

Abstract
Cortical thickness, surface area and volumes vary with age and cognitive function, and in neurological and psychiatric diseases. Here we report heritability, genetic correlations and genome-wide associations of these cortical measures across the whole cortex, and in 34 anatomically predefined regions. Our discovery sample comprises 22,824 individuals from 20 cohorts within the Cohorts for Heart and Aging Research in Genomic Epidemiology (CHARGE) consortium and the UK Biobank. We identify genetic heterogeneity between cortical measures and brain regions, and 160 genome-wide significant associations pointing to wnt/β-catenin, TGF-β and sonic hedgehog pathways. There is enrichment for genes involved in anthropometric traits, hindbrain development, vascular and neurodegenerative disease and psychiatric conditions. These data are a rich resource for studies of the biological mechanisms behind cortical development and aging.
Subject terms: Genome-wide association studies, Genetics of the nervous system, Neurology
Cortex morphology varies with age, cognitive function, and in neurological and psychiatric diseases. Here the authors report 160 genome-wide significant associations with thickness, surface area and volume of the total cortex and 34 cortical regions from a GWAS meta-analysis in 22,824 adults.
Introduction
The cortex is the largest part of the human brain, associated with higher brain functions, such as perception, thought, and action. Brain cortical thickness (CTh), cortical surface area (CSA), and cortical volume (CV) are morphological markers of cortical structure obtained from magnetic resonance imaging (MRI). These measures change with age1–3 and are linked to cognitive functioning4,5. The human cortex is also vulnerable to a wide range of disease or pathologies, ranging from developmental disorders and early onset psychiatric and neurological diseases to neurodegenerative conditions manifesting late in life. Abnormalities in global or regional CTh, CSA, and CV have been observed in neurological and psychiatric disorders, such as Alzheimer’s disease6, Parkinson’s disease7, multiple sclerosis8, schizophrenia9, bipolar disorder9, depression10, and autism11. The best method to study human cortical structure during life is using brain MRI. Hence, understanding the genetic determinants of the most robust MRI cortical markers in apparently normal adults could identify biological pathways relevant to brain development, aging, and various diseases. Neurons in the neocortex are organized in columns which run perpendicular to the surface of the cerebral cortex12; and, according to the radial unit hypothesis, CTh is determined by the number of cells within the columns and CSA is determined by the number of columns13.
Thus, CTh and CSA reflect different mechanisms in cortical development13,14 and are likely influenced by different genetic factors15–18. CV, which is the product of CTh and CSA, is determined by a combination of these two measures, but the relative contribution of CTh and CSA to CV may vary across brain regions. CTh, CSA, and CV are all strongly heritable traits15–21 with estimated heritability of 0.69–0.81 for global CTh, and from 0.42 to 0.90 for global CSA15,16,18. Across different cortical regions, however, there is substantial regional variation in heritability of CTh, CSA, and CV15–21.
Since CTh, CSA, and CV are differentially heritable and genetically heterogeneous, we explore the genetics of each of these imaging markers using genome-wide association analyses (GWAS) in large population-based samples. We study CTh, CSA, and CV in the whole cortex and in 34 cortical regions in 22,824 individuals from 21 discovery cohorts and replicate the strongest associations in 22,363 persons from the Enhancing Neuroimaging Genetics through Meta-analysis (ENIGMA) consortium. Our analyses reveal 160 genome-wide significant associations pointing to wnt/β-catenin, TGF-β, and sonic hedgehog pathways. We observe genetic heterogeneity between cortical measures and brain regions and find enrichment for genes involved in anthropometric traits, hindbrain development, vascular and neurodegenerative disease, and psychiatric conditions.
Results
Genome-wide association analysis
The analyses of global CTh, CSA, and CV included 22,163, 18,617, and 22,824 individuals, respectively. After correction for multiple testing (pDiscovery < 1.09 × 10−9), we identified no significant associations with global CTh. However, we identified 12 independent loci associated with global CSA (n = 6) and CV (n = 6). These are displayed in Supplementary Data 1 and Supplementary Figs. 1 and 2. Five of the 6 CSA loci were replicated in an external (ENIGMA consortium) sample22. The ENIGMA consortium only analyzed CSA and CTh.
GWAS of CTh, CSA, and CV in 34 cortical regions of interest (ROIs) identified 148 significant associations. There were 16 independent loci across 8 chromosomes determining CTh of 9 regions (Supplementary Data 2), 54 loci across 16 chromosomes associated with CSA of 21 regions (Supplementary Data 3), and 78 loci across 17 chromosomes determining CV of 23 cortical regions (Supplementary Data 4). We replicated 57 out of 64 regional CTh and CSA loci that were available in the ENIGMA consortium sample22 using a conservative replication threshold of pReplication = 3.1 × 10−4, 0.05/160. Region-specific variants with the strongest association at each genomic locus are shown in Tables 1–3. Chromosomal ideograms showing genome-wide significant associations with global and regional cortical measures in the discovery stage are presented in Fig. 1.
Table 2.
Genome-wide significant associations (pDiscovery < 1.09 × 10−9) of global and regional CSA.
| Lobe | Region | Locus | Position | Lead SNP | Nearest gene | Annotation | N | pDiscovery | pReplication | ppooled |
|---|---|---|---|---|---|---|---|---|---|---|
| Global | 17q21.31 | 44787313 | rs538628 | NSF | Intronic | 18,617 | 1.78E−23 | 4.45E−22 | 1.00E−43 | |
| 6q22.32 | 126792095 | rs11759026 | MIR588 | Intergenic | 18,617 | 5.21E−22 | 1.45E−14 | 3.50E−34 | ||
| 6q22.33 | 127204623 | rs9375477 | RSPO3 | Intergenic | 18,617 | 4.86E−13 | 1.60E−08 | 1.23E−19 | ||
| 6q21 | 109000316 | rs9398173 | FOXO3 | Intronic | 18,617 | 6.84E−10 | 2.96E−03 | 2.05E−10 | ||
| Frontal | Superior frontal | 5q14.3 | 92187932 | rs17669337 | NR2F1-AS1 | Intergenic | 18,272 | 1.40E−11 | 2.05E−06 | 8.07E−16 |
| Caudal middle frontal | 6q22.32 | 126876580 | rs9388500 | RSPO3 | Intergenic | 17,891 | 2.35E−11 | NA | NA | |
| Pars opercularis | 5q23.3 | 128734008 | rs12187568 | ADAMTS19 | Intergenic | 16,632 | 1.19E−16 | NA | NA | |
| Pars triangularis | 3q24 | 147106319 | rs2279829 | ZIC4 | UTR3 | 18,265 | 6.32E−20 | 1.94E−27 | 1.20E−45 | |
| 7q21.3 | 96175094 | rs10458281 | LOC100506136 | Intergenic | 18,265 | 1.15E−17 | 2.42E−11 | 1.20E−26 | ||
| Precentral | 15q14 | 39634222 | rs1080066 | C15orf54 | Intergenic | 18,267 | 8.45E−109 | 2.53E−95 | 1.00E−200 | |
| 6q15 | 92002569 | rs9345124 | MAP3K7 | Intergenic | 18,267 | 5.50E−11 | 2.73E−14 | 9.91E−24 | ||
| Temporal | Superior temporal | 2p16.3 | 48274592 | rs386645843 | FBXO11 | Intergenic | 18,269 | 9.51E−12 | 8.42E−07 | 1.71E−16 |
| 4q26 | 119249835 | rs55699931 | PRSS12 | Intronic | 18,269 | 2.08E−11 | 2.72E−02 | 6.96E−10 | ||
| 2q23.2 | 150022681 | rs13008194 | LYPD6B | Intronic | 18,269 | 5.94E−11 | 2.54E−07 | 1.92E−16 | ||
| Middle temporal | 6q22.32 | 126964510 | rs4273712 | RSPO3 | Intergenic | 18,269 | 6.93E−10 | 1.07E−04 | 1.99E−12 | |
| Banksts | 14q23.1 | 59072226 | rs186347 | KIAA0586 | Intergenic | 18,265 | 4.11E−10 | 1.83E−09 | 4.93E−18 | |
| Fusiform | 17q21.31 | 44822662 | rs199535 | NSF | Intronic | 18,269 | 1.01E−13 | 1.14E−06 | 8.13E−18 | |
| Transverse temporal | 2q23.2 | 150012936 | rs2046268 | LYPD6B | Intronic | 18,264 | 9.09E−10 | 3.21E−10 | 1.78E−18 | |
| Parietal | Superior parietal | 15q14 | 39632013 | rs71471500 | C15orf54 | Intergenic | 18,270 | 3.85E−24 | 5.55E−19 | 5.88E−41 |
| 19p13.2 | 13109763 | rs68175985 | NFIX | Intronic | 17,324 | 8.84E−11 | 2.68E−17 | 2.90E−26 | ||
| Inferior parietal | 20q13.2 | 52448936 | rs6097618 | SUMO1P1 | Intergenic | 18,267 | 1.78E−16 | NA | NA | |
| 12q14.3 | 65797096 | rs2336713 | MSRB3 | Intronic | 18,267 | 1.24E−12 | 2.99E−12 | 2.85E−23 | ||
| 2p25.2 | 4563477 | rs669952 | LINC01249 | Intergenic | 18,267 | 4.47E−10 | 1.37E−08 | 4.73E−17 | ||
| Supramarginal | 15q14 | 39633904 | rs2033939 | C15orf54 | Intergenic | 18,272 | 9.07E−27 | 1.61E−28 | 1.59E−53 | |
| 14q23.1 | 59627631 | rs2164950 | DAAM1 | Intergenic | 18,272 | 1.25E−13 | 3.79E−14 | 3.46E−26 | ||
| 3q24 | 147106319 | rs2279829 | ZIC4 | UTR3 | 18,272 | 7.38E−12 | 4.24E−16 | 2.29E−26 | ||
| Postcentral | 15q14 | 39634222 | rs1080066 | C15orf54 | Intergenic | 18,265 | 5.65E−47 | 2.44E−36 | 1.87E−80 | |
| 3q24 | 147106319 | rs2279829 | ZIC4 | UTR3 | 18,265 | 1.90E−21 | 1.69E−26 | 2.92E−46 | ||
| 9q21.13 | 76144318 | rs67286026 | ANXA1 | Intergenic | 18,265 | 3.58E−12 | 8.04E−06 | 7.82E−16 | ||
| Precuneus | 14q23.1 | 59628609 | rs74826997 | DAAM1 | Intergenic | 18,270 | 2.40E−24 | 4.41E−18 | 4.59E−40 | |
| 6q23.3 | 138866268 | rs9376354 | NHSL1 | Intronic | 18,270 | 7.80E−13 | 4.12E−08 | 7.28E−19 | ||
| 3q26 | 190666643 | rs1159211 | SNAR-I | Intergenic | 18,270 | 4.49E−10 | 2.04E−05 | 1.59E−13 | ||
| Occipital | Lateral occipital | 14q23.1 | 59627631 | rs2164950 | DAAM1 | Intergenic | 18,269 | 3.04E−26 | 2.92E−15 | 2.25E−38 |
| Lingual | 14q23.1 | 59628679 | rs76341705 | DAAM1 | Intergenic | 18,270 | 1.57E−20 | 8.67E−13 | 9.96E−31 | |
| Cuneus | 14q23.1 | 59625997 | rs73313052 | DAAM1 | Intergenic | 18,267 | 1.90E−32 | 3.19E−15 | 2.96E−43 | |
| 13q31.1 | 80191873 | rs9545155 | LINC01038 | Intergenic | 18,267 | 5.15E−10 | 2.98E−05 | 3.91E−13 | ||
| Pericalcarine | 14q23.1 | 59628679 | rs76341705 | DAAM1 | Intergenic | 18,267 | 4.67E−24 | 2.56E−19 | 3.35E−41 | |
| 5q12.1 | 60117723 | rs6893642 | ELOVL7 | Intronic | 18,267 | 1.40E−13 | 1.68E−08 | 6.29E−20 | ||
| 3q13.11 | 104724787 | rs971550 | ALCAM | Intergenic | 18,267 | 2.18E−10 | 1.31E−06 | 4.49E−15 | ||
| 6q22.33 | 127185801 | rs9375476 | RSPO3 | Intergenic | 18,267 | 2.20E−10 | 2.24E−08 | 4.32E−17 | ||
| 1p13.2 | 113239478 | rs2999158 | MOV10 | Intronic | 18,267 | 6.46E−10 | 8.39E−10 | 3.49E−18 | ||
| 13q31.1 | 80191873 | rs9545155 | LINC01068 | Intergenic | 18,267 | 7.51E−10 | 7.53E−09 | 4.05E−17 | ||
| Posterior cingulate | 5q12.3 | 66104105 | rs17214309 | MAST4 | Intronic | 18,268 | 7.84E−11 | 1.52E−05 | 4.04E−14 | |
| Insula | 10q25.3 | 118704077 | rs1905544 | SHTN1 | Intronic | 17,599 | 4.06E−12 | 3.65E−03 | 1.28E−11 |
N number of individuals in meta-analysis, pDiscovery two-sided p-value of discovery GWAS meta-analysis in CHARGE, pReplication two-sided p-value of replication meta-analysis in ENIGMA, ppooled two-sided p-value of pooled discovery and replication meta-analysis, p-values are not adjusted for multiple comparisons, banksts banks of the superior temporal sulcus.
