

Abstract
Background
Human cases of Madariaga virus (MADV) infection were first detected during an outbreak in 2010 in eastern Panama, where Venezuelan equine encephalitis virus (VEEV) also circulates. Little is known about the long-term consequences of either alphavirus infection.
Methods
A follow-up study of the 2010 outbreak was undertaken in 2015. An additional survey was carried out 2 weeks after a separate 2017 alphavirus outbreak in a neighboring population in eastern Panama. Serological studies and statistical analyses were undertaken in both populations.
Results
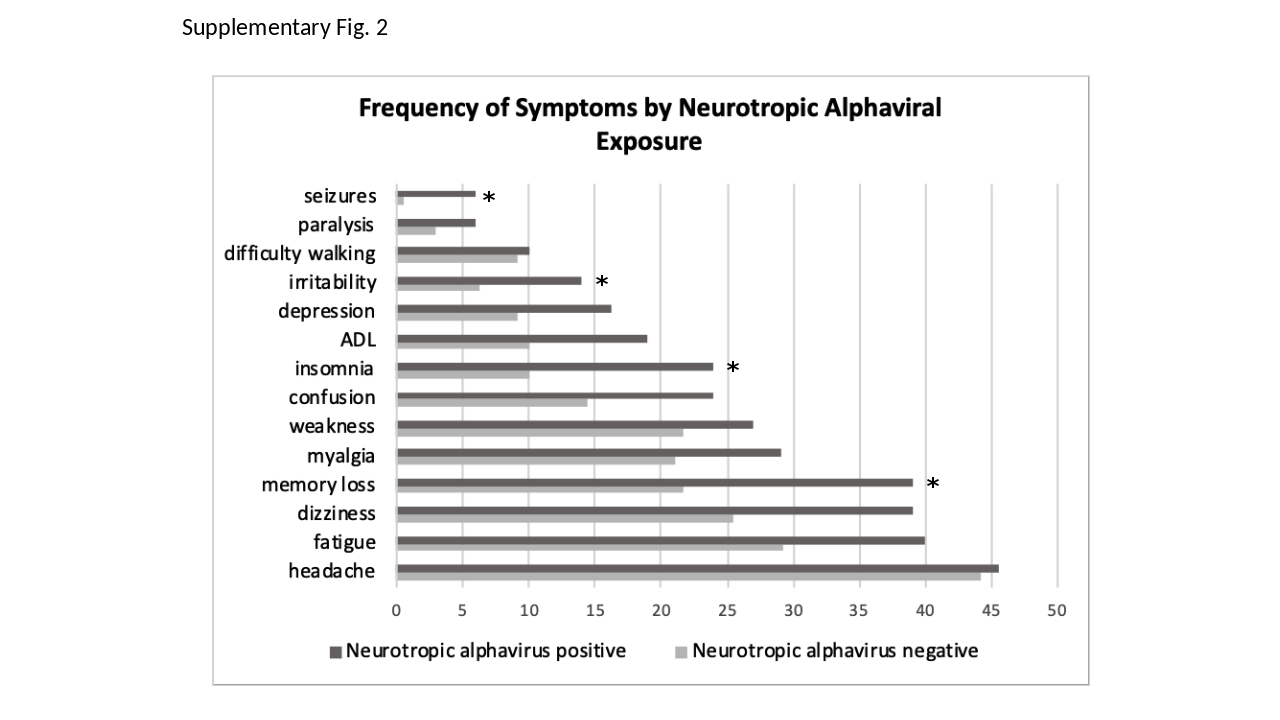
Among the originally alphavirus-seronegative participants (n = 35 of 65), seroconversion was observed at a rate of 14.3% (95% CI, 4.8%–30.3%) for MADV and 8.6% (95% CI, 1.8%–23.1%) for VEEV over 5 years. Among the originally MADV-seropositive participants (n = 14 of 65), VEEV seroconversion occurred in 35.7% (95% CI, 12.8%–64.9%). In the VEEV-seropositive participants (n = 16 of 65), MADV seroconversion occurred in 6.3% (95% CI, 0.2%–30.2%). MADV seroreversion was observed in 14.3% (95% CI, 1.8%–42.8%) of those who were originally seropositive in 2010. VEEV seroconversion in the baseline MADV-seropositive participants was significantly higher than in alphavirus-negative participants. In the population sampled in 2017, MADV and VEEV seroprevalence was 13.2% and 16.8%, respectively. Memory loss, insomnia, irritability, and seizures were reported significantly more frequently in alphavirus-seropositive participants than in seronegative participants.
Conclusions
High rates of seroconversion to MADV and VEEV over 5 years suggest frequent circulation of both viruses in Panama. Enhanced susceptibility to VEEV infection may be conferred by MADV infection. We provide evidence of persistent neurologic symptoms up to 5 years following MADV and VEEV exposure.
