

Abstract
Tropical forests vary in composition, structure, and function such that not all forests have similar ecological value. This variability is caused by natural and anthropogenic disturbance regimes, which influence the ability of forests to support biodiversity, store carbon, mediate water yield, and facilitate human wellbeing. While international environmental agreements mandate protecting and restoring forests, only forest extent is typically considered, while forest quality is ignored. Consequently, the locations and loss rates of forests of high ecological value are unknown and coordinated strategies for conserving these forests remain undeveloped. Here, we map locations high in forest structural integrity as a measure of ecological quality, based on recently developed fine-resolution maps of three-dimensional forest structure, integrated with human pressure across the global moist tropics. Our analyses reveal that tall forests with closed-canopies and low human pressure typical of natural conditions comprise half of the global humid or moist tropical forest estate, largely limited to the Amazon and Congo basins. The vast majority of these forests have no formal protection and, given recent rates of loss, are at significant risk. With the rapid disappearance of these ‘best of the last’ forests at stake, we provide a policy-driven framework for their conservation and restoration, and recommend locations to maintain protections, add new protections, mitigate deleterious human impacts, and restore forest structure.
As moist tropical forests continue to be rapidly lost1, there have been increasing calls for conservation efforts to focus on the most valuable remaining forests2. Parties to the Convention on Biological Diversity (CBD)3, the 2030 Agenda for Sustainable Development (2030 Agenda)4, and the UN Framework Convention on Climate Change (UNFCCC)5 have committed to increase forest extent by reducing deforestation and increasing forest restoration. Yet, under these current commitments, only forest extent is typically considered, while forest quality is ignored7. The coming year is a ‘biodiversity super year’ in which Parties to the CBD will agree on a new Global Biodiversity Framework for the next decade. Parties to the UNFCCC will also have a final chance to revise their Nationally Determined Contributions to reduce emissions before the Paris Climate Agreement goes into effect. Both these milestones will impact efforts to advance the nature-based Sustainable Development Goals of the 2030 Agenda. Providing the best science on forest quality in a policy-relevant framework can offer a powerful asset to shape these global commitments on biodiversity and climate change.
The ecological value of forests can be quantified based on the concept of ecological integrity6. This term refers to the composition, structure, and function of an ecosystem in relation to the system’s natural or historical range of variation7. Human pressures can alter these ecosystem properties and reduce ecological integrity8. Thus, evaluation of elements of forest composition, structure, and function relative to reference conditions allows objective assessment of the extent to which forests may support the biodiversity and ecosystem services characteristic of a region9.
In humid tropical forest ecosystems, undisturbed forests (often referred to as primary forests) tend to have tall, multi-story canopies and high variation in plant size, often including large emergent trees10. We refer to such forests as having high structural condition. The positive influence of high structural condition forests on biodiversity, ecological function, and ecosystem services is increasingly well understood11. Such forests provide high microclimate and habitat niche diversity and thus support high species diversity12,13. For example, biodiversity value is 41% lower in degraded forests (including selectively logged forests, secondary forests, and forests converted into various forms of agriculture) than in primary forest across the humid tropics14. Forests of high structural condition are also relatively high in productivity and carbon storage15,16. Primary forests in Brazil, Democratic Republic of the Congo (DRC), and Indonesia were found to be 38–59% taller in canopy height, have 100–183% greater aboveground biomass, and store 279–866% more carbon than other dense tree cover1. Tall multi-storied forests also influence water provisioning, providing higher levels of evapotranspiration that enhance regional precipitation and maintain the conditions for dense humid forests to persist17.
The quality of forests of high structural condition can be reduced by human activities2. Hunting decimates wildlife populations without direct effects on habitat, sometimes leading to ‘empty forests’ that are devoid of vertebrates18. Selective logging and burning can remove trees of high wood density and reduce carbon storage, alter microhabitat and niche diversity, and decrease relative humidity, thus increasing fire incidences19. Human settlements, roads, and clearings create edge effects that can extend well into adjacent forests20. These edge effects include invasive species, livestock grazing, increased fire and wind disturbance, and tree mortality. In sum, the effects of these types of anthropogenic disturbances on biodiversity may exceed that of deforestation alone21.
To identify and conserve forests of high ecological value, monitoring efforts must consider forest structural condition and human-induced disturbances. We recently drew on advances in remote sensing to map forests of high structural condition and low human pressure, termed high structural integrity forests22, across the Tropical and Subtropical Moist Broadleaf Forests (TSMBF) Biome23. We did so by integrating remotely-sensed measures of canopy cover, canopy height, and year of stand-replacing disturbance (referred to as loss year)24 to develop the Structural Condition Index (SCI). The SCI is a globally consistent, fine-scale measure of forest structure and allows identification of taller, older, more closed-canopy humid tropical forests. By integrating the SCI with the Human Footprint Index (HFP)25, we generated a Forest Structural Integrity Index (FSII). Forests with high structural condition and low human pressure are of high ecological value because of the ecological benefits of well-developed forest structure and lack of alteration by human activities.
In this analysis, we apply the SCI and FSII to identify the geographic regions and countries with high representation of forests with high structural condition and integrity. We also summarize the representation of high SCI and FSII forests in protected areas (IUCN I-IV Categories26) and quantify forest loss from 2013–2018. We assess the results in three classes of forest: (1) high FSII (high SCI and low human pressure); (2) high SCI (high SCI but higher human pressure than in Class 1); and (3) low SCI. We then provide a policy-driven framework to conserve these forests through actions to maintain protections, add new protections, mitigate deleterious human impacts, and restore forest structure.