NA, SNP or region not available in the ENIGMA sample.
In bold: significant replication—pReplication < 3.1 × 10−4 (= 0.05/Nl, Nl = 160, total number of lead SNPs).
Table 1.
Genome-wide significant associations (pDiscovery < 1.09 × 10−9) of regional CTh.
| Lobe | Region | Locus | Position | Lead SNP | Nearest gene | Annotation | N | pDiscovery | pReplication | ppooled |
|---|---|---|---|---|---|---|---|---|---|---|
| Temporal | Superior temporal | 16q24.2 | 87225139 | rs4843227 | LOC101928708 | Intergenic | 21,887 | 2.79E−12 | 2.45E−05 | 2.31E−15 |
| 17q21.31 | 44861003 | rs199504 | WNT3 | Intronic | 21,887 | 1.30E−10 | 1.17E−04 | 5.85E−13 | ||
| Middle temporal | 14q23.1 | 59072144 | rs10782438 | KIAA0586 | Intergenic | 21,559 | 2.17E−13 | 2.76E−08 | 8.99E−21 | |
| Inferior temporal | 2q35 | 217332057 | rs284532 | SMARCAL1 | Intronic | 21,885 | 1.03E−09 | 2.64E−01 | 3.04E−07 | |
| Banksts | 14q23.1 | 59074878 | rs160458 | KIAA0586( | Intergenic | 18,342 | 9.39E−10 | 2.42E−09 | 6.45E−18 | |
| Parietal | Superior parietal | 16q24.2 | 87225101 | rs9937293 | LOC101928708 | Intergenic | 21,886 | 2.68E−14 | 1.64E−13 | 2.27E−27 |
| 1q41 | 215141570 | rs10494988 | KCNK2 | Intergenic | 21,886 | 2.60E−12 | 3.66E−08 | 2.63E−19 | ||
| Postcentral | 15q14 | 39633904 | rs2033939 | C15orf54 | Intergenic | 21,885 | 1.17E−73 | 5.18E−68 | 7.73E−136 | |
| Occipital | Lateral occipital | 5q14.1 | 79933093 | rs245100 | DHFR | Intronic | 21,886 | 2.68E−11 | 3.77E−06 | 1.16E−15 |
| Cuneus | 14q23.1 | 59624317 | rs4901904 | DAAM1 | Intergenic | 21,885 | 4.02E−14 | 3.17E−10 | 2.88E−23 | |
| Insula | 16q12.1 | 51449978 | rs7197215 | SALL1 | Intergenic | 21,560 | 1.45E−13 | 2.00E−02 | 6.42E−12 | |
| 9q31.3 | 113679617 | rs72748157 | LPAR1 | Intronic | 21,560 | 1.46E−10 | 1.38E−04 | 5.16E−13 |
N number of individuals in meta-analysis, pDiscovery two-sided p-value of discovery GWAS meta-analysis in CHARGE, pReplication two-sided p-value of replication meta-analysis in ENIGMA, ppooled two-sided p-value of pooled discovery and replication meta-analysis, p-values are not adjusted for multiple comparisons, banksts banks of the superior temporal sulcus.
in bold: significant replication—pReplication < 3.1 × 10−4 (= 0.05/Nl, Nl=160, total number of lead SNPs).
Table 3.
Genome-wide significant associations (pDiscovery < 1.09 × 10−9) of global and regional CV.
| Lobe | Region | Locus | Position | Lead SNP | Nearest gene | Annotation | N | pDiscovery |
|---|---|---|---|---|---|---|---|---|
| Global | 6q22.32 | 126792095 | rs11759026 | MIR588 | Intergenic | 22,410 | 6.31E−19 | |
| 17q21.31 | 44790203 | rs169201 | NSF | Intronic | 22,784 | 2.11E−13 | ||
| 17q21.32 | 43549608 | rs149366495 | PLEKHM1 | Intronic | 22,099 | 8.18E−13 | ||
| 12q14.3 | 66358347 | rs1042725 | HMGA2 | 3’UTR | 22,784 | 7.04E−11 | ||
| 12q23.2 | 102921296 | rs11111293 | IGF1 | Intergenic | 22,784 | 5.45E−10 | ||
| 6q22 | 109002042 | rs4945816 | FOXO3 | 3’UTR | 22,784 | 8.93E−10 | ||
| Frontal | Superior frontal | 5q14.3 | 92186429 | rs888814 | NR2F1-AS1 | Intergenic | 22,692 | 3.29E−13 |
| Rostral middle frontal | 15q14 | 39636227 | rs17694988 | C15orf54 | Intergenic | 22,793 | 3.15E−11 | |
| Caudal middle frontal | 2q12.1 | 105460333 | rs745249 | LINC01158 | ncRNA_intronic | 22,726 | 2.35E−11 | |
| 6q22.32 | 127068983 | rs853974 | RSPO3 | Intergenic | 22,351 | 4.82E−11 | ||
| Pars opercularis | 5q23.3 | 128734008 | rs12187568 | ADAMTS19 | Intergenic | 20,753 | 4.27E−18 | |
| 15q14 | 39639898 | rs4924345 | C15orf54 | Intergenic | 22,758 | 1.97E−14 | ||
| Pars triangularis | 3q24 | 147106319 | rs2279829 | ZIC4 | UTR3 | 22,759 | 3.16E−23 | |
| 7q21.3 | 96196906 | rs67055449 | LOC100506136 | Intergenic | 22,759 | 4.03E−19 | ||
| 15q14 | 39633904 | rs2033939 | C15orf54 | Intergenic | 22,759 | 8.49E−14 | ||
| 7q21.3 | 96129071 | rs62470042 | C7orf76 | Intronic | 22,759 | 7.38E−13 | ||
| 6q15 | 91942761 | rs12660096 | MAP3K7 | Intergenic | 22,759 | 4.74E−10 | ||
| Lateral orbitofrontal | 14q22.2 | 54769839 | rs6572946 | CDKN3 | Intergenic | 22,801 | 2.29E−10 | |
| Precentral | 15q14 | 39634222 | rs1080066 | C15orf54 | Intergenic | 22,699 | 5.84E−125 | |
| 10q25.3 | 118648841 | rs3781566 | SHTN1 | Intronic | 22,699 | 4.68E−11 | ||
| Temporal | Superior temporal | 3q26.32 | 177296448 | rs13084960 | LINC00578 | ncRNA_intronic | 22,681 | 1.12E−11 |
| Banksts | 14q23.1 | 59072226 | rs186347 | KIAA0586 | Intergenic | 22,727 | 1.15E−15 | |
| Fusiform | 14q23.1 | 59833172 | rs1547199 | DAAM1 | Intronic | 22,605 | 4.58E−10 | |
| 1p33 | 47980916 | rs6658111 | FOXD2 | Intergenic | 22,605 | 7.78E−10 | ||
| Transverse temporal | 2q23.2 | 150012936 | rs2046268 | LYPD6B | Intronic | 22,786 | 2.55E−12 | |
| Parahippocampal | 2q33.1 | 199809716 | rs966744 | SATB2 | Intergenic | 22,747 | 2.23E−10 | |
| Parietal | Superior parietal | 15q14 | 39633904 | rs2033939 | C15orf54 | Intergenic | 22,723 | 4.28E−23 |
| 16q24.2 | 87225139 | rs4843227 | LOC101928708 | Intergenic | 22,723 | 1.16E−13 | ||
| 19p13.2 | 13109763 | rs68175985 | NFIX | Intronic | 21,777 | 3.27E−11 | ||
| 5q15 | 92866553 | rs62369942 | NR2F1-AS1 | ncRNA_intronic | 21,664 | 4.32E−10 | ||
| Inferior parietal | 20q13.2 | 52448936 | rs6097618 | SUMO1P1 | Intergenic | 22,701 | 2.09E−17 | |
| 12q14.3 | 65797096 | rs2336713 | MSRB3 | Intronic | 22,701 | 2.47E−13 | ||
| 3q13.11 | 104724634 | rs971551 | ALCAM | Intergenic | 22,701 | 2.34E−10 | ||
| Supramarginal | 15q14 | 39632013 | rs71471500 | THBS1 | Intergenic | 22,645 | 9.71E−28 | |
| 14q23.1 | 59627631 | rs2164950 | DAAM1 | Intergenic | 22,645 | 3.59E−20 | ||
| 3q24 | 147106319 | rs2279829 | ZIC4 | UTR3 | 22,645 | 5.36E−18 | ||
| Postcentral | 15q14 | 39633904 | rs2033939 | THBS1 | Intergenic | 22,662 | 4.34E−133 | |
| 3q24 | 147106319 | rs2279829 | ZIC4 | UTR3 | 22,662 | 2.54E−17 | ||
| 9q21.13 | 76144318 | rs67286026 | ANXA1 | Intergenic | 22,662 | 5.03E−11 | ||
| 2q36.3 | 226563259 | rs16866701 | NYAP2 | Intergenic | 22,545 | 5.69E−11 | ||
| Precuneus | 14q23.1 | 59628609 | rs74826997 | DAAM1 | Intergenic | 22,803 | 4.85E−20 | |
| 3q28 | 190663557 | rs35055419 | OSTN | Intergenic | 22,428 | 2.02E−10 | ||
