

Abstract
While feeding, mammalian browsers (primarily eat woody plants) encounter secondary metabolites such as tannins. Browsers may bind these tannins using salivary proteins, whereas mammalian grazers (primarily eat grasses that generally lack tannins) likely would not. Ruminant browsers rechew their food (ruminate) to increase the effectiveness of digestion, which may make them more effective at binding tannins than nonruminants. Few studies have included a sufficient number of species to consider possible scaling with body mass or phylogenetic effects on salivary proteins. Controlling for phylogeny, we ran inhibition radial diffusion assays of the saliva of 28 species of African herbivores that varied in size, feeding strategy, and digestive system. We could not detect the presence of salivary proline‐rich proteins that bind tannins in any of these species. However, using the inhibition radial diffusion assay, we found considerable abilities to cope with tannins in all species, albeit to varying degrees. We found no differences between browsers and grazers in the effectiveness of their salivary proteins to bind to and precipitate tannins, nor between ruminants and nonruminants, or scaling with body mass. Three species bound all tannins, but their feeding niches included one browser (gray duiker), one mixed feeder (bush pig), and one grazer (red hartebeest). Five closely related species of small ruminant browsers were very effective in binding tannins. Megaherbivores, considered generalists on account of their large body size, were capable of binding tannins. However, the grazing white rhinoceros was almost as effective at binding tannins as the megaherbivore browsers. We conclude, contrary to earlier predictions, that there were no differences in the relative salivary tannin‐binding capability that was related to common ancestry (phylogeny) or to differences in body size.
Keywords: African megaherbivores, Afrotheria, hindgut fermenters, ruminants, salivary proteins, tannins
Mammalian browsers frequently encounter secondary metabolites such as tannins. Browsers may bind these tannins using salivary proteins, while mammalian grazers likely would not. Controlling for phylogeny, we ran inhibition radial diffusion assays of the saliva of 28 species of African herbivores (browsers, grazers, ruminants, and nonruminants) that differed considerably in size. We conclude, contrary to earlier predictions, that there were no differences in the relative salivary tannin‐binding capability of browsers and grazers, nor ruminants versus nonruminants, that were related to common ancestry (phylogeny) or to differences in body size.

1. INTRODUCTION
Large mammalian herbivores foraging in savannas encounter both grasses and woody plants (Searle & Shipley, 2008). Grasses provide lower nutritional value than woody plants because they have a higher dietary fiber content, lower dry matter digestibility, and lower protein content (Demment & Van Soest, 1985; Owen‐Smith, 1982). Woody plants tend to invest less in cell walls but have relatively higher lignin contents than grasses (Ellis, 1990; Iason, Hodgson, & Barry, 1995; McNaughton, Tarrants, McNaughton, & Davis, 1985; Van Soest, 1994). Woody plants tend to defend themselves with plant secondary metabolites (PSMs) unlike grasses (Cooper & Owen‐Smith, 1985; Hofmann, 1989; Lundberg & Palo, 1993; Orians & Ward, 2010; Rohner & Ward, 1997). As a consequence of these fundamental differences between grasses and woody plants, many large mammalian herbivores are adapted to either be grazers (consume primarily (>70%) grasses and forbs) or browsers (consume primarily (>70%) woody plants) (Clauss, Hume, & Hummel, 2010; Clauss & Lechner‐Doll, 2001; Gordon & Illius, 1994; Hofmann, 1989; Hofmann & Stewart, 1972; Robbins, Hanley, et al., 1987; Robbins, Mole, Hagerman, & Hanley, 1987). Hofmann (1973, 1989) proposed that mammalian grazers evolved larger relative stomach capacity, more subdivision of chambers, and smaller openings to those chambers than browsers to increase the retention time of ingesta to improve fiber digestion capability (Clauss & Lechner‐Doll, 2001). Browsing herbivores, on the other hand, benefit from consumption of higher protein in woody plants than in grasses, but often this protein is chemically defended by plant secondary metabolites (PSMs) (Cooper & Owen‐Smith, 1985; Hofmann, 1989; Lundberg & Palo, 1993; Orians & Ward, 2010; Rohner & Ward, 1997; Shrader, Bell, Bertolli, & Ward, 2012). Tannins, a type of PSM, are known to be an antiherbivory defense in many woody plant species (Rhoades & Cates, 1976) and can have numerous deleterious effects when consumed (Shimada, 2006), although their effects depend on the plant species and on the ability of the animal to cope with tannins (Clausen, Provenza, Burritt, Reichardt, & Bryant, 1990; Mole, 1993; Mole, Butler, & Iason, 1990; Mole, Rogler, Morell, & Butler, 1990). The most notable of the effects of plant tannins on mammalian herbivores is a reduction in the amount of protein available and dry matter digestibility (Robbins, Hanley, et al., 1987; Robbins, Mole, et al., 1987). Grasses, on the other hand, are generally low in tannin content, and hydrolyzable tannins are notably absent from them and many other monocots (Chesselet, Wolfson, & Ellis, 1992; Ellis, Foo, & Porter, 1983; Ellis, 1990).
