

Abstract
1-octanol is a valuable molecule in the chemical industry, where it is used as a plasticizer, as a precursor in the production of linear low-density polyethylene (LLDPE), and as a growth inhibitor of tobacco plant suckers. Due to the low availability of eight-carbon acyl chains in natural lipid feedstocks and the selectivity challenges in petrochemical routes to medium-chain fatty alcohols,1-octanol sells for the highest price among the fatty alcohol products. As an alternative, metabolic engineers have pursued sustainable 1-octanol production via engineered microbes. Here, we report demonstration of gram per liter titers in the model bacterium Escherichia coli via the development of a pathway composed of a thioesterase, an acyl-CoA synthetase, and an acyl-CoA reductase. In addition, the impact of deleting fermentative pathways was explored E. coli K12 MG1655 strain for production of octanoic acid, a key octanol precursor. In order to overcome metabolic flux barriers, bioprospecting experiments were performed to identify acyl-CoA synthetases with high activity towards octanoic acid and acyl-CoA reductases with high activity to produce 1-octanol from octanoyl-CoA. Titration of expression of key pathway enzymes was performed and a strain with the full pathway integrated on the chromosome was created. The final strain produced 1-octanol at 1.3 g/L titer and a >90% C8 specificity from glycerol. In addition to the metabolic engineering efforts, this work addressed some of the technical challenges that arise when quantifying 1-octanol produced from cultures grown under fully aerobic conditions where evaporation and stripping are prevalent.
Keywords: Octanol, octanoic acid, thioesterase, Escherichia coli, oleochemical, metabolic engineering
Introduction
1-octanol is an aliphatic molecule widely used in the chemical industry as a plasticizer, as a precursor to surfactants and fatty amines, as a plant sucker growth inhibitor in the tobacco industry (USDA,A.A.D. for the U.N.O.P., 2016), as a biodiesel additive (Akhtar et al., 2015), and as an intermediate to make a co-monomer (via dehydration to 1-octene) (Van Leeuwen, et al., 2011) for the production of linear low-density polyethylene (LLDPE). Octanol often sells for the highest price (~$1.5/lb) among fatty alcohols (C12 and C16 chain alcohols sell for ~$0.8/lb) (Pfleger et al., 2015), in part because octanoic acid and other eight-carbon oleochemicals are not abundant in common lipid sources (Yan and Pfleger, 2020). Instead, 1-octanol is produced alongside other linear alcohols via iterative oligomerization chemistries from smaller petrochemicals. However, , the Ziegler process produces an alcohol stream containing only ~17% octanol when fed ethylene, hydrogen, and aluminum (Falbe et al., 2011). A proportional quantity of aluminum hydroxide is also produced in the process thereby limiting its scale. Thus, the lack of a sustainable, low-cost supply motivates a search for alternative routes of production.
Biological conversion of sugars and other feedstocks to 1-octanol is potentially feasible at current sugar prices if high yields are achieved (Pfleger et al., 2015). However, unlike other products (i.e. short-chain alcohols, short-chain acids, amino acids, among others) which are advanced in their development, production of 1-octanol remains in the proof-of-concept stage (Akhtar et al., 2015; Sattayawat et al., 2020). Microbial synthesis of oleochemicals has advanced substantially in the last decade (Yan and Pfleger, 2020), yet the Ziegler process remains the dominant technology for the industrial production of medium-chain oleochemicals. Chain-length selectivity remains a challenge because highly active enzymes that selectively target C8-C12 intermediates in fatty acid biosynthesis and β-oxidation are not common in nature (Dellomonaco et al., 2011; Mehrer et al., 2018). In addition, medium-chain products are inherently more toxic to the cell than more common long-chain oleochemicals because they disrupt the structure and function of microbial lipid membranes. Addressing these issues are two of the main focus points in current efforts to engineer medium-chain oleochemical production (Sarria et al., 2017).
1-octanol can be made biologically from the reduction 1-octanal via an aldehyde reductase (Figure 1). In a cellular context, fatty aldehydes are often a by-product of reactive oxygen species and long-chain phospholipids. Fatty aldehydes have a propensity to form adducts with amino groups in proteins and DNA causing cellular damage. To counteract this, cells have evolved very active aldehyde reductases (Adh) that protect them from these adduct-forming reactions (Perez et al., 2008). Therefore, in producing 1-octanol from 1-octanal the major concern is how to synthesize the latter with high titer and chain-length specificity. As can be seen in Figure 1, 1-octanal can be made enzymatically from three metabolic precursors: octanoyl-ACP (Acyl-Carrier Protein), octanoyl-CoA (Coenzyme A), and octanoic acid. These metabolites are derived either from fatty acid biosynthesis (FAB) or via thiolase-driven pathways such as the reversal of the β-oxidation (rβOX) (Dellomonaco et al., 2011). Three different pathways have been demonstrated for the production of octanol from octanoyl-ACP. In the simplest approach, acyl-ACPs can be directly reduced using an acyl-ACP reductase (AAR) (Liu et al., 2014). In a second approach, octanoyl-ACP can be hydrolyzed to octanoic acid via a C8-ACP thioesterase (TE), followed by direct reduction with a carboxylic acid reductase (CAR) (Akhtar et al., 2015). Alternatively, after free octanoic acid is produced from octanoyl-ACP, the acyl moiety can be activated to octanoyl-CoA using an acyl-CoA synthetase (ACS) and subsequently reduced with an acyl-CoA reductase (ACR) (Youngquist et al., 2013). The enzymes, regulation, and engineering of FAB and rβOX pathways have been thoroughly reviewed elsewhere (Cronan, 2003; d’Espaux et al., 2015; Lennen and Pfleger, 2013, 2012; Marella et al., 2018; Pfleger et al., 2015; Sarria et al., 2017; Yan and Pfleger, 2020). The highest yielding routes would use AAR or ACR to directly convert intermediates in the FAB and rβOX pathways to octanal. Unfortunately, enzymes capable of selectively performing these reactions at high rates have yet to be discovered or engineered. Therefore, pools of eight carbon molecules must be created and allowed to accumulate to be converted by downstream reductive enzymes. Thioesterases, particularly those from plants, have been shown to be highly selective towards specific chain-lengths. When β-oxidation is disrupted, chain-length selective acyl-ACP thioesterases can be used to create pools of desired fatty acids or acyl-CoAs with reactivation by acyl-CoA synthetases. Terminal reductions can then be performed by substrate promiscuous CAR or ACR enzymes. In this strategy, the thioesterase dictates selectivity and the lack of other acyl-CoA metabolism enables substrates to build to levels where CAR/ACR activity is substantial.
Figure 1. Oleochemical Metabolism and Routes to 1-Octanol.