| 2p22.2 | 37818236 | rs2215605 | CDC42EP3 | Intergenic | 22,803 | 3.43E−10 | ||
| 3q13.11 | 104713881 | rs12495603 | ALCAM | Intergenic | 22,803 | 9.71E−10 | ||
| Occipital | Lateral occipital | 14q23.1 | 59627631 | rs2164950 | DAAM1 | Intergenic | 22,799 | 6.89E−16 |
| Lingual | 14q23.1 | 59625997 | rs73313052 | DAAM1 | Intergenic | 22,805 | 1.06E−20 | |
| 6q22.32 | 127089401 | rs2223739 | RSPO3 | Intergenic | 22,805 | 1.75E−10 | ||
| Cuneus | 14q23.1 | 59625997 | rs73313052 | DAAM1 | Intergenic | 22,799 | 4.59E−43 | |
| 11p15.3 | 12072213 | rs11022131 | DKK3 | Intergenic | 22,799 | 5.96E−12 | ||
| 13q31.1 | 80192236 | rs9545156 | LINC01068 | Intergenic | 22,799 | 4.09E−10 | ||
| Pericalcarine | 14q23.1 | 59628679 | rs76341705 | DAAM1 | Intergenic | 22,824 | 1.39E−29 | |
| 13q31.1 | 80191873 | rs9545155 | LINC01068 | intergenic | 22,824 | 2.25E−13 | ||
| 11p14.1 | 30876113 | rs273594 | DCDC5 | Intergenic | 22,824 | 3.51E−13 | ||
| 1p13.2 | 113208039 | rs12046466 | CAPZA1 | Intronic | 22,824 | 2.36E−12 | ||
| 1p33 | 47980916 | rs6658111 | FOXD2 | Intergenic | 22,824 | 3.85E−11 | ||
| 11q22.3 | 104012656 | rs1681464 | PDGFD | Intronic | 22,824 | 7.51E−11 | ||
| 6q22.32 | 127096181 | rs9401907 | RSPO3 | Intergenic | 22,824 | 2.11E−10 | ||
| 7p21.1 | 18904400 | rs12700001 | HDAC9 | Intronic | 22,824 | 2.12E−10 | ||
| 5q12.1 | 60315823 | rs10939879 | NDUFAF2 | Intronic | 22,824 | 2.92E−10 | ||
| Caudal anterior cingulate | 5q14.3 | 82852578 | rs309588 | VCAN | Intronic | 22,748 | 2.60E−10 | |
| Insula | 11q23.1 | 110949402 | rs321403 | C11orf53 | Intergenic | 22,543 | 9.58E−12 | |
| 8q24.12 | 120596023 | rs10283100 | ENPP2 | Exonic | 21,481 | 8.29E−11 |
Fig. 1. Chromosomal ideogram of genome-wide significant associations with measures of cortical structure.

Cortical surface areas, cortical volumes and cortical thickness. Each point represents the significantly associated variant, the colors correspond to the different cortical regions and the shape to different type of measument (pDiscovery < 1.09 × 10−9).
If we had used a more stringent threshold of pDiscovery < 4.76 × 10−10 = 5 × 10−8/105, correcting for all the 105 GWAS analyses performed, we would have identified 142 significant associations (Supplementary Data 1–4).
The strongest associations with CTh and CV were observed for rs2033939 at 15q14 (pDiscovery, CTh = 1.17 × 10−73 and pDiscovery, CV = 4.34 × 10−133) in the postcentral (primary somatosensory) cortex, and for CSA with rs1080066 at 15q14 (pDiscovery, CSA = 8.45 × 10−109) in the precentral (primary motor) cortex. Figure 2 shows the lowest p-value of each cortical region. The postcentral cortex was also the region with the largest number of independent associations, mainly at a locus on 15q14. The corresponding regional association plots are presented in Supplementary Fig. 3. Quantile-quantile plots of all meta-analyses are presented in Supplementary Figs. 4–7 and the corresponding genomic inflation factors (λGC), LD score regression (LDSR) intercepts, and ratios are shown in Supplementary Data 5. Although we observe inflated test statistics for some traits with λGC between 1.02 and 1.11, LDSR intercepts between 0.98 and 1.02 indicate that the inflation is mainly due to polygenicity. For traits with λGC > 1.05, the LDSR ratios range between 0.00 and 0.15 which means that a maximum of 15% of the inflation is due to other causes.
Fig. 2. Lowest discovery meta-analysis p-value of CSA, CTh, and CV in each cortical region.

a Lowest pDiscovery of CSA, b lowest pDiscovery of CTh, c lowest pDiscovery of CV.
Associations across cortical measures and with other traits
Supplementary Data 6 presents variants that are associated with the CSA or the CV across multiple regions. We observed 25 single nucleotide polymorphisms (SNPs) that determined both the CSA and CV of a given region, 4 SNPs that determined CTh and CV of the same region, but no SNPs that determined both the CTh and CSA of any given region (Supplementary Data 7). We also checked the overlap between our findings and two previous GWAS studies, including 842823 and 19,62124 individuals from the UK Biobank, which among other phenotypes, investigate CTh, CSA, and CV (Supplementary Data 8). Regarding CTh, one variant, rs2033939 at 15q14, was associated with CTh of the postcentral gyrus in both studies. For CSA and CV, we found 11 associations at 15q14, 14q23.1 and 3q24, and 14 associations at 15q14, 14q23.1, 3q24, 8q24.1, 12q14.3, and 20q13.2, respectively, with the same cortical region as in our study. Out-of-sample polygenic risk score (PRS) analyses showed associations (pPRS < 4.76 × 10−3) with all investigated cortical measures in all cortical regions in 7800 UK Biobank individuals (Supplementary Data 9). For CTh, we observed the maximum phenotypic variance explained by the PRS (RPRS2) in the global cortex (RPRS2 = 0.015, pPRS = 1.05 × 10−26), and for CSA and CV in the pericalcarine cortex (RPRS2,CSA = 0.029, pPRS,CSA = 1.29 × 10−50; RPRS2,CV = 0.032, pPRS,CV = 5.30 × 10−56). When assessing genetic overlap with other traits, we observed that SNPs determining these cortical measures have been previously associated with anthropometric (height), neurologic (Parkinson’s disease, corticobasal degeneration, and Alzheimer’s disease), psychiatric (neuroticism and schizophrenia) and cognitive performance traits as well as with total intracranial volume (TIV) on brain MRI (Supplementary Data 10–12).
Gene identification
Positional mapping based on ANNOVAR showed that most of the lead SNPs were intergenic and intronic (Fig. 3). One variant, rs2279829, which was associated with both CSA and CV of the pars triangularis, postcentral and supramarginal cortices, is located in the 3′UTR of ZIC4 at 3q24. We also found an exonic variant, rs10283100, in gene ENPP2 at 8q24.12 associated with CV of the insula.
Fig. 3. Functional annotation categories for global and regional CTh, CSA, and CV.
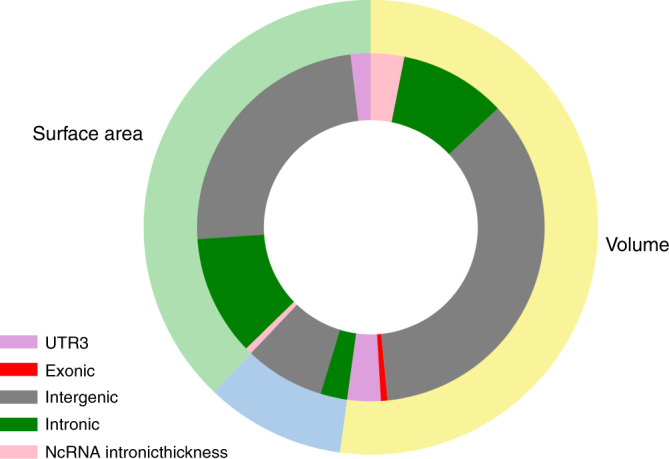
Proportion of functional annotation categories for global and regional cortical thickness (blue), surface area (light green), and volume (yellow) assigned by ANNOVAR.
