

Abstract
Activity-dependent persistent changes in neuronal intrinsic excitability and synaptic strength are widely thought to underlie learning and memory. Voltage-gated KCNQ/Kv7 potassium channels have been of great interest as the potential targets for memory disorders due to the beneficial effects of their antagonists in cognition. Importantly, de novo dominant mutations in their neuronal subunits KCNQ2/Kv7.2 and KCNQ3/Kv7.3 are associated with epilepsy and neurodevelopmental disorder characterized by developmental delay and intellectual disability. The role of Kv7 channels in neuronal excitability and epilepsy has been extensively studied. However, their functional significance in neural plasticity, learning, and memory remains largely unknown. Here, we review recent studies that support the emerging roles of Kv7 channels in intrinsic and synaptic plasticity, and their contributions to cognition and behavior.
Keywords: KCNQ channel, Kv7, intrinsic excitability, synaptic transmission, neural plasticity, learning, memory, behavior
Introduction
Voltage-gate channel potassium (K+) subfamily Q member 1–5 (KCNQ 1–5) encodes Kv7.1–Kv7.5 channels (Gutman et al., 2005) that are critical regulators of excitability in neurons, muscles, and sensory cells (Soldovieri et al., 2011). All Kv7 subunits have six transmembrane segments (S1–S6; Robbins, 2001). The S1–S4 comprise a voltage-sensing domain with the S4 being the main voltage-sensor (Robbins, 2001). The pore domain consists of the S5-S6 flanking the pore loop important for K+ ion selectivity (Sun and MacKinnon, 2017). The gate is formed by the intersection of four S6 segments (Cui, 2016; Sun and MacKinnon, 2017). Upon depolarization, the electric field on the basic residues of S4 promotes its translational rotation and outward displacement, which leads to the opening of the gate (Cui, 2016; Sun and MacKinnon, 2017). All Kv7 channels require phosphatidylinositol-4,5-bisphosphate (PIP2) in the plasma membrane for channel opening (Zhang et al., 2003; Suh and Hille, 2008; Zaydman and Cui, 2014), and PIP2 is proposed to couple the voltage-sensing domain to the pore domain in Kv7.1 (Zaydman et al., 2013; Sun and MacKinnon, 2020). Each Kv7 subunit also has a short intracellular N-terminal domain and a long intracellular C-terminal tail that harbors four helices (helices A–D; Haitin and Attali, 2008). Helices A and B bind to calmodulin (CaM; Strulovich et al., 2016; Sun and MacKinnon, 2017), whereas helices C–D mediate subunit assembly (Haitin and Attali, 2008).
In neurons, Kv7 channels open at subthreshold potentials around −60 mV and produce slowly-activating and non-inactivating outward K+ currents that potently suppress repetitive and burst firing of action potentials (APs; Brown and Passmore, 2009). Their functional significance in inhibiting neuronal excitability is underscored by the fact that mutations in their subunits cause epilepsy (Nappi et al., 2020), whereas Kv7 agonist retigabine inhibits seizures in rodents and humans (Miceli et al., 2008). Importantly, emerging new evidence suggests that Kv7 channels may contribute to activity-dependent persistent changes in neuronal intrinsic excitability and synaptic strength that are widely thought to underlie learning and memory. This review will summarize the function of Kv7 channels in the hippocampus and discuss recent studies that investigate their contributions to hippocampal plasticity, cognition, and behavior.
Brain Distribution of Kv7 Subunits and Their Channelopathies
Kv7.2, Kv7.3, and Kv7.5 are the major neuronal Kv7 subunits (Table 1). Kv7.2 and Kv7.3 show strong overlapping expression in the cerebral cortex, hippocampal formation, amygdala, basal ganglia, and hypothalamus (Wang et al., 1998; Cooper et al., 2001; Klinger et al., 2011). Kv7.5 is highly expressed in the brain stem and to less extent in the cerebral cortex, hippocampus, occipital, frontal, and temporal lobes (Lerche et al., 2000; Schroeder et al., 2000; Tzingounis et al., 2010; Fidzinski et al., 2015). While Kv7.1 and Kv7.4 are mainly found in the heart and cochlear hair cells, respectively (Wang et al., 1996; Kubisch et al., 1999), they are also detected at low level in multiple regions of the brain (Casimiro et al., 2001; Hansen et al., 2006; Goldman et al., 2009; Su et al., 2019; Table 1).
Table 1.
Distribution of Kv7 subunit in the brain and the diseases associated with its pathogenic variants.
| Gene | Protein | Primary location | Distribution in the brain | Pathogenic variants | ||
|---|---|---|---|---|---|---|
| Regions | Reference | Associated diseases | Reference | |||
| KCNQ1 | Kv7.1 | Heart | CTX, HPF, MB, CB, BS | Casimiro et al., 2001; Goldman et al., 2009. AIBS, THPA | Long QT syndrome 1, JLNS, familial atrial fibrillation epilepsy, SUDEP, ASD, developmental disorder | ClinVar, LOVD, denovo-db |
| KCNQ2 | Kv7.2 | Nervous system | CTX, HPF, A, HY, TH, OA, MD, SN, P, MY, CB | Wang et al., 1998; Cooper et al., 2001; Devaux et al., 2004; Klinger et al., 2011; D’Este et al., 2016; Galvin et al., 2020. AIBS, THPA | BFNE, EE, ASD, intellectual disability, developmental disorder, sporadic infantile spasm syndrome | ClinVar, RIKEE, denovo-db |
| KCNQ3 | Kv7.3 | Nervous system | CTX, HPF, A, HY, TH, OA, MD, SN, P, MY, CB | Wang et al., 1998; Devaux et al., 2004; Klinger et al., 2011; Galvin et al., 2020. AIBS, THPA | BFNE, EE, ASD, intellectual disability, developmental disorder | ClinVar, RIKEE, denovo-db |
| KCNQ4 | Kv7.4 | Inner ear | BS, OA, MD, RN, NA, MY, VTA, P | Hansen et al., 2006; Su et al., 2019. AIBS, THPA | DFNA2, ASD | ClinVar, denovo-db |
| KCNQ5 | Kv7.5 | Nervous system | CTX, HPF, BS, CB | Lerche et al., 2000; Schroeder et al., 2000; Tzingounis et al., 2010; Fidzinski et al., 2015; Galvin et al., 2020. AIBS, THPA | EE, ASD, intellectual disability, schizophrenia | ClinVar, RIKEE, denovo-db |
Brain regions: CTX, cortex; OA, olfactory areas; HPF, hippocampal formation; A, amygdala; NA, nucleus accumbens; BS, brain stem; TH, thalamus; HY, hypothalamus; MB, midbrain; RN, raphe nuclei; SN, substantia nigra; VTA, ventral tegmental area; PAL, pallidum; HB, hindbrain; CB, cerebellum; P, pons; MY, medulla. Kv7 channelopathies: JLNS, Jervell and Lange-Nielsen syndrome; SUDEP, sudden unexpected death in epilepsy; BFNE, benign familial neonatal epilepsy; EE, epileptic encephalopathy; ASD, autism spectrum disorder; DFNA2, nonsyndromic sensorineural deafness type 2. Database website: Allen Institute for Brain Science (AIBS, https://alleninstitute.org/what-we-do/brain-science/), The Human Protein Atlas (THPA, https://www.proteinatlas.org/), ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/), Leiden Open Variation Database (LOVD, https://research.cchmc.org/LOVD2/home.php), denovo-db (http://denovo-db.gs.washington.edu/denovo-db/index.jsp), and Rational Intervention for KCNQ2/3 Epileptic Encephalopathy (RIKEE, https://www.rikee.org/).
