

Land-cover change exerts more influence on soil respiration in boreal and temperate regions than climate change.
Abstract
Soil respiration (Rs) represents the largest flux of CO2 from terrestrial ecosystems to the atmosphere, but its spatial and temporal changes as well as the driving forces are not well understood. We derived a product of annual global Rs from 2000 to 2014 at 1 km by 1 km spatial resolution using remote sensing data and biome-specific statistical models. Different from the existing view that climate change dominated changes in Rs, we showed that land-cover change played a more important role in regulating Rs changes in temperate and boreal regions during 2000–2014. Significant changes in Rs occurred more frequently in areas with significant changes in short vegetation cover (i.e., all vegetation shorter than 5 m in height) than in areas with significant climate change. These results contribute to our understanding of global Rs patterns and highlight the importance of land-cover change in driving global and regional Rs changes.
INTRODUCTION
Large uncertainties in the global carbon budget are associated with the terrestrial carbon cycle, and reducing these uncertainties requires an improved capacity to estimate carbon fluxes between the atmosphere and terrestrial ecosystems (1). Over two-thirds of terrestrial carbon is stored belowground, and a significant amount of the atmospheric CO2 assimilated by plants is respired by roots and microbes in soils (2, 3). Soil respiration (Rs), consisting of root (autotrophic) respiration (Ra) and microbial (heterotrophic) respiration (Rh), is therefore a key process in the terrestrial carbon cycle. However, Rs is difficult to estimate at the global scale because of the limited understanding of the complex interactions of physical, chemical, and biological processes and the resulting high spatiotemporal dynamics (4).
Compared to the large number of studies on plant productivity in terrestrial carbon cycling, there are few studies on global Rs dynamics (4, 5). Global Rs can be estimated using top-down or bottom-up approaches. Top-down approaches typically estimate Rs as the residual of the carbon balance, and thus, the indirect estimates can naturally capture average soil carbon fluxes over large areas, although the results inevitably suffer from errors and uncertainties from any of the source datasets (6). By contrast, bottom-up methods, including process-based models and empirical statistical models, use direct observational data (6, 7). Process-based models rely on knowledge of the interactions among different physical and ecological processes, but many models rarely simulate Rs explicitly and instead generate only an explicit output of Rh (8). Global Rh may provide some information on Rs, but the use of different model structures and parameters has resulted in large discrepancies among them (3, 9). Compared to process-based models, statistical models are simpler and can provide data-oriented estimates of Rs using various biotic or abiotic factors as input, such as temperature, moisture, plant productivity, and soil properties (2, 3, 9–17). Previous studies using statistical models have produced mean annual global Rs estimates from 78 to 108 Pg C year−1, and the error associated with each estimation was high (table S1). One study (7) noted that the lack of large-scale, observation-driven Rs estimates was a major problem in constraining regional-scale to global-scale C fluxes.
Given that multiple regional-scale studies (18, 19) have shown that remote sensing data can be used to estimate Rs with high accuracy, we expect that it is also possible to estimate Rs beyond the regional scale. The openly available remote sensing data of high spatial resolution provide large-scale observations, and remote sensing is anticipated to play an increasingly important role in carbon cycle research in the future (20). For instance, current remote sensing products provide a range of key ecosystem variables that accurately reflect the spatiotemporal variations in surface temperature, moisture, and plant productivity of terrestrial ecosystems (21–23). Adachi et al. (14) provided insights into the estimation of global Rs by combining empirical models derived from field studies with remote sensing data. However, Adachi et al. (14) used one empirical model from a site-specific study for all ecosystem types, which is unlikely to be representative and may lead to large uncertainty in global Rs estimation.
Rs change can be affected by many factors, and climate factors (e.g., air temperature and precipitation) have been commonly investigated because of their direct or indirect effects on Rs metabolism (9–15). Land-cover change can also greatly affect Rs by changing vegetation structure, plant species composition, local microclimate, and soil properties (24, 25). For instance, global greening and the associated vegetation structural change could affect Rs by altering biogeophysical processes (26, 27). However, few studies have comprehensively considered both climate and land-cover change effects on the spatial and temporal variations in global Rs. As soil nutrient availability ultimately depends on photosynthate supply (28), we assume that the estimation of global Rs can be achieved by including three factors: temperature, moisture, and plant productivity.
With the number of measurements of Rs rapidly increasing worldwide, future modeling efforts that are focused on global Rs estimation should take advantage of remote sensing data and data mining techniques to link observations at different scales. In this study, we have developed an annual global Rs dataset at 1 km by 1 km spatial resolution for the time period from 2000 to 2014. The product was generated by combining globally distributed in situ Rs measurements, satellite remote sensing data, and biome-specific statistical models, including parametric models and nonparametric machine learning models. We analyzed spatial and temporal variations in global Rs. We then investigated how the changing trends in Rs were related to changes in climate (temperature, precipitation, and drought) and land cover.
RESULTS
Deriving annual global Rs product from 2000 to 2014
On the basis of a 10-fold cross-validation method, we evaluated the accuracy of the four statistical models [i.e., multiple nonlinear regression (MNLR), random forest regression (RFR), support vector regression (SVR), and artificial neural network (ANN) models] by comparing RMSE (root mean square error) and R2 (coefficient of determination) (table S2) using available measured Rs datasets as a reference. We determined the biome-scale model for the estimation of annual Rs of each biome by selecting the model with the highest cross-validation accuracy (table S3). Although the selected models were not the same across the 10 biomes, they were all machine learning algorithm models. The selected models explained 62 to 84% of the interannual and intersite variabilities in annual Rs with an RMSE ranging from 107 to 413 g C m−2 year−1 (table S3). On the basis of the selected biome-scale models (tables S3 and S4), we produced the annual global Rs dataset at the global scale from 2000 to 2014.
Spatial and temporal patterns of global Rs
The regions with high Rs are located in the tropics, such as the Amazon Basin, Central Africa, and Southeast Asia. Low Rs values are widely distributed in the Northern Hemisphere high-latitude regions, western United States, Canada, Central Asia, northern Mongolia, northeast China, Argentina, and Australia (Fig. 1). Boreal, temperate, and tropical regions contributed 15, 24, and 61%, respectively, to the total mean annual global Rs. Our estimated mean annual global Rs from 2000 to 2014 is 72.6 Pg C year−1 [95% confidence interval (CI) = 69.8-75.4 Pg C year−1], which was lower than the estimated global Rs for recent years (table S1). The estimated annual global Rs showed fluctuations over time (fig. S2), with the lowest value (70.6 Pg C year−1) occurring in 2000 and the highest value (74.5 Pg C year−1) occurring in 2010.
Fig. 1. Global distribution of mean annual Rs between 2000 and 2014.
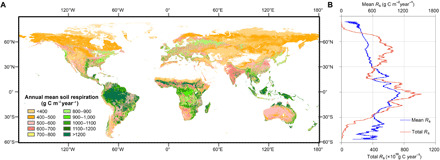
(A) Global map of mean annual Rs at 1 km by 1 km spatial resolution derived using satellite data. (B) Latitudinal distribution of mean annual Rs (blue line) and total annual Rs (orange line). All land grids along a latitudinal row in the global map were averaged to derive mean Rs and summed to derive total Rs.
