

Abstract
Metabolic reprogramming is a physiological cellular adaptation to intracellular and extracellular stimuli that couples to cell polarization and function in multiple cellular subsets. Pathological conditions associated to nutrients overload, such as dyslipidaemia, may disturb cellular metabolic homeostasis and, in turn, affect cellular response and activation, thus contributing to disease progression. At the vascular/immune interface, the site of atherosclerotic plaque development, many of these changes occur. Here, an intimate interaction between endothelial cells (ECs), vascular smooth muscle cells (VSMCs) and immune cells, mainly monocytes/macrophages and lymphocytes, dictates physiological versus pathological response. Furthermore, atherogenic stimuli trigger metabolic adaptations both at systemic and cellular level that affect the EC layer barrier integrity, VSMC proliferation and migration, monocyte infiltration, macrophage polarization, lymphocyte T and B activation. Rewiring cellular metabolism by repurposing “metabolic drugs” might represent a pharmacological approach to modulate cell activation at the vascular immune interface thus contributing to control the immunometabolic response in the context of cardiovascular diseases.
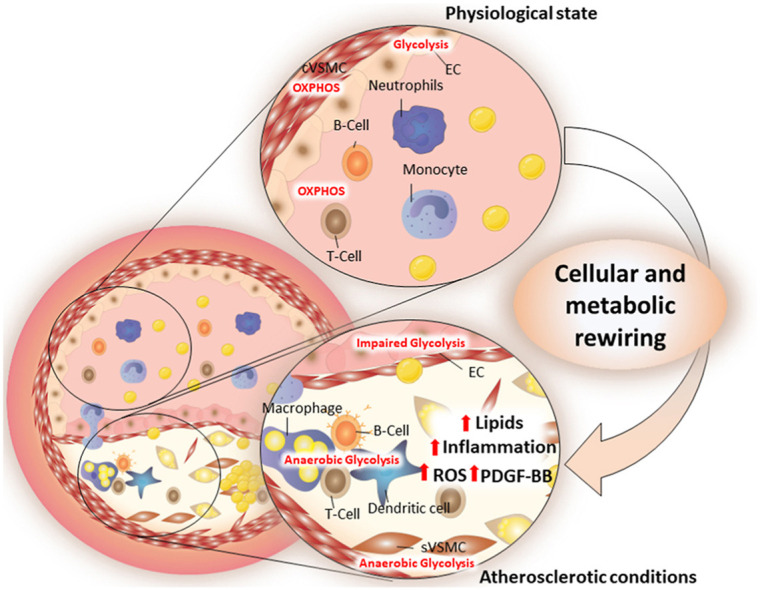
Graphical abstract
1. Metabolic adaptations in vascular wall cells
Atherosclerosis is a chronic-inflammatory disease of the large and medium-size arteries eventually leading to severe clinical manifestations, including myocardial infarction, angina, heart failure and stroke. Low-density lipoprotein cholesterol (LDL-C) plasma levels are directly correlated with their accumulation in the intima of the arteries where they promote endothelial dysfunction, immune cell recruitment, followed by smooth muscle cells migration and proliferation which contribute to extracellular matrix production and chronicization of the immune-inflammatory response in the plaque. Aim of this review is to discuss the relevance of cellular metabolic adaptations occurring at the vascular-immune interface during atherogenesis and how they impact disease progression. Understanding of these novel physio-pathological mechanisms may unveil potential innovative pharmacological targets and therapeutic opportunities for cardiovascular diseases.
1.1. Endothelial cells
Endothelial cells (ECs) serve as a selective barrier between blood and underlining tissues, control oxygen and nutrients supply, and participate to immune surveillance. Depending on the vascular bed, ECs are endowed with tissue-specific functions (Kalucka et al., 2020) that for the arteries include the regulation of vascular tone and redox balance, the control of haemostasis, thrombosis and of the inflammatory response (Gimbrone and Garcia-Cardena, 2016). These activities are achieved by orchestrating the biosynthesis and degradation of vasoactive mediators, extracellular matrix components, growth factors, cytokines and hormone-like molecules, prostaglandins and autacoids and the transport and metabolism of lipoproteins (Gimbrone and Garcia-Cardena, 2016). These multiple mechanisms which participate to tissue homeostasis, largely explain why ECs dysfunction represents a critical step for the development of atherosclerosis.
1.1.1. ECs metabolism and function and their relation to hemodynamic forces
Under physiological conditions, ECs maximize energy production by anaerobic glycolysis, that accounts for almost 85% of ATP production as compared to oxidation of glucose, fatty acids or amino acids by the electron transport chain (Eelen et al., 2018). This metabolic preference is also preserved by hemodynamic forces that in concert shape ECs morphology and function. Indeed, laminar shear stress contributes to maintain a quiescent state of ECs by inducing the expression of Kruppel-like factor 2 (KLF2), that besides vascular barrier integrity (Dekker et al., 2006), controls cellular glycolytic rate by modulating glucose uptake and phosphofructokinase-2/fructose-2,6-bisphosphatase-3 (PFKFB3) transcription (Doddaballapur et al., 2015; Sathanoori et al., 2015) a rate limiting enzyme in the aerobic glycolytic cascade, thus ensuring an adequate level of glycolytic flux to sustain EC homeostasis. In parallel, shear stress, by promoting cytosolic recruitment and activation of the GTPase dynamin-related protein 1(Drp1) (Breton-Romero et al., 2014), supports the formation of small and globular mitochondria, a phenomena known as fission which is associated to a low oxidative metabolism and reactive oxygen species (ROS) production as opposed to lactate (the final product of anaerobic glycolysis) which is increased. The latter appears particularly relevant in hypoxic conditions as lactate promotes EC sprouting and new blood vessel development (Ruan and Kazlauskas, 2013), an event particularly relevant in poorly oxygenated tissues and during the recovery phase after acute myocardial infarction.
At arterial branch points and curvatures, blood flow turns from laminar and unidirectional (typical of the straight parts of the arterial tree) to oscillatory, thus resulting in a minimal but continuous shear stress on ECs. At these sites, which are regarded as “athero-prone” vascular beds, ECs acquire a round-shape, with a non-uniform orientation, and alter the production of vasoactive molecules, turning ECs toward a pro-thrombotic and inflamed layer with increased permeability, favouring lipid deposition within the intima of the arterial wall (Cahill and Redmond, 2016). At this site the presence of turbulent blood flow impacts endothelial function by different mechanisms (Brandes et al., 2014). First, low shear stress could alter the function of mechano-sensing receptors located at the apical or basal pole or at the lateral border of ECs, among them alterations in the function and localization of the transmembrane glycoprotein receptor CD31, that impacts survival (Cheung et al., 2015; Gao et al., 2003) and maintenance of EC barrier integrity (Flynn et al., 2013; Privratsky and Newman, 2014), vascular tone (Bagi et al., 2005; Russell-Puleri et al., 2017), angiogenesis (Park et al., 2010), and lowers thrombogenicity, ROS production (Ji et al., 2002; Saragih et al., 2014), adhesion and inflammatory molecules expression (Cepinskas et al., 2003; Couty et al., 2007); second, low shear stress upregulates NADPH oxidase (NOX) expression which causes NADPH shortage. Given the critical role of NADPH in supporting endothelial nitric oxidase (eNOS) function, under NADPH shortage, eNOS becomes “uncoupled” and switches to the generation of superoxide anions (O2 −), that can react with NO to form peroxynitrile (ONOO−), a highly reactive ROS.
At the cellular level, proper EC functionality is maintained through a strict control on metabolic fluxes. Side pathways of glycolysis such as the pentose phosphate pathway (PPP) or the hexosamine biosynthetic pathway (HBP) are critical to reduce NADP+ to NADPH and support glutathione regeneration (GSH), or to generate glucosamine-6-phosphate (GlcN6P), that is converted to uridine diphosphate N-acetylglucosamine (UDP-GlcNAc), which is required for O- and N- protein glycosylation (Koo et al., 2013). Glycosylation of key ECs receptors, such as CD31, vascular endothelial growth factor receptor 2 (VEGFR2) and VE-cadherin (vascular endothelial cadherin), is central for EC angiogenesis, survival, proliferation and migration (Chandler et al., 2019; Imamaki et al., 2018; Park et al., 2016). For instance, reduction of N-glycosylation, as a consequence of inhibition of glycolysis by 2-Deoxy-d-glucose (2-DG) (a glucose analogue that competitively blocks phosphoglucose isomerase), dampens angiogenesis (Merchan et al., 2010). Also, the maintenance of the endothelial glycocalyx, a negatively charged and organized mesh of membranous glycoproteins, proteoglycans and associated plasma proteins (of about 100–750 nm thickness), that contributes to vascular wall protection (Yilmaz et al., 2019), is dependent on glycolysis-related protein glycosylation (Fig. 1 ).
Fig. 1.

In blood vessels characterized by high shear stress and laminar flow, ECs utilise a glycolytic metabolism that not only provides energy (ATP), but fuels collateral pathway as the HBP and PPP, required for efficient protein glycosylation, glycocalix maintenance, and antioxidant activities. In addition, conversion of pyruvate to lactate, which occur under low oxygen conditions, is a stimulus for EC sprouting for new vessel formation, while acetylCoA derived both from pyruvate or exogenous fatty acids oxidation is not only consumed in TCA cycle but partecipates to epigenetic remodeling and nucleotide synthesis. Physiologically also eNOS regulates vascular tone by converting L-arginine to NO. However, when blood flow is disturbed, glycolytic flux is reduced and endothelial barrier integrity is compromised, thus resulting in increased lipid deposition. These events contribute to EC dysfunction and lead to increased inflammation, oxidative stress and reduced NO production.
Similarly, cellular fatty acids (FAs) synthesis contributes to the maintenance of vascular endothelial tone by providing palmitic acid which is required for eNOS palmitoylation (Wei et al., 2011), a post-translational modification required for its efficient function (Liu et al., 1996), that allows eNOS association to membrane caveolae (50–100 nm omega-shaped pits enriched in cholesterol and sphingolipids (Galbiati et al., 2001)). In addition, while FA synthesis is critical for some EC activities, the intracellular pool of FAs may also derive from the uptake of albumin-bound FAs or from the lipolysis of triglyceride-rich lipoproteins followed by selective uptake via CD36 and fatty acid transport proteins (FATPs); extracellular FAs are critical to provide linoleic acid (18:2) that, converted into arachidonic acid (AA) (C20:4), is pivotal to the generation of prostaglandins (PGD2, PGE2 or PGF2), thromboxane A2 (TXA2), prostacyclin (PGI2) and leukotriens (Marcus et al., 1982).