We used multiple strategies beyond positional annotation to identify specific genes implicated by the various GWAS associated SNPs. FUMA identified 232 genes whose expression was determined by these variants (eQTL) and these and other genes implicated by chromatin interaction mapping are shown in Supplementary Data 13–15. MAGMA gene-based association analyses revealed 70 significantly associated (p < 5.87 × 10−8) genes (Supplementary Data 16–18). For global CSA and CV, 7 of 9 genes associated with each measure overlapped, but there was no overlap with global CTh. For regional CSA and CV, we found 28 genes across 13 cortical regions that determined both measures in the same region. Figure 4 summarizes the results of GTEx eQTL, chromatin interaction, positional annotation, and gene-based mapping strategies for all regions. While there are overlapping genes identified using different approaches, only DAAM1 gene (Chr14q23.1) is identified by all types of gene mapping for CV of insula. eQTL associations of our independent lead SNPs in the Religious Orders Study Memory and Aging Project (ROSMAP) dorsolateral frontal cortex gene expression dataset are presented in Supplementary Data 19.
Fig. 4. Number of overlapping genes between gene mapping methods.

Number of overlapping genes between FUMA eQTL mapping, FUMA chromatin interaction mapping, ANNOVAR chromosome positional mapping, and MAGMA gene-based analysis for all cortical regions combined for cortical surface area (a), thickness (b) and volume (c).
Pathway analysis
MAGMA gene set analyses identified 7 pathways for CTh, 3 pathways for CSA and 9 pathways for CV (Supplementary Data 20). Among them are the gene ontology (GO) gene sets hindbrain morphogenesis (strongest association with thickness of middle temporal cortex), forebrain generation of neurons (with surface area of precentral cortex), and central nervous system neuron development (with volume of transverse temporal cortex). However, after Bonferroni correction only one significant pathway (p < 1.02 × 10−7) remained: regulation of catabolic process for CTh of the inferior temporal cortex. InnateDB pathway analyses of genes mapped to independent lead SNPs by FUMA showed a significant overlap between CTh and CSA genes and the Wnt signaling pathway (Supplementary Figs. 8 and 9) as well as a significant overlap between CV genes and the basal cell carcinoma pathway (Supplementary Fig. 10).
Heritability
Heritability estimates (h2) of global CTh were 0.64 (standard error (se) = 0.12; pSOLAR = 3 × 10−7) in the ASPS-Fam study and 0.45 (se = 0.08; pGCTA = 2.5 × 10−7) in the Rotterdam study (RS). For CSA, h2 was 0.84 (se = 0.12; pSOLAR = 2.63 × 10−11) in ASPS-Fam and 0.33 (se = 0.08, pGCTA = 1 × 10−4) in RS, and for CV, h2 was 0.80 (se = 0.11; pSOLAR = 1.10 × 10−9) in ASPS-Fam and 0.32 (se = 0.08; pGCTA = 1 × 10−4) in RS. There was a large range in heritability estimates of regional CTh, CSA, and CV (Supplementary Data 21).
Heritability based on common SNPs as estimated with LDSR was 0.25 (se = 0.03) for global CTh, 0.29 (se = 0.04) for global CSA and 0.30 (se = 0.03) for global CV. LDSR heritability estimates of regional CTh, CSA, and CV are presented in Supplementary Data 21 and Supplementary Fig. 11. For the regional analyses, the estimated heritability ranged from 0.05 to 0.18 for CTh, from 0.07 to 0.36 for CSA and from 0.06 to 0.32 for CV. Superior temporal cortex (h2CTh = 0.18, h2CSA = 0.30, h2CV = 0.26), precuneus (h2CTh = 0.16, h2CSA = 0.29, h2CV = 0.28) and pericalcarine (h2CTh = 0.15, h2CSA = 0.36, h2CV = 0.32) are among the most genetically determined regions.
The results of partitioned heritability analyses for global and regional CTh, CSA, and CV with functional annotation and additionally with cell-type-specific annotation are presented in Supplementary Data 22 and 23. For global CTh, we found enrichment for super-enhancers, introns and histone marks. Repressors and histone marks were enriched for global CSA, and introns, super-enhancers, and repressors for global CV. For regional CSA and CV the highest enrichment scores (>18) were observed for conserved regions.
Genetic correlation
We found high genetic correlation (rg) between global CSA, and global CV (rg = 0.81, pLDSR = 1.2 × 10−186) and between global CTh and global CV (rg = 0.46, pLDSR = 1.4 × 10−14), but not between global CTh and global CSA (rg = −0.02, pLDSR = 0.82). Whereas the genetic correlation between CSA and CV was strong (rg > 0.7) in most of the regions (Supplementary Data 24 and Supplementary Fig. 12), it was generally weak between CSA and CTh with rg < 0.3, and ranged from 0.09 to 0.69 between CTh and CV. The postcentral and lingual cortices were the two regions with the highest genetic correlations between both CTh and CV, as well as CTh and CSA.
Genetic correlation across the various brain regions for CTh (Supplementary Fig. 13, Supplementary Data 25), CSA (Supplementary Fig. 14, Supplementary Data 26), and CV (Supplementary Fig. 15, Supplementary Data 27) showed a greater number of correlated regions for CTh and greater inter-regional variation for CSA and CV. Supplementary Data 28–30 and Supplementary Figs. 16–18 show genome-wide genetic correlations between the cortical measures and anthropometric, neurological and psychiatric, and cerebral structural traits.
Discussion
In our genome-wide association study of up to 22,824 individuals for MRI determined cortical measures of global and regional thickness, surface area, and volume, we identified 160 genome-wide significant associations across 19 chromosomes. Heritability was generally higher for cortical surface area and volume than for thickness, suggesting a greater susceptibility of cortical thickness to environmental influences. We observed strong genetic correlations between surface area and volume, but weak genetic correlation between surface area and thickness. We identified the largest number of novel genetic associations with cortical volumes, perhaps due to our larger sample size for this phenotype, which was assessed in all 21 discovery samples.
It is beyond the scope of our study to discuss each of the 160 associations identified. A large number of the corresponding genes are involved in pathways that regulate morphogenesis of neurons, neuronal cell differentiation, and cell growth, as well as cell migration and organogenesis during embryonic development. At a molecular level, the wnt/β-catenin, TGF-β, and sonic hedgehog pathways are strongly implicated. Gene-set-enrichment analyses revealed biological processes related to brain morphology and neuronal development.
Broad patterns emerged showing that genes determining cortical structure are also often implicated in development of the cerebellum and brainstem (KIAA0586, ZIC4, ENPP2) as well as the neural tube (one carbon metabolism genes DHFR and MSRBB3, the latter also associated with hippocampal volumes25). These genes determine development of not only neurons but also astroglia (THBS1) and microglia (SALL1). They determine susceptibility or resistance to a range of insults: inflammatory, vascular (THBS1, ANXA1, ARRDC3-AS126) and neurodegenerative (C15orf53, ZIC4, ANXA1), and have been associated with pediatric and adult psychiatric conditions (THBS1).
There is a wealth of information in the supplementary tables that can be mined for a better understanding of brain development, connectivity, function and pathology. We highlight this potential by discussing in additional detail, the possible significance of 6 illustrative loci, 5 of which, at 15q14, 14q23.1, 6q22.32, 17q21.31, and 3q24, associate with multiple brain regions at low p-values, while the locus at 8q24.12 identifies a plausible exonic variant.
The Chr15q14 locus was associated with cortical thickness, surface area, and volumes in the postcentral gyrus as well as with surface area or volume across six other regions in the frontal and parietal lobes. Lead SNPs at this locus were either intergenic between C15orf53 and C15orf54, or intergenic between C15orf54 and THBS1 (Thrombospondin-1). C15orf53 has been associated with an autosomal recessive form of spastic paraplegia showing intellectual disability and thinning of the corpus callosum (hereditary spastic paraparesis 11, or Nakamura Osame syndrome). Variants of THBS1 were reported to be related to autism27 and schizophrenia28. The protein product of THBS1 is involved in astrocyte induced synaptogenesis29, and regulates chain migration of interneuron precursors migrating in the postnatal radial migration stream to the olfactory bulb30. Moreover, THBS1 is an activator of TGFβ signaling, and an inhibitor of pro-angiogenic nitric oxide signaling, which plays a role in several cancers and immune-inflammatory conditions.
Variants at Chr14q23.1 were associated with cortical surface area and volume of all regions in the occipital lobe, as well as with thickness, surface area, and volume of the middle temporal cortex, banks of the superior temporal sulcus, fusiform, supramarginal and precuneus regions, areas associated with discrimination and recognition of language or visual form. These variants are either intergenic between KIAA0586, the product of which is a conserved centrosomal protein essential for ciliogenesis, sonic hedgehog signaling and intracellular organization, and DACT1, the product of which is a target for SIRT1 and acts on the wnt/β-catenin pathway. KIAA0586 has been associated with Joubert syndrome, another condition associated with abnormal cerebellar development. Other variants are intergenic between DACT1 and DAAM1 or intronic in DAAM1. DAAM1 has been associated with occipital lobe volume in a previous GWAS31.
Locus 6q22.32 contains various SNPs associated with cortical surface area and volume globally, and also within some frontal, temporal and occipital regions. The SNPs are intergenic between RSPO3 and CENPW. RSPO3 and CENPW have been previously associated with intracranial32,33 and occipital lobe volumes31. RSPO3 is an activator of the canonical Wnt signaling pathway and a regulator of angiogenesis.
Chr17q21.31 variants were associated with global cortical surface area and volume and with regions in temporal lobe. These variants are intronic in the genes PLEKHM1, CRHR1, NSF, and WNT3. In previous GWAS analyses, these genes have been associated with general cognitive function34 and neuroticism35. CRHR1, NSF, and WNT3 were additionally associated with Parkinson’s disease36 and intracranial volume32,33,37. The NSF gene also plays a role in Neuronal Intranuclear Inclusion Disease38 and CRHR1 is involved in anxiety and depressive disorders39. This chromosomal region also contains the MAPT gene, which plays a role in Alzheimer’s disease, Parkinson’s disease, and frontotemporal dementia40,41.
The protein product of the gene ZIC4 is a C2H2 zinc finger transcription factor that has an intraneuronal, non-synaptic expression and auto-antibodies to this protein have been associated with subacute sensory neuronopathy, limbic encephalitis, and seizures in patients with breast, small cell lung or ovarian cancers. ZIC4 null mice have abnormal development of the visual pathway42 and heterozygous deletion of the gene has also been associated with a congenital cerebellar (Dandy-Walker) malformation43, thus implicating it widely in brain development as well as in neurodegeneration. C2H2ZF transcription factors are the most widely expressed transcription factors in eukaryotes and show associations with responses to abiotic (environmental) stressors. Another transcription factor, FOXC1, also associated with Dandy-Walker syndrome has been previously shown to be associated with risk of all types of ischemic stroke and with stroke severity. Thus, ZIC4 might be a biological target worth pursuing to ameliorate neurodegenerative disorders.
