

Abstract
Mesenchymal stem cells (MSCs) are a reliable source for cell-based regenerative medicine owing to their multipotency and biological functions. However, aging-induced systemic homeostasis disorders in vivo and cell culture passaging in vitro induce a functional decline of MSCs, switching MSCs to a senescent status with impaired self-renewal capacity and biased differentiation tendency. MSC functional decline accounts for the pathogenesis of many diseases and, more importantly, limits the large-scale applications of MSCs in regenerative medicine. Growing evidence implies that epigenetic mechanisms are a critical regulator of the differentiation programs for cell fate and are subject to changes during aging. Thus, we here review epigenetic dysregulations that contribute to MSC aging and osteoporosis. Comprehending detailed epigenetic mechanisms could provide us with a novel horizon for dissecting MSC-related pathogenesis and further optimizing MSC-mediated regenerative therapies.
1. Introduction
1.1. Mesenchymal Stem Cells (MSCs)
MSCs are adult stem cells distributed in various mesenchymal tissues, which are derived from the mesoderm in the embryonic stage. MSCs exist in diverse tissues, such as bone marrow, umbilical cord blood, placenta, and adipose tissue [1]. Since MSCs are firstly isolated and defined from bone marrow, it has been traditionally accepted that bone marrow is the prevailing source of MSCs in humans [2, 3]. MSCs from different origins possess unique self-renewal capacities and can differentiate into multilineage cell types, including osteocytes, adipocytes, chondrocytes, and even endothelial cells or hepatocytes under certain given culture medium [4–6]. Apart from the aforementioned two basic characteristics, MSCs also exhibit various positive effects through paracrine action and immunomodulation during tissue repair, including regulating angiogenesis and osteoclastogenesis and guiding immune communication [7–9]. These properties signify that MSCs could perform extensive and active interactions with tissue-specific stem cell niches and represent an ideal and promising tool for tissue regeneration.
Although tentative therapeutic applications of MSCs have been carried out in the past years, disadvantages such as poor cell sources from diseased or aged hosts and in vitro passaging-induced senescent hypofunction both impair their therapeutic efficacy in tissue regeneration and hinder their large-scale clinical trials. MSC senescence manifests as division arrest, reflected by impaired proliferation and biased differentiation from osteoblasts towards adipocytes. Therein, biased differentiation can be induced by the imbalance between runt-related transcription factor 2 (Runx2) and peroxisome proliferator-activated receptor γ (PPARγ) pathway. These changes during senescence underlie bone mass loss and fat accumulation in aged or diseased skeletal tissues [10–13]. MSC aging is molecularly characterized by upregulated expression of senescence-associated genes such as p53, p21, p16INK4a, and β-galactosidase genes [14]. Notably, epigenetic regulation has emerged as a vital contributor to MSC aging and hypofunction, thus perturbing stem cell niche homeostasis and harming tissue health. Intriguingly, epigenetic alterations have also been demonstrated to modulate canonical senescence-associated genes directly or indirectly. Accordingly, therapeutic strategies based on epigenetic regulation may remedy tissue disorder in aging and diseases and further maximize the advantages of MSC-mediated tissue regeneration. In this review, we mainly focus on epigenetic marks and modifiers in regulating MSC aging in vivo or in vitro, in order to clarify the interactive link between epigenetic regulation and aging-related tissue diseases such as osteoporosis, and offer some clues for future utilization of epigenetics mediated tissue regeneration [15].
1.2. Epigenetic Regulation
Epigenetic regulation refers to altering phenotype through gene differential expression without changing DNA sequence and is a characteristic of heritability, reversibility, and no gene changes [11, 16]. Epigenetic alterations in cells happen in response to extrinsic environmental stimuli and cellular intrinsic inheritance to maintain cell and niche homeostasis. Accordingly, MSC aging or senescence in vivo or in vitro is also influenced by its own intrinsic dysregulation and microenvironmental factors from MSC niche, in the process of which typical epigenetic marks could be detected. In MSCs, epigenetic profile reflects dynamically transforming chromatin structure and corresponding transcriptional activity of genes; the major epigenetic mechanisms include DNA methylation, histone modifications, and chromatin remodeling [17]. In addition, posttranscriptional processing through mRNA and noncoding RNAs (ncRNAs) also takes part in epigenetic regulation of MSCs [18] (Figure 1). It has been widely documented that these epigenetic marks all have profound influences on MSC fate at multiple levels. Hence, further rationalizing and understanding the function mechanism of different epigenetic marks and modifiers occurring in MSC aging are of instructive importance to analyze the pathogenesis of aged and diseased tissue disorders and explore more effective therapeutic or regenerative strategies.
Figure 1.
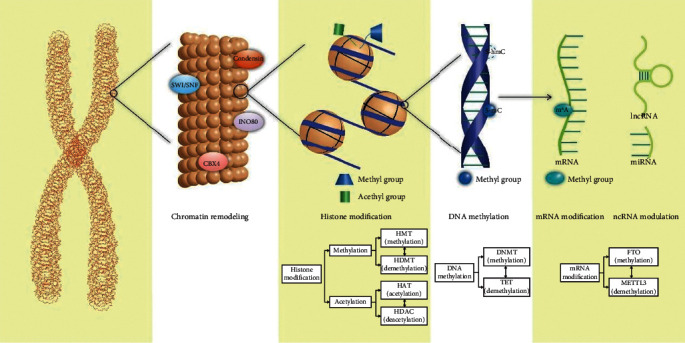
Factors in epigenetic regulation. Epigenetic regulation occurs in several steps of transcription (such as chromatin remodeling, DNA methylation, and histone modification) and posttranscription (such as mRNA processing and ncRNA regulation). Specific regulatory factors participate in each process.
2. Epigenetic Regulation in MSC Aging and Osteoporosis
2.1. DNA Methylation in MSC Aging and Osteoporosis
DNA methylation refers to the covalent binding of methyl to the 5th carbon of cytosine at CpG dinucleotide to form 5-methylcytosine (5-mC) under the catalysis of DNA methyltransferase (DNMT) [19]. This process can be reversed by demethylation relying on the catalysis of ten-eleven translocation protein (TET), which catalyzes the transformation of the 5th carbon of cytosine into 5-hydroxymethylcytosine (5-hmC) [20]. In most cases, methylation of gene promoters or enhancers represents repressed expression (Table 1).
Table 1.
DNA methylation in MSC aging and related diseases.
| Regulatory factors | Mechanism | In vivo or in vitro | Consequence | Materials | Ref. |
|---|---|---|---|---|---|
| Senescence and aging | |||||
| DNMT1↓ DNMT3b↓ |
Decreases methylation in the promoter region of miRNA targeting Ezh2 to decrease Ezh2 expression, thus inhibiting repressive H3K27me3 formation at p16's promoter; directly decreases methylation in CDK inhibitor genes p16 and p21 | In vitro | Aging | hUCSCs | [28] |
| DNMT↓ | Insufficient to methylate Tert promoter; thus upregulated TERT would repair the shortened telomeres with replication | In vitro | Antisenescence | hBMSCs | [30] |
| Skeletal diseases | |||||
| DNMT1 changes | Aberrant methylation of ABCB1 gene leads to a dysregulation of glucocorticoid | In vitro | ONFH | hBMSCs | [33] |
| TET1 and TET2↓ | The depletion of TET causes impeded demethylation of P2rX7 promoter and incapable exosome release, which leads to intracellular accumulated miR-297 targeting Runx2 signaling pathway | Both | Osteoporosis | hBMSCs, mouse BMSCs, mouse model | [34] |
No matter in vitro or in vivo, alteration of DNA methylation profile gradually emerges as a close connection to MSC aging. Recently, researchers have successfully detected gene sites with methylation changes in the aging process by BeadChip microarray and found that alteration of methylation overlaps in aged MSCs in vivo and in vitro [21]. Afterward, in 2015, more than 10000 hypermethylation CpG sites and 40000 hypomethylation CpG sites were uncovered, many of which are associated with homeobox genes related to cell differentiation. For example, Hox and Runx2, as key transcription factors for osteogenesis, are hypermethylated in aged MSCs [21–23]. Moreover, in 2017, enhanced reduced representation bisulfite sequencing was used to depict a more precise DNA methylation profile, which finds that transcription factor binding sites (TFBS) for silent regulator 6 (Sirt6), E2F transcription factor 6 (E2F6), JunB proto-oncogene, and signal transducer and activator of transcription (Stat5) genes were hypermethylated along with the culture process, while TFBS for Stat3 gene were hypomethylated. Besides, transcription factors influencing chromatin structure, such as SMARCs and SIN3A, are also differently methylated [24]. In general, the degree of methylation generally decreases in the process of aging [22]. On the other hand, these DNA methylation sites have also been discovered to be related to repressive and promotive histone modification, respectively. In MSCs, a large number of hypomethylated CpG sites are enriched in the region of active histone mark methylation of lysine 4 on histone H3 (H3K4me) indicating that H3K4 methylation is accompanied by DNA hypomethylation, and both of them are signs of increased transcription activity. By contrast, the hypermethylation CpG DNA region mainly overlaps with the repressive chromatin marks trimethylation of lysine 27 on histone H3 (H3K27me3), H3K9me3, and enhancer of zeste2 (EZH2), all of which play synergistic roles in inhibiting transcription activity [22, 25]. Particularly, H3K9me-promoted DNA hypermethylation of gene p16INK4a has also been proved to serve as a significant signature for epigenetic senescence in human MSCs (hMSCs) [26]. In MSCs isolated from aged individuals, 5-hmC also appears in specific CpG sites and, coincidentally, corresponds mostly to the hypomethylation region in the aged MSCs [27].
In the complicated regulatory network of DNA methylation, some regulatory factors and sites have been identified to have a direct relationship with MSC aging. For instance, 5-azacytidine (5-AzaC), as an inhibitor of DNMT1 and DNMT3b, exacerbates cell senescence by downregulating polycomb group proteins (PcGs) including B cell-specific Moloney murine leukemia virus integration site 1 (BMI1) and EZH2 through miRNAs. Since these PcGs are responsible for repressive mark H3K27me3 formation at the p16 gene promoter, 5-AzaC finally facilitates p16's activation in cell senescence. Moreover, inhibition of DNMT also directly demethylates cyclin-dependent kinase (CDK) inhibitor genes p16 and p21 [28]. However, 5-AzaC and another DNMT inhibitor RG108 were also reported to alleviate senescence by preventing reactive oxygen species (ROS) accumulation and maintaining telomerase reverse transcriptase (Tert) activity in aged human bone marrow MSCs (hBMSCs) [29, 30].
The methylation status of DNA is also responsible for differentiation potential and further influences the process of skeletal diseases. For example, in MSCs with high stemness, stemness-related transcription factors Octamer-binding transcription factor (OCT4) and NANOG protein both directly bind to the promoter region of DNMT1 and then transcriptionally activate it, followed by methylation and silencing of senescence-related genes [31]. On the contrary, when MSCs are treated with a DNMT1 inhibitor 5-AzaC, downregulation of DNMT1 is accompanied by hypomethylation of genomic DNA and increased expression of osteogenic genes such as Runx2, Osteocalcin (Ocn), distal-less homeobox 5, and Osterix (Osx), which is more visually presented by enhanced alkaline phosphatase (ALP) activity and mineralization [32]. Clinically, it has been reported that DNMT1 dysfunction also influences skeletal metabolic homeostasis. Aberrant CpG hypermethylation at gene ATP-binding cassette subfamily B member 1 (ABCB1, the encoding gene of P-glycoprotein) leads to osteonecrosis of the femoral head (ONFH) [33]. Besides, depletion of demethylase TET1 and TET2 causes osteopenia phenotype in mice by impeding demethylation of P2rX7 promoter; P2rX7 deficiency further leads to MSC incapability of exosome release, which results in intracellular accumulated miR-297 targeting Runx2 signaling pathway [34].
2.2. Histone Modification in MSC Aging and Osteoporosis
Histone modification is closely related to transcriptional activities of genes surrounding it. There are many different covalent modification types of the N-terminal amino acids of histone lysine, including acetylation, methylation, phosphorylation, ubiquitylation, and SUMO modification. Generally speaking, the first two are related to transcription activation, while the latter three often dominate transcription inhibition, with the exception of H3K4me3 [35]. In cases of aging and diseases related to MSCs, histone acetylation levels depending on the balance between histone deacetylases (HDACs) and histone acetyltransferases (HATs) and methylation levels relying on histone methyltransferases (HMTs) and histone demethylases (HDMTs) both make a critical difference [36, 37] (Table 2).
