

Lactobacillus salivarius Li01 improved TAA‐induced acute liver injury and hyperammonemia which was mediated by restoring destroyed gut microbiota and barrier function, improving TAA‐induced liver damage and systemic inflammation, and alleviating hyperammonaemia, neuro‐inflammation and damaged cognitive function.
Summary
The gut microbiota plays pivotal roles in liver disease onset and progression. The protective effects of Lactobacillus salivarius Li01 on liver diseases have been reported. In this study, we aimed to detect the protective effect of L. salivarius Li01 on thioacetamide (TAA)‐induced acute liver injury and hyperammonaemia. C57BL/6 mice were separated into three groups and given a gavage of L. salivarius Li01 or phosphate‐buffered saline for 7 days. Acute liver injury and hyperammonaemia were induced with an intraperitoneal TAA injection. L. salivarius Li01 decreased mortality and serum transaminase levels and improved histological liver damage caused by TAA. Serum inflammatory cytokine and chemokine and lipopolysaccharide‐binding protein (LBP) concentrations, nuclear factor κB (NFκB) pathway activation and macrophage and neutrophil infiltration into the liver were significantly alleviated by L. salivarius Li01. L. salivarius Li01 also reinforced gut barrier and reshaped the perturbed gut microbiota by upregulating Bacteroidetes and Akkermansia richness and downregulating Proteobacteria, Ruminococcaceae_UCG_014 and Helicobacter richness. Plasma and faecal ammonia levels declined noticeably in the Li01 group, accompanied by improvements in cognitive function, neuro‐inflammation and relative brain‐derived neurotrophic factor (BDNF) gene expression. Our results indicated that L. salivarius Li01 could be considered a potential probiotic in acute liver injury and hepatic encephalopathy (HE).
Introduction
Acute liver failure (ALF) is characterized by explosive hepatocyte injury that can be caused by numerous insults, such as drug toxicity, hepatitis viral infection and autoimmune hepatitis (Bernal et al., 2015). ALF and its complications frequently produce fatal outcomes in previously healthy people. Although many advanced therapeutic strategies for the treatment of patients with ALF have been proposed, liver transplantation is always the most effective method but is used in only approximately 30% of ALF patients (Stravitz and Lee, 2019) and is limited by a shortage in liver donors (O'Grady, 2012). ALF still presents a high mortality rate (Wlodzimirow et al., 2012) and seriously threatens human health.
Hepatic encephalopathy (HE) is a major complication of ALF and an important predictor of prognosis. After recovery from ALF, neuropsychological dysfunction might persist (Stravitz and Lee, 2019), which can affect the quality of life. The precise pathogenesis of HE is still unclear. Hyperammonaemia has generally been considered the main driver of HE (Gorg et al., 2013). Furthermore, the concentration of plasma ammonia is associated with the severity of HE (Patidar and Bajaj, 2015) and outcome of ALF (Yanny et al., 2019).
Accumulating evidence has demonstrated that an intimate relationship exists between the intestinal microbiota and liver disease (Wiest et al., 2017). Normally, the intestinal microbiota can protect the host against potential pathogens and their components, such as lipopolysaccharide (LPS), through a mechanism termed colonization resistance (Litvak and Baumler, 2019). Lipopolysaccharide‐binding protein (LBP) is a stable index that reflects LPS exposure (Asada et al., 2019). When the liver experiences injury, the balance of the gut microbiota is destroyed, which may participate in the onset and progression of liver injury. Deaminase and urease activation in the intestinal microbiota represent the major external sources of ammonia (Nicaise et al., 2008). The non‐absorbable disaccharide lactulose and poorly absorbed antibiotic rifaximin are the most common treatments for HE and function by decreasing ammonia production and absorption separately. However, the therapeutic effects are not obvious, and side‐effects, such as bloating, nausea and infection, cannot be ignored (Gluud et al., 2016; Yanny et al., 2019).
Previous studies have demonstrated that lactobacilli can efficiently treat hyperammonaemia and HE by remodelling the intestinal microbiota and reducing the production and absorption of ammonia (Nicaise et al., 2008; Singh et al., 2018). Lactobacillus salivarius Li01 (CGMCC 7045) found in healthy humans can significantly alleviate carbon tetrachloride (CCL4)‐induced liver cirrhosis and d‐galactosamine‐induced liver injury (Lv et al., 2014; Shi et al., 2017). In addition, L. salivarius Li01 exhibits tolerance to bile stress and possesses antibacterial and antifungal effects (Lv et al., 2017).
Thioacetamide (TAA), a classic hepatotoxic agent, has been extensively used for several years to develop liver injury models by bioactivating inducible nitric oxide synthase (INOS) and nuclear factor κB (NFκB), which can lead to inflammatory cytokine expression and centrilobular necrosis (Amanzada et al., 2014). Moreover, after TAA injection, the resulting neurobehavioral abnormalities resemble HE in patients (Shen et al., 2015).
Lactobacillus salivarius Li01 has been confirmed to improve liver cirrhosis and liver injury, and HE is a major complication of liver cirrhosis and liver injury (Lv et al., 2014; Shi et al., 2017). However, the protective effects of L. salivarius Li01 on hyperammonaemia are unclear. Therefore, we conducted this study to detect the therapeutic value of L. salivarius Li01 in TAA‐induced acute liver injury and hyperammonaemia and the potential mechanisms.
Results
L. salivarius Li01 reduced mortality and liver damage in a TAA‐induced mouse model
We administered a high‐dose TAA treatment to mice pretreated with phosphate‐buffered saline (PBS) or L. salivarius Li01 to detect the influence of L. salivarius Li01 on the mortality of TAA‐induced acute liver injury. The results indicated that the mice pretreated with L. salivarius Li01 had lower mortality than the mice pretreated with PBS (log‐rank test P < 0.01; Fig. 1B). After the administration of a lower dose of TAA, four of the ten mice in the positive control group (TP group, treated with TAA + PBS) died. However, the mortality in the probiotic intervention group (Li01 group, treated with TAA + L. salivarius Li01) was 20%, which was lower than the 40% mortality rate observed in the TP group. Haematoxylin and eosin (H&E) staining of liver sections showed evident destruction of the liver architecture, inflammatory cell infiltration and necrosis in the periportal areas, while L. salivarius Li01 could rehabilitate this type of liver damage (Fig. 1C). Consistent with the H&E staining, the dramatically elevated serum alanine aminotransferase (ALT; P < 0.001) and aspartate aminotransferase (AST; P < 0.001) levels induced by TAA could also be significantly reduced by L. salivarius Li01 (P < 0.001 and P < 0.05 respectively; Fig. 1E and F). The above results indicated that L. salivarius Li01 could improve TAA‐induced liver damage.
Fig. 1.

Lactobacillus salivarius Li01 intervention prolonged survival and improved liver injury. (A) Experimental scheme. CP group, normal mice treated with PBS; TP group, liver‐damaged mice treated with PBS; Li01 group, liver‐damaged mice treated with L. salivarius Li01. (B) Kaplan–Meier survival curves for the TP and Li01 groups, which were given high‐dose TAA. (C) Representative H&E staining of the liver. (D) Pathological scores of livers. (E) Serum levels of ALT. (F) Serum levels of AST. Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
L. salivarius Li01 pretreatment changed the structure of the gut microbiota
To explore the influence of L. salivarius Li01 on the structure of the gut microbiota, we conducted 16S rRNA gene sequencing of mouse faeces collected before the TAA injection. The rarefaction curve shown in Fig. S1 indicated that the amount of sequencing data is reasonable.
After the L. salivarius Li01 intervention, the α‐diversity of the gut commensals did not significantly change compared with that in the TP group (data not shown). The microbiome structure variance among the three groups was tested by UniFrac PCoA, permutational multivariate analysis (PERMANOVA or Adonis) and multiple response permutation procedure (MRPP). The L. salivarius Li01 intervention significantly changed the gut microbiome structure (Fig. 2A, Tables S2 and S3). The PCoA image showed that the microbiome clusters in the Li01 group were clearly distinct from those in the CP (treated with vehicle + PBS) and TP groups.
Fig. 2.