We found an exonic SNP within the gene ENPP2 (Autotaxin) at 8q24.12 to be associated with insular cortical volume. This gene is differentially expressed in the frontal cortex of Alzheimer patients44 and in mouse models of Alzheimer disease, such as the senescence-accelerated mouse prone 8 strain (SAMP8) mouse. Autotaxin is a dual-function ectoenzyme, which is the primary source of the signaling lipid, lysophosphatidic acid. Besides Alzheimer disease, changes in autotaxin/lysophosphatidic acid signaling have also been shown in diverse brain-related conditions, such as intractable pain, pruritus, glioblastoma, multiple sclerosis, and schizophrenia. In the SAMP8 mouse, improvements in cognition noted after administration of LW-AFC, a putative Alzheimer remedy derived from the traditional Chinese medicinal prescription ‘Liuwei Dihuang’ decoction, are correlated with restored expression of four genes in the hippocampus, one of which is ENPP2.
Among the other genetic regions identified, many have been linked to neurological and psychiatric disorders, cognitive functioning, cortical development, and cerebral structure (detailed listing in Supplementary Data 31).
Heritability estimates are, as expected, generally higher in the family-based Austrian Stroke Prevention-Family study (ASPS-Fam) than in the Rotterdam Study (RS) for CTh (average h2ASPS-Fam = 0.52; h2RS = 0.26), CSA (0.62 and 0.30) and CV (0.57 and 0.23). This discrepancy is explained by the different heritability estimation methods: pedigree-based heritability in ASPS-Fam versus heritability based on common SNPs that are in LD with causal variants45 in RS.
Average heritability over regions is also higher for surface area and volume, than for thickness. The observed greater heritability of CSA compared to CTh is consistent with the previously articulated hypothesis, albeit based on much smaller numbers, that CSA is developmentally determined to a greater extent with smaller subsequent decline after young adulthood, whereas CTh changes over the lifespan as aging, neurodegeneration and vascular injuries accrue1,3. It is also interesting that brain regions more susceptible to early amyloid deposition (e.g., superior temporal cortex and precuneus) have a higher heritability.
We found no or weak genetic correlation between CTh and CSA, globally and regionally, and no common lead SNPs, which indicates that these two morphological measures are genetically independent, a finding consistent with prior reports15,16. In contrast, we found strong genetic correlation between CSA and CV and identified common lead SNPs for CSA and CV globally, and in 12 cortical regions. Similar findings have been reported in a previous publication16. The genetic correlation between CTh and CV ranged between 0.09 and 0.77, implying a common genetic background in some regions (such as the primary sensory postcentral and lingual cortices), but not in others. For CTh, we observed genetic correlations between multiple regions within each of the lobes, whereas for CSA and CV, we found genetic correlations mainly between different regions of the occipital lobe. Chen et al.46 have also reported strong genetic correlation for CSA within the occipital lobe. There were also a few genetic correlations observed for regions from different lobes, suggesting similarities in cortical development transcended traditional lobar boundaries.
A limitation of our study is the heterogeneity of the MR phenotypes between cohorts due to different scanners, field strengths, MR protocols and MRI analysis software. This heterogeneity as well as the different age ranges in the participating cohorts may have caused different effects over the cohorts. We nevertheless combined the data of the individual cohorts to maximize the sample size as it has been done in previous CHARGE GWAS analyses31–33. To account for the heterogeneity we used a sample-size weighted meta-analysis that does not provide overall effect estimates. This method has lower power to detect associations compared to inverse-variance weighted meta-analysis and we therefore may have found less associations.Our inability to replicate 8 of the 76 genome-wide significant findings for CTh and CSA could be caused by false-positive results but may also be explained by insufficient power due to a too small sample size. Moreover, our sample comprises of mainly European ancestry, limiting the generalizability to other ethnicities. Strengths of our study are the population-based design, the large age range of our sample (20–100 years), use of three cortical measures as phenotypes of cortical morphometry, and the replication of our CTh and CSA findings in a large and independent cohort. In conclusion, we identified patterns of heritability and genetic associations with various global and regional cortical measures, as well as overlap of MRI cortical measures with genetic traits and diseases that provide new insights into cortical development, morphology, and possible mechanisms of disease susceptibility.
Methods
Study population
The sample of this study consist of up to 22,824 participants from 20 population-based cohort studies collaborating in the Cohorts of Heart and Aging Research in Genomic Epidemiology (CHARGE) consortium and the UK Biobank (UKBB). All the individuals were stroke- and dementia free, aged between 20 and 100 years, and of European ancestry, except for ARIC AA with African ancestry. Supplementary Data 32 provides population characteristics of each cohort and the Supplementary Methods provide a short description of each study. Each study secured approval from institutional review boards or equivalent organizations, and all participants provided written informed consent. Our results were replicated using summary GWAS findings of 22,635 individuals from the ENIGMA consortium.
Genotyping and imputation
Genotyping was conducted using various commercially available genotyping arrays across the study cohorts. Prior to imputation, extensive quality control was performed in each cohort. Genotype data were imputed to the 1000 Genomes reference panel (mainly phase 1, version 3) using validated software. Details on genotyping, quality control and imputation can be found in Supplementary Data 33.
Phenotype definition
This study investigated CTh, CSA, and CV globally in the whole cortex and in 34 cortical regions. Global and regional CTh was defined as the mean thickness of the left and the right hemisphere in millimeter (mm). Global CSA was defined as the total surface area of the left and the right hemisphere in mm2, while regional CSA was defined as the mean surface area of the left and the right hemisphere in mm2. Global and regional CV was defined as the mean volume of the left and the right hemisphere in mm3. The 34 cortical regions are listed in the Supplementary Methods. High resolution brain magnetic resonance imaging (MRI) data was obtained in each cohort using a range of MRI scanners, field strengths and protocols. CTh, CSA, and CV were generated using the Freesurfer software package47 in all cohorts except for FHSucd, where an in-house segmentation method was used. MRI protocols of each cohort can be found in Supplementary Data 35 and descriptive statistics of CTh, CSA, and CV can be found in Supplementary Data 36–38.
Genome-wide association analysis
Based on a predefined analysis plan, each study fitted linear regression models to determine the association between global and regional CTh, CSA, and CV and allele dosages of SNPs. Additive genetic effects were assumed and the models were adjusted for sex, age, age2, and if needed for study site and for principal components to correct for population stratification. Cohorts including related individuals calculated linear mixed models to account for family structure. Details on association software and covariates for each cohort are shown in Supplementary Data 33. Models investigating regional CTh, CSA, and CV were additionally adjusted for global CTh, global CSA and global CV, respectively. Quality control of the summary statistics shared by each cohort was performed using EasyQC48. Genetic variants with a minor allele frequency (MAF) <0.05, low imputation quality (R2 < 0.4), and which were available in less than 10,000 individuals were removed from the analyses. Details on quality control are provided in the Supplementary Methods.
We then used METAL49 to perform meta-analyses using the z-scores method, based on p-values, sample size, and direction of effect, with genomic control correction. To estimate the number of independent tests for the p-value threshold correction, we used a non-parametric permutation testing procedure50–53 in the combined Rotterdam Study cohort (N = 4442) and UK Biobank (N = 8213). First, we generated a random independent variable, to insure that there is no true relationship between brain measurements and this variable. Second, we ran linear regression analyses between this variable and all brain measurements one-by-one in each of the cohorts separately (104 regressions in total per cohort). Third, we saved the minimum p-value obtained from those 104 regressions. Then, as suggested in literature54, we repeated this procedure 10.000 times. Therefore, at the end we had 10.000 minimum p-values per cohort. The minimum p-value distribution follows a Beta distribution Beta(m,n), where m = 1 and n is the degree of freedom, which represents the number of independent tests in case of permutation testing. Using python statistical library we fitted the Beta function with the saved minimum p-values, and found n for Rotterdam Study and UK Biobank identically equal to 46. Based on the permutation test results, the genome-wide significance threshold was set a priori at 1.09 × 10−9 (=5 × 10−8/46). We used the clumping function in PLINK55 (linkage disequilibrium (LD) threshold: 0.2, distance: 300 kb) to identify the most significant SNP in each LD block. We used LDSR to calculate genomic inflation factors (λGC), LDSR intercepts and LDSR ratios for each meta-analysis. The LDSR intercept was estimated to differentiate between inflation due to a polygenic signal and inflation due to population stratification56. The LDSR ratio represents the amount of inflation that is due to other causes than polygenicity such as population stratification or cryptic relatedness.
For replication of our genome-wide significant CTh and CSA associations, we used GWAS meta-analysis results from the ENIGMA consortium22 for all SNPs that were associated at a p-value <5 × 10−8 and performed a pooled meta-analysis. The p-value threshold for replication was set to 3.1 × 10−4 (=0.05/160: nominal significance threshold divided by total number of lead SNPs). CV was not available in the ENIGMA results. PRS analysis was performed for 7800 out of sample subjects (not included in the current GWAS) from UK Biobank cohort using the PRSice-2 software57 with standard settings. The significance threshold for the association between the PRS and the phenotype was set to 4.76 × 10−3 (=0.05/105: nominal significance divided by number GWAS phenotypes). The NHGRI-EBI Catalog of published GWAS58 was searched for previous SNP-trait associations at a p-value of 5 × 10−8 of lead SNPs. Regional association plots were generated with LocusZoom59, and the chromosomal ideogram with PHENOGRAM (http://visualization.ritchielab.org/phenograms/plot).
Annotation of genome-wide significant variants was performed using the ANNOVAR software package60 and the FUMA web application61. FUMA eQTL mapping uses information from three data repositories (GTEx, Blood eQTL browser, and BIOS QTL browser) and maps SNPs to genes based on a significant eQTL association. We used a false discovery rate threshold (FDR) of 0.05 divided by number of tests (46) to define significant eQTL associations. Gene-based analyses, to combine the effects of SNPs assigned to a gene, and gene set analyses, to find out if genes assigned to significant SNPs were involved in biological pathways, were performed using MAGMA62 as implemented in FUMA. The significance threshold was set to 5.87 × 10−8 (=0.05/18522*46: FDR threshold divided by number of genes and independent tests) for gene-based analyses and to 1.02 × 10−7 (=0.05/10651: FDR threshold divided by the number of gene sets) for the gene set analyses. Additionally, FUMA was used to investigate a significant chromatin interaction between a genomic region in a risk locus and promoter regions of genes (250 bp upstream and 500 bp downstream of a TSS). We used an FDR of 1 × 10−6 to define significant interactions.