1.1.2. Impact of systemic metabolic alterations on endothelial barrier integrity and function
Altered shear stress and enhanced turbulent blood flow at inner curvatures and branch points of arteries increase the chances to trap LDL and other cholesterol-rich apoB-containing particles into the intima of the vessels through transcytosis and/or tight junction dysfunction (Mundi et al., 2018). This process is triggered by enhanced endothelial permeability, coupled to hypercholesterolemia and is considered the critical starting point driving EC switch to a pro-inflammed, pro-thrombotic and atherogenic layer. Recently attention was focused on understanding how lipid accumulation could influence EC metabolism together with promoting EC activation. Lipids carried by lipoproteins (mainly in their oxidized form) induce a strong activation of the inflammatory response through the activation of the endothelial NF-kB cascade; this event leads to the production of pro-inflammatory cytokines (TNFα, IL-1β), chemoatractant proteins (CCL1, CX3CL1, CCL5), and leukocyte adhesion molecules (E-selectin and VCAM-1), that are pivotal for immune cell recruitment at the vessel wall. Incubation of ECs with triglycerides-rich lipoproteins (TGRLs) promotes the expression of adhesion molecules (VACM-1, E-selectin and CD31) (Norata et al., 2006) and pro-inflammatory genes (PAI-1, CCL2 and IL-6), an effect more pronounced with TGRL from patients with hypertrygliceridemia or increased levels of small dense atherogenic LDL (sdLDL) (Norata et al., 2007; Norata et al., 2009). Interestingly, removal of cholesterol from ECs, such as by high-density lipoproteins (HDL) exerts, anti-inflammatory, anti-thrombotic and anti-proteolytic activity on ECs and thus counteracts the pro-atherogenic potential induced by lipid accumulation (Norata and Catapano, 2005). The athero-protective role of HDL is further confirmed by the observation that these mechanisms of defence are lost in case of oxidative modifications of HDL, as observed in patients with coronary artery disease (CAD) (Kratzer et al., 2014), thus turning HDL to pro-inflammatory mediators (Callegari et al., 2006; Norata et al., 2004; Pirillo et al., 2019).
Beyond hypercholesterolemia, also increased plasma glucose levels were shown to affect EC metabolism and function. In diabetic patients, hyperglycaemia favours EC intracellular glucose uptake that, however, is diverted into the polyol pathway to produce sorbitol and fructose, that ultimately promotes the nonenzymatic generation of AGEs; a negative effect also supported by marked activation of insulin signalling pathways which further fuels glucose uptake via intracellular trafficking of GLUT4 (de Zeeuw et al., 2015). Excessive glucose uptake also affects mitochondrial dynamics, with an increase in organelle fission leading to mitochondrial fragmentation, elevated ROS production which could contribute to EC toxicity (Kluge et al., 2013).
Dietary nutrients which are metabolized by the gut microbiota also impact endothelial function and CVD. Trimethylamine N-Oxide (TMAO), which is derived from conversion in the liver of TMA (produced by gut microbiota from choline/phosphatidylcholine and L-carnitine), activates NF-KB signalling in ECs thus triggering a pro-inflammatory response, followed by the recruitment and adhesion of leukocytes to ECs (Boini et al., 2017). This mechanism appears to be critical to explain the proatherogenic role of this molecule and the association of its plasma levels with CAD. On the contrary, other microbiota-derived molecules exert beneficial effects on endothelial integrity and function. This is the case of short-chain fatty acids (SCFAs), fatty acids with less than six carbon atoms, such as acetic acid, propionic acid and butyric acid or gaseous metabolites, as Hydrogen Sulfide (H2S), that improves vascular tone (Amedei and Morbidelli, 2019).
In summary endothelial function is deeply related to its metabolism under physiological conditions as well as when metabolic capacity is exceeding (such as in presence of excessive extracellular nutrients). In the latter case, ECs may undergo specific adaptations that impact the maintenance of barrier integrity and vascular tone and in the long term contributes to atherogenesis also by influencing the function of other cells present in the arteries such vascular smooth muscle cells and/or promoting immune cell recruitment.
1.2. Vascular smooth muscle cells
Lying below the endothelial layer of the tunica media of vascular vessel, vascular smooth muscle cells (VSMCs) are highly contractile cells involved in the maintenance of vessel structure and vascular tone. Under physiological conditions, autocrine production of TGFβ maintains the contractile phenotype, by promoting the transcription of the co-activator Myocardin (MYOCD), that controls the expression of contractile smooth muscle cell (SMC) proteins, such as smooth muscle cell myosin heavy chain (MYH11), SM22a/Transgelin, actin (ACTA2) and smoothelin. Under perturbed and damaged conditions, VSMCs may turn to a synthetic phenotype state to preserve vasal functionality and promote vascular remodeling; this phenomenon is known as VSMCs plasticity and is tightly regulated at the transcriptional level (Owens et al., 2004).
1.2.1. Adaptations of VSMC metabolism from physiological to pro-atherogenic conditions
Contractile SMCs mainly rely on aerobic metabolism via OXPHOS, where carbohydrates are used through the glycolytic pathway to support the mitochondrial cycle of tricarboxylic acids (TCA) and the production of energy by the electron transport chain. Functional mitochondria are therefore key for energy production in VSMCs. Of note, mitochondria are highly dynamic organelles, continuously switching between fused and fixed forms. The fused form of mitochondria is characterized by a higher metabolic activity due to the presence of a complex network of cristae and more electron transport chain complexes. On the contrary, fixed mitochondria are less metabolically active and more prone to be recycled toward the autophagic process called mitophagy. Fission allows the formation of smaller and more globular mitochondria and is mediated by the activation of dynamins, such as Dynamin Related Protein 1 (Drp1), which are cytosolic proteins recruiting mitochondrial fission 1 (FIS1) protein at the mitochondrial membrane, to form a spiral around the outer membrane of mitochondria (OMM) constricting and cleaving mitochondria into two parts. At the opposite, fusion is characterized by the formation of a complex mitochondrial network where mitofusin1 (MFN1) and mitofusin2 (MFN2) mediate the fusion of the OMM, while optic atrophy 1 (OPA1) mediates that of the internal mitochondrial membrane (IMM). This mitochondrial dynamism has a huge impact on cellular functionality and differentiation, with contractile VSMCs showing more fused mitochondria and increased need of FAs as a source of acetyl-CoA for TCA cycle (Kuznetsov et al., 2009).
VMSCs plasticity is particularly relevant during atherogenesis when VSMCs turn to the synthetic phenotype, characterized by highly proliferative and migratory potential. MicroRNAs such as mir143/145 are instrumental in supporting a pro-atherogenic switch toward a synthetic phenotype (Norata et al., 2012, 2013) which occurs when TGFβ production is reduced while platelet-derived grow factor BB (PDGF-BB), produced locally by SMCs and inflammatory cells, is increased, thus promoting VSMC migration to the intima, neointimal hyperproliferation and LDL uptake. Moreover, PDGF-BB stimulation coupled to the accumulation of cholesterol and oxidized phospholipids promotes Kruppel-Like Factor 4 (KLF4) activation (Deaton et al., 2009), a pluripotent transcription factor, that in turn, upregulates the expression of proinflammatory proteins, proliferative genes, ultimately leading to VSMC switch to “macrophage-like foam cells” (Tang et al., 2011) with phagocytotic ability (Shankman et al., 2015).
Metabolically, VSMCs polarization toward a synthetic state is strictly dependent on cellular metabolic rewiring, with increased anabolic and catabolic rate to support macromolecules production and energy generation. Proliferation of synthetic VSMCs requires the production of building blocks necessary for the synthesis of new macromolecules. This effect is mediated by PDGF-BB-dependent PI3K/Akt activation (Perez et al., 2010) and the subsequent increase in the expression of genes involved in anaerobic glycolysis including glucose transporter type 4 (GLUT4), hexokinase (HK2) and lactate dehydrogenase (LDH), which support a faster generation of ATP in response to proliferative stimulus as compared to glycolysis coupled to OXPHOS. Of note, this metabolic adaptation resembles the Warburg effect described in cancer cells, where increased glycolytic flux also fuels side pathways that are used for the biosynthesis of new macromolecules, including ribose 5-phosphate, a precursor of nucleotide synthesis, and NADPH for FA production, both critical for cell proliferation and differentiation (Werle et al., 2005). Increased glycolytic rate also provides the intermediate 3-phosphoglycerate, used for the synthesis of new amino acids and, therefore, proteins. Interestingly, proliferating VSMCs adopt anaerobic glycolysis (Barron and Parrillo, 1995) both in hypoxic condition, an effect that depends on the activation of hypoxia-inducible factor 1-alpha (HIF1a) (Liu et al., 2017), and even under normal oxygen levels. Of note, the oxidation of lactate from pyruvate produces NAD+ from NADH, a critical cofactor of glycolysis, which further fuels anaerobic metabolism. Under these conditions, the excess of lactate is eliminated extracellularly by specific transporters and can act as signalling molecule by activating mTOR, AMPK and PI3K/Akt in a paracrine fashion. PDGF-BB also impacts mitochondrial plasticity by reducing MFN2 expression, thus limiting mitochondrial fusion, and supporting mitochondrial fragmentation (fixation). The latter status induces the preferential use of FAs as a metabolic substrate for energy production through fatty acid β-oxidation rather than using Acetyl-CoA and pyruvate coming from glucose metabolism. The increased metabolic flux thought β-oxidation coupled to mitochondrial fixation favors the generation of ROS (Turrens, 2003), that may drive lipid peroxidation and promote mitochondrial damage (Fig. 2 ).
Fig. 2.

Lying below the endothelial layer of vascular vessel, vascular smooth muscle cells (VSMCs) are highly contractile cells involved in the maintenance of vessel structure and vascular tone. Under physiological condition contractile VSMCs use aerobic glycolysis to fuel oxidative phosphorylation (OXPHOS) to generate energy, present a more fused mitochondrial network characterized by an increased expression of Mitofusin2 (MFN2). During atherogenesis, VSMCs turn to a synthetic phenotype characterized by highly proliferative and migratory potential. To maintain the increased anabolic demands including nucleotide and amino acids synthesis, synthetic VSMCs preferentially switch to anaerobic glycolysis with a fixed mitochondrial network.
In summary, atherogenic conditions induce PDGF-BB production which promotes VSMCs phenotype switch by acting both on cellular metabolism and mitochondrial dynamics. The infiltration of immune cells in the tunica intima plays a key role in redirecting VMSCs fate toward a proliferative phenotype as they lose their primary function of contraction in the tunica media, migrate in the intima in response to pro-inflammatory activation of ECs thus participating to atheroprogression.
2. Metabolic adaptation in innate and adaptive immune cells
Accumulation of lipoproteins and lipids in the intima of arterial walls elicits an immune response where pro-inflammatory leukocytes are recruited to the vascular interface thanks to the engagement with chemoattractant and adhesion molecules expressed by endothelial cells. This response, which initially is aimed at clearing potentially damaging lipid accumulation, turns to a chronic inflammatory response that perpetuates immune cell recruitment and activation in the atherosclerotic plaque (Yvan-Charvet et al., 2019). These cells undergo specific immunometabolic reprogramming as a consequence of different substrate availability in the plaque, together with the need to acquire/maintain the activation status and to support innate and adaptive immune responses (Fig. 3 ).
Fig. 3.

Metabolic status of immune cells differs from physiological to atherosclerotic conditions. Under physiological conditions, circulating monocytes and lymphocytes (both T and B cells) mainly rely on oxidative metabolism (OXPHOS). During atherosclerosis, increased cholesterol levels trigger lipid accumulation both in circulating monocytes as well as in atherosclerotic plaque macrophages thus leading to cell reprogramming and the switch from oxidative metabolism to anaerobic glycolysis, which in turn guides their pro-inflammatory activation. In dendritic cells, cholesterol accumulates in plasma membrane and cytoplasm leading to increased antigen presenting function and pro-inflammatory cytokines release. In adaptive immune cells, lipid overload and the presence of atherosclerosis-related antigens contribute to B and T cells activation, favouring OSE antibodies production in B cells and the polarization toward an effector phenotype in T cells; both cells are characterized by faster glycolytic metabolism and increased demand of cholesterol to sustain cell proliferation.
