre heureusement faible de décès après PPE n’a pas conféré à cette étude la puissance souhaitée. Les résultats cliniques étaient cependant très rassurants.
Une deuxième étude, sérologique celle-là, a étudié les taux circulants d’anticorps chez des personnes consultant pour PPE à l’IPC après morsure par un chien confirmé enragé [44]. Une étude comparant le devenir de personnes après un protocole expérimental raccourci à 3 sessions versus un protocole validé de 4 sessions contre une maladie 100 % létale étant hors de question, une équipe conjointe de l’IPC et de l’Institut Pasteur de Paris a donc comparé les titres d’anticorps neutralisants immédiatement avant et deux semaines après la 4e session (j28 vs. j42). Ces titres circulants n’étant pas plus élevés, l’étude a donc montré que l’on pouvait se passer de la 4e session et réduire le schéma PPE à une semaine seulement (Fig. 2 ).
Figure 2.
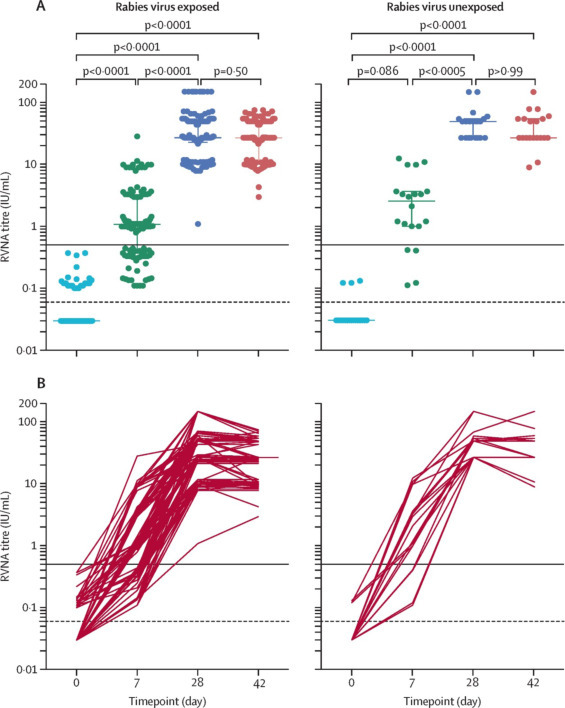
Titres d’anticorps neutralisant le virus de la rage avant et 2 semaines après la quatrième séance de prophylaxie post-exposition chez les participants exposés à la rage et non exposés à la rage. Au total, 116 participants avaient été mordus par un chien infecté par le virus rabique et 20 par un chien séronégatif. La ligne noire indique le seuil de titre en anticorps de 0,5 UI/mL considéré comme conférant une protection. La ligne en pointillés indique le seuil de détection du test (0,06 UI/mL). Les valeurs négatives ont été arbitrairement fixées à 0,03 UI/mL à des fins de visualisation. (A) titres d’anticorps neutralisants tels que déterminés par un test de réduction des foyers de fluorescence (RFFIT) ; la médiane et l’IQR sont indiqués, avec des points représentant les échantillons des patients individuels. B. Évolution des titres d’anticorps neutralisants par individu. RVNA : anticorps neutralisant le virus de la rage [44].
Ce nouveau schéma vaccinal intradermique de 3 sessions sur une semaine a été baptisé « Protocole IPC ». Ces informations ont été partagées avec l’OMS dans le cadre de la participation en 2016–2017 à un comité technique d’experts pour la révision des recommandations vaccinales PPE. L’OMS a pris note de la validation en octobre 2017 par le groupe indépendant SAGE. Émises en avril 2018, les nouvelles recommandations de l’OMS placent en première position le protocole intradermique IPC de 3 sessions sur une semaine [45]. C’est la première fois qu’un schéma de PPE sur une semaine est recommandé.
Le protocole IPC a, depuis juin 2018, été administré à plus de 60 000 personnes au Cambodge, sans décès identifié par la recherche active de cas qui est menée systématiquement auprès des personnes mordues par des chiens enragés ou non-testés mais malades. Il a été adopté officiellement dans les recommandations vaccinales au Cambodge, aux Philippines, au Bangladesh et en Côte d’Ivoire, est désormais utilisé dans certains centres vaccinaux en Inde (Bangalore, Himachal Pradesh), au Pakistan (Karachi), à Madagascar, en Tanzanie et au Mali. Son utilisation est à l’étude au Vietnam, au Tchad, au Cameroun et au Népal.
Le raccourcissement du schéma à doses réduites grâce aux travaux – menés de manière pragmatique et frugale – de ces équipes pasteuriennes à Phnom Penh et Paris permet de diminuer les coûts directs de la vaccination. Il diminue les coûts indirects de déplacement et d’hébergement de populations exposées, essentiellement rurales et pauvres, vers les centres où la PPE est disponible [46]. Il permet aussi d’économiser et de mieux répartir les doses de vaccin disponibles et ainsi d’améliorer l’équité vaccinale dans les pays endémiques, notamment grâce au soutien financier de GAVI [47].
