

Abstract
In 2016, tropical screech owl (Megascops choliba) (Tso) chicks were suddenly found dead in a Japanese breeding facility. We autopsied a 9-day-old Tso and discovered white spots scattered on the liver surface. Multifocal necrosis was diffused, and macrophages had infiltrated the necrotic hepatic lesions. Hepatocytes contained numerous intranuclear inclusion bodies. Immunohistochemical staining detected Adenovirus antigen only in the liver. Next, PCR and sequencing (LC536616) identified Tso Adenovirus (TsoAd). Basic Local Alignment Search Tool (BLAST) and phylogenic analyzes suggested TsoAd is an owl Aviadenovirus. Our study contributes to an improved understanding of infectious disease among captive raptors.
Keywords: Aviadenovirus, inclusion body hepatitis, Japan, tropical screech owl
Adenoviruses infect numerous species of mammals, birds, reptiles, and amphibians [6]. Adenoviridae is divided into five groups, with Aviadenovirus and Atadenovirus being infectious-disease agents in birds [6].
Aviadenovirus is a major cause of inclusion body hepatitis [12], gizzard erosion, and ulceration [4] in chickens. Inclusion body hepatitis has high mortality [9], and outbreaks have been reported worldwide [19], including in Japan [17]. Although inclusion body hepatitis caused by Aviadenovirus infection has been observed in birds other than chicken, including turkey, duck [26], racing pigeon [8], and Gambel’s quail [1], it is limited in raptors, except in falcon [2, 15, 20, 21, 23].
The tropical screech owl (Megascops choliba) is a common raptor occasionally kept as a pet. Endemic to South America, pet screech owls are typically imported to Japan, although some are bred domestically. Although this species is widespread, we know little regarding infectious diseases that affect them, especially in breeding facilities.
In this study, we report the detection of inclusion body hepatitis in the tropical screech owl, caused by an owl Aviadenovirus. Our research aims to improve current understanding of owl infectious disease in captive conditions, using pathological and phylogenetic data.
From March to May 2016, numerous tropical screech owls (age range 1–20 days) in a breeding facility were presented anorexia, vomiting, diarrhea, and sudden mortality. Owls were treated with toltrazuril as an anticoccidial agent and manually fed. Most birds died within one week. The facility houses other raptors, such as Eurasian eagle owl (Bubo bubo), rock eagle owl (Bubo bengalensis), barn owl (Tyto alba), and several falcon species. Most were imported, with some individuals used for breeding. Juvenile screech owls were housed in independent cages that caretakers disinfected with chlorine once a day. Their diet was a commercial feed comprising minced chicks of chicken and Japanese quail.
A 9-day-old tropical screech owl that died suddenly without external symptoms was moved to Aichi Prefectural Chuo Livestock Hygiene Service Center by a veterinary hospital and was autopsied (Fig. 1a). We observed white spots scattered across the liver surface (Fig. 1b) and hemorrhaging in the gizzard. No other gross lesions were detected. Intestinal contents did not contain coccidian oocysts.
Fig. 1.