A further subdivision, independent of browsers and grazers, pertains to the fact that herbivores consume plants that have low protein and energy content in comparison with carnivores that consume animals. Much of a plant contains cell walls that are relatively indigestible (Demment & Van Soest, 1985). Hindgut fermenters (or monogastric herbivores) can break down plant material in their hindgut (cecum and colon) and can obtain energy when food is abundant and of high quality (Duncan, Foose, Gordon, Gakahu, & Lloyd, 1990; Steuer et al., 2013). When food becomes limiting and is of low quality, ruminants (or foregut fermenters; “chew the cud”) have four‐chambered stomachs (rumen, omasum, abomasum, and reticulum) to more efficiently break down cell walls (Van Soest, 1994). A third category, known as pseudoruminants (including hippopotamus Hippopotamus amphibius), have foregut fermentation, but have only three sections to the foregut (they are missing the rumen, but have the omasum, abomasum, and reticulum) and are not as efficient as ruminants (Clauss et al., 2003).
An additional trait that affects the foraging ecology of mammalian herbivores is body size (Clauss et al., 2003; Demment & Van Soest, 1985). According to the Jarman–Bell principle (Bell, 1971; Geist, 1974; Jarman, 1974), small‐bodied herbivorous mammals (e.g., dik‐dik Madoqua kirkii (Manser & Brotherton, 1995)) must be more selective and eat higher‐quality foods (high energy and protein) to maintain their fitness compared to larger‐bodied herbivores because smaller animals have higher mass‐specific metabolic rates and require more protein and energy per unit body mass than larger animals (Van Soest, 1994). Furthermore, small animals have smaller digestive systems which would constrain their abilities to digest fiber (Demment & Van Soest, 1985).
We studied 28 African ungulate species that varied in body size, digestive system, and feeding guild. A number of these African ungulate species are small‐bodied (<15 kg), including red duiker Cephalophus natalensis, gray duiker Sylvicapra grimmia, blue duiker Philantomba monticola, steenbok Raphicerus campestris, oribi Ourebia ourebi, and klipspringer Oreotragus oreotragus (Estes, 1999). We also included five key megaherbivores (weigh >1,000 kg), viz. the black rhinoceros Diceros bicornis, white rhinoceros Ceratotherium simum, African elephant Loxodonta africana (Figure 1), giraffe Giraffa camelopardalis, and the hippopotamus (Owen‐Smith, 1988). Many have assumed that large animals can consume large amounts of low‐quality food because they have a low mass‐specific energy requirement and have a large digestive system that can hold a lot of food (Demment & Van Soest, 1985). The Jarman–Bell principle predicts that megaherbivore species should be able to maintain their fitness by being nonselective and eating larger amounts of lower‐quality foods than smaller‐bodied herbivores because larger animals require less energy and protein per unit body mass than smaller animals (Geist, 1974). In the case of the hippopotamus and the white rhinoceros, this would not be difficult because they are grazers and can consume large quantities of food. However, the other species are browsers (black rhinoceros and giraffe) or mixed feeders (also termed intermediate feeders (Hofmann, 1989) African elephant) and consume large quantities of plant biomass to meet their metabolic requirements (Schmitt, Ward, & Shrader, 2016; Shrader et al., 2012). These megaherbivores have high absolute energy requirements, but they also have relatively low mass‐specific energy requirements and have large digestive systems that allow them to consume a lot more forage and effectively digest fiber.
Figure 1.