Keywords: Madariaga virus, neurological sequelae, Panama, Venezuelan equine encephalitis virus
Madariaga (MADV; formerly known as South American eastern equine encephalitis) and Venezuelan equine encephalitis viruses (VEEV) are single-stranded RNA arthropod-borne zoonotic viruses (Togaviridae: Alphavirus), with circulation throughout much of the Americas [1]. Enzootic subtypes of the VEE antigenic complex are associated with human endemic, and sometimes fatal, infections in the Americas [2]. Human infections with these subtypes occur via spillover from enzootic cycles that involve sylvatic rodents and mosquitoes of the subgenus Culex (Melanoconion). VEEV epizootic/epidemic subtypes (IAB, IC) are associated with large and explosive equine-amplified epidemics in South America, and the available evidence suggests that epizootic strains evolve from enzootic ancestors via mosquito- or equine-adaptive mutations [3]. In Panama, enzootic/endemic VEEV subtype ID infection is highly prevalent in the easternmost province of Darien, resulting in up to 75% seroprevalence in some villages [4]. In this setting, MADV first emerged in the human population in 2010 in the eastern province of Darien, Panama [5]. Enzootic VEEV (subtype ID) was simultaneously circulating in the same area, causing significant neurologic morbidity and mortality [5].
Clinically, most human VEEV infections are symptomatic with dengue or influenza-like illness [6]. VEEV is underdiagnosed in Latin America, where it has been estimated that 0.1%–7% of dengue cases are in fact VEEV infections [2]. In the Americas, 30–50 million dengue infections were estimated to have occurred in 2010 [7]. By extension, then, VEEV could account for 300 000 to 3.5 million cases per year. Around 15% of VEEV cases develop neurologic disease, of which 1% are fatal [2]. VEEV infection manifests most frequently as a self-limited febrile illness, with a smaller portion of cases involving encephalitis, seizures, spontaneous abortions, and gastrointestinal hemorrhage [2, 8–10].
In contrast to VEEV, only 3 human cases of MADV were identified in the Americas before the 2010 outbreak in Panama, despite extensive research and epidemiologic surveillance in enzootic areas [11–13]. Mosquito vectors of MADV are also members of the subgenus Culex (Melanoconion). However, the main reservoir in Latin America is still unknown [14]. We previously reported on the acute clinical presentation, which included a nonspecific febrile illness, meningoencephalitis, and encephalitis usually accompanied by seizures, hemiparesis, and psychomotor retardation [5]. The severe symptoms were primarily seen in children.
As MADV is an emerging virus in Latin America, its long-term sequelae remain unknown. Our literature search revealed little information about long-term neurologic sequelae of enzootic VEEV infections. Following an outbreak in Texas in 1971, Bowen et al. (1976) described signs and symptoms in 86 patients hospitalized with VEEV [6]. None of the affected children reported any sequelae, but 7 of 9 adults examined 9 months later complained of fatigue.
The objective of this study was to estimate seroconversion rates and clinical consequences of MADV and VEEV infections by following up on probable and confirmed cases and their household contacts identified during the 2010 outbreak. We also examine the neurological symptoms of a neighboring population according to alphavirus exposure status. This is the first report, to our knowledge, of the long-term neurological symptoms following MADV, and only the second such report for VEEV.
METHODS
This study was undertaken in the easternmost province of Darien, Panama, which borders Colombia. Alphaviral encephalitis outbreaks were reported in Darien in 2010 [5] and 2017 [15]. A follow-up study of patients (suspected, probable, or confirmed) from the 2010 outbreak and their household contacts was undertaken in 2015. Detailed information on the 2010–2015 cohorts is provided in Tables 1 and 2. An additional population survey was undertaken in 2017 in Mogue, a community in Darien that was not included in the passive surveillance efforts of 2010. Samples were processed at Panama’s reference laboratory, the Gorgas Memorial Institute, located in Panama City.
Table 1.
Characteristics of the Study Populations in the 2010 Outbreak Cohort Studied in 2015 (n = 65) and the Mogue Study in 2017 (n = 243)
| Characteristic | 2015, No. (%) | 2017, No. (%) | |
|---|---|---|---|
| Sex | |||
| Male | 37 (56.9) | 120 (49.4) | |
| Female | 28 (43.1) | 123 (50.6) | |
| Age (tercile), y | |||
| 1–17 | 24 (36.9) | 129 (53.1) | |
| 19–43 | 19 (29.2) | 64 (26.3) | |
| 45–90 | 22 (33.9) | 50 (20.6) | |
| Occupation | |||
| Professional | 3 (4.6) | 8 (3.3) | |
| Technician | 1 (1.5) | 1 (0.4) | |
| Construction worker | 2 (3.1) | 0 (0.0) | |
| Housewife | 15 (23.1) | 56 (23.1) | |
| Student | 15 (23.1) | 93 (38.4) | |
| Rancher | 13 (20.0) | 48 (19.8) | |
| Farmer | 11 (16.9) | 48 (19.8) | |
| Preschool-aged child | 1 (1.5) | 24 (9.9) | |
| Other | 12 (18.5) | 8 (3.3) |
Table 2.
MADV and VEEV Seroprevalence by Survey Year as Determined by Plaque Reduction Neutralization Tests
| Year | Alphavirus Seronegative, No. (%) | MADV Seropositive, No. (%) | VEEV Seropositive, No. (%) | Dual Infection, No. (%) |
|---|---|---|---|---|
| 2010 (n = 65) | 35 (53.8) | 14 (21.5) | 16 (24.6) | 0 |
| 2015 (n = 65) | 29a (44.6) | 18a (27.7) | 24 (36.9) | 6 (9.2) |
Abbreviations: MADV, Madariaga virus; VEEV, Venezuelan equine encephalitis virus.
aThere were 28 alphavirus-seronegative participants, but 1 MADV seroreversion rendered that individual also seronegative. The other individual with MADV seroreversion became VEEV positive in 2015.