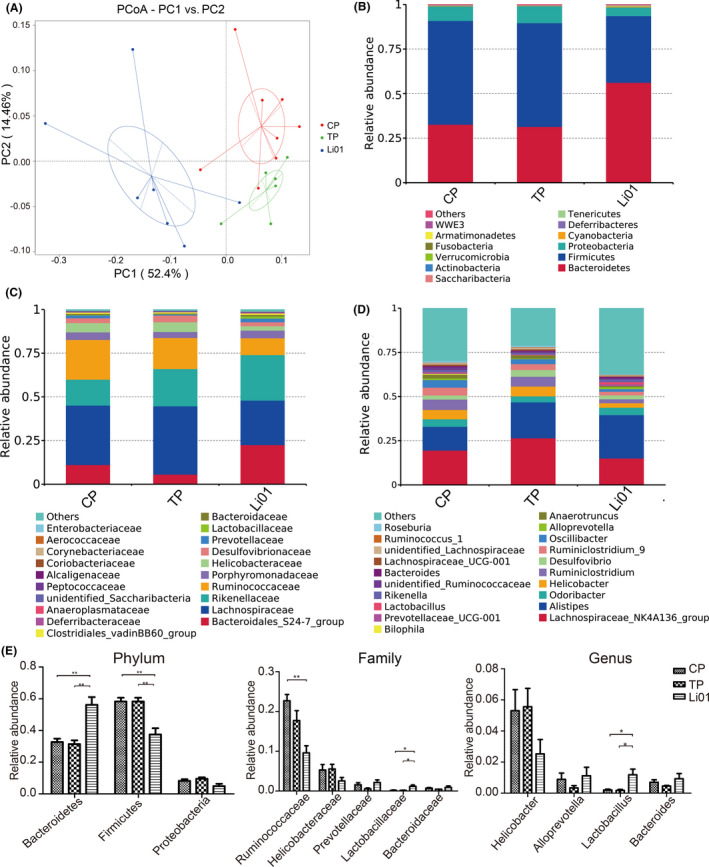
Lactobacillus salivarius Li01 intervention changed the structure of gut microbiota. (A) PCoA plot based on the weighted UniFrac metric. (B) Relative abundance of abundant taxa at phylum level. (C) Relative abundance of top twenty most abundant taxa at family level. (D) Relative abundance of top twenty most abundant taxa at genus level. (E) The specific taxa abundance at phylum (left), family (middle), genus (right) levels. Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
We detected the taxonomic abundance at the phylum, family and genus levels among the three groups (Fig. 2B–E). According to the results, compared with either the CP group or TP group, the L. salivarius Li01 supplementation significantly upregulated the relative abundance of Bacteroidetes and downregulated the richness of Firmicutes. Although there was no significant difference of Proteobacteria richness after the L. salivarius Li01 treatment, an elevated trend was observed. After the L. salivarius Li01 pretreatment, Ruminococcaceae, Helicobacteraceae and Helicobacter exhibited downward trends, and the richness of Lactobacillaceae and Lactobacillus was significantly increased.
L. salivarius Li01 pretreatment reshaped the aberrations in the gut microbiota caused by the TAA administration
To explore the change in the intestinal microbiota in the TAA‐induced mouse model of acute liver injury, we conducted 16S rRNA gene sequencing of mouse faeces collected 48 h after the TAA administration. The rarefaction curve reflected that the sequencing depth was reasonable (Fig. S2).
The α‐diversity of the gut microbiota in each of the three groups after the TAA administration was calculated by the Shannon index and Chao 1 metric. The TAA treatment increased the α‐diversity of gut commensals as indicated by noticeably increased Shannon and Chao 1 indices (P < 0.001 Fig. 3A and B). Compared with the TP group, the Li01 group exhibited reduced α‐diversity according to the Chao 1 indices.
Fig. 3.
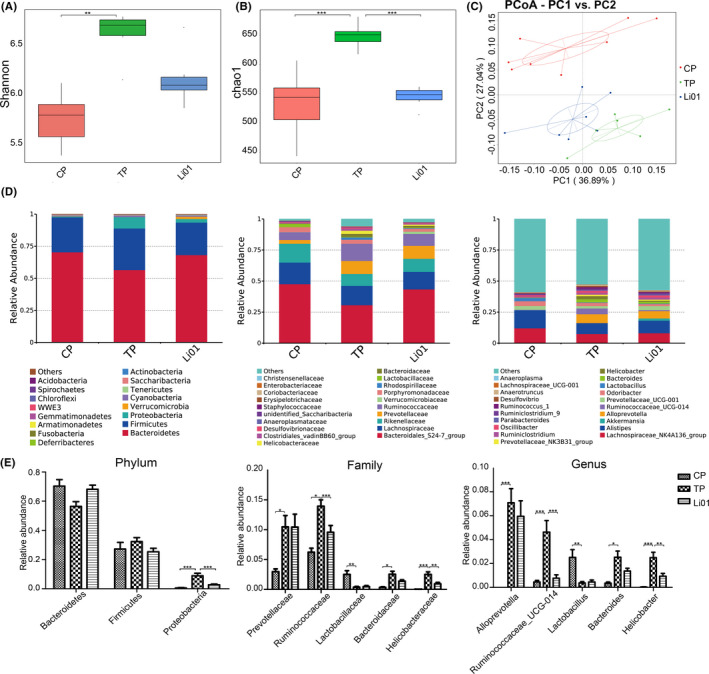
Alterations in the gut microbiota after TAA injection. (A) Shannon index. (B) Chao1 metric. (C) PCoA plot based on the weighted UniFrac metric. (D) Relative abundance of top twenty most abundant taxa at phylum (left), family (middle), genus (right) levels and bar charts of the specific taxa abundance. Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
There were marked differences in the commensal composition among the three groups after TAA injection (Tables S4 and S5). The PCoA image shows that the microbiome clusters of the three groups were clearly separated (Fig. 3C). The microbiome profile of the CP group was noticeably different from those of the other two groups. However, the distance in the microbiome profile between the Li01 group and CP group was shorter than that between the TP group and CP group, which corresponded with the R2 value and expected delta. The results indicated that the TAA injection could change the structure of the intestinal microbiota and that L. salivarius Li01 could partially restore the disordered microbiota.
At the phylum level, the TAA injection obviously increased the relative abundance of Proteobacteria (Fig. 3D, P < 0.001). However, compared with the TP group, the richness of Proteobacteria in the Li01 group was significantly decreased. At the family level, there were significant higher the relative abundances of Prevotellaceae, Ruminococcaceae, Bacteroidaceae, and Helicobacteraceae in the TP group than those in the CP group after the TAA injection (P < 0.05, P < 0.05, P < 0.05 and P < 0.001 respectively). The L. salivarius Li01 treatment dramatically reduced the relative abundances of Ruminococcaceae and Helicobacteraceae (P < 0.001 and P < 0.01). At the genus level, the TAA injection significantly increased the relative abundances of Alloprevotella, Ruminococcaceae_UCG‐014, Bacteroides and Helicobacter (P < 0.001, P < 0.001, P < 0.05 and P < 0.001 respectively), but compared with the TP group, the relative abundances of Ruminococcaceae_UCG‐014 and Helicobacter in the Li01 group were significantly reduced (P < 0.001 and P < 0.01). In addition, the TAA injection could significantly reduce the relative abundances of Lactobacillaceae and Lactobacillus (P < 0.01).
To further explore the key bacteria associated with TAA‐induced acute liver injury and the L. salivarius Li01 treatment, we conducted an LEfSe analysis (Fig. 4A‐D). The mice in the CP and TP groups had higher relative abundances of Bacteroidetes and Proteobacteria respectively. The families Ruminococcaceae (containing the genus Ruminococcaceae_UCG_014) and Prevotellaceae and the genus Alloprevotella were enriched in the TP group compared with those in the CP group. The other taxa enriched in the TP group, such as the order Campylobacterales, family Helicobacteraceae and genus Helicobacter, all belonged to the phylum Proteobacteria. In addition, the TAA injection could significantly reduce the abundance of the genus Lactobacillus. However, after the L. salivarius Li01 intervention, the reduced levels of the phylum Bacteroidetes and upregulated enrichment of the phylum Proteobacteria and family Ruminococcaceae were partially restored. In addition, Akkermansia plays a protective role in many liver diseases, such as alcoholic liver disease, non‐alcoholic fatty liver disease and immune‐mediated liver injury (Wu et al., 2017; Zhu et al., 2017; Grander et al., 2018). The L. salivarius Li01 administration increased the abundance of the genus Akkermansia. The results indicated that the L. salivarius Li01 intervention could reshape the disorder in the microbiota induced by TAA.
Fig. 4.