The high SCI and high FSII classes as formulated in this application are scaled to the tallest forests in the biome. The natural structural condition of moist tropical forests varies among ecoregions. Flooded wetland forests may be naturally lower in structural condition than terra firma forests. Our objective was to identify the forests with the most well-developed structure within the TSMBF biome because they are arguably the highest in ecological value (e.g. for carbon storage), and consequently the highest priority for international conservation planning. Conservation planners within countries, in contrast, may choose to scale SCI and FSII to the natural potential of ecoregions within their national boundaries, and develop plans to conserve those forests with the highest structural condition typical of each ecoregion.
Forests of high SCI were defined as >75% canopy cover, >15 m in height, and were not disturbed during 2000–201824. We selected this threshold to represent high structural condition forests based on map accuracy, and representing levels of structure typical of undisturbed natural forests in the biome. In an accuracy assessment, we found that the SCI adequately distinguished forest structure levels up to this threshold level, but the relationship saturated above that level22. We note that some secondary and selectively logged forests have structural characteristics that place them with the high SCI class. In our validation of SCI, we found that approximately 20% of older secondary forests were within the high SCI class22. While such disturbed forests are unlikely to have all of the ecological quality attributes associated with forests that have never undergone human-induced disturbances, current remote sensing capabilities do not allow discriminating them from unlogged primary forests.
We defined high FSII as high SCI and HFP ≤4. This range of HFP values is consistent with the values identified as relevant to responses of vertebrate species endangerment trends to human pressure27. This approach for proportional representation assumes that the TSMBF Biome was entirely forested prior to human disturbance, consistent with the definition of the biome23.
Results and Discussion
Our analyses reveal that of the 67% of the TSMBF biome that remains forested, more than half of these forests are either low in SCI (33%) or are high in SCI but with substantial human pressure (20%) (Figure 1, Supplementary Table 1). Thus, only 47% of the moist tropical forest estate is high in FSII (high SCI and low human pressure). The majority (76%) of the remaining high FSII forest is in the Americas, particularly in the Amazon Basin in the countries of Brazil (45%), Peru (10%), Colombia (6%), and Venezuela (5%) (Figure 2A, Supplementary Table 1). In Africa, just 30% of the remaining moist tropical forest is high in FSII. The DRC and the Republic of the Congo together contain 9% of the global total forest high in FSII. In Melanesia, which harbors 20% of the global high FSII forest, Papua New Guinea supports 3% of the global total. Asia/Melanesia also have large areas of high SCI forests that have potential for high ecological value if human pressure is managed effectively. Importantly, several countries in the TSMBF biome have relatively low levels of humid forest remaining (<50%) and virtually no high FSII forests (e.g. Vietnam, Mozambique, Cote d’Ivoire, Nigeria, and Bangladesh; Supplementary Table 1).
Figure 1.

Distribution of forest types across the Tropical and Subtropical Moist Broadleaf Forest (TSMBF) Biome. Proportion of forest types in the biome is shown in the inset.
Figure 2.

Extent of forest types within countries among the continents (A). Proportion of forest legally protected by forest type among the three continents (B). Loss rates by forest type during 2013–2018 (C).
Strict legal protections extend to only 10% of the remaining forest in the TSMBF biome (Figure 2B, Supplementary Table Table 2). Among these protected forests, 1.7% are low in SCI, 1.8% are high in SCI, and 6.5% are high in FSII. The largest proportion of protected high FSII forests is in South America. However, 86% of the global high FSII forests are not under strict legal protection.
High FSII forest loss rates during 2013–2018 were substantial, but lower than for other forest classes. Some 4.0% of all remaining forests were lost. Rates of loss were 1.3% for high FSII forests, 4.0% for high SCI forests, and 7.5% for low SCI (Figure 2D, Supplementary Table Table 3). Asia/Melanesia had the highest rates of high FSII forest loss (1.7%). Total loss rates (Supplementary Table Table 3) were relatively high in Madagascar (14.6%), Cote d’Ivoire (11.6%), Nicaragua (11.3%), Malaysia (10.0%), and Laos (9.8%).
The relatively lower rates of loss for high FSII forests, compared with other classes, is likely due to their remoteness. These forests are relatively distant from previous development and forest loss (Figure 3), which are known to be strong correlates with deforestation1,28. The forests closer to roads and previous loss were by definition more likely to fall within the high SCI or low SCI classes, largely due to edge effects of human development.
Figure 3.
