

Abstract
Myocardial infarction (MI) is one of the leading causes of death globally. The aim of the present study was to find valuable microRNAs (miRNAs/miRs) and target mRNAs in order to contribute to our understanding of the pathology of MI. miRNA and mRNA data were downloaded for differential expression analysis. Then, a regulatory network between miRNAs and mRNAs was established, followed by function annotation of target mRNAs. Thirdly, prognosis and diagnostic analysis of differentially methylated target mRNAs were performed. Finally, an in vitro experiment was used to validate the expression of selected miRNAs and target mRNAs. A total of 19 differentially expressed miRNAs and 1,007 differentially expressed mRNAs were identified. Several regulatory interaction pairs between miRNA and mRNAs were identified, such as hsa-miR-142-2p-long-chain-fatty-acid-CoA ligase 1 (ACSL1), hsa-miR-15a-3p-nicotinamide phosphoribosyltransferase (NAMPT), hsa-miR-33b-5p-regulator of G-protein signaling 2 (RGS2), hsa-miR-17-3p-Jun dimerization protein 2 (JDP2), hsa-miR-24-1-5p-aquaporin-9 (AQP9) and hsa-miR-34a-5p-STAT1/AKT3. Of note, it was demonstrated that ACSL1, NAMPT, RGS2, JDP2, AQP9, STAT1 and AKT3 had diagnostic and prognostic values for patients with MI. In addition, STAT1 was involved in the ‘chemokine signaling pathway’ and ‘Jak-STAT signaling pathway’. AKT3 was involved in both the ‘MAPK signaling pathway’ and ‘T cell receptor signaling pathway’. Reverse transcription-quantitative PCR validation of hsa-miR-142-3p, hsa-miR-15a-3p, hsa-miR-33b-5p, ACSL1, NAMPT, RGS2 and JDP2 expression was consistent with the bioinformatics analysis. In conclusion, the identified miRNAs and mRNAs may be involved in the pathology of MI.
Keywords: myocardial infarction, differentially expressed microRNAs, differentially expressed mRNAs, diagnose, prognosis
Introduction
Myocardial infarction (MI) is the death of cardiomyocytes caused by obstruction of one or more arteries supplying the heart (1,2). Clinically, patients with MI often show a number of different symptoms including pain, fever and increased white blood cell count (3). The pivotal pathological characteristic after MI is cardiac fibrosis, which leads to cardiac remodeling and failure (4,5). In addition, there are various complications of MI, such as arrhythmia, pericarditis, heart rupture and papillary muscle rupture (6). MI pathology can also be complicated by cardiogenic shock, which can be life threatening (7). The recurrence rate of MI is high, and the time of recurrence is the highest within two years after remission. Moreover, each relapse can aggravate the disease (8).
The pathological mechanism of MI is complex. Frangogiannis et al (9) and Nian et al (10) found that MI was related to innate immunity programming, which is crucial in cardiac remodeling. Angiogenesis is also a key event for re-establishing the blood supply to the surviving myocardium following MI, which contributes to recovery of cardiac function (11). In addition, age, sex, smoking status, hypertension, atherosclerosis, dyslipidemia, diabetes mellitus and hypercholesterolemia are reported to be the primary risks of MI (12,13).
MicroRNAs (miRNAs/miRs) can negatively regulate target mRNA expression. Previous reports demonstrated that miRNAs could be crucial regulators in cardiovascular diseases, including hypertrophy, heart failure and ischemic cardiomyopathy (14–17). It is worth noting that some miRNAs, such as hsa-miR-1, hsa-miR-21, hsa-miR-206 and hsa-miR-499-5p, are significantly dysregulated in the hearts of patients with acute MI (18–22). In view of the important roles of miRNAs in MI, the aim of the present study was to find potential miRNAs and target mRNAs in MI, which may be useful in understanding the molecular mechanism of MI pathology. The present findings demonstrated that the epigenetic modifications of hsa-miR-142-2p-ACSL1, hsa-miR-15a-3p-NAMPT, hsa-miR-17-3p-JDP2, hsa-miR-33-5p-RGS2, hsa-miR- 24-1-5p-AQP9 and hsa-miR-34a-5p-STAT1/AKT3 and four signaling pathways (‘chemokine signaling pathway’, ‘Jak-STAT signaling pathway’, ‘MAPK signaling pathway’ and ‘T cell receptor signaling pathway’) may be associated with the development of MI. In addition, ACSL1, NAMPT, RGS2, JDP2, AQP9, STAT1 and AKT3 could help monitor the recurrence risk of patients with MI.
Materials and methods
Data retrieval
The miRNA and mRNA expression profiles were downloaded from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo). The key words of ‘Myocardial infarction’ and ‘Homo sapiens’ was used to search for related datasets. Screening requirements for the included datasets were as follows: i) The selected datasets must be genome-wide miRNA/mRNA transcriptome expression data; and ii) both standardized and original datasets were considered in this study. According to the aforementioned screening criteria, one miRNA dataset (GSE24548) and four mRNA datasets (GSE97320, GSE24519, GSE66360 and GSE60993) were selected for integrated analysis, which is listed in Table I.
Table I.
One miRNA dataset and four mRNA datasets used for integrated analysis.
| First author, year | GEO ID | Platform | Samples | Type | Omics | (Refs.) |
|---|---|---|---|---|---|---|
| Bellin et al, 2017 | GSE24548 | GPL 8227 | MI:Control, 4:3 | Blood | miRNA | Unpublished |
| Fan et al, 2017 | GSE97320 | GPL 570 | MI:Control, 3:3 | Blood | mRNA | (84) |
| Bellin et al, 2017 | GSE24519 | GPL 2895 | MI:Control, 34:4 | Blood | mRNA | Unpublished |
| Muse et al, 2015 | GSE66360 | GPL 570 | MI:Control, 49:50 | Blood | mRNA | (85) |
| Park et al, 2015 | GSE60993 | GPL6884 | MI:Control, 17:7 | Blood | mRNA | (86) |
MI, myocardial infarction; GEO, Gene Expression Omnibus; miRNA, microRNA.
Identification of differentially expressed miRNAs and mRNAs. Firstly, scale standardization was performed for all datasets. The limma (version 3.36.5) (23) and metaMA (version 3.1.2) (24) packages in R were then used to identify differentially expressed miRNAs and mRNAs. The inverse normal method was used to combine the P-value in metaMA. The false discovery rate (FDR) was performed for multiple testing corrections of raw P-values through the Benjamin and Hochberg method (25,26). The threshold of differentially expressed miRNAs and mRNAs was set as FDR<0.01 and FDR<0.05, respectively. The pheatmap R package (cran.r-project.org/web/packages/pheatmap/index.html) was used for hierarchical clustering analysis of the differentially expressed miRNAs and mRNAs (27).
Correlation analysis between differentially expressed miRNAs and mRNAs
The miRWalk3.0 (http://mirwalk.umm.uni-heidelberg.de/) software was used to predict the differentially expressed target mRNAs of differentially expressed miRNAs. The establishment of a miRNA-mRNA regulatory network was visualized using Cytoscape 3.7.1 software (28).
Functional analysis of differentially expressed mRNAs
Gene Ontology (GO) (29,30) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) (31) pathway enrichment via GeneCodis 3 (http://genecodis.cnb.csic.es) software were used to analyze the biological functions of the differentially expressed target mRNAs of differentially expressed miRNAs. The criterion for selecting significantly enriched functional terms of target differentially expressed mRNAs was P<0.05.
Screening of differentially expressed target mRNAs associated with MI prognosis
In the process of screening for datasets that contained miRNA and mRNA expression profiles, the GSE59867, GSE97320, GSE24519, GSE66360 and GSE60993 datasets were initially retrieved. As the GSE59867 dataset recorded the mRNA expression profile in the blood of patients at admission (111 cases), at discharge (101 cases), one month after discharge (95 cases) and six months after discharge (83 cases), this dataset was used for screening mRNAs associated with prognosis from identified targeted differentially expressed mRNAs, with the aim of finding reliable biomarkers to monitor the risk of recurrence in patients with MI.
