

Abstract
In order to evaluate the genome evolution and systematics, karyotype analysis of mitotic metaphase chromosomes in 51 taxa of Epimedium and two species of Vancouveria was conducted. The 53 taxa were clustered, based on their karyotype similarity coefficients. Results showed that the 53 taxa studied were all diploid with 12 chromosomes (2n = 2x = 12). Each taxon had one pair of satellites located on pair I of homologous chromosomes. Moreover, the karyotype types of the 53 taxa studied were all type 1A or 2A of Stebbins. It can be concluded that the karyotypes between species are indeed very similar and the genome of Epimedium was conservative in evolution. The cluster analysis of karyotype similarity coefficients could provide valuable clues for the systematics and taxonomy of Epimedium. Results of the cluster analysis strongly supported the previous taxonomic division of E. subg. Rhizophyllum and E. subg. Epimedium. The results also showed that the interspecific relationship was closely correlated with geographical distribution in E. subg. Epimedium and the taxa native to east Asia had the highest genetic diversity in Epimedium. Finally, the origin of the modern geographical distribution of Epimedium was inferred. Results of the present study have significant scientific values in further studies on resource utilisation, taxonomy and phylogeny in Epimedium.
Keywords: Cluster analysis, cytogenetics, Ranunculales , similarity coefficient, systematics, Vancouveria
Introduction
Barrenwort, (Epimedium L., Berberidaceae), is an important traditional medicinal plant in China. It is effective in strengthening kidney and curing rheumatism (Guo et al. 2019), widely used in the treatment of osteoporosis, hypertension and coronary heart disease and also used to strengthen immunity and prevent dementia (Guo et al. 2008). Recent studies have verified that Epimedium can inhibit the growth of cancer cells in vitro (Wang et al. 2015). Moreover, Epimedium has become very popular in horticulture due to its excellent characteristics, such as exotic flower shape, variable flower colour, perennial evergreen plant etc. In recent years, studies on Epimedium have become considerably more common (Xu et al. 2019; Qu et al. 2020).
Currently, over 60 species of Epimedium are globally recognised (Qu et al. 2020). China is the main distribution centre, with more than 50 species and six varieties reported (Stearn 2002). The genus ranges from Japan in east Asia to Algeria in north Africa (Ying 2002), with the distribution mainly divided into two regions: 1) the Mediterranean and West Asia; and 2) East Asia. As such, the genus is typical from temperate areas (Ying 2002). For a long time, Epimedium has been neglected in systematics and taxonomy works due to its numerous species of difficult circumscription (Guo et al. 2008; Sheng and Wang 2010; Qu et al. 2020).
Stearn (2002) provided the most comprehensive taxonomic account of Epimedium, based on the geographical distribution, number of leaves on the stem, flower size, petal shape, relative size between petal and sepal and chromosomal C-banding. In the taxonomic system of Stearn (2002), Epimedium was divided into two subgenera: E. subg. Epimedium and E. subg. Rhizophyllum. Epimedium subg. Epimedium was divided into four sections: E. sect. Diphyllon (native to China), E. sect. Macroceras (native to Japan, Korea, north-eastern China and the far east of Russia), E. sect. Polyphyllon (native to West Himalaya) and E. sect. Epimedium (native to Europe, Caucasus and northern Turkey). Epimedium sect. Diphyllon (native to China), the most complex group in the genus, was further divided into four series: E. ser. Campanulatae, E. ser. Davidianae, E. ser. Dolichocerae and E. ser. Brachycerae.
Although Stearn’s (2002) taxonomic system is very comprehensive and the distinction between categories is clear, urgent problems remain unsolved regarding the taxonomy and systematics of Epimedium. For example: 1) the interspecific relationships are still unclear, especially in E. sect. Diphyllon (native to China); and 2) China is the diversity centre of this genus, but the process of diversification is still unclear, especially regarding the modern discontinuous distribution (Nakai et al. 1996; Sun et al. 2004; 2014; Guo et al. 2019; Sheng and Wang 2010).
Karyotype analysis has been extensively used in plant taxonomy and systematics. It can provide significant cytological data for the studies on origin, evolution and interspecific relationship in plants (Sheng and Chen 2007a). Since the 1980s, many cytological works have been conducted in Epimedium. Krikorian et al. (1983) reported the karyotypes of 11 Epimedium species native to Japan. Tanaka and Takahashi (1981) and Takahashi (1989) reported the chromosome C-banding of 26 Epimedium taxa, successively. The comparative study on karyotypes of 18 Epimedium species was carried out by Sheng and Chen (2007a, b). These results showed that all Epimedium were diploids with 12 chromosomes (2n = 2x = 12). All the karyotypes were symmetric to Stebbins’ type 2A or 1A and similar to each other with one pair of middle satellite chromosomes. It can be seen that differences between homologous chromosomes were insufficient and karyotypes between species were very similar. Despite many relevant studies on the karyology of Epimedium (Sheng et al. 2010; Yan et al. 2016; Zhang et al. 2018), all results further confirmed that the karyotypes are very similar and a traditional analysis could not provide more valuable cytological evidence for studies on taxonomy and systematics.
In the past 20 years, authors of the present study have collected 51 Epimedium and conducted their karyotype analysis of mitotic metaphase chromosomes in root tips. However, traditional karyotype analysis is very limited in studies focusing on the taxonomy and systematics of Epimedium since the karyotypes are very similar between the species. In the present study, the cluster analysis of karyotype similarity coefficients in the 53 taxa was conducted to try to provide cytological evidence for further studies on systematics and evolution of the genus.
Materials and methods
Materials studied
Fifty-three experimental taxa, including 51 Epimedium taxa and two Vancouveria species (the most closely-related genus) were collected from China, Japan, Germany and the United States. The details, including collection location and voucher specimens of these experimental materials, are detailed in Table 1. All the voucher specimens were conserved in the Institute of Karst Research, Guizhou Normal University. Parts of materials also were cultivated in a greenhouse of the Institute.
Table 1.
The location and vouchers of the material studied.