Over the 15 years of the study period, the Rs trends varied at different spatial scales (Fig. 2A). Rs showed a significantly increasing trend globally (0.13 ± 0.02 Pg C year−1, P < 0.05), in the boreal region (0.05 ± 0.01 Pg C year−1, P < 0.05), and in the tropical region (0.11 ± 0.02 Pg C year−1, P < 0.05) and a nonsignificantly decreasing trend in the temperate region (−0.02 ± 0.01 Pg C year−1, P = 0.42). The tropics accounted for approximately 85% of the total increase in the global Rs. The largest Rs net change occurred in the tropical region (1.54 Pg C, 95% CI = 1.30-1.78 Pg C; +3.66% change relative to the global Rs in 2000), followed by those in the boreal region (0.65 Pg C, 95% CI = 0.49-0.81 Pg C; +6.80%) and temperate region (−0.27 Pg C, 95% CI = −0.43-−0.11 Pg C; −1.53%).
Fig. 2. Trends in annual Rs from 2000 to 2014.
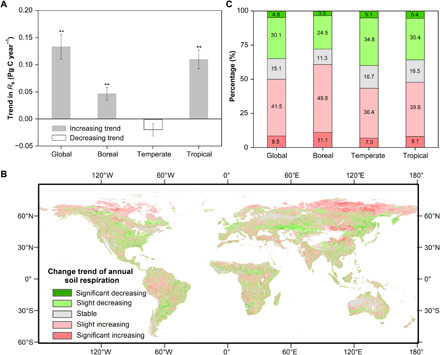
(A) Overall trends in annual Rs. To calculate the overall trend at the global scale, global average Rs for each year was first derived. Then, a two-sided Mann-Kendall test and a Theil-Sen median trend analysis were performed. Trends at the regional scales were derived following the same method as the global trend. Gray bars in the upward direction indicate an increasing trend, whereas the hollow bar in the downward direction indicates a decreasing trend, with two asterisks denoting significant trends (P < 0.05). Error bars represent the 95% CIs estimated via a 1000-bootstrap analysis. (B) Spatially explicit trends in annual Rs at 1 km by 1 km spatial resolution. Similar to overall trends in (A), per-pixel trend was characterized using a two-sided Mann-Kendall test and a Theil-Sen median trend analysis. Significant increasing and decreasing trends correspond to positive and negative Theil-Sen estimators, respectively, with significant Mann-Kendall test (P < 0.05). Slightly increasing and decreasing trends correspond to positive and negative Theil-Sen estimators, respectively, with nonsignificant Mann-Kendall test results (P > 0.05). The residual pixels belong to a stable class. (C) Normalized frequency distribution of trends derived using the map presented in (B). The color scheme of the histogram bars matches that of the map legend.
At the global scale, 50% of our study area experienced an increasing trend in Rs, and the regions with a significant increasing trend (8.5%) were spatially aggregated and mainly located at the high latitudes (e.g., northern Canada, northern Russia, and northern Europe), northern Mongolia, Loess Plateau and northeastern Tibetan Plateau of China, India, Amazon Basin, and Congo Basin (Fig. 2, B and C). The annual Rs significantly decreased in less than 5% of the study regions; these regions were mainly scattered in southeastern Russia, midlatitude temperate regions (i.e., United States, Kazakhstan, and northeast China), southern Africa, and Argentina (Fig. 2B). At the regional scale, the area with the highest proportion of increasing Rs (60.7%, 11.1% with a significant increase) was the boreal region, and the area with the highest proportion of decreasing Rs (39.9%, 5.1% with a significant decrease) was the temperate region (Fig. 2C).
The relationships between Rs and its driving factors at global and regional scales
Global Rs showed a significant positive partial correlation with annual mean air temperature (TEM) from 2000 to 2014 (R = 0.67, P < 0.05; Fig. 3). A significant positive partial correlation between Rs and TEM was also found in the tropical region over the study period (R = 0.83, P < 0.05; Fig. 3). However, for boreal and temperate regions, land-cover factors, especially the short vegetation (SV) cover, showed consistently significant positive partial correlation with Rs during 2000–2014 (Fig. 3). Through the statistical analyses of the per-pixel significant change trends in Rs and the six driving factors over the globe and three regions (Fig. 2B and figs. S4 and S5), we found that the highest proportion of the areas where Rs changed significantly was located in areas with significant change in TEM at the global scale and in the tropical region (Fig. 4). However, in temperate and boreal regions, the significantly changed Rs was more commonly located in areas with significant changes in SV cover than in areas with significant climate change (Fig. 4). Separating both effects (Fig. 5), climate change accounted for the majority of the observed Rs change at the global scale (56%) and in tropical regions (66%), but land-cover change contributed the most to the Rs change in temperate (58%) and boreal regions (55%).
Fig. 3. Relationships between spatially averaged annual Rs and six driving factors from 2000 to 2014.
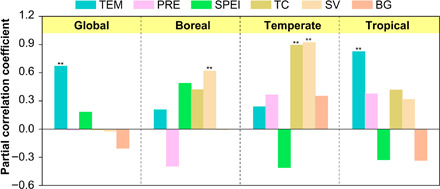
The plot shows partial correlation coefficient between spatially averaged annual Rs and each of the six driving factors at global and regional scales. The six driving factors are annual mean air temperature (TEM), annual precipitation (PRE), annual mean standardized precipitation-evapotranspiration (ET) index (SPEI), tree canopy (TC) cover, short vegetation (SV) cover, and bare ground (BG) cover. All variables (i.e., 15-year spatially averaged annual Rs, TEM, PRE, SPEI, TC cover, SV cover, and BG cover at the global and regional scales) were detrended before conducting partial correlation analysis. Two asterisks indicate that the partial correlation is significant at the 0.05 level (two-tailed).
Fig. 4. Proportion of colocated annual Rs change and each of the six driving factors at global and regional scales from 2000 to 2014.
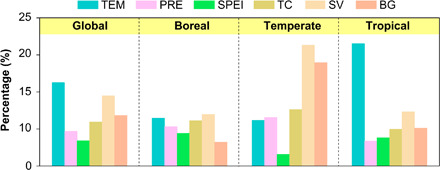
The plot represents the percentage of the areas with both significant climate (or land-cover) change and significant annual Rs change to the areas with significant annual Rs change. Climate factors include TEM, PRE, and SPEI. Land-cover factors include TC cover, SV cover, and BG cover. This plot indicates that the significantly changed Rs in temperate and boreal regions was more commonly located in areas with significant changes in SV cover than in areas with significant climate change.
Fig. 5. Contributions of climate change and land-cover change to changes in annual Rs from 2000 to 2014.
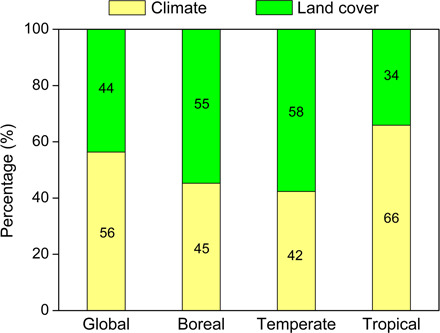
The plot indicates that climate change dominates global Rs change, but it does not have consistent influence on Rs change at the regional scale.
DISCUSSION
Comparison to previous studies on global Rs
Differences in estimated mean annual global Rs observed between recent studies and the present work (table S1) can be explained largely by differences in the spatial extent of study areas. Our study does not consider Rs from the permanent bare ground (BG) land-cover types (LCTs) because there are very few measurements of the annual Rs from bare lands. On the basis of the Moderate Resolution Imaging Spectroradiometer (MODIS) LCT data, global BG lands have a mean area of 20.6 × 106 km2 between 2000 and 2014. If the annual Rs of BG lands were 421.3 ± 167.9 g C m−2 year−1 (2), then mean annual Rs of global BG lands would range from 5.2 to 12.1 Pg C year−1. This result explains, to some extent, the underestimation of our global annual Rs and demonstrated that the estimated annual global Rs is likely in line with previous estimates.