Diagnostic analysis of differentially expressed target mRNAs associated with prognosis
In order to study whether identified differentially expressed target mRNAs associated with prognosis have diagnostic value, receiver operating characteristic (ROC) analysis was performed using pROC package in R language (32). The area under the curve (AUC) under binomial exact confidence interval was calculated and the ROC curve was generated.
In vitro validation of key miRNAs and target mRNAs
A total of five patients with MI and five normal controls were enrolled from October to November 2019 at Jinan Jigang Hospital. The inclusion criteria of patients with MI was as follows: i) Time of chest pain or distress was >30 min within 24 h, and the levels of creatine kinase-MB and troponin T were higher than the normal range; ii) no history of MI; iii) no drug treatment or surgical treatment before admission; iv) patients had to provide blood samples before hospitalization, at discharge and 6 months after MI; and v) patients had to provide complete clinical data, including sex, age, height and weight. The exclusion criteria was as follows: i) Patients with myocarditis and other diseases caused by chest pain or distress; ii) patients had a history of renal failure, advanced liver disease, malignant tumor and other inflammatory diseases, such as psoriasis and rheumatoid arthritis; iii) patients with recurring MI; iv) patients with incomplete clinical data; and v) patients who did not provide blood samples before hospitalization, at discharge and 6 months after MI. Ethical approval was obtained from the Ethics Committee of Jinan Jigang Hospital (approval no. JG-001) and informed written consent was obtained from all individuals.
Total RNA from the blood samples was extracted using the RNAliquid reagent (Tiangen Biotech Co., Ltd.), according to the manufacturer's protocols. RNA (1 µg) was applied to synthesize cDNA for 3 min at 42°C, followed by 15 min at 42°C and 3 min at 95°C using FastQuant RT kit (Tiangen Biotech Co., Ltd.). Then, reverse transcription-quantitative PCR (RT-qPCR) was performed in an ABI 7300 Real-time PCR system with SYBR-Green PCR Master Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.). The thermocycling conditions consisted of: i) Initial denaturation of 15 min at 95°C; iii) 40 cycles of 10 sec at 95°C, 30 sec at 55°C, 32 sec at 72°C; and iii) 15 sec at 95°C, 60 sec at 60°C, then 15 sec at 95°C. All reactions were carried out in triplicate β-actin and GAPDH were used as the internal reference genes. Relative gene expression was analyzed by the fold-change method (33). The sequences of reverse and forward primers for all of the genes analyzed were as follows: i) ACSL1-forward, 5′-TGTGGGACCGGCTGATCTT-3′; ii) ACSL1-reverse, 5′-AGTGCACTCTGTCTGTCCGT-3′; iii) NAMPT-forward, 5′-TCTTCTGAGGCAGCGGTTG-3′; iv) NAMPT-reverse, 5′-GGCTGGAGTAGTGGGAAAGC-3′; v) RGS2-forward, 5′-AAGATTGGAAGACCCGTTTGAG-3′; vi) RGS2-reverse, 5′-GCAAGACCATATTTGCTGGCT-3′; vii) JDP2-forward, 5′-AGACCCAGATTGAGGAGCTG-3′; JDP2-reverse, 5′-AGTGGGTTGCCTTCTGACCTC-3′; viii) hsa-miR-142-3p-forward, 5′-TGTAGTGTTTCCTACTTTATGGA-3; ix) hsa-miR-15a-3p-forward, 5′-CAGGCCATATTGTGCTGCCTCA-3; x) hsa-miR-33b-3p-forward, 5′-GTGCATTGCTGTTGCATTGC-3; xi) GAPDH-forward, 5′-GGAGCGAGATCCCTCCAAAAT-3′; xii) GAPDH-reverse, 5′-GGCTGTTGTCATACTTCTCATGG-3′; xiii) ACTB-forward, 5′-CATGTACGTTGCTATCCAGGC-3′; and vix) ACTB-reverse, 5′-CTCCTTAATGTCACGCACGAT-3′. The sequences of the reverse primers for hsa-miR-142-3p, hsa-miR-15a-3p and hsa-miR-33b-3p were not disclosed by the manufacturer.
Statistical analysis
All statistical analyses were performed using GraphPad Prism (version 2.0; GraphPad Software, Inc.). For the result described in Figs. 7 and 9, one-way ANOVA, followed by Bonferroni correction, were used to assess statistical significance among different groups. Results are presented as the mean ± SD. All experiments were repeated independently at least three times. P<0.05 was considered to indicate a statistically significant difference.
Figure 7.

Box plots of ACSL1, NAMPT, RGS2, AQP9, JDP2, STAT1 and AKT3 expression. The expression levels of ACSL1, NAMPT, RGS2, AQP9, JDP2, STAT1 and AKT3 were determined in the blood of patients with MI at admission (n=111), at discharge (n=101), 1 month after discharge (n=95) and 6 months after discharge (n=83). ACSL1, long-chain-fatty-acid-CoA ligase 1; NAMPT, nicotinamide phosphoribosyltransferase; RGS2, regulator of G-protein signaling 2; JDP2, Jun dimerization protein 2; AQP9, aquaporin-9; MI, myocardial infarction.
Figure 9.

In vitro reverse transcription-quantitative PCR validation of differentially expressed miRNAs and their target mRNAs. One-way ANOVA, followed by Bonferroni correction, were used to assess statistical significance among different groups. Data are presented as the mean ± SD of the fold-change with respect to the control. Fold-change >1 and fold-change <1 represent upregulation and downregulation, respectively. n=5. miRNA/miR, microRNA; ACSL1, long-chain-fatty-acid-CoA ligase 1; NAMPT, nicotinamide phosphoribosyltransferase; RGS2, regulator of G-protein signaling 2; JDP2, Jun dimerization protein 2.
Results
Expression pattern of miRNA and mRNA
A total of 19 differentially expressed (one upregulated and 18 downregulated) miRNAs and 1,007 differentially expressed (287 upregulated and 720 downregulated) mRNAs were identified in MI. All differentially expressed miRNAs and the top 20 differentially expressed mRNAs are shown in Tables II and III, respectively. The heat map of all differentially expressed miRNAs and the top 100 differentially expressed mRNAs are shown in Figs. 1 and 2, respectively.
Table II.
Differentially expressed miRNAs in MI.
| Symbol | LogFC | Average expression | P-value | FDR |
|---|---|---|---|---|
| hsa-miR-32-5p | −1.72 | 4.66 | 3.17×10−05 | 4.93×10−03 |
| hsa-miR-545-3p | −1.50 | 4.37 | 3.76×10−05 | 4.93×10−03 |
| hsa-miR-219a-5p | −1.34 | 3.30 | 5.23×10−05 | 4.93×10−03 |
| hsa-miR-29b-3p | −1.46 | 7.45 | 1.04×10−04 | 6.01×10−03 |
| hsa-miR-548a-5p | −1.05 | 2.59 | 1.11×10−04 | 6.04×10−03 |
| hsa-miR-17-3p | −1.04 | 6.45 | 2.03×10−04 | 6.04×10−03 |
| hsa-miR-33a-5p | −1.34 | 6.14 | 2.05×10−04 | 6.04×10−03 |
| hsa-miR-33b-5p | −1.17 | 2.70 | 2.14×10−04 | 6.04×10−03 |
| hsa-miR-186-3p | −0.90 | 2.14 | 2.32×10−04 | 6.04×10−03 |
| hsa-miR-101-3p | −1.19 | 7.41 | 2.46×10−04 | 6.04×10−03 |
| hsa-miR-142-3p | −1.08 | 1.05 | 2.57×10−04 | 6.04×10−03 |
| hsa-miR-598-3p | −1.14 | 4.69 | 2.84×10−04 | 6.04×10−03 |
| hsa-miR-362-3p | −1.51 | 3.30 | 2.91×10−04 | 6.04×10−03 |
| hsa-miR-590-5p | −1.20 | 7.49 | 3.10×10−04 | 6.04×10−03 |
| hsa-miR-24-1-5p | −1.07 | 5.30 | 3.20×10−04 | 6.04×10−03 |
| hsa-miR-34a-5p | 1.10 | 4.99 | 5.55×10−04 | 9.75×10−03 |
| hsa-miR-15a-3p | −0.84 | 2.89 | 5.86×10−04 | 9.75×10−03 |
| hsa-miR-301a-3p | −0.91 | 7.82 | 6.47×10−04 | 9.90×10−03 |
| hsa-miR-19a-3p | −1.06 | 8.77 | 6.65×10−04 | 9.90×10−03 |
LogFC>1 and LogFC<1 represents upregulation and downregulation, respectively. LogFC, log fold-change; FDR, false discovery rate; miRNA, microRNA; MI, myocardial infarction.