Importantly, >300 dominant mutations in KCNQ2 and KCNQ3 cause epilepsy including benign familial neonatal epilepsy (BFNE) and epileptic encephalopathy (EE; Rikee and ClinVar database). KCNQ2 is the second most frequently mutated gene in neurodevelopmental disorder (Traynelis et al., 2017; Coe et al., 2019) characterized by cognitive and behavioral deficits (Mullin et al., 2013). A few mutations in KCNQ1, KCNQ4, and KCNQ5 have been associated with epilepsy, autism, schizophrenia, and developmental disorder (Table 1). Haploinsufficiency in Kv7 function seems to underlie BFNE variants that cause the transient appearance of neonatal seizures (Soldovieri et al., 2011). EE patients display severe and often drug-resistant neonatal seizures and psychomotor retardation (Weckhuysen et al., 2012), and de novo EE mutations in KCNQ2 and KCNQ3 induce multiple defects in current and surface expression of Kv7 channels (Weckhuysen et al., 2012, 2013; Milh et al., 2013; Miceli et al., 2015; Kim et al., 2018; Zhang et al., 2020).
General Properties and Regulation of Kv7 Currents
Kv7.1 assembles with auxiliary β subunit KCNE1 to produce the slow delayed rectifier K+ current (IKs) important for the repolarization of cardiac APs (Barhanin et al., 1996). Importantly, coassembly with KCNE1 slows the activation kinetics of Kv7.1 channel, potentiates its current amplitude, and eliminates its voltage-dependent inactivation (Barhanin et al., 1996; Sanguinetti et al., 1996; Tristani-Firouzi and Sanguinetti, 1998). Homomeric Kv7.2 channels activate at −60 mV and produce slow-activating and non-inactivating currents (Biervert et al., 1998). In comparison, currents through Kv7.3 channels are negligible due to an Ala residue in the pore domain (Wang et al., 1998; Gomez-Posada et al., 2010). Kv7.5 activates at −60 mV with slower kinetics than Kv7.2 and Kv7.3 (Schroeder et al., 2000; Gamper et al., 2003). Kv7.4 activates at −40 mV with slower activation kinetics than other Kv7 channels (Kubisch et al., 1999).
Neuronal Kv7 channels are mostly heterotetrameric channels composed of Kv7.2 and Kv7.3, and to a lesser extent Kv7.3 and Kv7.5 (Wang et al., 1998; Shah et al., 2002; Table 1). Compared to homomeric channels, significantly larger currents are generated by Kv7.2/Kv7.3 channels (Schroeder et al., 1998; Wang et al., 1998; Schwake et al., 2000) and Kv7.3/Kv7.5 channels (Schroeder et al., 2000; Gilling et al., 2013). Kv7.2/Kv7.3 channels produce M-current (I M; Wang et al., 1998), which potently suppresses neuronal hyperexcitability (Wang et al., 1998; Yue and Yaari, 2004). I M is inhibited by muscarinic acetylcholine receptor activation (Selyanko et al., 2000) and the depletion of PIP2 (Suh and Hille, 2002; Zhang et al., 2003). Kv7 channels are also inhibited by other G-protein coupled receptors, including substance P, bradykinin, serotonin, angiotensin, luteinizing hormone-releasing hormone, opioid, and metabotropic glutamate receptors (Marrion, 1997). General properties and diverse regulation of Kv7 channels are described in detail in a previous review (Soldovieri et al., 2011).
Role of Kv7 Channels in Intrinsic Excitability and Plasticity in the Hippocampus
Brown and Adams have first reported in 1980 that inhibition of I M upon stimulation of muscarinic acetylcholine receptor results in repetitive firing of APs in bullfrog sympathetic ganglion neurons (Brown and Adams, 1980). In the hippocampus, strong expression of Kv7.2, Kv7.3, and Kv7.5 is detected in pyramidal neurons (Schroeder et al., 2000; Cooper et al., 2001; Devaux et al., 2004). Kv7 antagonists XE991 and linopirdine depolarize resting membrane potential (RMP) and reduce AP threshold of hippocampal CA1 pyramidal neurons, resulting in spontaneous AP firing (Aiken et al., 1995; Shah et al., 2008; Figure 1A). Kv7 antagonists also increase intrinsic excitability (Yue and Yaari, 2004, 2006; Shah et al., 2008), contribute to medium and slow afterhyperpolarization (AHP) currents (Gu et al., 2005), reduce spike frequency adaptation (Aiken et al., 1995), and ultimately lead to an increased AP firing rate (Lezmy et al., 2020; Figure 1A). Consistent with pharmacologic inhibition, suppression of Kv7 current by overexpressing Kv7.2 containing dominant-negative pore mutation G279S enhances intrinsic excitability and reduces spike frequency adaptation and mAHP in CA1 neurons (Peters et al., 2005). Similarly, conditional homozygous deletion of KCNQ2 increases CA1 excitability due to longer-lasting spike afterdepolarization (ADP) and reduced medium AHP (Soh et al., 2014; Figure 1A). Thus, Kv7 channels serve as critical “brakes” on neuronal excitability (Soldovieri et al., 2011).
Figure 1.
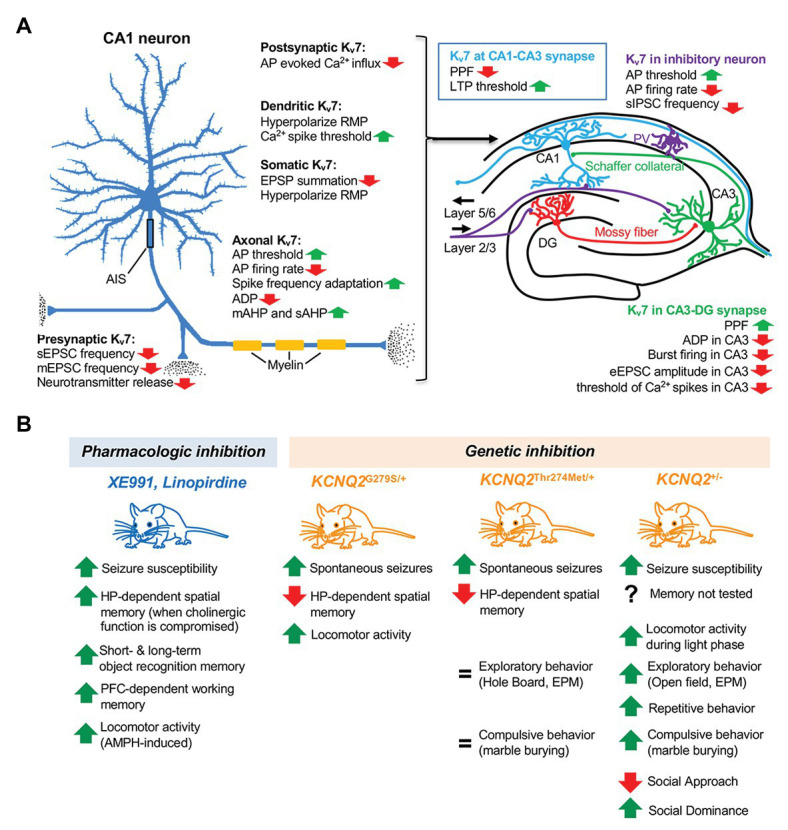
The role of Kv7 channels in hippocampal neurons, memory, and behavior. (A) Function of Kv7 channels in excitatory pyramidal neuron and GABAergic inhibitory neuron in the hippocampus. AP, action potential; RMP, resting membrane potential; EPSP, excitatory postsynaptic potential; ADP, afterdepolarization; mAHP, medium afterhyperpolarization; sAHP, slow afterhyperpolarization; sEPSC, spontaneous excitatory postsynaptic current; mEPSC, miniature excitatory postsynaptic current; PPF, paired-pulse facilitation; LTP, long-term potential; sIPSC, spontaneous inhibitory postsynaptic current; PV, parvalbumin; and eEPSC, evoked excitatory postsynaptic current. (B) Effects of pharmacological or genetic inhibition of Kv7 channels on memory and behavior. HP, hippocampus; PFC, pre-frontal cortex; AMPH, amphetamine; and EPM, elevated plus maze.