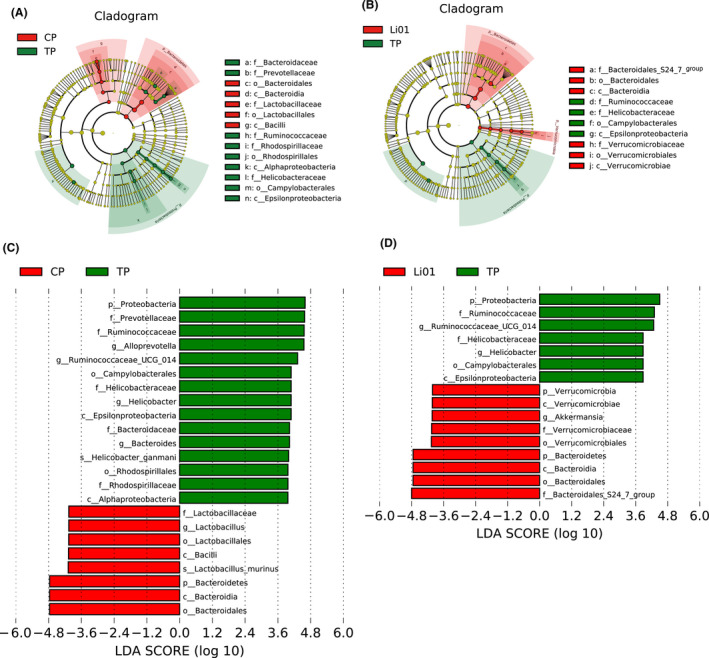
Effects of pretreatment with Lactobacillus salivarius Li01 on the alterations of gut bacterial taxonomic abundance after TAA injection. (A and B) LEfSe cladogram. (C and D) Discriminative biomarkers with an LDA score > 3.8. Red represents the CP or Li01 group, and green represents the TP group.
L. salivarius Li01 intervention partially restored the gut barrier function
The overgrowth of gut bacteria and damage to the intestinal barrier has been shown to be correlated with liver injury caused by TAA (Harputluoglu et al., 2012). Hence, we detected the colonic mRNA expression of zonula occludens‐1 (ZO‐1), claudin‐1 and mucin2 (MUC2) to assess the gut barrier function. The TAA administration destroyed the gut barrier as evidenced by a decreased expression of ZO‐1 and claudin‐1 (P < 0.001 and P < 0.05 respectively; Fig. 5A and B); however, the L. salivarius Li01 intervention ameliorated this impact as indicated by significant improvements in the ZO‐1 and MUC2 transcriptional levels (P < 0.05). Although the upregulation of claudin‐1 expression was not significant, an increasing trend existed. To further test our hypothesis, we conducted immunofluorescence staining for ZO‐1(Fig. 5D). This result was consistent with the mRNA expression results for ZO‐1 in the colon, indicating that the breakdown of ZO‐1 induced by TAA was partially restored by the L. salivarius Li01 intervention.
Fig. 5.
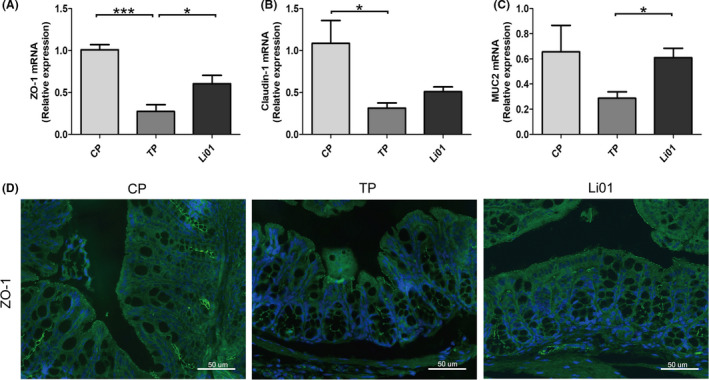
Lactobacillus salivarius Li01 reinforced gut barrier function. (A–C) Relative mRNA expression of ZO‐1, Claudin‐1 and MUC2 in the colon. (D) Representative immunofluorescence staining for ZO‐1 in the colon (scale bar: 50 µm). Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
L. salivarius Li01 attenuated the serum LBP levels and the TAA‐induced activation of the NFκB pathway
When the gut barrier is damaged, microorganisms and their components, represented by LPS, break through the gut barrier. LPS is related to the onset and progression of liver diseases and can activate the NFκB pathway via Toll‐like receptor 4 (TLR4) and myeloid differentiation 88 (MyD88). LBP is a stable index that reflects LPS exposure (Asada et al., 2019). Therefore, we detected the serum concentration of LBP. The results showed that the LBP levels in the Li01 group were significantly lower (Fig. 6A, P < 0.05) than those in the TP group, indicating that L. salivarius Li01 could improve the upregulation of the LPS level induced by TAA (P < 0.01, Fig. 4A). In addition, the relative mRNA expression of TLR4, CD14 (a co‐receptor of TLR4) and MyD88 was assessed by RT‐PCR and could be downregulated by the administration of L. salivarius Li01 (P < 0.01, P < 0.05 and P < 0.05 respectively; Fig. 6B–D). In addition, we assessed phospho‐NFκB p65 nuclear translocation in the liver by immunofluorescence. We found that after the TAA administration, mice had an increased fluorescence intensity for phospho‐NFκB p65 in the nucleus (Fig. 4E). Moreover, L. salivarius Li01 could downregulate the nuclear translocation of phospho‐NFκB p65 induced by the TAA injection. Therefore, we hypothesized that L. salivarius Li01 could improve the LPS levels and downregulate NFκB pathway activation to alleviate TAA‐induced acute liver injury.
Fig. 6.
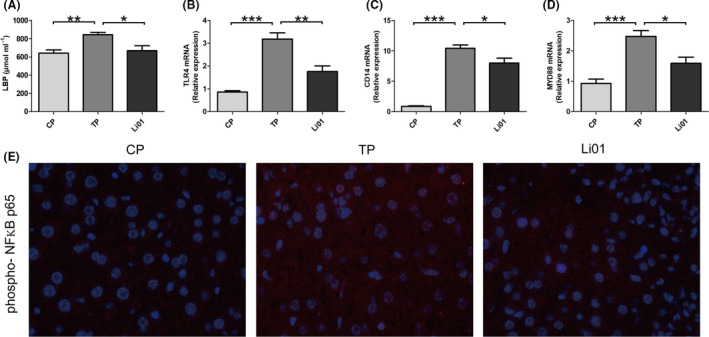
Lactobacillus salivarius Li01 alleviated serum LBP levels and suppressed the activation of the NFKB pathway. (A) Serum LBP levels. (B) Relative hepatic mRNA expression of TLR4, CD14 and MYD88. (C) Representative immunofluorescence staining for phospho‐NFκB p65 in the liver (scale bar: 50 µm). Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
L. salivarius Li01 mitigated the TAA‐induced inflammatory response and recruitment of macrophages and neutrophils into the liver
Thioacetamide induces the activation of the NFκB pathway, leading to the expression of pro‐inflammatory factors and chemokines (Shapiro et al., 2006). Can L. salivarius Li01 improve TAA‐induced inflammation? To address this question, we investigated the pro‐inflammatory cytokine and chemokine mRNA levels in the liver. The mice in the Li01 group exhibited a significantly lower relative expression of Interleukin‐6 (IL‐6), monocyte chemotactic protein 1(MCP1) and chemokine (C‐X‐C motif) ligand 1(CXCL1) than the mice in the TP group (P < 0.001, P < 0.001 and P < 0.01 respectively; Fig. 7C). In addition, we detected the serum levels of some pro‐inflammatory cytokines. Forty‐eight hours after the TAA challenge, the serum concentrations of pro‐inflammatory factors (Interleukin (IL)‐17A and IL‐6) and chemokines (MCP‐1, CXCL1 and RANTES) were significantly upregulated (Fig. 7A and B). However, compared with those in the TP group, the cytokine concentrations in the Li01 group were evidently mitigated.
Fig. 7.