Table III.
Top 20 differentially expressed mRNAs in MI.
| Symbol | Combined ES | P-value | FDR | Up/down |
|---|---|---|---|---|
| ACSL1 | 1.59 | <4.44×10−15 | <6.76×10−12 | Up |
| S100A12 | 1.44 | <4.44×10−15 | <6.76×10−12 | Up |
| IL1R2 | 1.27 | 4.44×10−15 | 6.76×10−12 | Up |
| NAMPT | 1.24 | 1.22×10−14 | 1.69×10−11 | Up |
| RGS2 | 1.60 | 2.51×10−14 | 3.18×10−11 | Up |
| AQP9 | 1.19 | 3.22×10−14 | 3.77×10−11 | Up |
| JDP2 | 1.20 | 4.13×10−14 | 4.49×10−11 | Up |
| DUSP1 | 1.24 | 2.57×10−13 | 2.17×10−10 | Up |
| HAL | 1.12 | 5.64×10−13 | 4.30×10−10 | Up |
| S100A8 | 1.88 | 6.36×10−13 | 4.61×10−10 | Up |
| PLA2G12A | −1.34 | 2.09×10−13 | 1.87×10−10 | Down |
| GIMAP6 | −1.08 | 1.55×10−12 | 9.43×10−10 | Down |
| PAQR8 | −1.32 | 1.85×10−12 | 1.08×10−09 | Down |
| CHST12 | −1.17 | 1.67×10−11 | 7.83×10−09 | Down |
| GZMA | −1.03 | 3.45×10−11 | 1.31×10−08 | Down |
| RNASEH2B | −1.20 | 4.46×10−11 | 1.61×10−08 | Down |
| DENND2D | −1.03 | 4.95×10−11 | 1.71×10−08 | Down |
| RPAP2 | −1.12 | 8.16×10−11 | 2.53×10−08 | Down |
| GIMAP7 | −1.00 | 1.33×10−10 | 3.76×10−08 | Down |
| TRIM68 | −1.04 | 2.13×10−10 | 5.68×10−08 | Down |
ES, effect size; FDR, false discovery rate; MI, myocardial infarction.
Figure 1.
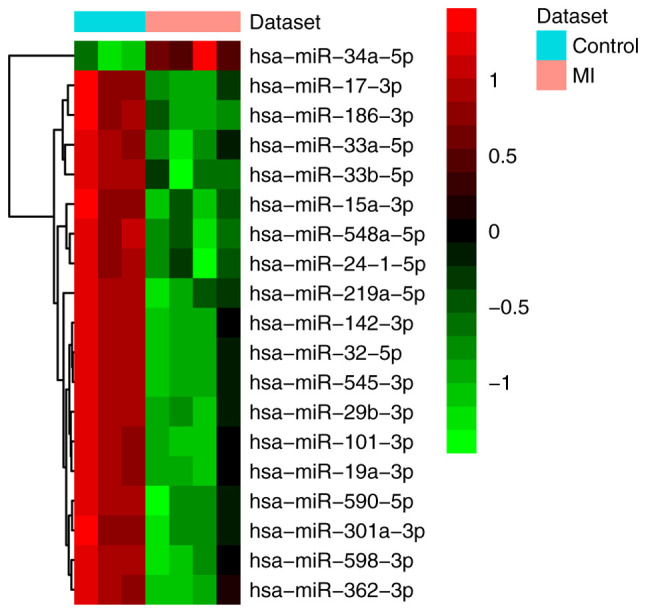
Heat map of all differentially expressed miRNAs in MI. The differentially expressed miRNA clustering tree is shown on the right. Red, below the reference channel; green, higher than the reference. miRNA/miR, microRNA; MI, myocardial infarction.
Figure 2.

Heat map of top 100 differentially expressed mRNAs in MI. The differentially expressed mRNA clustering tree is shown on the right. Red, below the reference channel; green, higher than the reference. MI, myocardial infarction.
Associations between miRNAs and mRNAs
Depending on the target analysis, 2,137 miRNA-mRNA pairs, including 19 miRNAs and 481 mRNAs, were identified. The regulatory network of miRNA-mRNA is shown in Fig. 3. In addition, the regulatory subnetworks of miRNAs-mRNAs between hsa-miR-142-3p, hsa-miR-15a-3p, hsa-miR-17-3p, hsa-miR-33b-5p, hsa-miR-24-1-5p, hsa-miR-34a-5p and their target mRNAs are shown in Fig. 4.
Figure 3.

Regulatory network of miRNA-mRNAs between 19 differentially expressed miRNAs and 481 differentially expressed mRNAs in myocardial infarction. The rhombuses and circles represent the differentially expressed miRNAs and the differentially expressed target mRNAs, respectively. The red and blue color represent upregulation and downregulation, respectively. The black borders represent the top 10 up- and downregulated mRNAs. miRNA/miR, microRNA.
Figure 4.

Regulatory subnetworks of miRNAs-mRNAs between hsa-miR-142-3p, hsa-miR-15a-3p, hsa-miR-17-3p, hsa-miR-33b-5p, hsa-miR-24-1-5p, hsa-miR-34a-5p and their target mRNAs in myocardial infarction. The triangles and circles represent the differentially expressed miRNAs and differentially expressed target mRNAs, respectively. The red and green color represent upregulation and downregulation, respectively. The black borders represents the top 10 up- and downregulated mRNAs. miRNA/miR, microRNA.
Functional analysis of differentially expressed target mRNAs
In order to understand the biological function of 481 differentially expressed target mRNAs of 19 differentially expressed miRNAs, GO and KEGG pathway analysis were performed. The top 5 significantly enriched GO terms and top 12 KEGG terms are shown in Figs. 5 and 6, respectively. The biological process, cell component and molecular function ‘regulation of transcription, DNA dependent’, ‘nucleus’ and protein binding’ were significantly enriched, respectively (Fig. 5). For KEGG terms, it was found that STAT1 was involved in the ‘chemokine signaling pathway’ and ‘Jak-STAT signaling pathway’. In addition, AKT3 was involved in the ‘MAPK signaling pathway’ and ‘T cell receptor signaling pathway’ (Fig. 6).
Figure 5.

Top 5 significant enrichment GO terms of the differentially expressed target mRNAs in myocardial infarction. The z-score clustering in the GO terms of the differentially expressed target mRNAs is shown at the bottom. The red and blue color represent upregulated and downregulated mRNAs, respectively. BP, biological process; CC, cellular component; MF, molecular function; GO, Gene Ontology.
Figure 6.

Top 12 Kyoto Encyclopedia of Genes and Genomes signaling pathways of the differentially expressed target mRNAs in myocardial infarction.