2.1. Metabolic changes associated to monocyte and macrophage activation
The vast majority of macrophages present in the atherosclerotic plaque derive from circulating monocytes. Monocytosis is an independent risk factor for CVD and is associated to increased atherogenesis and impaired resolution of inflammation (Flynn et al., 2019). Hypercholesterolemia and cholesterol enrichment in bone marrow and splenic hematopoietic cells increase the rate of monocyte precursor proliferation (Yvan-Charvet et al., 2010), and similar mechanisms have been reported in hyperglycaemic conditions (Nagareddy et al., 2013). When cellular cholesterol metabolism is impaired, such as is the case for defects of cholesterol efflux, monocytes acquire a pro-inflammatory state that is probably a consequence of epigenetic priming occurring in myeloid precursors in the bone marrow (Christ et al., 2018). This epigenetic priming extends also to genes associated to cellular metabolism, resulting in a preferential use of aerobic glycolysis toward oxidative phosphorylation that contributes to maintain a prolonged state of hyperactivation that increases the ability of monocytes to reach the atherosclerotic plaque and become macrophages.
Monocyte derived-macrophages, which are specialized cells committed to recognize “damage” signals and engulf “not self” material, localize in the intima to clear the tissue from modified and oxidized lipoproteins by engaging scavenger receptors (SR), including scavenger receptor A (SR-A), SR-B1, SREC-1, MARCO, CD36, LOX-1 and scavenger receptor for phosphatidylserine and oxidized LDL (also known as CXCL16) (Kzhyshkowska et al., 2012). The expression of these receptors, unlike the LDL-R, is not regulated by cellular cholesterol content and therefore, under continuous lipid overload, macrophages turn into foam cells that are trapped within the arterial intima where they can ultimately die causing necrotic core formation together with the release of cholesterol crystals and other cellular debris. At the molecular level, oxLDL trigger the inflammatory “priming” of macrophages leading to increased expression of IL-1β mRNA and the activation of the nucleotide-binding domain–like receptors Family Pyrin Domain Containing 3 (NLRP3) inflammasome (Sheedy et al., 2013). Of note, a proper NLRP3 activity requires to boost both fatty acid synthesis (FAS) and fatty acid oxidation (FAO), with FAS promoting inflammatory gene transcription and FAO boosting ROS production (Moon et al., 2015, 2016). In parallel, pro-inflammatory activation of macrophages is characterized by increased glycolytic flux to sustain rapid demand of energy for ATP production and, in line with this, monocyte-derived macrophages from patients with coronary artery disease (CAD) display elevated glucose utilization compared to control subjects (Shirai et al., 2016); indeed imaging of glucose uptake in the plaque is used to detect macrophage infiltration. This metabolic preference associates with increased mitochondrial stress and production of IL-6 and IL-1β, that is typical of M1 pro-inflammatory macrophages while anti-inflammatory or pro-resolutive M2 macrophages prefer oxidative phosphorylation. It is important to stress that immune cells continuously undergo metabolic adaptations, that in macrophages is explified by the engagement of SREBP1-dependent lipid metabolism, after a first glycolytic phase.; De novo lipid synthesis is required both to support the interaction between membrane lipid rafts and actin cytoskeleton for an efficient phagocytosis (Lee et al., 2018), and to generate anti-inflammatory lipids during the pro-resolutive phase (Oishi et al., 2017), such as resolvins, bioactive molecules that stimulate self-limited innate responses, that enhance innate microbial killing and clearance, and are organ-protective (Serhan and Levy, 2018). Of note, by controlling ATP-citrate lyase (ACLY) (Sato et al., 2000), SREBP also regulates the amount of acetyl-CoA that can be used for histone acetylation and thus influences the expression of several pro-inflammatory genes (Lauterbach et al., 2019). In parallel, in the context of the atherosclerotic plaque, hypoxia activates the HIF-1α pathway which in turn, promotes glycolysis, NLRP3 activation (Folco et al., 2014), sterol synthesis and suppresses cholesterol efflux. The latter mechanisms contribute to the accumulation of cellular cholesterol which ultimately leads to cell death.
Adaptations in cellular metabolism also occur during efferocytosis, a process used by phagocytes for the clearance of apoptotic macrophages, indeed, first the conversion of arginine to ornithine and putrescine is pivotal to drive Rac1 activation (Yurdagul et al., 2020) and second the activation of lysosomal acid lipase (LAL) guides the hydrolysis of esterified lipids derived from apoptotic cell engulfment, and provides the substrate for the synthesis of 25-hydroxycholesterol and 27-hydroxycholesterol, both endogenous ligands of liver-X-receptor (LXR), a transcription factor controlling cholesterol efflux (Gomaraschi et al., 2019; Viaud et al., 2018); these mechanisms are critical to promote macrophage pro-resolutive activity.
In parallel with monocytes and macrophages, other innate immune cells populate the atherosclerotic plaque, including neutrophils, natural killer cells (NKs) and mast cells. While the role of NKs is still controversial (Winkels and Ley, 2018), both mast cells and neutrophils contribute to disease severity (Drechsler et al., 2010; Kovanen and Bot, 2017). Neutrophils are recruited at early stages of atherosclerosis where they foster monocyte recruitment and macrophage activation by cytotoxic mechanisms that are exacerbated under hypercholesterolaemic and hyperglycaemic conditions through a feedback mechanism increasing neutrophil generation in the bone marrow and at extramedullary sites.
2.2. Immunometabolic regulation of adaptive response
Dendritic cells (DCs), are normally found in healthy arteries, accumulate in atherosclerotic lesions where engulf “atherosclerosis-related antigens” and thereafter migrate to lymphatic tissues, including tertiary lymphoid organs and vessel adventitia in a CCL19/21-CCR7 dependent manner, to instruct lymphocytes (Trogan et al., 2006; Zernecke, 2015). Also, DC activity is controlled by cellular cholesterol metabolism; indeed, a proper cholesterol content in lipid rafts is required to maintain a functional MHCII localization and apolipoprotein E works in an autocrine fashion to maintain this equilibrium (Bonacina et al., 2018). During atherosclerosis, a massive cellular lipid accumulation triggers NLRP3 activation and supports DCs antigen-presentation and lymphocyte activation (Bonacina et al., 2018; Ito et al., 2016; Westerterp et al., 2017) which occurs also in the aorta (MacRitchie et al., 2020). Lymphocyte activation, in turn, is generally accompanied by a metabolic switch from lipid oxidation, characteristic of naïve state, to cellular glycolysis which is orchestrated by mTOR, an evolutionarily conserved ser/thr kinase that integrates nutrient sensing and antigen-receptor signalling (Marelli-Berg et al., 2012). Upon TCR engagement, T cells upregulate glucose transporters and increase glycolytic flux for both rapid ATP production and generation of biosynthetic precursors for amino acids and nucleic acids via the pentose phosphate pathway (PPP) (Lunt and Vander Heiden, 2011). This metabolic rewiring allows both clonal expansion and the acquisition of effector functions; on the contrary, the inhibition of glycolysis selectively reduces the expression of cell cycle proteins and the production of IFNγ and granzyme, in both mouse and human CD8 T cells (Cham et al., 2008; Cham and Gajewski, 2005; Renner et al., 2015). Besides, T cells polarization, differentiation, migration and the acquisition of a memory phenotype are controlled by cellular metabolism (Norata et al., 2015; Vuononvirta et al., 2020). In patients with atherosclerotic diseases and in experimental mice, lymphocytes are more polarized toward T effectors subsets (Ammirati et al., 2012); whether this is the result of the atherosclerotic plaque inflammatory milieu or could depend on a direct metabolic effect of hypercholesterolaemia on lymphocyte sterol metabolism is debated. However, cholesterol has been shown to be critical in proliferating T cells to support their anabolic needs, including cell membrane production, while, when too abundant, becomes cytotoxic. Homeostasis of cellular cholesterol is controlled by two classes of transcription factors: LXRs and SREBPs. T cell proliferation is associated with a downregulation of pathways involved in cholesterol efflux and promotion of lipid synthesis (LXRs repression and SREBPs activation), as a strategy to increase the biomass for generation of new cells (Bensinger et al., 2008; Kidani et al., 2013). In parallel the increased availability of cholesterol in plasma membrane favors TCR clustering, thus enhancing T cell signalling, and improves effector CD8 antitumor function (Yang et al., 2016). By contrast, in thymocytes, lipid raft content, that is crucial for cell responsiveness to apoptosis-inducing signals for elimination of autoreactive T cells, is modulated by LXR activation (Chan et al., 2020).
Also, the function of B lymphocytes is regulated at the metabolic level. These cells have been involved in the production of antibodies to both naïve and oxidation-specific epitopes (OSEs) on LDL particles that might attenuate disease development although other B cells subsets might play a pro-atherogenic role (Ait-Oufella et al., 2010; Binder et al., 2003; Caligiuri et al., 2002; Major et al., 2002). Similar to T cells, BCR engagement reprograms B cell metabolism by activation of the nutrient sensor mTOR that in turn is crucial for B cell development, differentiation, and function (Iwata et al., 2017). After antigen stimulation and B cell activation, increased glucose uptake results in the upregulation of OXPHOS and TCA cycle; under this condition, TCA is supported by anapleurotic fuels, such as glutamine, while pyruvate, rather than being metabolized to lactate, is converted into acetyl-CoA for biomass synthesis (Waters et al., 2018). Also, cholesterol biosynthetic pathway is increased upon cell activation (including co-stimulatory interaction between CD40–CD40L), and is required for cell growth, organelle biogenesis, and cellular replication (Shimabukuro-Vornhagen et al., 2014). Bioproducts of cholesterol metabolism, such as the oxysterols 7α,25-HC and 7α,27-HC, are highly expressed in B cells and act as ligands of the orphan receptor EBI2, thus affecting B cell migration to outer follicles in lymphoid organs after antigen stimulation (Kelly et al., 2011).
3. Concluding remarks: therapeutic opportunities
Several lines of evidence indicate that during atherosclerosis cellular fate is controlled by metabolic reprogramming.
Interestingly, different cell types utilize specific metabolic pathways to adapt to the new milieu and/or to accommodate for the metabolic requests to fulfil their activities under resting or activated conditions. These adaptations play a key role in directing the interaction of cells at vascular-immune interface and allow to identify selective “metabolic checkpoints” that could be exploited to control cellular activation. In this scenario, “metabolic drugs” would be beneficial in specific pathological settings by modulating cellular metabolic responses, beyond ameliorating plasma biomarker profile.