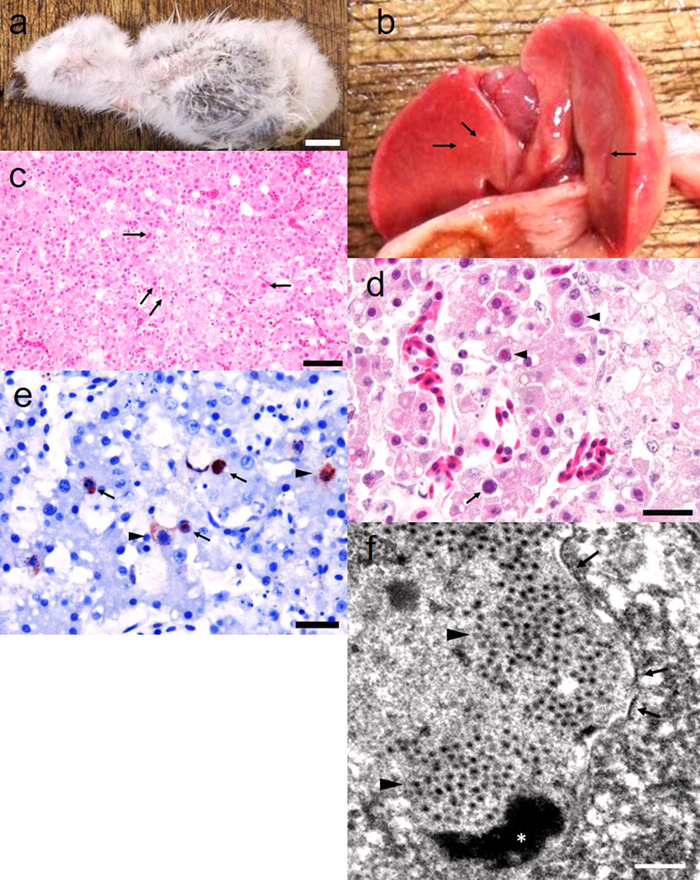
a) Nine-day-old tropical screech owl chick, found dead in the breeding facility. Bar=1 cm. b) White spots were scattered on the liver surface (arrows). c) Hepatic necrosis. Intranuclear inclusion bodies were detected in surrounding necrotic area (arrows). Hematoxylin and eosin (H&E) staining. Bar=50 µm. d) Full basophilic (arrow) and Cowdry type A eosinophilic (arrowheads) inclusion bodies in the liver. H&E staining. Bar=20 µm. e) Immunohistochemical staining detected numerous positive reactions in hepatocyte nuclei (arrows) and cytoplasm of infiltrated macrophages (arrowheads). Bar=20 µm. f) Transmission electron microscopy of TsoAd. Nonenveloped isometric viral particles (~90 nm in size) were arrayed in hepatocyte nuclei (arrowheads). The nuclear membrane was deformed (arrows). *Chromatin. Bar=500 nm.
At autopsy, the liver, kidney, heart, lung, pancreas, trachea, stomach, intestine, and brain tissue samples were fixed in 10% neutral-buffered formalin. Fixed tissues were then embedded in paraffin wax, sectioned (~3 µm), then stained with hematoxylin and eosin (H&E) for histological examination. Gram staining was also performed on the liver, kidney, and gizzard sections. For immunohistochemical examination, formalin-fixed paraffin-embedded (FFPE) sections of the liver, gizzard, pancreas, and intestines were deparaffinized and incubated with 3% hydrogen peroxide in methanol solution to suppress endogenous peroxidase activity. Antigen retrieval was conducted using 0.1% actinase E solution in phosphate buffered saline at 37°C for 20 min. After adding 10% normal goat serum to block non-specific reactions, sections were incubated with rabbit anti-Aviadenovirus antibody [18] for 1 hr. Sections were incubated with secondary antibody (Histofine Simple Stain MAX-PO Multi; Nichirei Bioscience Inc., Tokyo, Japan) for 30 min at room temperature, then treated with aminoethyl carbazole (AEC) substrate solution (Histofine Simple Stain AEC solution; Nichirei Bioscience Inc.), also at room temperature. Finally, sections were counterstained with hematoxylin.
To identify the virus associated with intranuclear inclusion bodies, formalin-fixed liver tissue was observed under transmission electron microscopy (TEM) (JEM-1400 Flash; JOEL, Tokyo, Japan), as described previously [14].
Histopathological examination revealed that multifocal necrosis was randomly diffused in the liver (Fig. 1c). Macrophages had infiltrated into necrotic lesions, and we also observed a few granulomatous necrotizing lesions. Hepatocytes in the surrounding necrotic areas contained numerous full, basophilic, and Cowdry type A, eosinophilic intranuclear inclusion bodies (Fig. 1d). Kidney interstitium exhibited infiltration of lymphocytes and a few macrophages. Gizzard surface displayed hemorrhaging and heterophil infiltration. Gram staining revealed a few gram-positive bacteria on the surface of the gizzard although no bacteria were detected in the liver and kidney.
Immunohistochemical staining revealed positive reactions in hepatocyte nuclei and infiltrated macrophage cytoplasm (Fig. 1e). We did not observe reactions in other tissues.