Identification of differentially expressed target mRNAs associated with prognosis
Through comparative analysis between mRNAs in the GSE59867 dataset and identified 481 differentially expressed target mRNAs, it was found that 185 differentially expressed target mRNAs, including long-chain-fatty-acid-CoA ligase 1 (ACSL1), nicotinamide phosphoribosyltransferase (NAMPT), regulator of G-protein signaling 2 (RGS2), Jun dimerization protein 2 (JDP2), aquaporin-9 (AQP9), STAT1 and AKT3, were associated with prognosis. The expression levels of ACSL1, NAMPT, RGS2, AQP9, JDP2, STAT1 and AKT3 were measured in the blood of patients with MI at admission, discharge, one month after discharge and six months after discharge (Fig. 7). The results showed that the expression levels of ACSL1 (P=2.2×10−11; P<2.22×10−16; P<2.22×10−16), NAMPT (P=1.1×10−07; P=2.5×10−10; P=5.6×10−11), RGS2 (P=3.8×10−05; P=1.6×10−07; P=1.3×10−05), AQP9 (P=9×10−12; P<2.22×10−16; P<2.22×10−16) and JDP2 (P=0.00031; P=1.3×10−08; P=1.3×10−07) in MI were significantly downregulated at discharge, one month after discharge and six months after discharge compared with that at admission. Whereas, it was found that the expression levels of STAT1 (P=8.1×10−11; P=2.1×10−13; P=7×10−10) and AKT3 (P=0.0036; P=5.6×10−06; P=1×10−09) in MI were significantly upregulated at discharge, one month after discharge and six months after discharge compared with that at admission. This suggested that ACSL1, NAMPT, RGS2, AQP9, JDP2, STAT1 and AKT3 expression could be used to monitor the recurrence risk of patients with MI.
Diagnosis prediction of differentially expressed target mRNAs associated with prognosis
ROC curve analysis was performed to assess the diagnostic ability of ACSL1, NAMPT, RGS2, AQP9, JDP2, STAT1 and AKT3 (Fig. 8). Of note, AUC values of the aforementioned differentially expressed target mRNAs were all >0.6, suggesting that they have a potential diagnostic value for MI.
Figure 8.

ROC curves of ACSL1, NAMPT, RGS2, AQP9, JDP2, STAT1 and AKT3 expression in myocardial infarction. The ROC curves were used to show the diagnostic value of these genes with 1-specificity and sensitivity. ACSL1, long-chain-fatty-acid-CoA ligase 1; NAMPT, nicotinamide phosphoribosyltransferase; RGS2, regulator of G-protein signaling 2; JDP2, Jun dimerization protein 2; AQP9, aquaporin-9; ROC, receiver operating characteristic; AUC, area under curve.
RT-qPCR
A total of five patients with MI and five normal controls were included in the present study. Clinical information of these individuals is presented in Table IV. In addition, three differentially expressed miRNAs (hsa-miR-142-3p, hsa-miR-15a-3p and hsa-miR-33b-5p) and their target mRNAs (ACSL1, NAMPT, RGS2 and JDP2) were randomly selected for validation via RT-qPCR (Fig. 9). The relative expression levels of hsa-miR-142-3p, hsa-miR-15a-3p and hsa-miR-33b-5p were downregulated in the blood of patients with MI. The relative expression levels of ACSL1, NAMPT, RGS2 and JDP2 were upregulated in the blood of patients with MI. These results were consistent with the bioinformatics analysis.
Table IV.
Clinical information of patients with MI and healthy controls.
| A, Patients with MI | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sex | Age, years | Height, m | Weight, kg | Hypertension history | Diabetes history | Smoking history | Drinking history | LDL | HDL | TC | TG | CTnT, ng/ml | CK-MB, µg/l |
| Female | 75 | 1.56 | 50 | Yes | No | No | No | 3.34 | 1.20 | 5.20 | 1.46 | 1.52 | 87 |
| Male | 84 | 1.70 | 70 | Yes | No | No | No | 3.05 | 1.17 | 4.56 | 0.75 | 0.07 | 46 |
| Male | 31 | 1.73 | 75 | Yes | Yes | No | Yes | 4.04 | 1.12 | 6.12 | 0.86 | 1.35 | 82 |
| Female | 65 | 1.61 | 61 | Yes | No | Yes | No | 3.51 | 1.31 | 5.69 | 1.24 | 1.21 | 49 |
| Male | 56 | 1.58 | 54 | Yes | Yes | Yes | No | 3.45 | 1.07 | 6.31 | 1.36 | 0.07 | 65 |
| B, Healthy controls | |||||||||||||
| Female | 83 | 1.55 | 80 | Yes | No | No | No | 1.26 | 1.03 | 3.12 | 1.83 | <0.02 | <1l |
| Male | 82 | 170 | 81 | Yes | Yes | No | Yes | 2.74 | 1.01 | 4.40 | 1.42 | 0.33 | 2.48 |
| Female | 67 | 1.58 | 60 | No | No | No | No | 2.20 | 0.61 | 3.42 | 1.35 | 0.01 | 1.00 |
| Male | 59 | 1.75 | 85 | Yes | No | Yes | No | 3.45 | 1.14 | 5.21 | 1.89 | 0.01 | 2.23 |
| Male | 41 | 1.73 | 68 | No | No | No | No | 3.89 | 1.23 | 6.01 | 0.75 | 0.02 | 2.45 |
MI, myocardial infarction; HC, healthy controls; LDL, low density lipoprotein; HDL, high density lipoprotein; TC, total cholesterol; TG, triglycerides; CTnT, troponin T; CK-MB, creatine kinase-MB.
Discussion
Decreased expression of hsa-miR-142-3p was found in patients with heart failure (34). Su et al (35) demonstrated that upregulating hsa-miR-142-3p could ameliorate myocardial ischemic injury. Of note, hsa-miR-142-3p is considered to be a biomarker for acute MI (36). In the present study, it was found that hsa-miR-142-3p was downregulated in the blood of patients with MI. Additionally, ACSL1 was found to be one of the target mRNAs of hsa-miR-142-3p and the most upregulated gene. Moreover, ACSL1 was shown to have a diagnostic and prognostic value for patients with MI. ACSL1 catalyzes the synthesis of acyl-CoA from fatty acid synthase (37). Overexpression of ACSL1 can lead to severe cardiomyopathy (38). The expression of ACSL1 is significantly related to the degree of atherosclerosis in the coronary artery (39). It has been observed that ACSL1 is upregulated in peripheral blood leukocytes of patients with acute MI (39). The present study further demonstrated the important roles of hsa-miR-142-3p and ACSL1 in MI.
Mature hsa-miR-15a-3p influences vascular function, including the expression of vascular endothelial growth factor receptor 2 (40–43). hsa-miR-15a-3p is notably downregulated in the atrial myocardial tissue of patients with congenital heart defect (44). The present study found decreased expression of hsa-miR-15a-3p in the blood of patients with MI; NAMPT was shown to be regulated by hsa-miR-15a-3p. Furthermore, NAMPT could be regarded as a diagnostic and prognostic biomarker for patients with MI. NAMPT is involved in atherosclerosis and myocardial failure (45). Increased expression of NAMPT has been found in MI (46). In addition, FK866 (a specific inhibitor of NAMPT) reduced infarct size following MI (47). The present results suggested that hsa-miR-15a-3p may play a critical role in the process of MI by targeting NAMPT.
Upregulation of hsa-miR-17-3p was demonstrated to promote the viability of cardiomyocytes (48). Overexpression of hsa-miR-17-3p can inhibit fibrosis of cardiac fibroblasts (49). In addition, hsa-miR-17-3p is related to atherosclerosis and angiogenesis (50–52). In the present study, it was found that hsa-miR-17-3p was downregulated in the blood of patients with MI. In addition, JDP2 was shown to be the target of hsa-miR-17-3p. It is worth mentioning that JDP2 was associated with diagnosis and prognosis. The expression of JDP2 was revealed to be increased in patients with heart failure and MI (46,53). Of note, JDP2 is considered to be a candidate marker for the risk stratification of patients who may develop heart failure following MI (54). The present results indicated that JDP2 may be involved in the development of MI under the regulation of hsa-miR-17-3p.
hsa-miR-33b-5p is a hypoxia-regulated miRNA. The methylation level of the encoding region of hsa-miR-33b-5p is related to lipid levels (55). The present study found that hsa-miR-33b-5p was downregulated in the blood of patients with MI. Moreover, upregulated RGS2 with diagnostic and prognostic value was shown to be regulated by hsa-miR-33b-5p. RGS2 is involved in regulating vascular smooth muscle cell contraction and blood pressure (56,57). Dysfunction of RGS2 is associated with cardiac hypertrophy (58). The present results suggested that hsa-miR-33b-5p could be associated with MI pathology by targeting RGS2.