The increasing global Rs over the period 2000–2014 was consistent with an increasing trend in global Rh in the periods 1998–2012 (29) and 2000–2015 (30). Tropical regions contributing the largest proportion to global Rs change is consistent with earlier studies (9, 11). Rs in the temperate region showed a nonsignificant decreasing trend between 2000 and 2014 (Fig. 2A), which was different from the temperature-associated significant increases in temperate Rs found by previous studies (9, 11). The differences in the time period considered for these estimates (table S1) could partially explain this result. The relatively greater Rs increase in the boreal region may be explained by the larger magnitude of temperature increase in this region (fig. S3A) because the cold regions exhibited a higher Rs temperature sensitivity and larger carbon stock than those of warm regions (31–33).
Global Rs in relation to climate and land-cover changes
On a global scale, Rs correlated significantly with TEM (Fig. 3). This result was consistent with the findings of previous studies (9, 11), which exhibited an increasing temporal trend for the annual global Rs, primarily driven by air temperature anomalies. Temperature also played an important role in explaining the interannual variation in Rs in the tropical region (Fig. 3). The positive response of Rs to temperature in the tropical region supported the findings of Fernández-Martínez et al. (33) that increasing temperature reduced the carbon sink capacity of the tropical region because of a greater stimulation of ecosystem respiration than photosynthesis at higher temperatures (34). However, vegetation change in SV showed stronger link with Rs than climate change in both boreal and temperate regions (Fig. 3). Furthermore, the globally dominant, coupled land-cover changes are the changes in tree canopy (TC) cover colocated with changes in SV and changes in SV colocated with BG (fig. S4) (35). The close link between TC and SV cover (table S5) explained why the partial correlation coefficient between Rs and SV cover had a similar magnitude as that between Rs and TC cover at the regional scale, especially in the temperate region (Fig. 3).
Overall, although climate change dominated global Rs change, it did not have consistent influences on the change in Rs for the period of 2000–2014 at the regional scale (Fig. 5). Land-cover change, especially changes in SV cover, exerted more effects on Rs in both boreal and temperate regions than in tropical regions (Figs. 3 and 4). However, at the pixel scale, the specific Rs drivers are diverse and interactive (36), as discussed in detail below.
Distinct effects of climate change on Rs change
In most tropical regions, including the Amazon Basin, Congo Basin, India, Myanmar, and large areas of Australia, the widespread increase in Rs might be attributed to increased temperature (Fig. 2B and fig. S5A), which was further verified by the positive partial correlation between annual Rs and TEM at the pixel scales (Fig. 6A). However, increased Rs in central and eastern China (Fig. 2B) may be due to increased temperature and precipitation (fig. S5, A and B). A slightly increased annual mean standardized precipitation-evapotranspiration (ET) index (SPEI) (fig. S3C) may explain the spatial hot spots of increased Rs in the western United States where limiting soil moisture has been constraining Rs (37).
Fig. 6. Spatial patterns of partial correlation coefficient between annual Rs and its six driving factors from 2000 to 2014.
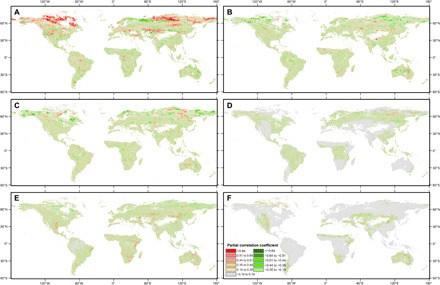
Partial correlation coefficient (R) between detrended annual Rs and detrended driving factors are shown in (A) Rs and TEM, (B) Rs and PRE, (C) Rs and SPEI, (D) Rs and TC cover, (E) Rs and SV cover, and (F) Rs and BG cover. R = ±0.64, R = ±0.51, R = ±0.44, R = ±0.35, and R = ±0.19 correspond to the 0.01, 0.05, 0.1, 0.2, and 0.5 significance levels, respectively. To reduce the effects from the data acquisition error in the land-cover data, only the per-pixel percent cover of TC, SV, and BG greater than 25% (61) was used to conduct the partial correlation analysis.
Some previous studies (11, 15) have shown that warmer air temperatures are associated with lower Rs values in the boreal region. However, our results found that annual Rs showed a strong positive partial correlation with TEM throughout most of the northern high latitudes (over 55°N) and the Tibetan Plateau (Fig. 6A). A pixel-based partial correlation analysis revealed that 65.6% of the boreal region showed a positive partial correlation between annual Rs and TEM, and 17.8% was statistically significant at the 95% confidence level (P < 0.05; Fig. 6A). This result supports earlier findings that air temperature is one of the dominant factors constraining Rs in high-latitude and high-altitude carbon cycling (3, 33).
Complex regional interactive effects of climate and land-cover changes on Rs
Hot spots of increasing Rs, found spatially coincident with increases in SV cover, SPEI, and annual precipitation (PRE) and decreases in BG cover, were observed in northern Canada, northern Mongolia, Loess Plateau of China, India, and eastern Australia (Fig. 2B and figs. S4, B and C, and S5, B and C). The widespread distribution of positive partial correlation between Rs and each of the four variables (i.e., SV cover, SPEI, PRE, and BG cover) also supported this finding (Fig. 6, B to F, respectively). This suggests that an increase in SV cover from BG cover coupled with increased moisture jointly drive the increase in Rs through increasing carbon inputs to the soil (28).
Most of the increasing Rs also showed a significant positive partial correlation with interannual variations in SPEI, TC, and SV cover in the boreal region, such as central Russia and northwest Canada (Fig. 6, C to E, respectively). These two areas experienced a widespread increase in SPEI and TC cover and decrease in SV cover (figs. S4, A and B, and S5C), which were probably related to woody expansion (27) and permafrost thawing (38) induced by climate warming. This result indicates the important effect of moisture on Rs (16) and that transformation of SV into woody vegetation may increase Rs in the boreal region (15). However, central Quebec, Canada also experienced widespread SV loss and TC gain, but Rs in this area showed a decreasing trend (Fig. 2B and fig. S4, A and B), possibly due to the effect of drought stress (39). The alteration of forests and woodlands by deforestation into cultivation is pervasive across tropical regions (40), which could increase soil temperature and thus increase Rs (41).
Decreasing Rs have been observed to be widely distributed in water-limited areas (42, 43), including the central United States, western Europe, northeast China, Kazakhstan, Argentina, east Brazil, east Africa, south Africa, and western Australia (Fig. 2B). These drought-prone areas spatially matched the areas with decreasing trends in SV cover (fig. S4B), with most of these areas corresponding to the areas with decreasing trends in PRE and SPEI (fig. S5, B and C) and some of these areas (i.e., Kazakhstan, Argentina, east Brazil, east Africa, and south Africa) corresponding to widespread increases in BG cover (fig. S4C). The widely distributed positive partial correlations between Rs and SV (or PRE or SPEI; Fig. 6, B, C, and E) in these areas indicated that SV loss and decreased water availability jointly determined the decrease in Rs in these water-limited regions. This phenomenon was particularly pronounced in the temperate region (Fig. 6, B, C, and E), which could help explain the decreased Rs trend in this region (Fig. 2A). The mechanism behind these changes may be attributed to the response of Rs to drought. Drought can lead to a significant decline in vegetation productivity by reducing water availability for plant growth (44), inhibiting soil Rh and extracellular enzymes diffusion (45), and thus inducing a substantial reduction in Rs (16).