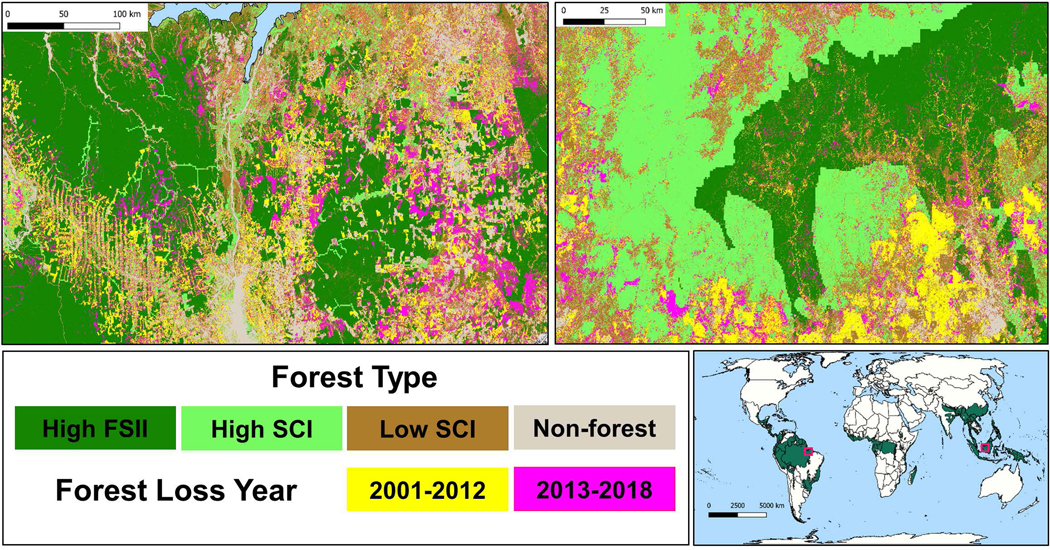
Landscapes showing the distribution of three types of forest and locations of past forest lost during two time periods in the Amazon (top left) and Southeast Asia (top right). These maps illustrate that forest loss during 2013–2018 was located in close proximity to past forest lost (2001–2012) and was biased towards the Low SCI forest that surrounded this past development.
Policy Implications and a Management Framework.
Reducing rates of tropical forest loss, especially that of forests of the highest ecological value, is now critical29. It has been suggested, for example, that forest loss in the Amazon is approaching a tipping point where negative synergies among deforestation, climate change, and human-induced fires may cause forest to flip to degraded savanna30. The fact that remoteness is associated with lower loss rates of high FSII forests has important implications for maintaining these forests. Protection of forests high in FSII may contribute to ensuring natural systems do not cross critical ecological tipping points. Expanding measures to explicitly protect remaining high FSII forests in the post-2020 Global Biodiversity Framework of the CBD and in Nationally Determined Contributions to the Paris Climate Agreement can build political will to limit loss rates of these remaining ‘best of the last’ humid tropical forests. In their implementation of these commitments, it will be important for nations proactively avoid development that increases accessibility to high FSII forests. The current relative isolation of these forests from human development would likely be quickly lost through various proposed mega-road and other economic growth-inducing development schemes31, exposing these forests to dramatically higher rates of loss.
Restoring degraded forests can also play a critical role in nations’ efforts to implement commitments to reduce biodiversity loss under the CBD and mitigate climate change within the UNFCCC. Given the rapid rates of forest regeneration and growth under the climatic conditions within the TSMBF biome, forest structure can be restored within years to decades. Thus, forest restoration strategies are viable options within policy-relevant timeframes. Finally, modification of human behaviors can be effective in mitigating some of the harmful effects of human pressure on humid tropical forests11, with positive implications for nations’ efforts to deliver on the CBD, UNFCCC, and 2030 Agenda. Restoration of forest structure and modification of human behaviors are of particularly high priority in the moist tropical forests of SE Asia and Africa where the majority of forests are either low to moderate in SCI and relatively high in human footprint.
The focus of our analysis on forest structure, human pressure, and protected status provides the basis for a framework prioritizing the conservation of tropical forests (Figure 4). The framework includes four strategies: (1) maintaining current protections, (2) adding new protections, (3) restoring forest structure, and (4) restoring forest integrity by mitigating harmful human activities.
Figure 4.
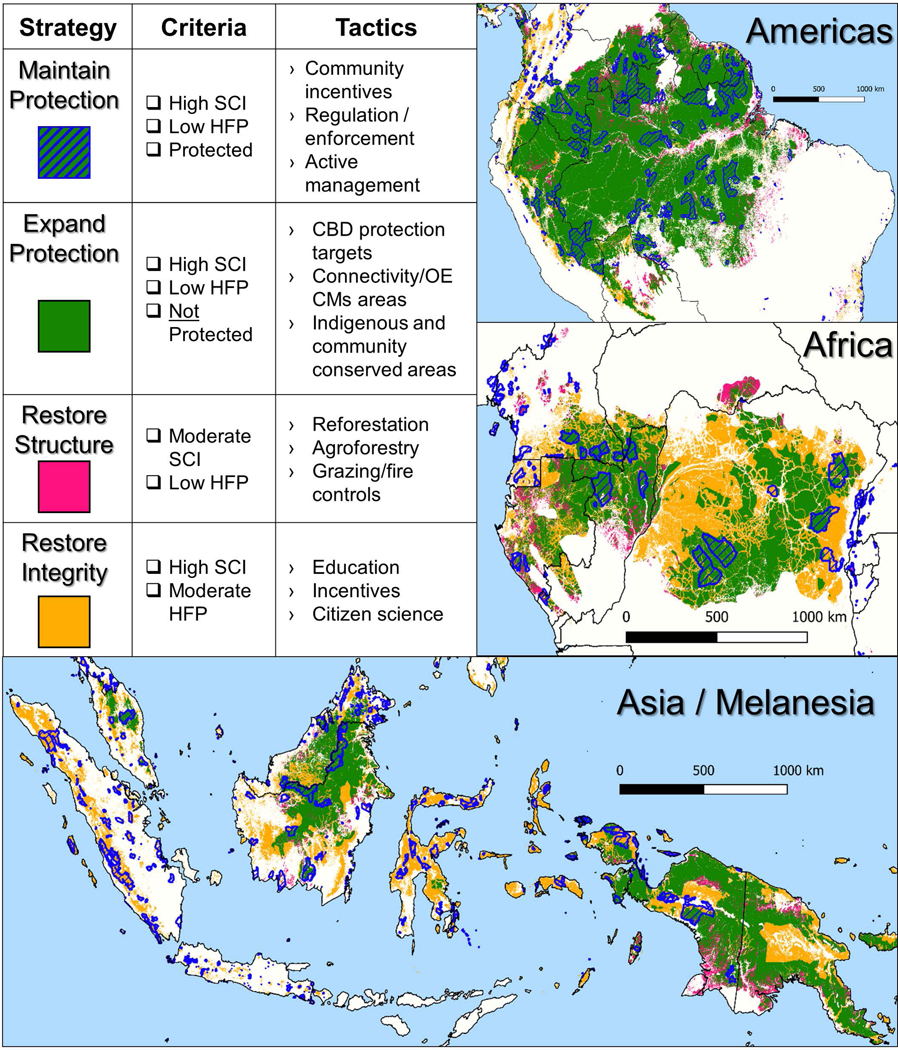
Recommended framework for conservation of tropical forests. Shown are four conservation strategies and associated criteria for mapping, tactics for management, and locations across the biome. Criteria include structural condition (SCI), human footprint (HFP), and protection status.