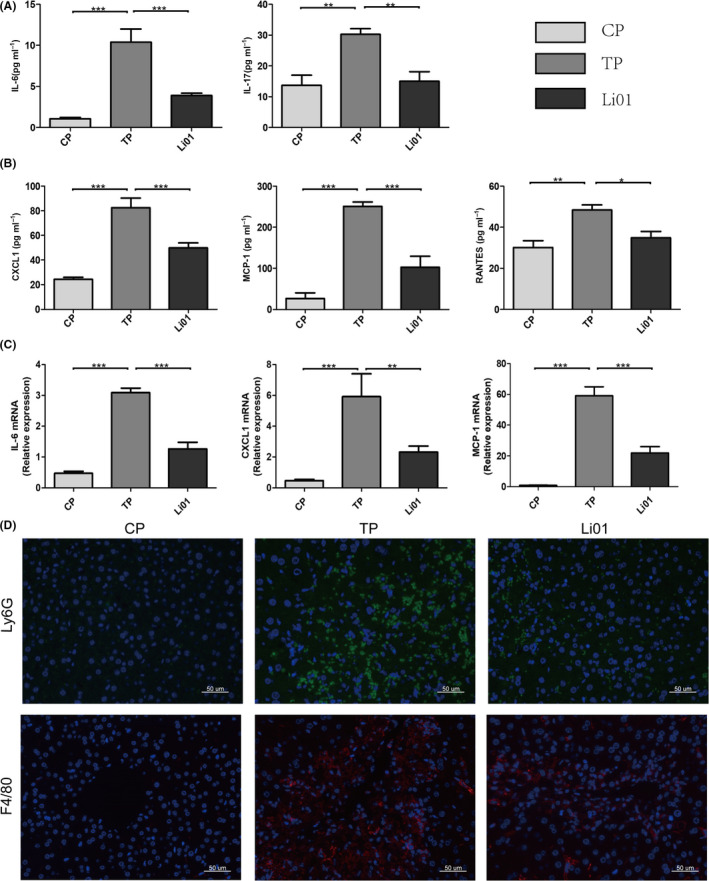
Lactobacillus salivarius Li01 relieved TAA‐induced systemic and hepatic inflammation, and reduced the hepatic recruitment of macrophages and neutrophils. (A) Expression of the cytokines IL‐6 and IL‐17A in the serum. (B) Expression of the chemokines CXCL1, MCP1 and RANTES in the serum. (C) Relative mRNA expression of IL‐6, CXCL1 and MCP1 in the liver. (D) Representative immunofluorescence staining for F4/80 (macrophage marker) and Ly6G (neutrophil marker) in the liver. Scale bar: 50 µm. Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
Thioacetamide injection can induce the accumulation of macrophages and neutrophils in the liver (Amanzada et al., 2014). MCP1 and RANTES can regulate macrophage recruitment, and CXCL1 is a chemoattractant of neutrophils. In our study, L. salivarius Li01 improved the upregulation of the levels of these chemokines. Whether L. salivarius Li01 can decrease the infiltration of macrophages and neutrophils requires further exploration. For this reason, we conducted immunofluorescence staining for macrophages (F4/80+) and neutrophils (Ly6G+) in the liver, and the images show that the TAA injection could promote macrophage (F4/80+) and neutrophil (Ly6G+) migration into the liver (Fig. 7D). Moreover, in the Li01 group, the infiltration of macrophages and neutrophils was sporadic.
L. salivarius Li01 intervention led to reductions in blood and faecal ammonia concentrations and neuro‐inflammation levels and restored cognitive function
Acute liver injury is often accompanied by hyperammonaemia, which is closely related to HE (Blei et al., 2001; Nicaise et al., 2008). The gut is regarded as an important source of circulating ammonia (van de Poll et al., 2008). Therefore, the faecal and plasma ammonia concentrations were tested. The blood and faecal ammonia levels were significantly higher in the TP group than those in the CP group (P < 0.001 and P < 0.001 respectively; Fig. 8A). However, after the L. salivarius Li01 administration, the faecal ammonia levels were evidently decreased (P < 0.05). Consistent with the improvement in the faecal ammonia level, the reduction in the blood ammonia level was also significant (P < 0.01).
Fig. 8.
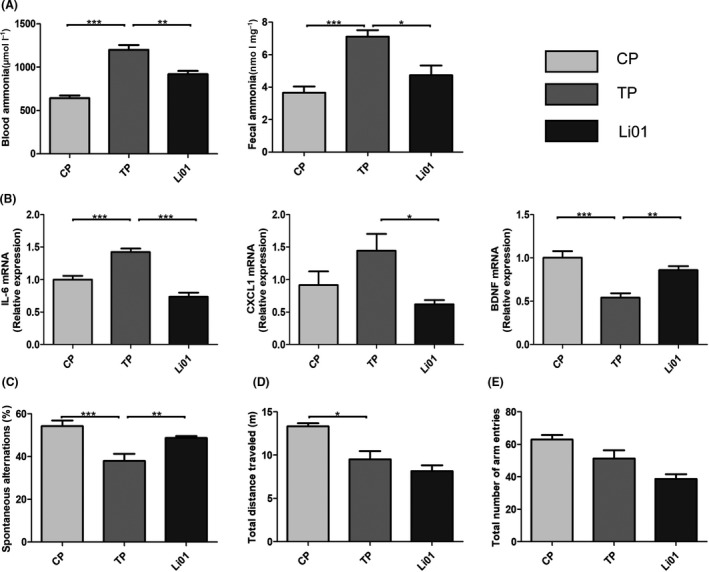
Lactobacillus salivarius Li01 downregulated plasma and faecal ammonia levels and improved neuro‐inflammation and cognitive function. (A) Plasma and faecal ammonia levels. (B) Relative expression of the IL‐6, CXCL1 and BDNF genes in the cortex. (C) Testing of spontaneous alternation with a Y maze at 48 h after TAA administration. (D) Total distance travelled. (E) Total number of arm entries within 8 min. Data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
The relative mRNA expression of IL‐6 and CXCL1 in the cortex increased after the TAA administration (Fig. 8B), whereas the L. salivarius Li01 intervention drastically improved this impact (P < 0.001 and P < 0.05 respectively).
Brain‐derived neurotrophic factor (BDNF) plays a pivotal role in not only the growth and survival of neurons (Barde et al., 1982; Leibrock et al., 1989) but also the acquisition, maintenance and recall of spatial memory (Mizuno et al., 2003). As shown in Fig. 8B, the TAA administration noticeably downregulated BDNF mRNA expression in the cortex (P < 0.001); however, the downregulation of BDNF expression was attenuated by the L. salivarius Li01 intervention (P < 0.001).
We conducted a Y maze test to evaluate the cognitive impairment caused by TAA and the protective effect of L. salivarius Li01. Our results illustrate that compared with the CP group, the TAA challenge significantly decreased spontaneous alternations in the TP group (P < 0.001, Fig. 8C). However, L. salivarius Li01 had a significant protective effect on these spontaneous alternations (P < 0.05). The locomotor activity of the mice was quantified by measuring the total arm entries and distance travelled. The mice in the CP group travelled a markedly longer distance than those in the TP group (P < 0.05). Nevertheless, the mice treated with either L. salivarius Li01 or PBS did not display a sufficient difference in the total distance travelled. In addition, the total arm entries did not change after the TAA administration or L. salivarius Li01 treatment. L. salivarius Li01 possessed protective effects on only cognitive function in the mice but had no beneficial effect on locomotor activity.
Correlations between the gut microbiota and gut barrier markers and systemic, hepatic and neuro‐inflammation
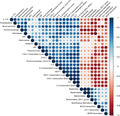
To further assess the influence of the altered gut microbiota, we conducted a correlation analysis between discriminative bacterial genera and injury parameters (Fig. 9). Markers of systemic and hepatic inflammation, including IL‐6 and MCP‐1, were significantly positively correlated with the relative abundances of Proteobacteria, Helicobacter and Ruminococcaceae_UCG.014 (P < 0.05), which were enriched in the TP group. In addition, the population density of the Bacteroidales_S24.7_group, which was prominently abundant in the Li01 group, was negatively associated with the serum levels of CXCL1 and IL‐6 and the expression of the IL‐6 and MCP1 genes in the liver (P < 0.05). On the one hand, the plasma ammonia concentrations had a significant positive relationship with the ammonia levels in the faeces (P < 0.05) and negative relationships with the mRNA levels of MUC2 (P < 0.05) and ZO1 (P < 0.01) in the colon. On the other hand, the neuro‐inflammation parameters (IL‐6 and CXCL1 mRNA levels) and BDNF mRNA levels were closely tied to the blood ammonia levels, systemic inflammation, liver function and the enrichment in Ruminococcaceae_UCG.014. Additionally, the spontaneous alternations had negative correlations with Proteobacteria (P < 0.01); Ruminococcaceae (P < 0.01); Helicobacter (P < 0.001); serum levels of ALT, IL‐17A, IL‐6, MCP1 and CXCL1; and hepatic mRNA expression of IL‐6 and MCP1 and a positive correlation with the Bacteroidales_S24.7_group (P < 0.05). Therefore, we speculate that L. salivarius Li01 modified‐gut microbiota played a pivotal role in limiting the detrimental effects of the TAA challenge.
Fig. 9.