| No. | Taxon | location | Voucher |
|---|---|---|---|
| Epimedium | |||
| subgenus Epimedium | |||
| Section Diphyllon (native to China) | |||
| Series Campanulatae | |||
| 1 | E. ecalcaratum G.Y.Zhong | Baoxing, Sichuan, China | 2008060210BX |
| 2 | E. shuichengense S.Z.He | Liupanshui, Guizhou, China | 2009022501LPS |
| 3 | E. platypetalum K.Meyer | Hanzhong, Shanxi, China | 2016030201HZ |
| Series Davidianae | |||
| 4 | E. davidii Franch. | Kunming, Yunnan, China | 2007070301 KM |
| 5 | E. pauciflorum K.C.Yen | Maoxian, Sichuan, China | 2008060210MX |
| 6 | E. flavum Stearn | Tianquan, Sichuan, China | 2014050902TQ |
| 7 | E. ilicifolium Stearn | Zhenping, Shanxi, China | 2015040501ZP |
| 8 | E. mikinorii Stearn | Enshi, Hubei, China | 2016030102ES |
| Series Dolichocerae | |||
| 9 | E. membranaceum K.Meyer | Shunso Garden, Japan | 2015032005SH |
| 10 | E. lishihchenii Stearn | Yishan, Tongren, Guizhou, China | 2008030201YS |
| 11 | E. acuminatum Franch. | Kaiyang, Guizhou, China | 2004050101KY |
| 12 | E. wushanense T.S.Ying | Kaili, Guizhou, China | 2007030201KL |
| 13 | E. leptorrhizum Stearn | Wuchuan, Guizhou, China | 2009020101WC |
| 14 | E. baojingense Q.L.Chen & B.M.Yang | Baojing, Hunan, China | 2008100601BJ |
| 15 | E. chlorandrum Stearn | Baoxing, Sichuan, China | 2008060208BX |
| 16 | E. luodianense M.Y.Sheng | Luodian, Guizhou, China | 2007050505LD |
| 17 | E. pudingense S.Z.He | Kaiyang, Guizhou, China | 2007110405KY |
| 18 | E. glandulosopilosum H.R.Liang | Wushan, Chongqing, China | 2014050201WS |
| 19 | E. pseudowushanense B.L.Guo | Leishan, Guizhou, China | 2014050601LS |
| 20 | E. franchetii Stearn | Badong, Hubei, China | 2015030201BD |
| 21 | E. enshiense B.L.Guo & Hsiao | Enshi, Hubei, China | 2016030101ES |
| 22 | E. sutchuenense Franch. | Zhuxi, Hubei, China | 2016030201ZX |
| 23 | E. zhushanense K.F.Wu & S.X.Qian | Zhushan, Hubei, China | 2016030302ZS |
| Series Brachycerae | |||
| 24 | E. pubescens Maxim. | Mt. Qingcheng, Sichuan, China | 2008030201QC |
| 25 | E. sagittatum (Sieb. & Zucc.) Maxim. | Guiyang Arboretum, Guizhou, China | 2007052001GY |
| 26 | E. sagittatum var. glabratum T.S.Ying | Enshi, Hubei, China | 2016030105ES |
| 27 | E. dolichostemon Stearn | Lichuan, Hubei, China | 2008110502LH |
| 28 | E. truncatum H.R.Liang | Dayong, Hunan, China | 2015041901DY |
| 29 | E. brevicornu Maxim. | Taibai, Shanxi, China | 2015042001TB |
| 30 | E. myrianthum Stearn | Jiangkou, Guizhou, China | 2016060301JK |
| 31 | E. stellulatum Stearn | Shiyan, Hubei, China | 2015032602SY |
| 32 | E. fargesii Franch. | Chengkou, Chongqing, China | 2015031503CK |
| 33 | E. elachyphyllum Stearn | Songtao, Guizhou, China | 2013060201ST |
| Section Macroceras (native to Japan, Korea, northeastern China, and the far east of Russia) | |||
| 34 | E. koreanum Nakai | Tonghua, Jilin, China | 2012030201TH |
| 35 | E. grandiflorum Morr. | Kochi, Japan | 2016062503KO |
| 36 | E. grandiflorum var. thunbergianum (Miq.) Nakai | Miyagi, Japan | 2016092530MI |
| 37 | E. grandiflorum var. higoense T.Shimizu | Kumamoto, Japan | 2015100102KU |
| 38 | E. grandiflorum var. coelestre (Nakai) T.Shimizu | Gunma, Japan | 2016120201GU |
| 39 | E. sempervirens Nakai | Fukui, Japan | 2016102320FU |
| 40 | E. sempervirens var. hypoglaucum (Makino) Ohwi | Ishikawa, Japan | 2015100205IS |
| 41 | E. sempervirens var. multifoliolatum T.Shimizu | Nara, Japan | 2016051002NA |
| 42 | E. trifoliatobinatum (Koidz.) Koidz | Kochi, Japan | 2012050301KO |
| 43 | E. diphyllum (Morren et Decne.) Lodd. | Hiroshima, Japan | 2011080602HI |
| 44 | E. cremeum Nakai & F.Maek | Iwate, Japan | 2011060502IW |
| 45 | E. kitamuranum Yamanaka | Tokushima, Japan | 2010080502TO |
| 46 | E. setosum Koidz | Okayama,, Japan | 2010080508OK |
| Section Epimedium (native Europe, Caucasia, and northern Turkey) | |||
| 47 | E. alpinum L. | Munich, Germany | 2008121401MU |
| 48 | E. pubigerum (DC.) Morren & Decne. | Munich, Germany | 2008121501MU |
| Subgenus Rhizophyllum (native to Caucasia and North Africa) | |||
| 49 | E. pinnatum subsp. colchicum Boiss | Arnold Arboretum, USA | 2013060501AR |
| 50 | E. pinnatum cv. “Elegans” | ||
| 51 | E. perralderianum Coss. | Arnold Arboretum, USA | 2013060508AR |
| Genus Vancouveria | |||
| 52 | V. hexandra (Hook.) Morren & Decne. | Arnold Arboretum, USA | 2013060509AR |
| 53 | V. chrysantha Greene | Arnold Arboretum, USA | 2013060510AR |
Preparation of chromosome spreads and karyotype parameters
The preparation method of chromosome slides referred to the method of Sheng and Chen (2007a). Chromosome slides obtained were examined and photographed by a tri-ocular microscope with a CCD camera (BX52-DP72, Olympus, Japan). At least ten well-spread metaphase plates of each experimental taxa were selected to calculate the karyotype parameters. Long arm (L), short arm (S), total chromosome length (TCL), relative length (%), arm ratio (L/S), arm index and location of the centromere were measured and calculated. RLR (the relative length ratio of the longest and shortest chromosome) and P (the proportion of chromosomes with arm ratio over 2:1) were also calculated. The classification of chromosome type was conducted according to Levan et al. (1964) and the karyotype type was determined by Stebbins’ (1971) criteria.
Calculation and cluster analysis of karyotype similarity coefficients
The calculation formula of karyotype similarity coefficients referred to Tan and Wu (1993):
(1)
(2)
(3)
In formula (3), D is the sum of distances and d is the square root of the product of inner distance (di) and outer distance (dn):
(4)
(5)
(6)
(7)
According to the above formulae, the karyotype similarity coefficients of mitotic metaphase chromosomes of all the 53 experimental taxa were calculated. Furthermore, the 53 experimental taxa were clustered, based on the karyotype similarity coefficients by the UPGMA method (Tan and Wu 1993). The cluster analysis was conducted and the dendrogram was drawn by the NTSYS-pc software (Version 2.10e).