De nouveaux vaccins pourraient changer la donne, mais plutôt dans la voie d’administration (voie cutanée, intranasale, etc.). Ceci peut influer sur l’observance, mais pas dans la décision de raccourcir ou non les schémas. La recherche clinique sur la rage doit donc se poursuivre dans les pays fortement endémiques, au Sud pour l’essentiel, afin d’informer les pratiques dans le monde. Cependant, et malgré l’excellence des vaccins antirabiques actuels, il est à craindre que nous entrions dans une « zone rouge » [35].
Il est extrêmement souhaitable de réduire encore le nombre ou le volume des doses dans la PPE. Cependant, la réduction du nombre de doses au cours de la période de quelques jours qui détermine probablement le risque d’infection et donc de décès est un exercice périlleux. Le changement souhaitable de schéma avec une réduction à 2 sessions, voire une seule, devra se baser sur des travaux d’immunologie mais aussi sur une évaluation clinico-épidémiologique extrêmement rigoureuse, le seul critère d’évaluation dans la rage étant la survie ou la mort, à moins de réussir à mesurer la réplication du virus in situ. Or, les décès restent heureusement rares, « naturellement » en raison de la transmission inconstante même par un chien confirmé enragé et grâce surtout à la PPE avec des vaccins très efficaces, associés ou non aux immunoglobulines. Par ailleurs, un essai clinique randomisé serait non-éthique. Cela impose donc se pencher sur le devenir clinique des patients n’ayant pas suivi la PPE à son terme [35].
Ce pourcentage de non-complétude va se réduire en raison d’un protocole désormais plus court. Il faudra donc disposer de très importantes bases de données patients. Ceci est tout à fait envisageable grâce, notamment, à des outils électroniques améliorés, le nécessaire renforcement de la pharmacovigilance et la mise en place d’un réseau mondial à l’heure où les instances internationales visent à éliminer la rage humaine transmise par le chien d’ici 2030.
Remplacement des anticorps polyclonaux par des anticorps monoclonaux
Les anticorps polyclonaux font partie de la préconisation de la PPE pour les victimes considérées de catégorie III, notamment ayant subi morsures ou griffures uniques ou multiples ayant traversé le derme, léchage de la peau lésée, contamination des muqueuses par de la salive après léchage, contact avec des chauves-souris [27]. Les anticorps polyclonaux, ou immunoglobuline antirabique, indiqués, sont des anticorps produits et purifiés à partir du sérum de personnes ou de chevaux immunisés contre la rage (human rabies immunoglobulin [HRIG], equine rabies immunoglobulin [ERIG]) [45].
Même si ces immunoglobulines font toujours partie des mesures prophylactiques contre la rage recommandées par l’OMS, on estime que moins de 10 % des patients de la catégorie III les reçoivent dans le cadre de la PPE [48]. Cette faible couverture peut être expliquée par plusieurs facteurs : la production mondiale d’immunoglobuline antirabique est assez restreinte, soit par les difficultés d’obtention de donneurs humains pour la production de HRIG, soit par l’abandon du marché par des producteurs de ERIG ; le prix d’un traitement avec ces immunoglobulines est très élevé, pouvant atteindre plusieurs centaines de dollars américains ; de nombreux patients nécessitant un traitement avec ces immunoglobulines se trouvent dans des pays à faible revenu et dans des régions géographiquement isolées (régions rurales d’Asie et d’Afrique). En plus, leur durée de conservation est courte et il existe des difficultés pour assurer la qualité entre les différents lots [49].
Vu les limitations d’utilisation des HRIG et ERIG, l’OMS recommande donc aussi l’utilisation d’anticorps monoclonaux (monoclonal Antibodies, mAbs) comme alternative aux immunoglobulines antirabiques dans le cadre de la PPE [45], [48], [49]. En général, les mAbs présentent l’avantage d’être plus puissants, d’avoir un spectre d’action neutralisante plus large et d’être efficaces à des concentrations plus faibles que les anticorps polyclonaux. Cependant, vu la mono-spécificité des mAbs, l’OMS préconise l’utilisation de cocktails contenant au moins deux mAbs se liant à des épitopes non chevauchants, de façon à éviter le risque d’échec dû à différentes souches ou variantes virales [50].
Le premier mAb mis à disposition dans le marché a été le SII RMAb (aussi dénommé HuMAb 17C7 ou RAB-1), un unique anticorps monoclonal recombinant humain anti-glycoprotéine du virus de la rage [51] disponible sur le nom de Rabishield depuis 2007 [52]. Très récemment, le Contrôleur général des médicaments de l’Inde (Drug Controller General of India, DCGI) a autorisé la commercialisation du Twinrab (précédemment nommé RABIMABs), qui contient cette fois-ci deux mAbs (M777-16-3/62-71-3) [53]. Deux autres produits sont actuellement en essai clinique : SYN023 [54] et Rabies mAb [48], et d’autres cocktails de mAbs comme RVC20/RVC58 [55], [56], CL184 (CR57/CR4098) [57] et CR57/RV08/RV3A5 [58] ont aussi été décrits comme possibles remplaçants des HRIG et ERIG et sont en cours d’étude.