Human Survey and Clinical Evaluation in 2015
Participants were located using records from 2010. Upon obtaining written consent, each participant was interviewed and examined by a physician using a demographic and focused medical history questionnaire. The presence or absence of neurological symptoms within the preceding 2 weeks was noted. Blood samples were obtained by peripheral venipuncture. Serum samples were placed in cryogenic tubes for storage in liquid nitrogen and then transported to the Gorgas Memorial Institute for analysis.
Human Survey and Clinical Evaluation in 2017
The survey was carried out from July 18 to 22, 2017, 3 weeks after confirmation of a fatal MADV case on June 30, 2017. A cross-sectional outbreak investigation was undertaken, and blood sampling and epidemiological surveys were carried out including demographic characteristics, potential risk factors, and clinical information. The presence or absence of neurological symptoms within the preceding 2 weeks was evaluated and recorded by a physician. Detailed information on surveys, laboratory testing, risk factors, and serologic results have been previously described [15]. The purpose of this 2017 survey was to describe alphaviral epidemiology and neurological sequelae in a community in which a fatal MADV case was confirmed. The epidemiological results are reported separately [15]. We include the 2017 neurological symptom survey here to validate the findings from our 2010 cohort.
Patient Consent Statement
An outbreak investigation in 2017 was undertaken during a public health outbreak response, and ethical approval for alphavirus and encephalitis surveillance and investigation was obtained from the Gorgas Memorial Institute Committee (IRB, CBI/ICGES/2015 and CIB/ICGES/2018). Study participation was voluntary, and written informed consent was obtained from adults (18 years of age and older) and from parents or guardians of children aged 2 to 17 years. In addition, verbal assent was obtained from children aged 7 to 12 years, and written informed consent was obtained from children aged 13 to 17 years.
Laboratory
Samples were also tested in duplicate for IgM and IgG antibodies against MADV and VEEV using enzyme-linked immunosorbent assays (ELISAs) for recent and past antibody response detection [16]. All samples were also run using virus-specific plaque-reduction neutralization tests (PRNTs). For the PRNT, a positive sample was reported as the reciprocal of the highest dilution that reduced plaque counts by >80% (PRNT80). ELISA antigens were prepared from eastern equine encephalitis virus (EEEV) using the sucrose-acetone technique (prepared by Dr. Robert Shope at the Yale Arbovirus Research Unit in August 1989) and VEE complex strain 78V-3531 from infected mouse brain. Strains used for the PRNT were the chimeric SINV/MADV [17], MADV wild-type strain PA2010-247168 (UTMB/WRCEVA), and TC83, an attenuated vaccine strain of VEEV closely related to the subtype ID strains that circulate in Panama [18].
Case Definition
We deemed IgM-positive samples to represent infection within 12 months. While there are no studies to our knowledge on the persistence of alphavirus IgM, dengue IgM wanes for most people by 1 year [19]. Samples positive for IgM and negative for IgG were classified as recent infection (within 2 weeks). Samples positive for IgG alone indicated past exposure. Due to substantial cross-reactivity between MADV and VEEV by ELISA, virus-specific infection was determined by PRNT80 (a titer of ≥1:20 was determined to be positive). In the event that PRNTs for both viruses were positive, we deemed a 4-fold difference in titers to discriminate between the causative virus vs cross-reactivity. If less than a 4-fold difference was present, we classified this as exposure to both viruses.
Statistical Analysis
Seroconversion rates for both viruses were calculated for each alphaviral exposure group from the 2010 cohort and reported with exact binomial 95% confidence intervals. The probability of seroconversion by alphaviral exposure group was determined by calculating risk ratios with Fisher exact P values given the small sample sizes. To determine whether neurological sequelae were reported at higher frequencies in the alphavirus-exposed group, we conducted a univariate logistic regression analysis, followed by a multivariable logistic regression controlling for sex and age. The outcome variable, alphaviral exposure, was defined as having been exposed to MADV, VEEV, or both viruses using the laboratory criteria described above. P values with alpha <.05 were considered significant. All analyses were undertaken using the statistical package Stata, version 14 (StataCorp, College Station, TX, USA).
RESULTS
Characteristics of the Population
During the original 2010 outbreak, 190 cases with febrile or neuroinvasive disease and 72 household contacts were included. In the 2015 follow-up study, 165 of these participants were excluded due to incomplete baseline information, and an additional 32 cases could not be located. A total of 65 cases were ultimately included in the 2015 follow-up serosurvey (Supplementary Figure 1). Characteristics of the 2015 and Mogue 2017 study populations are described in Table 1. There was a slight preponderance of men compared with women in the 2015 follow-up study, with an even age distribution. The most frequent occupational categories included housewife, student, and farmer. In contrast, the Mogue 2017 population was more heavily weighted toward younger participants (53.1% younger than 17 years), with a correspondingly higher rate of students.