Maintaining current protections is suggested for forests high in SCI, low in human pressure, and legally protected. Such forests, covering just 6.5% of the forested area (Supplementary Table Table 4), are high in FSII and are at the lowest risk of loss. Nonetheless, these forests face increasing human use and loss of legal protection. Human pressure increased in 55% of protected areas during 1992–2009, and currently 33% of protected land is under intense human pressure32.
In addition to the loss of ecological value, increased human pressure contributes to the widespread loss of legal protections through downgrading, degazettement, and downsizing33,34. Given these threats, protected area agencies can best maintain high FSII forests with incentives, regulation, and active management35. Incentives could include support for sustainable commodity supply chains and resource-generating activities for indigenous and local communities. Regulation could include enforcing current protections, preventing loss of legal protections, and providing titles to traditional lands to increase custodianship by indigenous peoples and local communities. Management could be aimed at reducing human pressure and implementing adaptive ecosystem planning to mitigate land use and climate change effects.
New protections should be considered for the 40.5% of the forest area that is high in FSII and unprotected. Mechanisms for doing so include expansion of current protected areas, formal designation of new protected areas, conservation easements on private lands, and inclusion in CBD-mandated “other effective area-based conservation measures” such as indigenous and community conserved areas (ICCAs) and connectivity conservation areas36–38.
Forest restoration is a high priority for the 6.8% of forest area that is intermediate in SCI and low in human pressure. Many of these high SCII forests are secondary forests recovering after agricultural land abandonment, shifts to agroforestry, and decreases in human pressure due to rural-to-urban migration and civil unrest24. Restoration of structure is also recommended for secondary forests that were classified as high FSII but that may not have the structural attributes of primary forest. There are increasing incentives for forest restoration national habitat compensation programs, carbon accounting, and ecotourism39. Forest restoration is also critical to meeting current CBD targets and mitigating climate change under the UNFCCC. The rapid forest growth rates in this biome result in substantial increases in structural condition within decades40. Techniques for actively restoring tropical forests are increasingly well-developed41. These include reforestation with native species, selective thinning, maintenance of coarse woody debris, management of grazing and fire, and land-owner education on these techniques.
Managing human activities that degrade forests should be considered for the 19.0% of forest area that has high SCI and moderate human pressure. Techniques exist to promote more sustainable fuel-wood gathering, fishing, hunting, forestry, agriculture, and management of livestock11. Such sustainable forest use can be incentivized through carbon credits and other payments for ecosystem services42 and be facilitated through community education programs. Better enforcement of existing laws can likewise contribute to overall ecosystem health by reducing illegal logging and poaching.
Conclusion.
Many countries have lands suitable for one or more of the above four conservation strategies (Supplementary Table Table 4), pointing to the potential for diverse conservation portfolios to meet international commitments to the CBD, UNFCCC, and 2030 Agenda. Opportunities for maintaining current legal protections and adding new legal protections are particularly important in the countries of the Amazon and Congo Basins (strategies 1 and 2). Restoration of forest structure has high potential in the Central African Republic and Gabon in Africa, and New Caledonia in Asia (strategy 3). Opportunity to enhance forest integrity through managing human activities are relatively high in both Africa and Asia/Melanesia (strategy 4). Countries striving to meet existing CBD and UNFCCC targets can use optimization methods43 to best determine how to distribute these four conservation strategies within their boundaries. Global financing mechanisms, particularly those of the UNFCCC, could support these conservation strategies, given the global benefits of these forests as part of natural climate solutions44.
Our conservation framework focuses on identifying the subsets of tropical forest globally that are the highest priorities for each of the four conservation strategies, based on feasibility of achieving effective conservation results in policy-relevant timeframes. In addition to these highest priority conservation locations, the four strategies would likely provide benefits throughout the TSMBF biome. For example, lowland forests in Borneo and Sumatra are not within any of the four conservation strategy zones due to high human pressure and deforestation. Yet these forests are known to be high in biodiversity and have considerable potential for restoration of carbon sequestration and ecosystem function. Therefore, we urge policy makers and forest managers to modify our conservation framework to best suit national and global conservation needs.
Our approach for integrating remotely-sensed global observations to quantify forest structural integrity should be seen as a first step towards quantifying other components of ecological integrity and incorporating them into forest conservation planning. As the big data revolution continues to evolve, it will be possible to integrate remotely-sensed metrics to map ecosystem structure, function, and composition across all forested biomes. The resulting indices of ecological integrity would enhance the current suite of “Essential Biodiversity Variables” used by the international community to monitor and evaluate global ecological change45. Robust measures of ecological integrity would allow full consideration of forest structure, function, and composition for prioritizing specific forest locations for protection and restoration.