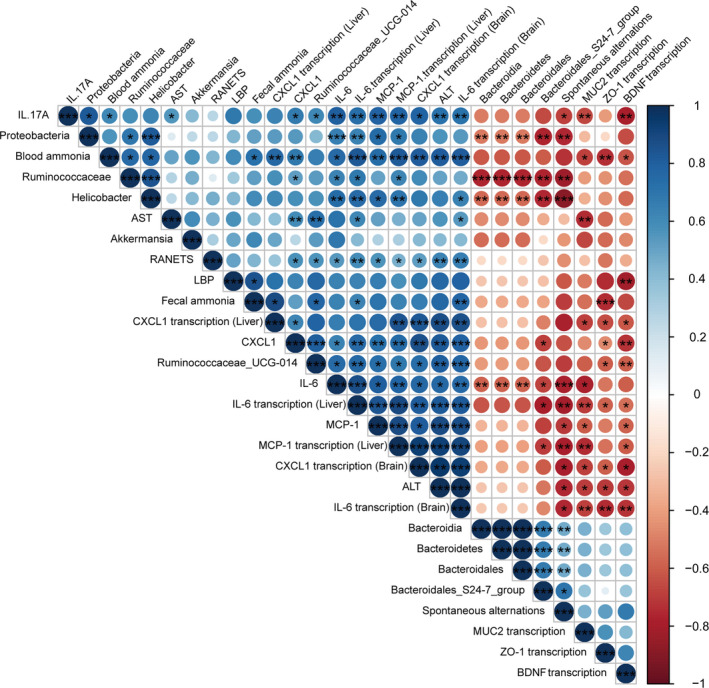
Correlation analysis. Spearman rank correlation analysis was used to detect associations between the discriminative gut microbiota and injury‐related parameters. The colour key and circle size show the strength of the correlation. Dark blue represents a positive correlation; red represents a negative correlation. *P < 0.05, **P < 0.01, ***P < 0.001.
Discussion
The perturbed gut‐liver‐brain axis has emerged as a pivotal component of liver diseases and complication HE (Wiest et al., 2017). Dysbiosis of the gut microbiota not only contributes to some bowel diseases but also influences the onset and progression of some extraintestinal diseases, including acute liver injury. Probiotic intervention is considered a potential therapeutic strategy.
Our results indicated that the L. salivarius Li01 pretreatment decreased mortality and alleviated liver injury by downregulating the serum transaminase levels and reducing the histological damage. The protective effects of L. salivarius Li01 partially depended on reshaping the intestinal microbiota, strengthening the intestinal barrier and improving systemic and hepatic inflammation. Moreover, L. salivarius Li01 ameliorated HE by alleviating liver damage and decreasing the plasma and faecal ammonia levels.
The gut microbiota, mucins, the continuously regulated turnover of intestinal epithelial cells and tight junction complexes are essential components of the gut barrier (Cliffe et al., 2005; McDole et al., 2012; Ramanan and Cadwell, 2016). Our results showed that the TAA administration destroyed the gut barrier as evidenced by the decreased expression of ZO‐1 and Claudin‐1, and the L. salivarius Li01 treatment restored the gut barrier manifested as the increase in ZO‐1 and MUC2. Microorganisms, microbial components and metabolites can all affect extraintestinal organs by disrupting the intestinal barrier (Nakamoto et al., 2017). L. salivarius Li01 reshaped the gut microbiota, including Akkermansia and Alloprevotella, which are associated with the gut barrier function (Bian et al., 2019; Xing et al., 2019). Our findings demonstrate that L. salivarius Li01 decreased the serum IL‐6, IL‐17, CXCL1, MCP1 and RANTES levels which may be a pivotal medium for damage to the gut barrier function (Soares et al., 2010). The effect of L. salivarius Li01 on reshaping the gut microbiota and alleviating systemic inflammation may contribute to restoring the destroyed gut barrier damaged by TAA.
Lipopolysaccharide, also known as endotoxin, is located in the bacterial cell wall and is a unique component of gram‐negative bacteria. Normally, LPS transfer to extraintestinal organs can be prevented by an intact intestinal barrier. However, in our TAA‐induced ALF mouse model, the gut barrier was damaged, and LPS could easily break through the impaired intestinal barrier as evidenced by the elevated serum LBP levels which is a stable index that reflects LPS exposure (Yang et al., 2017). LBP and CD14 are essential mediators of the recognition of LPS by the TLR4‐MD2 complex (Ryu et al., 2017). Therefore, the expression of TLR4 and CD14 in the liver was increased in TP group. Subsequently, the expression of MYD88 was increased in the TP group, MYD88‐dependent signalling was activated, the translocation of phosphorylated NFκB p65 into the nucleus was increased, and the NFκB signalling pathway was activated after the TAA treatment (Kagan and Medzhitov, 2006). However, L. salivarius Li01 restored the gut microbiota and barrier function, which may contribute to the lower level of serum LPS, and further activation of the NFκB signalling pathway induced by the TAA injection was inhibited as evidenced by the decreased serum LBP levels, hepatic TLR4, CD14 and MYD88 mRNA levels, and the translocation of phosphorylated NFκB p65 into the nucleus. The activation of the NFκB signalling pathway can induce the release of cytokines and chemokines and the recruitment of inflammatory cells (Amanzada et al., 2014). In our study, the relative mRNA abundances of IL‐6, MCP1 and CXCL1 in the liver and those of the systemic inflammatory parameters, including IL‐6, IL‐17, MCP1, KC and RANENTS, were upregulated after the TAA injection. Numerous chemokines and cytokines are secreted by Kupffer cells, liver myofibroblasts (LMFs) and biliary cells, which are mostly located in the hepatic portal area (Sprenger et al., 1997; Amanzada et al., 2014), which might explain why the liver damage mostly presented in the periportal tracts as shown by the H&E staining. Macrophages can both accentuate inflammation and mediate inflammation resolution and tissue repair during liver injury (Li et al., 2019). The generation of reactive oxygen species (ROS) is considered to contribute neutrophils to liver injury (Jaeschke and Hasegawa, 2006). Chemokines play an important role in recruiting inflammatory cells. MCP1, RANTES and KC are known chemoattractant of inflammatory cells. Our results showed that the infiltration of macrophages and neutrophils was increased, which was consistent with the elevations in the MCP1, RANTES and CXCL1 levels in the positive control group (TP group). However, after the L. salivarius Li01 intervention, the release of chemokines and recruitment of inflammatory cells were both significantly alleviated. Therefore, we speculated that the improvement in the gut barrier function by L. salivarius Li01 reduced the serum LPS level, inhibited the activation of the NFκB signalling pathway, and further alleviated systemic and hepatic inflammation, which may play an important role in the mechanism by which L. salivarius Li01 improved TAA‐induced acute liver injury.
The gut microbiota, which contains many organisms with different genomes, is considered a previously forgotten organ that plays important roles in physiological processes, such as nutrient digestion and self‐defence. In our study, we revealed that the TAA‐induced acute liver injury in our mouse model destroyed the intestinal microbiota homeostasis, which was accompanied by Lactobacillus deficiency. However, the L. salivarius Li01 intervention changed the structure of the gut microbiota and increased the relative abundance of Lactobacillus. The TAA injection favoured Proteobacteria and Helicobacter, which include many pathogens, have been related to liver injury in a previous study (Gu et al., 2020), and were positively correlated with systematic inflammation in our study. However, the L. salivarius Li01 pretreatment reduced these bacteria. A previous study demonstrated that Alloprevotella was associated with intestinal villus damage (Xing et al., 2019). Notably, the relative abundance of Alloprevotella was increased in the TP group in which the intestinal barrier function was damaged. In addition, Ruminococcaceae_UCG‐014 was negatively correlated with the short‐chain fatty acid level in the faeces, which plays a protective role in non‐alcoholic steatohepatitis (Zhang et al., 2019). An overgrowth of Ruminococcaceae_UCG‐014 was found in the TP group and was closely related to systemic inflammation and liver function. Supplementing with L. salivarius Li01 could recover the TAA‐induced depletion of Bacteroidetes and favour Akkermansia. Accumulating studies have elucidated that Akkermansia, a butyrate‐producing genus, has beneficial effects on liver damage and the gut barrier function (Wu et al., 2017; Bian et al., 2019). Our results demonstrate that L. salivarius Li01 reshaped the disturbed gut microbiota caused by the TAA treatment, which could alleviate acute liver injury.