We investigated cis (<1 Mb) and trans (>1 MB or on a different chromosome) expression quantitative trait loci (eQTL) for genome-wide significant SNPs in 724 post-mortem brains from ROSMAP63,64 stored in the AMP-AD database. The samples were collected from the gray matter of the dorsolateral prefrontal cortex. The significance threshold was set to 0.001 (=0.05/46: FDR threshold divided by the number of independent tests). For additional pathway analyses of genes that were mapped to independent lead SNPs by FUMA, we searched the InnateDB database65. The STRING database66 was used for visualizing protein–protein interactions. Only those protein subnetworks with five or more nodes are shown.
Heritability
Additive genetic heritability (h2) of CTh, CSA, and CV was estimated in two studies: the Austrian Stroke Prevention Family Study (ASPS-Fam; n = 365) and the Rotterdam Study (RS, n = 4472). In the population-based family study ASPS-Fam, the ratio of the genotypic variance to the phenotypic variance was calculated using variance components models in SOLAR67. In case of non-normalty, phenotype data were inverse-normal transformed. In RS, SNP-based heritability was computed with GCTA68. These heritability analyses were adjusted for age and sex.
Heritability and partitioned heritability based on GWAS summary statistics was calculated from GWAS summary statistics using LDSR) implemented in the LDSC tool (https://github.com/bulik/ldsc). Partitioned heritability analysis splits genome-wide SNP heritability into 53 functional annotation classes (e.g., coding, 3′UTR, promoter, transcription factor binding sites, conserved regions etc.) and additionally to 10 cell-type specific classes (e.g., central nervous system, cardiovascular, liver, skeletal muscle, etc.) as defined by Finucane et al.69 to estimate their contributions to heritability. The significance threshold was set to 2.05 × 10−5 (=0.05/53*46: nominal significance divided by number of functional annotation classes and number of independent tests) for heritability partitioned on functional annotation classes and 2.05 < 10−6 (=0.05/53*10*46: nominal significance divided by number of functional annotation classes, number of cell types and number of independent tests) for heritability partitioned on annotation classes and cell types.
Genetic correlation
LDSR genetic correlation70 between CTh, CSA, and CV was estimated globally and within each cortical region. The significance threshold was set to 7.35 × 10−4 (nominal threshold (0.05) divided by number of regions (34) and by number of correlations (CSA and CV, CSA and CTh). Genetic correlation was also estimated between all 34 cortical regions for CTh, CSA, and CV, with the significance threshold set to 8.91 × 10−5 (nominal threshold (0.05) divided by number of regions (34) times the number of regions −1 (33) divided by 2 (half of the matrix). Additionally, the amount of genetic correlation was quantified between CTh, CSA, and CV and physical traits (height, body mass index), neurological and psychiatric diseases (e.g., Alzheimer’s disease, Parkinson’s disease), cognitive traits and MRI volumes (p-value threshold (0.05/46/number of GWAS traits). As recommended by the LDSC tool developers, only HapMap3 variants were included in these analyses, as these tend to be well-imputed across cohorts.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Supplementary information
Descriptions of Additional Supplementary Files
Acknowledgements
We provide all investigator and study-specific acknowledgements in Supplementary Note 1.
Author contributions
Drafting of the manuscript: E.H., G.V.R., R.S., and S.S.; Genotype and phenotype data acquisition: H.S., N.A., P.R.S., M.J.W., C.E.F., P.S.S., K.A.M, N.J.A., J.B.K., O.A.A, A.M.D, M.C.N, U.V., R.S., S.M., B.M., J.J., J.T., G.B.P., H.B., A.T., D.A., R.B, J.T.B., O.L.L., C.T., and P.A.; Imaging, genetic and bioinformatics analysis: E.H., G.V.R., H.H.H.A., M.J.K., Y.S., R.X., J.C.B., S.A., S.L., S.J.v.d.L., Q.Y., C.L.S., H.L., J.J.H., H.Z., M.L., M.S., N.J.A., M.W.V., A.T., K.W., M.B., A.M., N.A.G., M.L., P.R.S., X.J., O.C., A.S.B, L.P., St. S., P.M., C.dC., S.K., L.L., M.B., W.W., F.B., A.V.W., M.H., J.J., and F.C. Cohort PIs: P.S.S., W.S.K., J.A.W., A.V., C.M.v. D, H.J.G., W.T.L.Jr, M.F., T.P., S.D., M.A.I., H.S., R.S., S.S., J.I.R., B.M.P., I.J.D., M.L., A.H., A.G.U., W.J.N., Z.P., M.S.P., T.H.M., J.T., C.E.F., M.J.L., and C.T.; ENIGMA Study design: K.L.G., N.J., J.N.P., L.C.-C., J.B., D.P.H., P.A.L., F.P., J.L.S., P.M.T., and S.E.M.; Critical revision of the manuscript: all authors.
Data availability
The genome-wide summary statistics that support the findings of this study are available via the CHARGE Summary Results portal at the NCBI dbGaP website https://www.omicsdi.org/dataset/dbgap/phs000930 upon publication, or from the corresponding authors R.S. and S.S. upon reasonable request. The summary statistics may be used for all scientific purposes except for the study of potentially sensitive and potentially stereotyping phenotypes such as intelligence and addiction, since this is proscribed by the consent terms for the NHLBI cohorts. Individual level data or study-specific summary results are only available through controlled access. Data for the Framingham Study are available through dbGaP, where qualified researchers can apply for authorization to access (https://www.ncbi.nlm.nih.gov/projects/gap/cgi-bin/study.cgi?study_id=phs000007.v30.p11). Individual level data for the ARIC and CHS studies are also available through dbGaP. Data of European and Australian cohorts are available upon request, in keeping with data sharing guidelines in the EU General Data Protection Regulation. Data from UK Biobank can be accessed at http://www.ukbiobank.ac.uk and for the ENIGMA consortium from medlandse@gmail.com. Individual level data for VETSA is not available due to consent restrictions.
Competing interests
Dr. Dale is a founder of and holds equity in CorTechs Labs, Inc, and serves on its Scientific Advisory Board. He is a member of the Scientific Advisory Board of Human Longevity, Inc. and receives funding through research agreements with General Electric Healthcare and Medtronic, Inc. The terms of these arrangements have been reviewed and approved by UCSD in accordance with its conflict of interest policies. W. Niessen is co-founder and shareholder of Quantib BV. H. Brodaty is an advisory board member of Nutricia. The other authors declare no competing interests.
Footnotes
Peer review information Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. Peer review reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Edith Hofer, Gennady V. Roshchupkin, Hieab H.H. Adams.
These authors jointly supervised this work: Reinhold Schmidt, Sudha Seshadri.
A list of authors and their affiliations appears at the end of the paper.
Contributor Information
Reinhold Schmidt, Email: reinhold.schmidt@medunigraz.at.
Sudha Seshadri, Email: suseshad@bu.edu.