Limitations and future work
This analysis has a number of limitations. First, a lack of representativeness of in situ observations of annual Rs is the largest source of uncertainty in the global Rs estimation using bottom-up methods (7). Our study is based on an integrated global dataset of Rs, which had more records from temperate regions but a distinct lack of field data from boreal and tropical regions (fig. S1). Without proper correction of the effects of spatial sampling, the heterogeneous distribution of the measurements may bias global Rs estimates toward the rates of the most sampled biomes. Thus, our method carries uncertainties, particularly in the globally undersampled regions. Future efforts to improve our method for Rs estimation would likely benefit most from reduced uncertainty in sampling error. In this study, we established the optimal statistical model for each biome by a combination of remote sensing data and in situ data and then used remote sensing data to estimate global Rs. This approach can efficiently reduce spatial sampling error because use of space-based remote sensing techniques can reduce sampling bias in boreal and tropical regions by providing dense sampling in space and time to characterize the heterogeneity of specific ecosystem properties and parameters [e.g., (46)].
Second, these selected biome-specific models explained 62 to 84% of the spatiotemporal variability in the global Rs (table S3), highlighting the limitations of the input datasets and the resulting empirical models in accurately predicting global Rs spatiotemporal patterns. Despite the uncertainties in the remote sensing datasets used for global Rs estimation (table S3), our approach has the potential to provide better estimates of global Rs with improvements to remote sensing–based estimates of the input variables [e.g., gross primary production, land surface temperature (LST), and vegetation index (VI)] (20). To better understand spatial and temporal variability of global Rs, more information about the uncertainty in the remote sensing inputs (table S3) and driving datasets (i.e., climate and land-cover data) is needed.
Furthermore, multiple uncertainties remain in modeling global Rs. These uncertainties may be due to the omission of potentially important factors, such as transition periods from a land cover to another (24, 25), nitrogen deposition (47), increasing atmospheric CO2 (48), soil organic carbon (49), and ecological disturbances (50). Last, data at subannual time scales (e.g., monthly and daily data) are important for upscaling global Rs (17). Estimation of global Rs at a temporal resolution finer than the current annual time scale may derive different conclusions and deserves further exploration.
CONCLUSIONS
Identifying the factors that affect the contribution of the soil surface CO2 efflux to the atmosphere will help increase confidence in future projections of the terrestrial carbon cycle in response to global climate change. Until recently, the analysis on driving forces of Rs change was largely limited to climate factors (9, 10, 15), and the role of land-cover change was rarely investigated. In this study, global annual Rs estimates were obtained from satellite data–driven statistical models that were well constrained by 1292 site years of in situ measurements of Rs at a per-biome scale. Consistent with previous studies, we found an increasing trend in global Rs due to climate change (9, 11, 15), and our results supported earlier findings that increased temperatures were negatively associated with the terrestrial C sink (33). However, unlike earlier analyses, our study, using statistical models and 1 km by 1 km remote sensing data, showed that land-cover change played a major role in driving changes in annual Rs in boreal and temperate regions. This result further confirmed that considering only climate change could not adequately explain the spatial and temporal variations in Rs, especially in temperate and boreal regions. We caution that there are uncertainties in the estimations of global Rs, such as sampling error, uncertainties in remotely sensed input data, ignoring potentially important factors, and the lack of subannual data. Further efforts should be made toward reducing these uncertainties.
MATERIALS AND METHODS
The data, statistical prediction models, and analytical approaches used in this study are described here in sufficient detail to understand the analysis. Additional methodological details needed to reproduce the results are presented in the Supplementary Materials.
Data
Integration of field-based Rs databases
Three available Rs databases covering global and regional scales (51–53) were collected to generate a centralized field database. These existing datasets were derived on the basis of comprehensive literature surveys of field (not laboratory) Rs measurements. To guarantee data quality for our analysis and to eliminate the mismatch between field-measured Rs and satellite remote sensing data, we applied the following selective criteria to the available records to create the integrated field database: (i) The database strictly focused on annual (i.e., year-round) Rs measurements, rather than seasonal (e.g., during the growing season) measurements. (ii) Only studies using an infrared gas analyzer or gas chromatograph were selected, and annual Rs values collected from nonagricultural biomes were limited to those without experimental manipulation. (iii) Some low-quality data records were removed, such as those from studies where the annual Rs value was extrapolated from low-frequency measurements or obtained from other sources (not original measurements). (iv) When more than one set of year-round Rs measurements were made at one site and for 1 year, those data were averaged to estimate the mean annual Rs for that site and year. As a general rule, averaging was avoided among different vegetation types. For instance, when the records included data from more than one location at the same site (i.e., site with different vegetation cover types), each location corresponding to a vegetation type was incorporated separately into our database. In addition to the annual Rs estimates, other site variables, such as latitude, longitude, study year, TEM, PRE, and vegetation type, were also included in the integrated database. The final database used in this study contained 1292 annual Rs observations collected from 701 sites from 2000 to 2014. These sites were distributed from 43°12′S to 78°10.20′N and from 155°56.40′W to 175°46.20′E, covering most of the global biomes and climate zones (fig. S1).
Remote sensing data products
We collected the global LST, gross primary productivity (GPP), VI, ET, and LCT products from the available MODIS data. Among these products, the 1-km MOD11A2 LST product (https://lpdaac.usgs.gov/products/mod11a2v006/), 500-m MOD17A2H GPP product (https://lpdaac.usgs.gov/products/mod17a2hv006/), 500-m MOD13A1 VI product (https://lpdaac.usgs.gov/products/mod13a1v006/), and 500-m MOD16A2 ET product (https://lpdaac.usgs.gov/products/mod16a2v006/) provided 8-day average values for every year between 2000 and 2014. The MCD12Q1 LCT product with a 500-m spatial resolution (https://lpdaac.usgs.gov/products/mcd12q1v006/) provided annual values for 2001 to 2014. We collected root-zone soil moisture (RZSM) products from the assimilation of the NASA Advanced Microwave Scanning Radiometer (AMSR-E) data and a two-layer Palmer water balance model (23), with global coverage from June 2002 to December 2010 at a spatial resolution of 25 km and a temporal resolution of 1 day. We included the Tropical Rainfall Measuring Mission (TRMM) 3B43 gridded global precipitation product with a monthly temporal resolution and 0.25° by 0.25° spatial resolution for 2000 to 2014 (54).
The LCT data in 2000 were missing in the LCT product, and for this, we substituted LCT data for 2001. The spatial resolution of the remote sensing data products is diverse, ranging from 500 m to 25 km. For site-scale analyses, we selected the pixels centered on each site from the remote sensing data products by a nearest-neighbor algorithm to match the geographic coordinates (latitude and longitude) of the Rs observation sites. Low-quality data were removed using the available quality flags in each data product. For the spatial analysis at the global scale, we created a 1 km by 1 km spatial resolution dataset from the original remote sensing data products based on nearest-neighbor resampling. All satellite remote sensing data products with 1-day, 8-day, and monthly temporal resolutions were averaged to obtain the corresponding values at annual, spring, and summer time scales.
Climate and land-cover change data
We extracted gridded monthly air temperature and precipitation time series data at a resolution of 0.5° by 0.5° from the website of the Center for Climate Research, University of Delaware (http://climate.geog.udel.edu/~climate/html_pages/download.html#ghcn_T_P_clim2). We also used global standardized precipitation-ET index data (http://SPEI.csic.es/database.html) as a measure of drought intensity (spatial resolution, 0.5° by 0.5°). Three climatic variables—TEM, PRE, and SPEI—were calculated for each year and each pixel from 2000 to 2014 over the globe to analyze the response of Rs to climate change. In addition, we used an annual global land-cover product by Song et al. (35) to analyze the response of Rs to land-cover change. This land-cover product provided proportional estimates of TC cover, SV cover, and BG cover at a 0.05° by 0.05° spatial resolution over the globe for each year during 2000–2014. Trees are defined as all vegetation taller than 5 m in height. SV refers to all vegetation other than trees, including shrubs, herbaceous vegetation, and mosses, while BG represents the proportion of the land surface not covered by vegetation.