There is an urgent need to identify and conserve the remaining high FSII forests before they are completely lost to deforestation and other human disturbances. Managing forests wisely in the TSMBF biome is critical to avoiding dramatic shifts in climate, vegetation, and loss of ecosystem services25. Forest structural integrity can be used in conservation planning to spatially design measures to best maintain and restore ‘best of the last’ forests of high ecological value. Given the high historic and current rates of loss of forest, and the important contributions of these forests to both nature and people, this metric can provide important input into national and international policy processes. As Parties work to develop the CBD’s post-2020 Global Biodiversity Framework, and develop strategies for national implementation, forest structural condition and integrity can provide a key measure to support the delivery of outcomes related to conservation and sustainable use of biodiversity, especially considering ecosystem targets and outcomes are likely to be important. Likewise, sustaining forests high in FSII and restoring forests high in SCI will enhance carbon storage and mitigate climate change, providing a critical strategy that can support the Nationally Determined Commitments of the Paris Climate Agreement5. Forest structural integrity can also guide policymakers to better identify where implementing measures to achieve forest-related nature and climate commitments can advance the nature-based Sustainable Development Goals of the 2030 Agenda. We strongly recommend a framework for prioritizing measures that incorporate ecological value, including forest structural integrity, be incorporated in global efforts to implement the CBD, UNFCCC and 2030 Agenda.
Methods
We conducted the analysis across the Tropical and Subtropical Moist Broadleaf Forests (TSMBF) Biome23 (Supplementary Table Figure 1). This region includes complex gradients in climates, landforms, and soils that influence natural and human disturbance regimes, forest growth rates, and forest structure. For example, in South America, the study area spans from Pacific coastal rain forests eastward across the Andes montane forests, and across the Amazon lowland humid forests. In Africa, humid forests lie between the drier biomes of the Sahel to the north and savanna biomes to the south. Southeast Asia has sharp coastal to mountain gradients that support lowland, wetland, and montane moist forests.
Forests of the TSMBF Biome occur as large, discontinuous patches centered on the equatorial belt and between the Tropics of Cancer and Capricorn46. Covering just 14% of global land area, these forests support at least 50% of the world’s species and more ecoregions than any other biome23. This biome also has among the highest forest growth rates16, carbon storage47, and a disproportionate influence on global climate48. The remarkable variety of human foods and medicinal plants produced by these forests have sustained many indigenous cultures and provided genetic resources for drugs and the breeding stock for cultivated crops globally.
Continental groupings (Americas, Africa, Asia/Melanesia) and countries are the spatial sub-units for our analyses. Country boundaries were obtained from the simplified versions of the Large Scale International Boundaries (LSIB) for Africa and the Americas (http://geonode.state.gov/layers/geonode%3AAfrica_Americas_LSIB7a_gen_polygons) as well as Eurasia and Oceania (http://geonode.state.gov/layers/geonode%3AEurasia_Oceania_LSIB7a_gen_polygons). These boundaries are current as of March 30, 2017. We merged these two shapefiles to form one set of country boundaries. Next, we spatially intersected the country boundaries with the TSMBF biome23 and dissolved single polygons into multipart polygons based on the country name field. We manually removed a small number (area) of polygons in the southern portion of the biome which did not overlap tree height map data. We uploaded the resulting shapefile to Google Earth Engine (GEE)49 and used it for summing area totals of different forest classes and conservation strategies at 30 m spatial resolution. All maps shown in this paper use the Equal Earth projection, an equal-area pseudocylindrical projection for world maps50.
Distribution of high Forest Structural Integrity forests.
Two forest metrics were used in the analysis, Forest Structural Condition Index (SCI) and Forest Structural Integrity Index (FSII)22. The SCI quantifies canopy stature, cover, and disturbance history. The SCI is derived from canopy cover, canopy height, and time since forest loss24,51. The reference year is 2013, with canopy cover from 2010, forest loss expressed as year of loss prior to 2018, and canopy height for 2012. The index ranges from 1 to 1822, with the lowest value assigned to stands less than 5 m tall, disturbed since 2012 or with canopy cover less than 25%. The highest value is for stands not undergoing loss since 2000 that are tall in stature and closed canopy. The index spans from short, open-canopy, recently disturbed forests to tall, closed-canopy stands that have not been disturbed during 2000 to 201822. Forest stature and canopy cover are products of both the biophysical potential of a local site and of disturbance history52,53. The tallest, densest forests are found in settings with favorable climate and soils but with low levels of natural or human disturbance. Our maps of SCI identify locations in the humid tropics of tall, dense forests resulting from high biophysical potential and low disturbance rates. The SCI validated well (R2 = 0.93) against estimates of foliage height diversity derived from airborne lidar across gradients in forest structure from recently disturbed forests, plantations, older secondary forest, and primary forest22.
The FSII is derived from overlaying the Human Footprint Index (HFP) of human pressure54 on the SCI22. The Human Footprint was the first high-resolution (1 km) measure of the cumulative, in-situ pressures humans exert on natural areas across terrestrial Earth55. The dataset was created through the integration of earth observation and survey data sources, including: 1) extent of built environments, 2) crop land, 3) night-time lights 4) navigable waterways, 5) pasture land, 6) human population density, 7) railways, and 8) roads. The original 1993 version was updated to 200954 and more recently to 201325. The data set underwent rigorous validation and has been widely used for global human pressure and biodiversity analyses32,56.