Table 2.
Histone modification in MSC aging and related diseases.
| Epigenetic changes | Factors | Mechanism | In vivo or in vitro | Consequence | Material | Ref. |
|---|---|---|---|---|---|---|
| Senescence and aging | ||||||
| Histone acetylation | HDAC↓ | Directly upregulates JMJD3 and indirectly downregulates PcGs through RB/E2F pathway to inhibit H3K27me3 at p16INK4A | In vitro | Aging | hADSCs, hUCSCs | [38] |
| Histone acetylation | HDAC↓ | Promotes the transcription of p21CIP1/WAF1 through increasing H3 and H4 acetylation | In vitro | Aging: decreased differentiation ability and proliferation rate | hADSCs, hUCSCs | [40] |
| Histone acetylation | SIRT6↓ | Insufficient SIRT6 causes increased H3K56ac and compromised recruitment of RNAP II complex to Hmox-1 gene promoter, leading to decrease in Hmox-1 expression and impaired cellular redox homeostasis | Both | Senescence, dysregulated redox metabolism, and increased sensitivity to oxidative stress | Human embryoid bodies MSC, mouse model | [42] |
| Histone methylation | TWIST1↓ | Insufficient to prevent senescence by recruiting EZH2 and form repressive H3K27me3 at p16/p14 promoters; upregulates E47 that binds to p16 promoter and promotes transcription activity | In vitro | Senescence | hBMSCs | [39] |
| Histone methylation | BMI1↓ | Fails to recruit and stabilize PRC2 which protects H3K27me3 of p16INK4A | In vitro | Aging | hADSCs, hUSCSs | [38] |
| Histone methylation | EZH2↓ | Fails to methylate H3K27 as catalytic subunit of PRC2; insufficient H3K27me3 cannot suppress p16 and p14 expression | In vitro | Aging | hADSCs, hUSCSs | [38] |
| Histone methylation | G9a↓ | (Unclear) | In vitro | Aging: decreased differentiation ability and proliferation rate | Rat BMSCs | [149] |
| Skeletal diseases | ||||||
| Histone methylation | EZH2↑ | Promotes H3K27me3 on Wnt1, Wnt6, and Wnt10a promoters to silence Wnt signaling pathway | Both | Osteoporosis | hBMSCs, mouse BMSCs, mouse model | [47] |
| Histone methylation | KDM5A↑ | Increases H3K4me3 levels on promoters of Runx2 | Both | Osteoporosis | hBMSCs, mouse BMSCs, mouse model | [45] |
| Histone methylation | ASH1L↓ | Fails to mediate H3K4me3 recruitment at the transcription start sites of Osx, Runx2, Sox9, and Creb genes | Both | Osteoporosis | hBMSCs, mouse BMSCs, mouse model | [46] |
| Histone methylation | KDM2B↓ | Unable to be recruited to the promoter of AP-2α and inhibit AP-2α expression via removing H3K4me3 | Both | Oculofaciocardiodental (OFCD) syndrome | hBMSCs, mouse BMSCs, mouse model | [51] |
| Histone acetylation | GCN5 (KAT2A)↓ | Insufficient to increase H3K9 acetylation on the promoters of Wnt genes | Both | Osteoporosis | hBMSCs, mouse BMSCs, mouse model | [48] |
| Histone acetylation | PCAF (KAT2B)↓ | Insufficient to acetylate H3K9 at promoters of BMP2, BMP4, BMPR2B, and Runx2 | Both | Osteoporosis | hBMSCs, mouse BMSCs, mouse model | [49] |
Histone modification regulates senescence by affecting the transcription activity of surrounding DNA related to cell cycle. For instance, the most direct regulators of repressive H3K27me3 can be divided into two categories: one is HMT such as G9a and PcGs including BMI1, EZH2, and SUZ12, and the other is HDMT such as lysine-specific demethylase 1 (LSD1) and jumonji domain-containing protein 3 (JMJD3). Both upregulation of Jmjd3 gene and downregulation of PcG genes suppress H3K27me3 at the promoter of p14 and p16, which triggers the activation of corresponding proteins and then exacerbates MSC aging [38]. On the other hand, HDAC and Twist1 exert their influence in MSC aging at the upstream of PcGs and JMJD3. Downregulation of Twist1 gene expression in aged MSCs is followed by Ezh2 suppression and E47 promotion. Then, the upregulated E47 protein directly binds to the p16 promoter to increase its transcriptional activity, producing a synergistic action with insufficient EZH2 protein [39]. In comparison to Twist1, the regulatory mechanism of HDAC is more specific. In normal cells, HDAC restrains Jmjd3 expression through deacetylating histone near its promoter region and oppositely promotes PcG proteins and c-MYC activity via the RB/E2F pathway. However, in aging MSCs, HDAC deficiency induces hypophosphorylation of retinoblastoma-like protein (RB). This prompts RB to bind to E2F and further compromises the overall expression of PcGs genes. Finally, alterations of JMJD3 and PcGs level bring about cell cycle arrest by demethylating H3K27 at the p16INK4A promoter [38]. Lee et al. also reported that HDAC inhibitors valproic acid and sodium butyrate both promote the acetylation of histone H3 and H4 to activate the transcription of p21CIP1/WAF1, but the p16 expression level remains unchanged unexpectedly [40]. By contrast, another group found that low concentrations of HDAC inhibitor largazole or trichostatin A induce improved proliferation, suppressed differentiation, and delayed aging of hUCSCs. The underlying mechanism is based on histone H3 acetylation and methylation around Tert, Nanog, Oct4, Alp, Opn, and Cxcr4 genes [41]. SIRT6 protein is another regulator with HDAC activity. As an NAD-dependent H3K9 and H3K56-specific deacetylase, SIRT6 deficiency causes acetylated H3K56 accumulation and compromised recruitment of RNA polymerase II (RNAP II) complex to heme oxygenase 1 (Hmox-1) gene promoter. More importantly, without the cooperation of SIRT6, expression of RNAP II, Nrf2, and Hmox-1 genes declines, resulting in impaired cellular redox homeostasis [42].
In terms of diseases, an imbalance between histone modifications of osteogenic and lipogenic genes is a possible mechanism. Intriguingly, as for histone modification, the regulatory effects of the same factor on osteogenic differentiation or adipogenic differentiation are not necessarily opposite. In other words, factors that promote adipogenesis may either inhibit or promote the biological osteogenic process [43]. For instance, mixed lineage leukemia protein (MLL), general control non-derepressible5 (GCNs, namely, KAT2A), and P300/CBP-associated factor (PCAF, namely, KAT2B) can promote both osteogenesis and lipogenesis, while HDAC1 can inhibit both. Moreover, SET domain bifurcated 1 protein, lysine-specific demethylase 4B/6B (KDM4B/6B), and HDAC3 all promote osteogenesis but inhibit lipogenesis; oppositely, EZH2 and HDAC6 promote lipogenesis but inhibit osteogenesis [43, 44]. Thus, homeostasis of bone tissues relies largely on coordinating and orderly expression in spatial-temporal dimensions. And histone modification dysregulation in osteoporosis is closely related to the break of balance among associated regulatory factors. For instance, in osteoporosis, the upregulated EZH2 and KDM5A and downregulated absent, small, or homeotic 1-like (ASH1L) genes suppress Wnt and Runx2 pathways by altering H3K4me3 and H3K27me3 levels [45–47]. Similarly, H3K9 acetyltransferase GCN5 and PCAF (namely, KAT2A and KAT2B) gliding deacetylates H3K9 on the promoter of Wnt, BMP, and Runx2 genes [48–50]. As to the mechanism of oculofaciocardiodental (OFCD) syndrome characterized by extremely long dental roots and craniofacial defects, recruitment restriction of KDM mediated by BCL-6 corepressor increases the H3K4me3 level and promotes upregulation of AP-2α, whose osteogenesis-fortifying function is overactivated, leading to osteogenic hyperfunction in OFCD syndrome [51].
2.3. Chromatin Remodeling in MSC Aging and Osteoporosis
In a narrow sense, chromatin remodeling is an ATP-dependent process catalyzed by chromatin-remodeling complexes. The core component of the complexes is an ATPase subunit from the SNF2 family including SWI/SNF (switch/sucrose nonfermentable) and INO80 [SWI2/SNF2 related (SWR)] subfamilies [52]. In a broad sense, all factors that bring about chromatin structural alterations, including the relaxing or packing of chromatin by histone modification, contribute to chromatin remodeling (Table 3).
Table 3.
Chromatin remodeling in MSC aging and related diseases.
| Regulatory factors | Mechanism | In vivo or in vitro | Material | Consequence | Ref. |
|---|---|---|---|---|---|
| Senescence and aging | |||||
| BRG1↓ | Inhibits Nanog gene expression by facilitating DNMT recruitment and methylation; induces γ-form heterochromatin formation and p53 pathway activation | In vitro | hBMSCs | Senescence | [53, 54] |
| Condensin↓ | Fails to alleviate DNA damage by chromatin reorganization | Both | hBMSCs, mouse BMSCs, mouse model | Aging, bone aging | [56] |
| KAP1↓ | Cannot be recruited by insufficient CBX4 at nucleolar rDNA, enhancing the excessive expression of rRNAs and destabilizing nucleolar heterochromatin | Both | hMSCs derived from embryonic cell culture, mouse BMSCs, mouse model | Premature cellular senescence, osteoarthritis | [58] |
| Skeletal diseases | |||||
| INO80↓ | Incapable of interacting with Wdr5 that catalyzes H3K4me3 formation, which promotes Wnt pathway activity | Both | hBMSCs, mouse model | Osteoporotic phenotypes | [59] |
During the process of aging, the protein encoded by Brahma-related gene 1 (Brg1), the ATPase subunit of SWI/SNF chromatin remodeling complex, has been regarded as an essential factor in global modulation. Both the upregulation and downregulation lead to acceleration of cell senescence. On the one hand, when Brg1 is silenced, the chromatin compaction cannot be completed. This facilitates DNMT recruitment and methylation at Nanog promoter and eventually induces transcription inhibition [53]. Besides, Brg1 insufficiency-induced senescence is also linked to γ-isoforms of heterochromatin formation and p53 activation-induced cell cycle arrest [53, 54]. On the other hand, overexpression of Brg1 also induces an increasing portion of programmed cell death, despite the fact that the specific mechanism is not clarified [55].
Apart from chromatin remodeling complexes, factors directly related to the chromatin structure, such as condensin and KRAB-associated protein 1 (KAP1), also contribute to chromatin remodeling. It has been reported that condensin I/II can alleviate DNA damage by chromatin reorganization in normal cells. However, hypermethylation around NCAPD2/NCAPG2, the encoding genes of the core components of condensin, leads to condensin shortage and DNA damage accumulation during aging [56]. Loss of heterochromatin is proved to be a potential cause of MSC aging [57]. For example, abnormality of the heterochromatin component KAP1 promotes MSC aging via a chromobox4- (CBX4-) dependent manner. When CBX4 declines in aged hMSCs, fibrillarin (FBL) and KAP1 cannot be recruited at nucleolar rDNA, leading to excessive expression of rRNAs, then trigger detrimental ribosome biogenesis and destabilize nucleolar heterochromatin [58].
As to osteoporosis, the INO80 chromatin remodeling complex interacts with WD repeat-containing protein 5 (Wdr5) protein that catalyzes H3K4me3 formation to positively regulate the canonical Wnt pathway. Correspondingly, after INO80 gene is knocked down, the osteogenic potential of MSCs decreases both in in vitro and in ectopic transplantation models, reproducing a similar phenotype as osteoporosis [59].
In general, chromatin remodeling is intertwined with DNA methylation and histone modification; the DNA and histone modification status directly determines the accessibility and structure of chromatin. Histone acetylation may “open” chromatin by neutralizing the positive charges of lysine to increase site exposure of the surrounding negatively charged DNA [60]. Meanwhile, histone methylation modulates the synthesis of chromatin remodeling-related proteins [56]. Thus, the functional importance of the interactive mechanism is to realize orderly integration and feedback of all three processes.