3.1. Pharmacological modulation of lipid metabolism
Statins inhibit the enzyme β-hydroxy β-methylglutaryl-coenzyme A (HMG-CoA) reductase, the rate-limiting step for cholesterol synthesis and promote the expression of the LDL-R in the liver, thus favouring LDL-C reduction and cardiovascular protection (Pirillo et al., 2020). Large clinical trials have demonstrated a cardiovascular benefit that goes beyond LDL-C reduction and could be attributed to their immunomodulatory functions via controlling cholesterol availability, as suggested by the reduction in markers of systemic inflammation such as CRP (Catapano et al., 2017). In line with this observation, statin treatment has been reported to reduce disease activity and inflammation in subjects with multiple sclerosis and systemic lupus erythematosus (Chataway et al., 2014; Ulivieri and Baldari, 2014), while more recently, were shonw to decrease the inflammatory response and lower the risk of death in Covid-19 patients during hospitalization, further pointing to an immunomodulatory role of these molecules (Zhang et al., 2020). However, it's still debated whether the anti-inflammatory potential of statin is mediated at cellular level (ECs, VSMCs, or/and immune cells) or rather depends on systemic reduction of LDL-C levels; the latter is supported by the evidence that the dramatic drop in plasma cholesterol levels mediated by anti-PCSK9 antibodies is associated with a lower inflammatory phenotype of circulating monocytes (Bernelot Moens et al., 2017). However, numerous in vitro studies have demonstrated additional effects of statins exerted at cellular level. Statins treatment enhances endothelial function and NO availability, reduces the expression of NADH oxidase, thus limiting the production of ROS (Wassmann et al., 2002), and dampens pro-inflammatory cytokine and metalloproteinase expression in monocytes and macrophages (Rezaie-Majd et al., 2002). Statins reduce the polarization of macrophage toward M1 phenotype and counteract the trained immunity phenotype induced by oxLDL treatment in primary human monocytes by epigenetic reprogramming (Bekkering et al., 2018; Riksen and Netea, 2020). Similarly, in lymphocytes, inhibition of cholesterol synthesis, by dampening the activation of Activator Protein (AP-)1 and NF-kb expression (Cheng et al., 2010), reduces cellular proliferation and cytokine production and promotes Treg differentiation (Jameel et al., 2013; Kagami et al., 2009).
Bempedoic acid is a recently approved hypocholesterolemic pro-drug that, after being activated in the liver, inhibits ATP-citrate lyase (ACLY), an enzyme upstream of HMG-CoA reductase in the cholesterol biosynthetic pathway, thus resulting in the reduction of cholesterol plasma levels as a consequence of increased LDL-R expression and lipoprotein uptake (Laufs et al., 2019). Similar to statins, treatment with bempedoic acid reduces CRP levels (Laufs et al., 2019; Ruscica et al., 2020), thus suggesting that the immunomodulatory effect could depend on the inhibition of cholesterol biosynthesis in the liver and the reduction of lipoprotein burden. ACLY has been recently shown to be critical also in contributing the acetyl-CoA pool required for histone acetylation and the regulation of TLR-driven gene expression after LPS stimulation (Lauterbach et al., 2019; Williams and O'Neill, 2020). This implies that approaches able to target ACLY specifically in macrophages in the atherosclerotic plaque could reduce their activation. At present this is not possible with bempedoic acid, as its conversion to the active compound only occurs in the liver, but support the need for investigating novel ACLY inhibitors with a different tissue/cell selectivity.
Fibrates are selective agonists of PPAR-α, a transcription factor highly expressed in liver and skeletal muscle, that, by promoting FAO and inducing lipoprotein lipase and apoA-I expression, regulate energy homeostasis and lipoprotein metabolism (Duval et al., 2007); this results in the reduction of plasma triglyceride levels. The cardiovascular benefit derived from PPAR-α agonism has also been associated to their anti-inflammatory properties, exerted by repressing STAT, AP-1, NF-κB, and NFAT expression and directly inducing the expression of IL-1 receptor antagonist (Mandard and Patsouris, 2013). The main anti-inflammatory effect is mediated in the context of liver by directly targeting hepatocytes and Kupffer cells (Stienstra et al., 2007), endothelial cells (Mandard and Patsouris, 2013) and T cells by inhibiting IFN-γ and promoting IL-4 secretion (Lovett-Racke et al., 2004; Marx et al., 2004).
3.2. Pharmacological modulation of glucose metabolism
Metformin is the first-line medication for the treatment of diabetes. Acting via both AMP-activated protein kinase (AMPK)-dependent and AMPK-independent mechanisms, metformin reduces hepatic glucose production and increases the sensitivity to insulin of peripheral tissues, thus contributing to normalize plasma glucose levels (Rena et al., 2017). AMPK-depend inhibition of mTORC1 also explains the benefit in the context of cancer and cardiovascular diseases as well as the immunomodulatory effect (Howell et al., 2017). Metformin inhibits monocyte to macrophage differentiation and restrains the inflammatory response of activated macrophages; reduces NETosis formation by neutrophils and increases differentiation of T cells into both regulatory and memory T cells (Schuiveling et al., 2018). Treatment with metformin increases the resistance to many Gram negative and Gram positive pathogens, and contribute to reduce auto-immune disease burden in humans. Metformin treatment improves insulin resistance both in endothelial cells, resulting in an increase vasodilation (Nafisa et al., 2018), and in VSMCs, thus controlling proliferation, migration and inflammation (Lu et al., 2013) by PGC-1β upregulation (Guo et al., 2013).
Thiazolidinediones are PPAR-γ agonists regulating the expression of several genes involved in the regulation of glucose, lipid and protein metabolism with the net effect of improving the response to insulin in skeletal muscle and adipose tissue (increasing glucose uptake) and in the liver (decreasing hepatic glucose production). PPAR-γ agonism has been associated with anti-inflammatory effects in other pathological conditions, including inflammatory bowel disease, CNS inflammation, and LPS-induced cardiac and pulmonary inflammation (Mandard and Patsouris, 2013). In adipose tissue, PPAR-γ activation favours the alternative activation of macrophages (Odegaard et al., 2007) promoting lipid metabolism (Kratz et al., 2014). PPAR-γ expression by visceral AT-resident Treg cells is instrumental for their ability to control the inflammatory state of AT and thereby insulin sensitivity (Cipolletta et al., 2012).
PFKFB3 inhibitors reduce anaerobic glycolysis by blocking the 6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 enzyme. This protein is highly expressed by tumour cells, explaining the interest of PFKB3 inhibitors for cancer therapy (Lu et al., 2017), but also in endothelial cells. Indeed, inhibition of PFKB3 reduces endothelial cell proliferation and angiogenesis (Cantelmo et al., 2016; Schoors et al., 2014), but plays also an anti-inflammatory effect by limiting NLRP3 activation (Finucane et al., 2019).
Translating experimental pharmacology to the clinic will imply that these drugs should target vascular cells and/or immune cells in vivo to a reasonable concentration without heading directly to the liver (primary site of action) or exerting side effects due to the elevated levels required to target peripheral cells. This suggests that pharmacology should explore novel strategies to vehiculate metabolic drugs selectively at cellular level, thus helping in improving disease outcome and control immunometabolic responses in the context of cardiovascular diseases. In this scenario, the use of nanoparticle-based drug delivery systems, via conjugation with immunotherapeutic agents will improve the pharmacokinetic profile and facilitate the delivery of multiple therapeutic’ combinations at cellular level. Recently, advanced nanoparticle-based drug delivery systems were successfully tested in cancer immunotherapy and show to reduce toxic side effects and immune-related adverse events, as well as maximize the pharmacological benefit (Lim et al., 2019). This suggests that also repurposing nanoparticles as delivery system of immune/metabolic-therapeutic agents to cells at the vascular/immune interphase would offer an additional pharmacological approach to cardiovascular diseases.
Acknowledgements
Authors are supported by Fondazione Cariplo (Italy) [grant number 2019-1560 to FB and 2016-0852 to GDN]; Telethon Foundation (Italy) [grant number GGP19146 to GDN], Progetti di Rilevante Interesse Nazionale (Italy) [grant number PRIN 2017H5F943 to ALC and PRIN 2017K55HLC to GDN]; ERANET (European Union) [grant number ER-2017-2364981 to ALC].
References
- Ait-Oufella H., Herbin O., Bouaziz J.D., Binder C.J., Uyttenhove C., Laurans L., Taleb S., Van Vre E., Esposito B., Vilar J., Sirvent J., Van Snick J., Tedgui A., Tedder T.F., Mallat Z. B cell depletion reduces the development of atherosclerosis in mice. J. Exp. Med. 2010;207(8):1579–1587. doi: 10.1084/jem.20100155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amedei A., Morbidelli L. Circulating metabolites originating from gut microbiota control endothelial cell function. Molecules. 2019;24(21) doi: 10.3390/molecules24213992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ammirati E., Cianflone D., Vecchio V., Banfi M., Vermi A.C., De Metrio M., Grigore L., Pellegatta F., Pirillo A., Garlaschelli K., Manfredi A.A., Catapano A.L., Maseri A., Palini A.G., Norata G.D. Effector memory T cells are associated with atherosclerosis in humans and animal models. J Am Heart Assoc. 2012;1(1):27–41. doi: 10.1161/JAHA.111.000125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagi Z., Frangos J.A., Yeh J.C., White C.R., Kaley G., Koller A. PECAM-1 mediates NO-dependent dilation of arterioles to high temporal gradients of shear stress. Arterioscler. Thromb. Vasc. Biol. 2005;25(8):1590–1595. doi: 10.1161/01.ATV.0000170136.71970.5f. [DOI] [PubMed] [Google Scholar]