Malgré l’urgence médicale que constitue la fourniture de produits biologiques de qualité pour lutter contre la rage et le fait que le marché des anticorps monoclonaux destinés à la médecine humaine soit en pleine expansion, y compris dans le domaine des maladies infectieuses [59], [60], [61], peu de produits sont actuellement disponibles [62]. Selon une étude du marché des produits pour la rage, un producteur de mAbs qui est entré en activité en 2017 a déclaré une production annuelle de 2 millions de doses de mAbs, soit quatre fois plus élevée que la production estimée de HRIG/ERIG dans le monde [62]. Actuellement, seul Rabishield est disponible, en plus du Twinrab qui vient d’être autorisé, tous les deux pour une commercialisation exclusive limitée à l’Inde.
De nombreuses raisons peuvent expliquer le fait que le remplacement des HRIG et ERIG par des mAbs ne soit pas encore une réalité. Parmi celles-ci, on peut distinguer le fait que la rage soit une maladie présente particulièrement dans des pays à faibles ressources économiques du fait que le développement et la production de mAbs sont coûteux, que les phases d’essais cliniques pour cette maladie sont compliquées et nécessitent de très grands effectifs. Cependant, en vue des nombreux avantages inhérents à l’utilisation des mAbs dans la PPE, l’OMS encourage l’utilisation de mAbs comme alternative aux HRIG et ERIG selon le contexte géographique et épidémiologique, mais recommande également qu’un registre soit tenu pour surveiller l’utilisation clinique des mAbs [27].
Guérir de la rage ? Vers un changement de paradigme dans le traitement chez l’Homme
La rage est une encéphalomyélite aiguë considérée fatale dans plus de 99 % des cas après l’apparition des signes cliniques, ce qui est corrélé à l’arrivée du virus dans le système nerveux central [7], [63]. Actuellement, la seule approche thérapeutique décrite reste le protocole Milwaukee. Après plusieurs modifications depuis sa première utilisation en 2004, aujourd’hui il comprend, entre autres, l’induction d’un coma artificiel et l’injection de kétamine et d’amantadine (deux antagonistes compétitifs des récepteurs NMDA qui empêcheraient la libération des particules virales) [1], [8], [64]. Néanmoins, son efficacité est discutable, car au moins 31 échecs documentés ont été signalés [7], [8]. Dans ce contexte, la recherche pour un traitement efficace après l’apparition des signes cliniques reste d’extrême urgence.
Malgré de nombreuses études visant la prévention de la rage et l’amélioration de la PPE, les efforts déployés pour la recherche d’un traitement contre la maladie établie sont moins nombreux. Idéalement, les antiviraux contre la rage devraient bloquer la réplication virale et éliminer le virus présent chez des patients ayant déjà montré des signes cliniques neurologiques [64], [65], [66].
À ce jour, plusieurs molécules antivirales ont été testées contre la rage, et malgré une action antivirale efficace in vitro, les tests in vivo chez la souris ou le hamster syrien n’ont pas été fructueux. C’est le cas notamment pour la ribavirine, un analogue de la guanosine à large spectre antiviral [67], pour des composés dérivés de la ribavirine (EICAR, EICNR) [68], pour le favipiravir, un inhibiteur de virus à ARN à large spectre [69], pour la pyriméthamine, une molécule utilisée dans le traitement de maladies à protozoaires comme la toxoplasmose ou le paludisme [70], ou la pour la ranpirnase, une ribonucléase utilisée comme agent antitumoral [71].
Récemment, des études proposant des associations larges et complexes entre antiviraux (ribavirine et favipiravir), inhibiteurs de la réponse pro-inflammatoire de l’hôte (inhibiteurs de TNF-α, de CASP-1 et de MAPK), HRIG, interférons α/β et mannitol (agent promoteur de l’ouverture de la barrière hématoencéphalique) ont été proposés chez la souris [72], [73]. Certaines combinaisons ont été capables de prolonger la survie de souris infectées mais de façon limitée, et seulement dans le cadre d’une administration précoce après l’infection. Même si ces antiviraux à large spectre, seuls ou en association, présentent une certaine action contre le RABV, des antiviraux spécifiques ciblant le complexe de réplication du virus de la rage semblent toutefois être la piste la plus prometteuse [64].
D’autres approches incluent des peptides analogues au nAChRα1 (sous-unité alpha 1 du récepteur nicotinique d’acétylcholine) [74] ou même des extraits de plantes comme Datura metel (famille des Solanacées) [75]. De plus, malgré les risques sur l’atténuation incomplète ou le retour à la virulence, l’utilisation de vaccins à virus vivant atténué reste aussi une possibilité [5].