The inhibitory effects of Kv7 currents on neuronal excitability are largely attributed to axonal Kv7 channels. Kv7.2/Kv7.3 channels are preferentially enriched at the axonal plasma membrane compared to the somatodendritic plasma membrane in hippocampal neurons (Chung et al., 2006) with the highest concentration at the axonal initial segments (AIS; Chung et al., 2006; Pan et al., 2006) where AP initiates (Clark et al., 2009). CaM binding to Kv7.2 is critical for targeting Kv7.2/Kv7.3 channels to the axonal surface (Cavaretta et al., 2014), whereas disruption of this binding decreases I M and increases hippocampal neuronal excitability (Shahidullah et al., 2005). Furthermore, disrupting the enrichment of Kv7 channels at the AIS by blocking their interaction with ankyrin-G results in spontaneous firing of CA1 neurons by depolarizing RMP and reducing AP threshold (Shah et al., 2008).
In contrast to the well-documented function of axonal Kv7 channels discussed above, the existence and role of dendritic Kv7 channels are still in debate. Non-inactivating Kv7 current sensitive to muscarinic agonist is detected in the distal apical dendrites of CA1 neurons (Chen and Johnston, 2004). Dendritic Kv7 current can increase the threshold for initiating calcium (Ca2+) spikes and induce spike bursts only in hyperexcitable conditions that promote Ca2+ electrogenesis in these dendrites (Yue and Yaari, 2006). However, XE991 and linopirdine do not affect input resistance of CA1 dendrites (Shah et al., 2008), and focal inhibition of dendritic I M has no effect on the excitatory postsynaptic potential (EPSP) summation and excitability of CA1 neuron (Hu et al., 2007), indicating very low level of dendritic Kv7 current.
There is accumulating evidence for activity-dependent modulation of Kv7 channels and their contribution to persistent changes in intrinsic excitability termed “intrinsic plasticity.” In the pilocarpine model of temporal lobe epilepsy, reduced Kv7 function and expression may contribute to muscarinic-dependent ictogenesis (Maslarova et al., 2013). However, acute induction of seizures increases KCNQ2 and KCNQ3 transcripts in the hippocampi as a homeostatic response to suppress neuronal hyperexcitability, and this regulation requires activation of L-type voltage-gated Ca2+ channels (Zhang and Shapiro, 2012). Enhancing neuronal activity by Kv7 inhibition with XE991 also results in homeostatic suppression of firing rate over 48 h (Lezmy et al., 2020). In contrast, prolonged blockade of neuronal activity or N-methyl-D-aspartate (NMDA) receptors increases firing rate and reduces in KCNQ3 transcript and Kv7 current in hippocampal neurons (Lee and Chung, 2014; Lee et al., 2015). In the avian cochlear neurons, depriving afferent inputs induces a switch from fast activating Kv1 to slow activating Kv7.2 channels at the AIS, resulting in enhanced excitability (Kuba et al., 2015). This activity-dependent regulation of Kv7 transcript and distribution offers a powerful means to control intrinsic excitability.
Role of Kv7 Channels in Synaptic Transmission and Plasticity in the Hippocampus
Since the discovery of long-term potentiation (LTP) in the dentate gyrus of the hippocampus (Bliss and Lomo, 1973), persistent modification in synaptic strength termed “synaptic plasticity” has attracted significant attention as the cellular correlate of learning and memory (Nicoll, 2017). LTP at excitatory synapses can exert destabilizing influence on neural circuits by generating unconstrained synaptic strengthening (Turrigiano, 2012). Homeostatic plasticity counteracts such destabilizing condition by allowing neurons to adjust their synaptic strength (Turrigiano, 2012). While activity-dependent modulation of glutamate release and glutamate receptors serves as key mechanisms for LTP expression (Turrigiano, 2012; Humeau and Choquet, 2019), Kv7 channels and upstream muscarinic acetylcholine receptors have emerged as important regulators of excitatory synaptic transmission and plasticity.
Synaptic functions of Kv7 channels have been extensively studied at the excitatory synapses formed by hippocampal CA1 and CA3 pyramidal neurons (Figure 1A). These neurons show strong expression of Kv7.2 and Kv7.3 (Cooper et al., 2001; Pan et al., 2006). Conditional deletion of KCNQ2 and KCNQ3 increases the frequency of spontaneous excitatory postsynaptic currents (EPSC) in CA1 neurons (Soh et al., 2018), suggesting enhanced presynaptic release at CA1–CA3 synapses. Consistent with this notion, application of Kv7 antagonist XE991 increases whereas Kv7 agonist Flupirtine decreases miniature EPSC frequency in CA1 neurons (Sun and Kapur, 2012). Furthermore, Kv7 inhibition with linopirdine and XE991 treatment also increases neurotransmitter release (Nickolson et al., 1990; Martire et al., 2004; Peretz et al., 2007). While Kv7 current restrains AP-evoked Ca2+ influx into the presynaptic terminal and decreases the paired pulse ratio of evoked EPSCs at the mossy fiber–CA3 synapses (Martinello et al., 2019), paired pulse facilitation of EPSP is higher at CA1–CA3 synapses in XE991-treated mice (Fontan-Lozano et al., 2011), suggesting differential roles of Kv7 channels in short-term plasticity at two different synapses.
Kv7.2 and Kv7.3 are expressed in GABAergic neurons including parvalbumin (PV)- and somatostatin (SST)-positive interneurons in the hippocampus (Cooper et al., 2001; Lawrence et al., 2006). Application of XE991 abolishes I M, depolarizes RMP, and increases AP firing in SST+ interneurons (Lawrence et al., 2006) and enhances intrinsic excitability of PV+ interneurons (Soh et al., 2018; Figure 1A). Furthermore, conditional deletion of KCNQ2 and KCNQ3 from PV+ interneurons increases their firing and spontaneous inhibitory postsynaptic current (sIPSC) frequency of CA1 neurons in the hippocampus (Soh et al., 2018).
These studies highlight the presynaptic influence of Kv7 channels at glutamatergic and GABAergic synapses. Given that increased firing rate and burst firing can enhance neurotransmitter release probabilities (Hansen et al., 2008), Kv7 inhibition may increase neurotransmitter release as a consequence of increased axonal excitability (Devaux et al., 2004; Shah et al., 2008; Klinger et al., 2011). Indeed, when CA3 neurons are depolarized upon elevating extracellular K+ concentration, XE991 enhances EPSP amplitude in CA1 neurons as a consequence of increasing spike ADP and burst firing of CA3 neurons (Vervaeke et al., 2006). Alternatively, Kv7 channels at the presynaptic terminals (Cooper et al., 2001; Martire et al., 2004; Regev et al., 2009) may directly counteract the depolarization of the presynaptic membrane necessary for synaptic vesicle fusion and neurotransmitter release.