A previous report found that hsa-miR-24-1-5p reduced cardiac fibrosis following MI (59). In the current study, decreased expression of hsa-miR-24-1-5p was found in the blood of patients with MI. In addition, upregulated AQP9 was one of the targets of hsa-miR-24-1-5p. Moreover, AQP9 had a diagnostic and prognostic value for patients with MI. It has been reported that increased AQP9 is associated with the development of atherosclerotic lesions (60). It has been demonstrated that AQP9 is notably upregulated in patients with heart failure and MI (46,61). In addition, silencing of the AQP9 gene was revealed to inhibit apoptosis of myocardial cells and improve cardiac function following MI (62). Thus, it has been demonstrated that hsa-miR-24-1-5p and AQP9 may play a significant role in cardiac function following MI.
has-miR-34a-5p, which is associated with cardiac fibrosis and regeneration, is upregulated during atherosclerosis progression (63,64). Increased expression of hsa-miR-34a-5p in circulation is related to cardiac remodeling and heart failure following MI (65–67). In the present study, it was found that hsa-miR-34a-5p was upregulated in the blood of patients with MI. In addition, downregulation of STAT1 and AKT3 was shown to be regulated by hsa-miR-34a-5p. Furthermore, STAT1 and AKT3 were associated with the diagnosis and prognosis of patients with MI. Activation of STAT1 has been demonstrated to induce cardiomyocyte apoptosis following ischemia/reperfusion injury by enhancing the expression of pro-apoptotic proteins, which suggests that STAT1 plays a key role in the pathology of MI (68–70). AKT3 is involved in cardiomyocyte cell death (71). In rats, the expression of AKT3 is decreased in the spinal cord following myocardial ischemia-reperfusion injury (72). In addition, AKT3 deficiency is related to an increase in atherosclerosis (73).
A side from being a target of has-miR-34a-5p, it was also found that STAT1 was involved in the ‘chemokine signaling pathway’ and ‘Jak-STAT signaling pathway’ according to KEGG. AKT3 was found to be involved in the ‘MAPK signaling pathway’ and ‘T cell receptor signaling pathway’ according to KEGG. Previous studies revealed that chemokines were expressed in the infarcted myocardium and play an important role in myocardial inflammation and healing (74,75). The Jak/STAT signaling pathway plays a key role in the modulation of cardioprotection against ischemia/reperfusion injury (76,77). Activation of p38 MAPK has been observed in cardiac hypertrophy induced by MI, and inhibition of the signing pathway protects the heart against MI (78). In addition, the p38-JNK-MAPK signaling pathway is associated with the inflammatory response in MI (79–81). T cells, the main components of cell-mediated inflammation, are associated with the development of atherosclerotic plaques (82). Notably, Abbate et al (83) detected activated T cells in the infarct and remote areas of the heart tissue of patients with MI.
In conclusion, the bioinformatics analysis from the GEO database led to the finding of several potential differentially expressed miRNAs and mRNAs. The epigenetic modifications of hsa-miR-142-2p-ACSL1, hsa-miR-15a-3p-NAMPT, hsa-miR-17-3p-JDP2, hsa-miR-33-5p-RGS2, hsa-miR- 24-1-5p-AQP9 and hsa-miR-34a-5p-STAT1/AKT3 and four signaling pathways (‘chemokine signaling pathway’, ‘Jak-STAT signaling pathway’, ‘MAPK signaling pathway’ and ‘T cell receptor signaling pathway’) may be associated with the development of MI. In addition, ACSL1, NAMPT, RGS2, JDP2, AQP9, STAT1 and AKT3 could monitor the recurrence risk of patients with MI. However, there is a limitation of the present study. The underlying mechanism of the identified differentially expressed miRNAs and mRNAs in MI requires further study in vivo and in vitro.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets generated and/or analyzed during the current study are available in the GEO repository under accession nos. GSE24548 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE24548), GSE97320 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE97320), GSE24519 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE24519), GSE66360 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE66360) and GSE60993 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE60993).
Authors' contributions
SW drafted the manuscript. SW and NC carried out the experiments and analyzed and interpreted the data. SW designed the study. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Ethics approval was obtained from the Ethics Committee of Jinan Jigang Hospital (approval no. JG-001) and informed written consent was obtained from all individuals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Bainey KR, Fresco C, Zheng Y, Halvorsen S, Carvalho A, Ostojic M, Goldstein P, Gershlick AH, Westerhout CM, Van de Werf F, et al. Implications of ischaemic area at risk and mode of reperfusion in ST-elevation myocardial infarction. Heart. 2016;102:527–533. doi: 10.1136/heartjnl-2015-308075. [DOI] [PubMed] [Google Scholar]
- 2.Goto Y, Masaki I, Yamada A, Uno M, Nakamori S, Nagata M, Ichikawa Y, Kitagawa K, Ito M, Sakuma H. Native T1 mapping allows for the accurate detection of the segments with chronic myocardial infarction in patients with known or suspected coronary artery disease. J Cardiovasc Magn Reson. 2016;18:P70. doi: 10.1186/1532-429X-18-S1-P70. [DOI] [Google Scholar]
- 3.Orlic D, Ostojic M, Beleslin B, Milasinovic D, Tesic M, Borovic M, Vukcevic V, Stojkovic S, Nedeljkovic M, Stankovic G. The randomized physiologic assessment of thrombus aspiration in patients with acute ST-segment elevation myocardial infarction trial (PATA STEMI): Study rationale and design. J Interv Cardiol. 2014;27:341–347. doi: 10.1111/joic.12105. [DOI] [PubMed] [Google Scholar]
- 4.Furtado MB, Costa MW, Rosenthal NA. The cardiac fibroblast: Origin, identity and role in homeostasis and disease. Differentiation. 2016;92:93–101. doi: 10.1016/j.diff.2016.06.004. [DOI] [PubMed] [Google Scholar]
- 5.Kurose H, Mangmool S. Myofibroblasts and inflammatory cells as players of cardiac fibrosis. Arch Pharm Res. 2016;39:1100–1113. doi: 10.1007/s12272-016-0809-6. [DOI] [PubMed] [Google Scholar]
- 6.Gao M, Yin D, Chen J, Qu X. Activating the interleukin-6-Gp130-STAT3 pathway ameliorates ventricular electrical stability in myocardial infarction rats by modulating neurotransmitters in the paraventricular nucleus. BMC Cardiovasc Disord. 2020;20:60. doi: 10.1186/s12872-020-01363-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ouweneel DM, Eriksen E, Sjauw KD, van Dongen IM, Hirsch A, Packer EJ, Vis MM, Wykrzykowska JJ, Koch KT, Baan J, et al. Percutaneous mechanical circulatory support versus intra-aortic balloon pump in cardiogenic shock after acute myocardial infarction. J Am Coll Cardiol. 2017;69:278–287. doi: 10.1016/j.jacc.2016.10.022. [DOI] [PubMed] [Google Scholar]