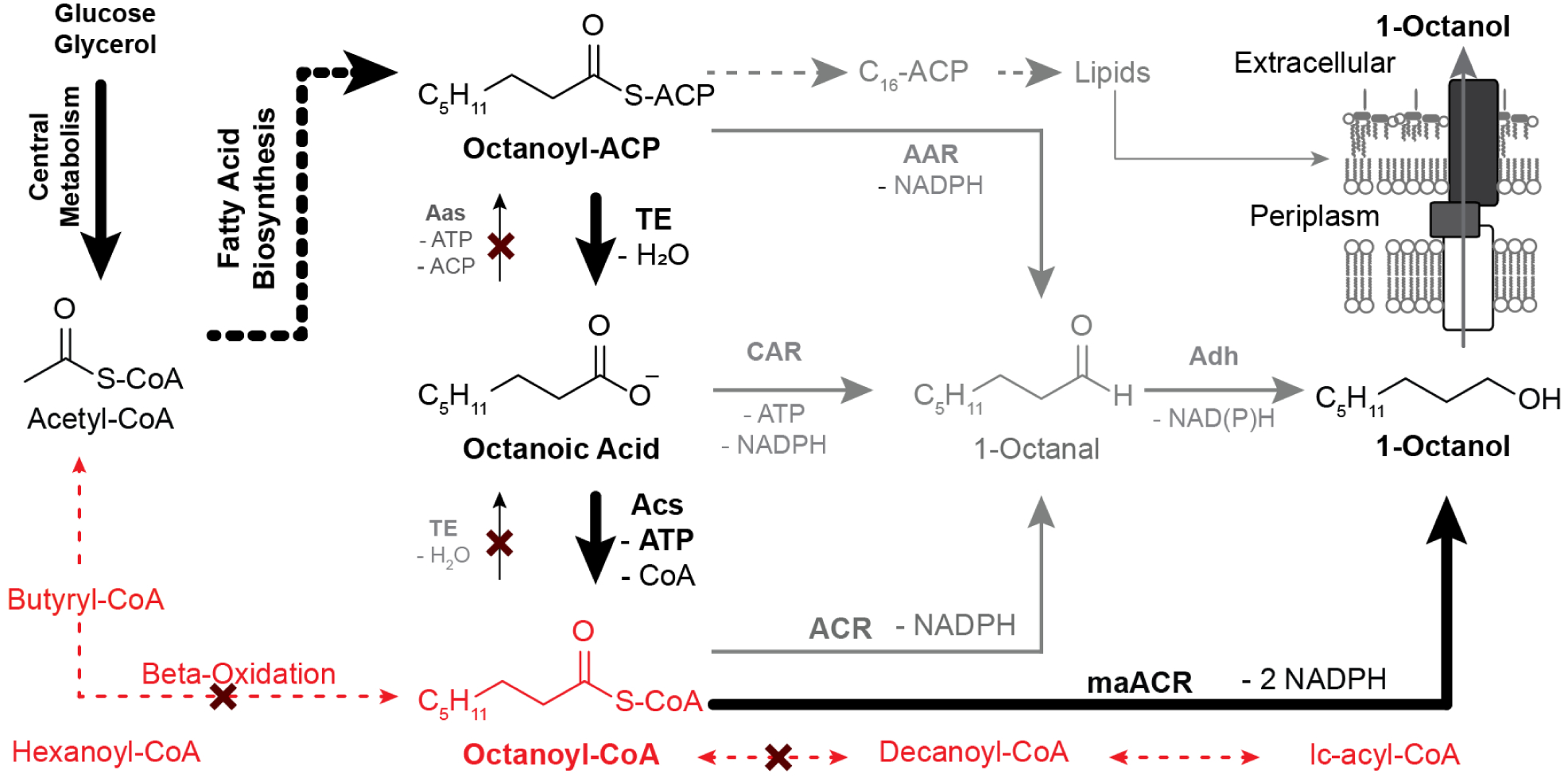
Oleochemicals are derived from acetyl-CoA that is generated in central metabolism from exogenous feedstocks (e.g. glucose, glycerol). Longer acyl-chains are synthesized via fatty acid biosynthesis (black hashed lines) or beta-oxidation (red hashed lines). The strategy applied in this project uses a heterologous thioesterase with activity towards octanoyl-ACP to divert flux from fatty acid biosynthesis to octanoic acid. Octanoic acid is then activated to octanoyl CoA by an acyl-CoA synthase (ACS). Beta oxidation is blocked such that a pool of octanoyl-CoA accumulates. Octanoyl-CoA is reduced twice by a dual-function acyl-CoA reductase (ACR) and aldehyde reductase (Adh) encoded by the Marinobacter aquaeolei MaACR. Alternative routes (in grey), including 1.) CAR mediated reduction of octanoic acid, 2.) direct reduction of octanoyl-ACP to octanal, and 3.) reduction of octanoyl-CoA to octanal by a distinct ACR were not pursued.
In previous work, we engineered an acyl-ACP thioesterase (CpFatB1) from the shrub Cuphea palustris to increase its activity while maintaining its ~90% specificity for the C8 acyl-chains (Hernandez-Lozada et al., 2018). The improved variant was isolated via a genetic selection from an error-prone library of the CpFatB1 thioesterase. The selection strain, E. coli MG1655 ΔlipB, lacked the ability to synthesize a lipoylated E2 domain of pyruvate dehydrogenase from octanoyl-ACP. The deletion forces the ΔlipB strain to use a salvage pathway, through LplA and LipA enzymes, that relies on exogenous octanoic or lipoic acids. Without these compounds present in the media, cells cannot grow in aerobic conditions (Zhao et al., 2003). Active thioesterases provided a route to produce the required octanoic acid. The best variant isolated from the selection, CpFatB1.2-M4–287, which hereafter will be referred as CpFatB1*, demonstrated a 15-fold improvement in kcat. The strong activity allowed us to express a single chromosomal copy of CpFatB1* in the fadD loci and produce octanoic acid at titers up to 1.7 g/L in rich media. This level of performance motivated development of the downstream components of the pathway to produce octanol and other 8-carbon oleochemical products. Recently, Jones and colleagues demonstrated the CAR-mediated route to produce octanol using our improved thioesterase (Sattayawat et al., 2020).
In this manuscript, we explored the variables that lead to rapid conversion of octanoic acid to octanol via the ACS/ACR route and the genetic and environmental changes that lead to high flux to octanol from sugars. We first explored fermentation variables including host strain, fermentative pathway knockouts, and growth media to maximize production of octanoic acid, the key metabolic intermediate, from sugars. Next, we tested a series of acyl-CoA synthetases (ACS) to replace E. coli FadD which has been shown to have very low activity on octanoic acid (Iram & Cronan, 2006). In addition, we characterized the pattern of evaporation of 1-octanol in a shake flask under our fermentation conditions and found that dodecane can be used to alleviate evaporation issues. We then screened acyl-CoA reductases (ACR) in order to identify variants with high catalytic activity towards octanoyl-CoA under aerobic conditions. Finally, we assembled the full pathway to demonstrate 1.3 g/L 1-octanol production from sugars in shake flasks from a strain with the 1-octanol pathway fully incorporated in the chromosome. These results represent a 2.3-fold improvement to the best result in the literature and the first demonstration of 1-octanol production at gram per liter titers in a plasmid-free strain.
Materials and Methods
Chemicals, Strains and Plasmids
Chemicals were purchased from Sigma Aldrich (St. Louis, MO) and Fisher Scientific (Waltham, MA). Oligonucleotides and gene fragments were purchased from Integrated DNA Technologies (Coralville, IA). Enzymes were purchased from New England Biolabs (Ipswich, MA). DNA purification kits were purchased from Qiagen (Venlo, Netherlands). All cultures were started from single colonies grown on LB agar isolated from freezer stocks stored in 15% glycerol. Overnight cultures of strains were grown in LB media at 37°C in a rotary shaker at 250 r.p.m. In experiments where plasmids were used, media was supplemented with the appropriate antibiotics (carbenicillin, 100 μg/mL; kanamycin, 50 μg/mL; chloramphenicol, 34 μg/mL). See Table S1 for a detailed list of strains and Table S2 for a list of plasmids used in this study. Annotated sequence maps are provided for all plasmids as supplementary files.