2.4. mRNA Modification in MSC Aging and Osteoporosis
There are only a few studies about mRNA modification of MSCs during aging. Even so, the biological activity of MSCs in a normal bone is closely related to RNA N6-methyladenosine (m6A) modification. Osteogenesis induced by methyltransferase like 3 (METTL3) is counterbalanced with lipogenesis promoted by demethyltransferase fat mass and obesity-associated protein (FTO) [61]. During the aging process, expression of Fto gene increases and inhibits m6A formation on Pparγ mRNA, which results in lipogenesis through the GDF11-FTO-PPARγ axis [62]. However, the adipogenesis and osteoporosis process could be prevented by METTL3 application [63].
2.5. ncRNA in MSC Aging and Osteoporosis
2.5.1. ncRNA in MSC Aging
Alterations of miRNA and long noncoding RNA (lncRNA) abundance are closely associated with the aging process of MSCs either in vivo or in vitro. Researchers compared miRNA profiles in aged MSCs with younger generations to figure out up- or downregulated miRNA types (Table 4). Data from different organizations varied widely, probably because of different sources of MSCs or the experimental conditions [64–66]. Notably, the variation among MSC miRNA expression profiles from different tissues indicates that their regulatory mechanisms are relatively tissue-specific [67, 68].
Table 4.
ncRNAs in MSC aging and related diseases.
| Regulatory factors | Mechanism | In vivo or in vitro | Consequence | Material | Ref. |
|---|---|---|---|---|---|
| Senescence and aging | |||||
| miR-10a↓ | Insufficient to target Klf4 and repress its function | Both | Senescence, decreased differentiation | hBMSCs | [74, 75] |
| miR-20b-5p and miR-106a-5p↓ | Insufficient to inhibit Smads/p21/CDK/E2F pathway, which alleviates suspension of DNA synthesis during oxidative stress-induced premature senescence | In vitro | Premature senescence | hBMSCs | [77] |
| miR-22 and miR-485-5p↑ | Targets CKS1 to downregulate CDK1 and cyclin B | In vitro | Senescence | SHED | [69] |
| miR-31a-5p↑ | Targets E2F2 and promotes SAHF formation | Both | Senescence | Rat BMSCs | [71] |
| miR-27b↑ | Upregulates p16 expression and MAPK pathway activation | In vitro | Senescence | Pig ADSCs | [98] |
| miR-29c-3p↑ | Targets CNOT6 thus inducing senescence via p53/p21 and p16/pRB pathways | In vitro | Senescence | hBMSCs | [72, 73] |
| miR-34a↑ | Reduces CDK2, CDK4, CDK6, and cyclin D and E expression to hinder the SOX2-related self-renewal ability | In vitro | Senescence | hADSCs | [70] |
| miR-34a↑ | Targets Sirt1 to induce senescence via Sirt1/FoxO3a pathway, induces mitochondrial dysfunction | In vitro | Senescence and intrinsic apoptosis | Mouse BMSCs, rat BMSCs | [89, 90] |
| miR-141-3p↑ | Targets Zmpste24 transcripts, causing prelamin A accumulation in nuclear envelope and intracellular DNA damage | In vitro | Senescence | hUCSCs | [92] |
| miR-141-3p↑ | Targets YAP to inhibit proliferation and accelerate senescence | In vitro | Senescence | Human papilla apical stem cells | [93] |
| miR-142↑ | Targets Epas1 to downregulate pexophagic activity and induce ROS accumulation | In vitro | Aging | Mouse BMSCs | [81] |
| miR-155-5p↑ | Targets Cab39 and then reduces mitochondrial fission and increases mitochondrial fusion via the Cab39/AMPK signaling pathway | Both | Aging | Mouse model, hBMSCs | [86] |
| miR-155-5p↑ | Targets Bag5 that encodes partner protein of PINK1, to inhibit mitophagy and dysfunctional mitochondria elimination | In vitro | Aging | hBMSCs | [87] |
| miR-155-5p↑ | Targets the common transcription factor C/EBP-β thus repressing antioxidant genes and inducing ROS production | Both | Aging | hBMSCs/mouse BMSCs | [82] |
| miR-182↑ | Targets FoxO1, which is critical to protecting cells from ROS | In vitro | Aging, decreased proliferation, and osteogenesis | hBMSCs | [83] |
| miR-183-5p↑ | Targets Hmox-1 to impair response to oxidative stress | In vitro | Senescence | Mouse BMSCs | [84] |
| miR-188↑ | Targets HDAC9 and RICTOR | Both | Aging, decreased proliferation | Mouse BMSCs, mouse model | [94, 95] |
| miR-195↑ | Targets Tert and prevents TERT to repair the shortened telomeres with replication | In vitro | Aging | Mouse BMSCs | [91] |
| miR-199b-5p↓ | Insufficient to repress LAMC1 | In vitro | Senescence | hBMSCs | [80] |
| miR-206↑ | Targets Alpl, which is essential for the intracellular ATP level and AMPK pathway | Both | Premature senescence | Rat BMSCs, rat model | [96] |
| miR-363-3p↑ | Targets TRAF3, which inhibits adipogenic differentiation and senescence | In vitro | Senescence, upregulated adipogenesis | Rat BMSCs | [99] |
| miR-486-5p↑ | Targets Sirt1 | In vitro | Senescence | hADSCs | [85] |
| miR-543 and miR-590-3p↓ | Insufficient to target AIMP3/p18 to inhibit expression, inducing an increase in CDK inhibitors p16INK4A and p21CIP1/WAF1 | In vitro | Senescence | hUCSCs | [78] |
| miR-1292↑ | Targets Wnt receptor FZD4, thus hindering the Wnt/β-catenin/TCF/LEF1 pathway | In vitro | Senescence, downregulated osteogenesis | hADSCs | [100] |
| lncRNA-Bmncr↓ | Insufficient to serve as a scaffold to facilitate the interaction of ABL and transcriptional coactivator with TAZ, hindering the assembly of the TAZ and RUNX2/PPARγ transcriptional complex | Both | Aging, transition from osteogenesis to adipogenesis | hBMSCs, mouse BMSCs | [103] |
| lncRNA-HOTAIR↑ | Modulates senescence-associated changes in gene expression and DNA methylation via triple helix DNA-DNA-RNA formation | In vitro | Senescence | hBMSCs | [102] |
| Skeletal diseases | |||||
| miR-21↓ | Insufficient to target Spry1, which negatively regulates osteogenesis via FGF and MAPK | Both | Osteoporosis | hBMSCs, mouse model | [112] |
| miR-21a↓ | Insufficient to target Pten and PTEN downregulates AKT pathway to induce osteocyte apoptosis | Both | Glucocorticoid-induced osteonecrosis | hUCSCs, mouse model | [116] |
| miR-23b↑ | Targets Runx2 | Both | Osteoporosis | hBMSCs, mouse model | [106] |
| miR-27↓ | Insufficient to target Mef2c, which facilitates the adipogenic differentiation | Both | Osteoporosis | hBMSCs, mouse model | [113] |
| miR-181a↓ | Leads to the accumulation of FasL from BMSCs, followed by CD4+ T cell apoptosis | Both | Osteoporosis | Mouse BMSCs, mouse model | [115] |
| miR-212 and miR-384↑ | Targets Runx2 | Both | Osteoporosis | Mouse BMSCs, mouse model | [108] |
| miR-542-3p↓ | Insufficient to inhibit sFRP1 expression, which is a negative regulator of Wnt pathway | Both | Osteoporosis | HEK293T cells, rat BMSCs, rat model | [111] |
| miR-596↑ | Targets Smad3 to inhibit Runx2 expression and osteogenesis | In vitro | ONFH | hBMSCs | [109] |
| miR-705 and miR-3077-5p↑ | Respectively, targets HOXA10 and Runx2 mRNA, leading to MSC lineage commitment transition to adipocytes | In vitro | Osteoporosis | hBMSCs | [107] |
| miR-708↑ | Targets Smad3 to inhibit Runx2 expression | In vitro | ONFH | hBMSCs | [110] |
| miR-1263↓ | Insufficient to suppress Mob-1/YAP/Hippo signaling pathway-induced apoptosis | Both | Disused osteoporosis | hUCSCs, rat model | [114] |
| lncRNA-MALAT1↓ | Insufficient to inhibit miR-143, whose target is Osx | In vitro | Osteoporosis | hBMSCs | [117] |
| lncRNA-ORLNC1↑ | Endogenously competes with miR-296 and eliminates miR-296's suppression of Pten, which is a negative regulator of osteogenesis | Both | Osteoporosis | hBMSCs, mouse model | [119] |
| lncRNA-DEPTOR↑ | Binds to ncRNA-MEG3's promoter and reduces its function to activate BMP4 pathway | Both | Osteoporosis | Mouse BMSCs, mouse model | [120] |
| lncRNA-H19↓ | Insufficient to inhibit MSC proliferation and osteogenic differentiation via suppressing miR-19b-3p | In vitro | Inhibits osteoporosis | hBMSCs | [121] |
| lncRNA-HOTAIR↑ | Suppresses miR-17-5p to elevate Smad7 pathway | In vitro | Inhibits ONFH | hBMSCs | [122] |
ABL1: oncogene homolog 1; AIMP3: aminoacyl-tRNA synthetase-interacting multifunctional protein-3; CKS1: cyclin-dependent kinase regulatory subunit 1; Mef2c: myocyte enhancer factor 2c; LAMC1: laminin gamma 1; PRC2: polycomb repressive complex 2; SAHF: senescence-associated heterochromatin foci; sFRP1: secreted Frizzled-related protein-1; Spry1: sprouty homolog 1.
Some upregulated miRNAs maintain senescent cells in a proliferation-disabled state by binding to the transcripts of genes related to the cell cycle. For instance, miR-22 and miR-485-5p directly target the cyclin-dependent kinase regulatory subunit 1 gene, thus impeding synthesis and function of CDK and cyclin B and cause G2/M phase arrest [69]. miR-34a also targets CDK2, CDK4, CDK6, cyclin D, cyclin E, and RBP2 to hinder self-renewal ability [70]. In addition, miR-31a-5p can bind to the 3′UTR of E2F2 mRNA and bring about senescence-associated heterochromatin foci formation in aged rat BMSCs [71]. Besides, CNOT6 encodes deadenylase subunits of the Ccr4-Not complex; miR-29c-3p-induced CNOT6 downregulation can induce responsive elevation of p53, p21, and p16 expression followed by arrest of cell cycle [72, 73].
Correspondingly, with replicative pressure due to serial passages, miRNAs that play roles in repressing senescence-inducing proteins are downregulated. Downregulation of these miRNAs initially leads to dysregulation of global gene regulatory network and eventually fosters the aging process. For example, downregulation of miR-10a makes it insufficient to suppress senescence-inducing function of Krüppel-like factor 4 (KLF4) [74, 75]. And intriguingly, downregulation of miR-17 family (including miR-17, miR-20b-5p, and miR-106a-5p) regulates the activities of various genes, including Smad ubiquitination regulatory factor-1 (Smurf1), p21, CCND1, and E2F1 genes in aging [76, 77]. Besides, other downregulated miRNAs including miR-543, miR-590-3p, and miR-24a have been discovered to modulate p18/p21 and p16 activity separately [78, 79]. In addition, downregulation of miR-199b-5p also promotes cell cycle arrest indirectly via laminin gamma 1 [80].
Accumulation of reactive oxygen species (ROS) is another important mechanism of stem cell aging. Several miRNAs influence the production or elimination of ROS epigenetically. In younger cells, the amount of peroxisome is restricted by pexophagy. However, in aged cells, upregulated miR-142 targets the endothelial PAS domain protein (Epas1) gene, a positive regulator of pexophagy, inducing ROS accumulation [81]. Moreover, miR-155-5p can repress antioxidant genes by targeting the common transcription factor CCAAT/enhancer-binding protein β (C/EBP-β) gene and then induces ROS generation [82]. miR-182 antagonizes osteoblast proliferation and differentiation by targeting FoxO1 gene, which protects hBMSCs from ROS-induced harm [83]. Besides, miR-183-5p, which belongs to the same cluster to miR-182, also increases in extracellular vesicles (EVs) derived from the bone marrow of aged mice. It accelerates cell senescence by impairing HMOX-1 protein's responsive capacity to oxidative stress [84].
miRNA also affects cell aging through mitochondrial or telomere mechanisms. miR-34a targeting Sirt1 and miR-155 targeting calcium-binding protein 39 (Cab39) are all contributors in mitochondrial mechanisms [85–90]. When it comes to the telomere hypothesis of aging, miR-195 binds to the 3′UTR of Tert mRNA, thus preventing TERT protein from repairing the shortened telomeres due to replicative senescence [91].
miRNAs such as miR-27, mi-R141-3p, and miR-1292 can also induce aging through multiple cellular pathways involved in cell differentiation and metabolism. The more detailed information is listed in Table 4 [92–100].