Thioacetamide injection can induce acute progressive liver damage with parallel involvement of hyperammonaemia and is recognized as a reliable mouse model of acute liver injury and HE (Shen et al., 2015). However, the damaged liver cannot eliminate gut‐derived ammonia as evidenced by the positive relationship between the blood ammonia levels and the severity of HE in the clinic (Weissenborn et al., 2007). In our study, the decreased spontaneous alternations and expression of BDNF and the increased expression of IL‐6 in the TP group indicated that the TAA treatment‐induced neuro‐inflammation and destroyed cognitive function. Plasm ammonia was significantly upregulated in the TP group and closely correlated with neuro‐inflammation and the relative expression of the BNDF gene. Therefore, we speculate that hyperammonaemia plays an important role in HE. The gut microbiota can produce ammonia, which is regarded as a toxin from the gut associated with neurotoxicity based on highly convincing evidence (Norenberg, 1996). This theory was confirmed in our study by the elevated faecal ammonia level in the TP group and the positive correlation between blood and faecal ammonia. The common medicines used to treat hyperammonaemia include lactulose and rifaximin, and their mechanisms are based on regulating the intestinal microbiome (Gluud et al., 2016). However, the existing treatment schemes have some shortcomings that limit their clinical use. Interventions with probiotics are considered potentially effective as a treatment for HE. In this study, L. salivarius Li01 not only alleviated the TAA‐induced acute liver injury but also improved the complication HE as evidenced by the elevated spontaneous alternations and BDNF mRNA level and reduced expression of IL‐6 and CXCL1 in the brain. On the one hand, the correlation analysis showed the close correlation among blood ammonia, faecal ammonia and microbiota. The L. salivarius Li01 intervention restored the gut microbiota, which may contribute to the simultaneous declines in the blood and faecal ammonia levels. Accordingly, we hypothesized that L. salivarius Li01 decreases the level of gut‐derived ammonia to reduce the blood ammonia levels and further improve HE. On the other hand, as L. salivarius Li01 relieved the TAA‐induced liver injury, the ability to eliminate ammonia in the liver was improved.
Nevertheless, some limitations exist in our study. Although our results indicate that close relationships exist between the gut microbiota and some parameters of systemic, hepatic and neuro‐inflammation, the mechanisms need to be fully explored and verified.
Conclusion
Our findings show the beneficial role of L. salivarius Li01 in TAA‐induced acute liver injury and HE in mice. We hypothesize that L. salivarius Li01 improves the damage induced by TAA via reshaping the gut microbiota, reinforcing the gut barrier function, downregulating NFκB pathway activation, alleviating systemic inflammation, reducing inflammatory cell infiltration into the liver and decreasing ammonia levels. Therefore, these results indicate that L. salivarius Li01 may be a potential probiotic that can be used against acute liver injury and its complication HE.
Experimental procedures
Strains and culture conditions
Lactobacillus salivarius Li01 isolated from the faeces of healthy individuals was grown anaerobically in MRS medium for 24 h at 37 °C (Lv et al., 2017). The cultures were centrifuged at 8000 g for 10 min. Then, the supernatant was discarded, and the precipitate was washed twice with sterile PBS and resuspended in PBS at a final concentration of 3 × 109 CFUs ml−1 by testing the absorbance at 630 nm (O.D. range from 0.6 to 0.8; Bian et al., 2019). Fresh L. salivarius Li01 suspension was used in the animal experiment.
Animals
Male C57BL/6 mice (6–8 weeks, Shanghai SLAC Laboratory Animal, Co, Ltd, Shanghai, China) were housed for 7 days for adaptation to conventional laboratory conditions (room temperature: 20–22 °C, 12‐h light/dark cycle, free access to food and drinking water). Subsequently, the mice were randomly assigned to three groups and treated with 0.2 ml vehicle (groups CP n = 8 and TP n = 10) or a fresh L. salivarius Li01 suspension (3 × 109 CFUs ml−1, group Li01 n = 10) per day by intragastric infusion for one week (Fig. 1A). The acute liver injury and hyperammonaemia model in the TP and Li01 groups were induced via a single intraperitoneal (i.p.) injection of 300 mg kg−1 TAA (Sigma‐Aldrich, St. Louis, MO, United States). Before the TAA injection, a faecal sample was collected from each mouse. The Y maze neurobehavioral test was conducted 48 h after the TAA administration. Then, the mice were sacrificed, and the liver, colon, brain tissue, blood and intestinal contents were collected for the subsequent tests.
Survival curves
The mice were given a single high dose of TAA (600 mg kg−1) by i.p. injection after 7 days of L. salivarius Li01 suspension (3 × 109 CFUs ml−1 in PBS) or PBS administration. The survival time of the mice was recorded (Shen et al., 2015).
Neurobehavioral test
The memory and spatial learning of the mice were assessed by the Y maze test, which was conducted with an apparatus with three identical black plastic arms. Normally, a mouse would investigate new arms. Before the test, the mice were acclimated to the testing environment for 30 min. Each mouse was released at the end of one arm and allowed to navigate freely for 8 min. All mice were placed in the same place. The total number of arm entries, distance travelled and spontaneous alternations (entering the three different arms sequentially) were recorded. The percentage of spontaneous alternations was defined by the following formula: [(number of alternations)/(total arm entries‐2)] × 100 (Shen et al., 2015; Ghafouri et al., 2016).
Biochemical assays
The serum was separated from blood samples by centrifugation at 3000 g for 10 min at 4 °C. Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were evaluated with a dry chemistry analyser (FUJI DRI‐CHEM 7000V, FUJIFILM, Tokyo, Japan). The LBP levels were determined with LBP ELISA kits (Guduo, Shanghai, China). The serum cytokine levels were measured with magnetic bead suspension arrays (Bio‐Rad, Hercules, CA, USA).
Plasma and faecal ammonia
Blood was treated with the anticoagulant heparin and centrifuged at 1000 g for 10 min at 4 °C to obtain plasma. The plasma ammonia concentrations were measured using an Ammonia Assay kit (Abcam, Cambridge, MA, USA). The faecal samples (100 mg) were suspended in 1 ml assay buffer, and the supernatants were obtained by centrifugation at 13 000 g for 10 min at room temperature (Shen et al., 2015). The faecal ammonia levels were detected according to the manufacturer's protocols.
Histopathology and immunofluorescence
Appropriately sized samples were obtained from the left liver and colon and immediately fixed in 10% formalin for 24 h. Then, paraffin was used to embed the tissue. H&E staining and a histological activity index analysis were performed to estimate liver damage (Knodell et al., 1981).
The liver samples were cut into 4‐µm‐thick sections and stained with anti‐phospho‐p65, anti‐Ly6G, or anti‐F4/80 antibodies. The colon samples were cut and stained with anti‐ZO‐1 (Proteintech, Rosemont, IL, USA) antibodies according to a previously reported immunofluorescence protocol (Chung et al., 2014). The images were visualized by P250 FLASH (3D HISTECH, Budapest, Hungary).
RNA extraction and real‐time PCR analysis
The liver cortex and colon samples were collected at sacrifice and stored at −80 °C. The total RNA was extracted by using a RNeasy Mini Kit (QIAGEN, Hilden, Germany). The detailed steps are described in the product manuals. PrimeScript RT Master Mix (TAKARA Biomedicals, Kusatsu, Japan) was applied to accomplish reverse transcription. The mRNA relative abundance was measured by SYBR Premix Ex Taq II reagent (TAKARA Biomedicals, Kusatsu, Japan) using an Applied Biosystems VIIA7 Real‐time PCR system (see Table S1 for the primer information). We included two biological replicates for each sample in the test. GAPDH was employed as an internal control for the liver and brain samples. β‐actin was used as the internal control for the colon samples.
Analysis of the microbiota
Faecal pellets were collected before the TAA injection and sacrifice and immediately stored in a −80 °C freezer. We performed the stool DNA extraction with the QIAamp Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany). Briefly, 300 µl 0.1 mm zirconia silica beads and 1 ml InhibitEX buffer were added to the samples, and then, the samples were homogenized twice at 5000 rpm for 30 s. The next steps referred to the manufacturer’s recommendations. The distinct region V3‐V4 of 16S rRNA was amplified using Phusion® High‐Fidelity PCR Master Mix (New England Biolabs). The PCR products were mixed and then purified with a GeneJET™ Gel Extraction Kit (Thermo Scientific). Sequencing was performed using an Ion S5™ XL platform (Thermo Scientific). The data were split and filtered, and the chimaeras were removed to obtain clean reads. Clean reads with ≥ 97% sequence similarity were clustered into operational taxonomic units (OTUs) by Uparse software. The species annotation was performed using the SILVA database. The alpha diversity was analysed by QIIME (Version 1.7.0). The principal coordinate analysis (PCoA) was conducted with R software (Version 2.15.3) according to a distance matrix of weighted UniFrac. The linear discriminant analysis effect size (LefSe) analysis was performed online (Bajaj et al., 2014).