Chromosomal integrations
Chromosome modifications (deletions, insertions, replacements) were performed with a two plasmid system, as described previously (Mehrer et al., 2018). Briefly, a 50 mL cell culture containing the pMP11 vector was inoculated with 1 mL of dense overnight culture, grown at 30 °C for an hour, and induced with 1% (w/v) arabinose to express the λ Red genes. The culture was grown to mid-exponential phase (OD600 = 0.6) and prepared for electroporation of the double-stranded DNA repair template and the guide RNA corresponding to the chromosomal target locus. After electroporation, the transformants were recovered with SOC media, grown at 30 °C for 3 hours, and plated on antibiotic selection plates. Integrants were screened by colony PCR and verified with Sanger sequencing. The gRNA and pMP11 plasmids were subsequently cured from the successful integrant. DNA repair templates were cloned using the BW25141/pKD13 system and were designed with 500 base pairs of homology upstream and downstream of the integration site (Datsenko et al., 2000, Haldimann et al., 2001). Guide RNAs were designed with the Atum designer tool (atum.bio).
Free fatty acid experiments
For experiments designed to produce free fatty acids (Figure S1), single colonies were grown overnight in LB media supplemented with the appropriate antibiotics. For minimal media experiments (Figure S1F), 50 μL of cultures grown overnight in LB media was used to inoculate 5 mL minimal media seed cultures that were ultimately used to inoculate experimental cultures. Seed cultures from three independent colonies were used to inoculate 50 mL of media in a 250 mL shake flask at an initial OD600 = 0.05. Cells were allowed to grow at 37 °C to OD600 = 0.2 before induction with IPTG; at which point cultures were incubated with shaking at 30 °C for 24 h. IPTG concentrations used were as follows: for Figure S1A–C induction is indicated on X axis; for Figure S1D–F, 50 μM IPTG was used which corresponds to the optimal induction level for the pTRC99a-CpFatB1* plasmid (see Figure S1C); for Figures 2 and 3, 1 mM IPTG was used. Media used for Figure S1A–D was MOPS rich media supplemented with glycerol as described elsewhere (Kim et al., 2015), but with the following changes: 1.39 mM Na2HPO4, no biotin, thiamine or sodium selenite added). For Figure S1E, we replaced the glycerol from the MOPS rich media described above with glucose. For Figure S1F we used MOPS minimal media phosphate limited as described elsewhere (Youngquist et al., 2013). Experiments in which octanoic acid was supplied externally (Figures 2 and 3) were done in LB media.
Figure 2. Acyl-CoA Synthetase with activity on octanoic acid.
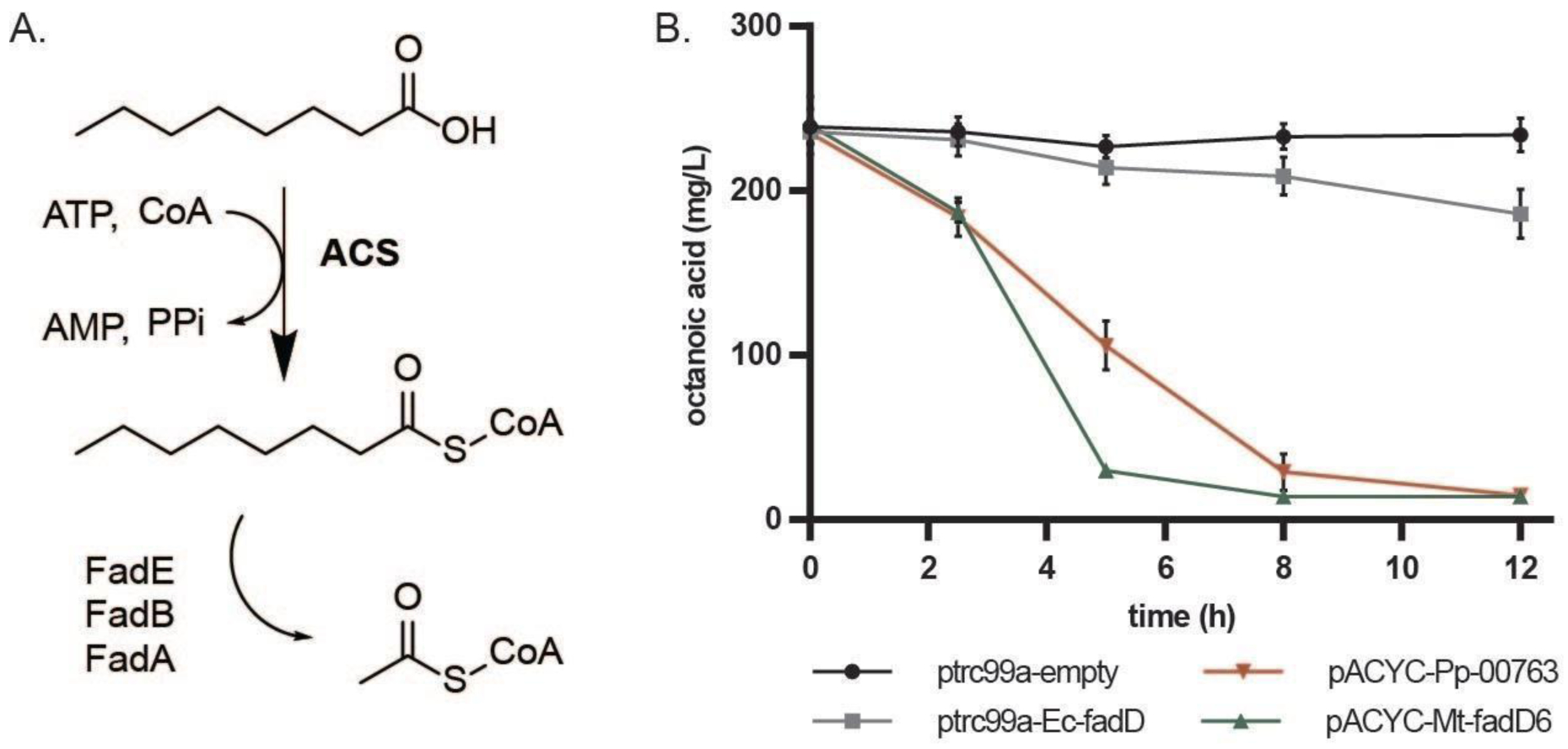
(A) E. coli MHS04 (MG1655 ΔfadR ΔfadD) cells carrying plasmids with various acyl-CoA synthases are tested for their ability to convert octanoic acid to acetyl-CoA via deregulated β-oxidation. (B) MHS04 carrying ACS plasmids were grown in LB media with 250 mg/L octanoic acid (see materials and methods), and the octanoic acid consumption was tracked over time. MtFadD6 was the best performer with most of the octanoic acid being consumed after 5 hours.
Figure 3. Production of 1-octanol from octanoic acid.