- Barron J.T., Parrillo J.E. Production of lactic acid and energy metabolism in vascular smooth muscle: effect of dichloroacetate. Am. J. Physiol. 1995;268(2 Pt 2):H713–H719. doi: 10.1152/ajpheart.1995.268.2.H713. [DOI] [PubMed] [Google Scholar]
- Bekkering S., Arts R.J.W., Novakovic B., Kourtzelis I., van der Heijden C., Li Y., Popa C.D., Ter Horst R., van Tuijl J., Netea-Maier R.T., van de Veerdonk F.L., Chavakis T., Joosten L.A.B., van der Meer J.W.M., Stunnenberg H., Riksen N.P., Netea M.G. Metabolic induction of trained immunity through the mevalonate pathway. Cell. 2018;172(1–2):135–146. doi: 10.1016/j.cell.2017.11.025. e139. [DOI] [PubMed] [Google Scholar]
- Bensinger S.J., Bradley M.N., Joseph S.B., Zelcer N., Janssen E.M., Hausner M.A., Shih R., Parks J.S., Edwards P.A., Jamieson B.D., Tontonoz P. LXR signaling couples sterol metabolism to proliferation in the acquired immune response. Cell. 2008;134(1):97–111. doi: 10.1016/j.cell.2008.04.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernelot Moens S.J., Neele A.E., Kroon J., van der Valk F.M., Van den Bossche J., Hoeksema M.A., Hoogeveen R.M., Schnitzler J.G., Baccara-Dinet M.T., Manvelian G., de Winther M.P.J., Stroes E.S.G. PCSK9 monoclonal antibodies reverse the pro-inflammatory profile of monocytes in familial hypercholesterolaemia. Eur. Heart J. 2017;38(20):1584–1593. doi: 10.1093/eurheartj/ehx002. [DOI] [PubMed] [Google Scholar]
- Binder C.J., Horkko S., Dewan A., Chang M.K., Kieu E.P., Goodyear C.S., Shaw P.X., Palinski W., Witztum J.L., Silverman G.J. Pneumococcal vaccination decreases atherosclerotic lesion formation: molecular mimicry between Streptococcus pneumoniae and oxidized LDL. Nat. Med. 2003;9(6):736–743. doi: 10.1038/nm876. [DOI] [PubMed] [Google Scholar]
- Boini K.M., Hussain T., Li P.L., Koka S. Trimethylamine-N-Oxide instigates NLRP3 inflammasome activation and endothelial dysfunction. Cell. Physiol. Biochem. 2017;44(1):152–162. doi: 10.1159/000484623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonacina F., Coe D., Wang G., Longhi M.P., Baragetti A., Moregola A., Garlaschelli K., Uboldi P., Pellegatta F., Grigore L., Da Dalt L., Annoni A., Gregori S., Xiao Q., Caruso D., Mitro N., Catapano A.L., Marelli-Berg F.M., Norata G.D. Myeloid apolipoprotein E controls dendritic cell antigen presentation and T cell activation. Nat. Commun. 2018;9(1):3083. doi: 10.1038/s41467-018-05322-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandes R.P., Weissmann N., Schroder K. Nox family NADPH oxidases in mechano-transduction: mechanisms and consequences. Antioxidants Redox Signal. 2014;20(6):887–898. doi: 10.1089/ars.2013.5414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breton-Romero R., Acin-Perez R., Rodriguez-Pascual F., Martinez-Molledo M., Brandes R.P., Rial E., Enriquez J.A., Lamas S. Laminar shear stress regulates mitochondrial dynamics, bioenergetics responses and PRX3 activation in endothelial cells. Biochim. Biophys. Acta. 2014;1843(11):2403–2413. doi: 10.1016/j.bbamcr.2014.07.003. [DOI] [PubMed] [Google Scholar]
- Cahill P.A., Redmond E.M. Vascular endothelium - gatekeeper of vessel health. Atherosclerosis. 2016;248:97–109. doi: 10.1016/j.atherosclerosis.2016.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caligiuri G., Nicoletti A., Poirier B., Hansson G.K. Protective immunity against atherosclerosis carried by B cells of hypercholesterolemic mice. J. Clin. Invest. 2002;109(6):745–753. doi: 10.1172/JCI07272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callegari E., Norata G.D., Inoue H., Catapano A.L. Oxidized-HDL3 modulates the expression of Cox-2 in human endothelial cells. Int. J. Mol. Med. 2006;18(1):209–213. [PubMed] [Google Scholar]
- Cantelmo A.R., Conradi L.C., Brajic A., Goveia J., Kalucka J., Pircher A., Chaturvedi P., Hol J., Thienpont B., Teuwen L.A., Schoors S., Boeckx B., Vriens J., Kuchnio A., Veys K., Cruys B., Finotto L., Treps L., Stav-Noraas T.E., Bifari F., Stapor P., Decimo I., Kampen K., De Bock K., Haraldsen G., Schoonjans L., Rabelink T., Eelen G., Ghesquiere B., Rehman J., Lambrechts D., Malik A.B., Dewerchin M., Carmeliet P. Inhibition of the glycolytic activator PFKFB3 in endothelium induces tumor vessel normalization, impairs metastasis, and improves chemotherapy. Canc. Cell. 2016;30(6):968–985. doi: 10.1016/j.ccell.2016.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catapano A.L., Pirillo A., Norata G.D. Vascular inflammation and low-density lipoproteins: is cholesterol the link? A lesson from the clinical trials. Br. J. Pharmacol. 2017;174(22):3973–3985. doi: 10.1111/bph.13805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cepinskas G., Savickiene J., Ionescu C.V., Kvietys P.R. PMN transendothelial migration decreases nuclear NFkappaB in IL-1beta-activated endothelial cells: role of PECAM-1. J. Cell Biol. 2003;161(3):641–651. doi: 10.1083/jcb.200212048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cham C.M., Driessens G., O'Keefe J.P., Gajewski T.F. Glucose deprivation inhibits multiple key gene expression events and effector functions in CD8+ T cells. Eur. J. Immunol. 2008;38(9):2438–2450. doi: 10.1002/eji.200838289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cham C.M., Gajewski T.F. Glucose availability regulates IFN-gamma production and p70S6 kinase activation in CD8+ effector T cells. J. Immunol. 2005;174(8):4670–4677. doi: 10.4049/jimmunol.174.8.4670. [DOI] [PubMed] [Google Scholar]
- Chan C.T., Fenn A.M., Harder N.K., Mindur J.E., McAlpine C.S., Patel J., Valet C., Rattik S., Iwamoto Y., He S., Anzai A., Kahles F., Poller W.C., Janssen H., Wong L.P., Fernandez-Hernando C., Koolbergen D.R., van der Laan A.M., Yvan-Charvet L., Sadreyev R.I., Nahrendorf M., Westerterp M., Tall A.R., Gustafsson J.A., Swirski F.K. Liver X receptors are required for thymic resilience and T cell output. J. Exp. Med. 2020;217(10) doi: 10.1084/jem.20200318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandler K.B., Leon D.R., Kuang J., Meyer R.D., Rahimi N., Costello C.E. N-Glycosylation regulates ligand-dependent activation and signaling of vascular endothelial growth factor receptor 2 (VEGFR2) J. Biol. Chem. 2019;294(35):13117–13130. doi: 10.1074/jbc.RA119.008643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chataway J., Schuerer N., Alsanousi A., Chan D., MacManus D., Hunter K., Anderson V., Bangham C.R., Clegg S., Nielsen C., Fox N.C., Wilkie D., Nicholas J.M., Calder V.L., Greenwood J., Frost C., Nicholas R. Effect of high-dose simvastatin on brain atrophy and disability in secondary progressive multiple sclerosis (MS-STAT): a randomised, placebo-controlled, phase 2 trial. Lancet. 2014;383(9936):2213–2221. doi: 10.1016/S0140-6736(13)62242-4. [DOI] [PubMed] [Google Scholar]
- Cheng S.M., Lai J.H., Yang S.P., Tsao T.P., Ho L.J., Liou J.T., Cheng C.C. Modulation of human T cells signaling transduction by lovastatin. Int. J. Cardiol. 2010;140(1):24–33. doi: 10.1016/j.ijcard.2008.10.044. [DOI] [PubMed] [Google Scholar]
- Cheung K., Ma L., Wang G., Coe D., Ferro R., Falasca M., Buckley C.D., Mauro C., Marelli-Berg F.M. CD31 signals confer immune privilege to the vascular endothelium. Proc. Natl. Acad. Sci. U.S.A. 2015;112(43):E5815–E5824. doi: 10.1073/pnas.1509627112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christ A., Gunther P., Lauterbach M.A.R., Duewell P., Biswas D., Pelka K., Scholz C.J., Oosting M., Haendler K., Bassler K., Klee K., Schulte-Schrepping J., Ulas T., Moorlag S., Kumar V., Park M.H., Joosten L.A.B., Groh L.A., Riksen N.P., Espevik T., Schlitzer A., Li Y., Fitzgerald M.L., Netea M.G., Schultze J.L., Latz E. Western diet triggers NLRP3-dependent innate immune reprogramming. Cell. 2018;172(1–2):162–175. doi: 10.1016/j.cell.2017.12.013. e114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cipolletta D., Feuerer M., Li A., Kamei N., Lee J., Shoelson S.E., Benoist C., Mathis D. PPAR-gamma is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature. 2012;486(7404):549–553. doi: 10.1038/nature11132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couty J.P., Rampon C., Leveque M., Laran-Chich M.P., Bourdoulous S., Greenwood J., Couraud P.O. PECAM-1 engagement counteracts ICAM-1-induced signaling in brain vascular endothelial cells. J. Neurochem. 2007;103(2):793–801. doi: 10.1111/j.1471-4159.2007.04782.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Zeeuw P., Wong B.W., Carmeliet P. Metabolic adaptations in diabetic endothelial cells. Circ. J. 2015;79(5):934–941. doi: 10.1253/circj.CJ-15-0230. [DOI] [PubMed] [Google Scholar]
- Deaton R.A., Gan Q., Owens G.K. Sp1-dependent activation of KLF4 is required for PDGF-BB-induced phenotypic modulation of smooth muscle. Am. J. Physiol. Heart Circ. Physiol. 2009;296(4):H1027–H1037. doi: 10.1152/ajpheart.01230.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dekker R.J., Boon R.A., Rondaij M.G., Kragt A., Volger O.L., Elderkamp Y.W., Meijers J.C., Voorberg J., Pannekoek H., Horrevoets A.J. KLF2 provokes a gene expression pattern that establishes functional quiescent differentiation of the endothelium. Blood. 2006;107(11):4354–4363. doi: 10.1182/blood-2005-08-3465. [DOI] [PubMed] [Google Scholar]
- Doddaballapur A., Michalik K.M., Manavski Y., Lucas T., Houtkooper R.H., You X., Chen W., Zeiher A.M., Potente M., Dimmeler S., Boon R.A. Laminar shear stress inhibits endothelial cell metabolism via KLF2-mediated repression of PFKFB3. Arterioscler. Thromb. Vasc. Biol. 2015;35(1):137–145. doi: 10.1161/ATVBAHA.114.304277. [DOI] [PubMed] [Google Scholar]
- Drechsler M., Megens R.T., van Zandvoort M., Weber C., Soehnlein O. Hyperlipidemia-triggered neutrophilia promotes early atherosclerosis. Circulation. 2010;122(18):1837–1845. doi: 10.1161/CIRCULATIONAHA.110.961714. [DOI] [PubMed] [Google Scholar]
- Duval C., Muller M., Kersten S. PPARalpha and dyslipidemia. Biochim. Biophys. Acta. 2007;1771(8):961–971. doi: 10.1016/j.bbalip.2007.05.003. [DOI] [PubMed] [Google Scholar]