African elephant Loxodonta africana browsing on Vachellia (formerly Acacia) tortilis. This species is considered a mixed feeder (consumes both woody plants and grasses). Credit: Megan E. Griffiths
The Jarman–Bell principle takes no account of the role of plant secondary metabolites, such as tannins. However, several authors (Freeland, 1991; Freeland & Janzen, 1974; Westoby, 1978) have argued that mammalian herbivores might be greatly affected by tannins and that they must choose a wide variety of foods to minimize their intake of tannins and sustain their body masses. This may be due to tannin reducing the amount of crude protein available for digestion per bite (Schmitt, 2017; Schmitt, Shuttleworth, Ward, & Shrader, 2018). A strategy for reducing the impact of tannins that mammalian herbivores might employ is the production of salivary tannin‐binding proteins (Hofmann, Streich, Fickel, Hummel, & Clauss, 2008; McArthur, Hagerman, & Robbins, 1991; Shimada, 2006). Tannin‐binding proteins bind to tannins released from foraged material during mastication in the oral cavity (Bennick, 2002; McArthur et al., 1991; Shimada, 2006). These proteins can neutralize some, if not all, of the negative effects of tannins (Harborne, 1993). Salivary tannin‐binding proteins are restricted to mammalian herbivores and omnivores (Harborne, 1993). Mammalian carnivores do not encounter tannins in their diets and do not have salivary proteins. Studies have indicated that browsers have higher levels of salivary proteins than grazers because the former are more likely to encounter tannins (Austin, Suchar, Robbins, & Hagerman, 1989; Hagerman, Robbins, Weerasuriya, Wilson, & McArthur, 1992; Hofmann et al., 2008; Robbins, Hanley, et al., 1987). Proline is a common salivary tannin‐binding protein found in many large mammalian herbivores (Austin et al., 1989; Mole, Butler, et al., 1990; Hagerman 1992). Several browser species have proline in their saliva (mule deer Odocoileus hemionus (Austin et al., 1989); moose Alces alces (Juntheikki, 1996)), whereas grazers such as sheep Ovis aries do not have these salivary proteins (Austin et al., 1989).
Large mammalian herbivores may differ in their investments in salivary tannin‐binding proteins not merely because of ecological differences (e.g., browsers versus grazers), their type of digestive system (ruminants vs. hindgut fermenters), or scaling with body size (Hofmann et al., 2008; Pérez‐Barberìa, Elston, Gordon, & Illius, 2004). A key issue of importance is that there are phylogenetic differences among mammal digestive systems (Clauss et al., 2003, 2010; Hofmann et al., 2008; Pérez‐Barberìa et al., 2004; Pérez‐Barberìa, Pérez‐Fernandez, Robertson, & Alvarez‐Enriquez, 2008). For example, ruminants and nonruminants belong to different parts of the mammal phylogeny (Bärmann, Rössner, & Wörheide, 2013; Gatesy, Yelon, DeSalle, & Vrba, 1992; Georgiadis, Kat, Oketch, & Patton, 1990; Hassanin & Douzery, 1999; Matthee & Davis, 2001; Matthee & Robinson, 1999; Figure 2), which may confound claims of adaptation and shared ancestry. However, there are several statistical requirements that need to be addressed before analyzing data that are nested in a hierarchically structured phylogeny. The reason for this is that the absolute values of the species’ values (= “tips”) may be similar because of a shared common ancestry. Such data cannot be assumed to have been drawn independently from the same distribution (Felsenstein, 1985; Huey, Garland, & Turelli, 2019; Ward, 2000; Ward & Seely, 1996). For this reason, phylogenetically independent contrasts are calculated as the absolute differences (“contrasts”) between the “tip” (actual) values that species have and standardized by the square root of the variance, followed by estimates earlier in the phylogeny by similar means (Felsenstein, 1985; Garland, Harvey, & Ives, 1992; Pérez‐Barberìa et al. 2001, 2004, 2008). We specifically examined the phylogenetically independent contrasts of the effectiveness of salivary tannin‐binding proteins for the phylogeny of African mammals belonging to the Afrotheria (Hedges, 2001; Meredith et al., 2011; Springer et al., 1997; Tabuce, Asher, & Lehmann, 2008).
Figure 2.