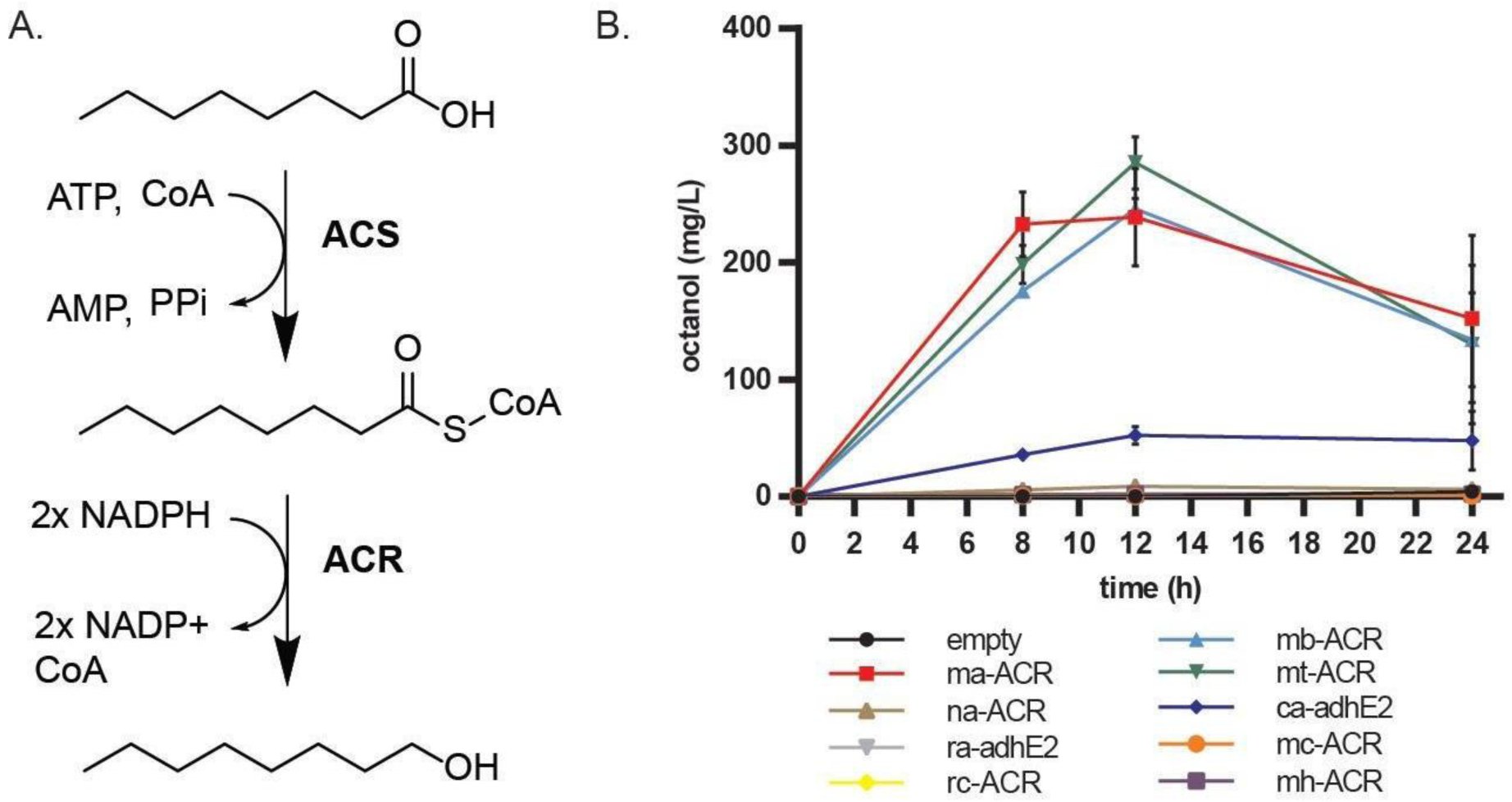
(A) E. coli DE (MG1655 ΔfadD ΔfadE) cells carrying plasmids with MtFadD6 ACS and various acyl-CoA reductases are tested for their ability to convert octanoic acid to 1-octanol. (B) DE cells carrying pACYC-MtFadD6 and different acyl-CoA reductases in a pBTRK plasmid were tested to determine an optimal ACR. Cells were grown in LB media with 500 mg/L octanoic acid (see materials and methods), and the 1-octanol production was tracked over time. MaACR, MtACR and MbACR were the best performers with similar peak production.
Fatty acid extraction, methylation and quantification
Fatty acid samples were prepared by collecting the equivalent of 2.5 mL culture in a 10 mL glass centrifuge tube, such that 2.5 mL was collected when no dodecane overlay was added to the culture, 2.75 mL when 1:10 dodecane to media ratio was used, and 3.0 mL when 1:5 dodecane to media was used. 50 μL of internal standard containing 12.5 mg/mL nonanoic acid and 1.25 mg/mL pentadecanoic acid was added to the samples. The extraction and methylation procedure was performed as described in previous work (Grisewood et al., 2017). For GC quantification, samples were analyzed using a GC-FID model Shimadzu GC-2010 equipped with an AOC-20i auto-injector and a 30 m, 0.25 mm ID RTX-5 column. The column oven temperature protocol was 100°C for 2 min, ramp to 125°C (at 7°C/min), ramp to 235°C (at 10°C/min), fast ramp to 250°C, and hold for 2 min. All error bars represent the standard error of three biological replicates.
Fatty alcohol experiments
For experiments designed to produce fatty alcohols (Figure 3B, Figure 4) single colonies were grown overnight in LB media supplemented with the appropriate antibiotics for maintaining plasmids. Overnight cultures from three single colonies were used to inoculate three 250 mL shake flasks containing 50 mL of media (for Figure 3B we used LB + 500 mg/L octanoic acid; for Figure 4 MOPS rich media glycerol described elsewhere (Kim et al., 2015) with the following changes: 1.39 mM Na2HPO4, no biotin, thiamine or sodium selenite added) at an initial OD600 = 0.05. Cells were allowed to grow at 37 °C to OD600 = 0.2 before inducing with 1mM IPTG (Figure 3B, Figure 4, last two panels ) or 50 μM IPTG (Figure 4, first three panels), 10 mL dodecane was added to control evaporation and the cultures were moved to 30 °C for up to 24 h after induction. For octanoic acid consumption experiments in the presence of ACS and ACR (Figure S4) 100 μL of a 250 mg/mL octanoic acid solution in ethanol was added immediately after induction.
Figure 4. Developing plasmid-free 1-octanol producing strain.