TEM observation revealed that hepatocyte nuclei had deformed membranes and contained nonenveloped isometric viral particles of approximately 90 nm in size (Fig. 1f).
Next, we conducted PCR on FFPE sections to investigate viruses associated with inclusion body hepatitis. Genomic DNA was extracted from liver sections using the QIAamp DNA FFPE Tissue kit (Qiagen GmbH, Hilden, Germany), following manufacturer protocol. Primers used targeted those encoding hexon [11], DNA-dependent Adenovirus DNA polymerase [25], and Herpesvirus DNA polymerase [24]; only Adenovirus DNA polymerase was amplified (516 bp). Amplicons of the first degenerate PCR were sequenced. Basic Local Alignment Search Tool (BLAST) searches of the National Center for Biotechnology Information (NCBI) revealed that partial nucleotide and amino acid sequences exhibited the highest similarity with those of fowl Aviadenovirus E (accession number: MK572855) (74.61%) and fowl Aviadenovirus A (accession number: APP94056) (86.63%). This result implies that the tropical screech owl Adenovirus (TsoAd) that we detected possibly represents a novel Aviadenovirus because the novelty threshold for an Aviadenovirus is a 5–15% sequence gap in the DNA-dependent DNA polymerase amino acid sequence [5]. However, the full-length sequence of DNA polymerase, genome organization, cross-neutralization, and other characteristics should be investigated for greater certainty [5]. The TsoAd sequence was deposited to the DNA Data Bank of Japan (accession number: LC536616).
We performed a phylogenetic analysis of the TsoAd amino acid sequence with the other aviadenoviruses. Sequences included were those of fowl, goose, duck, psittacine, pigeon, and owl adenoviruses, with egg drop syndrome virus set as an outgroup. Multiple alignments were performed in MEGA version 7.0, using Clustal W. All candidate sequences were retrieved from the NCBI nucleotide database. A neighbor-joining phylogenic tree was constructed in MEGA 7. The TsoAd amino acid sequence was classified as an Aviadenovirus and is closely related to barn owl Adenovirus (Fig. 2).
Fig. 2.
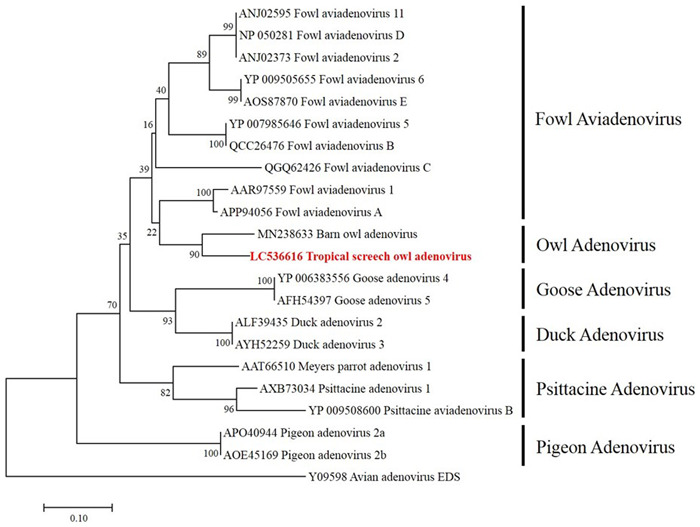
Phylogenic tree based on partial adenoviral DNA-dependent DNA polymerase amino acid sequences. The scale bar represents genetic distances among strains. Egg drop syndrome (EDS) virus (accession number: Y09598) was the outgroup.
These results confirmed Aviadenovirus infection in the dead specimen and provides an explanation for the outbreak of inclusion body hepatitis. Another recent study had also detected novel Adenovirus in tropical screech owls, but classified it as Atadenovirus and used fecal samples of owls without clinical symptoms [7]. Therefore, our findings are the first report of Aviadenovirus infection with clinical symptoms in the tropical screech owl. To the best of our knowledge, Aviadenovirus infection in an owl is limited [27]. Among a captive breeding colony of taita falcons that experienced an Aviadenovirus outbreak, adult females had started laying eggs before dying [2]. Pathogenicity of Aviadenovirus against chickens differed across strains [10] and age [16]. In our study, characteristic symptoms were diarrhea, sudden death, and high mortality. Our observations imply that the disease is highly virulent, as owls were probably too young to express other symptoms before dying.