The postsynaptic role of Kv7 channels is unclear. A recent electron microscopy study shows that Kv7.2, Kv7.3, and Kv7.5 colocalize with muscarinic acetylcholine receptors at dendritic spines in layer III pyramidal neurons of the primate prefrontal cortex (Galvin et al., 2020), although the specificity of the immunolabeling needs to be further validated. In the CA1–CA3 synapses, the mEPSC amplitude is unaltered by agonist nor antagonists of Kv7 channels (Sun and Kapur, 2012), suggesting their negligible role in regulating postsynaptic glutamate receptor function at this synapse (Figure 1A).
Nonetheless, accumulating evidence suggests that Kv7 channels regulate hippocampal synaptic plasticity. At CA1–CA3 synapses, XE991 induces LTP by subthreshold theta-burst stimulation (Petrovic et al., 2012). Systemic administration of XE991 also decreases the threshold for LTP induction in the hippocampal CA1 area in vivo without affecting the field EPSP amplitude (Song et al., 2009; Fontan-Lozano et al., 2011). Lastly, homeostatic increase in excitatory synaptic transmission in CA1 neurons has been observed upon conditional deletion of KCNQ2 and KCNQ3 from GABAergic interneurons (Soh et al., 2018), suggesting the contribution of Kv7 channels in synaptic scaling.
Role of Kv7 Channels in Hippocampus-Dependent Learning and Memory
Hippocampal LTP occurs during hippocampus-dependent learning and memory (Bliss et al., 2018) and its reduction is linked to memory loss in mouse models of Alzheimer’s disease (Mango et al., 2019). Facilitation of LTP induction by XE991 (Song et al., 2009; Fontan-Lozano et al., 2011; Petrovic et al., 2012) suggests that pharmacologic Kv7 inhibition may enhance learning and memory. Indeed, linopirdine enhances the performance of rats in a hippocampus-dependent active avoidance test (Cook et al., 1990). XE991 improves memory in object recognition task in wild-type mice and mouse models of dementia induced by cholinergic depletion and neurodegeneration (Fontan-Lozano et al., 2011; Ballinger et al., 2016; Dennis et al., 2016) despite its ability to induce seizures at a higher dose (Fontan-Lozano et al., 2011; Figure 1B). In contrast, Kv7 agonists have yielded mixed results on affecting memory in rodents (Li et al., 2014; Frankel et al., 2016).
The cognition-enhancing effect of linopirdine is correlated with the increased release of acetylcholine in the hippocampus (Nickolson et al., 1990; Fontana et al., 1994), and stimulation of muscarinic acetylcholine receptor inhibits I M in hippocampal neurons (Shah et al., 2002). Consistently, muscarinic agonist improves whereas anticholinergic agent scopolamine impairs performance in hippocampus-dependent memory tasks (Fontana et al., 1994; Fontan-Lozano et al., 2011). Muscarinic acetylcholine receptors in the prefrontal cortex also modulate working memory in primates via Kv7 channels (Galvin et al., 2020). Since cholinergic depletion and dysfunction in the hippocampus and prefrontal cortex are implicated in age-related cognitive decline and Alzheimer’s disease (Ballinger et al., 2016; Haam and Yakel, 2017), these studies support the therapeutic potential for Kv7 antagonists as cognitive enhancers.
Surprisingly, genetic inhibition or reduction of Kv7 currents induces an opposite effect on memory (Figure 1B). Deficits in hippocampal-dependent spatial memory and spontaneous seizures are observed in mice with conditional transgenic expression of dominant-negative mutant Kv7.2-G279S (Peters et al., 2005) and heterozygous knock-in mice for Kv7.2 containing epileptic encephalopathy loss-of-function variant T274M (Milh et al., 2020). Considering that Kv7 channels are critical for development and inhibition of neonatal brain (Peters et al., 2005; Soh et al., 2014), the memory impairment in these genetic models could be attributed to abnormal hippocampal morphology and/or hyperexcitability (Peters et al., 2005; Milh et al., 2020).
Kv7 channels also regulate multiple behaviors (Figure 1B). Behavioral phenotyping of the global or conditional homozygous KCNQ2 knock-out mice has not been possible due to their early postnatal lethality or premature death, respectively (Watanabe et al., 2000; Soh et al., 2014). However, heterozygous KCNQ2 knock-out mice are viable and display increased locomotor activity and exploratory behavior (Kim et al., 2020), consistent with behavioral hyperactivity induced by transgenic suppression of Kv7 currents (Peters et al., 2005) and amphetamine and XE991 (Sotty et al., 2009). These mice also show decreased sociability and increased repetitive and compulsive behavior (Kim et al., 2020), reminiscent of autism seen in some EE patients with dominant KCNQ2 mutations (Weckhuysen et al., 2012, 2013; Milh et al., 2013). However, the precise circuitries responsible for these abnormal behaviors remain unknown.
Future Perspectives
The studies discussed in this review support the emerging concept that Kv7 channels contribute to neural plasticity, memory, and behavior. However, there is a significant knowledge gap in our understanding of the underlying molecular and cellular mechanisms. Future studies should continue to investigate structure-function and subcellular targeting of Kv7 channels, which will provide mechanistic insights for developing specific modulators of their function and trafficking. Generation of mouse models in which deletion of a Kv7 subunit from specific neurons and subcellular localization with temporal control will be critical to delineate cell- and circuit-specific function of Kv7 channels in neural plasticity, cognition, and behavior.
Author Contributions
BCB and HC contributed to the conception and design of the manuscript. BCB, JZ, and HC also drafted and revised the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
Funding. This work was supported by the National Institute of Neurological Disorders and Stroke (NINDS) Research Project Grant NS083402 (HC). Understanding the Brain: Training the Next Generation of Researchers in Engineering and Deciphering of Miniature Brain Machinery. NSF training grant 1735252 (BCB).