As for lncRNAs, although they are regarded as crucial modulators of MSC-mediated ectopic tissue regeneration, only a few studies have reported the influence of lncRNAs on aging. lncRNAs regulate gene expression in diverse manners, such as serving as scaffolds to facilitate the assembly of specific transcriptional complexes or acting as sponges to reduce the availability of targeted miRNAs [101]. For instance, upregulated lncRNA-HOTAIR has been found to bring about senescence-associated changes, including differential expression of specific genes and abnormal DNA methylation by facilitating triple-helix DNA-DNA-RNA formation [102]. Besides, lncRNA-Bmncr serves as a scaffold to facilitate the interaction of Abelson murine leukemia viral oncogene homolog 1 protein and transcriptional coactivator with PDZ-binding motif (TAZ), guaranteeing the assembly of the TAZ and Runx2/Pparγ transcriptional complex to inhibit MSC adipogenic differentiation. When lncRNA-Bmncr expression decreases in aging MSCs, the tendency of lipogenesis increases [103].
2.5.2. ncRNAs in Osteoporosis
It has been widely reported that miRNAs and lncRNAs play significant roles in maintaining the balance between osteogenesis and adipogenesis. Scholars have summarized that miRNAs were involved in osteogenic regulation mainly through two patterns: affecting Runx2 expression via canonical Wnt pathway, TGF-β pathway, and BMP pathway or directly targeting Runx2 or Osx genes. On the other hand, ncRNAs influence adipogenesis through PPARγ and C/EBP-α [104, 105]. When these miRNA expression levels change and differentiation balance is broken, bone diseases such as osteoporosis will occur. For instance, it has been reported that in osteoporosis, upregulated miR-23b, miR-3077-5p, miR-212, and miR-384 inhibit osteogenesis by directly targeting Runx2 gene [106–108], while in ONFH, upregulated miR-596 and miR-708 hinder osteogenesis by binding to Smad3 transcripts to suppress Runx2 gene expression [109, 110]. In addition, miRNAs such as miR-542-3p, miR-21, and miR-27 target negative regulators in osteogenesis-related pathways; thus, their downregulation also leads to MSC lineage commitment transition from osteocytes to adipocytes [111–113]. In addition to modulating MSC differentiation, miR-181a and miR-1263 also influence skeletal homeostasis by regulating cell apoptosis through FasL accumulation and Mob-1/YAP/Hippo, respectively, [114, 115]. Besides, downregulated miR-21a is insufficient to inhibit phosphatase and tension homolog (Pten) gene expression, and abundant PTEN protein downregulates the AKT pathway to induce osteocyte apoptosis in glucocorticoid-induced osteonecrosis [116].
lncRNAs often exert their pathogenic effect via miRNAs or other ncRNAs. For example, miR-143, a direct inhibitor of Osx gene, is suppressed by lncRNA-MALAT1 in normal MSCs. However, in osteoporosis patients, downregulation of lncRNA-MALAT1 leads to decreased Osx gene expression and loss of bone mass [117]. Another team also proved that lncRNA-MALAT1 acts as a sponge of miR-34c to increase the expression of special AT-rich sequence binding protein 2 (SATB2), which is conducive to restoring osteogenesis in osteoporosis conditions [118]. Besides, elevated lncRNA-ORLNC1 endogenously competes with miR-296 and eliminates miR-296's suppression of Pten gene, which is a negative regulator of osteogenesis [119]. Moreover, in ONFH, upregulated lncRNA-DEPTOR binds to lncRNA-MEG3 promoter and prevents it from activating BMP4 pathway [120]. It is worth mentioning that in the process of skeletal diseases, alteration of lncRNA expression may work against bone destruction. For example, lncRNA-H19 inhibits MSC proliferation and osteogenic differentiation via suppressing miR-196-3p when estrogen exists. However, in postmenopausal osteoporosis, H19 expression level decreases [121]. Similarly, in ONFH, lncRNA-HOTAIR is upregulated and promotes osteogenesis via the miR-17-5p/Smad7 pathway [122].
In general, existing research reveals that in terms of mechanism, ncRNAs primarily regulate cell cycle or affect aging-related factors including ROS, telomere, and mitochondria to induce MSC aging. In the process of skeletal diseases, epigenetic factors promote disease progression through biased differentiation and cell apoptosis. Specifically speaking, the dysregulation of miRNA and lncRNA forecasts the dysfunction of MSCs, and notably, modulative effects of ncRNAs are not directly realized in a unidirectional manner.
2.6. The Interplay of Different Epigenetic Factors in MSC Aging
As we mentioned in the previous parts, MSC aging is epigenetically marked by heterochromatin loss, altered DNA methylation profile, and organized histone modification. Actually, it should be particularly noted that epigenetic factors do not perform their functions in a parallel and independent fashion. It has been extensively reported that a multitude of them mutually intertwine and influence. Firstly, DNA methylation and histone modification are closely related in many aspects. In aged MSCs, hypomethylated CpG sites always come together with H3K4me, and both of them are signs of increased transcription activity, while DNA regions enriched in hypermethylated CpG sites mainly overlap with the repressive chromatin marks H3K27me3 and H3K9me3, synergistically inhibiting transcription activity [22, 25]. Moreover, DNA demethylation-induced PcG downregulation can regulate H3K27 methylation [28]. Secondly, histone modifications can alter chromatin structure not only by influencing histone-DNA, histone-histone interactions, and chaperone-histone binding [60] but also through modulating the synthesis of chromatin remodeling-related proteins such as condensin [56]. Last, DNA methylation also influences chromatin structure by regulating transcription factors, such as SMARCs and SIN3A [24]. Besides, it has been discovered that considerable ncRNAs interplay with complicated molecular networks and pretranscriptional epigenetic marks in the aging process. For instance, depletion of DNA demethylase TET causes accumulated miR-297; miR-188 inhibits HDAC9-mediated histone deacetylation; and miR-31a-5p can bring about senescence-associated heterochromatin foci formation in aged rat BMSCs [34, 71, 95]. Pursuit in epigenetics about MSCs has never slowed down. It is believable and desirable that more interactive relationships between different epigenetic marks could be established and elaborated. However, according to existing researches and experimental technology, it is difficult and one-sided for us to rank a certain epigenetic mark as a more critical inducing factor for MSC aging. Instead, it is most likely that they function as an interlaced system in coordination and order. Although it has been reported that intervention of a single factor of epigenetic marks could delay or reverse MSC aging to some extent, we believe that other types of accompanied epigenetic regulation may initiate in the biological effect, and even imaginably, proper combinational modulation of two or more epigenetic marks could upgrade the efficacy of therapy for MSC-related diseases.
3. Application of Epigenetic Regulation in Skeletal Diseases and Engineering Regeneration
MSCs are an essential source for cell-based bone regeneration. The premise of bone regeneration is to maintain MSC stemness and promote their osteogenic differentiation. As mentioned above, it has been demonstrated that epigenetic markers and modifiers play fundamental roles in aging and diseases through modulating MSC function. Thus, MSC-mediated therapeutic or regenerative strategies based on epigenetic principle possess enormous potential in the treatment of aging-related bone disorders and defects. Despite the lack of experiments that mediate bone regeneration by means of epigenetic regulation in aging models, great efforts have been made in corresponding explorations with normal animal models. In turn, tentative application of epigenetic therapies in vivo further reinforces our understanding of the intrinsic mechanisms and makes it possible to realize our clinic utilization in the future. So far, there are three kinds of commonly used epigenetic interventions of tissue diseases or regeneration in vivo: exogenous inhibition of negative epigenetic regulators, exogenous supplement of positive regulators, and direct gene manipulation (Figure 2).
Figure 2.
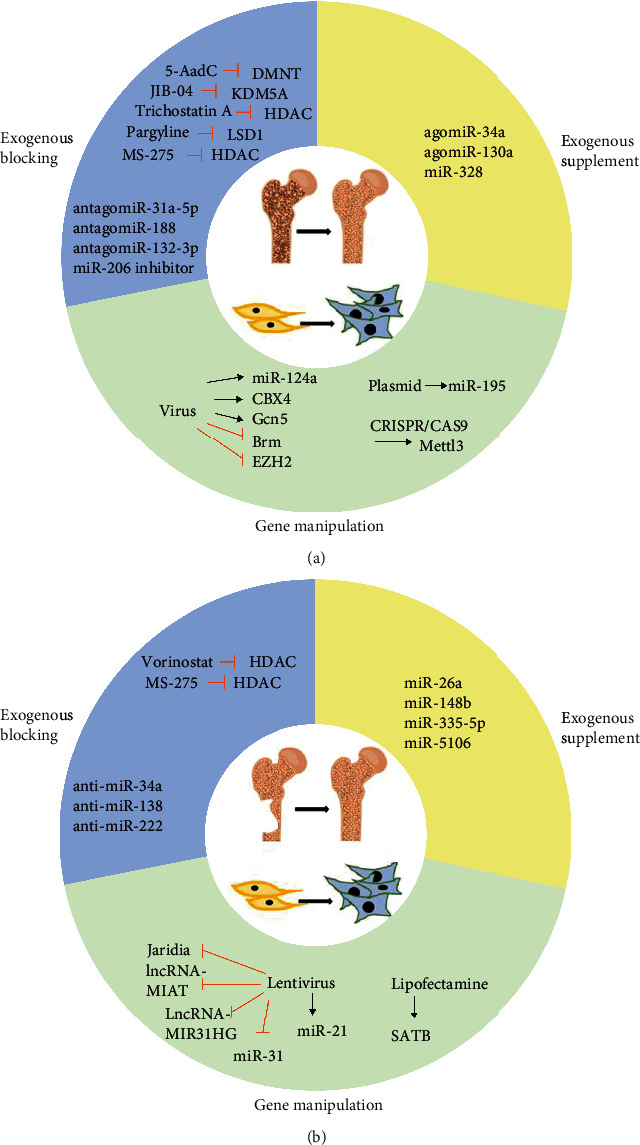
Application of epigenetic regulation in (a) skeletal diseases and (b) bone regeneration. Exogenous blocking or supplement and direct gene manipulation are separately used in both conditions.
3.1. Exogenous Blocking
One common method is to block epigenetic factor-induced MSC aging and diseases by using exogenous inhibitors. For bone tissue regeneration in nonaging animal models, a combination of collagen sponge and HDAC1/4 inhibitor MS-275 exerts a promotive effect in rat critical-sized calvarial defect healing [123]. Moreover, intraperitoneal injection of MS-275 avoids delayed cranial suture closure in Runx2-null mice [124]. Similarly, when vorinostat, another HDAC1 inhibitor, was intraperitoneally injected into mice, the number of osteoblasts in endocortical bone increased and OCN level in serum rose [125]. Except for that, miRNA-mediated bone regeneration usually proceeds with specific biomaterials [126]. For example, when hMSCs transfected with anti-miR-34a, anti-miR-138, or anti-miR-222 by lipofectamine are loaded on a hydroxyapatite/tricalcium phosphate ceramic powder, the ectopic bone formation on complex scaffolds increases compared to the untransfected group [127, 128]. Moreover, anti-miR-222 also manifests a promotive effect toward bone defect healing when directly loaded on the atelocollagen scaffold [129].