Multifocal necrosis was diffused widely in the liver. We detected inclusion bodies mainly around, rather than in, necrotic lesions. Immunohistochemical staining revealed that Adenovirus antigens were present only in the liver. This was not the case for other birds; infected taita falcons exhibited inclusion bodies or lesions in the liver, spleen [2, 23], and kidneys [2], while chickens had lesions in pancreatic acinar cells [13] and small-intestine epithelial cells [17]. In addition, despite being infected with the same Aviadenovirus infection, broiler and layer chickens differed in clinical symptoms and mortality rates [13]. This variation in lesion appearance may be partly attributable to our specimen’s rapid death, which may have prevented viral spread throughout the rest of the body. To verify this hypothesis, we require more research using other viral strains and owls of different ages.
We could not identify the source of Aviadenovirus infection. We initially thought the virus may have been introduced via the feed (containing minced chicken and quail), because owls were maintained in separate cages that are frequently disinfected with chlorine. However, BLAST and phylogenetic analysis indicated that the detected Aviadenovirus was different from chicken Aviadenovirus. Although chickens and quails do not appear susceptible to infection from falcon-derived Aviadenovirus [20], cross-species transmission between chickens and falcons has been documented [15]. We used the hexon gene [11] to compare our owl Aviadenovirus with strains isolated in Japan [12], but this PCR was unsuccessful, possibly due to the influence of formalin fixing [3, 22].
Although phylogenetic analysis indicated that TsoAd was close to barn owl Adenovirus, barn owls housed in the same facility did not express any symptoms. Some other owls and falcons were also housed in this facility, however, we were not able to investigate whether these raptors had adenoviruses or not. A previous study reported that wild and captive peregrine falcons presented high seropositivity rates for falcon Adenovirus (80% and 100%) and that they act as a primary reservoir for the virus [20]. Similarly, we suspect that imported barn owls or the other raptors may be subclinical carriers of adenoviruses. To clarify the mechanisms of cross-species transmission among tropical screech owl, chicken, and quail in Japan, we recommend conducting molecular epidemiological studies using DNA polymerase region and next-generation sequencing technology in addition to using hexon region [12].
In conclusion, we identified an owl Aviadenovirus that caused an outbreak of inclusion body hepatitis in captive tropical screech owls. Few reports are available regarding Adenovirus infection in this raptor species, despite its popularity as a companion animal. Our study provides important insight on diseases in tropical screech owls that should benefit both veterinarians and raptor breeders.
Acknowledgments
We thank Dr. N. Tanimura for advice on the manuscript, as well as Mr. M. Kobayashi and Ms. M. Shimada for histopathological assistance.