- Eelen G., de Zeeuw P., Treps L., Harjes U., Wong B.W., Carmeliet P. Endothelial cell metabolism. Physiol. Rev. 2018;98(1):3–58. doi: 10.1152/physrev.00001.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finucane O.M., Sugrue J., Rubio-Araiz A., Guillot-Sestier M.V., Lynch M.A. The NLRP3 inflammasome modulates glycolysis by increasing PFKFB3 in an IL-1beta-dependent manner in macrophages. Sci. Rep. 2019;9(1):4034. doi: 10.1038/s41598-019-40619-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flynn K.M., Michaud M., Canosa S., Madri J.A. CD44 regulates vascular endothelial barrier integrity via a PECAM-1 dependent mechanism. Angiogenesis. 2013;16(3):689–705. doi: 10.1007/s10456-013-9346-9. [DOI] [PubMed] [Google Scholar]
- Flynn M.C., Pernes G., Lee M.K.S., Nagareddy P.R., Murphy A.J. Monocytes, macrophages, and metabolic disease in atherosclerosis. Front. Pharmacol. 2019;10:666. doi: 10.3389/fphar.2019.00666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folco E.J., Sukhova G.K., Quillard T., Libby P. Moderate hypoxia potentiates interleukin-1beta production in activated human macrophages. Circ. Res. 2014;115(10):875–883. doi: 10.1161/CIRCRESAHA.115.304437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galbiati F., Razani B., Lisanti M.P. Emerging themes in lipid rafts and caveolae. Cell. 2001;106(4):403–411. doi: 10.1016/s0092-8674(01)00472-x. [DOI] [PubMed] [Google Scholar]
- Gao C., Sun W., Christofidou-Solomidou M., Sawada M., Newman D.K., Bergom C., Albelda S.M., Matsuyama S., Newman P.J. PECAM-1 functions as a specific and potent inhibitor of mitochondrial-dependent apoptosis. Blood. 2003;102(1):169–179. doi: 10.1182/blood-2003-01-0003. [DOI] [PubMed] [Google Scholar]
- Gimbrone M.A., Jr., Garcia-Cardena G. Endothelial cell dysfunction and the pathobiology of atherosclerosis. Circ. Res. 2016;118(4):620–636. doi: 10.1161/CIRCRESAHA.115.306301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomaraschi M., Bonacina F., Norata G.D. Lysosomal acid lipase: from cellular lipid handler to immunometabolic target. Trends Pharmacol. Sci. 2019;40(2):104–115. doi: 10.1016/j.tips.2018.12.006. [DOI] [PubMed] [Google Scholar]
- Guo Y., Fan Y., Zhang J., Chang L., Lin J.D., Chen Y.E. Peroxisome proliferator-activated receptor gamma coactivator 1beta (PGC-1beta) protein attenuates vascular lesion formation by inhibition of chromatin loading of minichromosome maintenance complex in smooth muscle cells. J. Biol. Chem. 2013;288(7):4625–4636. doi: 10.1074/jbc.M112.407452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howell J.J., Hellberg K., Turner M., Talbott G., Kolar M.J., Ross D.S., Hoxhaj G., Saghatelian A., Shaw R.J., Manning B.D. Metformin inhibits hepatic mTORC1 signaling via dose-dependent mechanisms involving AMPK and the TSC complex. Cell Metabol. 2017;25(2):463–471. doi: 10.1016/j.cmet.2016.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imamaki R., Ogawa K., Kizuka Y., Komi Y., Kojima S., Kotani N., Honke K., Honda T., Taniguchi N., Kitazume S. Glycosylation controls cooperative PECAM-VEGFR2-beta3 integrin functions at the endothelial surface for tumor angiogenesis. Oncogene. 2018;37(31):4287–4299. doi: 10.1038/s41388-018-0271-7. [DOI] [PubMed] [Google Scholar]
- Ito A., Hong C., Oka K., Salazar J.V., Diehl C., Witztum J.L., Diaz M., Castrillo A., Bensinger S.J., Chan L., Tontonoz P. Cholesterol accumulation in CD11c(+) immune cells is a causal and targetable factor in autoimmune disease. Immunity. 2016;45(6):1311–1326. doi: 10.1016/j.immuni.2016.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwata T.N., Ramirez-Komo J.A., Park H., Iritani B.M. Control of B lymphocyte development and functions by the mTOR signaling pathways. Cytokine Growth Factor Rev. 2017;35:47–62. doi: 10.1016/j.cytogfr.2017.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jameel A., Ooi K.G., Jeffs N.R., Galatowicz G., Lightman S.L., Calder V.L. Statin modulation of human T-cell proliferation, IL-1beta and IL-17 production, and IFN-gamma T cell expression: synergy with conventional immunosuppressive agents. Int. J. Inflamm. 2013:434586. doi: 10.1155/2013/434586. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji G., O'Brien C.D., Feldman M., Manevich Y., Lim P., Sun J., Albelda S.M., Kotlikoff M.I. PECAM-1 (CD31) regulates a hydrogen peroxide-activated nonselective cation channel in endothelial cells. J. Cell Biol. 2002;157(1):173–184. doi: 10.1083/jcb.200110056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kagami S., Owada T., Kanari H., Saito Y., Suto A., Ikeda K., Hirose K., Watanabe N., Iwamoto I., Nakajima H. Protein geranylgeranylation regulates the balance between Th17 cells and Foxp3+ regulatory T cells. Int. Immunol. 2009;21(6):679–689. doi: 10.1093/intimm/dxp037. [DOI] [PubMed] [Google Scholar]
- Kalucka J., de Rooij L., Goveia J., Rohlenova K., Dumas S.J., Meta E., Conchinha N.V., Taverna F., Teuwen L.A., Veys K., Garcia-Caballero M., Khan S., Geldhof V., Sokol L., Chen R., Treps L., Borri M., de Zeeuw P., Dubois C., Karakach T.K., Falkenberg K.D., Parys M., Yin X., Vinckier S., Du Y., Fenton R.A., Schoonjans L., Dewerchin M., Eelen G., Thienpont B., Lin L., Bolund L., Li X., Luo Y., Carmeliet P. Single-cell transcriptome atlas of murine endothelial cells. Cell. 2020;180(4):764–779. doi: 10.1016/j.cell.2020.01.015. e720. [DOI] [PubMed] [Google Scholar]
- Kelly L.M., Pereira J.P., Yi T., Xu Y., Cyster J.G. EBI2 guides serial movements of activated B cells and ligand activity is detectable in lymphoid and nonlymphoid tissues. J. Immunol. 2011;187(6):3026–3032. doi: 10.4049/jimmunol.1101262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidani Y., Elsaesser H., Hock M.B., Vergnes L., Williams K.J., Argus J.P., Marbois B.N., Komisopoulou E., Wilson E.B., Osborne T.F., Graeber T.G., Reue K., Brooks D.G., Bensinger S.J. Sterol regulatory element-binding proteins are essential for the metabolic programming of effector T cells and adaptive immunity. Nat. Immunol. 2013;14(5):489–499. doi: 10.1038/ni.2570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kluge M.A., Fetterman J.L., Vita J.A. Mitochondria and endothelial function. Circ. Res. 2013;112(8):1171–1188. doi: 10.1161/CIRCRESAHA.111.300233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koo A., Dewey C.F., Jr., Garcia-Cardena G. Hemodynamic shear stress characteristic of atherosclerosis-resistant regions promotes glycocalyx formation in cultured endothelial cells. Am. J. Physiol. Cell Physiol. 2013;304(2):C137–C146. doi: 10.1152/ajpcell.00187.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovanen P.T., Bot I. Mast cells in atherosclerotic cardiovascular disease - activators and actions. Eur. J. Pharmacol. 2017;816:37–46. doi: 10.1016/j.ejphar.2017.10.013. [DOI] [PubMed] [Google Scholar]
- Kratz M., Coats B.R., Hisert K.B., Hagman D., Mutskov V., Peris E., Schoenfelt K.Q., Kuzma J.N., Larson I., Billing P.S., Landerholm R.W., Crouthamel M., Gozal D., Hwang S., Singh P.K., Becker L. Metabolic dysfunction drives a mechanistically distinct proinflammatory phenotype in adipose tissue macrophages. Cell Metabol. 2014;20(4):614–625. doi: 10.1016/j.cmet.2014.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kratzer A., Giral H., Landmesser U. High-density lipoproteins as modulators of endothelial cell functions: alterations in patients with coronary artery disease. Cardiovasc. Res. 2014;103(3):350–361. doi: 10.1093/cvr/cvu139. [DOI] [PubMed] [Google Scholar]
- Kuznetsov A.V., Hermann M., Saks V., Hengster P., Margreiter R. The cell-type specificity of mitochondrial dynamics. Int. J. Biochem. Cell Biol. 2009;41(10):1928–1939. doi: 10.1016/j.biocel.2009.03.007. [DOI] [PubMed] [Google Scholar]
- Kzhyshkowska J., Neyen C., Gordon S. Role of macrophage scavenger receptors in atherosclerosis. Immunobiology. 2012;217(5):492–502. doi: 10.1016/j.imbio.2012.02.015. [DOI] [PubMed] [Google Scholar]
- Laufs U., Banach M., Mancini G.B.J., Gaudet D., Bloedon L.T., Sterling L.R., Kelly S., Stroes E.S.G. Efficacy and safety of bempedoic acid in patients with hypercholesterolemia and statin intolerance. J Am Heart Assoc. 2019;8(7) doi: 10.1161/JAHA.118.011662. e011662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauterbach M.A., Hanke J.E., Serefidou M., Mangan M.S.J., Kolbe C.C., Hess T., Rothe M., Kaiser R., Hoss F., Gehlen J., Engels G., Kreutzenbeck M., Schmidt S.V., Christ A., Imhof A., Hiller K., Latz E. Toll-like receptor signaling rewires macrophage metabolism and promotes histone acetylation via ATP-citrate lyase. Immunity. 2019;51(6):997–1011. doi: 10.1016/j.immuni.2019.11.009. e1017. [DOI] [PubMed] [Google Scholar]
- Lee J.H., Phelan P., Shin M., Oh B.C., Han X., Im S.S., Osborne T.F. SREBP-1a-stimulated lipid synthesis is required for macrophage phagocytosis downstream of TLR4-directed mTORC1. Proc. Natl. Acad. Sci. U.S.A. 2018;115(52):E12228–E12234. doi: 10.1073/pnas.1813458115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim S., Park J., Shim M.K., Um W., Yoon H.Y., Ryu J.H., Lim D.K., Kim K. Recent advances and challenges of repurposing nanoparticle-based drug delivery systems to enhance cancer immunotherapy. Theranostics. 2019;9(25):7906–7923. doi: 10.7150/thno.38425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J., Garcia-Cardena G., Sessa W.C. Palmitoylation of endothelial nitric oxide synthase is necessary for optimal stimulated release of nitric oxide: implications for caveolae localization. Biochemistry. 1996;35(41):13277–13281. doi: 10.1021/bi961720e. [DOI] [PubMed] [Google Scholar]
- Liu K., Fang C., Shen Y., Liu Z., Zhang M., Ma B., Pang X. Hypoxia-inducible factor 1a induces phenotype switch of human aortic vascular smooth muscle cell through PI3K/AKT/AEG-1 signaling. Oncotarget. 2017;8(20):33343–33352. doi: 10.18632/oncotarget.16448. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Lovett-Racke A.E., Hussain R.Z., Northrop S., Choy J., Rocchini A., Matthes L., Chavis J.A., Diab A., Drew P.D., Racke M.K. Peroxisome proliferator-activated receptor alpha agonists as therapy for autoimmune disease. J. Immunol. 2004;172(9):5790–5798. doi: 10.4049/jimmunol.172.9.5790. [DOI] [PubMed] [Google Scholar]
- Lu J., Ji J., Meng H., Wang D., Jiang B., Liu L., Randell E., Adeli K., Meng Q.H. The protective effect and underlying mechanism of metformin on neointima formation in fructose-induced insulin resistant rats. Cardiovasc. Diabetol. 2013;12:58. doi: 10.1186/1475-2840-12-58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu L., Chen Y., Zhu Y. The molecular basis of targeting PFKFB3 as a therapeutic strategy against cancer. Oncotarget. 2017;8(37):62793–62802. doi: 10.18632/oncotarget.19513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lunt S.Y., Vander Heiden M.G. Aerobic glycolysis: meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011;27:441–464. doi: 10.1146/annurev-cellbio-092910-154237. [DOI] [PubMed] [Google Scholar]