Une toute autre approche thérapeutique est l’immunothérapie anti-rage. Certains anticorps monoclonaux ont été développés et proposés comme alternatives pour la PPE [51], [54], [55], [57], [58]. Une étude utilisant le cocktail d’anticorps monoclonaux humains RVC20/RVC58, récemment publiée, montre que l’administration périphérique et intracérébroventriculaire de ces anticorps, même à des phases tardives de l’infection, a été capable de guérir des souris déjà présentant des signes neurologiques de la rage [76], ce qui pourrait être la prémisse d’un nouveau paradigme dans le traitement de la rage chez l’Homme.
Le traitement contre la rage humaine demeure un défi, car les molécules proposées (antiviraux ou anticorps) doivent franchir la barrière hématoencéphalique pour être actives sur le virus déjà présent dans le système nerveux central et – évidemment – ne pas présenter de toxicité et d’induire d’effets secondaires. Le contexte socio-épidémiologique de cette maladie montre que les victimes sont souvent très jeunes et très pauvres. Au vu de sa gravité clinique, le développement d’un traitement curatif efficace contre la rage est donc nécessaire de toute urgence.
Déclaration de liens d’intérêts
Les auteurs déclarent ne pas avoir de liens d’intérêts.
Footnotes
Étant donné le contexte sanitaire épidémique lié à la COVID-19 en 2020, la présentation de cette communication en séance à l’Académie a été reportée.
Références
- 1.Fooks A.R., Cliquet F., Finke S., Freuling C., Hemachudha T., Mani R.S. Rabies. Nat Rev Dis Primer. 2017;3(1):1–19. doi: 10.1038/nrdp.2017.91. [DOI] [PubMed] [Google Scholar]
- 2.Hampson K., Coudeville L., Lembo T., Sambo M., Kieffer A., Attlan M. Estimating the global burden of endemic canine rabies. PLoS Negl Trop Dis. 2015;9(4):e0003709. doi: 10.1371/journal.pntd.0003709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Taylor E., Banyard A.C., Bourhy H., Cliquet F., Ertl H., Fehlner-Gardiner C. Avoiding preventable deaths: the scourge of counterfeit rabies vaccines. Vaccine. 2019;37(17):2285–2287. doi: 10.1016/j.vaccine.2019.03.037. [DOI] [PubMed] [Google Scholar]
- 4.Dacheux L., Delmas O., Bourhy H. Human rabies encephalitis prevention and treatment: progress since pasteurs discovery. Infect Disord Drug Targets. 2011;11:251–299. doi: 10.2174/187152611795768079. [Cité 3 mai 2020. Disponible sur : http://www.eurekaselect.com/87337/article] [DOI] [PubMed] [Google Scholar]
- 5.Smith S.P., Wu G., Fooks A.R., Ma J., Banyard A.C. Trying to treat the untreatable: experimental approaches to clear rabies virus infection from the CNS. J Gen Virol. 2019;100(8):1171–1186. doi: 10.1099/jgv.0.001269. [DOI] [PubMed] [Google Scholar]
- 6.Willoughby R.E.J., Tieves K.S., Hoffman G.M., Ghanayem N.S., Amlie-Lefond C.M., Schwabe M.J. Survival after treatment of rabies with induction of coma. N Engl J Med. 2005;352:2508–2514. doi: 10.1056/NEJMoa050382. [DOI] [PubMed] [Google Scholar]
- 7.Jackson A.C. Rabies: a medical perspective. Rev Sci Tech. 2018;37(2):569. doi: 10.20506/rst.37.2.2825. [DOI] [PubMed] [Google Scholar]
- 8.Zeiler F.A., Jackson A.C. Critical appraisal of the milwaukee protocol for rabies: this failed approach should be abandoned. Can J Neurol Sci. 2016;43(1):44–51. doi: 10.1017/cjn.2015.331. [DOI] [PubMed] [Google Scholar]
- 9.Hossain MdA, Larrous F., Rawlinson S.M., Zhan J., Sethi A., Ibrahim Y. Structural elucidation of viral antagonism of innate immunity at the STAT1 interface. Cell Rep. 2019;29(7):1934e8–1945e8. doi: 10.1016/j.celrep.2019.10.020. [DOI] [PubMed] [Google Scholar]
- 10.Troupin C., Dacheux L., Tanguy M., Sabeta C., Blanc H., Bouchier C. Large-scale phylogenomic analysis reveals the complex evolutionary history of rabies virus in multiple carnivore hosts. PLOS Pathog. 2016;12(12):e1006041. doi: 10.1371/journal.ppat.1006041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bourhy H., Dautry-Varsat A., Hotez P.J., Salomon J. Rabies, still neglected after 125 years of vaccination. PLoS Negl Trop Dis. 2010;4(11):e839. doi: 10.1371/journal.pntd.0000839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Baele G., Suchard M.A., Rambaut A., Lemey P. Emerging concepts of data integration in pathogen phylodynamics. Syst Biol. 2017;66(1):e47–e65. doi: 10.1093/sysbio/syw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Brunker K., Nadin-Davis S., Biek R. Genomic sequencing, evolution and molecular epidemiology of rabies virus. Rev Sci Tech. 2018;37(2):401. doi: 10.20506/rst.37.2.2810. [DOI] [PubMed] [Google Scholar]