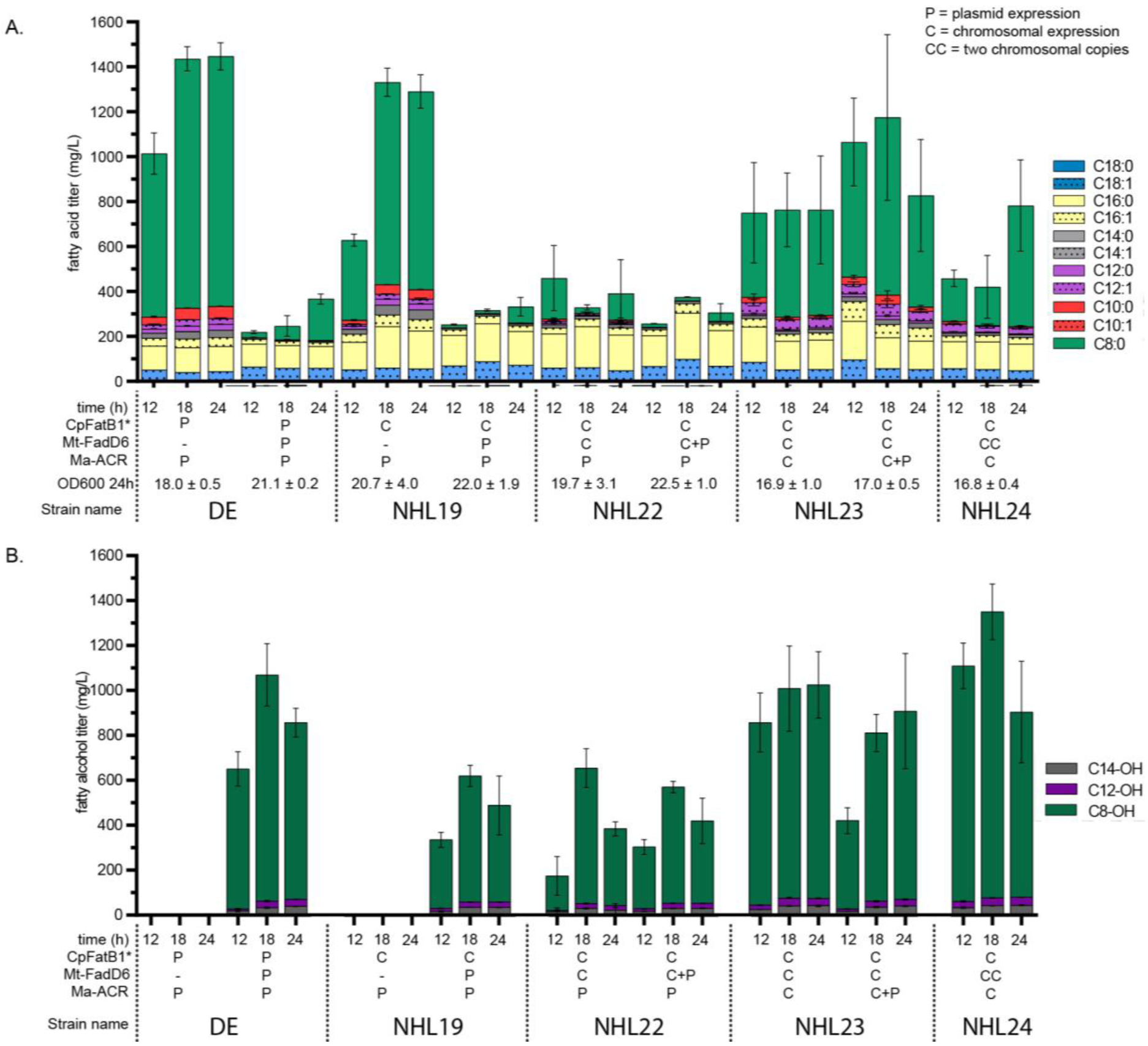
Strains containing 1-octanol pathway enzymes are first expressed exclusively from plasmids (strain DE, first panel) and then the genes were added systematically into the chromosome (strains NHL19 and NHL22; second and third panels, respectively) until reaching fully chromosomally expressed strain (NHL23, fourth panel). Finally, a second copy of Mt-FadD6 was added to the chromosome (NHL24, fifth panel) (A) indicates the fatty acid, and (B) fatty alcohol titer for strains DE (ΔfadD ΔfadE) (first panel); NHL19 (ΔfadE ΔfadD::PTRCCpFatB1*) (second panel); NHL22 (ΔfadE ΔfadD::PTRCCpFatB1* ΔackA-pta::PTRC-MtFadD6) (third panel); NHL23 (ΔfadD::PTRCCpFatB1* ΔfadE ΔackA-pta::PTRC-MtFadD6 ΔFadBA::PTRCMaACR) (fourth panel); and NHL24 (ΔfadD::PTRCCpFatB1* ΔfadE ΔackA-pta::PTRC-MtFadD6 ΔpoxB::PTRC-MtFadD6 ΔFadBA::PTRCMaACR) (fifth panel). Chromosomally expressed genes are indicated by a ‘C’ and plasmid expressed are denoted by a ‘P’. Genes being expressed both chromosomally and in plasmid are denoted as ‘C+P’. OD600 for 24 h time points are shown as well.
Fatty alcohol extraction and quantification
Before collecting fatty alcohol samples, 100 μL of an internal standard containing 125 mg/mL nonanol, and 12.5 mg/mL pentadecanol was added to the shake flasks and allowed to shake for 5 min to ensure mixing. Cultures in the presence of dodecane were constantly mixed right before the sample was taken. Fatty alcohol samples were prepared by collecting the equivalent of 2.5 mL culture (same as free fatty acid extraction section) into a 10 mL glass centrifuge tube. Samples were then extracted into 1 mL hexane by vortexing the solution for 3 min at max speed on a Vortex Genie 2 purchased from Scientific Industries (Bohemia, NY). Samples were centrifuged at 3000 x g for 10 min and the hexane layer was injected in a GC-FID. Samples were analyzed using a GC-FID model Shimadzu GC-2010 equipped with an AOC-20i auto-injector and a 30 m, 0.25 mm ID RTX-5 column. The column oven temperature protocol was 100°C for 2 min, ramp to 125°C (at 7°C/min), ramp to 235°C (at 10°C/min), fast ramp to 250°C, and hold for 2 min. All error bars represent the standard error of three biological replicates.
Results and Discussion
Optimizing Thioesterase Expression
In E. coli, long-chain acyl-ACPs are the key regulatory signal that controls flux through fatty acid biosynthesis. Therefore, we hypothesized that expression of the thioesterase CpFatB1*, which creates a competing metabolic sink and thereby depletes the abundance of long-chain acyl-ACPs, was a key step to optimize production of 1-octanol. In prior work, a MG1655 ΔfadD strain produced the most octanoic acid when expressing CpFatB1* at sub-saturating induction conditions (Hernandez-Lozada et al., 2018). Therefore, we explored thioesterase expression from the common K12 MG1655 and LS5218, an E. coli K12 strain obtained after selection on decanoate and then butyrate (Rand et al., 2017). LS5218 was a superior host for producing fatty alcohols under anaerobic conditions (Mehrer et al., 2018) and is widely used for producing poly(3-hydroxyalkanoates) (Agnew et al., 2012) under aerobic conditions.
We tested expression of CpFatB1* in these two strains using three vectors, 1.) integration of a PTRC-CpFatB1*expression cassette (strong ribosome binding site, RBS) in the fadD loci of each strain, 2.) expression of CpFatB1* with a strong RBS on low-copy pBTRK plasmids, and 3.) expression of CpFatB1* with a weak RBS on high-copy pTRC99A plasmids. Cells were inoculated from overnight cultures into 50 mL of rich glycerol media, induced at OD600 of 0.2 with 0 μM, 20 μM, 50 μM and 1000 μM IPTG and incubated with shaking at 30 °C for 24 h after induction. The titer of the plasmid-free strains both peaked at maximum induction with the MG1655 strain (1.7 g/L titer octanoic acid) outperforming the LS5218 strain (0.79 g/L) when maximally induced (Figure S1A). In prior studies, we have observed reduced titers and cell fitness as thioesterase expression was maximized (Lennen et al., 2010; Hoover et al., 2011; Youngquist et al., 2013). The lack of a maximum in octanoic acid titer suggested that further increases could be achieved with additional CpFatB1* expression. Therefore, we tested CpFatB1* expression from plasmids. When low copy plasmids were used but the RBS strength was held constant (Figure S1B), the titer from MG1655 cultures peaked at 1.5 g/L octanoic acid when uninduced while the titer of LS5218 cultures peaked when induced at 50 μM IPTG with 0.94 g/L. Finally, we lowered the RBS strength and moved the genes to a high copy plasmid, the MG1655 titer peaked when induced with 50 μM IPTG with 1.5 g/L whereas LS5218 titer peaked at 1 mM IPTG with 1.1 g/L (Figure S1C). These trends suggested that leaky expression from the low-copy plasmid was sufficient to produce the maximum amount of octanoic acid in MG1655 and further expression led to diminishing titers, analogous to prior studies. The preceding data shows that for all expression systems E. coli MG1655 outperformed LS5218 in terms of octanoic acid titer. Therefore, we abandoned further development of LS5218 for 1-octanol production and performed all further experiments in K12 MG1655.