REFERENCES
- 1.Bradley G. A., Shupe M. R., Reggiardo C., Noon T. H., Lozano-Alarcon F., Bicknell E. J.1994. Inclusion body hepatitis in Gambel’s quail (Callipepla gambelii). J. Wildl. Dis. 30: 281–284. doi: 10.7589/0090-3558-30.2.281 [DOI] [PubMed] [Google Scholar]
- 2.Dean J., Latimer K. S., Oaks J. L., Schrenzel M., Redig P. T., Wünschmann A.2006. Falcon adenovirus infection in breeding Taita falcons (Falco fasciinucha). J. Vet. Diagn. Invest. 18: 282–286. doi: 10.1177/104063870601800310 [DOI] [PubMed] [Google Scholar]
- 3.Feldman M. Y.1973. Reactions of nucleic acids and nucleoproteins with formaldehyde. Prog. Nucleic Acid Res. Mol. Biol. 13: 1–49. doi: 10.1016/S0079-6603(08)60099-9 [DOI] [PubMed] [Google Scholar]
- 4.Gjevre A. G., Kaldhusdal M., Eriksen G. S.2013. Gizzard erosion and ulceration syndrome in chickens and turkeys: a review of causal or predisposing factors. Avian Pathol. 42: 297–303. doi: 10.1080/03079457.2013.817665 [DOI] [PubMed] [Google Scholar]
- 5.Harrach B., Benko M., Both G. W., Brown M., Davison A. J., Echavarria M., Hess M., Jones M. S., Kajon A., Lehmkuhl H. D., Mautner V., Mittal S. K., Wadell G.2011. Family adenoviridae. pp. 125–141. In: Virus Taxonomy, 9th Report of the International Committee on Taxonomy of Viruses (King, A. M. Q., Lefkowitz, E., Adams, M. J. and Carstens, E. B. eds.), Elsevier, San Diego. [Google Scholar]
- 6.Harrach B., Tarján Z. L., Benkő M.2019. Adenoviruses across the animal kingdom: a walk in the zoo. FEBS Lett. 593: 3660–3673. doi: 10.1002/1873-3468.13687 [DOI] [PubMed] [Google Scholar]
- 7.Jejesky de Oliveira A. P., Valdetaro Rangel M. C. Z., Z Vidovszky M., Rossi J. L., Jr., Vicentini F., Harrach B., L Kaján G.2020. Identification of two novel adenoviruses in smooth-billed ani and tropical screech owl. PLoS One 15: e0229415. doi: 10.1371/journal.pone.0229415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ketterer P. J., Timmins B. J., Prior H. C., Dingle J. G.1992. Inclusion body hepatitis associated with an adenovirus in racing pigeons in Australia. Aust. Vet. J. 69: 90–91. doi: 10.1111/j.1751-0813.1992.tb15559.x [DOI] [PubMed] [Google Scholar]
- 9.Li L., Wang J., Chen P., Zhang S., Sun J., Yuan W.2018. Pathogenicity and molecular characterization of a fowl adenovirus 4 isolated from chicken associated with IBH and HPS in China. BMC Vet. Res. 14: 400. doi: 10.1186/s12917-018-1733-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Liu Y., Wan W., Gao D., Li Y., Yang X., Liu H., Yao H., Chen L., Wang C., Zhao J.2016. Genetic characterization of novel fowl aviadenovirus 4 isolates from outbreaks of hepatitis-hydropericardium syndrome in broiler chickens in China. Emerg. Microbes Infect. 5: e117. doi: 10.1038/emi.2016.115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mase M., Mitake H., Inoue T., Imada T.2009. Identification of group I-III avian adenovirus by PCR coupled with direct sequencing of the hexon gene. J. Vet. Med. Sci. 71: 1239–1242. doi: 10.1292/jvms.71.1239 [DOI] [PubMed] [Google Scholar]
- 12.Mase M., Nakamura K., Minami F.2012. Fowl adenoviruses isolated from chickens with inclusion body hepatitis in Japan, 2009–2010. J. Vet. Med. Sci. 74: 1087–1089. doi: 10.1292/jvms.11-0443 [DOI] [PubMed] [Google Scholar]
- 13.Matos M., Grafl B., Liebhart D., Hess M.2016. The outcome of experimentally induced inclusion body hepatitis (IBH) by fowl aviadenoviruses (FAdVs) is crucially influenced by the genetic background of the host. Vet. Res. 47: 69. doi: 10.1186/s13567-016-0350-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Matsubayashi M., Murakoshi N., Komatsu T., Tokoro M., Haritani M., Shibahara T.2015. Genetic identification of Entamoeba polecki subtype 3 from pigs in Japan and characterisation of its pathogenic role in ulcerative colitis. Infect. Genet. Evol. 36: 8–14. doi: 10.1016/j.meegid.2015.08.030 [DOI] [PubMed] [Google Scholar]