Available predictive variables for annual Rs estimation
Our model development assumes that temperature, moisture, and plant productivity are the predominant drivers of the spatiotemporal variation in global Rs. Remotely sensed LST has shown great potential in estimating Rs at the site and regional scales (19). In this study, temperature variables include MOD11A2 LST measurements during the daytime (LST_day) and nighttime (LST_night). Moisture availability, as it relates to temperature, may be better addressed using data representing actual ET or the ratio of actual to potential ET (ET_PET) (16) and can be accounted for via the difference between LST_day and LST_night (18). Thus, four parameters are used as a surrogate for soil moisture status, including the remotely sensed precipitation from TRMM 3B43 (RS_pre), difference between LST_day and LST_night (LST_diff), ET and ET_PET from MOD16A2, and RZSM, which were extracted from our collected remote sensing data products. GPP and VI can be used as quantitative indicators of plant productivity to estimate Rs (19). Thus, this study used the MOD17A2H GPP, enhanced VI (EVI), and normalized difference VI (NDVI) from MOD13A1 as measures of plant productivity.
Because annual Rs is affected by various biotic and abiotic factors at annual and seasonal time scales (55), this study used predictive variables from remotely sensed temperature, moisture, and plant productivity variables at annual, spring, and summer time scales for the estimation of global Rs. In this study, spring and summer correspond to March to May and June to August, respectively. In addition, the remotely sensed temperature variables used in this study include LST_day_annual, LST_day_spring, LST_day_summer, LST_night_annual, LST_night_spring, and LST_night_summer. The moisture variables include RS_pre_annual, RS_pre_spring, RS_pre_summer, LST_diff_annual, LST_diff_spring, LST_diff_summer, ET_annual, ET_spring, ET_summer, ET_PET_annual, ET_PET_spring, ET_PET_summer, RZSM_annual, RZSM_spring, and RZSM_summer. The plant productivity variables include GPP_annual, GPP_spring, GPP_summer, EVI_annual, EVI_spring, EVI_summer, NDVI_annual, NDVI_spring, and NDVI_summer.
Rs rates vary among major biomes because of the interactions between climate and vegetation at the global scale (14, 15). Therefore, it is necessary to establish the models at the biome scale for accurately estimating global Rs. Furthermore, the large dataset available from field measurements and satellite remote sensing technology at specific biomes enable the development of biome-specific models, which could be applied to provide global coverage. Thus, comprehensively considering the similarities in climate and vegetation types and the minimum number of data points needed for constructing a statistical model, we divided the globe into 10 biomes: temperate evergreen needleleaf forest, temperate evergreen broadleaf forest, temperate deciduous needleleaf forest, temperate deciduous broadleaf forest, temperate mixed forest, boreal vegetation region, tropical forest, grasslands, croplands, and shrublands. To achieve this analysis, we used global climatic data from the European Soil Data Centre (https://esdac.jrc.ec.europa.eu/) defined on the basis of the climatic classification of the Intergovernmental Panel on Climate Change (2006). The vegetation type data used are MCD12Q1 LCT data from 2000 to 2014.
Nonlinear regression models
The well-known Q10 model (Eq. 1) has been frequently used to describe the relationship between annual Rs (g C m−2 year−1) and temperature. The simple exponential temperature model has been widely applied to the modeling of the temperature sensitivity of Rs at the global scale (10, 12). However, a common criticism of the Q10 model is the fixed temperature sensitivity (Q10 value, the factor by which the respiration rate increases for a temperature increase of 10°C), which violates the fact that the Q10 value decreases with an increase in temperature. In this study, we used a more flexible second-order polynomial temperature model (Eq. 2), which has been reported to behave better than the simple exponential temperature model in modeling global Rs (9)
| (1) |
| (2) |
A quadratic model (Eq. 3) was used to quantify the dependence of annual Rs on moisture availability, which considered a reduction in microbial decomposition at very low or very high soil moisture content (16)
| (3) |
A linear model (Eq. 4) was used to describe the dependency of the annual Rs on plant productivity (7)
| (4) |
where Rs is the annual soil respiration (g C m−2 year−1), T is a temperature-related variable, and M is a variable referring to moisture availability. P is a quantitative indicator of plant productivity. θ1,…, θn are model parameters and differ among models. The parameters in nonlinear regression models are estimated by nonlinearly minimizing the sum of the squared residuals.
Predictive variables selected for estimating annual Rs
First, the abovementioned three types of variables (i.e., those related to temperature, moisture, and plant productivity) from remote sensing data products were used to implement nonlinear regressions (i.e., Eqs. 2 to 4). There could be a high correlation among the variables within the same type. As the spatiotemporal coverage and data quality of these global remote sensing data products were different, the available observation data for the fitting models (i.e., Eqs. 2 to 4) were different when one model from Eqs. 2 to 4 was driven by different input variables. To select the optimal variables for Rs estimations in Eqs. 2 to 4 for each biome, a performance metric (ρ) for model evaluation (Eq. 5) was used
| (5) |
where n is the number of observation data. RMSEmin is the minimum RMSE from one model driven by different input variables (i.e., temperature, moisture, or plant productivity variables) for one biome, and and nmax are the corresponding maximum R2 and n, respectively. ρ is a measure of the model’s explanatory power (i.e., and) and data representation () for one parameterized model for one biome. The weighting factor for data representation (i.e., 0.5) is greater than that for the model’s explanatory power (i.e., 0.25) because we have more confidence in the performance of a model with large n and such a model will tend to have a better spatiotemporal representation.
For each model, with Eqs. 2 to 4, each variable within the same type (temperature, moisture, or plant productivity) was used as one predictor, and the variable with the maximum ρ was selected for each biome. When the optimal temperature variable in Eq. 2 was determined, it was used in Eq. 3. The same data processing was used for Eq. 3 to select the optimal moisture variable. Then, the selected temperature and moisture variables were used in an MNLR model (Eq. 4), and the optimal plant productivity variable was determined for each biome. By gradually adding the optimal inputs from Eqs. 2 to 4, to what extent additional predictor variables would improve the model performance at the biome scale, and the importance of remotely sensed variables, could be determined. To ensure comparisons with the machine learning algorithm models, the same predictive variables (the optimal temperature, moisture, and plant productivity) were maintained for the analysis of the machine learning methods.
Applying machine learning algorithm models
Random forest regression
RFR is an ensemble machine learning algorithm that predicts a response from a set of predictors by creating multiple decision trees and aggregating their results (56). Each tree is constructed by a randomly selected subset of training data. The remaining training data, which are called out-of-bag data, are used to estimate the prediction error and variable importance.
Support vector regression
SVR, a regression version of support vector machine algorithms, with an improved generalization ability, uses unique and globally optimal architecture and can be rapidly trained (57). SVR projects the input space data into a feature space with a much larger dimension, enabling linearly nonseparable data to become separable in the feature space. It identifies optimum hyperplanes by using kernel functions and arrives at an optimum solution by iteratively adjusting the hyperplanes on the basis of their associated errors.
Artificial neural network
An ANN is a parallel-distributed information processing system that simulates the actions of neurons in the human brain and is able to learn from examples (58). In an ANN, information flows in a unidirectional forward mode from an input layer to an output layer via hidden layer(s). Network connection weights are adjusted if the separation of inputs incurs an error during training, and convergence proceeds until the reduction in error between iterations reaches a decay threshold.