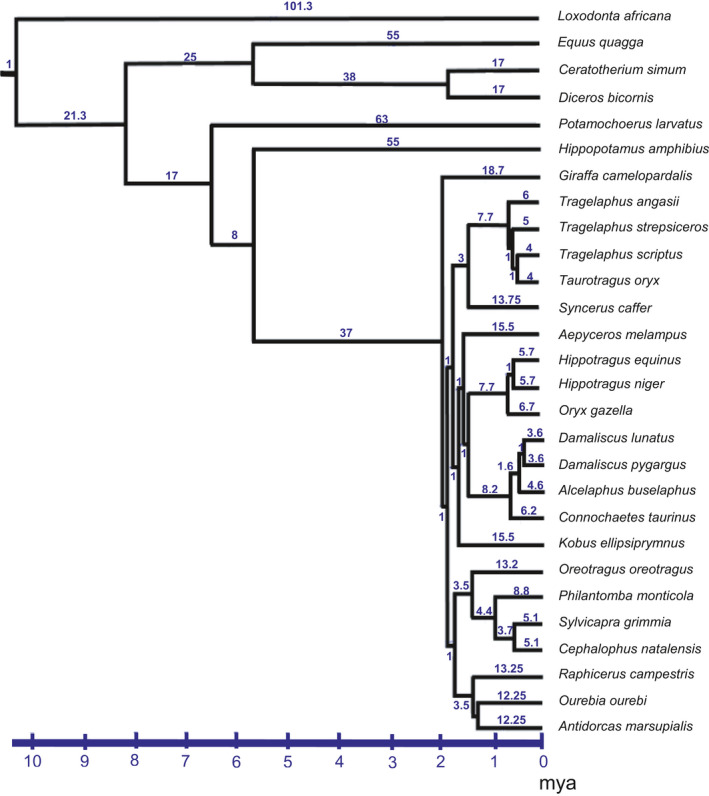
Pruned phylogeny for African ruminants and hindgut fermenters to include only those species that we studied. We used branch lengths (indicated above the branches) as indicated in the Materials and Methods, with the shortest branch lengths being set at 1
To explore the physiological response of herbivores to tannins, we tested whether any of the herbivore species have proline‐rich proteins in their saliva. Additionally, we explored the relative binding affinity of herbivore saliva to tannins in an inhibition radial diffusion assay, based on Hagerman's assay (Hagerman, 1987, 2011). We predicted that browsing herbivores may use salivary tannin‐binding proteins as an important mechanism to reduce the negative impact that tannins have on nutritional uptake, and that browsers would invest more in salivary proteins than grazers for the reasons outlined above (Figure 3).
Figure 3.
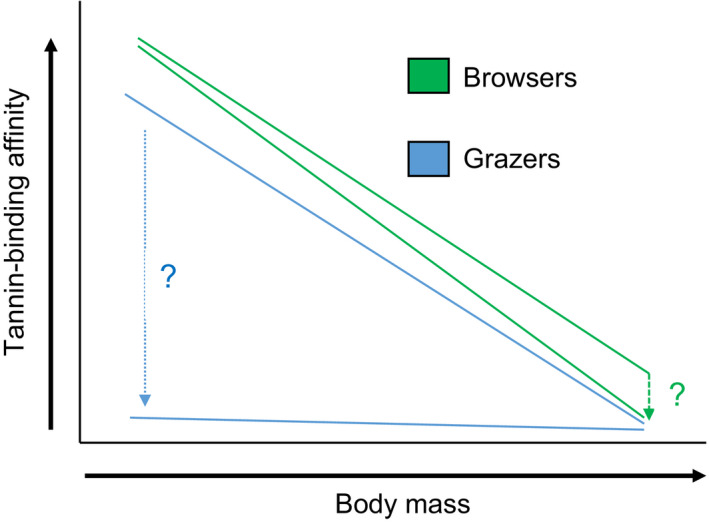
Schematic diagram of predicted differences in salivary tannin‐binding affinity between browsers and grazers relative to body size, assuming that the Jarman–Bell principle works for these salivary proteins too. If grazers do not produce salivary tannin‐binding proteins, they may have uniformly low values regardless of size. If very large herbivores (megaherbivores) are unselective, then tannin‐binding affinity of their saliva should be as low as that of grazers
We predicted that:
Proline‐rich proteins would be present in the saliva to bind with tannins (Beeley, Khoo, & Lamey, 1991). Browsers, which encounter tannins more frequently in their diets than grazers, would have higher concentrations of proline‐rich proteins than grazers which seldom encounter tannins.