- 15.Mohamed M. H. A., El-Sabagh I. M., Abdelaziz A. M., Al-Ali A. M., Alramadan M., Lebdah M. A., Ibrahim A. M., Al-Ankari A. S.2018. Molecular characterization of fowl aviadenoviruses species D and E associated with inclusion body hepatitis in chickens and falcons indicates possible cross-species transmission. Avian Pathol. 47: 384–390. doi: 10.1080/03079457.2018.1457769 [DOI] [PubMed] [Google Scholar]
- 16.Nakamura K., Mase M., Yamaguchi S., Yuasa N.2000. Induction of hydropericardium in one-day-old specific-pathogen-free chicks by adenoviruses from inclusion body hepatitis. Avian Dis. 44: 192–196. doi: 10.2307/1592524 [DOI] [PubMed] [Google Scholar]
- 17.Nakamura K., Mase M., Yamamoto Y., Takizawa K., Kabeya M., Wakuda T., Matsuda M., Chikuba T., Yamamoto Y., Ohyama T., Takahashi K., Sato N., Akiyama N., Honma H., Imai K.2011. Inclusion body hepatitis caused by fowl adenovirus in broiler chickens in Japan, 2009–2010. Avian Dis. 55: 719–723. doi: 10.1637/9813-052511-Case.1 [DOI] [PubMed] [Google Scholar]
- 18.Nakamura K., Shoyama T., Mase M., Imada T., Yamada M.2003. Reproduction of hydropericardium syndrome in three-week-old cyclophosphamide-treated specific-pathogen-free chickens by adenoviruses from inclusion body hepatitis. Avian Dis. 47: 169–174. doi: 10.1637/0005-2086(2003)047[0169:ROHSIT]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 19.Niu Y., Sun Q., Zhang G., Sun W., Liu X., Xiao Y., Shang Y., Liu S.2018. Epidemiological investigation of outbreaks of fowl adenovirus infections in commercial chickens in China. Transbound. Emerg. Dis. 65: e121–e126. doi: 10.1111/tbed.12691 [DOI] [PubMed] [Google Scholar]
- 20.Oaks J. L., Schrenzel M., Rideout B., Sandfort C.2005. Isolation and epidemiology of falcon adenovirus. J. Clin. Microbiol. 43: 3414–3420. doi: 10.1128/JCM.43.7.3414-3420.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schrenzel M., Oaks J. L., Rotstein D., Maalouf G., Snook E., Sandfort C., Rideout B.2005. Characterization of a new species of adenovirus in falcons. J. Clin. Microbiol. 43: 3402–3413. doi: 10.1128/JCM.43.7.3402-3413.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sikorsky J. A., Primerano D. A., Fenger T. W., Denvir J.2007. DNA damage reduces Taq DNA polymerase fidelity and PCR amplification efficiency. Biochem. Biophys. Res. Commun. 355: 431–437. doi: 10.1016/j.bbrc.2007.01.169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tomaszewski E. K., Phalen D. N.2007. Falcon adenovirus in an American kestrel (Falco sparverius). J. Avian Med. Surg. 21: 135–139. doi: 10.1647/1082-6742(2007)21[135:FAIAAK]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 24.VanDevanter D. R., Warrener P., Bennett L., Schultz E. R., Coulter S., Garber R. L., Rose T. M.1996. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 34: 1666–1671. doi: 10.1128/JCM.34.7.1666-1671.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wellehan J. F., Johnson A. J., Harrach B., Benkö M., Pessier A. P., Johnson C. M., Garner M. M., Childress A., Jacobson E. R.2004. Detection and analysis of six lizard adenoviruses by consensus primer PCR provides further evidence of a reptilian origin for the atadenoviruses. J. Virol. 78: 13366–13369. doi: 10.1128/JVI.78.23.13366-13369.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yugo D. M., Hauck R., Shivaprasad H. L., Meng X. J.2016. Hepatitis virus infections in poultry. Avian Dis. 60: 576–588. doi: 10.1637/11229-070515-Review.1 [DOI] [PubMed] [Google Scholar]
- 27.Zsivanovits P., Monks D. J., Forbes N. A., Ursu K., Raue R., Benkő M.2006. Presumptive identification of a novel adenovirus in a Harris Hawk (Parabuteo unicinctus), a Bengal Eagle Owl (Bubo bengalensis), and a Verreaux’s Eagle Owl (Bubo lacteus). J. Avian Med. Surg. 20: 105–112. doi: 10.1647/2005-008.1 [DOI] [Google Scholar]