Parameter optimizations of the three machine learning algorithm models
The RFR, SVR, and ANN parameters within a certain range (i.e., different numbers or types) were adjusted and tested to ensure that their performance was optimal. For the RFR model, the parameter set included the number of trees (NumTrees), a predictor-splitting algorithm, and the number of predictors to select at random for each split (NumPredictorsToSample). For SVR, the parameter set included half of the width of the epsilon-insensitive band (Epsilon), the flag to standardize the predictor data (Standardize), an optimization routine (Solver), a kernel scale parameter (KernelScale), and a kernel function (KernelFunction). For ANN, the parameter set included training algorithms (trainFcn), the number of hidden layer (N), and hidden layer size (hiddenSizes). An internationally recognized uniform method for parameter optimization for these machine learning algorithm models has not been established. In this study, a trial-and-error process was used to select the optimal parameters by grid search method, which results in the best performance model with the highest R2 and lowest RMSE.
Global Rs estimation
A 10-fold cross-validation (59) was used to evaluate the performance of the four statistical models for annual Rs estimation of each biome. One measured Rs dataset for each biome type was randomly partitioned into 10 equal-sized subsets. During each model fitting, one of the partitions was reserved for validation, while the other nine were used for training. This modeling process was repeated 10 times, and the performance metrics including R2 and RMSE were averaged to describe the final performance. For each biome, the model with the highest R2 and lowest RMSE was selected as the optimal model for annual Rs estimation. To confirm the uncertainties associated with the structure and parameters of the selected models, a Monte Carlo approach was used to propagate the model errors to the global estimates. A normal distribution with a 10% coefficient of variance was assumed for each quantitative parameter (13). Random sampling from all candidate values was used to analyze the uncertainty of qualitative parameters. For each trial (N = 500), new parameters were chosen from the uncertainty for each parameter, and a group of new models for the 10 biomes were generated to compute the annual global Rs for each year from 2000 to 2014. The means and 95% CIs of all the estimates of the annual global Rs were calculated to represent the model uncertainties. Notably, our “global” study area included only the regions with vegetation according to the definitions of the remotely sensed LCT products. Bare lands, water bodies, urban lands, and built-up lands were not included in our study area.
Trend and attribution analysis
The temporal variation in Rs from 2000 to 2014 was characterized by combining the results of a Mann-Kendall test and a Theil-Sen median trend analysis. The trend uncertainty was estimated by a 1000-bootstrap analysis. If the Mann Kendall test result was statistically significant (P < 0.05), then we applied the Theil-Sen estimator to derive the slope (annual change) of trend and provide the estimate of Rs net change between 2000 and 2014 (i.e., slope times 14 years). The direct response of Rs to climate and land-cover changes was examined using the partial correlation analysis between Rs and potential driving factors [i.e., climate (TEM, PRE, and SPEI) and land-cover (TC, SV, and BG cover) factors]. The partial correlation coefficient represents the correlation of each pair of variables after statistically controlling for all the other variables. Before conducting the partial correlation analysis, we first detrended all time series data, including Rs, the climate variables and the land-cover fractions, to avoid autocorrelation and to make the time series stationary. To achieve this, the linear trend derived from the least-squares method was removed for both Rs and its driving factors. All these analyses were performed at the global, regional, and pixel scales. To compare with previous studies on global and regional Rs estimates, we defined three regions based on annual air temperature following the method of Bond-Lamberty and Thomson (11): tropical [annual air temperature (T) > 17°C], temperate (2°C ≤ T ≤ 17°C), and boreal (T < 2°C).
The independent effects of climate and land-cover changes on the spatial and temporal variations in Rs were difficult to separate because these variables covaried at different scales (60). For instance, climate change is known to control Rs because of its close link with environmental factors (3, 10, 12), such as soil temperature, soil moisture, and substrate quality, while land-cover change (e.g., grassland to woodland) can be expected to change some or all of these environmental determinants of Rs (24, 25). To analyze the contributions of climate and land-cover changes to the observed global Rs change, land-cover changes between 2000 and 2014 were first derived using Mann-Kendall test and Theil-Sen median trend analysis, following the method of Song et al. (35). To approximately quantify the impact of climate change and land-cover change, we assumed that changes in Rs colocated with significant land-cover change (two-sided Mann-Kendall test, P < 0.05) were attributable to land-cover change, whereas Rs changes that occurred without land-cover change were attributable to climate change. Rs net change at different spatial scale was calculated by summing the per-pixel absolute Rs net change at the corresponding spatial scale. For example, Rs net change for one land-cover change type was calculated by summing the per-pixel absolute Rs net change over this land-cover change type. Last, the overall impact of land-cover change was defined as the proportion of the Rs net change in all significant land-cover change types. The overall climate change impact was defined as the residual of the land-cover change impact. All these analyses were conducted at global and regional scales.
Supplementary Material
Acknowledgments
Funding: This research was funded by the National Key Research and Development Program of China (no. 2017YFA0603002), the National Natural Science Foundation of China (nos. 41771465 and 41871347), and the Strategic Priority Research Program of the Chinese Academy of Sciences (no. XDA19030404). Author contributions: N.H., L.W., and Z.N. designed the study and analysis. N.H. performed the Rs model runs, analyzed the data, and wrote the manuscript. All authors contributed substantially to the data processing, writing, and discussion of the paper. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data processing and statistical analyses were conducted using MATLAB (MathWorks, Natick, MA). All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Code and raster datasets of model output have been deposited into the Dryad Data Repository at https://doi.org/10.5061/dryad.w3r2280nq. Additional data related to this paper maybe requested from the authors.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/6/41/eabb8508/DC1
REFERENCES AND NOTES