- MacRitchie N., Grassia G., Noonan J., Cole J.E., Hughes C.E., Schroeder J., Benson R.A., Cochain C., Zernecke A., Guzik T.J., Garside P., Monaco C., Maffia P. The aorta can act as a site of naive CD4+ T-cell priming. Cardiovasc. Res. 2020;116(2):306–316. doi: 10.1093/cvr/cvz102. [DOI] [PubMed] [Google Scholar]
- Major A.S., Fazio S., Linton M.F. B-lymphocyte deficiency increases atherosclerosis in LDL receptor-null mice. Arterioscler. Thromb. Vasc. Biol. 2002;22(11):1892–1898. doi: 10.1161/01.atv.0000039169.47943.ee. [DOI] [PubMed] [Google Scholar]
- Mandard S., Patsouris D. Nuclear control of the inflammatory response in mammals by peroxisome proliferator-activated receptors. PPAR Res. 2013;2013:613864. doi: 10.1155/2013/613864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcus A.J., Broekman M.J., Weksler B.B., Jaffe E.A., Safier L.B., Ullman H.L., Islam N., Tack-Goldman K. Arachidonic acid metabolism in endothelial cells and platelets. Ann. N. Y. Acad. Sci. 1982;401:195–202. doi: 10.1111/j.1749-6632.1982.tb25718.x. [DOI] [PubMed] [Google Scholar]
- Marelli-Berg F.M., Fu H., Mauro C. Molecular mechanisms of metabolic reprogramming in proliferating cells: implications for T-cell-mediated immunity. Immunology. 2012;136(4):363–369. doi: 10.1111/j.1365-2567.2012.03583.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marx N., Duez H., Fruchart J.C., Staels B. Peroxisome proliferator-activated receptors and atherogenesis: regulators of gene expression in vascular cells. Circ. Res. 2004;94(9):1168–1178. doi: 10.1161/01.RES.0000127122.22685.0A. [DOI] [PubMed] [Google Scholar]
- Merchan J.R., Kovacs K., Railsback J.W., Kurtoglu M., Jing Y., Pina Y., Gao N., Murray T.G., Lehrman M.A., Lampidis T.J. Antiangiogenic activity of 2-deoxy-D-glucose. PloS One. 2010;5(10) doi: 10.1371/journal.pone.0013699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon J.S., Lee S., Park M.A., Siempos, Haslip M., Lee P.J., Yun M., Kim C.K., Howrylak J., Ryter S.W., Nakahira K., Choi A.M. UCP2-induced fatty acid synthase promotes NLRP3 inflammasome activation during sepsis. J. Clin. Invest. 2015;125(2):665–680. doi: 10.1172/JCI78253. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Moon J.S., Nakahira K., Chung K.P., DeNicola G.M., Koo M.J., Pabon M.A., Rooney K.T., Yoon J.H., Ryter S.W., Stout-Delgado H., Choi A.M. NOX4-dependent fatty acid oxidation promotes NLRP3 inflammasome activation in macrophages. Nat. Med. 2016;22(9):1002–1012. doi: 10.1038/nm.4153. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Mundi S., Massaro M., Scoditti E., Carluccio M.A., van Hinsbergh V.W.M., Iruela-Arispe M.L., De Caterina R. Endothelial permeability, LDL deposition, and cardiovascular risk factors-a review. Cardiovasc. Res. 2018;114(1):35–52. doi: 10.1093/cvr/cvx226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nafisa A., Gray S.G., Cao Y., Wang T., Xu S., Wattoo F.H., Barras M., Cohen N., Kamato D., Little P.J. Endothelial function and dysfunction: impact of metformin. Pharmacol. Ther. 2018;192:150–162. doi: 10.1016/j.pharmthera.2018.07.007. [DOI] [PubMed] [Google Scholar]
- Nagareddy P.R., Murphy A.J., Stirzaker R.A., Hu Y., Yu S., Miller R.G., Ramkhelawon B., Distel E., Westerterp M., Huang L.S., Schmidt A.M., Orchard T.J., Fisher E.A., Tall A.R., Goldberg I.J. Hyperglycemia promotes myelopoiesis and impairs the resolution of atherosclerosis. Cell Metabol. 2013;17(5):695–708. doi: 10.1016/j.cmet.2013.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norata G.D., Banfi C., Pirillo A., Tremoli E., Hamsten A., Catapano A.L., Eriksson P. Oxidised-HDL3 induces the expression of PAI-1 in human endothelial cells. Role of p38MAPK activation and mRNA stabilization. Br. J. Haematol. 2004;127(1):97–104. doi: 10.1111/j.1365-2141.2004.05163.x. [DOI] [PubMed] [Google Scholar]
- Norata G.D., Caligiuri G., Chavakis T., Matarese G., Netea M.G., Nicoletti A., O'Neill L.A., Marelli-Berg F.M. The cellular and molecular basis of translational immunometabolism. Immunity. 2015;43(3):421–434. doi: 10.1016/j.immuni.2015.08.023. [DOI] [PubMed] [Google Scholar]
- Norata G.D., Catapano A.L. Molecular mechanisms responsible for the antiinflammatory and protective effect of HDL on the endothelium. Vasc. Health Risk Manag. 2005;1(2):119–129. doi: 10.2147/vhrm.1.2.119.64083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norata G.D., Grigore L., Raselli S., Redaelli L., Hamsten A., Maggi F., Eriksson P., Catapano A.L. Post-prandial endothelial dysfunction in hypertriglyceridemic subjects: molecular mechanisms and gene expression studies. Atherosclerosis. 2007;193(2):321–327. doi: 10.1016/j.atherosclerosis.2006.09.015. [DOI] [PubMed] [Google Scholar]
- Norata G.D., Grigore L., Raselli S., Seccomandi P.M., Hamsten A., Maggi F.M., Eriksson P., Catapano A.L. Triglyceride-rich lipoproteins from hypertriglyceridemic subjects induce a pro-inflammatory response in the endothelium: molecular mechanisms and gene expression studies. J. Mol. Cell. Cardiol. 2006;40(4):484–494. doi: 10.1016/j.yjmcc.2006.01.022. [DOI] [PubMed] [Google Scholar]
- Norata G.D., Pinna C., Zappella F., Elia L., Sala A., Condorelli G., Catapano A.L. MicroRNA 143-145 deficiency impairs vascular function. Int. J. Immunopathol. Pharmacol. 2012;25(2):467–474. doi: 10.1177/039463201202500216. [DOI] [PubMed] [Google Scholar]
- Norata G.D., Raselli S., Grigore L., Garlaschelli K., Vianello D., Bertocco S., Zambon A., Catapano A.L. Small dense LDL and VLDL predict common carotid artery IMT and elicit an inflammatory response in peripheral blood mononuclear and endothelial cells. Atherosclerosis. 2009;206(2):556–562. doi: 10.1016/j.atherosclerosis.2009.03.017. [DOI] [PubMed] [Google Scholar]
- Norata G.D., Sala F., Catapano A.L., Fernandez-Hernando C. MicroRNAs and lipoproteins: a connection beyond atherosclerosis? Atherosclerosis. 2013;227(2):209–215. doi: 10.1016/j.atherosclerosis.2012.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Odegaard J.I., Ricardo-Gonzalez R.R., Goforth M.H., Morel C.R., Subramanian V., Mukundan L., Red Eagle A., Vats D., Brombacher F., Ferrante A.W., Chawla A. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature. 2007;447(7148):1116–1120. doi: 10.1038/nature05894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oishi Y., Spann N.J., Link V.M., Muse E.D., Strid T., Edillor C., Kolar M.J., Matsuzaka T., Hayakawa S., Tao J., Kaikkonen M.U., Carlin A.F., Lam M.T., Manabe I., Shimano H., Saghatelian A., Glass C.K. SREBP1 contributes to resolution of pro-inflammatory TLR4 signaling by reprogramming fatty acid metabolism. Cell Metabol. 2017;25(2):412–427. doi: 10.1016/j.cmet.2016.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens G.K., Kumar M.S., Wamhoff B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004;84(3):767–801. doi: 10.1152/physrev.00041.2003. [DOI] [PubMed] [Google Scholar]
- Park E.J., Grabinska K.A., Guan Z., Sessa W.C. NgBR is essential for endothelial cell glycosylation and vascular development. EMBO Rep. 2016;17(2):167–177. doi: 10.15252/embr.201540789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park S., DiMaio T.A., Scheef E.A., Sorenson C.M., Sheibani N. PECAM-1 regulates proangiogenic properties of endothelial cells through modulation of cell-cell and cell-matrix interactions. Am. J. Physiol. Cell Physiol. 2010;299(6):C1468–C1484. doi: 10.1152/ajpcell.00246.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez J., Hill B.G., Benavides G.A., Dranka B.P., Darley-Usmar V.M. Role of cellular bioenergetics in smooth muscle cell proliferation induced by platelet-derived growth factor. Biochem. J. 2010;428(2):255–267. doi: 10.1042/BJ20100090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pirillo A., Catapano A.L., Norata G.D. Biological consequences of dysfunctional HDL. Curr. Med. Chem. 2019;26(9):1644–1664. doi: 10.2174/0929867325666180530110543. [DOI] [PubMed] [Google Scholar]
- Pirillo A., Norata G.D., Catapano A.L. Handbook of Experimental Pharmacology. Springer; Berlin, Heidelberg: 2020. LDL-Cholesterol-Lowering Therapy. [DOI] [Google Scholar]
- Privratsky J.R., Newman P.J. PECAM-1: regulator of endothelial junctional integrity. Cell Tissue Res. 2014;355(3):607–619. doi: 10.1007/s00441-013-1779-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rena G., Hardie D.G., Pearson E.R. The mechanisms of action of metformin. Diabetologia. 2017;60(9):1577–1585. doi: 10.1007/s00125-017-4342-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renner K., Geiselhoringer A.L., Fante M., Bruss C., Farber S., Schonhammer G., Peter K., Singer K., Andreesen R., Hoffmann P., Oefner P., Herr W., Kreutz M. Metabolic plasticity of human T cells: preserved cytokine production under glucose deprivation or mitochondrial restriction, but 2-deoxy-glucose affects effector functions. Eur. J. Immunol. 2015;45(9):2504–2516. doi: 10.1002/eji.201545473. [DOI] [PubMed] [Google Scholar]
- Rezaie-Majd A., Maca T., Bucek R.A., Valent P., Muller M.R., Husslein P., Kashanipour A., Minar E., Baghestanian M. Simvastatin reduces expression of cytokines interleukin-6, interleukin-8, and monocyte chemoattractant protein-1 in circulating monocytes from hypercholesterolemic patients. Arterioscler. Thromb. Vasc. Biol. 2002;22(7):1194–1199. doi: 10.1161/01.atv.0000022694.16328.cc. [DOI] [PubMed] [Google Scholar]