- 8.Rodríguez-Jiménez AE, Cruz-Inerarity H, Negrín-Valdés T, Fardales-Rodríguez R, Chávez-González E. Corrected QT-Interval Dispersion: An Electrocardiographic Tool to Predict Recurrence of Myocardial Infarction. MEDICC Rev. 2019;21:22–25. doi: 10.37757/MR2019.V21.N2-3.6. [DOI] [PubMed] [Google Scholar]
- 9.Frangogiannis NG, Smith CW, Entman ML. The inflammatory response in myocardial infarction. Cardiovasc Res. 2002;53:31–47. doi: 10.1016/S0008-6363(01)00434-5. [DOI] [PubMed] [Google Scholar]
- 10.Nian M, Lee P, Khaper N, Liu P. Inflammatory cytokines and postmyocardial infarction remodeling. Circ Res. 2004;94:1543–1553. doi: 10.1161/01.RES.0000130526.20854.fa. [DOI] [PubMed] [Google Scholar]
- 11.Landmesser U, Wollert KC, Drexler H. Potential novel pharmacological therapies for myocardial remodelling. Cardiovasc Res. 2009;81:519–527. doi: 10.1093/cvr/cvn317. [DOI] [PubMed] [Google Scholar]
- 12.Yamada Y, Ichihara S, Nishida T. Molecular genetics of myocardial infarction. Genomic Med. 2008;2:7–22. doi: 10.1007/s11568-008-9025-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Swirski FK, Nahrendorf M. Leukocyte behavior in atherosclerosis, myocardial infarction, and heart failure. Science. 2013;339:161–166. doi: 10.1126/science.1230719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cheng Y, Zhang C. MicroRNA-21 in cardiovascular disease. J Cardiovasc Transl Res. 2010;3:251–255. doi: 10.1007/s12265-010-9169-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Marques FZ, Vizi D, Khammy O, Mariani JA, Kaye DM. The transcardiac gradient of cardio-microRNAs in the failing heart. Eur J Heart Fail. 2016;18:1000–1008. doi: 10.1002/ejhf.517. [DOI] [PubMed] [Google Scholar]
- 16.Oliveira-Carvalho V, da Silva MM, Guimarães GV, Bacal F, Bocchi EA. MicroRNAs: new players in heart failure. Mol Biol Rep. 2013;40:2663–2670. doi: 10.1007/s11033-012-2352-y. [DOI] [PubMed] [Google Scholar]
- 17.Huang JB, Mei J, Jiang LY, Jiang ZL, Liu H, Zhang JW, Ding FB. miR-196a2 rs11614913 T>C polymorphism is associated with an increased risk of tetralogy of fallot in a Chinese population. Acta Cardiol Sin. 2015;31:18–23. doi: 10.6515/ACS20140310B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Meder B, Keller A, Vogel B, Haas J, Sedaghat-Hamedani F, Kayvanpour E, Just S, Borries A, Rudloff J, Leidinger P, et al. MicroRNA signatures in total peripheral blood as novel biomarkers for acute myocardial infarction. Basic Res Cardiol. 2011;106:13–23. doi: 10.1007/s00395-010-0123-2. [DOI] [PubMed] [Google Scholar]
- 19.Bostjancic E, Zidar N, Glavac D. MicroRNA microarray expression profiling in human myocardial infarction. Dis Markers. 2009;27:255–268. doi: 10.1155/2009/641082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liang H, Zhang C, Ban T, Liu Y, Mei L, Piao X, Zhao D, Lu Y, Chu W, Yang B. A novel reciprocal loop between microRNA-21 and TGFβRIII is involved in cardiac fibrosis. Int J Biochem Cell Biol. 2012;44:2152–2160. doi: 10.1016/j.biocel.2012.08.019. [DOI] [PubMed] [Google Scholar]
- 21.Shan ZX, Lin QX, Fu YH, Deng CY, Zhou ZL, Zhu JN, Liu XY, Zhang YY, Li Y, Lin SG, Yu XY. Upregulated expression of miR-1/miR-206 in a rat model of myocardial infarction. Biochem Biophys Res Commun. 2009;381:597–601. doi: 10.1016/j.bbrc.2009.02.097. [DOI] [PubMed] [Google Scholar]
- 22.Shi B, Guo Y, Wang J, Gao W. Altered expression of microRNAs in the myocardium of rats with acute myocardial infarction. BMC Cardiovasc Disord. 2010;10:11. doi: 10.1186/1471-2261-10-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, Smyth GK. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015;43:e47. doi: 10.1093/nar/gkv007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Marot G, Foulley JL, Mayer CD, Jaffrézic F. Moderated effect size and P-value combinations for microarray meta-analyses. Bioinformatics. 2009;25:2692–2699. doi: 10.1093/bioinformatics/btp444. [DOI] [PubMed] [Google Scholar]
- 25.Reiner-Benaim A. FDR control by the BH procedure for two-sided correlated tests with implications to gene expression data analysis. Biom J. 2007;49:107–126. doi: 10.1002/bimj.200510313. [DOI] [PubMed] [Google Scholar]
- 26.Benjamini Y, Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol. 1995;57:289–300. [Google Scholar]
- 27.Wu Y, Zhang L, Zhang Y, Zhen Y, Liu S. Bioinformatics analysis to screen for critical genes between survived and non-survived patients with sepsis. Mol Med Rep. 2018;18:3737–3743. doi: 10.3892/mmr.2018.9408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.The Gene Ontology Consortium, corp-author. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019;47:D330–D338. doi: 10.1093/nar/gky1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28:27–30. doi: 10.1093/nar/28.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Robin X, Turck N, Hainard A, Tiberti N, Lisacek F, Sanchez JC, Müller M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinformatics. 2011;12:77. doi: 10.1186/1471-2105-12-77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 34.Ellis KL, Cameron VA, Troughton RW, Frampton CM, Ellmers LJ, Richards AM. Circulating microRNAs as candidate markers to distinguish heart failure in breathless patients. Eur J Heart Fail. 2013;15:1138–1147. doi: 10.1093/eurjhf/hft078. [DOI] [PubMed] [Google Scholar]
- 35.Su Q, Liu Y, Lv XW, Ye ZL, Sun YH, Kong BH, Qin ZB. Inhibition of lncRNA TUG1 upregulates miR-142-3p to ameliorate myocardial injury during ischemia and reperfusion via targeting HMGB1- and Rac1-induced autophagy. Mol Cell Cardiol. 2019;133:12–25. doi: 10.1016/j.yjmcc.2019.05.021. [DOI] [PubMed] [Google Scholar]
- 36.Zhu Y, Lin Y, Yan W, Sun Z, Jiang Z, Shen B, Jiang X, Shi J. Novel biomarker MicroRNAs for subtyping of acute coronary syndrome: A Bioinformatics Approach. Biomed Res Int. 2016;2016:4618323. doi: 10.1155/2016/4618323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Durgan DJ, Smith JK, Hotze MA, Egbejimi O, Cuthbert KD, Zaha VG, Dyck JR, Abel ED, Young ME. Distinct transcriptional regulation of long-chain acyl-CoA synthetase isoforms and cytosolic thioesterase 1 in the rodent heart by fatty acids and insulin. Am J Physiol Heart Circ Physiol. 2006;290:H2480–H2497. doi: 10.1152/ajpheart.01344.2005. [DOI] [PubMed] [Google Scholar]
- 38.Chiu HC, Kovacs A, Ford DA, Hsu FF, Garcia R, Herrero P, Saffitz JE, Schaffer JE. A novel mouse model of lipotoxic cardiomyopathy. J Clin Invest. 2001;107:813–822. doi: 10.1172/JCI10947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yang L, Yang Y, Si D, Shi K, Liu D, Meng H, Meng F. High expression of long chain acyl-coenzyme A synthetase 1 in peripheral blood may be a molecular marker for assessing the risk of acute myocardial infarction. Exp Ther Med. 2017;14:4065–4072. doi: 10.3892/etm.2017.5091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sun CY, She XM, Qin Y, Chu ZB, Chen L, Ai LS, Zhang L, Hu Y. miR-15a and miR-16 affect the angiogenesis of multiple myeloma by targeting VEGF. Carcinogenesis. 2013;34:426–435. doi: 10.1093/carcin/bgs333. [DOI] [PubMed] [Google Scholar]