References
- Aiken S. P., Lampe B. J., Murphy P. A., Brown B. S. (1995). Reduction of spike frequency adaptation and blockade of M-current in rat CA1 pyramidal neurones by linopirdine (DuP 996), a neurotransmitter release enhancer. Br. J. Pharmacol. 115, 1163–1168. 10.1111/j.1476-5381.1995.tb15019.x, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballinger E. C., Ananth M., Talmage D. A., Role L. W. (2016). Basal forebrain cholinergic circuits and signaling in cognition and cognitive decline. Neuron 91, 1199–1218. 10.1016/j.neuron.2016.09.006, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barhanin J., Lesage F., Guillemare E., Fink M., Lazdunski M., Romey G. (1996). K(V)LQT1 and lsK (minK) proteins associate to form the I(Ks) cardiac potassium current. Nature 384, 78–80. 10.1038/384078a0, PMID: [DOI] [PubMed] [Google Scholar]
- Biervert C., Schroeder B. C., Kubisch C., Berkovic S. F., Propping P., Jentsch T. J., et al. (1998). A potassium channel mutation in neonatal human epilepsy. Science 279, 403–406. 10.1126/science.279.5349.403, PMID: [DOI] [PubMed] [Google Scholar]
- Bliss T., Collingridge G., Morris R., Reymann K. (2018). Long-term potentiation in the hippocampus: discovery, mechanisms and function. Neuroforum 24, 103–120. 10.1515/nf-2017-A059 [DOI] [Google Scholar]
- Bliss T. V., Lomo T. (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 232, 331–356. 10.1113/jphysiol.1973.sp010273, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown D. A., Adams P. R. (1980). Muscarinic suppression of a novel voltage-sensitive K+ current in a vertebrate neurone. Nature 283, 673–676. 10.1038/283673a0, PMID: [DOI] [PubMed] [Google Scholar]
- Brown D. A., Passmore G. M. (2009). Neural KCNQ (Kv7) channels. Br. J. Pharmacol. 156, 1185–1195. 10.1111/j.1476-5381.2009.00111.x, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casimiro M. C., Knollmann B. C., Ebert S. N., Vary J. C., Jr., Greene A. E., Franz M. R., et al. (2001). Targeted disruption of the Kcnq1 gene produces a mouse model of Jervell and Lange-Nielsen syndrome. Proc. Natl. Acad. Sci. U. S. A. 98, 2526–2531. 10.1073/pnas.041398998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavaretta J. P., Sherer K. R., Lee K. Y., Kim E. H., Issema R. S., Chung H. J. (2014). Polarized axonal surface expression of neuronal KCNQ potassium channels is regulated by calmodulin interaction with KCNQ2 subunit. PLoS One 9:e103655. 10.1371/journal.pone.0103655, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Johnston D. (2004). Properties of single voltage-dependent K+ channels in dendrites of CA1 pyramidal neurones of rat hippocampus. J. Physiol. 559, 187–203. 10.1113/jphysiol.2004.068114, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung H. J., Jan Y. N., Jan L. Y. (2006). Polarized axonal surface expression of neuronal KCNQ channels is mediated by multiple signals in the KCNQ2 and KCNQ3 C-terminal domains. Proc. Natl. Acad. Sci. U. S. A. 103, 8870–8875. 10.1073/pnas.0603376103, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark B. D., Goldberg E. M., Rudy B. (2009). Electrogenic tuning of the axon initial segment. Neuroscientist 15, 651–668. 10.1177/1073858409341973, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coe B. P., Stessman H. A. F., Sulovari A., Geisheker M. R., Bakken T. E., Lake A. M., et al. (2019). Neurodevelopmental disease genes implicated by de novo mutation and copy number variation morbidity. Nat. Genet. 51, 106–116. 10.1038/s41588-018-0288-4, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook L., Nickolson V., Steinfels G., Rohrbach K., Denoble V. (1990). Cognition enhancement by the acetylcholine releaser DuP 996. Drug Dev. Res. 19, 301–314. 10.1002/ddr.430190308 [DOI] [Google Scholar]
- Cooper E. C., Harrington E., Jan Y. N., Jan L. Y. (2001). M channel KCNQ2 subunits are localized to key sites for control of neuronal network oscillations and synchronization in mouse brain. J. Neurosci. 21, 9529–9540. 10.1523/JNEUROSCI.21-24-09529.2001, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui J. (2016). Voltage-dependent gating: novel insights from KCNQ1 channels. Biophys. J. 110, 14–25. 10.1016/j.bpj.2015.11.023, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennis S. H., Pasqui F., Colvin E. M., Sanger H., Mogg A. J., Felder C. C., et al. (2016). Activation of muscarinic M1 acetylcholine receptors induces long-term potentiation in the hippocampus. Cereb. Cortex 26, 414–426. 10.1093/cercor/bhv227, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Este E., Kamin D., Balzarotti F., Hell S. (2016). Ultrastructural anatomy of nodes of Ranvier in the peripheral nervous system as revealed by STED microscopy. Proc. Natl. Acad. Sci. U. S. A. 114, E191–E199. 10.1073/pnas.1619553114, [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devaux J. J., Kleopa K. A., Cooper E. C., Scherer S. S. (2004). KCNQ2 is a nodal K+ channel. J. Neurosci. 24, 1236–1244. 10.1523/JNEUROSCI.4512-03.2004, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fidzinski P., Korotkova T., Heidenreich M., Maier N., Schuetze S., Kobler O., et al. (2015). KCNQ5 K(+) channels control hippocampal synaptic inhibition and fast network oscillations. Nat. Commun. 6:6254. 10.1038/ncomms7254, PMID: [DOI] [PubMed] [Google Scholar]
- Fontana D. J., Inouye G. T., Johnson R. M. (1994). Linopirdine (DuP 996) improves performance in several tests of learning and memory by modulation of cholinergic neurotransmission. Pharmacol. Biochem. Behav. 49, 1075–1082. 10.1016/0091-3057(94)90267-4, PMID: [DOI] [PubMed] [Google Scholar]
- Fontan-Lozano A., Suarez-Pereira I., Delgado-Garcia J. M., Carrion A. M. (2011). The M-current inhibitor XE991 decreases the stimulation threshold for long-term synaptic plasticity in healthy mice and in models of cognitive disease. Hippocampus 21, 22–32. 10.1002/hipo.20717, PMID: [DOI] [PubMed] [Google Scholar]
- Frankel S., Medvedeva N., Gutherz S., Kulick C., Kondratyev A., Forcelli P. A. (2016). Comparison of the long-term behavioral effects of neonatal exposure to retigabine or phenobarbital in rats. Epilepsy Behav. 57, 34–40. 10.1016/j.yebeh.2016.01.018, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galvin V. C., Yang S. T., Paspalas C. D., Yang Y., Jin L. E., Datta D., et al. (2020). Muscarinic M1 receptors modulate working memory performance and activity via KCNQ potassium channels in the primate prefrontal cortex. Neuron 106, 649.e4–661.e4. 10.1016/j.neuron.2020.02.030, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamper N., Stockand J. D., Shapiro M. S. (2003). Subunit-specific modulation of KCNQ potassium channels by Src tyrosine kinase. J. Neurosci. 23, 84–95. 10.1523/JNEUROSCI.23-01-00084.2003, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilling M., Rasmussen H. B., Calloe K., Sequeira A. F., Baretto M., Oliveira G., et al. (2013). Dysfunction of the heteromeric KV7.3/KV7.5 potassium channel is associated with autism spectrum disorders. Front. Genet. 4:54. 