Blocking Alternative Carbon Sinks
When rapidly growing, E. coli is known to produce overflow metabolites, including acetate and lactate, that can reduce the final titer of desired products (Atsumi et al., 2008). In prior studies, we observed up to 7% of carbon flux being directed to acetate instead of the desired dodecanoic acid (Youngquist et al., 2012). Therefore, we created derivatives of MG1655 ΔfadD with additional gene deletions to block acetate and lactate secretion: phosphate acetyltransferase (pta), pyruvate oxidase (poxB), and lactate dehydrogenase (ldhA). These strains were transformed with pTRC99a-CpFatB1*. Cultures of each strain were grown in three media conditions, rich media with glycerol, rich media with glucose, and phosphate-limited minimal media with glucose. Cells were induced with 50 μM IPTG (optimal level from Figure S1C), and incubated with shaking at 30°C for 24 h after induction. In rich media, the deletion strains generated less octanoic acid than the base strain containing all fermentation pathways (Figure S1D–E). In phosphate-limited MOPS minimal media supplemented with glucose, all strains produced 0.40–0.49 g/L and no increase in titers was seen for having the additional deleted pathways (Figure S1F). Overall, we did not find that fermentative pathways were a major carbon sink when grown in batch cultures.
Acyl-CoA Synthetase
The next enzymatic step towards production of octanol is activation of octanoic acid via an acyl-CoA synthetase (ACS). We tested ACS homologs, Pseudomonas putida Pp-0763 (Wang et al., 2012; Agnew et al., 2012) and Mycobacterium tuberculosis MtFadD6 (Arora et al. 2005) to evaluate their activity towards octanoic acid in-vivo relative to that of E. coli EcFadD. These ACSs have been shown in the past to have wide substrate specificity, including medium-chain activity. In addition, Pp0763 was successfully used to synthesize C8 and C10 polyhydroxyalkanoates (Wang et al., 2012). In the past, we have tested these enzymes for dodecanoic acid activity but were unable to outperform EcFadD (Youngquist et al., 2013). However, EcFadD has particularly weak enzymatic activity towards acyl-chains of less than ten carbons and efforts to alter its substrate preference have been met with limited success (Iram and Cronan, 2006; Ford et al., 2015).
In this experiment, we expressed each ACS candidate from a plasmid in strain MHS04 (E. coli MG1655 ΔfadR ΔfadD) and cultured cells in the presence of octanoic acid (Youngquist et al., 2013). The ΔfadR mutation in MHS04 deregulates expression of enzymes in the β-oxidation pathway (Figure 2A) and the ΔfadD deletion makes plasmid-based ACS expression necessary for activation of octanoic acid to octanoyl-CoA. Cells were inoculated from LB media overnight cultures into 50 mL of LB media with 250 mg/L octanoic acid, induced at OD600 0.2 with 1 mM IPTG and grown at 30°C with shaking for an additional 12 h post induction. Cells carrying MtFadD6 were able to consume most of the octanoic acid by 5 h post induction, and cells carrying Pp00763 were able to consume most of the octanoic acid by 8 h (Figure 2B). Cells expressing EcFadD lagged behind and were unable to consume the octanoic acid in 12 hours. This data suggests that both MtFadD6 and Pp00763 are active on octanoic acid and are suitable enzymes for assembling the pathway toward 1-octanol. For the rest of this study, we used Mt-FadD6 when expressing an acyl-CoA synthetase.
1-Octanol evaporation
We measured the evaporative loss of octanol from similar environments to assess if accurate titering was possible. In preliminary studies, we detected 1-octanol odors in production experiments, however, in samples taken after 24 h of incubation, a 1-octanol peak was not detected by gas chromatography. We set up shake flasks containing 50 mL of LB media supplemented with 250 mg/L of 1-octanol, incubated the flasks at 37°C with shaking, and took samples at different times. After 12 h, 85% of the 1-octanol had been lost from the system, presumably to evaporation or air stripping from the LB media (Figure S2). Prior studies used dodecane in cell cultures as a distinct organic phase to provide a product sink (Youngquist et al., 2013). We explored the effect of dodecane on the loss of octanol from flasks. We repeated the experiment by adding 5 mL (1:10) and 10 mL (1:5) dodecane to the 50 mL LB media supplemented with 250 mg/L 1-octanol and tracked the evaporation of 1-octanol over time. Interestingly, addition of dodecane reduced octanol loss but led to underestimation of octanol titers by approximately 20–25%. The underestimation stems in part from challenges associated with sampling an accurate volume of both phases, something we corrected using an internal standard in the production flask (Figure S3). Based on these results, all 1-octanol quantification was performed using cultures containing 1:5 v:v ratio of dodecane:media.
Acyl-CoA reductase
We next sought an ACR that would allow us to convert the octanoyl-CoA generated from the activity of CpFatB1* and ACS to 1-octanol. In an initial test, we showed that both ACS and ACR heterologous activities were required for consumption of exogenous octanoic acid in strain DE (E. coli K12 MG1655 ΔfadD ΔfadE) (Figure S4). This test used the previously characterized ACR from Marinobacter aquaeolei (Willis et al., 2011) that our group used to produce dodecanol (Youngquist et al., 2013). In search of more effective enzymes, we tested eight additional acyl-CoA reductases identified in prior work (Mehrer et al., 2018) using the Enzyme Similarity Tool (Gerlt et al., 2015) and tested in a strain engineered to produce medium-chain fatty alcohols as anaerobic fermentation products. In order to test these variants, we fed octanoic acid to cultures of DE harboring ACS plasmid pACYC-MtfadD6 and various ACR variants on pTRCK plasmid while monitoring 1-octanol production over time (Figure 3B). In addition to MaACR, we found that MtACR from Methylibium sp., and MbACR from Marinobacter BSs20148 were able to have high turnover of octanoyl-CoA to 1-octanol. In the work by Mehrer and coworkers, MtACR was seen to express well and produce alcohols in anaerobic conditions, but MbACR did not express or produce alcohols. Interestingly, we found that both of them produce alcohols at the same levels as MaACR in aerobic conditions. It is not clear which of these enzymes produced the most 1-octanol from aerobic conditions since there is some evaporation occurring simultaneously. For this reason, we decided to continue using MaACR throughout the study. Furthermore, after 24 h the levels of 1-octanol accumulated was 30–50% percent lower than the 18 h time point suggesting that there is still considerable evaporation occurring even in the presence of dodecane and the actual 1-octanol production might be higher than the accumulation seen in the flask.
Assembly of the full pathway
With all enzymes selected, we assembled the complete pathway in order to test the ability of E. coli K12 MG1655 to produce 1-octanol from unrelated carbon sources. We initially expressed each enzyme from separate plasmids in E. coli strain DE (ΔfadD ΔfadE). The thioesterase CpFatB1* was expressed from pTRC99a, the ACS MtFadD6 from pACYC, and the ACR MaACR from pBTRK (see Tables S1 and S2 for a detailed description of strains and plasmids). A control strain lacking the ACS expression vector was created to track free fatty acid accumulation as a baseline. Cultures of E. coli DE containing each plasmid were grown in shake flasks containing 50 mL MOPS rich media and samples were collected at 12 h, 18 h, and 24 h to track accumulation of products as well as evaporation effects. Control strains lacking an ACS produced 1.1 g/L octanoic acid after 18 h under these conditions (Figure 4A) and did not produce any alcohols (Figure 4B, first panel). The cells also produced C16 and C18 saturated and unsaturated fatty acids at ratios consistent with prior E. coli studies (Lennen, 2010; Lennen, 2013). It should be noted that the acid-catalyzed methylation reaction used to produce quantifiable fatty acid methyl-esters also trans-esterifies lipid esters. When the MtFadD6 ACS was co-expressed in the DE strain, cultures accumulated 1.0 g/L 1-octanol after 18 h and minor amounts of other chain-lengths (Figure 4B, first panel). Interestingly, we observed a similar strain (ΔaraBAD ΔfadD) carrying the same pTRC99a-CpFatB1* expression vector and expressed under the same conditions (Figure S2C) produce 50% more octanoic acid. This suggests that there may be a burden associated with carrying three plasmids in the cell.