ENIGMA consortium:
Katrina L. Grasby, Neda Jahanshad, Jodie N. Painter, Lucía Colodro-Conde, Janita Bralten, Derrek P. Hibar, Penelope A. Lind, Fabrizio Pizzagalli, Christopher R. K. Ching, Mary Agnes B. McMahon, Natalia Shatokhina, Leo C. P. Zsembik, Ingrid Agartz, Saud Alhusaini, Marcio A. A. Almeida, Dag Alnæs, Inge K. Amlien, Micael Andersson, Tyler Ard, Nicola J. Armstrong, Allison Ashley-Koch, Manon Bernard, Rachel M. Brouwer, Elizabeth E. L. Buimer, Robin Bülow, Christian Bürger, Dara M. Cannon, Mallar Chakravarty, Qiang Chen, Joshua W. Cheung, Baptiste Couvy-Duchesne, Anders M. Dale, Shareefa Dalvie, Tânia K. de Araujo, Greig I. de Zubicaray, Sonja M. C. de Zwarte, Anouk den Braber, Nhat Trung Doan, Katharina Dohm, Stefan Ehrlich, Hannah-Ruth Engelbrecht, Susanne Erk, Chun Chieh Fan, Iryna O. Fedko, Sonya F. Foley, Judith M. Ford, Masaki Fukunaga, Melanie E. Garrett, Tian Ge, Sudheer Giddaluru, Aaron L. Goldman, Nynke A. Groenewold, Dominik Grotegerd, Tiril P. Gurholt, Boris A. Gutman, Narelle K. Hansell, Mathew A. Harris, Marc B. Harrison, Courtney C. Haswell, Michael Hauser, Stefan Herms, Dirk J. Heslenfeld, New Fei Ho, David Hoehn, Per Hoffmann, Laurena Holleran, Martine Hoogman, Jouke-Jan Hottenga, Masashi Ikeda, Deborah Janowitz, Iris E. Jansen, Tianye Jia, Christiane Jockwitz, Ryota Kanai, Sherif Karama, Dalia Kasperaviciute, Tobias Kaufmann, Sinead Kelly, Masataka Kikuchi, Marieke Klein, Michael Knapp, Annchen R. Knodt, Bernd Krämer, Max Lam, Thomas M. Lancaster, Phil H. Lee, Tristram A. Lett, Lindsay B. Lewis, Iscia Lopes-Cendes, Michelle Luciano, Fabio Macciardi, Andre F. Marquand, Samuel R. Mathias, Tracy R. Melzer, Yuri Milaneschi, Nazanin Mirza-Schreiber, Jose C. V. Moreira, Thomas W. Mühleisen, Bertram Müller-Myhsok, Pablo Najt, Soichiro Nakahara, Kwangsik Nho, Loes M. Olde Loohuis, Dimitri Papadopoulos Orfanos, John F. Pearson, Toni L. Pitcher, Benno Pütz, Anjanibhargavi Ragothaman, Faisal M. Rashid, Ronny Redlich, Céline S. Reinbold, Jonathan Repple, Geneviève Richard, Brandalyn C. Riedel, Shannon L. Risacher, Cristiane S. Rocha, Nina Roth Mota, Lauren Salminen, Arvin Saremi, Andrew J. Saykin, Fenja Schlag, Lianne Schmaal, Peter R. Schofield, Rodrigo Secolin, Chin Yang Shapland, Li Shen, Jean Shin, Elena Shumskaya, Ida E. Sønderby, Emma Sprooten, Lachlan T. Strike, Katherine E. Tansey, Alexander Teumer, Anbupalam Thalamuthu, Sophia I. Thomopoulos, Diana Tordesillas-Gutiérrez, Jessica A. Turner, Anne Uhlmann, Costanza Ludovica Vallerga, Dennis van der Meer, Marjolein M. J. van Donkelaar, Liza van Eijk, Theo G. M. van Erp, Neeltje E. M. van Haren, Daan van Rooij, Marie-José van Tol, Jan H. Veldink, Ellen Verhoef, Esther Walton, Mingyuan Wang, Yunpeng Wang, Joanna M. Wardlaw, Wei Wen, Lars T. Westlye, Christopher D. Whelan, Stephanie H. Witt, Katharina Wittfeld, Christiane Wolf, Thomas Wolfers, Clarissa L. Yasuda, Dario Zaremba, Zuo Zhang, Alyssa H. Zhu, Marcel P. Zwiers, Eric Artiges, Amelia A. Assareh, Rosa Ayesa-Arriola, Aysenil Belger, Christine L. Brandt, Gregory G. Brown, Sven Cichon, Joanne E. Curran, Gareth E. Davies, Franziska Degenhardt, Bruno Dietsche, Srdjan Djurovic, Colin P. Doherty, Ryan Espiritu, Daniel Garijo, Yolanda Gil, Penny A. Gowland, Robert C. Green, Alexander N. Häusler, Walter Heindel, Beng-Choon Ho, Wolfgang U. Hoffmann, Florian Holsboer, Georg Homuth, Norbert Hosten, Clifford R. Jack, Jr, MiHyun Jang, Andreas Jansen, Knut Kolskår, Sanne Koops, Axel Krug, Kelvin O. Lim, Jurjen J. Luykx, Daniel H. Mathalon, Karen A. Mather, Venkata S. Mattay, Sarah Matthews, Jaqueline Mayoral Van Son, Sarah C. McEwen, Ingrid Melle, Derek W. Morris, Bryon A. Mueller, Matthias Nauck, Jan E. Nordvik, Markus M. Nöthen, Daniel S. O’Leary, Nils Opel, Marie -. Laure Paillère Martinot, G. Bruce Pike, Adrian Preda, Erin B. Quinlan, Varun Ratnakar, Simone Reppermund, Vidar M. Steen, Fábio R. Torres, Dick J. Veltman, James T. Voyvodic, Robert Whelan, Tonya White, Hidenaga Yamamori, Marina K. M. Alvim, David Ames, Tim J. Anderson, Ole A. Andreassen, Alejandro Arias-Vasquez, Mark E. Bastin, Bernhard T. Baune, John Blangero, Dorret I. Boomsma, Henry Brodaty, Han G. Brunner, Randy L. Buckner, Jan K. Buitelaar, Juan R. Bustillo, Wiepke Cahn, Vince Calhoun, Xavier Caseras, Svenja Caspers, Gianpiero L. Cavalleri, Fernando Cendes, Aiden Corvin, Benedicto Crespo-Facorro, John C. Dalrymple-Alford, Udo Dannlowski, Eco J. C. de Geus, Ian J. Deary, Norman Delanty, Chantal Depondt, Sylvane Desrivières, Gary Donohoe, Thomas Espeseth, Guillén Fernández, Simon E. Fisher, Herta Flor, Andreas J. Forstner, Clyde Francks, Barbara Franke, David C. Glahn, Randy L. Gollub, Hans J. Grabe, Oliver Gruber, Asta K. Håberg, Ahmad R. Hariri, Catharina A. Hartman, Ryota Hashimoto, Andreas Heinz, Manon H. J. Hillegers, Pieter J. Hoekstra, Avram J. Holmes, L. Elliot Hong, William D. Hopkins, Hilleke E. Hulshoff Pol, Terry L. Jernigan, Erik G. Jönsson, René S. Kahn, Martin A. Kennedy, Tilo T. J. Kircher, Peter Kochunov, John B. J. Kwok, Stephanie Le Hellard, Nicholas G. Martin, Jean -. Luc Martinot, Colm McDonald, Katie L. McMahon, Andreas Meyer-Lindenberg, Rajendra A. Morey, Lars Nyberg, Jaap Oosterlaan, Roel A. Ophoff, Tomáš Paus, Zdenka Pausova, Brenda W. J. H. Penninx, Tinca J. C. Polderman, Danielle Posthuma, Marcella Rietschel, Joshua L. Roffman, Laura M. Rowland, Perminder S. Sachdev, Philipp G. Sämann, Gunter Schumann, Kang Sim, Sanjay M. Sisodiya, Jordan W. Smoller, Iris E. Sommer, Beate St Pourcain, Dan J. Stein, Arthur W. Toga, Julian N. Trollor, Nic J. A. Van der Wee, Dennis van ’t Ent, Henry Völzke, Henrik Walter, Bernd Weber, Daniel R. Weinberger, Margaret J. Wright, Juan Zhou, Jason L. Stein, Paul M. Thompson, and Sarah E. Medland
Supplementary information
Supplementary information is available for this paper at 10.1038/s41467-020-18367-y.
References
- 1.Storsve AB, et al. Differential longitudinal changes in cortical thickness, surface area and volume across the adult life span: regions of accelerating and decelerating change. J. Neurosci. 2014;34:8488–8498. doi: 10.1523/JNEUROSCI.0391-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hogstrom LJ, Westlye LT, Walhovd KB, Fjell AM. The structure of the cerebral cortex across adult life: age-related patterns of surface area, thickness, and gyrification. Cereb. Cortex. 2013;23:2521–2530. doi: 10.1093/cercor/bhs231. [DOI] [PubMed] [Google Scholar]
- 3.Fjell AM, et al. Development and aging of cortical thickness correspond to genetic organization patterns. Proc. Natl Acad. Sci. USA. 2015;112:15462–15467. doi: 10.1073/pnas.1508831112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Vuoksimaa E, et al. The genetic association between neocortical volume and general cognitive ability is driven by global surface area rather than thickness. Cereb. Cortex. 2015;25:2127–2137. doi: 10.1093/cercor/bhu018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vuoksimaa E, et al. Is bigger always better? The importance of cortical configuration with respect to cognitive ability. NeuroImage. 2016;129:356–366. doi: 10.1016/j.neuroimage.2016.01.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lerch JP, et al. Focal decline of cortical thickness in Alzheimer’s disease identified by computational neuroanatomy. Cereb. Cortex. 2005;15:995–1001. doi: 10.1093/cercor/bhh200. [DOI] [PubMed] [Google Scholar]
- 7.Uribe C, et al. Patterns of cortical thinning in nondemented Parkinson’s disease patients. Mov. Disord. 2016;31:699–708. doi: 10.1002/mds.26590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Steenwijk MD, et al. Cortical atrophy patterns in multiple sclerosis are non-random and clinically relevant. Brain. 2016;139:115–126. doi: 10.1093/brain/awv337. [DOI] [PubMed] [Google Scholar]
- 9.Rimol LM, et al. Cortical volume, surface area, and thickness in schizophrenia and bipolar disorder. Biol. Psychiatry. 2012;71:552–560. doi: 10.1016/j.biopsych.2011.11.026. [DOI] [PubMed] [Google Scholar]
- 10.Schmaal L, et al. Cortical abnormalities in adults and adolescents with major depression based on brain scans from 20 cohorts worldwide in the ENIGMA Major Depressive Disorder Working Group. Mol. Psychiatry. 2017;22:900–909. doi: 10.1038/mp.2016.60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.van Rooij D, et al. Cortical and subcortical brain morphometry differences between patients with autism spectrum disorder and healthy individuals across the lifespan: results from the ENIGMA ASD Working Group. Am. J. Psychiatry. 2018;175:359–369. doi: 10.1176/appi.ajp.2017.17010100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mountcastle VB. The columnar organization of the neocortex. Brain. 1997;120(Pt 4):701–722. doi: 10.1093/brain/120.4.701. [DOI] [PubMed] [Google Scholar]
- 13.Rakic P. A small step for the cell, a giant leap for mankind: a hypothesis of neocortical expansion during evolution. Trends Neurosci. 1995;18:383–388. doi: 10.1016/0166-2236(95)93934-P. [DOI] [PubMed] [Google Scholar]
- 14.Rakic P. Evolution of the neocortex: a perspective from developmental biology. Nat. Rev. Neurosci. 2009;10:724–735. doi: 10.1038/nrn2719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Panizzon MS, et al. Distinct genetic influences on cortical surface area and cortical thickness. Cereb. Cortex. 2009;19:2728–2735. doi: 10.1093/cercor/bhp026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Winkler AM, et al. Cortical thickness or grey matter volume? The importance of selecting the phenotype for imaging genetics studies. NeuroImage. 2010;53:1135–1146. doi: 10.1016/j.neuroimage.2009.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rimol LM, et al. Cortical thickness is influenced by regionally specific genetic factors. Biol. psychiatry. 2010;67:493–499. doi: 10.1016/j.biopsych.2009.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Eyler LT, et al. Genetic and environmental contributions to regional cortical surface area in humans: a magnetic resonance imaging twin study. Cereb. Cortex. 2011;21:2313–2321. doi: 10.1093/cercor/bhr013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kremen WS, et al. Genetic and environmental influences on the size of specific brain regions in midlife: the VETSA MRI study. Neuroimage. 2010;49:1213–1223. doi: 10.1016/j.neuroimage.2009.09.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Joshi AA, et al. The contribution of genes to cortical thickness and volume. Neuroreport. 2011;22:101–105. doi: 10.1097/WNR.0b013e3283424c84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wen W, et al. Distinct genetic influences on cortical and subcortical brain structures. Sci. Rep. 2016;6:32760. doi: 10.1038/srep32760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grasby, K. L. et al. The genetic architecture of the human cerebral cortex. Science. 10.1126/science.aay6690 (2020).