10.3389/fgene.2013.00054, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman A. M., Glasscock E., Yoo J., Chen T. T., Klassen T. L., Noebels J. L. (2009). Arrhythmia in heart and brain: KCNQ1 mutations link epilepsy and sudden unexplained death. Sci. Transl. Med. 1:2ra6. 10.1126/scitranslmed.3000289, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez-Posada J. C., Etxeberria A., Roura-Ferrer M., Areso P., Masin M., Murrell-Lagnado R. D., et al. (2010). A pore residue of the KCNQ3 potassium M-channel subunit controls surface expression. J. Neurosci. 30, 9316–9323. 10.1523/JNEUROSCI.0851-10.2010, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu N., Vervaeke K., Hu H., Storm J. F. (2005). Kv7/KCNQ/M and HCN/h, but not KCa2/SK channels, contribute to the somatic medium after-hyperpolarization and excitability control in CA1 hippocampal pyramidal cells. J. Physiol. 566, 689–715. 10.1113/jphysiol.2005.086835, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutman G. A., Chandy K. G., Grissmer S., Lazdunski M., Mckinnon D., Pardo L. A., et al. (2005). International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol. Rev. 57, 473–508. 10.1124/pr.57.4.10, PMID: [DOI] [PubMed] [Google Scholar]
- Haam J., Yakel J. L. (2017). Cholinergic modulation of the hippocampal region and memory function. J. Neurochem. 142, 111–121. 10.1111/jnc.14052, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haitin Y., Attali B. (2008). The C-terminus of Kv7 channels: a multifunctional module. J. Physiol. 586, 1803–1810. 10.1113/jphysiol.2007.149187, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen H. H., Ebbesen C., Mathiesen C., Weikop P., Ronn L. C., Waroux O., et al. (2006). The KCNQ channel opener retigabine inhibits the activity of mesencephalic dopaminergic systems of the rat. J. Pharmacol. Exp. Ther. 318, 1006–1019. 10.1124/jpet.106.106757, PMID: [DOI] [PubMed] [Google Scholar]
- Hansen H. H., Waroux O., Seutin V., Jentsch T. J., Aznar S., Mikkelsen J. D. (2008). Kv7 channels: interaction with dopaminergic and serotonergic neurotransmission in the CNS. J. Physiol. 586, 1823–1832. 10.1113/jphysiol.2007.149450, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu H., Vervaeke K., Storm J. F. (2007). M-channels (Kv7/KCNQ channels) that regulate synaptic integration, excitability, and spike pattern of CA1 pyramidal cells are located in the perisomatic region. J. Neurosci. 27, 1853–1867. 10.1523/JNEUROSCI.4463-06.2007, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humeau Y., Choquet D. (2019). The next generation of approaches to investigate the link between synaptic plasticity and learning. Nat. Neurosci. 22, 1536–1543. 10.1038/s41593-019-0480-6, PMID: [DOI] [PubMed] [Google Scholar]
- Kim E. C., Patel J., Zhang J., Soh H., Rhodes J. S., Tzingounis A. V., et al. (2020). Heterozygous loss of epilepsy gene KCNQ2 alters social, repetitive and exploratory behaviors. Genes Brain Behav. 19:e12599. 10.1111/gbb.12599, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim E. C., Zhang J., Pang W., Wang S., Lee K. Y., Cavaretta J. P., et al. (2018). Reduced axonal surface expression and phosphoinositide sensitivity in Kv7 channels disrupts their function to inhibit neuronal excitability in Kcnq2 epileptic encephalopathy. Neurobiol. Dis. 118, 76–93. 10.1016/j.nbd.2018.07.004, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klinger F., Gould G., Boehm S., Shapiro M. S. (2011). Distribution of M-channel subunits KCNQ2 and KCNQ3 in rat hippocampus. Neuroimage 58, 761–769. 10.1016/j.neuroimage.2011.07.003, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuba H., Yamada R., Ishiguro G., Adachi R. (2015). Redistribution of Kv1 and Kv7 enhances neuronal excitability during structural axon initial segment plasticity. Nat. Commun. 6:8815. 10.1038/ncomms9815, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubisch C., Schroeder B. C., Friedrich T., Lutjohann B., El-Amraoui A., Marlin S., et al. (1999). KCNQ4, a novel potassium channel expressed in sensory outer hair cells, is mutated in dominant deafness. Cell 96, 437–446. 10.1016/S0092-8674(00)80556-5, PMID: [DOI] [PubMed] [Google Scholar]
- Lawrence J. J., Saraga F., Churchill J. F., Statland J. M., Travis K. E., Skinner F. K., et al. (2006). Somatodendritic Kv7/KCNQ/M channels control interspike interval in hippocampal interneurons. J. Neurosci. 26, 12325–12338. 10.1523/JNEUROSCI.3521-06.2006, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee K. Y., Chung H. J. (2014). NMDA receptors and L-type voltage-gated Ca2+ channels mediate the expression of bidirectional homeostatic intrinsic plasticity in cultured hippocampal neurons. Neuroscience 277, 610–623. 10.1016/j.neuroscience.2014.07.038, PMID: [DOI] [PubMed] [Google Scholar]
- Lee K. Y., Royston S. E., Vest M. O., Ley D. J., Lee S., Bolton E. C., et al. (2015). N-methyl-D-aspartate receptors mediate activity-dependent down-regulation of potassium channel genes during the expression of homeostatic intrinsic plasticity. Mol. Brain 8:4. 10.1186/s13041-015-0094-1, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerche C., Scherer C. R., Seebohm G., Derst C., Wei A. D., Busch A. E., et al. (2000). Molecular cloning and functional expression of KCNQ5, a potassium channel subunit that may contribute to neuronal M-current diversity. J. Biol. Chem. 275, 22395–22400. 10.1074/jbc.M002378200, PMID: [DOI] [PubMed] [Google Scholar]
- Lezmy J., Gelman H., Katsenelson M., Styr B., Tikochinsky E., Lipinsky M., et al. (2020). M-current inhibition in hippocampal excitatory neurons triggers intrinsic and synaptic homeostatic responses at different temporal scales. J. Neurosci. 40, 3694–3706. 10.1523/JNEUROSCI.1914-19.2020, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C., Huang P., Lu Q., Zhou M., Guo L., Xu X. (2014). KCNQ/Kv7 channel activator flupirtine protects against acute stress-induced impairments of spatial memory retrieval and hippocampal LTP in rats. Neuroscience 280, 19–30. 10.1016/j.neuroscience.2014.09.009, PMID: [DOI] [PubMed] [Google Scholar]
- Mango D., Saidi A., Cisale G. Y., Feligioni M., Corbo M., Nistico R. (2019). Targeting synaptic plasticity in experimental models of Alzheimer’s disease. Front. Pharmacol. 10:778. 10.3389/fphar.2019.00778, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marrion N. V. (1997). Control of M-current. Annu. Rev. Physiol. 59, 483–504. 10.1146/annurev.physiol.59.1.483, PMID: [DOI] [PubMed] [Google Scholar]
- Martinello K., Giacalone E., Migliore M., Brown D. A., Shah M. M. (2019). The subthreshold-active KV7 current regulates neurotransmission by limiting spike-induced Ca2+ influx in hippocampal mossy fiber synaptic terminals. Commun. Biol. 2:145. 10.1038/s42003-019-0408-4, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martire M., Castaldo P., D’amico M., Preziosi P., Annunziato L., Taglialatela M. (2004). M channels containing KCNQ2 subunits modulate norepinephrine, aspartate, and GABA release from hippocampal nerve terminals. J. Neurosci. 24, 592–597. 10.1523/JNEUROSCI.3143-03.2004, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maslarova A., Salar S., Lapilover E., Friedman A., Veh R. W., Heinemann U. (2013). Increased susceptibility to acetylcholine in the entorhinal cortex of pilocarpine-treated rats involves alterations in KCNQ channels. Neurobiol. Dis. 56, 14–24. 10.1016/j.nbd.2013.02.016, PMID: [DOI] [PubMed] [Google Scholar]