For this reason, we created strains to integrate all genes to the chromosome of E. coli. We constructed E. coli NHL19 (ΔfadE, ΔfadD::PTRCCpFatB1*) and repeated the experiment as described above with pTRC99a empty plasmid taking the place of the pTRC99a-CpFatB1* plasmid. NHL19 lacking MtFadD6 ACS accumulated 0.9 g/L octanoic acid after 18 h (Figure 4A, second panel) while not producing any alcohols (Figure 4B second panel). When the MtFadD6 ACS was included, NHL19 accumulated 0.6 g/L 1-octanol after 18 h and minor amounts of other chain-lengths (Figure 4B, second panel). Interestingly, the 1-octanol titers observed were substantially less than the octanoic acid titers produced in the corresponding control strains. We suspect that this difference is caused by a combination of factors including 1-octanol evaporation and the extra metabolic cost (1 ATP and 2 NADPH) required to convert octanoic acid to 1-octanol.
Next, we moved the MtFadD6 ACS to the chromosome, generating NHL22 (ΔfadE, ΔfadD::PTRCCpFatB1*, ΔackA-pta::PTRC-MtFadD6). We repeated the production experiment as described above with empty pACYC and with pACYC-MtFadD6 in order to compare chromosomal expression against the higher copy expression vector in the same strain. Interestingly, strain NHL22 maintained the same level of 1-octanol production made by NHL19 (Figure 4B, third panel). Additional plasmid expression of MtFadD6 did not increase the performance of the strain in terms of 1-octanol production after 18 h, albeit the earlier 12 h time point shows less 1-octanol. Moreover, little accumulation of octanoic acid was observed in this strain, suggesting that chromosomal expression of ACS was sufficient to process the octanoic acid available. Up to this point, these strains maintained three plasmids (empty vectors when expression was tested from the chromosome). However, as more genes were moved to the chromosome (strains NHL23 and NHL24) the plasmid load was reduced.
Last, we moved MaACR to the chromosome creating a fully chromosomal 1-octanol production strain, NHL23 (ΔfadE, ΔfadD::PTRCCpFatB1*, ΔackA-pta::PTRCMtFadD6, ΔFadBA::PTRCMaACR). This strain was tested with an empty pBTRCK and with additional pBTRCK-MaACR expression vector (Figure 4A–B, fourth panel). NHL23 produced over 1 g/L 1-octanol and 0.6 g/L of octanoic acid. Additional expression of MaACR in NHL23 did not increase octanol titers. Given this data, we hypothesized that ACS activity was a bottleneck in the conversion of octanoic acid to octanol. Therefore, we introduced a second copy of the ACS expression cassette on the chromosome, creating strain NHL24 (ΔfadE, ΔfadD::PTRCCpFatB1*, ΔackA-pta::PTRC-MtFadD6, ΔpoxB::PTRC-MtFadD6, ΔFadBA::PTRCMaACR). This strain accumulated 1.3 g/L 1-octanol after 18 h, with lower accumulation of octanoic acid (Figure 4A–B, fifth panel). It is interesting to note that the 1-octanol titer produced by E. coli NHL24 was substantially higher than that of NHL23. The major difference between these strains is the removal of all plasmids, which likely removed a substantial metabolic burden from the cell as seen elsewhere (Glick, 1995; Wu et al., 2016). On the other hand, we attribute the reductions in titer observed in the 24h time points to be due to evaporation. Moreover, we speculate that combining these strains with a fermentation system to capture all the 1-octanol being produced will result in higher observed titers. However, efforts to develop such a system have proven difficult. In aerated bioreactors, gas flow strips volatile products from the aqueous fermentation broth at higher rates than evaporation from shake flasks. Capture of stripped products can theoretically be accomplished by condensation, adsorption on solid resins, or absorption by chilled solvents. We have made multiple attempts to capture the 1-octanol using small scale condensation or absorption systems but technical challenges such as the high aeration rate, humidity of the off-gas stream, clogged lines due to ice formation, absorber temperature, and the composition of the absorber organic layer have led to failures in each case (not shown). Nevertheless, shake-flask titers reported here represent a 2.3-fold increase to the highest 1-octanol titer reported in the literature (see Table S3 for a summary of previous medium-chain alcohol efforts). Importantly, NHL24 is a fully chromosomally integrated strain, a feature that distinguishes it from previous approaches to produce 1-octanol.
Conclusions
In this work, we demonstrated 1-octanol production from simple sugars using a set of E. coli strains engineered to direct flux through fatty acid biosynthesis, a C8-specific thioesterase, an acyl-CoA synthetase, and an acyl-CoA reductase/aldehyde reductase. Our experiments determined that 1.) MG1655 was a superior base strain to LS5218 for aerobic 1-octanol production; 2.) deletion of acetate and lactate pathways had little impact on octanoic acid titers, likely because they are not the current bottlenecks of the pathway; 3.) expression of the thioesterases (CpFatB1*) needed to be low, 4.) two heterologous acyl-CoA synthetases from P. putida and M. tuberculosis were superior to native E. coli FadD for activating octanoic acid, and 5.) the pathway can be expressed fully from the chromosome. Our best plasmid-free strain produced 1.3 g/L 1-octanol in shake flasks. In addition to the strain development findings, we alleviated challenges associated with quantifying octanol by adding an organic overlay and internal standards. Future work will focus on the scaling up of this system to bioreactor experiments as well as developing a 1-octanol capture mechanism.
Supplementary Material
Acknowledgements
Research described in this manuscript was supported by the National Science Foundation (CBET-1703504) and the Dow Chemical Company through a project grant to B.F.P. N.H.L. is the recipient of a NIH Chemistry-Biology Interface Training Program fellowship (No. T32 GM008505) and a Graduate Engineering Research Scholars fellowship from the UW-Madison College of Engineering.
Footnotes
Declaration of competing interest
The authors declare no competing interests.