Results
General characteristics of karyotype
All 51 studied Epimedium taxa are diploid with 12 chromosomes (2n = 2x = 12). Pair I of homologous chromosomes in each studied taxon has one pair of satellites. The mitotic metaphase chromosomes of the 51 Epimedium taxa studied were illustrated in Fig. 1 (Material 1–24) and Fig. 2 (Material 25–51), respectively. The karyotype parameters of chromosome length, arm ratio, centromere index, RLR, P and karyotype are all detailed in Table 2. The average lengths of the six pairs of homologous chromosomes were 9.82, 9.04, 8.59, 8.02, 7.56 and 6.98 μm, respectively. The average arm ratios of the six pairs of homologous chromosomes were 1.28, 1.31, 1.44, 1.98, 2.12 and 2.06, respectively. In all 51 Epimedium taxa studied, the majority of chromosomes were metacentric (m) chromosomes or submetacentric (sm) chromosomes. Subtelocentric (st) chromosomes were few and telocentric (t) chromosomes were not found. The average values of RLR and P of the 51 taxa studied were 1.41 and 0.33, respectively. According to Stebbins’ (1971) classification, the karyotypes of all Epimedium taxa tested were highly symmetrical with type 2A or 1A. It also can be seen that the karyotypes between species were very similar in Epimedium species.
Figure 1.

Mitotic metaphase chromosomes in 24 Epimedium species. 1E. ecalcaratum2E. shuichengense3E. platypetalum4E. davidii5E. pauciflorum6E. flavum7E. ilicifolium8E. mikinorii9E. membranaceum10E. lishihchenii11E. acuminatum12E. wushanense13E. leptorrhizum14E. baojingense15E. chlorandrum16E. luodianense17E. pudingense18E. glandulosopilosum19E. pseudowushanense20E. franchetii21E. enshiense22E. sutchuenense23E. zhushanense24E. pubescens. Scale bars: 5 μm.
Figure 2.
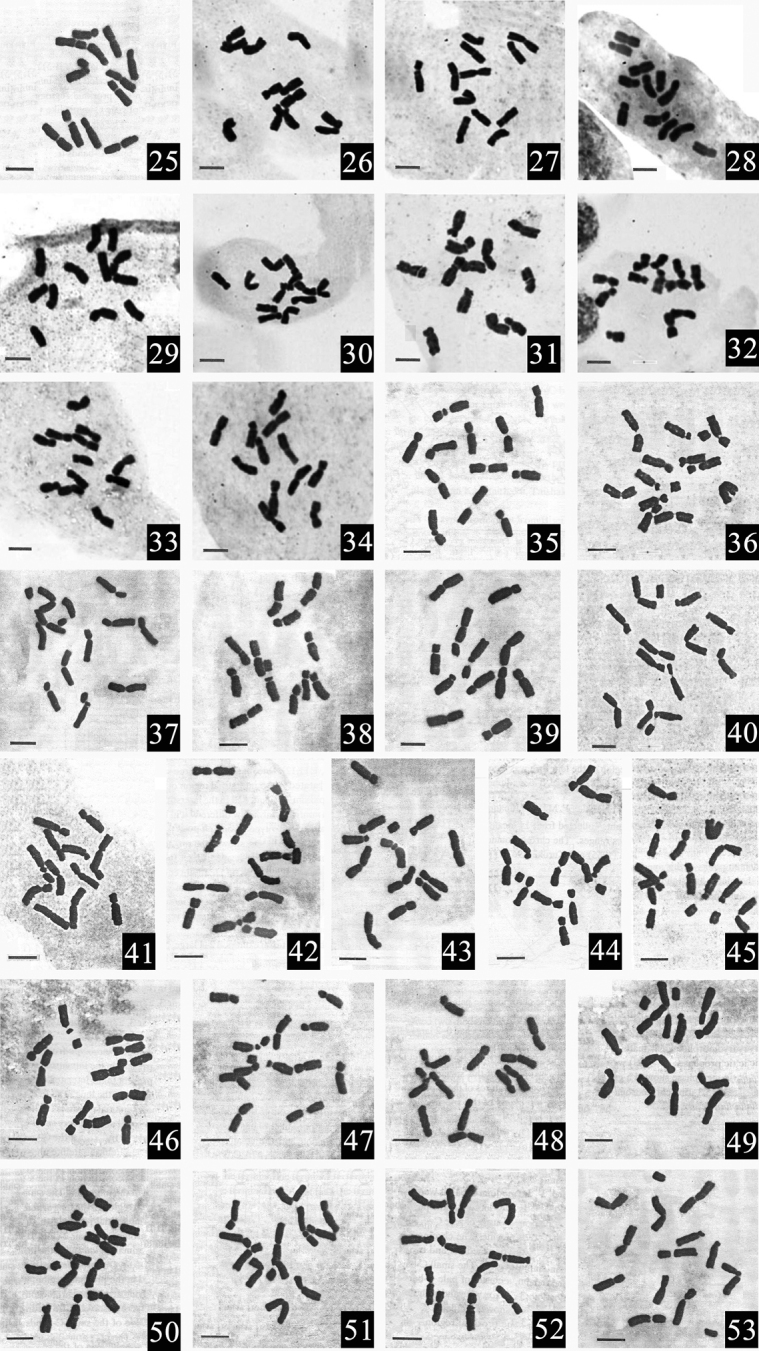
Mitotic metaphase chromosomes in 27 Epimedium taxa and two Vancouveria species. 25E. sagittatum26E. sagittatum var. glabratum27E. dolichostemon28E. truncatum29E. brevicornu30E. myrianthum31E. stellulatum32E. fargesii33E. elachyphyllum34E. koreanum35E. grandiflorum var. grandiflorum36E. grandiflorum var. thunbergianum37E. grandiflorum var. higoense38E. grandiflorum var. coelestre39E. sempervirens40E. sempervirens var. hypoglaucum41E. sempervirens var. multifoliolatum42E. trifoliatobinatum43E. diphyllum44E. cremeum45E. kitamuranum46E. setosum47E. alpinum48E. pubigerum49E. pinnatum subsp. colchicum50E. pinnatum cv. “Elegans” 51E. perralderianum52V. hexandra53V. chrysantha. Scale bars: 5 μm.
Table 2.
Karyomorphological features of mitotic metaphase chromosomes in 51 Epimedium taxa and two Vancouveria species.