- Miceli F., Soldovieri M. V., Martire M., Taglialatela M. (2008). Molecular pharmacology and therapeutic potential of neuronal Kv7-modulating drugs. Curr. Opin. Pharmacol. 8, 65–74. 10.1016/j.coph.2007.10.003, PMID: [DOI] [PubMed] [Google Scholar]
- Miceli F., Striano P., Soldovieri M. V., Fontana A., Nardello R., Robbiano A., et al. (2015). A novel KCNQ3 mutation in familial epilepsy with focal seizures and intellectual disability. Epilepsia 56, e15–e20. 10.1111/epi.12887, PMID: [DOI] [PubMed] [Google Scholar]
- Milh M., Boutry-Kryza N., Sutera-Sardo J., Mignot C., Auvin S., Lacoste C., et al. (2013). Similar early characteristics but variable neurological outcome of patients with a de novo mutation of KCNQ2. Orphanet J. Rare Dis. 8:80. 10.1186/1750-1172-8-80, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milh M., Roubertoux P., Biba N., Chavany J., Spiga Ghata A., Fulachier C., et al. (2020). A knock-in mouse model for KCNQ2-related epileptic encephalopathy displays spontaneous generalized seizures and cognitive impairment. Epilepsia 61, 868–878. 10.1111/epi.16494, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mullin A. P., Gokhale A., Moreno-De-Luca A., Sanyal S., Waddington J. L., Faundez V. (2013). Neurodevelopmental disorders: mechanisms and boundary definitions from genomes, interactomes and proteomes. Transl. Psychiatry 3:e329. 10.1038/tp.2013.108, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nappi P., Miceli F., Soldovieri M. V., Ambrosino P., Barrese V., Taglialatela M. (2020). Epileptic channelopathies caused by neuronal Kv7 (KCNQ) channel dysfunction. Pflugers Arch. 472, 881–898. 10.1007/s00424-020-02404-2, PMID: [DOI] [PubMed] [Google Scholar]
- Nickolson V., William Tam S., Myers M., Cook L. (1990). DuP 996 (3,3-bis(4-pyrindinylmethyl)-1-phenylindolin-2-one) enhances the stimulus-induced release of acetylcholine from rat brain in vitro and in vivo. Drug Dev. Res. 19, 285–300. 10.1002/ddr.430190307 [DOI] [Google Scholar]
- Nicoll R. A. (2017). A brief history of long-term potentiation. Neuron 93, 281–290. 10.1016/j.neuron.2016.12.015, PMID: [DOI] [PubMed] [Google Scholar]
- Pan Z., Kao T., Horvath Z., Lemos J., Sul J. Y., Cranstoun S. D., et al. (2006). A common ankyrin-G-based mechanism retains KCNQ and NaV channels at electrically active domains of the axon. J. Neurosci. 26, 2599–2613. 10.1523/JNEUROSCI.4314-05.2006, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peretz A., Sheinin A., Yue C., Degani-Katzav N., Gibor G., Nachman R., et al. (2007). Pre- and postsynaptic activation of M-channels by a novel opener dampens neuronal firing and transmitter release. J. Neurophysiol. 97, 283–295. 10.1152/jn.00634.2006, PMID: [DOI] [PubMed] [Google Scholar]
- Peters H. C., Hu H., Pongs O., Storm J. F., Isbrandt D. (2005). Conditional transgenic suppression of M channels in mouse brain reveals functions in neuronal excitability, resonance and behavior. Nat. Neurosci. 8, 51–60. 10.1038/nn1375, PMID: [DOI] [PubMed] [Google Scholar]
- Petrovic M. M., Nowacki J., Olivo V., Tsaneva-Atanasova K., Randall A. D., Mellor J. R. (2012). Inhibition of post-synaptic Kv7/KCNQ/M channels facilitates long-term potentiation in the hippocampus. PLoS One 7:e30402. 10.1371/journal.pone.0030402, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Regev N., Degani-Katzav N., Korngreen A., Etzioni A., Siloni S., Alaimo A., et al. (2009). Selective interaction of syntaxin 1A with KCNQ2: possible implications for specific modulation of presynaptic activity. PLoS One 4:e6586. 10.1371/journal.pone.0006586, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robbins J. (2001). KCNQ potassium channels: physiology, pathophysiology, and pharmacology. Pharmacol. Ther. 90, 1–19. 10.1016/S0163-7258(01)00116-4, PMID: [DOI] [PubMed] [Google Scholar]
- Sanguinetti M. C., Curran M. E., Zou A., Shen J., Spector P. S., Atkinson D. L., et al. (1996). Coassembly of K(V)LQT1 and minK (IsK) proteins to form cardiac I(Ks) potassium channel. Nature 384, 80–83. 10.1038/384080a0, PMID: [DOI] [PubMed] [Google Scholar]
- Schroeder B. C., Hechenberger M., Weinreich F., Kubisch C., Jentsch T. J. (2000). KCNQ5, a novel potassium channel broadly expressed in brain, mediates M-type currents. J. Biol. Chem. 275, 24089–24095. 10.1074/jbc.M003245200, PMID: [DOI] [PubMed] [Google Scholar]
- Schroeder B. C., Kubisch C., Stein V., Jentsch T. J. (1998). Moderate loss of function of cyclic-AMP-modulated KCNQ2/KCNQ3 K+ channels causes epilepsy. Nature 396, 687–690. 10.1038/25367, PMID: [DOI] [PubMed] [Google Scholar]
- Schwake M., Pusch M., Kharkovets T., Jentsch T. J. (2000). Surface expression and single channel properties of KCNQ2/KCNQ3, M-type K+ channels involved in epilepsy. J. Biol. Chem. 275, 13343–13348. 10.1074/jbc.275.18.13343, PMID: [DOI] [PubMed] [Google Scholar]
- Selyanko A. A., Hadley J. K., Wood I. C., Abogadie F. C., Jentsch T. J., Brown D. A. (2000). Inhibition of KCNQ1-4 potassium channels expressed in mammalian cells via M1 muscarinic acetylcholine receptors. J. Physiol. 522, 349–355. 10.1111/j.1469-7793.2000.t01-2-00349.x, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shah M. M., Migliore M., Valencia I., Cooper E. C., Brown D. A. (2008). Functional significance of axonal Kv7 channels in hippocampal pyramidal neurons. Proc. Natl. Acad. Sci. U. S. A. 105, 7869–7874. 10.1073/pnas.0802805105, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shah M., Mistry M., Marsh S. J., Brown D. A., Delmas P. (2002). Molecular correlates of the M-current in cultured rat hippocampal neurons. J. Physiol. 544, 29–37. 10.1113/jphysiol.2002.028571, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahidullah M., Santarelli L. C., Wen H., Levitan I. B. (2005). Expression of a calmodulin-binding KCNQ2 potassium channel fragment modulates neuronal M-current and membrane excitability. Proc. Natl. Acad. Sci. U. S. A. 102, 16454–16459. 10.1073/pnas.0503966102, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soh H., Pant R., Loturco J. J., Tzingounis A. V. (2014). Conditional deletions of epilepsy-associated KCNQ2 and KCNQ3 channels from cerebral cortex cause differential effects on neuronal excitability. J. Neurosci. 34, 5311–5321. 10.1523/JNEUROSCI.3919-13.2014, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soh H., Park S., Ryan K., Springer K., Maheshwari A., Tzingounis A. V. (2018). Deletion of KCNQ2/3 potassium channels from PV+ interneurons leads to homeostatic potentiation of excitatory transmission. Elife 7:e38617. 10.7554/eLife.38617, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soldovieri M. V., Miceli F., Taglialatela M. (2011). Driving with no brakes: molecular pathophysiology of Kv7 potassium channels. Physiology 26, 365–376. 10.1152/physiol.00009.2011, PMID: [DOI] [PubMed] [Google Scholar]