- 41.Jackstadt R, Hermeking H. MicroRNAs as regulators and mediators of c-MYC function. Biochim Biophys Acta. 2015;1849:544–553. doi: 10.1016/j.bbagrm.2014.04.003. [DOI] [PubMed] [Google Scholar]
- 42.Chamorro-Jorganes A, Araldi E, Penalva LO, Sandhu D, Fernández-Hernando C, Suárez Y. MicroRNA-16 and microRNA-424 regulate cell-autonomous angiogenic functions in endothelial cells via targeting vascular endothelial growth factor receptor-2 and fibroblast growth factor receptor-1. Arterioscler Thromb Vasc Biol. 2011;31:2595–2606. doi: 10.1161/ATVBAHA.111.236521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chan LS, Yue PY, Wong YY, Wong RN. MicroRNA-15b contributes to ginsenoside-Rg1-induced angiogenesis through increased expression of VEGFR-2. Biochem Pharmacol. 2013;86:392–400. doi: 10.1016/j.bcp.2013.05.006. [DOI] [PubMed] [Google Scholar]
- 44.Abu-Halima M, Poryo M, Ludwig N, Mark J, Marsollek I, Giebels C, Petersen J, Schäfers HJ, Grundmann U, Pickardt T, et al. Differential expression of microRNAs following cardiopulmonary bypass in children with congenital heart diseases. J Transl Med. 2017;15:117. doi: 10.1186/s12967-017-1213-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dahl TB, Holm S, Aukrust P, Halvorsen B. Visfatin/NAMPT: A multifaceted molecule with diverse roles in physiology and pathophysiology. Annu Rev Nutr. 2012;32:229–243. doi: 10.1146/annurev-nutr-071811-150746. [DOI] [PubMed] [Google Scholar]
- 46.Qiu L, Liu X. Identification of key genes involved in myocardial infarction. Eur J Med Res. 2019;24:22. doi: 10.1186/s40001-019-0381-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Montecucco F, Bauer I, Braunersreuther V, Bruzzone S, Akhmedov A, Lüscher TF, Speer T, Poggi A, Mannino E, Pelli G, et al. Inhibition of nicotinamide phosphoribosyltransferase reduces neutrophil-mediated injury in myocardial infarction. Antioxid Redox Signal. 2013;18:630–641. doi: 10.1089/ars.2011.4487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang X, Chen J, Huang X. Rosuvastatin attenuates myocardial ischemia-reperfusion injury via upregulating miR-17-3p-mediated autophagy. Cell Reprogram. 2019;21:323–330. doi: 10.1089/cell.2018.0053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tao H, Chen ZW, Yang JJ, Shi KH. MicroRNA-29a suppresses cardiac fibroblasts proliferation via targeting VEGF-A/MAPK signal pathway. Int J Biol Macromol. 2016;88:414–423. doi: 10.1016/j.ijbiomac.2016.04.010. [DOI] [PubMed] [Google Scholar]
- 50.Romaine SP, Tomaszewski M, Condorelli G, Samani NJ. MicroRNAs in cardiovascular disease: An introduction for clinicians. Heart. 2015;101:921–928. doi: 10.1136/heartjnl-2013-305402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dews M, Homayouni A, Yu D, Murphy D, Sevignani C, Wentzel E, Furth EE, Lee WM, Enders GH, Mendell JT, Thomas-Tikhonenko A. Augmentation of tumor angiogenesis by a Myc-activated microRNA cluster. Nat Genet. 2006;38:1060–1065. doi: 10.1038/ng1855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Landskroner-Eiger S, Qiu C, Perrotta P, Siragusa M, Lee MY, Ulrich V, Luciano AK, Zhuang ZW, Corti F, Simons M, et al. Endothelial miR-17~92 cluster negatively regulates arteriogenesis via miRNA-19 repression of WNT signaling. Proc Natl Acad Sci USA. 2015;112:12812–12817. doi: 10.1073/pnas.1507094112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Maciejak A, Kiliszek M, Michalak M, Tulacz D, Opolski G, Matlak K, Dobrzycki S, Segiet A, Gora M, Burzynska B. Gene expression profiling reveals potential prognostic biomarkers associated with the progression of heart failure. Genome Med. 2015;7:26. doi: 10.1186/s13073-015-0149-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Heger J, Bornbaum J, Würfel A, Hill C, Brockmann N, Gáspár R, Pálóczi J, Varga ZV, Sárközy M, Bencsik P, et al. JDP2 overexpression provokes cardiac dysfunction in mice. Sci Rep. 2018;8:7647. doi: 10.1038/s41598-018-26052-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Pfeiffer L, Wahl S, Pilling LC, Reischl E, Sandling JK, Kunze S, Holdt LM, Kretschmer A, Schramm K, Adamski J, et al. DNA methylation of lipid-related genes affects blood lipid levels. Circ Cardiovasc Genet. 2015;8:334–342. doi: 10.1161/CIRCGENETICS.114.000804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Heximer SP, Knutsen RH, Sun X, Kaltenbronn KM, Rhee MH, Peng N, Oliveira-dos-Santos A, Penninger JM, Muslin AJ, Steinberg TH, et al. Hypertension and prolonged vasoconstrictor signaling in RGS2-deficient mice. J Clin Invest. 2003;111:445–452. doi: 10.1172/JCI200318099C. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tang KM, Wang GR, Lu P, Karas RH, Aronovitz M, Heximer SP, Kaltenbronn KM, Blumer KJ, Siderovski DP, Zhu Y, Mendelsohn ME. Regulator of G-protein signaling-2 mediates vascular smooth muscle relaxation and blood pressure. Nat Med. 2003;9:1506–1512. doi: 10.1038/nm958. [DOI] [PubMed] [Google Scholar]
- 58.Wieland T, Lutz S, Chidiac P. Regulators of G protein signalling: A spotlight on emerging functions in the cardiovascular system. Curr Opin Pharmacol. 2007;7:201–207. doi: 10.1016/j.coph.2006.11.007. [DOI] [PubMed] [Google Scholar]
- 59.Wang J, Huang W, Xu R, Nie Y, Cao X, Meng J, Xu X, Hu S, Zheng Z. MicroRNA-24 regulates cardiac fibrosis after myocardial infarction. J Cell Mol Med. 2012;16:2150–2160. doi: 10.1111/j.1582-4934.2012.01523.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Inouye M, Ripatti S, Kettunen J, Lyytikäinen LP, Oksala N, Laurila PP, Kangas AJ, Soininen P, Savolainen MJ, Viikari J, et al. Novel Loci for metabolic networks and multi-tissue expression studies reveal genes for atherosclerosis. PLoS Genet. 2012;8:e1002907. doi: 10.1371/journal.pgen.1002907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Thuny F, Textoris J, Amara AB, Filali AE, Capo C, Habib G, Raoult D, Mege JL. The gene expression analysis of blood reveals S100A11 and AQP9 as potential biomarkers of infective endocarditis. PLoS One. 2012;7:e31490. doi: 10.1371/journal.pone.0031490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Huang X, Yu X, Li H, Han L, Yang X. Regulation mechanism of aquaporin 9 gene on inflammatory response and cardiac function in rats with myocardial infarction through extracellular signal-regulated kinase1/2 pathway. Heart Vessels. 2019;34:2041–2051. doi: 10.1007/s00380-019-01452-8. [DOI] [PubMed] [Google Scholar]
- 63.Yang Y, Cheng HW, Qiu Y, Dupee D, Noonan M, Lin YD, Fisch S, Unno K, Sereti KI, Liao R. MicroRNA-34a plays a key role in cardiac repair and regeneration following myocardial infarction. Circ Res. 2015;117:450–459. doi: 10.1161/CIRCRESAHA.117.305962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Huang Y, Qi Y, Du JQ, Zhang DF. MicroRNA-34a regulates cardiac fibrosis after myocardial infarction by targeting Smad4. Expert Opin Ther Targets. 2014;18:1355–1365. doi: 10.1517/14728222.2014.961424. [DOI] [PubMed] [Google Scholar]