- Riksen N.P., Netea M.G. Immunometabolic control of trained immunity. Mol. Aspect. Med. 2020:100897. doi: 10.1016/j.mam.2020.100897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruan G.X., Kazlauskas A. Lactate engages receptor tyrosine kinases Axl, Tie2, and vascular endothelial growth factor receptor 2 to activate phosphoinositide 3-kinase/Akt and promote angiogenesis. J. Biol. Chem. 2013;288(29):21161–21172. doi: 10.1074/jbc.M113.474619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruscica M., Corsini A., Ferri N., Banach M., Sirtori C.R. Clinical approach to the inflammatory etiology of cardiovascular diseases. Pharmacol. Res. 2020;159:104916. doi: 10.1016/j.phrs.2020.104916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell-Puleri S., Dela Paz N.G., Adams D., Chattopadhyay M., Cancel L., Ebong E., Orr A.W., Frangos J.A., Tarbell J.M. Fluid shear stress induces upregulation of COX-2 and PGI2 release in endothelial cells via a pathway involving PECAM-1, PI3K, FAK, and p38. Am. J. Physiol. Heart Circ. Physiol. 2017;312(3):H485–H500. doi: 10.1152/ajpheart.00035.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saragih H., Zilian E., Jaimes Y., Paine A., Figueiredo C., Eiz-Vesper B., Blasczyk R., Larmann J., Theilmeier G., Burg-Roderfeld M., Andrei-Selmer L.C., Becker J.U., Santoso S., Immenschuh S. PECAM-1-dependent heme oxygenase-1 regulation via an Nrf2-mediated pathway in endothelial cells. Thromb. Haemostasis. 2014;111(6):1077–1088. doi: 10.1160/TH13-11-0923. [DOI] [PubMed] [Google Scholar]
- Sathanoori R., Rosi F., Gu B.J., Wiley J.S., Muller C.E., Olde B., Erlinge D. Shear stress modulates endothelial KLF2 through activation of P2X4. Purinergic Signal. 2015;11(1):139–153. doi: 10.1007/s11302-014-9442-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato R., Okamoto A., Inoue J., Miyamoto W., Sakai Y., Emoto N., Shimano H., Maeda M. Transcriptional regulation of the ATP citrate-lyase gene by sterol regulatory element-binding proteins. J. Biol. Chem. 2000;275(17):12497–12502. doi: 10.1074/jbc.275.17.12497. [DOI] [PubMed] [Google Scholar]
- Schoors S., De Bock K., Cantelmo A.R., Georgiadou M., Ghesquiere B., Cauwenberghs S., Kuchnio A., Wong B.W., Quaegebeur A., Goveia J., Bifari F., Wang X., Blanco R., Tembuyser B., Cornelissen I., Bouche A., Vinckier S., Diaz-Moralli S., Gerhardt H., Telang S., Cascante M., Chesney J., Dewerchin M., Carmeliet P. Partial and transient reduction of glycolysis by PFKFB3 blockade reduces pathological angiogenesis. Cell Metabol. 2014;19(1):37–48. doi: 10.1016/j.cmet.2013.11.008. [DOI] [PubMed] [Google Scholar]
- Schuiveling M., Vazirpanah N., Radstake T., Zimmermann M., Broen J.C.A. Metformin, A new era for an old drug in the treatment of immune mediated disease? Curr. Drug Targets. 2018;19(8):945–959. doi: 10.2174/1389450118666170613081730. [DOI] [PubMed] [Google Scholar]
- Serhan C.N., Levy B.D. Resolvins in inflammation: emergence of the pro-resolving superfamily of mediators. J. Clin. Invest. 2018;128(7):2657–2669. doi: 10.1172/JCI97943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shankman L.S., Gomez D., Cherepanova O.A., Salmon M., Alencar G.F., Haskins R.M., Swiatlowska P., Newman A.A., Greene E.S., Straub A.C., Isakson B., Randolph G.J., Owens G.K. KLF4-dependent phenotypic modulation of smooth muscle cells has a key role in atherosclerotic plaque pathogenesis. Nat. Med. 2015;21(6):628–637. doi: 10.1038/nm.3866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheedy F.J., Grebe A., Rayner K.J., Kalantari P., Ramkhelawon B., Carpenter S.B., Becker C.E., Ediriweera H.N., Mullick A.E., Golenbock D.T., Stuart L.M., Latz E., Fitzgerald K.A., Moore K.J. CD36 coordinates NLRP3 inflammasome activation by facilitating intracellular nucleation of soluble ligands into particulate ligands in sterile inflammation. Nat. Immunol. 2013;14(8):812–820. doi: 10.1038/ni.2639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimabukuro-Vornhagen A., Zoghi S., Liebig T.M., Wennhold K., Chemitz J., Draube A., Kochanek M., Blaschke F., Pallasch C., Holtick U., Scheid C., Theurich S., Hallek M., von Bergwelt-Baildon M.S. Inhibition of protein geranylgeranylation specifically interferes with CD40-dependent B cell activation, resulting in a reduced capacity to induce T cell immunity. J. Immunol. 2014;193(10):5294–5305. doi: 10.4049/jimmunol.1203436. [DOI] [PubMed] [Google Scholar]
- Shirai T., Nazarewicz R.R., Wallis B.B., Yanes R.E., Watanabe R., Hilhorst M., Tian L., Harrison D.G., Giacomini J.C., Assimes T.L., Goronzy J.J., Weyand C.M. The glycolytic enzyme PKM2 bridges metabolic and inflammatory dysfunction in coronary artery disease. J. Exp. Med. 2016;213(3):337–354. doi: 10.1084/jem.20150900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang Y., Yang X., Friesel R.E., Vary C.P., Liaw L. Mechanisms of TGF-beta-induced differentiation in human vascular smooth muscle cells. J. Vasc. Res. 2011;48(6):485–494. doi: 10.1159/000327776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trogan E., Feig J.E., Dogan S., Rothblat G.H., Angeli V., Tacke F., Randolph G.J., Fisher E.A. Gene expression changes in foam cells and the role of chemokine receptor CCR7 during atherosclerosis regression in ApoE-deficient mice. Proc. Natl. Acad. Sci. U.S.A. 2006;103(10):3781–3786. doi: 10.1073/pnas.0511043103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turrens J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003;552(Pt 2):335–344. doi: 10.1113/jphysiol.2003.049478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulivieri C., Baldari C.T. Statins: from cholesterol-lowering drugs to novel immunomodulators for the treatment of Th17-mediated autoimmune diseases. Pharmacol. Res. 2014;88:41–52. doi: 10.1016/j.phrs.2014.03.001. [DOI] [PubMed] [Google Scholar]
- Viaud M., Ivanov S., Vujic N., Duta-Mare M., Aira L.E., Barouillet T., Garcia E., Orange F., Dugail I., Hainault I., Stehlik C., Marchetti S., Boyer L., Guinamard R., Foufelle F., Bochem A., Hovingh K.G., Thorp E.B., Gautier E.L., Kratky D., Dasilva-Jardine P., Yvan-Charvet L. Lysosomal cholesterol hydrolysis couples efferocytosis to anti-inflammatory oxysterol production. Circ. Res. 2018;122(10):1369–1384. doi: 10.1161/CIRCRESAHA.117.312333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vuononvirta J., Marelli-Berg F.M., Poobalasingam T. Metabolic regulation of T lymphocyte motility and migration. Mol. Aspect. Med. 2020:100888. doi: 10.1016/j.mam.2020.100888. [DOI] [PubMed] [Google Scholar]
- Wassmann S., Laufs U., Muller K., Konkol C., Ahlbory K., Baumer A.T., Linz W., Bohm M., Nickenig G. Cellular antioxidant effects of atorvastatin in vitro and in vivo. Arterioscler. Thromb. Vasc. Biol. 2002;22(2):300–305. doi: 10.1161/hq0202.104081. [DOI] [PubMed] [Google Scholar]
- Waters L.R., Ahsan F.M., Wolf D.M., Shirihai O., Teitell M.A. Initial B cell activation induces metabolic reprogramming and mitochondrial remodeling. iScience. 2018;5:99–109. doi: 10.1016/j.isci.2018.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei X., Schneider J.G., Shenouda S.M., Lee A., Towler D.A., Chakravarthy M.V., Vita J.A., Semenkovich C.F. De novo lipogenesis maintains vascular homeostasis through endothelial nitric-oxide synthase (eNOS) palmitoylation. J. Biol. Chem. 2011;286(4):2933–2945. doi: 10.1074/jbc.M110.193037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werle M., Kreuzer J., Hofele J., Elsasser A., Ackermann C., Katus H.A., Vogt A.M. Metabolic control analysis of the Warburg-effect in proliferating vascular smooth muscle cells. J. Biomed. Sci. 2005;12(5):827–834. doi: 10.1007/s11373-005-9010-5. [DOI] [PubMed] [Google Scholar]
- Westerterp M., Gautier E.L., Ganda A., Molusky M.M., Wang W., Fotakis P., Wang N., Randolph G.J., D'Agati V.D., Yvan-Charvet L., Tall A.R. Cholesterol accumulation in dendritic cells links the inflammasome to acquired immunity. Cell Metabol. 2017;25(6):1294–1304. doi: 10.1016/j.cmet.2017.04.005. e1296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams N.C., O'Neill L.A. ACLY-matizing macrophages to histone modification during immunometabolic reprogramming. Trends Immunol. 2020;41(2):93–94. doi: 10.1016/j.it.2019.12.009. [DOI] [PubMed] [Google Scholar]
- Winkels H., Ley K. Natural killer cells at ease: atherosclerosis is not affected by genetic depletion or hyperactivation of natural killer cells. Circ. Res. 2018;122(1):6–7. doi: 10.1161/CIRCRESAHA.117.312289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W., Bai Y., Xiong Y., Zhang J., Chen S., Zheng X., Meng X., Li L., Wang J., Xu C., Yan C., Wang L., Chang C.C., Chang T.Y., Zhang T., Zhou P., Song B.L., Liu W., Sun S.C., Liu X., Li B.L., Xu C. Potentiating the antitumour response of CD8(+) T cells by modulating cholesterol metabolism. Nature. 2016;531(7596):651–655. doi: 10.1038/nature17412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yilmaz O., Afsar B., Ortiz A., Kanbay M. The role of endothelial glycocalyx in health and disease. Clin Kidney J. 2019;12(5):611–619. doi: 10.1093/ckj/sfz042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yurdagul A., Jr., Subramanian M., Wang X., Crown S.B., Ilkayeva O.R., Darville L., Kolluru G.K., Rymond C.C., Gerlach B.D., Zheng Z., Kuriakose G., Kevil C.G., Koomen J.M., Cleveland J.L., Muoio D.M., Tabas I. Macrophage metabolism of apoptotic cell-derived arginine promotes continual efferocytosis and resolution of injury. Cell Metabol. 2020;31(3):518–533. doi: 10.1016/j.cmet.2020.01.001. e510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yvan-Charvet L., Bonacina F., Guinamard R.R., Norata G.D. Immunometabolic function of cholesterol in cardiovascular disease and beyond. Cardiovasc. Res. 2019;115(9):1393–1407. doi: 10.1093/cvr/cvz127. [DOI] [PubMed] [Google Scholar]
- Yvan-Charvet L., Pagler T., Gautier E.L., Avagyan S., Siry R.L., Han S., Welch C.L., Wang N., Randolph G.J., Snoeck H.W., Tall A.R. ATP-binding cassette transporters and HDL suppress hematopoietic stem cell proliferation. Science. 2010;328(5986):1689–1693. doi: 10.1126/science.1189731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zernecke A. Dendritic cells in atherosclerosis: evidence in mice and humans. Arterioscler. Thromb. Vasc. Biol. 2015;35(4):763–770. doi: 10.1161/ATVBAHA.114.303566. [DOI] [PubMed] [Google Scholar]
- Zhang X.J., Qin J.J., Cheng X., Shen L., Zhao Y.C., Yuan Y., Lei F., Chen M.M., Yang H., Bai L., Song X., Lin L., Xia M., Zhou F., Zhou J., She Z.G., Zhu L., Ma X., Xu Q., Ye P., Chen G., Liu L., Mao W., Yan Y., Xiao B., Lu Z., Peng G., Liu M., Yang J., Yang L., Zhang C., Lu H., Xia X., Wang D., Liao X., Wei X., Zhang B.H., Zhang X., Yang J., Zhao G.N., Zhang P., Liu P.P., Loomba R., Ji Y.X., Xia J., Wang Y., Cai J., Guo J., Li H. In-hospital use of statins is associated with a reduced risk of mortality among individuals with COVID-19. Cell Metabol. 2020;32(2):176–187. doi: 10.1016/j.cmet.2020.06.015. e174. [DOI] [PMC free article] [PubMed] [Google Scholar]