- Song M. K., Cui Y. Y., Zhang W. W., Zhu L., Lu Y., Chen H. Z. (2009). The facilitating effect of systemic administration of Kv7/M channel blocker XE991 on LTP induction in the hippocampal CA1 area independent of muscarinic activation. Neurosci. Lett. 461, 25–29. 10.1016/j.neulet.2009.05.042, PMID: [DOI] [PubMed] [Google Scholar]
- Sotty F., Damgaard T., Montezinho L. P., Mork A., Olsen C. K., Bundgaard C., et al. (2009). Antipsychotic-like effect of retigabine [N-(2-amino-4-(fluorobenzylamino)-phenyl)carbamic acid ester], a KCNQ potassium channel opener, via modulation of mesolimbic dopaminergic neurotransmission. J. Pharmacol. Exp. Ther. 328, 951–962. 10.1124/jpet.108.146944, PMID: [DOI] [PubMed] [Google Scholar]
- Strulovich R., Tobelaim W. S., Attali B., Hirsch J. A. (2016). Structural insights into the M-channel proximal C-terminus/calmodulin complex. Biochemistry 55, 5353–5365. 10.1021/acs.biochem.6b00477, PMID: [DOI] [PubMed] [Google Scholar]
- Su M., Li L., Wang J., Sun H., Zhang L., Zhao C., et al. (2019). Kv7.4 channel contribute to projection-specific auto-inhibition of dopamine neurons in the ventral tegmental area. Front. Cell. Neurosci. 13:557. 10.3389/fncel.2019.00557, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suh B. C., Hille B. (2002). Recovery from muscarinic modulation of M current channels requires phosphatidylinositol 4,5-bisphosphate synthesis. Neuron 35, 507–520. 10.1016/S0896-6273(02)00790-0, PMID: [DOI] [PubMed] [Google Scholar]
- Suh B. C., Hille B. (2008). PIP2 is a necessary cofactor for ion channel function: how and why? Annu. Rev. Biophys. 37, 175–195. 10.1146/annurev.biophys.37.032807.125859, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun J., Kapur J. (2012). M-type potassium channels modulate Schaffer collateral-CA1 glutamatergic synaptic transmission. J. Physiol. 590, 3953–3964. 10.1113/jphysiol.2012.235820, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun J., Mackinnon R. (2017). Cryo-EM structure of a KCNQ1/CaM complex reveals insights into congenital long QT syndrome. Cell 169, 1042.e1049–1050.e1049. 10.1016/j.cell.2017.05.019, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun J., Mackinnon R. (2020). Structural basis of human KCNQ1 modulation and gating. Cell 180, 340.e349–347.e349. 10.1016/j.cell.2019.12.003, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traynelis J., Silk M., Wang Q., Berkovic S. F., Liu L., Ascher D. B., et al. (2017). Optimizing genomic medicine in epilepsy through a gene-customized approach to missense variant interpretation. Genome Res. 27, 1715–1729. 10.1101/gr.226589.117, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tristani-Firouzi M., Sanguinetti M. C. (1998). Voltage-dependent inactivation of the human K+ channel KvLQT1 is eliminated by association with minimal K+ channel (minK) subunits. J. Physiol. 510, 37–45. 10.1111/j.1469-7793.1998.037bz.x, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turrigiano G. (2012). Homeostatic synaptic plasticity: local and global mechanisms for stabilizing neuronal function. Cold Spring Harb. Perspect. Biol. 4:a005736. 10.1101/cshperspect.a005736, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzingounis A. V., Heidenreich M., Kharkovets T., Spitzmaul G., Jensen H. S., Nicoll R. A., et al. (2010). The KCNQ5 potassium channel mediates a component of the afterhyperpolarization current in mouse hippocampus. Proc. Natl. Acad. Sci. U. S. A. 107, 10232–10237. 10.1073/pnas.1004644107, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vervaeke K., Gu N., Agdestein C., Hu H., Storm J. F. (2006). Kv7/KCNQ/M-channels in rat glutamatergic hippocampal axons and their role in regulation of excitability and transmitter release. J. Physiol. 576, 235–256. 10.1113/jphysiol.2006.111336, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q., Curran M. E., Splawski I., Burn T. C., Millholland J. M., Vanraay T. J., et al. (1996). Positional cloning of a novel potassium channel gene: KVLQT1 mutations cause cardiac arrhythmias. Nat. Genet. 12, 17–23. 10.1038/ng0196-17, PMID: [DOI] [PubMed] [Google Scholar]
- Wang H. S., Pan Z., Shi W., Brown B. S., Wymore R. S., Cohen I. S., et al. (1998). KCNQ2 and KCNQ3 potassium channel subunits: molecular correlates of the M-channel. Science 282, 1890–1893. 10.1126/science.282.5395.1890, PMID: [DOI] [PubMed] [Google Scholar]
- Watanabe H., Nagata E., Kosakai A., Nakamura M., Yokoyama M., Tanaka K., et al. (2000). Disruption of the epilepsy KCNQ2 gene results in neural hyperexcitability. J. Neurochem. 75, 28–33. 10.1046/j.1471-4159.2000.0750028.x, PMID: [DOI] [PubMed] [Google Scholar]
- Weckhuysen S., Ivanovic V., Hendrickx R., Van Coster R., Hjalgrim H., Moller R. S., et al. (2013). Extending the KCNQ2 encephalopathy spectrum: clinical and neuroimaging findings in 17 patients. Neurology 81, 1697–1703. 10.1212/01.wnl.0000435296.72400.a1, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weckhuysen S., Mandelstam S., Suls A., Audenaert D., Deconinck T., Claes L. R., et al. (2012). KCNQ2 encephalopathy: emerging phenotype of a neonatal epileptic encephalopathy. Ann. Neurol. 71, 15–25. 10.1002/ana.22644, PMID: [DOI] [PubMed] [Google Scholar]
- Yue C., Yaari Y. (2004). KCNQ/M channels control spike afterdepolarization and burst generation in hippocampal neurons. J. Neurosci. 24, 4614–4624. 10.1523/JNEUROSCI.0765-04.2004, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yue C., Yaari Y. (2006). Axo-somatic and apical dendritic Kv7/M channels differentially regulate the intrinsic excitability of adult rat CA1 pyramidal cells. J. Neurophysiol. 95, 3480–3495. 10.1152/jn.01333.2005, PMID: [DOI] [PubMed] [Google Scholar]
- Zaydman M. A., Cui J. (2014). PIP2 regulation of KCNQ channels: biophysical and molecular mechanisms for lipid modulation of voltage-dependent gating. Front. Physiol. 5:195. 10.3389/fphys.2014.00195, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaydman M. A., Silva J. R., Delaloye K., Li Y., Liang H., Larsson H. P., et al. (2013). Kv7.1 ion channels require a lipid to couple voltage sensing to pore opening. Proc. Natl. Acad. Sci. U. S. A. 110, 13180–13185. 10.1073/pnas.1305167110, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H., Craciun L. C., Mirshahi T., Rohacs T., Lopes C. M., Jin T., et al. (2003). PIP(2) activates KCNQ channels, and its hydrolysis underlies receptor-mediated inhibition of M currents. Neuron 37, 963–975. 10.1016/S0896-6273(03)00125-9, PMID: [DOI] [PubMed] [Google Scholar]
- Zhang J., Kim E. C., Chen C., Procko E., Pant S., Lam K., et al. (2020). Identifying mutation hotspots reveals pathogenetic mechanisms of KCNQ2 epileptic encephalopathy. Sci. Rep. 10:4756. 10.1038/s41598-020-61697-6, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J., Shapiro M. S. (2012). Activity-dependent transcriptional regulation of M-Type (Kv7) K(+) channels by AKAP79/150-mediated NFAT actions. Neuron 76, 1133–1146. 10.1016/j.neuron.2012.10.019, PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]