Alphavirus Epidemiological and Serological Profiles
At baseline in 2010, 14 (21.5%) of the 65 participants were seropositive for MADV only, 16 (24.6%) were seropositive for VEEV only, and 0 were seropositive for both as determined by PRNT (Table 3).
Table 3. .
Neutralizing Antibody Titers Measured in 2010 and 2015 Among the 6 Individuals Found to Have Antibodies to Both MADV and VEEV in 2010
| Neutralizing Antibodies | ||||
|---|---|---|---|---|
| 2010 | 2015 | |||
| ID | MADV | VEEV | MADV | VEEV |
| 37 | 1:20 | 1:640 | <1:20 | 1:160 |
| 40 | 1:20 | 1:640 | <1:20 | 1:320 |
| 44 | <1:20 | 1:640 | <1:20 | 1:80 |
| 44a | 1:20 | 1:640 | ||
| 47 | 1:40 | 1:160 | <1:20 | 1:80 |
| 49 | 1:80 | 1:640 | <1:20 | >1:640 |
| 54 | 1:20 | 1:640 | <1:20 | 1:320 |
| 54a | 1:40 | 1:640 |
Abbreviations: MADV, Madariaga virus; VEEV, Venezuelan equine encephalitis virus.
aSecond convalescent serum sample obtained ~15 days after onset of illness.
Between 2010 and 2015, MADV seroconversion occurred in 6 participants. One of these participants was MADV IgM positive (and IgG positive). VEEV seroconversion occurred in 8 participants. One of these participants was VEEV IgM positive (and IgG positive). Among the 35 participants who were alphavirus-seronegative in 2010, seroconversion was observed for MADV in 5 of 35 (14.3%; 95% CI, 4.8%–30.3%) and for VEEV in 3 of 35 (8.6%; 95% CI, 1.8%–23.1%) by 2015 (Table 2).
Among the 14 participants who were MADV-seropositive in 2010, seroconversion to VEEV was observed in 2015 in 5 of 14 (35.7%; 95% CI, 12.8%–64.9%). Only 1 of the 16 participants who were VEEV-seropositive in 2010 seroconverted to MADV by 2015 (6.3%; 95% CI, 0.2%–30.2%). Seroreversion (disappearance of detectable antibodies) was observed for MADV in 2 of 14 participants (14.3%; 95% CI, 1.8%–42.8%). No VEEV seroreversions were observed. The risk ratio for seroconversion to MADV in VEEV-seropositive (in 2010) participants vs previously MADV- and VEEV-seronegative subjects was 0.4 (95% CI, 0.1–3.4; Fisher exact P = .38). The risk ratio for seroconversion to VEEV in MADV-seropositive (in 2010) subjects vs MADV- and VEEV-seronegative subjects was 4.2 (95% CI, 1.1–15.1; Fisher exact P = .03).
A total of 6 participants were found to have neutralizing antibodies to both MADV and VEEV in 2010, though in each instance titers were at least 4-fold greater against VEEV (Table 3). Convalescent serum sample results (>15 days after symptom onset) were obtained for 2 of these participants (ID 44 and 54) in 2010. These demonstrate a rise in MADV titer in relation to the acute samples. Each of these participants with low MADV titers became MADV PRNT80–negative (≥1:20) by 2015 (Table 3).
The mean age of MADV seroconverters was 42.0 years, spanning a large range (10 to 75 years). Most were men (n = 4, 66.7%), and half were cattle ranchers (n = 3, 50.0%, compared with 20.0% of the total sample). Cases were clustered around the township Meteti, located on the Panamerican Highway in the middle of Darien Province. The mean age of VEEV seroconverters (range) was lower, at 14.1 (7–25) years. The cases were evenly distributed between men and women. This group consisted of mostly school-age children (n = 6, 75.0%) and did not include any cattle ranchers or farmers. Three of the participants were also from the township of Meteti; others were scattered throughout the province.
Seroprevalence in the Mogue Cross-Sectional Survey
A total of 243 individuals were surveyed in Mogue, Darien Province, during 2017, with seroprevalence of 31/243 (12.8%) for MADV and 42/243 (17.3%) for VEEV. The MADV-positive participants had a mean age (range) of 35.3 (5–78) years and an equal distribution of men (n = 16) and women (n = 15). Most positive participants were housewives (n = 11, 35.5%), farmers (n = 9, 29%, and students (n = 9, 25.8%). The VEEV-positive participants had a mean age (range) of 37.9 (2–97) years and were mostly women (n = 26, 61.9%). The majority were housewives (n = 19, 45.2%), followed by farmers (n = 12, 28.6%) and students (n = 8, 19.1%).
Neurological Symptoms
Memory loss, dizziness, fatigue, difficulty concentrating, confusion, depression, irritability, myalgia, insomnia, seizures, and impairment in activities of daily living were more frequent in participants exposed to VEEV and/or MADV (Table 4; Supplementary Figure 2). We repeated this survey in a different population in Darien, Panama, in 2017, with roughly similar results (Table 4, right-hand columns). After adjusting for sex and age, the association with prior alphaviral (MADV and/or VEEV) exposure and seizures (OR, 14.5; 95% CI, 1.6–130.1), irritability (OR, 2.7; 95% CI, 1.1–6.3), insomnia (OR, 2.5; 95% CI, 1.3–5.0), and memory loss (OR, 1.9; 95% CI, 1.1–3.3) remained statistically significant (Table 5).