| Materials No. | Chromosome length (μm) | Arm ratio | Centromeric index | RLR | P | KC | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | ||||
| 1 | 9.39 | 9.03 | 8.53 | 7.93 | 7.69 | 7.44 | 1.75 | 1.55 | 2.18 | 2.61 | 1.17 | 2.21 | 36.36 | 39.19 | 31.43 | 27.69 | 46.03 | 31.15 | 1.26 | 0.50 | 2A |
| 2 | 9.79 | 8.87 | 8.53 | 8.52 | 8.07 | 6.22 | 2.15 | 1.20 | 1.64 | 1.55 | 2.33 | 1.84 | 31.76 | 45.45 | 37.84 | 39.19 | 30.00 | 35.19 | 1.57 | 0.33 | 2A |
| 3 | 9.86 | 9.15 | 8.31 | 8.08 | 7.48 | 7.13 | 1.02 | 1.20 | 1.26 | 2.58 | 2.00 | 2.16 | 49.40 | 45.45 | 44.29 | 27.94 | 33.33 | 31.67 | 1.38 | 0.33 | 2A |
| 4 | 9.40 | 9.29 | 9.18 | 7.52 | 7.30 | 7.30 | 1.30 | 1.40 | 1.68 | 2.58 | 2.14 | 1.75 | 43.53 | 41.67 | 37.35 | 27.94 | 31.82 | 36.36 | 1.29 | 0.33 | 2A |
| 5 | 9.26 | 9.14 | 8.79 | 7.96 | 7.60 | 7.25 | 1.52 | 1.41 | 1.24 | 2.53 | 2.05 | 2.21 | 39.74 | 41.56 | 44.59 | 28.36 | 32.81 | 31.15 | 1.28 | 0.50 | 2A |
| 6 | 10.22 | 8.93 | 8.30 | 7.98 | 7.45 | 7.12 | 1.82 | 1.63 | 3.11 | 1.27 | 2.68 | 2.72 | 35.42 | 38.10 | 24.36 | 44.00 | 27.14 | 26.87 | 1.44 | 0.50 | 2A |
| 7 | 9.51 | 9.38 | 9.12 | 7.94 | 7.55 | 6.51 | 1.35 | 1.32 | 1.41 | 1.77 | 2.05 | 1.78 | 42.47 | 43.06 | 41.43 | 36.07 | 32.76 | 36.00 | 1.46 | 0.17 | 2A |
| 8 | 9.49 | 9.39 | 8.22 | 7.83 | 7.63 | 7.44 | 1.31 | 1.13 | 1.63 | 2.20 | 2.71 | 3.00 | 43.30 | 46.88 | 38.10 | 31.25 | 26.92 | 25.00 | 1.28 | 0.50 | 2A |
| 9 | 10.14 | 8.87 | 8.48 | 7.60 | 7.50 | 7.41 | 1.04 | 1.33 | 1.29 | 3.11 | 3.05 | 2.04 | 49.04 | 42.86 | 43.68 | 24.36 | 24.68 | 32.89 | 1.37 | 0.50 | 2A |
| 10 | 9.76 | 8.70 | 8.47 | 8.35 | 7.53 | 7.18 | 1.59 | 1.55 | 1.32 | 2.23 | 1.56 | 2.21 | 38.55 | 39.19 | 43.06 | 30.99 | 39.06 | 31.15 | 1.36 | 0.33 | 2A |
| 11 | 10.00 | 9.30 | 8.70 | 7.90 | 7.30 | 6.80 | 1.22 | 1.21 | 2.48 | 1.26 | 2.32 | 2.58 | 45.00 | 45.16 | 28.74 | 44.30 | 30.14 | 27.94 | 1.47 | 0.50 | 2A |
| 12 | 9.90 | 8.79 | 8.30 | 8.02 | 7.55 | 7.42 | 1.05 | 1.22 | 1.39 | 1.71 | 2.21 | 2.16 | 48.75 | 45.07 | 41.79 | 36.92 | 31.15 | 31.67 | 1.33 | 0.33 | 2A |
| 13 | 9.28 | 9.00 | 8.86 | 8.17 | 7.75 | 6.92 | 1.09 | 1.03 | 1.06 | 1.19 | 1.24 | 1.78 | 47.76 | 49.23 | 48.44 | 45.76 | 44.64 | 36.00 | 1.34 | 0.00 | 1A |
| 14 | 9.75 | 8.86 | 8.48 | 7.85 | 7.72 | 7.34 | 1.41 | 1.41 | 2.05 | 1.30 | 3.07 | 1.76 | 41.56 | 41.43 | 32.84 | 43.55 | 24.59 | 36.21 | 1.33 | 0.33 | 2A |
| 15 | 10.52 | 8.33 | 8.06 | 7.93 | 7.65 | 7.51 | 1.41 | 1.10 | 1.68 | 2.05 | 1.67 | 1.62 | 41.56 | 47.54 | 37.29 | 32.76 | 37.50 | 38.18 | 1.40 | 0.17 | 2A |
| 16 | 10.83 | 8.46 | 8.31 | 8.30 | 8.01 | 6.08 | 1.28 | 2.17 | 1.67 | 1.43 | 1.84 | 2.42 | 43.84 | 31.58 | 37.50 | 41.07 | 35.19 | 29.27 | 1.78 | 0.33 | 2A |
| 17 | 9.39 | 8.68 | 8.43 | 8.31 | 7.59 | 7.58 | 1.69 | 1.06 | 1.26 | 2.14 | 2.00 | 1.86 | 37.18 | 48.61 | 44.29 | 31.88 | 33.33 | 34.92 | 1.24 | 0.17 | 2A |
| 18 | 9.68 | 9.45 | 9.22 | 8.43 | 7.52 | 5.70 | 1.50 | 1.68 | 1.79 | 1.96 | 2.14 | 1.17 | 40.00 | 37.35 | 35.80 | 33.78 | 31.82 | 46.00 | 1.70 | 0.17 | 2A |
| 19 | 9.28 | 9.27 | 8.82 | 8.70 | 6.96 | 6.95 | 1.29 | 1.05 | 2.04 | 1.78 | 2.16 | 2.33 | 43.75 | 48.75 | 32.89 | 36.00 | 31.67 | 30.00 | 1.33 | 0.50 | 2A |
| 20 | 10.43 | 9.01 | 8.75 | 7.82 | 7.11 | 6.87 | 1.15 | 1.17 | 1.00 | 2.14 | 2.16 | 2.41 | 46.59 | 46.05 | 50.00 | 31.82 | 31.67 | 29.31 | 1.52 | 0.50 | 2A |
| 21 | 9.23 | 8.78 | 8.43 | 8.42 | 7.74 | 7.39 | 1.05 | 2.04 | 1.28 | 1.35 | 1.68 | 2.37 | 48.75 | 32.89 | 43.84 | 42.47 | 37.31 | 29.69 | 1.25 | 0.33 | 2A |
| 22 | 9.41 | 8.77 | 8.76 | 8.66 | 7.36 | 7.03 | 1.23 | 1.31 | 1.53 | 1.29 | 1.34 | 1.95 | 44.83 | 43.21 | 39.51 | 43.75 | 42.65 | 33.85 | 1.34 | 0.00 | 1A |
| 23 | 10.73 | 8.45 | 8.32 | 7.91 | 7.90 | 6.70 | 1.05 | 1.86 | 1.00 | 2.47 | 1.81 | 1.78 | 48.75 | 34.92 | 50.00 | 28.81 | 35.39 | 36.00 | 1.60 | 0.17 | 2A |
| 24 | 9.50 | 9.23 | 8.40 | 8.21 | 8.12 | 6.55 | 1.10 | 1.56 | 2.14 | 1.34 | 2.14 | 2.23 | 47.57 | 39.00 | 31.87 | 42.70 | 31.82 | 30.99 | 1.45 | 0.50 | 2A |
| 25 | 10.04 | 8.61 | 8.41 | 8.40 | 7.75 | 6.79 | 1.06 | 1.31 | 1.05 | 2.52 | 1.89 | 2.38 | 48.57 | 43.33 | 48.86 | 28.41 | 34.57 | 29.58 | 1.48 | 0.33 | 2A |
| 26 | 9.18 | 8.99 | 8.98 | 8.42 | 7.56 | 6.88 | 1.46 | 1.19 | 1.09 | 1.10 | 1.82 | 1.32 | 40.63 | 45.74 | 47.87 | 47.73 | 35.44 | 43.06 | 1.33 | 0.00 | 1A |
| 27 | 10.46 | 9.76 | 8.35 | 7.64 | 6.94 | 6.84 | 1.00 | 1.16 | 1.02 | 1.17 | 2.14 | 1.06 | 50.00 | 46.39 | 49.40 | 46.05 | 31.88 | 48.53 | 1.53 | 0.17 | 2A |