- 14.Velasco-Villa A., Escobar L.E., Sanchez A., Shi M., Streicker D.G., Gallardo-Romero N.F. Successful strategies implemented towards the elimination of canine rabies in the Western Hemisphere. Antiviral Res. 2017;143:1–12. doi: 10.1016/j.antiviral.2017.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Banyard A.C., McElhinney L.M., Johnson N., Fooks A.R. Introduction history of rabies control by vaccination. Rev Sci Tech Int. 2018;37(2):305–322. doi: 10.20506/rst.37.2.2804. [DOI] [PubMed] [Google Scholar]
- 16.Lembo T., Hampson K., Kaare M.T., Ernest E., Knobel D., Kazwala R.R. The feasibility of canine rabies elimination in Africa: dispelling doubts with data. PLoS Negl Trop Dis. 2010;4(2):e626. doi: 10.1371/journal.pntd.0000626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lavan R.P., King AIMacG, Sutton D.J., Tunceli K. Rationale and support for a One Health program for canine vaccination as the most cost-effective means of controlling zoonotic rabies in endemic settings. Vaccine. 2017;35(13):1668–1674. doi: 10.1016/j.vaccine.2017.02.014. [DOI] [PubMed] [Google Scholar]
- 18.Cleaveland S., Thumbi S.M., Sambo M., Lugelo A., Lushasi K., Hampson K. Proof of concept of mass dog vaccination for thecontrol and elimination of canine rabies. Rev Sci Tech Int. 2018;37(2):559–568. doi: 10.20506/rst.37.2.2824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Abela-Ridder B., Balogh de K., Kessels J.A., Dieuzy-Labaye I., Torres G. Global rabies control: the role of international organisations and the Global Strategic Plan to eliminate dog-mediated human rabies. Rev Sci Tech Int. 2018;37(2):741–749. doi: 10.20506/rst.37.2.2837. [DOI] [PubMed] [Google Scholar]
- 20.Morters M.K., McKinley T.J., Restif O., Conlan A.J.K., Cleaveland S., Hampson K. The demography of free-roaming dog populations and applications to disease and population control. J Appl Ecol. 2014;51(4):1096–1106. doi: 10.1111/1365-2664.12279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chidumayo N.N. System dynamics modelling approach to explore the effect of dog demography on rabies vaccination coverage in Africa. PLoS ONE. 2018;13(10):e0205884. doi: 10.1371/journal.pone.0205884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rattanavipapong W., Thavorncharoensap M., Youngkong S., Genuino A.J., Anothaisintawee T., Chaikledkaew U. The impact of transmission dynamics of rabies control: systematic review. Vaccine. 2019;37:A154–A165. doi: 10.1016/j.vaccine.2018.11.035. [DOI] [PubMed] [Google Scholar]
- 23.Talbi C., Lemey P., Suchard M.A., Abdelatif E., Elharrak M., Jalal N. Phylodynamics and human-mediated dispersal of a zoonotic virus. PLOS Pathog. 2010;6(10):e1001166. doi: 10.1371/journal.ppat.1001166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bourhy H., Nakouné E., Hall M., Nouvellet P., Lepelletier A., Talbi C. Revealing the micro-scale signature of endemic zoonotic disease transmission in an African urban setting. PLOS Pathog. 2016;12(4):e1005525. doi: 10.1371/journal.ppat.1005525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zinsstag J., Lechenne M., Laager M., Mindekem R., Naïssengar S., Oussiguéré A. Vaccination of dogs in an African city interrupts rabies transmission and reduces human exposure. Sci Transl Med. 2017;9(421) doi: 10.1126/scitranslmed.aaf6984. [DOI] [PubMed] [Google Scholar]
- 26.Cori A., Nouvellet P., Garske T., Bourhy H., Nakouné E., Jombart T. A graph-based evidence synthesis approach to detecting outbreak clusters: an application to dog rabies. PLOS Comput Biol. 2018;14(12):e1006554. doi: 10.1371/journal.pcbi.1006554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.World Health Organization . WHO Expert Consultation on Rabies. Second Report. WHO; Geneva: 2018. WHO Technical Expert Series. [PubMed] [Google Scholar]