- 1.Friedlingstein P., Jones M. W., O’Sullivan M., Andrew R. M., Hauck J., Peters G. P., Peters W., Pongratz J., Sitch S., Le Quéré C., Bakker D. C. E., Canadell J. G., Ciais P., Jackson R. B., Anthoni P., Barbero L., Bastos A., Bastrikov V., Becker M., Bopp L., Buitenhuis E., Chandra N., Chevallier F., Chini L. P., Currie K. I., Feely R. A., Gehlen M., Gilfillan D., Gkritzalis T., Goll D. S., Gruber N., Gutekunst S., Harris I., Haverd V., Houghton R. A., Hurtt G., Ilyina T., Jain A. K., Joetzjer E., Kaplan J. O., Kato E., Klein Goldewijk K., Korsbakken J. I., Landschützer P., Lauvset S. K., Lefèvre N., Lenton A., Lienert S., Lombardozzi D., Marland G., McGuire P. C., Melton J. R., Metzl N., Munro D. R., Nabel J. E. M. S., Nakaoka S., Neill C., Omar A. M., Ono T., Peregon A., Pierrot D., Poulter B., Rehder G., Resplandy L., Robertson E., Rödenbeck C., Séférian R., Schwinger J., Smith N., Tans P. P., Tian H., Tilbrook B., Tubiello F. N., van der Werf G. R., Wiltshire A. J., Zaehle S., Global carbon budget 2019. Earth Syst. Sci. Data 11, 1783–1838 (2019). [Google Scholar]
- 2.Warner D. L., Bond-Lamberty B., Jian J., Stell E., Vargas R., Spatial predictions and associated uncertainty of annual soil respiration at the global scale. Glob. Biogeochem. Cycles 33, 1733–1745 (2019). [Google Scholar]
- 3.Tang X., Fan S., Du M., Zhang W., Gao S., Liu S., Chen G., Yu Z., Yang W., Spatial-and temporal-patterns of global soil heterotrophic respiration in terrestrial ecosystems. Earth Syst. Sci. Data 12, 1037–1051 (2020). [Google Scholar]
- 4.Bond-Lamberty B., New techniques and data for understanding the global soil respiration flux. Earth Future 6, 1176–1180 (2018). [Google Scholar]
- 5.Anav A., Friedlingstein P., Beer C., Ciais P., Harper A., Jones C., Murray-Tortarolo G., Papale D., Parazoo N. C., Peylin P., Piao S., Sitch S., Viovy N., Wiltshire A., Zhao M., Spatiotemporal patterns of terrestrial gross primary production: A review. Rev. Geophys. 53, 785–818 (2015). [Google Scholar]
- 6.Konings A. G., Bloom A. A., Liu J., Parazoo N. C., Schimel D. S., Bowman K. W., Global satellite-driven estimates of heterotrophic respiration. Biogeosciences 16, 2269–2284 (2019). [Google Scholar]
- 7.Xu M., Shang H., Contribution of soil respiration to the global carbon equation. J. Plant Physiol. 203, 16–28 (2016). [DOI] [PubMed] [Google Scholar]
- 8.Phillips C. L., Bond-Lamberty B., Desai A. R., Lavoie M., Risk D., Tang J. W., Todd-Brown K., Vargas R., The value of soil respiration measurements for interpreting and modeling terrestrial carbon cycling. Plant Soil 413, 1–25 (2017). [Google Scholar]
- 9.Hashimoto S., Carvalhais N., Ito A., Migliavacca M., Nishina K., Reichstein M., Global spatiotemporal distribution of soil respiration modeled using a global database. Biogeosciences 12, 4121–4132 (2015). [Google Scholar]
- 10.Raich J. W., Potter C. S., Bhagawati D., Interannual variability in global soil respiration, 1980-94. Glob. Chang. Biol. 8, 800–812 (2002). [Google Scholar]
- 11.Bond-Lamberty B., Thomson A., Temperature-associated increases in the global soil respiration record. Nature 464, 579–582 (2010). [DOI] [PubMed] [Google Scholar]
- 12.Chen S., Huang Y., Xie W., Zou J., Lu Y., Hu Z., A new estimate of global soil respiration from 1970 to 2008. Chin. Sci. Bull. 58, 4153–4160 (2013). [Google Scholar]
- 13.Hashimoto S., A new estimation of global soil greenhouse gas fluxes using a simple data-oriented model. PLOS ONE 7, e41962 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Adachi M., Ito A., Yonemura S., Takeuchi W., Estimation of global soil respiration by accounting for land-use changes derived from remote sensing data. J. Environ. Manag. 200, 97–104 (2017). [DOI] [PubMed] [Google Scholar]
- 15.Zhao Z., Peng C., Yang Q., Meng F.-R., Song X., Chen S., Epule T., Li P., Zhu Q., Model prediction of biome-specific global soil respiration from 1960 to 2012. Earth Future 5, 715–729 (2017). [Google Scholar]
- 16.Hursh A., Ballantyne A., Cooper L., Maneta M., Kimball J., Watts J., The sensitivity of soil respiration to soil temperature, moisture, and carbon supply at the global scale. Glob. Chang. Biol. 23, 2090–2103 (2017). [DOI] [PubMed] [Google Scholar]
- 17.Jian J. S., Steele M. K., Thomas R. Q., Day S. D., Hodges S. C., Constraining estimates of global soil respiration by quantifying sources of variability. Glob. Chang. Biol. 24, 4143–4159 (2018). [DOI] [PubMed] [Google Scholar]
- 18.Jägermeyr J., Gerten D., Lucht W., Hostert P., Migliavacca M., Nemani R., A high-resolution approach to estimating ecosystem respiration at continental scales using operational satellite data. Glob. Chang. Biol. 20, 1191–1210 (2014). [DOI] [PubMed] [Google Scholar]
- 19.Huang N., Gu L. H., Black T. A., Wang L., Niu Z., Remote sensing-based estimation of annual soil respiration at two contrasting forest sites. J. Geophys. Res. 120, 2306–2325 (2015). [Google Scholar]
- 20.Xiao J., Chevallier F., Gomez C., Guanter L., Hicke J. A., Huete A. R., Ichii K., Ni W. J., Pang Y., Rahman A. F., Sun G., Yuan W., Zhang L., Zhang X., Remote sensing of the terrestrial carbon cycle: A review of advances over 50 years. Remote Sens. Environ. 233, 111383 (2019). [Google Scholar]
- 21.Running S. W., Thornton P. E., Nemani R., Glassy J. M., Global terrestrial gross and net primary productivity from the earth observing system. Methods Ecosyst. Sci. 44–57 (2000). [Google Scholar]
- 22.Wan Z. M., New refinements and validation of the MODIS Land-Surface Temperature/Emissivity products. Remote Sens. Environ. 112, 59–74 (2008). [Google Scholar]
- 23.Bolten J. D., Crow W. T., Zhan X. W., Jackson T. J., Reynolds C. A., Evaluating the utility of remotely sensed soil moisture retrievals for operational agricultural drought monitoring. IEEE J. Sel. Top. Appl. Earth Observ. 3, 57–66 (2010). [Google Scholar]
- 24.Alekseev A., Alekseeva T., Kalinin P., Hajnos M., Soils response to the land use and soil climatic gradients at ecosystem scale: Mineralogical and geochemical data. Soil Tillage Res. 180, 38–47 (2018). [Google Scholar]
- 25.Thomas A. D., Elliott D. R., Dougill A. J., Stringer L. C., Hoon S. R., Sen R., The influence of trees, shrubs, and grasses on microclimate, soil carbon, nitrogen, and CO2 efflux: Potential implications of shrub encroachment for Kalahari rangelands. Land Degrad. Dev. 29, 1306–1316 (2018). [Google Scholar]
- 26.Chen J. M., Ju W., Ciais P., Viovy N., Liu R., Liu Y., Lu X., Vegetation structural change since 1981 significantly enhanced the terrestrial carbon sink. Nat. Commun. 10, 4259 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Piao S., Wang X., Park T., Chen C., Lian X., He Y., Bjerke J. W., Chen A., Ciais P., Tømmervik H., Nemani R. R., Myneni R. B., Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 1, 14–17 (2020). [Google Scholar]
- 28.Mitra B., Miao G., Minick K., McNulty S. G., Sun G., Gavazzi M., King J. S., Noormets A., Disentangling the effects of temperature, moisture, and substrate availability on soil CO2 efflux. J. Geophys. Res. 124, 2060–2075 (2019). [Google Scholar]
- 29.Piao S., Huang M., Liu Z., Wang X., Ciais P., Canadell J. G., Wang K., Bastos A., Friedlingstein P., Houghton R. A., Quéré C. L., Liu Y., Myneni R. B., Peng S., Pongratz J., Sitch S., Yan T., Wang Y., Zhu Z., Wu D., Wang T., Lower land-use emissions responsible for increased net land carbon sink during the slow warming period. Nat. Geosci. 11, 739–743 (2018). [Google Scholar]