| 28 | 9.97 | 9.47 | 8.85 | 7.85 | 7.61 | 6.23 | 1.05 | 1.17 | 1.15 | 1.25 | 2.21 | 1.78 | 48.75 | 46.05 | 46.48 | 44.44 | 31.15 | 36.00 | 1.60 | 0.17 | 2A |
| 29 | 9.75 | 9.26 | 8.48 | 7.70 | 7.61 | 7.22 | 1.22 | 1.44 | 2.00 | 1.55 | 1.36 | 1.24 | 45.00 | 41.05 | 33.33 | 39.24 | 42.31 | 44.59 | 1.35 | 0.00 | 1A |
| 30 | 9.04 | 9.03 | 8.48 | 8.47 | 7.49 | 7.48 | 1.00 | 1.65 | 1.20 | 2.50 | 1.96 | 2.24 | 50.00 | 37.80 | 45.45 | 28.57 | 33.82 | 30.88 | 1.21 | 0.33 | 2A |
| 31 | 9.84 | 9.11 | 8.93 | 7.58 | 7.31 | 7.22 | 1.27 | 1.06 | 1.06 | 2.36 | 2.52 | 1.76 | 44.04 | 48.51 | 48.48 | 29.76 | 28.40 | 36.25 | 1.36 | 0.33 | 2A |
| 32 | 10.31 | 8.68 | 8.67 | 8.66 | 7.38 | 6.32 | 2.03 | 1.31 | 1.11 | 1.96 | 1.86 | 1.84 | 32.95 | 43.24 | 47.30 | 33.78 | 34.92 | 35.19 | 1.63 | 0.17 | 2A |
| 33 | 9.03 | 8.59 | 8.48 | 8.37 | 8.26 | 7.27 | 1.41 | 1.69 | 1.41 | 1.17 | 1.88 | 1.75 | 41.46 | 37.18 | 41.56 | 46.05 | 34.67 | 36.36 | 1.24 | 0.00 | 1A |
| 34 | 9.67 | 9.25 | 8.84 | 7.93 | 7.19 | 7.10 | 1.44 | 1.20 | 1.18 | 1.74 | 2.48 | 2.07 | 41.03 | 45.54 | 45.79 | 36.46 | 28.74 | 32.56 | 1.36 | 0.33 | 2A |
| 35 | 9.57 | 9.01 | 8.78 | 7.84 | 7.83 | 6.96 | 1.33 | 1.24 | 1.09 | 1.68 | 2.54 | 2.03 | 42.98 | 44.74 | 47.75 | 37.37 | 28.28 | 32.95 | 1.38 | 0.33 | 2A |
| 36 | 10.18 | 9.19 | 8.12 | 7.96 | 7.58 | 6.97 | 1.18 | 1.07 | 2.03 | 1.12 | 2.54 | 2.14 | 45.86 | 48.33 | 33.02 | 47.12 | 28.28 | 31.87 | 1.46 | 0.50 | 2A |
| 37 | 9.85 | 9.16 | 8.70 | 7.63 | 7.56 | 7.10 | 1.22 | 1.35 | 1.19 | 1.86 | 2.54 | 2.00 | 44.96 | 42.50 | 45.61 | 35.00 | 28.28 | 33.33 | 1.39 | 0.17 | 2A |
| 38 | 9.53 | 9.36 | 8.86 | 7.69 | 7.36 | 7.19 | 1.15 | 1.20 | 1.36 | 1.97 | 2.52 | 2.44 | 46.49 | 45.54 | 42.54 | 33.70 | 28.41 | 29.10 | 1.33 | 0.33 | 2A |
| 39 | 10.09 | 8.74 | 8.67 | 7.78 | 7.77 | 6.95 | 1.01 | 1.13 | 1.11 | 2.35 | 2.15 | 2.32 | 49.63 | 47.01 | 47.41 | 29.81 | 31.73 | 30.11 | 1.45 | 0.50 | 2A |
| 40 | 9.75 | 9.04 | 8.54 | 7.75 | 7.74 | 7.17 | 1.09 | 1.07 | 1.16 | 2.27 | 2.09 | 2.45 | 47.79 | 48.41 | 46.22 | 30.56 | 32.41 | 29.00 | 1.36 | 0.50 | 2A |
| 41 | 9.48 | 9.11 | 8.74 | 8.15 | 7.70 | 6.82 | 1.33 | 1.37 | 1.11 | 2.55 | 1.97 | 2.17 | 42.97 | 42.28 | 47.46 | 28.18 | 33.65 | 31.52 | 1.39 | 0.33 | 2A |
| 42 | 9.71 | 9.57 | 9.14 | 7.50 | 7.35 | 6.71 | 1.16 | 1.13 | 1.17 | 2.28 | 2.32 | 2.24 | 46.32 | 47.01 | 46.09 | 30.48 | 30.10 | 30.85 | 1.45 | 0.50 | 2A |
| 43 | 10.31 | 9.02 | 8.94 | 7.89 | 7.00 | 6.85 | 1.17 | 1.29 | 1.27 | 2.38 | 2.48 | 1.66 | 46.09 | 43.75 | 44.14 | 29.59 | 28.74 | 37.65 | 1.51 | 0.33 | 2A |
| 44 | 10.52 | 9.10 | 8.22 | 8.05 | 7.89 | 6.20 | 1.11 | 1.22 | 2.29 | 1.08 | 1.97 | 2.50 | 47.33 | 45.13 | 30.39 | 48.00 | 33.67 | 28.57 | 1.70 | 0.33 | 2A |
| 45 | 9.96 | 8.92 | 8.85 | 8.11 | 7.22 | 6.93 | 1.14 | 1.05 | 1.45 | 3.40 | 1.97 | 2.24 | 46.67 | 48.76 | 40.83 | 22.73 | 33.67 | 30.85 | 1.44 | 0.33 | 2A |
| 46 | 9.66 | 9.34 | 8.14 | 7.99 | 7.67 | 7.19 | 1.28 | 1.13 | 1.08 | 3.00 | 2.31 | 2.21 | 43.80 | 47.01 | 48.04 | 25.00 | 30.21 | 31.11 | 1.34 | 0.50 | 2A |
| 47 | 10.65 | 9.92 | 8.13 | 7.16 | 7.15 | 7.00 | 1.08 | 1.10 | 1.04 | 2.26 | 2.52 | 2.07 | 48.09 | 47.54 | 49.00 | 30.68 | 28.41 | 32.56 | 1.52 | 0.50 | 2A |
| 48 | 9.68 | 9.27 | 8.46 | 8.13 | 7.64 | 6.83 | 1.29 | 1.07 | 1.04 | 2.45 | 2.76 | 2.36 | 43.70 | 48.25 | 49.04 | 29.00 | 26.60 | 29.76 | 1.42 | 0.50 | 2A |
| 49 | 10.18 | 8.47 | 8.32 | 7.95 | 7.57 | 7.51 | 1.11 | 1.07 | 1.29 | 2.06 | 2.29 | 2.16 | 47.45 | 48.25 | 43.75 | 32.71 | 30.39 | 31.68 | 1.36 | 0.50 | 2A |
| 50 | 9.60 | 8.88 | 8.80 | 8.08 | 7.60 | 7.04 | 1.03 | 1.27 | 1.29 | 2.48 | 1.97 | 2.03 | 49.17 | 44.14 | 43.64 | 28.71 | 33.68 | 32.95 | 1.36 | 0.33 | 2A |
| 51 | 9.94 | 8.89 | 8.43 | 7.76 | 7.61 | 7.38 | 1.24 | 1.41 | 1.29 | 2.68 | 1.59 | 2.38 | 44.70 | 41.53 | 43.75 | 27.18 | 38.61 | 29.59 | 1.35 | 0.33 | 2A |
| 52 | 9.47 | 9.10 | 8.95 | 8.28 | 7.17 | 7.01 | 1.31 | 1.07 | 1.31 | 1.64 | 1.82 | 1.69 | 43.31 | 48.36 | 43.33 | 37.84 | 35.42 | 37.23 | 1.35 | 0.00 | 1A |
| 53 | 9.82 | 9.12 | 8.42 | 7.93 | 7.43 | 7.30 | 1.03 | 1.13 | 1.07 | 1.22 | 2.03 | 1.48 | 49.29 | 46.92 | 48.33 | 45.13 | 33.02 | 40.38 | 1.35 | 0.17 | 2A |
Note: RLR, the relative length ratio of the longest and shortest chromosome; P, proportion of chromosomes with arm ratio over 2:1; KC, karyotype classification of Stebbins (1971).