- 28.Ferguson E.A., Hampson K., Cleaveland S., Consunji R., Deray R., Friar J. Heterogeneity in the spread and control of infectious disease: consequences for the elimination of canine rabies. Sci Rep. 2015;5:18232. doi: 10.1038/srep18232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dellicour S., Rose R., Faria N.R., Vieira L.F.P., Bourhy H., Gilbert M. Using viral gene sequences to compare and explain the heterogeneous spatial dynamics of virus epidemics. Mol Biol Evol. 2017;34(10):2563–2571. doi: 10.1093/molbev/msx176. [DOI] [PubMed] [Google Scholar]
- 30.Dellicour S., Troupin C., Jahanbakhsh F., Salama A., Massoudi S., Moghaddam M.K. Using phylogeographic approaches to analyse the dispersal history, velocity and direction of viral lineages – Application to rabies virus spread in Iran. Mol Ecol. 2019;28(18):4335–4350. doi: 10.1111/mec.15222. [DOI] [PubMed] [Google Scholar]
- 31.Hampson K., Dushoff J., Cleaveland S., Haydon D.T., Kaare M., Packer C. Transmission dynamics and prospects for the elimination of canine rabies. PLOS Biol. 2009;7(3):e1000053. doi: 10.1371/journal.pbio.1000053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gsell A.S., Knobel D.L., Cleaveland S., Kazwala R.R., Vounatsou P., Zinsstag J. Domestic dog demographic structure and dynamics relevant to rabies control planning in urban areas in Africa: the case of Iringa, Tanzania. BMC Vet Res. 2012;8(1):236. doi: 10.1186/1746-6148-8-236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Colombi D., Poletto C., Nakouné E., Bourhy H., Colizza V. Long-range movements coupled with heterogeneous incubation period sustain dog rabies at the national scale in Africa. PLOS Neglected Tropical Diseases. 2020;14(5):e0008317. doi: 10.1371/journal.pntd.0008317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tarantola A. Four thousand years of concepts relating to rabies in animals and humans, its prevention and its cure. Trop Med Infect Dis. 2017;2(2) doi: 10.3390/tropicalmed2020005. [Cité 3 mai 2020. Disponible sur : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6082082/] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tarantola A., Tejiokem M.C., Briggs D.J. Evaluating new rabies post-exposure prophylaxis (PEP) regimens or vaccines. Vaccine. 2019;37:A88–A93. doi: 10.1016/j.vaccine.2018.10.103. [DOI] [PubMed] [Google Scholar]
- 36.Use of a Reduced (4-Dose) Vaccine Schedule for Postexposure Prophylaxis to Prevent Human Rabies [Internet]. [cité 3 mai 2020]. Disponible sur: https://www.cdc.gov/mmwr/preview/mmwrhtml/rr5902a1.htm.
- 37.Vodopija I., Sureau P., Smerdel S., Lafon M., Baklaić Ž, Ljubičić M. Interaction of rabies vaccine with human rabies immunoglobulin and reliability of a 2-1-1 schedule application for postexposure treatment. Vaccine. 1988;6(3):283–286. doi: 10.1016/0264-410x(88)90225-3. [DOI] [PubMed] [Google Scholar]
- 38.Nicholson K.G., Cole P.J., Turner G.S., Harrison P. Immune responses of humans to a human diploid cell strain of rabies virus vaccine: lymphocyte transformation, production of virus-neutralizing antibody, and induction of interferon. J Infect Dis. 1979;140(2):176–182. doi: 10.1093/infdis/140.2.176. [DOI] [PubMed] [Google Scholar]
- 39.Warrell M.J., Warrell D.A., Nicholson K.G., Suntharasamai P., Chanthavanich P., Viravan C. Economical multiple-site intradermal immunisation with human diploid-cell-strain vaccine is effective for post-exposure rabies prophylaxis. Lancet. 1985;325(8437):1059–1062. doi: 10.1016/s0140-6736(85)92367-0. [DOI] [PubMed] [Google Scholar]
- 40.Chutivongse S., Wilde H., Supich C., Baer G.M., Fishbein D.B. Postexposure prophylaxis for rabies with antiserum and intradermal vaccination. Lancet. 1990;335(8694):896–898. doi: 10.1016/0140-6736(90)90488-q. [DOI] [PubMed] [Google Scholar]
- 41.Kessels J., Tarantola A., Salahuddin N., Blumberg L., Knopf L. Rabies post-exposure prophylaxis: a systematic review on abridged vaccination schedules and the effect of changing administration routes during a single course. Vaccine. 2019;37:A107–A117. doi: 10.1016/j.vaccine.2019.01.041. [DOI] [PubMed] [Google Scholar]
- 42.Khawplod P., Wilde H., Chomchey P., Benjavongkulchai M., Yenmuang W., Chaiyabutr N. What is an acceptable delay in rabies immune globulin administration when vaccine alone had been given previously? Vaccine. 1996;14(5):389–391. doi: 10.1016/0264-410x(95)00213-k. [DOI] [PubMed] [Google Scholar]
- 43.Tarantola A., Ly S., Chan M., In S., Peng Y., Hing C. Intradermal rabies post-exposure prophylaxis can be abridged with no measurable impact on clinical outcome in Cambodia, 2003–2014. Vaccine. 2019;37:A118–A127. doi: 10.1016/j.vaccine.2018.10.054. [DOI] [PubMed] [Google Scholar]