- 30.Bond-Lamberty B., Bailey V. L., Chen M., Gough C. M., Vargas R., Globally rising soil heterotrophic respiration over recent decades. Nature 560, 80–83 (2018). [DOI] [PubMed] [Google Scholar]
- 31.Lloyd J., Taylor J. A., On the temperature dependence of soil respiration. Funct. Ecol. 8, 315–323 (1994). [Google Scholar]
- 32.Koven C. D., Hugelius G., Lawrence D. M., Wieder W. R., Higher climatological temperature sensitivity of soil carbon in cold than warm climates. Nat. Clim. Chang. 7, 817–822 (2017). [Google Scholar]
- 33.Fernández-Martínez M., Sardans J., Chevallier F., Ciais P., Obersteiner M., Vicca S., Canadell J. G., Bastos A., Friedlingstein P., Sitch S., Piao S. L., Janssens I. A., Peñuelas J., Global trends in carbon sinks and their relationships with CO2 and temperature. Nat. Clim. Chang. 9, 73–82 (2019). [Google Scholar]
- 34.Liang J. Y., Xia J. Y., Liu L. L., Wan S. Q., Global patterns of the responses of leaf-level photosynthesis and respiration in terrestrial plants to experimental warming. J. Plant Ecol. 6, 437–447 (2013). [Google Scholar]
- 35.Song X.-P., Hansen M. C., Stehman S. V., Potapov P. V., Tyukavina A., Vermote E. F., Townshend J. R., Global land change from 1982 to 2016. Nature 560, 639–643 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Reynolds L. L., Johnson B. R., Pfeifer-Meister L., Bridgham S. D., Soil respiration response to climate change in Pacific Northwest prairies is mediated by a regional Mediterranean climate gradient. Glob. Chang. Biol. 21, 487–500 (2015). [DOI] [PubMed] [Google Scholar]
- 37.Thomey M. L., Collins S. L., Vargas R., Johnson J. E., Brown R. F., Natvig D. O., Friggens M. T., Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland. Glob. Chang. Biol. 17, 1505–1515 (2011). [Google Scholar]
- 38.Chadburn S. E., Burke E. J., Cox P. M., Friedlingstein P., Hugelius G., Westermann S., An observation-based constraint on permafrost loss as a function of global warming. Nat. Clim. Chang. 7, 340–344 (2017). [Google Scholar]
- 39.Girardin M. P., Hogg E. H., Bernier P. Y., Kurz W. A., Guo X. J., Cyr G., Negative impacts of high temperatures on growth of black spruce forests intensify with the anticipated climate warming. Glob. Chang. Biol. 22, 627–643 (2016). [DOI] [PubMed] [Google Scholar]
- 40.Tyukavina A., Baccini A., Hansen M. C., Potapov P. V., Stehman S. V., Houghton R. A., Krylov A. M., Turubanova S., Goetz S. J., Aboveground carbon loss in natural and managed tropical forests from 2000 to 2012. Environ. Res. Lett. 10, 074002 (2015). [Google Scholar]
- 41.Mapanda F., Mupini J., Wuta M., Nyamangara J., Rees R. M., A cross-ecosystem assessment of the effects of land cover and land use on soil emission of selected greenhouse gases and related soil properties in Zimbabwe. Eur. J. Soil Sci. 61, 721–733 (2010). [Google Scholar]
- 42.Dai A. G., Characteristics and trends in various forms of the Palmer Drought Severity Index during 1900-2008. J. Geophys. Res.-Atmos. 116, D12115 (2011). [Google Scholar]
- 43.Yuan W. P., Zheng Y., Piao S. L., Ciais P., Lombardozzi D., Wang Y. P., Ryu Y., Chen G. X., Dong W. J., Hu Z. M., Jain A. K., Jiang C. Y., Kato E., Li S. H., Lienert S., Liu S. G., Nabel J. E. M. S., Qin Z. C., Quine T., Sitch S., Smith W. K., Wang F., Wu C. Y., Xiao Z. Q., Yang S., Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 5, eaax1396 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Xu C., Mcdowell N., Fisher R. A., Wei L., Sevanto S., Christoffersen B., Weng E., Middleton R., Increasing impacts of extreme droughts on vegetation productivity under climate change. Nat. Clim. Chang. 9, 948–953 (2019). [Google Scholar]
- 45.Jassal R. S., Black T. A., Novak M. D., Gaumont-Guay D., Nesic Z., Effect of soil water stress on soil respiration and its temperature sensitivity in an 18-year-old temperate Douglas-fir stand. Glob. Chang. Biol. 14, 1305–1318 (2008). [Google Scholar]
- 46.Schimel D., Pavlick R., Fisher J. B., Asner G. P., Saatchi S., Townsend P., Miller C., Frankenberg C., Hibbard K., Cox P., Observing terrestrial ecosystems and the carbon cycle from space. Glob. Chang. Biol. 21, 1762–1776 (2015). [DOI] [PubMed] [Google Scholar]
- 47.Janssens I. A., Dieleman W., Luyssaert S., Subke J.-A., Reichstein M., Ceulemans R., Ciais P., Dolman A. J., Grace J., Matteucci G., Papale D., Piao S. L., Schulze E.-D., Tang J., Law B. E., Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 3, 315–322 (2010). [Google Scholar]
- 48.King J. S., Hanson P. J., Bernhardt E., DeAngelis P., Norby R. J., Pregitzer K. S., A multiyear synthesis of soil respiration responses to elevated atmospheric CO2 from four forest FACE experiments. Glob. Chang. Biol. 10, 1027–1042 (2004). [Google Scholar]
- 49.Leinweber P., Jandl G., Baum C., Eckhardt K. U., Kandeler E., Stability and composition of soil organic matter control respiration and soil enzyme activities. Soil Biol. Biochem. 40, 1496–1505 (2008). [Google Scholar]
- 50.Borkhuu B., Peckham S. D., Ewers B. E., Norton U., Pendall E., Does soil respiration decline following bark beetle induced forest mortality? Evidence from a lodgepole pine forest. Agric. For. Meteorol. 214, 201–207 (2015). [Google Scholar]
- 51.Bond-Lamberty B., Thomson A., A global database of soil respiration data. Biogeosciences 7, 1915–1926 (2010). [Google Scholar]
- 52.Chen S. T., Zou J. W., Hu Z. H., Chen H. S., Lu Y. Y., Global annual soil respiration in relation to climate, soil properties and vegetation characteristics: Summary of available data. Agric. For. Meteorol. 198, 335–346 (2014). [Google Scholar]
- 53.Epule T. E., A new compendium of soil respiration data for Africa. Challenges 6, 1–10 (2015). [Google Scholar]
- 54.Huffman G. J., Bolvin D. T., Nelkin E. J., Wolff D. B., Adler R. F., Gu G., Hong Y., Bowman K. P., Stocker E. F., The TRMM Multisatellite Precipitation Analysis (TMPA): Quasi-global, multiyear, combined-sensor precipitation estimates at fine scales. J. Hydrometeorol. 8, 38–55 (2010). [Google Scholar]
- 55.Domínguez M. T., Smith A. R., Reinsch S., Emmett B. A., Inter-annual variability of soil respiration in wet shrublands: Do plants modulate its sensitivity to climate? Ecosystems 20, 796–812 (2017). [Google Scholar]
- 56.Breiman L., Random forests. Mach. Learn. 45, 5–32 (2001). [Google Scholar]
- 57.Vapnik V. N., An overview of statistical learning theory. IEEE Trans. Neural Netw. 10, 988–999 (1999). [DOI] [PubMed] [Google Scholar]
- 58.R. Rojas, Neural Networks: A Systematic Introduction (Springer-Verlag, 1996). [Google Scholar]
- 59.Kohavi R., A study of cross-validation and bootstrap for accuracy estimation and model selection. Int. Joint Conf. Artif. Intell. 14, 1137–1145 (1995). [Google Scholar]
- 60.Linscheid N., Estupinan-Suarez L. M., Brenning A., Carvalhais N., Cremer F., Gans F., Rammig A., Reichstein M., Sierra C. A., Mahecha M. D., Towards a global understanding of vegetation–climate dynamics at multiple time scales. Biogeosciences 17, 945–962 (2020). [Google Scholar]
- 61.Vina A., Mcconnell W. J., Yang H., Xu Z., Liu J., Effects of conservation policy on China’s forest recovery. Sci. Adv. 2, e1500965 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/6/41/eabb8508/DC1