Simultaneously, great progress has been made in MSC-dependent epigenetic therapeutics aimed at aging-related skeletal diseases. For histone acetylation, MS-275 subcutaneous injection rescues NF-κB-induced rapid bone loss by interrupting interactions between HDACs and DExH-box helicase Dhx36, which inhibits tissue-nonspecific alkaline phosphatase (TNAP) activity [123]. Pretreatment with KDM5A inhibitor JIB-04 partially rescues bone loss during osteoporosis by increasing the H3K4me3 level on the Runx2 promoter [45]. And HDAC inhibitor trichostatin prospers osteogenesis of rat adipose tissue-derived stem cells (hADSCs) by histone modifying on Runx2 promoter [130]; LSD1 inhibitor pargyline rescues osteogenic ability of BMSCs under osteoporosis conditions by modulating H3K4 methylation at the promoter region of Ocn and Runx2 genes [131]. Notably, miRNA inhibitors are also applied in osteoporosis treatment. For example, injection of antagomiR-31a-5p or antagomiR-188 into bone marrow cavity significantly alleviates fat accumulation and remedies bone loss in aged mice [71, 95]. Moreover, atagomiR-132-3p delivered by a bone-targeted (AspSerSer)6-cationic liposome system silences miRNA-132-3p expression in bone tissues, thus effectively preserving bone mass, bone structure, and strength in hindlimb-unloaded mice [132].
3.2. Exogenous Supplement
ncRNAs or their mimics can be exogenously supplemented to delay or reverse disease progression. It has been extensively reported that loading MSCs transfected by proosteogenic miRNAs (such as miR-26a, miR-148b, miR-5106, miR-335-5p, or their mimics) on biomaterials is an effective strategy to promote bone regeneration [133–136].
With regard to skeletal disease treatment, agomiR-130a intravenously injection reduces bone loss in elderly mice by targeting Smurf2 and Pparγ genes [137, 138], while collagen-based hydrogel containing agomir-34a elevates bone volume in mouse radiational bone injury area by downregulating Notch1 expression in BMSCs [139]. In addition, miR-328 is the antagonist of Axin1 gene, whose product AXIN1 protein is an inhibitor of Wnt signaling pathway. Thus, application of apoptotic bodies containing miR-328 significantly ameliorates osteopenia in OVX mice [140].
3.3. Gene Manipulation
In addition to the above two methods, virus transfection and CRISPR/Cas9 are applied to realize direct gene manipulation. Jumonji AT-rich interactive domain 1A (JARID1A) protein is a KDM5A component participating in Runx2-related H3K4 demethylation. Compared with the control group, scaffolds containing BMSCs transduced with si-Jarid1a increase bone volume and mineral density during the process of calvarial defect healing [141]. Besides, SATB2 protein is a nuclear matrix protein involved in chromatin remodeling, and the Satb2 gene overexpression by lipofectamine transfection enhances skeletal tissue regeneration and mineralization in mouse mandibular bone defects [142]. Moreover, anti-miR-31-expressing BMSCs/poly (glycerol sebacate) complex and miRNA-21-modified BMSCs/β-tricalcium phosphate composite both bring higher bone regeneration rate in rat bone defects [143, 144]. Similarly, knockdown lncRNA MIR31HG or MIAT with lentivirus significantly enhances ADSCs' bone formation capacity when implanted subcutaneously with biomaterials [145, 146].
As to therapeutic application in diseases, injection of lentiviruses encoding CBX4 protein into the joint capsules leads to upregulation of proliferation, bone growth-associated genes, and downregulation of inflammation and cell death-related genes [58]. Similarly, mammalian brahma (BRM) protein is a component of SWI/SNF complex with ATPase activity. Knockdown of Brm gene in mice helps it resist aging-related osteoporosis and reduces adiposity in bone marrow [147]. Lentivirus is also used to alter histone acetylation and methylation level in osteoporosis. In OVX mice, injection of lentiviruses expressing Gcn5 gene restores endogenous BMSC osteogenic potential by increasing H3K9ac on the promoters of Wnt genes [48]. Except for that, knockdown of Ezh2 gene by lentivirus-expressing shRNA decreases H3K27me3 on Wnt genes, reversing the abnormal MSC adipogenic lineage commitment in osteoporosis [47]. Moreover, when RNA N6-methyltransferase Mettl3 gene is knocked in transplanted MSCs with CRISPR/Cas9 and Cre/LoxP, mice are protected from OVX-induced osteoporosis [63]. When it comes to ncRNAs, bone defects completely healed with transplantation of BMSCs expressing miR-214 sponges transduced by baculovirus [148].
4. Conclusion
Epigenetic regulation of MSCs occurs in several steps of transcription, including chromatin remodeling, DNA methylation, and histone modification, and posttranscription, including mRNA processing and ncRNA regulation. Epigenetic markers and modifiers have been proved to play indispensable roles in MSC aging and fundamental homeostasis in vivo, both of which are related to the pathogenesis of tissue disorders in aging and diseases. Initial experimental attempts, roughly according to epigenetic clues, have been carried out to delay MSC aging or rejuvenate senescent MSCs, which is aimed at enhancing their self-renewal capacity and correct biased differentiation lineage. However, there remain several obstacles for translational application, including lack of sequential identification of spatiotemporal epigenetic alteration, and difficulties in precise translational intervention in vivo. Hopefully, many revolutionary technological progresses emerged just in the past years, including single-cell epigenomic analysis and CRISPR/Cas9, cell transplantation, and regenerative biomaterials. In this context, therapeutic or regenerative strategies based on epigenetic regulation of MSC aging stand a tremendous chance to restore MSC homeostasis in vivo and even boost translational application in tissue regeneration, especially among the elderly or people with bone diseases.
Acknowledgments
This work was supported by the projects of Ten-Thousand Talents Program (Y.L.), National Natural Science Foundation of China (No. 81871492 (Y.L.) and No. 81670957 (WR.L.)), and Beijing Municipal Natural Science Foundation (No. L182005 (Y.L.)).
Contributor Information
Yan Liu, Email: orthoyan@bjmu.edu.cn.
Weiran Li, Email: weiranli2003@163.com.
Conflicts of Interest
The authors declare no conflict of interest.
Authors' Contributions
Ruoxi Wang and Yu Wang contributed equally to this work.
References
- 1.Kolf C. M., Cho E., Tuan R. S. Mesenchymal stromal cells. Biology of adult mesenchymal stem cells: regulation of niche, self-renewal and differentiation. Arthritis Research & Therapy. 2007;9(1):p. 204. doi: 10.1186/ar2116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Pittenger M. F., Mackay A. M., Beck S. C., et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 3.Friedenstein A. J., Chailakhyan R. K., Latsinik N. V., Panasyuk A. F., Keiliss-Borok I. V. Stromal cells responsible for transferring the microenvironment of the hemopoietic tissues. Cloning in vitro and retransplantation in vivo. Transplantation. 1974;17(4):331–340. doi: 10.1097/00007890-197404000-00001. [DOI] [PubMed] [Google Scholar]
- 4.Mushahary D., Spittler A., Kasper C., Weber V., Charwat V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytometry. Part A. 2018;93(1):19–31. doi: 10.1002/cyto.a.23242. [DOI] [PubMed] [Google Scholar]
- 5.Wang C. H., Wang T. M., Young T. H., Lai Y. K., Yen M. L. The critical role of ECM proteins within the human MSC niche in endothelial differentiation. Biomaterials. 2013;34(17):4223–4234. doi: 10.1016/j.biomaterials.2013.02.062. [DOI] [PubMed] [Google Scholar]
- 6.Brückner S., Tautenhahn H. M., Winkler S., et al. Isolation and hepatocyte differentiation of mesenchymal stem cells from porcine bone marrow—“surgical waste” as a novel MSC source. Transplantation Proceedings. 2013;45(5):2056–2058. doi: 10.1016/j.transproceed.2013.01.101. [DOI] [PubMed] [Google Scholar]
- 7.Zacharek A., Chen J., Li A., et al. Angiopoietin1/Tie2 and VEGF/Flk1 induced by MSC treatment amplifies angiogenesis and vascular stabilization after stroke. Journal of Cerebral Blood Flow & Metabolism. 2007;27(10):1684–1691. doi: 10.1038/sj.jcbfm.9600475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bessout R., Sémont A., Demarquay C., Charcosset A., Benderitter M., Mathieu N. Mesenchymal stem cell therapy induces glucocorticoid synthesis in colonic mucosa and suppresses radiation-activated T cells: new insights into MSC immunomodulation. Mucosal Immunology. 2014;7(3):656–669. doi: 10.1038/mi.2013.85. [DOI] [PubMed] [Google Scholar]
- 9.Peng K. Y., Liu Y. H., Li Y. W., Yen B. L., Yen M. L. Extracellular matrix protein laminin enhances mesenchymal stem cell (MSC) paracrine function through αvβ3/CD61 integrin to reduce cardiomyocyte apoptosis. Jounal of Cellular and Molecular Medicine. 2017;21(8):1572–1583. doi: 10.1111/jcmm.13087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Turinetto V., Vitale E., Giachino C. Senescence in human mesenchymal stem cells: functional changes and implications in stem cell-based therapy. International Journal of Molecular Science. 2016;17(7):p. 1164. doi: 10.3390/ijms17071164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sui B. D., Zheng C. X., Li M., Jin Y., Hu C. H. Epigenetic regulation of mesenchymal stem cell homeostasis. Trends in Cell Biology. 2019;30(2):97–166. doi: 10.1016/j.tcb.2019.11.006. [DOI] [PubMed] [Google Scholar]
- 12.Shyh-Chang N., Daley G. Q., Cantley L. C. Stem cell metabolism in tissue development and aging. Development (Cambridge, England) 2013;140(12):2535–2547. doi: 10.1242/dev.091777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Almalki S. G., Agrawal D. K. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation; Research in Biological Diversity. 2016;92(1-2):41–51. doi: 10.1016/j.diff.2016.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Campisi J., di Fagagna F. d.' A. Cellular senescence: when bad things happen to good cells. Nature Reviews Molecular Cell Biology. 2007;8(9):729–740. doi: 10.1038/nrm2233. [DOI] [PubMed] [Google Scholar]
- 15.Yu B., Wang C. Y. Osteoporosis: the result of an 'aged' bone microenvironment. Trends in Cell Biology. 2016;22(8):641–644. doi: 10.1016/j.molmed.2016.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Avgustinova A., Benitah S. A. Epigenetic control of adult stem cell function. Nature Reviews Molecular Cell Biology. 2016;17(10):643–658. doi: 10.1038/nrm.2016.76. [DOI] [PubMed] [Google Scholar]
- 17.Ozkul Y., Galderisi U. The impact of epigenetics on mesenchymal stem cell biology. Journal of Cellular Physiology. 2016;231(11):2393–2401. doi: 10.1002/jcp.25371. [DOI] [PubMed] [Google Scholar]
- 18.Han J. W., Yoon Y. S. Epigenetic landscape of pluripotent stem cells. Antioxidants & Redox Signaling. 2012;17(2):205–223. doi: 10.1089/ars.2011.4375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Luo C., Hajkova P., Ecker J. R. Dynamic DNA methylation:in the right place at the right time. Science. 2018;361(6409):1336–1340. doi: 10.1126/science.aat6806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wu H., Zhang Y. Reversing DNA methylation: mechanisms, genomics, and biological functions. Cell. 2014;156(1-2):45–68. doi: 10.1016/j.cell.2013.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bork S., Pfister S., Witt H., et al. DNA methylation pattern changes upon long-term culture and aging of human mesenchymal stromal cells. Aging Cell. 2010;9(1):54–63. doi: 10.1111/j.1474-9726.2009.00535.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fernández A. F., Bayón G. F., Urdinguio R. G., et al. H3K4me1 marks DNA regions hypomethylated during aging in human stem and differentiated cells. Genome Research. 2015;25(1):27–40. doi: 10.1101/gr.169011.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bentivegna A., Roversi G., Riva G., et al. The effect of culture on human bone marrow mesenchymal stem cells: focus on dna methylation profiles. Stem Cells International. 2016;2016:12. doi: 10.1155/2016/5656701.5656701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pasumarthy K. K., Doni Jayavelu N., Kilpinen L., et al. Methylome analysis of human bone marrow MSCs reveals extensive age- and culture-induced changes at distal regulatory elements. Stem Cell Reports. 2017;9(3):999–1015. doi: 10.1016/j.stemcr.2017.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Schellenberg A., Lin Q., Schüler H., et al. Replicative senescence of mesenchymal stem cells causes DNA-methylation changes which correlate with repressive histone marks. Aging. 2011;3(9):873–888. doi: 10.18632/aging.100391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zheng Y., He L., Wan Y., Song J. H3K9me-enhanced DNA hypermethylation of the p16INK4a gene: an epigenetic signature for spontaneous transformation of rat mesenchymal stem cells. Stem Cells and Development. 2013;22(2):256–267. doi: 10.1089/scd.2012.0172. [DOI] [PubMed] [Google Scholar]