Statistical analysis
The differences in the Kaplan–Meier survival curves were analysed by the log‐rank test. The data are expressed as the mean ± SEM. The significance of the differences in the ALT, AST, LBP, serum cytokine, plasma ammonia and faecal ammonia levels and relative abundances were determined by a one‐way analysis of variance (ANOVA). Tukey’s or Dunnett’s tests were conducted to adjust for multiple comparisons. PERMANOVA and MRPP were applied to analyse the β‐diversity of the gut microbiota. The correlations between different variables were analysed by a Spearman rank correlation analysis using R software. SPSS 20.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad prism (GraphPad Software, Inc., CA, USA) were used to complete the statistical work. A P‐value < 0.05 was considered statistically significant.
Conflict of interest
The authors declare that they have no competing interests.
Supporting information
Fig. S1. The rarefaction curve reflected the sequencing depth of the faecal 16S rRNA sequencing analysis before TAA treatment.
Fig. S2. The rarefaction curve reflected the sequencing depth of the faecal 16S rRNA sequencing analysis after TAA treatment.
Table S1. Specific primers applied for RT‐PCR test.
Table S2. PERMANOVA test of community structure differences in the gut microbiota among groups after L. salivarius Li01 intervention.
Table S3. MRPP test of community structure differences in the gut microbiota among groups after L. salivarius Li01 intervention.
Table S4. PERMANOVA test of community structure differences in the gut microbiota among groups after TAA injection.
Table S5. MRPP test of community structure differences in the gut microbiota among groups after TAA injection.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (81790631), the National Key Research and Development Program of China (2018YFC2000500), and the National Natural Science Foundation of China (81570512).
The authors gratefully acknowledge Daohui Zhang from the Core Facilities Platform of the Zhejiang University School of Medicine for support and help with the neurobehavioral test.
Microbial Biotechnology (2020) 13(6), 1860–1876
Funding information
This study was supported by the National Natural Science Foundation of China (81790631), the National Key Research and Development Program of China (2018YFC2000500), and the National Natural Science Foundation of China (81570512).
Data availability statement
The 16S rRNA gene sequencing data were deposited in the sequence read archive (SRA) database (accession no. PRJNA636106).
References
- Amanzada, A. , Moriconi, F. , Mansuroglu, T. , Cameron, S. , Ramadori, G. , and Malik, I.A. (2014) Induction of chemokines and cytokines before neutrophils and macrophage recruitment in different regions of rat liver after TAA administration. Lab Invest 94: 235–247. [DOI] [PubMed] [Google Scholar]
- Asada, M. , Oishi, E. , Sakata, S. , Hata, J. , Yoshida, D. , Honda, T. , et al (2019) Serum lipopolysaccharide‐binding protein levels and the incidence of cardiovascular disease in a general Japanese population: the hisayama study. J Am Heart Assoc 8: e013628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajaj, J.S. , Heuman, D.M. , Hylemon, P.B. , Sanyal, A.J. , White, M.B. , Monteith, P. , et al (2014) Altered profile of human gut microbiome is associated with cirrhosis and its complications. J Hepatol 60: 940–947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barde, Y.A. , Edgar, D. , and Thoenen, H. (1982) Purification of a new neurotrophic factor from mammalian brain. EMBO J 1: 549–553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernal, W. , Lee, W.M. , Wendon, J. , Larsen, F.S. , and Williams, R. (2015) Acute liver failure: a curable disease by 2024? J Hepatol 62: S112–S120. [DOI] [PubMed] [Google Scholar]
- Bian, X. , Wu, W. , Yang, L. , Lv, L. , Wang, Q. , Li, Y. , et al (2019) Administration of akkermansia muciniphila ameliorates dextran sulfate sodium‐induced ulcerative colitis in mice. Front Microbiol 10: 2259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blei, A.T. , Córdoba, J. , and Practice Parameters Committee of the American College of Gastroenterology (2001) Hepatic encephalopathy. Am J Gastroenterol 96: 1968–1976. [DOI] [PubMed] [Google Scholar]
- Chung, C.Y. , Alden, S.L. , Funderburg, N.T. , Fu, P. , and Levine, A.D. (2014) Progressive proximal‐to‐distal reduction in expression of the tight junction complex in colonic epithelium of virally‐suppressed HIV+ individuals. PLoS Pathog 10: e1004198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cliffe, L.J. , Humphreys, N.E. , Lane, T.E. , Potten, C.S. , Booth, C. , and Grencis, R.K. (2005) Accelerated intestinal epithelial cell turnover: a new mechanism of parasite expulsion. Science 308: 1463–1465. [DOI] [PubMed] [Google Scholar]
- Ghafouri, S. , Fathollahi, Y. , Javan, M. , Shojaei, A. , Asgari, A. , and Mirnajafi‐Zadeh, J. (2016) Effect of low frequency stimulation on impaired spontaneous alternation behavior of kindled rats in Y‐maze test. Epilepsy Res 126: 37–44. [DOI] [PubMed] [Google Scholar]
- Gluud, L.L. , Vilstrup, H. , and Morgan, M.Y. (2016) Nonabsorbable disaccharides for hepatic encephalopathy: a systematic review and meta‐analysis. Hepatology 64: 908–922. [DOI] [PubMed] [Google Scholar]
- Gorg, B. , Schliess, F. , and Haussinger, D. (2013) Osmotic and oxidative/nitrosative stress in ammonia toxicity and hepatic encephalopathy. Arch Biochem Biophys 536: 158–163. [DOI] [PubMed] [Google Scholar]
- Grander, C. , Adolph, T.E. , Wieser, V. , Lowe, P. , Wrzosek, L. , Gyongyosi, B. , et al (2018) Recovery of ethanol‐induced Akkermansia muciniphila depletion ameliorates alcoholic liver disease. Gut 67: 891–901. [DOI] [PubMed] [Google Scholar]
- Gu, Z. , Wu, Y. , Wang, Y. , Sun, H. , You, Y. , Piao, C. , et al (2020) Lactobacillus rhamnosus granules dose‐dependently balance intestinal microbiome disorders and ameliorate chronic alcohol‐induced liver injury. J Med Food 23: 114–124. [DOI] [PubMed] [Google Scholar]
- Harputluoglu, M.M. , Demirel, U. , Gul, M. , Temel, I. , Gursoy, S. , Selcuk, E.B. , et al (2012) Effects of rifaximin on bacterial translocation in thioacetamide‐induced liver injury in rats. Inflammation 35: 1512–1517. [DOI] [PubMed] [Google Scholar]
- Jaeschke, H. , and Hasegawa, T. (2006) Role of neutrophils in acute inflammatory liver injury. Liver Int 26: 912–919. [DOI] [PubMed] [Google Scholar]
- Kagan, J.C. , and Medzhitov, R. (2006) Phosphoinositide‐mediated adaptor recruitment controls Toll‐like receptor signaling. Cell 125: 943–955. [DOI] [PubMed] [Google Scholar]
- Knodell, R.G. , Ishak, K.G. , Black, W.C. , Chen, T.S. , Craig, R. , Kaplowitz, N. , et al (1981) Formulation and application of a numerical scoring system for assessing histological activity in asymptomatic chronic active hepatitis. Hepatology 1: 431–435. [DOI] [PubMed] [Google Scholar]
- Leibrock, J. , Lottspeich, F. , Hohn, A. , Hofer, M. , Hengerer, B. , Masiakowski, P. , et al (1989) Molecular cloning and expression of brain‐derived neurotrophic factor. Nature 341: 149–152. [DOI] [PubMed] [Google Scholar]
- Li, M. , Sun, X. , Zhao, J. , Xia, L. , Li, J. , Xu, M. , et al (2019) CCL5 deficiency promotes liver repair by improving inflammation resolution and liver regeneration through M2 macrophage polarization. Cell Mol Immunol 17: 753–764. 10.1038/s41423-019-0279-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Litvak, Y. , and Baumler, A.J. (2019) Microbiota‐nourishing immunity: a guide to understanding our microbial self. Immunity 51: 214–224. [DOI] [PubMed] [Google Scholar]