Table 4. .
Frequency of Neurological Signs and Symptoms (%) Reported in 2 Study Populations in Darien, Panama
| Original Outbreak Cohort (n = 65) | 2017 Mogue Study (n = 243) | |||||||
|---|---|---|---|---|---|---|---|---|
| Symptoms | MADV (n = 13) |
VEEV (n = 17) |
MADV & VEEV (n = 7) |
No Alphaviral Exposure (n = 28) |
MADV (n = 22) |
VEEV (n = 33) |
MADV & VEEV (n = 9) |
No Alphaviral Exposure (n = 179) |
| Memory loss | 6 (46) | 8 (47) | 4 (57) | 8 (29) | 8 (36) | 10 (30) | 3 (33) | 37 (21) |
| Headache | 5 (42)a | 7 (41) | 4 (57) | 11 (39) | 9 (41) | 15 (45) | 6 (67) | 80 (45) |
| Dizziness | 5 (38) | 6 (35) | 4 (57) | 5 (18) | 7 (32) | 13 (39) | 5 (56) | 47 (26) |
| Fatigue | 4 (31) | 5 (29) | 3 (43) | 4 (14) | 10 (45) | 14 (42) | 5 (56) | 56 (31) |
| Confusion | 5 (38) | 5 (29) | 1 (14) | 2 (7) | 3 (14) | 8 (24) | 3 (33) | 27 (15) |
| Depression | 2 (17) | 5 (29) | 3 (43) | 3 (11) | 5 (23) | 1 (3) | 1 (11) | 15 (8) |
| Irritability | 4 (31) | 5 (29) | 1 (14) | 1 (4) | 1 (5) | 3 (9) | 1 (11) | 11 (6) |
| Myalgia | 2 (17) | 3 (18) | 3 (43) | 4 (14) | 8 (36) | 9 (27) | 4 (44) | 40 (22) |
| Insomnia | 3 (23) | 4 (24) | 2 (29) | 3 (11) | 3 (14) | 6 (18) | 6 (67) | 18 (10) |
| Difficulty with activities of daily living | 3 (23) | 1 (6) | 1 (14) | 1 (4) | 6 (27) | 6 (18) | 2 (22) | 20 (11) |
| Weakness | 1 (8) | 3 (18) | 1 (14) | 2 (7) | 9 (41) | 9 (27) | 4 (44) | 43 (24) |
| Seizures | 2 (15) | 0 (0) | 0 (0) | 0 (0) | 2 (9) | 2 (6) | 0 (0) | 1 (0.6) |
| Paralysis | 1 (8) | 0 (0) | 0 (0) | 0 (0) | 1 (5) | 4 (12) | 0 (0) | 6 (3) |
| Total | 13 | 17 | 7 | 28 | 22 | 33 | 9 | 179 |
Exposure status reflects seropositivity measured in 2015 for the original outbreak cohort and seropositivity measured in 2017 for the Mogue study. Note participants whose MADV titers seroreverted in 2015 are classified as MADV positive.
Abbreviations: MADV, Madariaga virus; VEEV, Venezuelan equine encephalitis virus.
aOne nonrespondent in this category yielded a denominator of 12.
Table 5. .
Univariate and Multivariable Logistic Regression Analysis of the Presence of Self-Reported Signs and Symptoms by Alphaviral Exposure (n = 308)
| Symptoms | No. (%) | Unadjusted Odd Ratio | 95% CI | P Value | Adjusted Odds Ratioa | 95% CI | P Value |
|---|---|---|---|---|---|---|---|
| Seizures | 7 (2.3) | 13.21 | 1.56–111.3 | .018 | 14.46 | 1.61–130.1 | .017 |
| Paralysis | 12 (3.9) | 2.14 | 0.68–6.83 | .195 | 1.5 | 0.43–5.30 | .525 |
| Difficulty walking | 29 (9.4) | 1.10 | 0.49–2.47 | .808 | 0.87 | 0.36–2.10 | .767 |
| Irritability | 27 (8.8) | 2.44 | 1.10–5.41 | .028 | 2.68 | 1.14–6.27 | .023 |
| Depressionb | 35 (11.4) | 1.92 | 0.94–3.91 | .074 | 1.73 | 0.81–3.65 | .154 |
| Impairment in ADL | 40 (13.0) | 2.08 | 1.06–4.09 | .032 | 1.47 | 0.71–3.03 | .297 |
| Insomnia | 45 (14.6) | 2.81 | 1.48–5.35 | .002 | 2.52 | 1.28–4.97 | .008 |
| Confusion | 54 (17.5) | 1.88 | 1.03–3.41 | .040 | 1.64 | 0.87–3.10 | .128 |
| Weakness | 72 (23.4) | 1.34 | 0.77–2.32 | .298 | 1.18 | 0.66–2.11 | .564 |
| Myalgia | 73 (23.7) | 1.52 | 0.88–2.63 | .131 | 1.10 | 0.61–1.98 | .757 |
| Memory loss | 84 (27.3) | 2.31 | 1.38–3.90 | .002 | 1.89 | 1.09–3.28 | .022 |
| Dizziness | 92 (29.9) | 1.87 | 1.12–3.10 | .016 | 1.48 | 0.87–2.54 | .148 |
| Fatigue | 1001 (32.8) | 1.61 | 0.98–2.65 | .063 | 1.27 | 0.74–2.16 | .376 |
| Headache | 137 (44.6) | 1.05 | 0.65–1.70 | .840 | 0.86 | 0.51–1.45 | .582 |
Abbreviations: ADL, activities of daily living.
aAdjusted by age and sex.
bn = 307.