References
- Agnew DE, Stevermer AK, Youngquist JT & Pfleger BF Engineering Escherichia coli for production of C12-C14 polyhydroxyalkanoate from glucose. Metab. Eng 14, 705–13 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akhtar MK, Dandapani H, Thiel K & Jones PR Microbial production of 1-octanol: A naturally excreted biofuel with diesel-like properties. Metab. Eng. Commun 2, 1–5 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arora P, Vats A, Saxena P, Mohanty D & Gokhale RS Promiscuous fatty acyl CoA ligases produce acyl-CoA and acyl-SNAC precursors for polyketide biosynthesis. J. Am. Chem. Soc 127, 9388–9389 (2005). [DOI] [PubMed] [Google Scholar]
- Atsumi S et al. Metabolic engineering of Escherichia coli for 1-butanol production. Metab. Eng 10, 305–311 (2008). [DOI] [PubMed] [Google Scholar]
- Cronan JE Bacterial Membrane Lipids: Where Do We Stand? Annu. Rev. Microbiol 57, 203–224 (2003). [DOI] [PubMed] [Google Scholar]
- Datsenko KA & Wanner BL One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U. S. A 97, 6640–6645 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dellomonaco C, Clomburg JM, Miller EN & Gonzalez R Engineered reversal of the β-oxidation cycle for the synthesis of fuels and chemicals. Nature 476, 355–9 (2011). [DOI] [PubMed] [Google Scholar]
- d’Espaux L, Mendez-Perez D, Li R & Keasling JD Synthetic biology for microbial production of lipid-based biofuels. Curr. Opin. Chem. Biol 29, 58–65 (2015). [DOI] [PubMed] [Google Scholar]
- Falbe J, Bahrmann H, Lipps W, Mayer D & Guido D Frey. in Ullmann’s Encyclopedia of Industrial Chemistry 235–261 (2011). [Google Scholar]
- Ford TJ & Way JC Enhancement of E. coli acyl-CoA synthetase FadD activity on medium chain fatty acids. PeerJ 3, (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerlt JA, Bouvier JT, Davidson DB, Imker HJ, Sadkhin B, Slater DR, Whalen KL Enzyme function initiative-enzyme similarity tool (EFI-EST): A web tool for generating protein sequence similarity networks Biochim. Biophys Acta: - Proteins Proteomics; 1854, 1019–1037 (2015). 10.1016/j.bbapap.2015.04.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glick BR (1995). Metabolic load and heterologous gene expression. Biotechnology Advances, 13(2), 247–261. 10.1016/0734-9750(95)00004-A [DOI] [PubMed] [Google Scholar]
- Grisewood MJ et al. Computational Redesign of Acyl-ACP Thioesterase with Improved Selectivity toward Medium-Chain-Length Fatty Acids. ACS Catal. 3837–3849 (2017). doi: 10.1021/acscatal.7b00408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haldimann A & Wanner BL Conditional-Replication , Integration , Excision , and Retrieval Plasmid-Host Systems for Gene Structure-Function Studies of Bacteria. J. Bacteriol 183, 6384–6393 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez-Lozada NJ et al. Highly active C8-acyl-ACP thioesterase variant isolated by a synthetic selection strategy. ACS Synth. Biol 2205–2215 (2018). doi: 10.1021/acssynbio.8b00215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoover SW, Marner WD, Brownson AK, Lennen RM, Wittkopp TM, Yoshitani J, Zulkifly S, Graham LE, Chaston SD, McMahon KD, Pfleger BF, 2011. Bacterial production of free fatty acids from freshwater macroalgal cellulose. Appl. Microbiol. Biotechnol 91, 435–446. 10.1007/s00253-011-3344-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iram SH & Cronan JE The Beta-Oxidation Systems of Escherichia coli and Salmonella enterica Are Not Functionally Equivalent. J. Bacteriol 188, 599–608 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S, Clomburg JM & Gonzalez R Synthesis of medium-chain length (C6–C10) fuels and chemicals via β-oxidation reversal in Escherichia coli. J. Ind. Microbiol. Biotechnol 42, 465–475 (2015). [DOI] [PubMed] [Google Scholar]
- Lennen RM, Braden DJ, West R. a, Dumesic J. a & Pfleger BF A process for microbial hydrocarbon synthesis: Overproduction of fatty acids in Escherichia coli and catalytic conversion to alkanes. Biotechnol. Bioeng 106, 193–202 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lennen RM & Pfleger BF Engineering Escherichia coli to synthesize free fatty acids. Trends Biotechnol 30, 659–67 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lennen RM & Pfleger BF Microbial production of fatty acid-derived fuels and chemicals. Curr. Opin. Biotechnol 24, 1044–53 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lennen RM & Pfleger BF Modulating membrane composition alters free fatty acid tolerance in Escherichia coli. PLoS One 8, e54031 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu R et al. Metabolic engineering of fatty acyl-ACP reductase-dependent pathway to improve fatty alcohol production in Escherichia coli. Metab. Eng 22, 10–21 (2014). [DOI] [PubMed] [Google Scholar]
- Marella ER, Holkenbrink C, Siewers V & Borodina I Engineering microbial fatty acid metabolism for biofuels and biochemicals. Curr. Opin. Biotechnol 50, 39–46 (2018). [DOI] [PubMed] [Google Scholar]
- Mehrer CR, Incha MR, Politz MC & Pfleger BF Anaerobic production of medium-chain fatty alcohols via a β-reduction pathway. Metab. Eng 48, 63–71 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pérez JM, Arenas FA, Pradenas GA, Sandoval JM & Vásquez CC Escherichia coli YqhD exhibits aldehyde reductase activity and protects from the harmful effect of lipid peroxidation-derived aldehydes. J. Biol. Chem 283, 7346–7353 (2008). [DOI] [PubMed] [Google Scholar]
- Pfleger BF, Gossing M & Nielsen J Metabolic engineering strategies for microbial synthesis of oleochemicals. Metab. Eng 29, 1–11 (2015). [DOI] [PubMed] [Google Scholar]
- Rand JM, Gordon GC, Mehrer CR & Pfleger BF Genome sequence and analysis of Escherichia coli production strain LS5218. Metab. Eng. Commun 5, 78–83 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarria S, Kruyer NS & Peralta-Yahya P Microbial synthesis of medium-chain chemicals from renewables. Nat. Biotechnol 35, 1158–1166 (2017). [DOI] [PubMed] [Google Scholar]
- Sattayawat P, Sofian Yunus I & Jones PR Bioderivatization as a concept for renewable production of chemicals that are toxic or poorly soluble in the liquid phase. Proc. Natl. Acad. Sci. U. S. A 117, 1404–1413 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- USDA, A.A.D. for the U.N.O.P., Fatty Alcohols (Octanol and Decanol). (2016). [Google Scholar]
- Van Leeuwen PWNM, Clément ND & Tschan MJL New processes for the selective production of 1-octene. Coord. Chem. Rev 255, 1499–1517 (2011). [Google Scholar]
- Willis RM, Wahlen BD, Seefeldt LC & Barney BM Characterization of a Fatty Acyl-CoA Reductase from Marinobacter aquaeolei VT8: A Bacterial Enzyme Catalyzing the Reduction of Fatty Acyl-CoA to Fatty Alcohol. ACS Biochem. 50, 10550–10558 (2011). [DOI] [PubMed] [Google Scholar]
- Wu G, Yan Q, Jones JA, Tang YJ, Fong SS, & Koffas MAG (2016). Metabolic Burden: Cornerstones in Synthetic Biology and Metabolic Engineering Applications. Trends in Biotechnology, 34(8), 652–664. 10.1016/j.tibtech.2016.02.010 [DOI] [PubMed] [Google Scholar]
- Yan Q & Pfleger BF Revisiting metabolic engineering strategies for microbial synthesis of oleochemicals. Metab. Eng 58, 35–46 (2020). [DOI] [PubMed] [Google Scholar]
- Youngquist JT et al. Kinetic modeling of free fatty acid production in Escherichia coli based on continuous cultivation of a plasmid free strain. Biotechnol. Bioeng 109, 1518–27 (2012). [DOI] [PubMed] [Google Scholar]
- Youngquist JT et al. Production of medium chain length fatty alcohols from glucose in Escherichia coli. Metab. Eng 20, 177–86 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Youngquist JT, Rose JP & Pfleger BF Free fatty acid production in Escherichia coli under phosphate-limited conditions. Appl. Microbiol. Biotechnol 97, 5149–59 (2013). [DOI] [PubMed] [Google Scholar]
- Wang Q, Tappel RC, Zhu C & Nomura CT Development of a new strategy for production of medium-chain-length polyhydroxyalkanoates by recombinant Escherichia coli via inexpensive non-fatty acid feedstocks. Appl. Environ. Microbiol 78, 519–527 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X, Miller JR, Jiang Y, Marletta MA & Cronan JE Assembly of the Covalent Linkage between Lipoic Acid and Its Cognate Enzymes. Chem. Biol 10, 1293–1302 (2003). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.