- Lv, L.X. , Hu, X.J. , Qian, G.R. , Zhang, H. , Lu, H.F. , Zheng, B.W. , et al (2014) Administration of Lactobacillus salivarius LI01 or Pediococcus pentosaceus LI05 improves acute liver injury induced by D‐galactosamine in rats. Appl Microbiol Biotechnol 98: 5619–5632. [DOI] [PubMed] [Google Scholar]
- Lv, L.X. , Yan, R. , Shi, H.Y. , Shi, D. , Fang, D.Q. , Jiang, H.Y. , et al (2017) Integrated transcriptomic and proteomic analysis of the bile stress response in probiotic Lactobacillus salivarius LI01. J Proteomics 150: 216–229. [DOI] [PubMed] [Google Scholar]
- McDole, J.R. , Wheeler, L.W. , McDonald, K.G. , Wang, B. , Konjufca, V. , Knoop, K.A. , et al (2012) Goblet cells deliver luminal antigen to CD103+ dendritic cells in the small intestine. Nature 483: 345–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizuno, M. , Yamada, K. , Takei, N. , Tran, M.H. , He, J. , Nakajima, A. , et al (2003) Phosphatidylinositol 3‐kinase: a molecule mediating BDNF‐dependent spatial memory formation. Mol Psychiatry 8: 217–224. [DOI] [PubMed] [Google Scholar]
- Nakamoto, N. , Amiya, T. , Aoki, R. , Taniki, N. , Koda, Y. , Miyamoto, K. , et al (2017) Commensal Lactobacillus controls immune tolerance during acute liver injury in mice. Cell Rep 21: 1215–1226. [DOI] [PubMed] [Google Scholar]
- Nicaise, C. , Prozzi, D. , Viaene, E. , Moreno, C. , Gustot, T. , Quertinmont, E. , et al (2008) Control of acute, chronic, and constitutive hyperammonemia by wild‐type and genetically engineered Lactobacillus plantarum in rodents. Hepatology 48: 1184–1192. [DOI] [PubMed] [Google Scholar]
- Norenberg, M.D. (1996) Astrocytic‐ammonia interactions in hepatic encephalopathy. Semin Liver Dis 16: 245–253. [DOI] [PubMed] [Google Scholar]
- O'Grady, J. (2012) Liver transplantation for acute liver failure. Best Pract Res Clin Gastroenterol 26: 27–33. [DOI] [PubMed] [Google Scholar]
- Patidar, K.R. , and Bajaj, J.S. (2015) Covert and overt hepatic encephalopathy: diagnosis and management. Clin Gastroenterol Hepatol 13: 2048–2061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van de Poll, M.C. , Ligthart‐Melis, G.C. , Olde Damink, S.W. , van Leeuwen, P.A. , Beets‐Tan, R.G. , Deutz, N.E. , et al (2008) The gut does not contribute to systemic ammonia release in humans without portosystemic shunting. Am J Physiol Gastrointest Liver Physiol 295: G760–G765. [DOI] [PubMed] [Google Scholar]
- Ramanan, D. , and Cadwell, K. (2016) Intrinsic defense mechanisms of the intestinal epithelium. Cell Host Microbe 19: 434–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryu, J.K. , Kim, S.J. , Rah, S.H. , Kang, J.I. , Jung, H.E. , Lee, D. , et al (2017) Reconstruction of LPS transfer cascade reveals structural determinants within LBP, CD14, and TLR4‐MD2 for efficient LPS recognition and transfer. Immunity 46: 38–50. [DOI] [PubMed] [Google Scholar]
- Shapiro, H. , Ashkenazi, M. , Weizman, N. , Shahmurov, M. , Aeed, H. , and Bruck, R. (2006) Curcumin ameliorates acute thioacetamide‐induced hepatotoxicity. J Gastroenterol Hepatol 21: 358–366. [DOI] [PubMed] [Google Scholar]
- Shen, T.C. , Albenberg, L. , Bittinger, K. , Chehoud, C. , Chen, Y.Y. , Judge, C.A. , et al (2015) Engineering the gut microbiota to treat hyperammonemia. J Clin Invest 125: 2841–2850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi, D. , Lv, L. , Fang, D. , Wu, W. , Hu, C. , Xu, L. , et al (2017) Administration of Lactobacillus salivarius LI01 or Pediococcus pentosaceus LI05 prevents CCl4‐induced liver cirrhosis by protecting the intestinal barrier in rats. Sci Rep 7: 6927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh, P. , Chung, H.J. , Lee, I.A. , D'Souza, R. , Kim, H.J. , and Hong, S.T. (2018) Elucidation of the anti‐hyperammonemic mechanism of Lactobacillus amylovorus JBD401 by comparative genomic analysis. BMC Genom 19: 292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soares, J.B. , Pimentel‐Nunes, P. , Roncon‐Albuquerque, R. , and Leite‐Moreira, A. (2010) The role of lipopolysaccharide/toll‐like receptor 4 signaling in chronic liver diseases. Hepatol Int 4: 659–672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sprenger, H. , Kaufmann, A. , Garn, H. , Lahme, B. , Gemsa, D. , and Gressner, A.M. (1997) Induction of neutrophil‐attracting chemokines in transforming rat hepatic stellate cells. Gastroenterology 113: 277–285. [DOI] [PubMed] [Google Scholar]
- Stravitz, R.T. , and Lee, W.M. (2019) Acute liver failure. Lancet 394: 869–881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weissenborn, K. , Ahl, B. , Fischer‐Wasels, D. , van den Hoff, J. , Hecker, H. , Burchert, W. , and Kostler, H. (2007) Correlations between magnetic resonance spectroscopy alterations and cerebral ammonia and glucose metabolism in cirrhotic patients with and without hepatic encephalopathy. Gut 56: 1736–1742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiest, R. , Albillos, A. , Trauner, M. , Bajaj, J.S. , and Jalan, R. (2017) Targeting the gut‐liver axis in liver disease. J Hepatol 67: 1084–1103. [DOI] [PubMed] [Google Scholar]
- Wlodzimirow, K.A. , Eslami, S. , Abu‐Hanna, A. , Nieuwoudt, M. , and Chamuleau, R.A. (2012) Systematic review: acute liver failure ‐ one disease, more than 40 definitions. Aliment Pharmacol Ther 35: 1245–1256. [DOI] [PubMed] [Google Scholar]
- Wu, W. , Lv, L. , Shi, D. , Ye, J. , Fang, D. , Guo, F. , et al (2017) Protective effect of akkermansia muciniphila against immune‐mediated liver injury in a mouse model. Front Microbiol 8: 1804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xing, S.C. , Huang, C.B. , Mi, J.D. , Wu, Y.B. , and Liao, X.D. (2019) Bacillus coagulans R11 maintained intestinal villus health and decreased intestinal injury in lead‐exposed mice by regulating the intestinal microbiota and influenced the function of faecal microRNAs. Environ Pollut 255: 113139. [DOI] [PubMed] [Google Scholar]
- Yang, R. , Zou, X. , Tenhunen, J. , and Tonnessen, T.I. (2017) HMGB1 and extracellular histones significantly contribute to systemic inflammation and multiple organ failure in acute liver failure. Mediators Inflamm 2017: 5928078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanny, B. , Winters, A. , Boutros, S. , and Saab, S. (2019) Hepatic encephalopathy challenges, burden, and diagnostic and therapeutic approach. Clin Liver Dis 23: 607–623. [DOI] [PubMed] [Google Scholar]
- Zhang, D. , Liu, H. , Wang, S. , Zhang, W. , Wang, J. , Tian, H. , et al (2019) Fecal microbiota and its correlation with fatty acids and free amino acids metabolism in piglets after a Lactobacillus strain oral administration. Front Microbiol 10: 785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, A. , Chen, J. , Wu, P. , Luo, M. , Zeng, Y. , Liu, Y. , et al (2017) Cationic polystyrene resolves nonalcoholic steatohepatitis, obesity, and metabolic disorders by promoting eubiosis of gut microbiota and decreasing endotoxemia. Diabetes 66: 2137–2143. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. The rarefaction curve reflected the sequencing depth of the faecal 16S rRNA sequencing analysis before TAA treatment.
Fig. S2. The rarefaction curve reflected the sequencing depth of the faecal 16S rRNA sequencing analysis after TAA treatment.
Table S1. Specific primers applied for RT‐PCR test.
Table S2. PERMANOVA test of community structure differences in the gut microbiota among groups after L. salivarius Li01 intervention.
Table S3. MRPP test of community structure differences in the gut microbiota among groups after L. salivarius Li01 intervention.
Table S4. PERMANOVA test of community structure differences in the gut microbiota among groups after TAA injection.
Table S5. MRPP test of community structure differences in the gut microbiota among groups after TAA injection.
Data Availability Statement
The 16S rRNA gene sequencing data were deposited in the sequence read archive (SRA) database (accession no. PRJNA636106).