- 27.Toraño E. G., Bayón G. F., del Real Á., et al. Age-associated hydroxymethylation in human bone-marrow mesenchymal stem cells. Journal of Translational Medicine. 2016;14(1):p. 207. doi: 10.1186/s12967-016-0966-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.So A. Y., Jung J. W., Lee S., Kim H. S., Kang K. S. DNA methyltransferase controls stem cell aging by regulating BMI1 and EZH2 through microRNAs. PloS One. 2011;6(5):p. e19503. doi: 10.1371/journal.pone.0019503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kornicka K., Marycz K., Maredziak M., Tomaszewski K. A., Nicpon J. The effects of the DNA methyltranfserases inhibitor 5-azacitidine on ageing, oxidative stress and DNA methylation of adipose derived stem cells. Journal of Cellurlar and Molecular Medicine. 2017;21(2):387–401. doi: 10.1111/jcmm.12972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Oh Y. S., Jeong S. G., Cho G. W. Anti-senescence effects of DNA methyltransferase inhibitor RG108 in human bone marrow mesenchymal stromal cells. Biotechnology and Applied Biochemistry. 2015;62(5):583–590. doi: 10.1002/bab.1393. [DOI] [PubMed] [Google Scholar]
- 31.Tsai C. C., Su P. F., Huang Y. F., Yew T. L., Hung S. C. Oct4 and Nanog directly regulate Dnmt1 to maintain self-renewal and undifferentiated state in mesenchymal stem cells. Molecular Cell. 2012;47(2):169–182. doi: 10.1016/j.molcel.2012.06.020. [DOI] [PubMed] [Google Scholar]
- 32.Zhou G. S., Zhang X. L., Wu J. P., et al. 5-Azacytidine facilitates osteogenic gene expression and differentiation of mesenchymal stem cells by alteration in DNA methylation. Cytotechnology. 2009;60(1-3):11–22. doi: 10.1007/s10616-009-9203-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sun Z., Yang S., Ye S., et al. Aberrant CpG islands' hypermethylation of ABCB1 in mesenchymal stem cells of patients with steroid-associated osteonecrosis. The Journal of Rheumatology. 2013;40(11):1913–1920. doi: 10.3899/jrheum.130191. [DOI] [PubMed] [Google Scholar]
- 34.Yang R., Yu T., Kou X., et al. Tet1 and Tet2 maintain mesenchymal stem cell homeostasis via demethylation of the P2rX7 promoter. Nature Communications. 2018;9(1):p. 2143. doi: 10.1038/s41467-018-04464-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Portela A., Esteller M. Epigenetic modifications and human disease. Nature Biotechnology. 2010;28(10):1057–1068. doi: 10.1038/nbt.1685. [DOI] [PubMed] [Google Scholar]
- 36.Barnes C. E., English D. M., Cowley S. M. Acetylation & co: an expanding repertoire of histone acylations regulates chromatin and transcription. Essays in Biochemistry. 2019;63(1):97–107. doi: 10.1042/EBC20180061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Brand M., Nakka K., Zhu J., Dilworth F. J. Polycomb/trithorax antagonism: cellular memory in stem cell fate and function. Cell Stem Cell. 2019;24(4):518–533. doi: 10.1016/j.stem.2019.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jung J. W., Lee S., Seo M. S., et al. Histone deacetylase controls adult stem cell aging by balancing the expression of polycomb genes and jumonji domain containing 3. Cellular and Molecular Life Sciences. 2010;67(7):1165–1176. doi: 10.1007/s00018-009-0242-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cakouros D., Isenmann S., Cooper L., et al. Twist-1 induces Ezh2 recruitment regulating histone methylation along the Ink4A/Arf locus in mesenchymal stem cells. Molecular and Cellular Biology. 2012;32(8):1433–1441. doi: 10.1128/MCB.06315-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lee S., Park J. R., Seo M. S., et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Proliferation. 2009;42(6):711–720. doi: 10.1111/j.1365-2184.2009.00633.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang Y., Chen T., Yan H., et al. Role of histone deacetylase inhibitors in the aging of human umbilical cord mesenchymal stem cells. Journal of Cellular Biochemistry. 2013;114(10):2231–2239. doi: 10.1002/jcb.24569. [DOI] [PubMed] [Google Scholar]
- 42.Pan H., Guan D., Liu X., et al. SIRT6 safeguards human mesenchymal stem cells from oxidative stress by coactivating NRF2. Cell Research. 2016;26(2):190–205. doi: 10.1038/cr.2016.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ren J., Huang D., Li R., Wang W., Zhou C. Control of mesenchymal stem cell biology by histone modifications. Cell and Bioscience. 2020;10:p. 11. doi: 10.1186/s13578-020-0378-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Deng P., Chen Q. M., Hong C., Wang C. Y. Histone methyltransferases and demethylases: regulators in balancing osteogenic and adipogenic differentiation of mesenchymal stem cells. International Journal of Oral Science. 2015;7(4):197–204. doi: 10.1038/ijos.2015.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wang C., Wang J., Li J., et al. KDM5A controls bone morphogenic protein 2-induced osteogenic differentiation of bone mesenchymal stem cells during osteoporosis. Cell Death & Disease. 2016;7(8, article e2335) doi: 10.1038/cddis.2016.238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yin B., Yu F., Wang C., Li B., Liu M., Ye L. Epigenetic control of mesenchymal stem cell fate decision via histone methyltransferase ASH1L. Stem Cells (Dayton, Ohio) 2019;37(1):115–127. doi: 10.1002/stem.2918. [DOI] [PubMed] [Google Scholar]
- 47.Jing H., Liao L., An Y., et al. Suppression of EZH2 prevents the shift of osteoporotic MSC fate to adipocyte and enhances bone formation during osteoporosis. Molecular Therapy: The Journal of the American Society of Gene Therapy. 2016;24(2):217–229. doi: 10.1038/mt.2015.152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jing H., Su X., Gao B., et al. Epigenetic inhibition of Wnt pathway suppresses osteogenic differentiation of BMSCs during osteoporosis. Cell Death & Disease. 2018;9(2):p. 176. doi: 10.1038/s41419-017-0231-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhang P., Liu Y., Jin C., et al. Histone H3K9 acetyltransferase PCAF is essential for osteogenic differentiation through bone morphogenetic protein signaling and may be involved in osteoporosis. Stem Cells (Dayton, Ohio) 2016;34(9):2332–2341. doi: 10.1002/stem.2424. [DOI] [PubMed] [Google Scholar]
- 50.Pérez-Campo F. M., Riancho J. A. Epigenetic mechanisms regulating mesenchymal stem cell differentiation. Current Genomics. 2015;16(6):368–383. doi: 10.2174/1389202916666150817202559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fan Z., Yamaza T., Lee J. S., et al. BCOR regulates mesenchymal stem cell function by epigenetic mechanisms. Nature Cell Biology. 2009;11(8):1002–1009. doi: 10.1038/ncb1913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hota S. K., Bruneau B. G. ATP-dependent chromatin remodeling during mammalian development. Development (Cambridge, England) 2016;143(16):2882–2897. doi: 10.1242/dev.128892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Squillaro T., Severino V., Alessio N., et al. De-regulated expression of the BRG1 chromatin remodeling factor in bone marrow mesenchymal stromal cells induces senescence associated with the silencing of NANOG and changes in the levels of chromatin proteins. Cell Cycle. 2015;14(8):1315–1326. doi: 10.4161/15384101.2014.995053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Alessio N., Squillaro T., Cipollaro M., Bagella L., Giordano A., Galderisi U. The BRG1 ATPase of chromatin remodeling complexes is involved in modulation of mesenchymal stem cell senescence through RB-P53 pathways. Oncogene. 2010;29(40):5452–5463. doi: 10.1038/onc.2010.285. [DOI] [PubMed] [Google Scholar]
- 55.Napolitano M. A., Cipollaro M., Cascino A., Melone M. A. B., Giordano A., Galderisi U. Brg1 chromatin remodeling factor is involved in cell growth arrest, apoptosis and senescence of rat mesenchymal stem cells. Journal of Cell Science. 2007;120(16):2904–2911. doi: 10.1242/jcs.004002. [DOI] [PubMed] [Google Scholar]
- 56.Huang B., Wang B., Yuk-Wai Lee W., et al. KDM3A and KDM4C regulate mesenchymal stromal cell senescence and bone aging via condensin-mediated heterochromatin reorganization. iScience. 2019;21:375–390. doi: 10.1016/j.isci.2019.10.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zhang W., Li J., Suzuki K., et al. Aging stem cells. A Werner syndrome stem cell model unveils heterochromatin alterations as a driver of human aging. Science. 2015;348(6239):1160–1163. doi: 10.1126/science.aaa1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ren X., Hu B., Song M., et al. Maintenance of nucleolar homeostasis by CBX4 alleviates senescence and osteoarthritis. Cell Reports. 2019;26(13):3643–3656.e7. doi: 10.1016/j.celrep.2019.02.088. [DOI] [PubMed] [Google Scholar]
- 59.Zhou C., Zou J., Zou S., Li X. INO80 is required for osteogenic differentiation of human mesenchymal stem cells. Scientific Reports. 2016;6(1, article 35924) doi: 10.1038/srep35924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tessarz P., Kouzarides T. Histone core modifications regulating nucleosome structure and dynamics. Nature Reviews Molecular Cell Biology. 2014;15(11):703–708. doi: 10.1038/nrm3890. [DOI] [PubMed] [Google Scholar]
- 61.Chen X., Hua W., Huang X., Chen Y., Zhang J., Li G. Regulatory role of RNA N(6)-methyladenosine modification in bone biology and osteoporosis. Frontiers in Endocrinology. 2019;10:p. 911. doi: 10.3389/fendo.2019.00911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Shen G. S., Zhou H. B., Zhang H., et al. The GDF11-FTO-PPARγ axis controls the shift of osteoporotic MSC fate to adipocyte and inhibits bone formation during osteoporosis. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 2018;1864(12):3644–3654. doi: 10.1016/j.bbadis.2018.09.015. [DOI] [PubMed] [Google Scholar]
- 63.Wu Y., Xie L., Wang M., et al. Mettl3-mediated m6A RNA methylation regulates the fate of bone marrow mesenchymal stem cells and osteoporosis. Nature Communications. 2018;9(1, article 4772) doi: 10.1038/s41467-018-06898-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kilpinen L., Parmar A., Greco D., et al. Expansion induced microRNA changes in bone marrow mesenchymal stromal cells reveals interplay between immune regulation and cell cycle. Aging. 2016;8(11):2799–2813. doi: 10.18632/aging.101088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lei Q., Liu T., Gao F., et al. Microvesicles as potential biomarkers for the identification of senescence in human mesenchymal stem cells. Theranostics. 2017;7(10):2673–2689. doi: 10.7150/thno.18915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wagner W., Horn P., Castoldi M., et al. Replicative senescence of mesenchymal stem cells: a continuous and organized process. PloS One. 2008;3(5, article e2213) doi: 10.1371/journal.pone.0002213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Meng X., Xue M., Xu P., Hu F., Sun B., Xiao Z. MicroRNA profiling analysis revealed different cellular senescence mechanisms in human mesenchymal stem cells derived from different origin. Genomics. 2017;109(3-4):147–157. doi: 10.1016/j.ygeno.2017.02.003. [DOI] [PubMed] [Google Scholar]