DISCUSSION
We provide new clinical and epidemiological findings on human infection with MADV and VEEV. Our seroconversion results suggest that MADV has become endemic, with co-circulation of VEEV in eastern Panama. Participants positive for MADV in 2010 had higher rates of VEEV seroconversion than those who were originally seronegative for alphaviruses. On the other hand, participants with prior exposure to VEEV tended to seroconvert to MADV at lower rates than those who were originally alphavirus seronegative. These data suggest that MADV exposure leads to enhanced susceptibility against VEEV, but not vice versa. The mechanism underlying this enhanced susceptibility to VEEV following MADV exposure is unclear. Increased risk of acquiring alphavirus sympatric infections may reflect their similar enzootic habitat and overlapping epidemiological risk of acquisition [5]. However, we have observed differences in MADV and VEEV vector and host usage, as well as different geographic distributions of disease [4, 15]. The seemingly increased susceptibility to VEEV conferred by prior exposure to MADV may also be explained by immune interference. Alphavirus vaccine studies have demonstrated that sequential vaccination with eastern equine encephalitis (EEE) and western equine encephalitis (WEE) vaccines before live-attenuated VEE vaccination resulted in a diminished VEEV-neutralizing antibody response [20, 21]. To our knowledge, this is the first epidemiological study to demonstrate asymmetric human cross-immunity findings, even though these viruses were identified almost a century ago in the 1930s [2,14].
In contrast, VEEV infection appears to generate a robust and durable immune response. Not only do titers remain detectable 5 years after exposure, but the low rate of seroconversion to MADV during this period suggests the presence of cross-protective immunity. The alphaviral vaccine literature provides ample evidence of in vivo examples of cross-protection. For example, hamsters inoculated with an attenuated strain of VEEV experienced a 37% reduction in mortality when subsequently exposed to western equine encephalitis virus (WEEV) and a 59% reduction in mortality when inoculated with EEEV [22]. However, passive transfer of neutralizing antibodies was not shown to confer protection to heterologous viruses [13, 23]. Cross-protection may therefore be attributable to a cellular or humoral response mediated by non-neutralizing antibodies [24].
Seroreversion was documented in 2 of the 14 2010 MADV-positive individuals. It is possible that neutralizing antibody titers waned over time and dropped below our limits of detection. Whether these individuals are newly at risk for MADV infection remains uncertain. In the case of hepatitis B, for example, it was demonstrated that in the face of declining antibody titers following vaccination, hepatitis B infection rates rose in Senegalese children [25].
Memory loss, dizziness, fatigue, difficulty concentrating, confusion, depression, irritability, myalgia, insomnia, and impairment of daily living activities were more frequent in participants exposed to VEEV and/or MADV. After adjusting for sex and age, increased seizures, irritability, insomnia, and memory loss remained statistically significant. Seizures and paralysis were mainly observed in severe cases of MADV encephalitis during the 2010 outbreak. However, seizures were not present in any of the 2010 VEE participants, in contrast to reports of VEE sequelae in children in Colombia [26]. Indeed, on the whole, our 2010–2015 cohort of VEEV-positive participants did not present with severe cases of neurological disease. This may reflect variations in virulence among VEEV strains [27], though there are historical reports of severe and fatal cases of enzootic VEE in Panama [18].
Seven of 9 adults reported fatigue 9 months after a VEEV outbreak in Texas in 1971 [6]. Our results more closely resemble descriptions of sequelae after WEEV infection [28], as well as West Nile virus [29, 30]. Memory loss, learning impairments, and behavioral changes are noted in approximately half of patients after acute illness due to neurotropic alphaviruses [31]. While there does not appear to be any precedent for cognitive testing of participants exposed to alphaviral infections, there have been several such studies in patients who recovered from West Nile virus (WNV). For example, Murray et al. [32] noted that at 8 years from initial infection (neuroinvasive and febrile WNV), 40% of patients still reported sequelae. We suspect that the true burden of MADV and VEEV extends substantially beyond the acute phase of illness. Understanding the full scope of such sequelae is paramount in this region, given the high prevalence of MADV and VEEV exposure.
Interestingly, the majority of MADV- and VEEV-positive individuals in our study did not recall having encephalitis or severe neurological signs/symptoms. Other studies have also suggested that the majority of the encephalitic alphaviral infections present as a self-limited febrile illness [2,33]. Thus, the high rates of self-reported neurological sequelae in this study suggest that long-term neurological sequelae occur even after mild to moderate clinical presentations.