Phylogenetic effects would differentiate salivary tannin‐binding proteins of ruminants and nonruminants (hindgut fermenters and pseudoruminants (hippopotamus)) (Demment & Van Soest, 1985) and browsers compared to grazers (Hofmann et al., 2008). Specifically, we predicted that ruminants would be more efficient than nonruminants and pseudoruminants and would have a smaller saliva‐ring size (i.e., bind tannins more effectively) than nonruminants, once the effects of phylogeny had been accounted for. Furthermore, we predicted that browsers, which encounter tannins more frequently than grazers, would have a smaller saliva‐ring size, due to a greater reduction in the effectiveness of tannins (Austin et al., 1989; Hagerman et al., 1992; Hofmann et al., 2008; Robbins, Hanley, et al., 1987; Schmitt et al., 2016; Windels & Hewitt, 2011; Figure 3).
Smaller herbivores would invest more in salivary tannin‐binding proteins than larger herbivores because they follow the Jarman–Bell principle (Bell, 1971; Demment & Van Soest, 1985; Jarman, 1974) and require highly digestible energy and protein (Figure 3). Consequently, there should be a significant negative correlation between the saliva‐ring size (i.e., bind tannins more effectively) and the body mass of the species, particularly if they are browsers (Figure 3). Furthermore, if grazers do not produce salivary tannin‐binding proteins, they may have uniformly low values regardless of size (Figure 3). If very large herbivores (megaherbivores) are unselective, then tannin‐binding affinity of their saliva should be as low as that of grazers (Figure 3).
2. MATERIALS AND METHODS
2.1. Research ethics
All aspects of this study were approved by the University of KwaZulu‐Natal's Animal Ethics Committee (095/13/Animal). Moreover, none of the animals were harmed or put under any additional stress during this study. Due to the noninvasive and voluntary nature of the saliva collection procedure from the human subjects (i.e., the authors and two additional graduate students—see details below), the BioMedical Research Ethics Committee of UKZN indicated that they did not need to review that experimental design or provide ethical approval. We obtained informed consent for the use of human saliva samples in our study. We confirm that all methods were performed in accordance with the relevant guidelines and regulations of the University of KwaZulu‐Natal, South Africa.
2.2. Sample collection
We acquired saliva samples from a combination of wild and semitame mammalian herbivores for this study. Saliva samples were collected during the dry season when resources are limited for herbivores in African savannas and when mixed feeders typically shift to feed on more browse (i.e., woody vegetation, which is chemically defended) than grass (i.e., less chemically defended). We would expect that for mixed feeders that shift to use more woody vegetation than grass, there might be the potential for them to have inducible salivary tannin‐binding proteins, which they would likely be using during this time (Ventura‐Cordero, Sandoval‐Castro, Torres‐Acosta, & Capetillo‐Leal, 2017). To collect samples from wild herbivores, we collected saliva from 28 species during planned game‐capture and translocation activities across South Africa. We collected the herbivore samples from wild individuals while they were immobilized by veterinarians during these routine game‐capture procedures. Although sedatives can alter salivary production in mammals (Scully, 2003), we did not observe excessive salivation in any of the individuals from which we collected samples, which has been noted as a common side effect of certain sedatives (Holz, Holz, & Barnett, 1994). The individuals from which we collected samples were captured by several veterinarians who used drug combinations that were mass‐ and species‐specific. Saliva samples were collected as soon as the individual was safe to approach after sedation and while the mouth environment was likely to be as normal as possible.
To collect the saliva, we used cotton swabs and sampled from the entire mouth, each swab touching each part of the oral cavity (tongue, cheeks, and sublingual region). Humans and domestic goats Capra hircus were sampled in a similar way. Additionally, to increase sample size, we collected saliva from 6 semitame elephants that forage naturally from Adventures with Elephants near Bela‐Bela, Limpopo Province, South Africa. These elephants were awake and willingly allowed us to take saliva samples. We immediately sealed the swabs in plastic Eppendorf vials and froze the samples until laboratory analysis. Samples were then thawed, and the saliva was separated from the cotton swabs by centrifuging them at 800 rpm (72 g) for 5 min. The herbivore species we collected, their sample sizes (i.e., number of individuals from which we took samples), feeding niches, and gut morphologies are listed in Table 1.
Table 1.