- 65.Matsumoto S, Sakata Y, Suna S, Nakatani D, Usami M, Hara M, Kitamura T, Hamasaki T, Nanto S, Kawahara Y, Komuro I. Circulating p53-responsive microRNAs are predictive indicators of heart failure after acute myocardial infarction. Circ Res. 2013;113:322–326. doi: 10.1161/CIRCRESAHA.113.301209. [DOI] [PubMed] [Google Scholar]
- 66.Bernardo BC, Gao XM, Winbanks CE, Boey EJ, Tham YK, Kiriazis H, Gregorevic P, Obad S, Kauppinen S, Du XJ, et al. Therapeutic inhibition of the miR-34 family attenuates pathological cardiac remodeling and improves heart function. Proc Natl Acad Sci USA. 2012;109:17615–17620. doi: 10.1073/pnas.1206432109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Bernardo BC, Gao XM, Tham YK, Kiriazis H, Winbanks CE, Ooi JY, Boey EJ, Obad S, Kauppinen S, Gregorevic P, et al. Silencing of miR-34a attenuates cardiac dysfunction in a setting of moderate, but not severe, hypertrophic cardiomyopathy. PLoS One. 2014;9:e90337. doi: 10.1371/journal.pone.0090337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Stephanou A, Brar BK, Scarabelli TM, Jonassen AK, Yellon DM, Marber MS, Knight RA, Latchman DS. Ischemia-induced STAT-1 expression and activation play a critical role in cardiomyocyte apoptosis. J Biol Chem. 2000;275:10002–10008. doi: 10.1074/jbc.275.14.10002. [DOI] [PubMed] [Google Scholar]
- 69.Stephanou A. Role of STAT-1 and STAT-3 in ischaemia/reperfusion injury. J Cell Mol Med. 2004;8:519–525. doi: 10.1111/j.1582-4934.2004.tb00476.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhang S, Liu W, Liu X, Qi J, Deng C. Biomarkers identification for acute myocardial infarction detection via weighted gene co-expression network analysis. Medicine (Baltimore) 2017;96:e8375. doi: 10.1097/MD.0000000000008375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chang PC, Lin SF, Chu Y, Wo HT, Lee HL, Huang YC, Wen MS, Chou CC. LCZ696 therapy reduces ventricular tachyarrhythmia inducibility in a myocardial infarction-induced heart failure rat model. Cardiovasc Ther. 2019;2019:6032631. doi: 10.1155/2019/6032631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Li SY, Li ZX, He ZG, Wang Q, Li YJ, Yang Q, Wu DZ, Zeng HL, Xiang HB. Quantitative proteomics reveal the alterations in the spinal cord after myocardial ischemia-reperfusion injury in rats. Int J Mol Med. 2019;44:1877–1887. doi: 10.3892/ijmm.2019.4341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Getz GS, Reardon CA. Do the Apoe−/- and Ldlr−/−Mice yield the same insight on atherogenesis? Arterioscler Thromb Vasc Biol. 2016;36:1734–1741. doi: 10.1161/ATVBAHA.116.306874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Frangogiannis NG. Chemokines in the ischemic myocardium: From inflammation to fibrosis. Inflamm Res. 2004;53:585–595. doi: 10.1007/s00011-004-1298-5. [DOI] [PubMed] [Google Scholar]
- 75.Frangogiannis NG, Entman ML. Chemokines in myocardial ischemia. Trends Cardiovasc Med. 2005;15:163–169. doi: 10.1016/j.tcm.2005.06.005. [DOI] [PubMed] [Google Scholar]
- 76.Boengler K, Hilfiker-Kleiner D, Drexler H, Heusch G, Schulz R. The myocardial JAK/STAT pathway: From protection to failure. Pharmacol Ther. 2008;120:172–185. doi: 10.1016/j.pharmthera.2008.08.002. [DOI] [PubMed] [Google Scholar]
- 77.Fuglesteg BN, Suleman N, Tiron C, Kanhema T, Lacerda L, Andreasen TV, Sack MN, Jonassen AK, Mjøs OD, Opie LH, Lecour S. Signal transducer and activator of transcription 3 is involved in the cardioprotective signalling pathway activated by insulin therapy at reperfusion. Basic Res Cardiol. 2008;103:444–453. doi: 10.1007/s00395-008-0728-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Liu YH, Wang D, Rhaleb NE, Yang XP, Xu J, Sankey SS, Rudolph AE, Carretero OA. Inhibition of p38 mitogen-activated protein kinase protects the heart against cardiac remodeling in mice with heart failure resulting from myocardial infarction. J Card Fail. 2005;11:74–81. doi: 10.1016/j.cardfail.2004.04.004. [DOI] [PubMed] [Google Scholar]
- 79.Ge ZW, Zhu XL, Wang BC, Hu JL, Sun JJ, Wang S, Chen XJ, Meng SP, Liu L, Cheng ZY. MicroRNA-26b relieves inflammatory response and myocardial remodeling of mice with myocardial infarction by suppression of MAPK pathway through binding to PTGS2. Int J Cardiol. 2019;280:152–159. doi: 10.1016/j.ijcard.2018.12.077. [DOI] [PubMed] [Google Scholar]
- 80.Liu X, Chen K, Zhuang Y, Huang Y, Sui Y, Zhang Y, Lv L, Zhang G. Paeoniflorin improves pressure overload-induced cardiac remodeling by modulating the MAPK signaling pathway in spontaneously hypertensive rats. Biomed Pharmacother. 2019;111:695–704. doi: 10.1016/j.biopha.2018.12.090. [DOI] [PubMed] [Google Scholar]
- 81.Li C, Li J, Xue K, Zhang J, Wang C, Zhang Q, Chen X, Gao C, Yu X, Sun L. MicroRNA-143-3p promotes human cardiac fibrosis via targeting sprouty3 after myocardial infarction. J Mol Cell Cardiol. 2019;129:281–292. doi: 10.1016/j.yjmcc.2019.03.005. [DOI] [PubMed] [Google Scholar]
- 82.Jia L, Zhu L, Wang JZ, Wang XJ, Chen JZ, Song L, Wu YJ, Sun K, Yuan ZY, Hui R. Methylation of FOXP3 in regulatory T cells is related to the severity of coronary artery disease. Atherosclerosis. 2013;228:346–352. doi: 10.1016/j.atherosclerosis.2013.01.027. [DOI] [PubMed] [Google Scholar]
- 83.Abbate A, Bonanno E, Mauriello A, Bussani R, Biondi-Zoccai GG, Liuzzo G, Leone AM, Silvestri F, Dobrina A, Baldi F, et al. Widespread myocardial inflammation and infarct-related artery patency. Circulation. 2004;110:46–50. doi: 10.1161/01.CIR.0000133316.92316.81. [DOI] [PubMed] [Google Scholar]
- 84.Fan L, Meng H, Guo X, Li X, Meng F. Differential gene expression profiles in peripheral blood in northeast chinese han people with acute myocardial infarction. Genet Mol Biol. 2018;41:59–66. doi: 10.1590/1678-4685-gmb-2017-0075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Muse ED, Kramer ER, Wang H, Barrett P, Parviz F, Novotny MA, Lasken RS, Jatkoe TA, Oliveira G, Peng H, et al. A whole blood molecular signature for acute myocardial infarction. Sci Rep. 2017;7:12268. doi: 10.1038/s41598-017-12166-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Park HJ, Noh JH, Eun JW, Koh YS, Seo SM, Park WS, Lee JY, Chang K, Seung KB, Kim PJ, Nam SW. Assessment and diagnostic relevance of novel serum biomarkers for early decision of ST-elevation myocardial infarction. Oncotarget. 2015;6:12970–12983. doi: 10.18632/oncotarget.4001. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available in the GEO repository under accession nos. GSE24548 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE24548), GSE97320 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE97320), GSE24519 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE24519), GSE66360 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE66360) and GSE60993 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE60993).