- 23.Elliott LT, et al. Genome-wide association studies of brain imaging phenotypes in UK Biobank. Nature. 2018;562:210–216. doi: 10.1038/s41586-018-0571-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhao B, et al. Genome-wide association analysis of 19,629 individuals identifies variants influencing regional brain volumes and refines their genetic co-architecture with cognitive and mental health traits. Nat. Genet. 2019;51:1637–1644. doi: 10.1038/s41588-019-0516-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hibar DP, et al. Novel genetic loci associated with hippocampal volume. Nat. Commun. 2017;8:13624. doi: 10.1038/ncomms13624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Irvin, M. R. et al. Genome-wide meta-analysis of SNP-by9-ACEI/ARB and SNP-by-thiazide diuretic and effect on serum potassium in cohorts of European and African ancestry. Pharmacogenomics J.10.1038/s41397-018-0021-9 (2018). [DOI] [PMC free article] [PubMed]
- 27.Lu L, et al. Common and rare variants of the THBS1 gene associated with the risk for autism. Psychiatr. Genet. 2014;24:235–240. doi: 10.1097/YPG.0000000000000054. [DOI] [PubMed] [Google Scholar]
- 28.Park HJ, Kim SK, Kim JW, Kang WS, Chung JH. Association of thrombospondin 1 gene with schizophrenia in Korean population. Mol. Biol. Rep. 2012;39:6875–6880. doi: 10.1007/s11033-012-1513-3. [DOI] [PubMed] [Google Scholar]
- 29.Christopherson KS, et al. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell. 2005;120:421–433. doi: 10.1016/j.cell.2004.12.020. [DOI] [PubMed] [Google Scholar]
- 30.Blake SM, et al. Thrombospondin-1 binds to ApoER2 and VLDL receptor and functions in postnatal neuronal migration. EMBO J. 2008;27:3069–3080. doi: 10.1038/emboj.2008.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.van der Lee SJ, et al. A genome-wide association study identifies genetic loci associated with specific lobar brain volumes. Commun. Biol. 2019;2:285. doi: 10.1038/s42003-019-0537-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ikram MA, et al. Common variants at 6q22 and 17q21 are associated with intracranial volume. Nat. Genet. 2012;44:539–544. doi: 10.1038/ng0612-732c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Adams HH, et al. Novel genetic loci underlying human intracranial volume identified through genome-wide association. Nat. Neurosci. 2016;19:1569–1582. doi: 10.1038/nn.4398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Davies G, et al. Study of 300,486 individuals identifies 148 independent genetic loci influencing general cognitive function. Nat. Commun. 2018;9:2098. doi: 10.1038/s41467-018-04362-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Luciano M, et al. Association analysis in over 329,000 individuals identifies 116 independent variants influencing neuroticism. Nat. Genet. 2018;50:6–11. doi: 10.1038/s41588-017-0013-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chang D, et al. A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nat. Genet. 2017;49:1511–1516. doi: 10.1038/ng.3955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hibar DP, et al. Common genetic variants influence human subcortical brain structures. Nature. 2015;520:224–229. doi: 10.1038/nature14101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pountney DL, Raftery MJ, Chegini F, Blumbergs PC, Gai WP. NSF, Unc-18-1, dynamin-1 and HSP90 are inclusion body components in neuronal intranuclear inclusion disease identified by anti-SUMO-1-immunocapture. Acta Neuropathologica. 2008;116:603–614. doi: 10.1007/s00401-008-0437-4. [DOI] [PubMed] [Google Scholar]
- 39.Muller MB, Wurst W. Getting closer to affective disorders: the role of CRH receptor systems. Trends Mol. Med. 2004;10:409–415. doi: 10.1016/j.molmed.2004.06.007. [DOI] [PubMed] [Google Scholar]
- 40.Desikan RS, et al. Genetic overlap between Alzheimer’s disease and Parkinson’s disease at the MAPT locus. Mol. Psychiatry. 2015;20:1588–1595. doi: 10.1038/mp.2015.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Spillantini MG, Goedert M. Tau pathology and neurodegeneration. Lancet Neurol. 2013;12:609–622. doi: 10.1016/S1474-4422(13)70090-5. [DOI] [PubMed] [Google Scholar]
- 42.Horng S, et al. Differential gene expression in the developing lateral geniculate nucleus and medial geniculate nucleus reveals novel roles for Zic4 and Foxp2 in visual and auditory pathway development. The. J. Neurosci. 2009;29:13672–13683. doi: 10.1523/JNEUROSCI.2127-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Grinberg I, et al. Heterozygous deletion of the linked genes ZIC1 and ZIC4 is involved in Dandy-Walker malformation. Nat. Genet. 2004;36:1053–1055. doi: 10.1038/ng1420. [DOI] [PubMed] [Google Scholar]
- 44.Umemura K, et al. Autotaxin expression is enhanced in frontal cortex of Alzheimer-type dementia patients. Neurosci. Lett. 2006;400:97–100. doi: 10.1016/j.neulet.2006.02.008. [DOI] [PubMed] [Google Scholar]
- 45.Wray NR, et al. Pitfalls of predicting complex traits from SNPs. Nat. Rev. Genet. 2013;14:507–515. doi: 10.1038/nrg3457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chen CH, et al. Genetic influences on cortical regionalization in the human brain. Neuron. 2011;72:537–544. doi: 10.1016/j.neuron.2011.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Desikan RS, et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. 2006;31:968–980. doi: 10.1016/j.neuroimage.2006.01.021. [DOI] [PubMed] [Google Scholar]
- 48.Winkler TW, et al. Quality control and conduct of genome-wide association meta-analyses. Nat. Protoc. 2014;9:1192–1212. doi: 10.1038/nprot.2014.071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Willer CJ, Li Y, Abecasis GR. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics. 2010;26:2190–2191. doi: 10.1093/bioinformatics/btq340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Purcell S, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007;81:559–575. doi: 10.1086/519795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Nichols TE. Multiple testing corrections, nonparametric methods, and random field theory. NeuroImage. 2012;62:811–815. doi: 10.1016/j.neuroimage.2012.04.014. [DOI] [PubMed] [Google Scholar]
- 52.Uppu S, Krishna A, Gopalan RP. A review on methods for detecting SNP interactions in high-dimensional genomic. Data. IEEE/ACM Trans. Comput Biol. Bioinform. 2018;15:599–612. doi: 10.1109/TCBB.2016.2635125. [DOI] [PubMed] [Google Scholar]
- 53.Alberton, B. A. V., Nichols, T. E., Gamba, H. R. & Winkler, A. M. Multiple testing correction over contrasts for brain imaging. NeuroImage. 10.1016/j.neuroimage.2020.116760 (2020). [DOI] [PMC free article] [PubMed]
- 54.Churchill GA, Doerge RW. Empirical threshold values for quantitative trait mapping. Genetics. 1994;138:963–971. doi: 10.1093/genetics/138.3.963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chang CC, et al. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience. 2015;4:7. doi: 10.1186/s13742-015-0047-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Bulik-Sullivan BK, et al. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat. Genet. 2015;47:291–295. doi: 10.1038/ng.3211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Choi, S. W. & O’Reilly, P. F. PRSice-2: Polygenic Risk Score software for biobank-scale data. Gigascience10.1093/gigascience/giz082 (2019). [DOI] [PMC free article] [PubMed]
- 58.MacArthur J, et al. The new NHGRI-EBI Catalog of published genome-wide association studies (GWAS Catalog) Nucleic Acids Res. 2017;45:D896–D901. doi: 10.1093/nar/gkw1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pruim RJ, et al. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 2010;26:2336–2337. doi: 10.1093/bioinformatics/btq419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wang K, Li M, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010;38:e164. doi: 10.1093/nar/gkq603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017;8:1826. doi: 10.1038/s41467-017-01261-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.de Leeuw CA, Mooij JM, Heskes T, Posthuma D. MAGMA: generalized gene-set analysis of GWAS data. PLoS Comput Biol. 2015;11:e1004219. doi: 10.1371/journal.pcbi.1004219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bennett DA, Schneider JA, Arvanitakis Z, Wilson RS. Overview and findings from the religious orders study. Curr. Alzheimer Res. 2012;9:628–645. doi: 10.2174/156720512801322573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bennett DA, et al. Overview and findings from the rush Memory and Aging Project. Curr. Alzheimer Res. 2012;9:646–663. doi: 10.2174/156720512801322663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Breuer K, et al. InnateDB: systems biology of innate immunity and beyond-recent updates and continuing curation. Nucleic acids Res. 2013;41:D1228–D1233. doi: 10.1093/nar/gks1147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Szklarczyk D, et al. The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic acids Res. 2017;45:D362–D368. doi: 10.1093/nar/gkw937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Almasy L, Blangero J. Multipoint quantitative-trait linkage analysis in general pedigrees. Am. J. Hum. Genet. 1998;62:1198–1211. doi: 10.1086/301844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Yang J, Lee SH, Goddard ME, Visscher PM. GCTA: a tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011;88:76–82. doi: 10.1016/j.ajhg.2010.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Finucane HK, et al. Partitioning heritability by functional annotation using genome-wide association summary statistics. Nat. Genet. 2015;47:1228–1235. doi: 10.1038/ng.3404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bulik-Sullivan B, et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015;47:1236–1241. doi: 10.1038/ng.3406. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Descriptions of Additional Supplementary Files
Data Availability Statement
The genome-wide summary statistics that support the findings of this study are available via the CHARGE Summary Results portal at the NCBI dbGaP website https://www.omicsdi.org/dataset/dbgap/phs000930 upon publication, or from the corresponding authors R.S. and S.S. upon reasonable request. The summary statistics may be used for all scientific purposes except for the study of potentially sensitive and potentially stereotyping phenotypes such as intelligence and addiction, since this is proscribed by the consent terms for the NHLBI cohorts. Individual level data or study-specific summary results are only available through controlled access. Data for the Framingham Study are available through dbGaP, where qualified researchers can apply for authorization to access (https://www.ncbi.nlm.nih.gov/projects/gap/cgi-bin/study.cgi?study_id=phs000007.v30.p11). Individual level data for the ARIC and CHS studies are also available through dbGaP. Data of European and Australian cohorts are available upon request, in keeping with data sharing guidelines in the EU General Data Protection Regulation. Data from UK Biobank can be accessed at http://www.ukbiobank.ac.uk and for the ENIGMA consortium from medlandse@gmail.com. Individual level data for VETSA is not available due to consent restrictions.