- 44.Cantaert T., Borand L., Kergoat L., Leng C., Ung S., In S. A 1-week intradermal dose-sparing regimen for rabies post-exposure prophylaxis (RESIST-2): an observational cohort study. Lancet Infect Dis. 2019;19(12):1355–1362. doi: 10.1016/S1473-3099(19)30311-1. [DOI] [PubMed] [Google Scholar]
- 45.World Health Organization Rabies vaccines: WHO position paper, April 2018 – Recommendations. Vaccine. 2018;36(37):5500–5503. doi: 10.1016/j.vaccine.2018.06.061. [DOI] [PubMed] [Google Scholar]
- 46.Tarantola A., Ly S., In S., Ong S., Peng Y., Heng N. Rabies vaccine and rabies immunoglobulin in cambodia: use and obstacles to use. J Travel Med. 2015;22(5):348–352. doi: 10.1111/jtm.12228. [DOI] [PubMed] [Google Scholar]
- 47.Wentworth D., Hampson K., Thumbi S.M., Mwatondo A., Wambura G., Chng N.R. A social justice perspective on access to human rabies vaccines. Vaccine. 2019;37:A3–A5. doi: 10.1016/j.vaccine.2019.01.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sparrow E., Torvaldsen S., Newall A.T., Wood J.G., Sheikh M., Kieny M.P. Recent advances in the development of monoclonal antibodies for rabies post exposure prophylaxis: a review of the current status of the clinical development pipeline. Vaccine. 2019;37:A132–A139. doi: 10.1016/j.vaccine.2018.11.004. [DOI] [PubMed] [Google Scholar]
- 49.O’Brien K.L., Nolan T. The WHO position on rabies immunization – 2018 updates. Vaccine. 2019;37:A85–A87. doi: 10.1016/j.vaccine.2018.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ilina E.N., Larina M.V., Aliev T.K., Dolgikh D.A., Kirpichnikov M.P. Recombinant monoclonal antibodies for rabies post-exposure prophylaxis. Biochem Mosc. 2018;83(1):1–12. doi: 10.1134/S0006297918010017. [DOI] [PubMed] [Google Scholar]
- 51.Gogtay N.J., Munshi R., Ashwath Narayana D.H., Mahendra B.J., Kshirsagar V., Gunale B. Comparison of a novel human rabies monoclonal antibody to human rabies immunoglobulin for postexposure prophylaxis: a phase 2/3, randomized, single-blind, noninferiority, controlled study. Clin Infect Dis. 2018;66(3):387–395. doi: 10.1093/cid/cix791. [DOI] [PubMed] [Google Scholar]
- 52.Serum Institute of India PVT. LTD. Rabishield [Internet]. [cité 1 mai 2020]. Disponible sur: https://www.seruminstitute.com/product_ind_rabishield.php#prosub-ind.
- 53.The Clinical Trials Registry India (CTRI) 2017. Randomized, multi-centric, open-label, comparator-controlled study to evaluate the efficacy and safety of RABIMABs administered in conjunction with Vaxirab N for post-exposure prophylaxis in patients following potential rabies exposure. [Report No.: CTRI/2017/07/009038] [Google Scholar]
- 54.Chao T., Zhang S., Tsao E., Rupprecht C.E. In vivo efficacy of SYN023, an anti-rabies monoclonal antibody cocktail, in post-exposure prophylaxis animal models. Trop Med Infect Dis. 2020;5(1):31. doi: 10.3390/tropicalmed5010031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.De Benedictis P., Minola A., Rota Nodari E., Aiello R., Zecchin B., Salomoni A. Development of broad-spectrum human monoclonal antibodies for rabies post-exposure prophylaxis. EMBO Mol Med. 2016;8(4):407–421. doi: 10.15252/emmm.201505986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hellert J., Buchrieser J., Larrous F., Minola A., de Melo G.D., Soriaga L. Structure of the prefusion-locking broadly neutralizing antibody RVC20 bound to the rabies virus glycoprotein. Nat Commun. 2020;11(1):1–8. doi: 10.1038/s41467-020-14398-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Franka R., Carson W.C., Ellison J.A., Taylor S.T., Smith T.G., Kuzmina N.A. In vivo efficacy of a cocktail of human monoclonal antibodies (CL184) against diverse North American bat rabies virus variants. Trop Med Infect Dis. 2017;2(3):48. doi: 10.3390/tropicalmed2030048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sun L., Liu Y., Li C., Li D., Liang M. Development of recombinant human monoclonal antibody cocktail for post-exposure rabies prophylaxis. Bing Xue Bao Chin J Virol. 2016;32(4):399–403. [PubMed] [Google Scholar]
- 59.Grilo A.L., Mantalaris A. The increasingly human and profitable monoclonal antibody market. Trends Biotechnol. 2019;37(1):9–16. doi: 10.1016/j.tibtech.2018.05.014. [DOI] [PubMed] [Google Scholar]