Our study has several limitations. Because this follow-up study was undertaken 5 years after the outbreak, we were not able to determine the timing of seroconversion during the 5-year period before data collection. It is therefore possible that the majority of the seroconversions occurred shortly after the 2010 outbreak. However, we did note that 1 of the participants presented with MADV IgM in 2015, suggesting recent infection. In addition, our sample size was small because our cohort was generated from participants originally tested during an outbreak, and there were many participants who could not be traced. Therefore, our data may not be generalizable. Longitudinal studies of a larger population are important to validate the seroconversion rates and cross-immunity we describe here. Furthermore, there may have been bias in reporting neurological signs and symptoms stemming from the fact that the participants were selected based on their inclusion in the 2010 outbreak studies. We sought to counteract this by including neurological symptom data from a separate cross-sectional study conducted in 2017.
In summary, our results demonstrate that MADV remains in circulation and is an important human pathogen in Panama. We further describe other novel findings, such as the decay in MADV antibodies in some individuals, the possibility of cross-protective immunity conferred by VEEV but not by MADV, and increased susceptibility to VEEV conferred by prior MADV exposure. These findings have implications for vaccine development and merit further study. In addition, we provide preliminary information on the persistence of neurologic symptoms following MADV and VEEV infection. Future investigation into the duration and magnitude of such sequelae, as well as underlying mechanisms and risk factors, may benefit not only those affected by these viruses but also the larger population exposed to similar neurotropic arboviruses.
Supplementary Data
Supplementary materials are available at Open Forum Infectious Diseases online. Consisting of data provided by the authors to benefit the reader, the posted materials are not copyedited and are the sole responsibility of the authors, so questions or comments should be addressed to the corresponding author.
{kind=link}
{kind=link}
Acknowledgments
We are deeply grateful to Mr. Abadia from the Ministry of Health at Taimatí, Darien, for help with tracing participants and navigating the waters of the Panamanian Pacific coast to far-flung villages. We also thank Dr. Sandra Lopez for helping us deliver laboratory results to the cohort participants and Jorge Maguiña for help with database maintenance.
Financial support. This work was supported by the Neglected Diseases Grant from the Ministry of Economy and Finance of Panama to J.M.P. (grant number 1.11.1.3.703.01.55.120) and by the World Reference Center for Emerging Viruses and Arboviruses, National Institutes of Health (grant number AI120942). J.P.C. is funded by the Clarendon Scholarship from the University of Oxford and the Lincoln-Kingsgate Scholarship from Lincoln College, University of Oxford (grant number SFF1920_CB2_MPLS_1293647). C.A.D. acknowledges the MRC Centre, which is jointly funded by the UK Medical Research Council (MRC) and the UK Department for International Development (DFID) under the MRC/DFID Concordat agreement and is also part of the EDCTP2 program supported by the European Union.
Disclaimer. The opinions expressed by authors contributing to this work do not necessary reflect the opinions of the Gorgas Memorial Institute of Health Studies, the Panamanian Government, or the institutions with which the authors are affiliated.
Potential conflicts of interest. All authors: no reported conflicts of interest. All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References
- 1. Navarro J-C, Carrera J-P, Liria J, et al. Alphaviruses in Latin America and the introduction of chikungunya virus. In: Ludert, JE, Pujol F, Arbiza J, eds. Human Virology in Latin America. Cham, Switzerland: Springer International Publishing; 2017:169–92. [Google Scholar]
- 2. Aguilar PV, Estrada-Franco JG, Navarro-Lopez R, Ferro C, Haddow AD, Weaver SC. Endemic Venezuelan equine encephalitis in the Americas: hidden under the dengue umbrella. Future Virol 2011; 6:721–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Powers AM, Oberste MS, Brault AC, et al. Repeated emergence of epidemic/epizootic Venezuelan equine encephalitis from a single genotype of enzootic subtype ID virus. J Virol 1997; 71:6697–705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Vittor AY, Armien B, Gonzalez P, et al. Epidemiology of emergent Madariaga encephalitis in a region with endemic Venezuelan equine encephalitis: initial host studies and human cross-sectional study in Darien, Panama. PLoS Neglect Trop Dis 2016; 10:e0004554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Carrera J-PJ-P, Forrester N, Wang E, et al. Eastern equine encephalitis in Latin America. New Eng J Med 2013; 369:732–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bowen GS, Fashinell TR, Dean PB, Gregg MB. Clinical aspects of human Venezuelan equine encephalitis in Texas. Bull Pan Am Health Organ 1976; 10:46–57. [PubMed] [Google Scholar]
- 7. Bhatt S, Gething PW, Brady OJ, et al. The global distribution and burden of dengue. Nature 2013; 496:504-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Rivas F, Diaz LA, Cardenas VM, et al. Epidemic Venezuelan equine encephalitis in La Guajira, Colombia, 1995. J Infect Dis 1997; 175:828-32. [DOI] [PubMed] [Google Scholar]