The two Vancouveria species studied also were diploid with 12 chromosomes (2n = 2x = 12). Furthermore, pair I of homologous chromosomes of each species also has one pair of satellites located. The mitotic metaphase chromosomes and the karyotype parameters of the two species studied are given in Fig. 2 (Material 52, 53) and Table 2 (Material 52, 53), respectively. The average lengths of the six pairs of homologous chromosomes were 9.65, 9.11, 8.69, 8.11, 7.30 and 7.16 μm, respectively. Furthermore, the average arm ratios of the six pairs of homologous chromosomes were 1.17, 1.10, 1.19, 1.43, 1.93 and 1.59, respectively. In the two Vancouveria species studied, only two chromosomal types, i.e. m and sm types, were found. The average RLR of the two species studied was 1.41. The P values of the two species studied were 0.00 and 0.17, respectively. Moreover, the karyotypes of the two species studied were 1A and 2A, respectively. It also can be seen that the karyotypes of the two Vancouveria species were very similar.
Cluster analysis of the karyotype similarity coefficients
The karyotype similarity coefficients of the 53 taxa studied were calculated (Suppl. material 1: Table S1). The 53 taxa are clustered into two groups, mostly corresponding to the two genera (Fig. 3). However, some species of Epimedium, i.e. E. truncatum, E. leptorrhizum and E. dolichostemon were clustered into the group of genus Vancouveria.
Figure 3.

Diagram of cluster analysis of karyotype similarity coefficients in 51 Epimedium taxa and two Vancouveria species.
Clustering results also showed Epimedium, firstly, split into two groups, E. subg. Epimedium and E. subg. Rhizophyllum. Epimedium subg. Epimedium is further split into two clusters, basically reflecting the geographical distribution, with one group mainly consisting of the taxa native to the Mediterranean and West Asia and the majority of the species of the other cluster native to east Asia. The clustering result also showed that the genetic diversity of Epimedium taxa native to east Asia was higher than those of Mediterranean and West Asian taxa. Finally, some species native to east Asia, i.e. E. brevicornu, E. trifoliatobinatum and E. flavum, were clustered into the group of the Mediterranean and West Asian taxa.
The karyotype similarity coefficients between the original species and its variant were significant, such as E. sempervirens var. sempervirens and E. sempervirens var. hypoglaucum, E. sempervirens var. multifoliolatum, E. sagittatum var. sagittatum and E. sagittatum var. glabratum and E. grandiflorum var. grandiflorum, E. grandiflorum var. thunbergianum, E. grandiflorum var. higoense and E. grandiflorum var. coelestre. It can be seen that the cluster analysis of karyotype similarity coefficients can provide reliable clues for studies on plant taxonomy and systematics, especially for those taxa with similar karyotypes between species and insufficiency differences between homologous chromosomes.
Discussion and conclusion
Relationships of Epimedium and related genera
Results of karyotype analysis showed that the genome of Epimedium was conservative in evolution and highly similar between species. The 51 Epimedium taxa tested are all diploid with the basic chromosomal number of 6 (2n = 2x = 12) and each of them has one pair of satellites located on pair I of homologous chromosomes. These results are consistent with the previous research reported (Tanaka and Takahashi 1981; Takahashi 1989; Yan et al. 2016; Zhang et al. 2018). Karyotypes of the 51 taxa studied are all highly symmetrical with the type 2A or 1A of Stebbins (1971). There has been the conclusion that species with symmetrical karyotypes usually are ancient and primitive in evolution and the karyotypes of evolutionary taxa are asymmetrical in spermatophytes (Stebbins 1971; Stace 2000). The highly symmetrical karyotypes in Epimedium species indicate that Epimedium should be conservative in evolution, consistent with other studies on the morphology (Stearn 2002; Xu et al. 2019), molecular biology (Kim and Jansen 1998; Kim et al. 2004), C-banding (Tanaka and Takahashi 1981; Takahashi 1989) and rDNA chromosomal location (Sheng and Wang 2010) in this genus. However, for a long time, studies on the systematics and taxonomy of Epimedium by using the traditional karyotype analysis have achieved little, because of the highly similar karyotypes between species and insufficient differences between homologous chromosomes (Guo et al. 2008).