- 68.Hackl M., Brunner S., Fortschegger K., et al. miR-17, miR-19b, miR-20a, and miR-106a are down-regulated in human aging. Aging Cell. 2010;9(2):291–296. doi: 10.1111/j.1474-9726.2010.00549.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Aung S. W., Abu Kasim N. H., Shamsuddin S. A. A., Ramasamy T. S. MicroRNAomic transcriptomic analysis reveal deregulation of clustered cellular functions in human mesenchymal stem cells during in vitro passaging. Stem Cell Reviews and Reports. 2020;16(1):222–238. doi: 10.1007/s12015-019-09924-0. [DOI] [PubMed] [Google Scholar]
- 70.Park H., Park H., Pak H. J., et al. miR-34a inhibits differentiation of human adipose tissue-derived stem cells by regulating cell cycle and senescence induction. Differentiation; Research in Biological Diversity. 2015;90(4-5):91–100. doi: 10.1016/j.diff.2015.10.010. [DOI] [PubMed] [Google Scholar]
- 71.Xu R., Shen X., Si Y., et al. MicroRNA-31a-5p from aging BMSCs links bone formation and resorption in the aged bone marrow microenvironment. Aging Cell. 2018;17(4, article e12794) doi: 10.1111/acel.12794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shang J., Yao Y., Fan X., et al. miR-29c-3p promotes senescence of human mesenchymal stem cells by targeting CNOT6 through p53-p21 and p16-pRB pathways. Biochimica et Biophysica Acta. 2016;1863(4):520–532. doi: 10.1016/j.bbamcr.2016.01.005. [DOI] [PubMed] [Google Scholar]
- 73.Mittal S., Aslam A., Doidge R., Medica R., Winkler G. S. The Ccr4a (CNOT6) and Ccr4b (CNOT6L) deadenylase subunits of the human Ccr4-not complex contribute to the prevention of cell death and senescence. Molecular Biology of the Cell. 2011;22(6):748–758. doi: 10.1091/mbc.e10-11-0898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Dong J., Zhang Z., Huang H., et al. miR-10a rejuvenates aged human mesenchymal stem cells and improves heart function after myocardial infarction through KLF4. Stem Cell Research & Therapy. 2018;9(1):p. 151. doi: 10.1186/s13287-018-0895-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Li J., Dong J., Zhang Z. H., et al. miR-10a restores human mesenchymal stem cell differentiation by repressing KLF4. Journal of Cellular Physiology. 2013;228(12):2324–2336. doi: 10.1002/jcp.24402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Liu W., Qi M., Konermann A., Zhang L., Jin F., Jin Y. The p53/miR-17/Smurf1 pathway mediates skeletal deformities in an age-related model via inhibiting the function of mesenchymal stem cells. Aging. 2015;7(3):205–218. doi: 10.18632/aging.100728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Tai L., Huang C. J., Choo K. B., Cheong S. K., Kamarul T. Oxidative stress down-regulates MiR-20b-5p, MiR-106a-5p and E2F1 expression to suppress the G1/S transition of the cell cycle in multipotent stromal cells. International Journal of Medical Science. 2020;17(4):457–470. doi: 10.7150/ijms.38832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lee S., Yu K. R., Ryu Y. S., et al. miR-543 and miR-590-3p regulate human mesenchymal stem cell aging via direct targeting of AIMP3/p18. Age. 2014;36(6, article 9724) doi: 10.1007/s11357-014-9724-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Philipot D., Guérit D., Platano D., et al. p16INK4a and its regulator miR-24 link senescence and chondrocyte terminal differentiation-associated matrix remodeling in osteoarthritis. Arthritis Research & Therapy. 2014;16(1):p. R58. doi: 10.1186/ar4494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Yoo J. K., Kim C. H., Jung H. Y., Lee D. R., Kim J. K. Discovery and characterization of miRNA during cellular senescence in bone marrow-derived human mesenchymal stem cells. Experimental Gerontology. 2014;58:139–145. doi: 10.1016/j.exger.2014.07.020. [DOI] [PubMed] [Google Scholar]
- 81.Houri K., Mori T., Onodera Y., et al. miR-142 induces accumulation of reactive oxygen species (ROS) by inhibiting pexophagy in aged bone marrow mesenchymal stem cells. Scientific Reports. 2020;10(1):p. 3735. doi: 10.1038/s41598-020-60346-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Onodera Y., Teramura T., Takehara T., Obora K., Mori T., Fukuda K. miR-155 induces ROS generation through downregulation of antioxidation-related genes in mesenchymal stem cells. Aging Cell. 2017;16(6):1369–1380. doi: 10.1111/acel.12680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kim K. M., Park S. J., Jung S. H., et al. miR-182 is a negative regulator of osteoblast proliferation, differentiation, and skeletogenesis through targeting FoxO1. Journal of Bone and Mineral Research. 2012;27(8):1669–1679. doi: 10.1002/jbmr.1604. [DOI] [PubMed] [Google Scholar]
- 84.Davis C., Dukes A., Drewry M., et al. MicroRNA-183-5p increases with age in bone-derived extracellular vesicles, suppresses bone marrow stromal (stem) cell proliferation, and induces stem cell senescence. Tissue Engineering Part A. 2017;23(21-22):1231–1240. doi: 10.1089/ten.tea.2016.0525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kim Y. J., Hwang S. H., Lee S. Y., et al. miR-486-5p induces replicative senescence of human adipose tissue-derived mesenchymal stem cells and its expression is controlled by high glucose. Stem Cells and Development. 2012;21(10):1749–1760. doi: 10.1089/scd.2011.0429. [DOI] [PubMed] [Google Scholar]
- 86.Hong Y., He H., Jiang G., et al. miR-155-5p inhibition rejuvenates aged mesenchymal stem cells and enhances cardioprotection following infarction. Aging Cell. 2020;19(4, article e13128) doi: 10.1111/acel.13128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Tsujimoto T., Mori T., Houri K., et al. miR-155 inhibits mitophagy through suppression of BAG5, a partner protein of PINK1. Biochemical and Biophysical Research and Communications. 2020;523(3):707–712. doi: 10.1016/j.bbrc.2020.01.022. [DOI] [PubMed] [Google Scholar]
- 88.Kujoth G. C., Hiona A., Pugh T. D., et al. Mitochondrial DNA mutations, oxidative stress, and apoptosis in mammalian aging. Science (New York, N.Y.) 2005;309(5733):481–484. doi: 10.1126/science.1112125. [DOI] [PubMed] [Google Scholar]
- 89.Zhang F., Cui J., Liu X., et al. Roles of microRNA-34a targeting SIRT1 in mesenchymal stem cells. Stem Cell Research & Therapy. 2015;6(1):p. 195. doi: 10.1186/s13287-015-0187-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Fulzele S., Mendhe B., Khayrullin A., et al. Muscle-derived miR-34a increases with age in circulating extracellular vesicles and induces senescence of bone marrow stem cells. Aging. 2019;11(6):1791–1803. doi: 10.18632/aging.101874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Okada M., Kim H. W., Matsu-ura K., Wang Y. G., Xu M., Ashraf M. Abrogation of age-induced microRNA-195 rejuvenates the senescent mesenchymal stem cells by reactivating telomerase. Stem Cells (Dayton, Ohio) 2016;34(1):148–159. doi: 10.1002/stem.2211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Yu K. R., Lee S., Jung J. W., et al. MicroRNA-141-3p plays a role in human mesenchymal stem cell aging by directly targeting ZMPSTE24. Journal of Cell Science. 2013;126(23):5422–5431. doi: 10.1242/jcs.133314. [DOI] [PubMed] [Google Scholar]
- 93.Li Z., Ge X., Lu J., et al. MiR-141-3p regulates proliferation and senescence of stem cells from apical papilla by targeting YAP. Experimental Cell Research. 2019;383(2, article 111562) doi: 10.1016/j.yexcr.2019.111562. [DOI] [PubMed] [Google Scholar]
- 94.Fafián-Labora J., Morente-López M., Sánchez-Dopico M. J., et al. Influence of mesenchymal stem cell-derived extracellular vesicles in vitro and their role in ageing. Stem Cell Research & Therapy. 2020;11(1):p. 13. doi: 10.1186/s13287-019-1534-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Li C.-J., Cheng P., Liang M.-K., et al. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. Journal of Clinical Investigation. 2015;125(4):1509–1522. doi: 10.1172/JCI77716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Liu X., Yang Z., Meng Q., et al. Downregulation of microRNA-206 alleviates the sublethal oxidative stress-induced premature senescence and dysfunction in mesenchymal stem cells via targeting alpl. Oxidative Medicine and Cellular Longevity. 2020;2020:15. doi: 10.1155/2020/7242836.7242836 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Liu W., Zhang L., Xuan K., et al. Alpl prevents bone ageing sensitivity by specifically regulating senescence and differentiation in mesenchymal stem cells. Bone Research. 2018;6(1):p. 27. doi: 10.1038/s41413-018-0029-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Meng Y., Eirin A., Zhu X. Y., et al. Micro-RNAS regulate metabolic syndrome-induced senescence in porcine adipose tissue-derived mesenchymal stem cells through the P16/MAPK pathway. Cell Transplantation. 2018;27(10):1495–1503. doi: 10.1177/0963689718795692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wang D., Cai G., Wang H., He J. TRAF3, a target of microRNA-363-3p, suppresses senescence and regulates the balance between osteoblastic and adipocytic differentiation of Rat bone marrow-derived mesenchymal stem cells. Stem Cells and Development. 2020;29(11):737–745. doi: 10.1089/scd.2019.0276. [DOI] [PubMed] [Google Scholar]
- 100.Fan J., An X., Yang Y., et al. MiR-1292 targets FZD4 to regulate senescence and osteogenic differentiation of stem cells in te/sj/mesenchymal tissue system via the Wnt/β-catenin pathway. Aging Disease. 2018;9(6):1103–1121. doi: 10.14336/AD.2018.1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Ransohoff J. D., Wei Y., Khavari P. A. The functions and unique features of long intergenic non-coding RNA. Nature Reviews Molecular Cell Biology. 2018;19(3):143–157. doi: 10.1038/nrm.2017.104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kalwa M., Hänzelmann S., Otto S., et al. The lncRNA HOTAIR impacts on mesenchymal stem cells via triple helix formation. Nucleic Acids Research. 2016;44(22):10631–10643. doi: 10.1093/nar/gkw802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Li C. J., Xiao Y., Yang M., et al. Long noncoding RNA Bmncr regulates mesenchymal stem cell fate during skeletal aging. Journal of Clinical Investigation. 2018;128(12):5251–5266. doi: 10.1172/JCI99044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Huang C., Geng J., Jiang S. MicroRNAs in regulation of osteogenic differentiation of mesenchymal stem cells. Cell and Tissue Research. 2017;368(2):229–238. doi: 10.1007/s00441-016-2462-2. [DOI] [PubMed] [Google Scholar]
- 105.Li B. MicroRNA regulation in osteogenic and adipogenic differentiation of bone mesenchymal stem cells and its application in bone regeneration. Current Stem Cell Research & Therapy. 2018;13(1):26–30. doi: 10.2174/1574888X12666170605112727. [DOI] [PubMed] [Google Scholar]
- 106.Deng L., Hu G., Jin L., Wang C., Niu H. Involvement of microRNA-23b in TNF-α-reduced BMSC osteogenic differentiation via targeting runx2. Journal of Bone and Mineral Metabolism. 2018;36(6):648–660. doi: 10.1007/s00774-017-0886-8. [DOI] [PubMed] [Google Scholar]
- 107.Liao L., Yang X., Su X., et al. Redundant miR-3077-5p and miR-705 mediate the shift of mesenchymal stem cell lineage commitment to adipocyte in osteoporosis bone marrow. Cell Death & Disease. 2013;4(4, article e600) doi: 10.1038/cddis.2013.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Zhang Y., Jiang Y., Luo Y., Zeng Y. Interference of miR-212 and miR-384 promotes osteogenic differentiation via targeting RUNX2 in osteoporosis. Experimental and Molecular Pathology. 2020;113:p. 104366. doi: 10.1016/j.yexmp.2019.104366. [DOI] [PubMed] [Google Scholar]