We used the 2013 version of the Human Footprint to coincide with the dates for the forest structure data. For the FSII, human pressure classes and weights were: Low (1), HFP ≤ 4; Medium (5), HFP > 4 and ≤15; and High (10), HFP>15. These threshold values of HFP are consistent with those identified as being highly relevant to responses of vertebrate species endangerment trends to human pressure25. We calculated FSII as:
The resulting values of the FSII range from 0.1 to 18 with the higher values representing forests high in structural complexity and low in human pressure22. Such forests are most likely to not be compromised in either the compositional or functional components of ecological integrity.
The forest types examined in the study were low SCI forest, high SCI forest, and high FSII forest. Low SCI forest was defined as tree cover >25% and canopy height >5 m22. We set SCI ≥14 as the threshold to denote high SCI. These forests are >75% canopy cover, >15 m in height, have not been disturbed during 2000 to 2018. The SCI ≥14 level was selected to represent high structure forests based on map accuracy and based on representing levels of structure typical of undisturbed natural forests in the biome. An accuracy assessment found that the SCI adequately distinguished forest structure levels up to SCI = 14, but the relationship saturated above that level22. High FSII was defined as high SCI and HFP ≤4. This value of HFP was consistent with those identified as being highly relevant to responses of vertebrate species endangerment trends to human pressure27. This approach for proportional representation assumes that the TSMBF Biome was entirely forested prior to human disturbance, consistent with the definition of the biome23.
We summarized the distribution of high FSII forest within the study area based on the proportion of the total areal extent of the forest type that occurred within continental grouping and countries. Because countries differ in the areal overlap within the TSMBF Biome, we also quantified the proportional representation of the forest types of interest within each country. We summed areas using the reduceRegions method in GEE at a spatial resolution of 30 m. See Code and Data Access section below for script access.
Representation in Protected Areas.
We used the IUCN World Database on Protected Areas26,57 to identify protected areas. We specifically included IUCN Categories I-IV, which represent allocations where conservation of biodiversity is a primary objective. We communicated with the World Commission on Protected Areas to ensure that the protected area data were up to date for countries within the study area. We summarized the proportional representation of forest, high SCI forest, and high FSII forest within and outside of protected areas at the national, continental, and global levels using GEE. See Code and Data Access section below for script access.
Loss rates.
Loss was defined as a stand-replacing disturbance, which can lead either to a permeant conversion to non-forest (deforestation) or to a secondary forest of lower structural condition24. While not all the input data for SCI and FSII are updated annually and the index is not time varying, loss rates of low SCI, high SCI and high FSII forests could be estimated for 2013–2018. This was done by defining high SCI forest for the year 2012. To do so, we used the input layers of canopy cover (2010), canopy height (2012) and forest loss prior to 2013 to identify forests >75% canopy cover, >15 m height, and not disturbed during 2000–2012. FSII for 2013 was defined based on high SCI 2012 and HFP 2013. Forest loss for 2013–2018 was then overlaid on the 2012 high SCI and 2013 high FSII forest layers to identify locations where deforestation had occurred. This was also done for forest that was not high in SCI (<14) in 2012 or FSII in 2013. The resulting rates of loss of high SCI, high FSII, and low SCI forests were then summarized by country using GEE. See Code and Data Access section below for script access.
Conservation strategies.
We mapped the four conservation strategies using rules found in Supplementary Table Table 5. The proportional representation of these strategies was then summarized within each country using GEE. See Code and Data Access section below for script access.
Scope and limitations.
SCI and FSII were designed specifically to provide integrated metrics of forest structure and human pressure of use in quantifying aspect of forest ecological quality. These indices overlap with previously published forest metrics that deal with canopy condition and human pressure such as forest intactness, forest loss/gain, large intact landscapes, and hinterland forest (see Table 1 in22). Our formulation is distinct from those previous metrics and was designed for our specific application.
A strength of our approach is that forest structural condition and forest structural integrity are mapped within fine-scale grids (30 m and 1 km, respectively) that can be aggregated to provide for ecological values that are expressed at landscape levels. Individual large trees and small patches of high structural integrity provide habitat for species with small home ranges and contribute to carbon storage and water retention. Thus, conservation benefits accrue from maintaining or restoring fragments of high structure forest within a degraded forest matrix. Other species and ecological processes are optimized in landscapes with large, intact patches of forest. Thus, SCI and FSII data can be used for conservation planning at multiple spatial scales: 30 m to 1 km cells can be used to conserve fine-scale forest features (e.g., small stands of high FSII forest); the data can also be aggregated to landscape scales for conservation planning regarding fragmentation and connectivity of large patches of high FSII forests.
As stated in the Main section of this paper, the SCI and FSII are scaled to the tallest forests in the biome. The natural structural condition of moist tropical forests varies among ecoregions. We identified the most well-developed structural forests in the biome because they are arguably the highest in ecological value (e.g. for carbon storage) and consequently the highest priorities for international conservation planning. We suggest that planners consider both this biome-wide scaled approach and an approach scaled to the natural potential within ecoregions to prioritize forests for conservation.