- 9. Johnson KM, Shelokov A, Peralta PH, Dammin GJ, Young NA. Recovery of Venezuelan equine encephalomyelitis virus in Panama. A fatal case in man. Am J Trop Med Hyg 1968; 17:432-40. [DOI] [PubMed] [Google Scholar]
- 10. Vilcarromero S, Laguna-Torres VA, Fernández C, et al. Venezuelan equine encephalitis and upper gastrointestinal bleeding in child. Emerg Infect Dis 2009; 15:323–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Corniou B, Ardoin P, Bartholomew C, et al. First isolation of a South American strain of eastern equine virus from a case of encephalitis in Trinidad. Trop Geogr Med 1972; 24:162–7. [PubMed] [Google Scholar]
- 12. Alice FJ. Infeccao humana pelo virus “leste” da encefalite equina. Bol Inst Biol da Bahia (Brazil) 1956; 3:3–9. [Google Scholar]
- 13. Aguilar PV, Robich RM, Turell MJ, et al. Endemic eastern equine encephalitis in the Amazon region of Peru. Am J Trop Med Hyg 2007; 76:293–8. [PubMed] [Google Scholar]
- 14. Arrigo NC, Adams AP, Weaver SC. Evolutionary patterns of eastern equine encephalitis virus in North versus South America suggest ecological differences and taxonomic revision. J Virol 2010; 84:1014–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Carrera JP, Cucunuba ZM, Neira K, et al. Endemic and epidemic human alphavirus infections in Eastern Panama, an analysis of population-based cross-sectional surveys. bioRxiv 901462 [Preprint]. 10 January 2020. Available at: 10.1101/2020.01.10.901462. Accessed 10 January 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Beaty BJ, Calisher CH, Shope RE. Arboviruses. In: Schmidt NJ, Emmons RW, eds. Diagnostic Procedures for Viral, Rickettsial and Chlamydial Infections. Washington, DC: American Public Health Association; 1989:797–855. [Google Scholar]
- 17. Johnson BW, Kosoy O, Wang E, et al. Use of sindbis/eastern equine encephalitis chimeric viruses in plaque reduction neutralization tests for arboviral disease diagnostics. Clin Vaccine Immunol 2011; 18:1486–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Quiroz E, Aguilar PV, Cisneros J, et al. Venezuelan equine encephalitis in Panama: fatal endemic disease and genetic diversity of etiologic viral strains. PLoS Neglect Trop Dis 2009; 3:e472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Chien YW, Liu ZH, Tseng FC, et al. Prolonged persistence of IgM against dengue virus detected by commonly used commercial assays. BMC Infect Dis 2018; 18:156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Pittman PR, Liu CT, Cannon TL, et al. Immune interference after sequential alphavirus vaccine vaccinations. Vaccine 2009; 27:4879–82. [DOI] [PubMed] [Google Scholar]
- 21. Calisher CH, Sasso DR, Sather GE. Possible evidence for interference with Venezuelan equine encephalitis virus vaccination of equines by pre-existing antibody to eastern or western equine encephalitis virus, or both. Appl Microbiol 1973; 26:485–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Cole FE Jr, McKinney RW. Cross-protection in hamsters immunized with group A arbovirus vaccines. Infect Immun 1971; 4:37–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Webb EM, Azar SR, Haller SL, et al. Effects of Chikungunya virus immunity on Mayaro virus disease and epidemic potential. Scientific Reports 2019; 9:20399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Linn ML, Mateo L, Gardner J, Suhrbier A. Alphavirus-specific cytotoxic T lymphocytes recognize a cross-reactive epitope from the capsid protein and can eliminate virus from persistently infected macrophages. J Virol 1998; 72:5146–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Coursaget P, Leboulleux D, Soumare M, et al. Twelve-year follow-up study of hepatitis B immunization of Senegalese infants. J Hepatol 1994; 21:250–4. [DOI] [PubMed] [Google Scholar]
- 26. León CA. Sequelae of Venezuelan equine encephalitis in humans: a four year follow-up. Int J Epidemiol 1975; 4:131–40. [DOI] [PubMed] [Google Scholar]
- 27. Greene IP, Paessler S, Austgen L, et al. Envelope glycoprotein mutations mediate equine amplification and virulence of epizootic Venezuelan equine encephalitis virus. J Virol 2005; 79:9128–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hollister AC, Longshore WA, Dean BH, Stevens IM. The 1952 outbreak of encephalitis in California; epidemiologic aspects. Calif Med 1953; 79:80–90. [PMC free article] [PubMed] [Google Scholar]
- 29. Murray KO, Nolan MS, Ronca SE, et al. The neurocognitive and MRI outcomes of West Nile virus infection: preliminary analysis using an external control group. Front Neurol 2018; 9:111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Samaan Z, Vaz SM, Bawor M, et al. Neuropsychological impact of West Nile virus infection: an extensive neuropsychiatric assessment of 49 cases in Canada. PLoS One 2016; 11:e0158364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ronca SE, Dineley KT, Paessler S. Neurological sequelae resulting from encephalitic alphavirus infection. Front Microbiol 2016; 7:959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Murray KO, Garcia MN, Rahbar MH, et al. Survival analysis, long-term outcomes, and percentage of recovery up to 8 years post-infection among the Houston West Nile virus cohort. PLoS One 2014; 9:e102953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Bowen GS, Calisher CH. Virological and serological studies of Venezuelan equine encephalomyelitis in humans. J Clin Microbiol 1976; 4:22–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.