Epimedium is phylogenetic related to Vancouveria (Tishler 1902; Berg 1972; Loconte and Estes 1989; Jin et al. 2018), being the species belonging to Vancouveria once classified into Epimedium (Stearn 1938). In the present study, results showed that the karyotypes of Epimedium and Vancouveria were highly similar. Karyotypes of the two Vancouveria species studies, with one pair of satellites located on pair I of homologous chromosomes, are very similar with some species of Epimedium. The two Vancouveria species, i.e. V. hexandra and V. chrysantha, were clustered into the group with E. truncatum, E. leptorrhizum and E. dolichostemon in the clustering of karyotype similarity coefficients. When compared with Diphylleia Michaux, Dysosma R. E. Woodson, Podophyllum L., and Sinopodophyllum Ying, the karyotype of Epimedium is significantly different and more symmetrical (Li 1986; Ma and Hu 1996), suggesting that the genus might be an ancient taxon in Berberidaceae and distantly related to these four genera. This conclusion also can be well supported by studies on morphology (Li et al. 2014), palynology (Zhang and Wang 1983; Wang et al. 2015), molecular markers (Wang et al. 2001; Sun et al. 2005; Zhang et al. 2014, 2016), isozymes (Sheng et al. 2011) and chemotaxonomy (Koga et al. 1991; Sheng et al. 2008). Therefore, it can be seen that karyotype analysis has important significance for studies on the relationships of Epimedium within Berberidaceae.
Relationships of Epimedium infrageneric categories and species
Cluster analysis of karyotype similarity coefficients showed that, although the genome was conservative in evolution and the karyotypes between species were highly similar, the cluster analysis of karyotype similarity coefficients still provided some valuable clues for studies on phylogenetics and taxonomy in Epimedium. Clustering results of karyotype similarity coefficients strongly support the classification of the two subgenera of E. subg. Rhizophyllum and E. subg. Epimedium. Based on morphological characteristics and geographical distribution, E. Subg. Epimedium is clustered into two groups: 1) Mediterranean and Western Asian taxa and 2) East Asian taxa. The present results supported this classification, consistent with the previous studies on the morphology (Stearn 2002), cytology (Sheng et al. 2010; Zhang et al. 2018), molecular biology (Kim et al. 2004; Guo et al. 2018; Sajad et al. 2018) and phytochemistry (Guo et al. 2008), all of which confirmed the conclusion that the systematic relationship between species is closely related to the specific geographical distribution in this genus. The present results also showed that, although the vast majority of East Asia taxa could be clustered into one group, there are a few species clustered into other groups of Epimedium or the group of Vancouveria, indicating that the genetic diversity of East Asian taxa are the highest in the genus.
Acknowledgements
We gratefully acknowledge the Support Plan for Excellent Young Science and Technology talents of Guizhou Province (Qiankehe Pingtai Rencai [2017]5638), the Support Plan for Science and Technology Top-notch Talents of Guizhou Higher Education Institutions (Qianjiaohe KY zi [2016]064), the Key Project of Guizhou Science and Technology Fund (Qiankehe Jichu [2020]1Z012) and the Key Project of Guizhou Cooperation Plan for Science and Technology (Qiankehe LH zi [2015]7779) for their financial support.
Citation
Wang L-J, Gao M-D, Sheng M-Y, Yin J (2020) Cluster analysis of karyotype similarity coefficients in Epimedium (Berberidaceae): insights in the systematics and evolution. PhytoKeys 161: 11–26. https://doi.org/10.3897/phytokeys.161.51046
Funding Statement
Guizhou Provincial Science and Technology Department Guizhou Provincial Education Department
Supplementary materials
Table S1. Karyotype resemblance-near coefficients in 51 Epimedium taxa and two Vancouveria species
This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Lin-Jiao Wang, Meng-Di Gao, Mao-Yin Sheng, Jie Yin
Data type
cytogenetic data
References
- Berg RY. (1972) Dispersal ecology of Vancouveria (Berberidaceae). American Journal of Botany 59(2): 109–122. 10.1002/j.1537-2197.1972.tb10069.x [DOI] [Google Scholar]
- Guo BL, Pei LK, Xiao PG. (2008) Further research on taxonomic significance of flavonoids in Epimedium (Berberidaceae). Journal of Systematics and Evolution 46: 874–885. [Google Scholar]
- Guo MY, Li R, Xu YQ, Liao BS, Song JY, Li Y, Mantri N, Guo BL, Chen SL, Pang XH. (2019) Development of plastid genomic resources for discrimination and classification of Epimedium wushanense (Berberidaceae). International Journal of Molecular Sciences 20(16): 4003 10.3390/ijms20164003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo M, Xu Y, Ren L, He S, Pang AX. (2018) A systematic study on DNA barcoding of medicinally important genus Epimedium L. (Berberidaceae). Genes 9(12): 637 10.3390/genes9120637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin ZY, Jocelyn ML, Ye WQ, Li P. (2018) The complete chloroplast genome of Vancouveria planipetala, with implication for the phylogeny of Ranunculales. Mitochondrial DNA. Part B, Resources 3(2): 628–629. 10.1080/23802359.2018.1473726 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim YD, Jansen RK. (1998) Chloroplast DNA restriction site variation and phylogeny of the Berberidaceae. American Journal of Botany 85(12): 1766–1788. 10.2307/2446511 [DOI] [PubMed] [Google Scholar]
- Kim YD, Kim SH, Kim CH, Jansen RK. (2004) Phylogeny of Berberidaceae based on sequence of the chloroplast gene ndhF. Biochemical Systematics and Ecology 32(3): 291–301. 10.1016/j.bse.2003.08.002 [DOI] [Google Scholar]
- Koga S, Shoyama Y, Nishioka I. (1991) Studies on Epimedium species: Flavonol glycosides and isozymes. Biochemical Systematics and Ecology 19(4): 315–318. 10.1016/0305-1978(91)90019-V [DOI] [Google Scholar]
- Krikorian AD, Connor SA, Fitter MS. (1983) Chromosome number variation and karyotype stability in cultures and culture-derived plants. In: Evans DA, Sharp WR, Ammirato PV, Yamata Y. (Eds) Handbook Of Plant Cell Culture.Macmillan Publishing Company, New York, 541–571.