A current limitation of the SCI and FSII data sets is that they are relatively static in time (centered on 2013). Currently analyses of change are restricted to change since 2013, as we report in this paper. Efforts are underway, however, to update the input data layers to allow change analysis for the 2000–2017 period, which will greatly enhance our ability to use the SCI and FSII framework to evaluate change over a more comprehensive period. A second limitation is that FSII has a coarser 1 km resolution compared with the 30 m SCI. This difference is due to the 1 km resolution of the global human footprint data set used to generate FSII. Some countries, such as Colombia, are now using the human footprint methodology to generate national 30 m maps of human pressure and include them in their reports to the CBD on national progress towards the Aichi Biodiversity Targets. When completed, these data could be used to generate FSII at finer spatial resolution for use in national conservation planning and policy development. A third limitation is that the accuracy of the height data used to generate SCI necessitates height classes being resolved in 5m height intervals. Canopy metrics resulting from the space-station based GEDI Lidar Mission58, which will become available over the next few years, are expected to be much more accurate.
Data Availability
Full details of the forest structural condition and forest structural integrity maps is available as a Data Descriptor[22]. Input and output datasets can be accessed via FigShare: https://figshare.com/account/home#/projects/72164.
Code Availability
The Google Earth Engine code is available at: https://code.earthengine.google.com/625bede18e265d81f6184b27129fecf8.
Supplementary Material
Acknowledgements
The work was funded by the NASA Biodiversity and Ecological Forecasting Program under the 2016 ECO4CAST solicitation through grant NNX17AG51G, the NASA Global Ecosystem Dynamics Investigation (NNL15AA03 to S.G.), and the NASA GEO solicitation (80NSSC18K0338 to P.J.).
Footnotes
Competing Interest Declaration
The authors declare no competing interests.
Additional information
Further information is provided as Supplementary Information.
References
- 1.Turubanova S, Potapov PV, Tyukavina A & Hansen MC Ongoing primary forest loss in Brazil, Democratic Republic of the Congo, and Indonesia. Environ. Res. Lett 13, 074028 (2018). [Google Scholar]
- 2.Watson JEM et al. The exceptional value of intact forest ecosystems. Nat. Ecol. Evol 2, 599–610 (2018). [DOI] [PubMed] [Google Scholar]
- 3.Convention on Biological Diversity (CBD). COP 11 Decision X/2. Strategic Plan for Biodiversity 2011–2020. (2010). [Google Scholar]
- 4.United Nations. Transforming Our World: The 2030 Agenda For Sustainable Development. A/RES/70/1 Resolution adopted by the United Nations General Assembly. (2015). [Google Scholar]
- 5.UNFCCC. Adoption of the Paris Agreement. Proposal by the President. Draft Decision -/CP.21. (2015). [Google Scholar]
- 6.Parks Canada Agency. Parks Canada Guide to Management Planning. (2008). [Google Scholar]
- 7.Parrish JD, Braun DP & Unnasch RS Are we conserving what we say we are? Measuring ecological integrity within protected areas. Bioscience 53, 851–860 (2003). [Google Scholar]
- 8.Anderson JE A conceptual framework for evaluating and quantifying naturalness. Conserv. Biol 5, 347–352 (1991). [Google Scholar]
- 9.Tierney GL, Faber-Langendoen D, Mitchell BR, Shriver WG & Gibbs JP Monitoring and evaluating the ecological integrity of forest ecosystems. Front. Ecol. Environ 7, 308–316 (2009). [Google Scholar]
- 10.Kricher J. Tropical Ecology. (Princeton University Press, 2011). [Google Scholar]
- 11.Lindenmayer DB & Franklin JF Conserving Forest Biodiversity: A Comprehensive Multiscaled Approach. (Island Press, 2002). [Google Scholar]
- 12.Rozendaal DMA et al. Biodiversity recovery of Neotropical secondary forests. Sci. Adv 5, eaau3114 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cortés-Gómez AM, Castro-Herrera F & Urbina-Cardona JN Small changes in vegetation structure create great changes in amphibian ensembles in the Colombian Pacific rainforest. Trop. Conserv. Sci 6, 749–769 (2013). [Google Scholar]
- 14.Gibson L. et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–381 (2011). [DOI] [PubMed] [Google Scholar]
- 15.Poorter L. et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr 24, 1314–1328 (2015). [Google Scholar]
- 16.Running SW et al. A continuous satellite-derived measure of global terrestrial primary production. Bioscience 54, 547–560 (2004). [Google Scholar]
- 17.Bonan GB & Doney SC Climate, ecosystems, and planetary futures: the challenge to predict life in Earth system models. Science 359, eaam8328 (2018). [DOI] [PubMed] [Google Scholar]
- 18.Symes WS, Edwards DP, Miettinen J, Rheindt FE & Carrasco LR Combined impacts of deforestation and wildlife trade on tropical biodiversity are severely underestimated. Nat. Commun 9, 4052 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lindenmayer DB, Laurance WF & Franklin JF Global decline in large old trees. Science 338, 1305–1306 (2012). [DOI] [PubMed] [Google Scholar]
- 20.Pfeifer M. et al. Creation of forest edges has a global impact on forest vertebrates. Nature 551, 187–191 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barlow J. et al. Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 535, 144–147 (2016). [DOI] [PubMed] [Google Scholar]
- 22.Hansen A. et al. Global humid tropics forest structural condition and forest structural integrity maps. Sci. Data 6, 232 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dinerstein E. et al. An ecoregion-based approach to protecting half the terrestrial realm. Bioscience 67, 534–545 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hansen MC et al. High-resolution global maps of 21st-century forest cover change. Science 342, 850–853 (2013). [DOI] [PubMed] [Google Scholar]
- 25.Venter O. et al. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun 7, 12558 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.IUCN and UNEP-WCMC. The World Database on Protected Areas (WDPA) Cambridge, UK. (2019). Available at: https://www.protectedplanet.net/.