- 60.Wagner E.K., Maynard J.A. Engineering therapeutic antibodies to combat infectious diseases. Curr Opin Chem Eng. 2018;19:131–141. doi: 10.1016/j.coche.2018.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Desoubeaux G., Pelegrin M. Anticorps monoclonaux en infectiologie – Des nouveaux partenaires dans l’arsenal thérapeutique. Med Sci. 2019;35(12):1008–1013. doi: 10.1051/medsci/2019200. [DOI] [PubMed] [Google Scholar]
- 62.Ives A., Dieuzy-Labaye I., Abela-Ridder B. Global characteristics of the rabies biologics market in 2017. Vaccine. 2019;37:A73–A76. doi: 10.1016/j.vaccine.2018.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ugolini G., Hemachudha T. Rabies: changing prophylaxis and new insights in pathophysiology. Curr Opin Infect Dis. 2018;31(1):93–101. doi: 10.1097/QCO.0000000000000420. [DOI] [PubMed] [Google Scholar]
- 64.Du Pont V., Plemper R.K., Schnell M.J. Status of antiviral therapeutics against rabies virus and related emerging lyssaviruses. Curr Opin Virol. 2019;35:1–13. doi: 10.1016/j.coviro.2018.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Koraka P., Martina B. Antivirals for human use against rabies and prospects for their future application. Rev Sci Tech Int. 2018;37(2):673–680. doi: 10.20506/rst.37.2.2832. [DOI] [PubMed] [Google Scholar]
- 66.Jochmans D., Neyts J. The path towards effective antivirals against rabies. Vaccine. 2019;37(33):4660–4662. doi: 10.1016/j.vaccine.2017.12.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Appolinario C., Prehaud C., Allendorf S., Azevedo de Paula Antunes J., Peres M., Lafon M. Ribavirin has an in vitro antiviral effect in rabies virus infected neuronal cells but fails to provide benefit in experimental rabies in mice. J Virol Antiviral Res. 2013 doi: 10.4172/2324-8955.1000111. [DOI] [Google Scholar]
- 68.Anindita P.D., Sasaki M., Okada K., Ito N., Sugiyama M., Saito-Tarashima N. Ribavirin-related compounds exert in vitro inhibitory effects toward rabies virus. Antiviral Res. 2018;154:1–9. doi: 10.1016/j.antiviral.2018.03.011. [DOI] [PubMed] [Google Scholar]
- 69.Banyard A.C., Mansfield K.L., Wu G., Selden D., Thorne L., Birch C. Re-evaluating the effect of Favipiravir treatment on rabies virus infection. Vaccine. 2019;37(33):4686–4693. doi: 10.1016/j.vaccine.2017.10.109. [DOI] [PubMed] [Google Scholar]
- 70.Rogée S., Larrous F., Jochmans D., Ben-Khalifa Y., Neyts J., Bourhy H. Pyrimethamine inhibits rabies virus replication in vitro. Antiviral Res. 2019;161:1–9. doi: 10.1016/j.antiviral.2018.10.016. [DOI] [PubMed] [Google Scholar]
- 71.Smith T.G., Jackson F.R., Morgan C.N., Carson W.C., Martin B.E., Gallardo-Romero N. Antiviral ranpirnase TMR-001 inhibits rabies virus release and cell-to-cell infection in vitro. Viruses. 2020;12(2):177. doi: 10.3390/v12020177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Smreczak M., Orłowska A., Marzec A., Trębas P., Kycko A., Reichert M. The effect of combined drugs therapy on the course of clinical rabies infection in a murine model. Vaccine. 2019;37(33):4701–4709. doi: 10.1016/j.vaccine.2018.04.003. [DOI] [PubMed] [Google Scholar]
- 73.Marosi A., Dufkova L., Forró B., Felde O., Erdélyi K., Širmarová J. Combination therapy of rabies-infected mice with inhibitors of pro-inflammatory host response, antiviral compounds and human rabies immunoglobulin. Vaccine. 2019;37(33):4724–4735. doi: 10.1016/j.vaccine.2018.05.066. [DOI] [PubMed] [Google Scholar]
- 74.Sajjanar B., Dhusia K., Saxena S., Joshi V., Bisht D., Thakuria D. Nicotinic acetylcholine receptor alpha 1(nAChRα1) subunit peptides as potential antiviral agents against rabies virus. Int J Biol Macromol. 2017;104:180–188. doi: 10.1016/j.ijbiomac.2017.05.179. [DOI] [PubMed] [Google Scholar]
- 75.Roy S., Samant L., Ganjhu R., Mukherjee S., Chowdhary A. Assessment of in vivo antiviral potential of datura metel linn. Extracts against rabies virus. Pharmacogn Res. 2018;10(1):109. doi: 10.4103/pr.pr_121_17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.de Melo G.D., Sonthonnax F., Lepousez G., Jouvion G., Minola A., Zatta F. A combination of two human monoclonal antibodies cures symptomatic rabies. EMBO Mol Med. 2020:e12628. doi: 10.15252/emmm.202012628. [DOI] [PMC free article] [PubMed] [Google Scholar]