- Levan A, Fredga K, Sandlberg A. (1964) Nomenclature for centromeric position on chromosomes. Hereditas 52(2): 201–220. 10.1111/j.1601-5223.1964.tb01953.x [DOI] [Google Scholar]
- Li LC. (1986) A study on the karyotypes and evolution of Dysosma pleiantha with its relatives. Yunnan Zhi Wu Yan Jiu 8: 451–457. [Google Scholar]
- Li C, Zhao J, Li M. (2014) Cladistic analysis of Epimedium Linn. (Berberidaceae) based on morphological characters. Journal of Plant Resources & Environment 23: 111–113. [Google Scholar]
- Loconte H, Estes J. (1989) Phylogenetic systematics of Berberidaceae and Ranunculales (Magnoliidae). Systematic Botany 14(4): 565–579. 10.2307/2419001 [DOI] [Google Scholar]
- Ma SB, Hu ZH. (1996) A karyotypes study on Podophylloideae (Berberidaceae). Yunnan Zhi Wu Yan Jiu 18: 325–330. [Google Scholar]
- Nakai R, Shoyama Y, Shirashi S. (1996) Genetic characterization of Epimedium species using random amplified polymorphic DNA (RAPD) and PCR-restriction fragment length polymorphism (RFLP) diagnosis. Biological & Pharmaceutical Bulletin 19(1): 67–70. 10.1248/bpb.19.67 [DOI] [PubMed] [Google Scholar]
- Qu LL, Huang CY, Li HX, Yang XB, Sun YC, Hu X. (2020) Chemical markers of four species of Epimedium used in drug Yin-Yang-Huo. Biochemical Systematics and Ecology 88: 103983. 10.1016/j.bse.2019.103983 [DOI]
- Sajad AL, Parvaiz HQ, Suphla G. (2018) Genetic diversity of Epimedium elatum (Morren & Decne) revealed by RAPD characterization. Current Biotica 9: 41–46. 10.25081/cb.2018.v9.3843 [DOI] [Google Scholar]
- Sheng MY, Chen QF. (2007a) Karymorphology of twelve species in Epimedium (Berberidaceae). Yunnan Zhi Wu Yan Jiu 29: 309–315. [Google Scholar]
- Sheng MY, Chen QF. (2007b) Karyotype analysis of six Epimedium species native to Guizhou. China. Guihaia 27: 440–443. [Google Scholar]
- Sheng MY, Chen QF, Yang QX. (2008) Variation in icariin and flavonoid contents of barrenwort accessions native to Guizhou, China. Biochemical Systematics and Ecology 36(9): 719–723. 10.1016/j.bse.2008.06.001 [DOI] [Google Scholar]
- Sheng MY, Wang LJ. (2010) Chromosomal localization of 45S and 5S rDNA in 14 species and the implications for genome evolution of genus Epimedium. Plant Systematics and Evolution 290(1–4): 65–73. 10.1007/s00606-010-0349-3 [DOI] [Google Scholar]
- Sheng MY, Wang LJ, Tian XJ. (2010) Karyomorphology of eighteen species of genus Epimedium (Berberidaceae) and its phylogenetic implications. Genetic Resources and Crop Evolution 57(8): 1165–1176. 10.1007/s10722-010-9556-6 [DOI] [Google Scholar]
- Sheng MY, Wang LJ, Xiong KN. (2011) Isozyme diversity and genetic structure of barrenwort (Berberidaceae) populations. Scientia Horticulturae 131: 58–66. 10.1016/j.scienta.2011.09.017 [DOI] [Google Scholar]
- Stace CA. (2000) Cytology and cytogenetics as a fundamental taxonomic resource for the 20th and 21st centuries. Taxon 49(3): 451–477. 10.2307/1224344 [DOI] [Google Scholar]
- Stearn WT. (1938) Epimedium and Vancouveria (Berberidaceae), a monograph. Botanical Journal of the Linnean Society 51(340): 409–535. 10.1111/j.1095-8339.1937.tb01914.x [DOI] [Google Scholar]
- Stearn WT. (2002) The genus Epimedium and other herbaceous Berberidaceae. Timber Press, Portland.
- Stebbins GL. (1971) Chromosomal evolution in higher plants. Edward Arnold Ltd, London.
- Sun Y, Fung KP, Leung PC, Shaw PC. (2005) A phylogenetic analysis of Epimedium (Berberidaceae) based on nuclear ribosomal DNA sequences. Molecular Phylogenetics and Evolution 35(1): 287–291. 10.1016/j.ympev.2004.12.014 [DOI] [PubMed] [Google Scholar]
- Sun Y, Fung KP, Leung PC, Shi D, Shaw PC. (2004) Characterization of medicinal Epimedium species by 5S rRNA gene spacer sequencing. Planta Medica 70(3): 287–288. 10.1055/s-2004-818928 [DOI] [PubMed] [Google Scholar]
- Sun W, Huang WJ, Li ZN, Song C, Liu D, Liu YL, Hayward A, Liu YF, Huang HW, Wang Y. (2014) Functional and evolutionary analysis of the AP1/SEP/AGL6 superclade of MADS-box genes in the basal eudicot Epimedium sagittatum. Annals of Botany 113(4): 653–668. 10.1093/aob/mct301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi C. (1989) Karyomorphological studies on speciation of Epimedium and its allied Vancouveria with special reference to C-bands. Journal of Science of the Hiroshima University 2: 159–269. [Google Scholar]
- Tan YD, Wu CM. (1993) Cluster analysis method of karyotype similarity coefficient. Journal of Genetics and Genomics 20: 305–311. [Google Scholar]
- Tanaka R, Takahashi C. (1981) Comparative karyotype analysis in Epimedium species by C-banding (1) E. sempervirens var. hypoglaucum and E. perralderianum. Shokubutsu Kenkyu Zasshi 56: 17–24.
- Tishler G. (1902) Die Berberidaceen und Podophyllaceen. Botanishe Jahrbucher fur Systematik 31: 596–727. [Google Scholar]
- Wang T, Su YJ, Zhu JM, Fan GK, Chen J. (2001) RAPD analysis on some species of Berberidaceae. Bulletin of Botanical Research 21: 428–43l.
- Wang YW, Sun QW, Yang W, Xu WF, He SZ, Guo BL, Yang XB. (2015) Pollen characteristics and taxonomic significance of Chinese Epimedium medicinal plant. Xibei Zhiwu Xuebao 35: 1367–1377. [Google Scholar]
- Xu YQ, Liu LJ, Liu SX, He YM, Li RQ, Ge F. (2019) The taxonomic relevance of flower colour for Epimedium (Berberidaceae), with morphological and nomenclatural notes for five species from China. PhytoKeys 118: 33–64. 10.3897/phytokeys.118.30268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan FL, He SZ, Xu WF, Li ZH. (2016) Cytotaxonomy of seven species of Epimedium medicinal plants from Chinese. Guihaia 36: 1039–1045. [Google Scholar]
- Ying JS. (2002) Petal evolution and distribution patterns of Epimedium L. (Berberidaceae). Zhiwu Fenlei Xuebao 40: 481–489. [Google Scholar]
- Zhang YJ, Du LW, Liu A, Chen JJ, Wu L, Hu WM, Zhang W, Kim K, Lee SC, Yang TJ, Wang Y. (2016) The complete chloroplast genome sequences of five Epimedium species: Lights into phylogenetic and taxonomic analyses. Frontiers of Plant Science 7: 306. 10.3389/fpls.2016.00306 [DOI] [PMC free article] [PubMed]
- Zhang C, Peng ZB, Nie ZL, Zhang DG, Li KG, Meng Y. (2018) Chromosome numbers and karyotypes of 18 species of Epimedium sect. Diphyllon (Berberidaceae) from central China. Caryologia 71(4): 1–10. 10.1080/00087114.2018.1499482 [DOI] [Google Scholar]
- Zhang JT, Wang PL. (1983) Study on the pollen morphology of the family Berberidaceae. Zhiwu Fenlei Xuebao 21: 130–141. [Google Scholar]
- Zhang YJ, Yang LL, Chen JJ, Sun W, Wang Y. (2014) Taxonomic and phylogenetic analysis of Epimedium L. based on amplified fragment length polymorphisms. Scientia Horticulturae 170: 284–292. 10.1016/j.scienta.2014.02.025 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Karyotype resemblance-near coefficients in 51 Epimedium taxa and two Vancouveria species
This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Lin-Jiao Wang, Meng-Di Gao, Mao-Yin Sheng, Jie Yin
Data type
cytogenetic data