- 109.Fu L., Liu H., Lei W. MiR-596 inhibits osteoblastic differentiation and cell proliferation by targeting Smad3 in steroid-induced osteonecrosis of femoral head. Journal of Orthopedic Surgery and Research. 2020;15(1):p. 173. doi: 10.1186/s13018-020-01688-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Hao C., Yang S., Xu W., et al. MiR-708 promotes steroid-induced osteonecrosis of femoral head, suppresses osteogenic differentiation by targeting SMAD3. Scientific Reports. 2016;6(1, article 22599) doi: 10.1038/srep22599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Zhang X., Zhu Y., Zhang C., et al. miR-542-3p prevents ovariectomy-induced osteoporosis in rats via targeting SFRP1. Journal of Cellular Physiology. 2018;233(9):6798–6806. doi: 10.1002/jcp.26430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Yang N., Wang G., Hu C., et al. Tumor necrosis factor α suppresses the mesenchymal stem cell osteogenesis promoter miR-21 in estrogen deficiency-induced osteoporosis. Journal of Bone and Mineral Research. 2013;28(3):559–573. doi: 10.1002/jbmr.1798. [DOI] [PubMed] [Google Scholar]
- 113.You L., Pan L., Chen L., Gu W., Chen J. MiR-27a is essential for the shift from osteogenic differentiation to adipogenic differentiation of mesenchymal stem cells in postmenopausal osteoporosis. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology. 2016;39(1):253–265. doi: 10.1159/000445621. [DOI] [PubMed] [Google Scholar]
- 114.Yang B. C., Kuang M. J., Kang J. Y., Zhao J., Ma J. X., Ma X. L. Human umbilical cord mesenchymal stem cell-derived exosomes act via the miR-1263/Mob1/Hippo signaling pathway to prevent apoptosis in disuse osteoporosis. Biochemical and Biophysical Research and Communications. 2020;524(4):883–889. doi: 10.1016/j.bbrc.2020.02.001. [DOI] [PubMed] [Google Scholar]
- 115.Shao B., Fu X., Yu Y., Yang D. Regulatory effects of miRNA-181a on FasL expression in bone marrow mesenchymal stem cells and its effect on CD4+T lymphocyte apoptosis in estrogen deficiency‑induced osteoporosis. Molecular Medicine Reports. 2018;18(1):920–930. doi: 10.3892/mmr.2018.9026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kuang M. J., Huang Y., Zhao X. G., et al. Exosomes derived from Wharton's jelly of human umbilical cord mesenchymal stem cells reduce osteocyte apoptosis in glucocorticoid-induced osteonecrosis of the femoral head in rats via the miR-21-PTEN-AKT signalling pathway. International Journal of Biological Sciences. 2019;15(9):1861–1871. doi: 10.7150/ijbs.32262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Gao Y., Xiao F., Wang C., et al. Long noncoding RNA MALAT1 promotes osterix expression to regulate osteogenic differentiation by targeting miRNA-143 in human bone marrow-derived mesenchymal stem cells. Journal of Cellular Biochemistry. 2018;119(8):6986–6996. doi: 10.1002/jcb.26907. [DOI] [PubMed] [Google Scholar]
- 118.Yang X., Yang J., Lei P., Wen T. LncRNA MALAT1 shuttled by bone marrow-derived mesenchymal stem cells-secreted exosomes alleviates osteoporosis through mediating microRNA-34c/SATB2 axis. Aging. 2019;11(20):8777–8791. doi: 10.18632/aging.102264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Yang L., Li Y., Gong R., et al. The long non-coding RNA-ORLNC1 regulates bone mass by directing mesenchymal stem cell fate. Molecular Therapy. 2019;27(2):394–410. doi: 10.1016/j.ymthe.2018.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Chen S., Jia L., Zhang S., Zheng Y., Zhou Y. DEPTOR regulates osteogenic differentiation via inhibiting MEG3-mediated activation of BMP4 signaling and is involved in osteoporosis. Stem Cell Research & Therapy. 2018;9(1):p. 185. doi: 10.1186/s13287-018-0935-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Xiaoling G., Shuaibin L., Kailu L. MicroRNA-19b-3p promotes cell proliferation and osteogenic differentiation of BMSCs by interacting with lncRNA H19. BMC Medical Genetetics. 2020;21(1):p. 11. doi: 10.1186/s12881-020-0948-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Wei B., Wei W., Zhao B., Guo X., Liu S. Long non-coding RNA HOTAIR inhibits miR-17-5p to regulate osteogenic differentiation and proliferation in non-traumatic osteonecrosis of femoral head. PloS One. 2017;12(2) doi: 10.1371/journal.pone.0169097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Kim H.-N., Lee J.-H., Bae S.-C., et al. Histone deacetylase inhibitor MS-275 stimulates bone formation in part by enhancing Dhx36-mediated TNAP transcription. Journal of Bone and Mineral Research. 2011;26(9):2161–2173. doi: 10.1002/jbmr.426. [DOI] [PubMed] [Google Scholar]
- 124.Bae H. S., Yoon W. J., Cho Y. D., et al. An HDAC inhibitor, entinostat/MS-275, partially prevents delayed cranial suture closure in heterozygous runx2 null mice. Journal of Bone and Mineral Research. 2017;32(5):951–961. doi: 10.1002/jbmr.3076. [DOI] [PubMed] [Google Scholar]
- 125.Xu S., De Veirman K., Evans H., et al. Effect of the HDAC inhibitor vorinostat on the osteogenic differentiation of mesenchymal stem cells in vitro and bone formation in vivo. Acta Pharmacologica Sinica. 2013;34(5):699–709. doi: 10.1038/aps.2012.182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Leng Q., Chen L., Lv Y. RNA-based scaffolds for bone regeneration: application and mechanisms of mRNA, miRNA and siRNA. Theranostics. 2020;10(7):3190–3205. doi: 10.7150/thno.42640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Chen L., HolmstrØm K., Qiu W., et al. MicroRNA-34a inhibits osteoblast differentiation and in vivo bone formation of human stromal stem cells. Stem Cells. 2014;32(4):902–912. doi: 10.1002/stem.1615. [DOI] [PubMed] [Google Scholar]
- 128.Chang C. C., Venø M. T., Chen L., et al. Global microRNA profiling in human bone marrow skeletal-stromal or mesenchymal-stem cells identified candidates for bone regeneration. Molecular therapy : the journal of the American Society of Gene Therapy. 2018;26(2):593–605. doi: 10.1016/j.ymthe.2017.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Yoshizuka M., Nakasa T., Kawanishi Y., et al. Inhibition of microRNA-222 expression accelerates bone healing with enhancement of osteogenesis, chondrogenesis, and angiogenesis in a rat refractory fracture model. Journal of Orthopaedic Science. 2016;21(6):852–858. doi: 10.1016/j.jos.2016.07.021. [DOI] [PubMed] [Google Scholar]
- 130.Hu X., Zhang X., Dai L., et al. Histone deacetylase inhibitor trichostatin A promotes the osteogenic differentiation of rat adipose-derived stem cells by altering the epigenetic modifications on Runx2 promoter in a BMP signaling-dependent manner. Stem Cells and Development. 2013;22(2):248–255. doi: 10.1089/scd.2012.0105. [DOI] [PubMed] [Google Scholar]
- 131.Lv L., Ge W., Liu Y., et al. Lysine-specific demethylase 1 inhibitor rescues the osteogenic ability of mesenchymal stem cells under osteoporotic conditions by modulating H3K4 methylation. Bone Research. 2016;4(1):p. 16037. doi: 10.1038/boneres.2016.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Hu Z., Zhang L., Wang H., et al. Targeted silencing of miRNA-132-3p expression rescues disuse osteopenia by promoting mesenchymal stem cell osteogenic differentiation and osteogenesis in mice. Stem Cell Research & Therapy. 2020;11(1):p. 58. doi: 10.1186/s13287-020-1581-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Wang Z., Zhang D., Hu Z., et al. MicroRNA-26a-modified adipose-derived stem cells incorporated with a porous hydroxyapatite scaffold improve the repair of bone defects. Molecular Medicine Reports. 2015;12(3):3345–3350. doi: 10.3892/mmr.2015.3795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Moncal K. K., Aydin R. S. T., Abu-Laban M., et al. Collagen-infilled 3D printed scaffolds loaded with miR-148b-transfected bone marrow stem cells improve calvarial bone regeneration in rats. Materials Science and Engineering: C. 2019;105:p. 110128. doi: 10.1016/j.msec.2019.110128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Xue Y., Guo Y., Yu M., Wang M., Ma P. X., Lei B. Monodispersed bioactive glass nanoclusters with ultralarge pores and intrinsic exceptionally high mirna loading for efficiently enhancing bone regeneration. Advanced Healthcare Materials. 2017;6(20) doi: 10.1002/adhm.201700630. [DOI] [PubMed] [Google Scholar]
- 136.Sui L., Wang M., Han Q., et al. A novel lipidoid-microRNA formulation promotes calvarial bone regeneration. Biomaterials. 2018;177:88–97. doi: 10.1016/j.biomaterials.2018.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Lin Z., He H., Wang M., Liang J. MicroRNA-130a controls bone marrow mesenchymal stem cell differentiation towards the osteoblastic and adipogenic fate. Cell Proliferation. 2019;52(6, article e12688) doi: 10.1111/cpr.12688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Seenprachawong K., Tawornsawutruk T., Nantasenamat C., Nuchnoi P., Hongeng S., Supokawej A. miR-130a and miR-27b Enhance Osteogenesis in Human Bone Marrow Mesenchymal Stem Cells via Specific Down-Regulation of Peroxisome Proliferator-Activated Receptor γ. Frontiers in Genetics. 2018;9:p. 543. doi: 10.3389/fgene.2018.00543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Liu H., Dong Y., Feng X., et al. miR-34a promotes bone regeneration in irradiated bone defects by enhancing osteoblastic differentiation of mesenchymal stromal cells in rats. Stem Cell Research & Therapy. 2019;10(1):p. 180. doi: 10.1186/s13287-019-1285-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Liu D., Kou X., Chen C., et al. Circulating apoptotic bodies maintain mesenchymal stem cell homeostasis and ameliorate osteopenia via transferring multiple cellular factors. Cell Research. 2018;28(9):918–933. doi: 10.1038/s41422-018-0070-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Deng Y., Guo T., Li J., Guo L., Gu P., Fan X. Repair of calvarial bone defect using Jarid1a-knockdown bone mesenchymal stem cells in rats. Tissue Engineering Part A. 2018;24(9-10):711–718. doi: 10.1089/ten.tea.2017.0168. [DOI] [PubMed] [Google Scholar]
- 142.Zhang J., Tu Q., Grosschedl R., et al. Roles of SATB2 in osteogenic differentiation and bone regeneration. Tissue Engineering Part A. 2011;17(13-14):1767–1776. doi: 10.1089/ten.tea.2010.0503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Deng Y., Bi X., You Z., Wang Y., Gu P., Fan X. Repair of critical-sized bone defects with anti-miR-31-expressing bone marrow stromal stem cells and poly(glycerol sebacate) scaffolds. European Cells & Materials. 2014;27:13–25. doi: 10.22203/eCM.v027a02. [DOI] [PubMed] [Google Scholar]
- 144.Yang C., Liu X., Zhao K., et al. miRNA-21 promotes osteogenesis via the PTEN/PI3K/Akt/HIF-1α pathway and enhances bone regeneration in critical size defects. Stem Cell Research & Therapy. 2019;10(1):p. 65. doi: 10.1186/s13287-019-1168-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Jin C., Zheng Y., Huang Y., Liu Y., Jia L., Zhou Y. Long non-coding RNAMIATknockdown promotes osteogenic differentiation of human adipose-derived stem cells. Cell Biology International. 2017;41(1):33–41. doi: 10.1002/cbin.10697. [DOI] [PubMed] [Google Scholar]
- 146.Jin C., Jia L., Huang Y., et al. Inhibition of lncRNA MIR31HG promotes osteogenic differentiation of human adipose-derived stem cells. Stem Cells (Dayton, Ohio) 2016;34(11):2707–2720. doi: 10.1002/stem.2439. [DOI] [PubMed] [Google Scholar]
- 147.Nguyen K. H., Xu F., Flowers S., Williams E. A. J., Fritton J. C., Moran E. SWI/SNF-mediated lineage determination in mesenchymal stem cells confers resistance to osteoporosis. Stem Cells. 2015;33(10):3028–3038. doi: 10.1002/stem.2064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Li K. C., Chang Y. H., Yeh C. L., Hu Y. C. Healing of osteoporotic bone defects by baculovirus-engineered bone marrow-derived MSCs expressing MicroRNA sponges. Biomaterials. 2016;74:155–166. doi: 10.1016/j.biomaterials.2015.09.046. [DOI] [PubMed] [Google Scholar]
- 149.Khanban H., Fattahi E., Talkhabi M. In vivo administration of G9a inhibitor A366 decreases osteogenic potential of bone marrow-derived mesenchymal stem cells. EXCLI Journal. 2019;18:300–309. doi: 10.17179/excli2019-1234. [DOI] [PMC free article] [PubMed] [Google Scholar]