- 27.Di Marco M, Venter O, Possingham HP & Watson JEM Changes in human footprint drive changes in species extinction risk. Nat. Commun 9, 4621 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Armenteras D, Espelta JM, Rodríguez N & Retana J. Deforestation dynamics and drivers in different forest types in Latin America: three decades of studies (1980–2010). Glob. Environ. Chang 46, 139–147 (2017). [Google Scholar]
- 29.Lenton TM et al. Climate tipping points - too risky to bet against. Nature 575, 592–595 (2019). [DOI] [PubMed] [Google Scholar]
- 30.Lovejoy TE & Nobre C. Amazon tipping point. Sci. Adv 4, eaat2340 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Laurance WF et al. A global strategy for road building. Nature 513, 229–232 (2014). [DOI] [PubMed] [Google Scholar]
- 32.Jones KR et al. One-third of global protected land is under intense human pressure. Science 360, 788–791 (2018). [DOI] [PubMed] [Google Scholar]
- 33.Golden Kroner RE et al. The uncertain future of protected lands and waters. Science 364, 881–886 (2019). [DOI] [PubMed] [Google Scholar]
- 34.Geldmann J, Manica A, Burgess ND, Coad L & Balmford A. A global-level assessment of the effectiveness of protected areas at resisting anthropogenic pressures. Proc. Natl. Acad. Sci 116, 23209–23215 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.DeFries R, Karanth KK & Pareeth S. Interactions between protected areas and their surroundings in human-dominated tropical landscapes. Biol. Conserv 143, 2870–2880 (2010). [Google Scholar]
- 36.Polak T. et al. Efficient expansion of global protected areas requires simultaneous planning for species and ecosystems. R. Soc. Open Sci 2, 150107 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Garnett ST et al. A spatial overview of the global importance of indigenous lands for conservation. Nat. Sustain 1, 369–374 (2018). [Google Scholar]
- 38.Jonas HD, Barbuto V, Jonas HC, Kothari A & Nelson F. New steps of change: looking beyond protected areas to consider other effective area-based conservation measures. Parks 20, 111–127 (2014). [Google Scholar]
- 39.Brancalion PHS et al. Global restoration opportunities in tropical rainforest landscapes. Sci. Adv 5, eaav3223 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chazdon RL et al. Rates of change in tree communities of secondary Neotropical forests following major disturbances. Philos. Trans. R. Soc. B Biol. Sci 362, 273–289 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chazdon RL Beyond deforestation: restoring forests and ecosystem services on degraded lands. Science 320, 1458–1460 (2008). [DOI] [PubMed] [Google Scholar]
- 42.Venter O. et al. Harnessing carbon payments to protect biodiversity. Science 326, 1368 (2009). [DOI] [PubMed] [Google Scholar]
- 43.Margules CR & Pressey RL Systematic conservation planning. Nature 405, 243–253 (2000). [DOI] [PubMed] [Google Scholar]
- 44.Griscom BW et al. National mitigation potential from natural climate solutions in the tropics. Philos. Trans. R. Soc. B Biol. Sci 375, 20190126 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pereira HM et al. Essential biodiversity variables. Science 339, 277–278 (2013). [DOI] [PubMed] [Google Scholar]
- 46.Olson DM & Dinerstein E. The Global 200: priority ecoregions for global conservation. Ann. Missouri Bot. Gard 89, 199–224 (2002). [Google Scholar]
- 47.Pan Y. et al. A large and persistent carbon sink in the world’s forests. Science 333, 988–993 (2011). [DOI] [PubMed] [Google Scholar]
- 48.Prevedello JA, Winck GR, Weber MM, Nichols E & Sinervo B. Impacts of forestation and deforestation on local temperature across the globe. PLoS One 14, e0213368 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gorelick N. et al. Google Earth Engine: planetary-scale geospatial analysis for everyone. Remote Sens. Environ 202, 18–27 (2017). [Google Scholar]
- 50.Šavrič B, Patterson T & Jenny B. The Equal Earth map projection. Int. J. Geogr. Inf. Sci 33, 454–465 (2019). [Google Scholar]
- 51.Tyukavina A. et al. Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012. Environ. Res. Lett 10, 074002 (2015). [Google Scholar]
- 52.Goetz S & Dubayah R. Advances in remote sensing technology and implications for measuring and monitoring forest carbon stocks and change. Carbon Manag 2, 231–244 (2011). [Google Scholar]
- 53.Hansen AJ, Phillips LB, Dubayah R, Goetz S & Hofton M. Regional-scale application of lidar: variation in forest canopy structure across the southeastern US. For. Ecol. Manage 329, 214–226 (2014). [Google Scholar]
- 54.Venter O. et al. Global terrestrial Human Footprint maps for 1993 and 2009. Sci. Data 3, 160067 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sanderson EW et al. The human footprint and the last of the wild. Bioscience 52, 891–904 (2002). [Google Scholar]
- 56.Tucker MA et al. Moving in the anthropocene: global reductions in terrestrial mammalian movements. Science 359, 466–469 (2018). [DOI] [PubMed] [Google Scholar]
- 57.Dudley N. Guidelines for Applying Protected Area Management Categories. (2008). [Google Scholar]
- 58.Dubayah R. et al. The Global Ecosystem Dynamics Investigation: high-resolution laser ranging of the Earth’s forests and topography. Sci. Remote Sens 1, 100002 (2020). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Full details of the forest structural condition and forest structural integrity maps is available as a Data Descriptor[22]. Input and output datasets can be accessed via FigShare: https://figshare.com/account/home#/projects/72164.