Large mammalian herbivores sampled for these analyses, indicating their feeding niches, gut morphologies, and their effects on tannin binding (mean ring diameter)
| Species | N | Feeding Niche | Digestive system | Mean ring diameter ± SD (mm) |
|---|---|---|---|---|
| Black Rhinoceros Diceros bicornis | 10 | Browser | Nonruminant | 0.4 ± 0.66 |
| Blesbok Damaliscus pygargus | 10 | Grazer | Ruminant | 1.8 ± 1.03 |
| Blue Duiker Philantomba monticola | 2 | Browser | Ruminant | 0.4 ± 0.53 |
| Buffalo Syncerus caffer | 10 | Grazer | Ruminant | 1.3 ± 0.95 |
| Bush Pig Potamochoerus larvatus | 2 | Mixed feeder | Nonruminant | 0.0 ± 0.00 |
| Bushbuck Tragelaphus sylvaticus | 9 | Browser | Ruminant | 0.8 ± 1.03 |
| Eland Taurotragus oryx | 13 | Mixed feeder | Ruminant | 1.5 ± 1.19 |
| Elephant Loxodonta africana | 10 | Mixed feeder | Nonruminant | 1.4 ± 0.46 |
| Gemsbok Oryx gazella | 4 | Browser | Ruminant | 3.0 ± 0.41 |
| Giraffe Giraffa camelopardalis | 10 | Browser | Ruminant | 0.6 ± 1.26 |
| Gray Duiker Sylvicapra grimmia | 2 | Browser | Ruminant | 0.0 ± 0.00 |
| Hippopotamus Hippopotamus amphibius | 4 | Grazer | Foregut fermenter | 2.4 ± 0.58 |
| Impala Aepyceros melampus | 10 | Mixed feeder | Ruminant | 1.3 ± 0.82 |
| Klipspringer Oreotragus oreotragus | 4 | Browser | Ruminant | 1.1 ± 1.31 |
| Kudu Tragelaphus strepsiceros | 10 | Browser | Ruminant | 0.1 ± 0.32 |
| Nyala Tragelaphus angasii | 10 | Browser | Ruminant | 0.7 ± 0.82 |
| Oribi Ourebia ourebi | 2 | Grazer | Ruminant | 1.8 ± 0.35 |
| Red Duiker Cephalophus natalensis | 10 | Browser | Ruminant | 0.9 ± 0.91 |
| Hartebeest Alcelaphus buselaphus | 3 | Grazer | Ruminant | 0.0 ± 0.00 |
| Roan Hippotragus equinus | 5 | Grazer | Ruminant | 0.1 ± 0.22 |
| Sable Hippotragus niger | 10 | Grazer | Ruminant | 1.2 ± 0.57 |
| Springbok Antidorcas marsupialis | 10 | Grazer | Ruminant | 1.2 ± 0.83 |
| Steenbok Raphicerus campestris | 4 | Browser | Ruminant | 2.3 ± 0.65 |
| Tsessebe Damaliscus lunatus | 10 | Grazer | Ruminant | 0.6 ± 0.76 |
| Waterbuck Kobus ellipsiprymnus | 10 | Grazer | Ruminant | 0.6 ± 0.78 |
| White Rhinoceros Ceratotherium simum | 4 | Grazer | Nonruminant | 0.8 ± 0.65 |
| Blue Wildebeest Connochaetes taurinus | 9 | Grazer | Ruminant | 1.3 ± 0.80 |
| Burchell's Zebra Equus quagga | 10 | Grazer | Nonruminant | 1.4 ± 0.57 |
A smaller ring means a greater effect on tannins. Nonruminant = hindgut fermenter. Mixed feeders consume both woody plants and grasses, albeit not necessarily in the same place (Codron et al., 2012; Schmitt et al., 2016; Shrader et al., 2012). N = sample size (number of individuals). We also sampled five human individuals (positive control) and five domestic goats (negative control) (see Section 2).
2.3. Proline‐rich proteins
To test for the presence of proline‐rich proteins in the herbivore saliva, we used two different approaches. First, we used a sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS‐PAGE) (Laemmli, 1970). We used a 12.5% running gel and 4% stacking gel to separate the various proteins present in the herbivore saliva. We used a Bradford’s (1976) analysis to assess the amount of protein in each sample. Thereafter, we diluted each saliva sample by 50% with phosphate‐buffered saline (PBS) to reduce the potential negative effects of salts in our gels. We ran reducing gels, so we added β‐mercaptoethanol, a reducing agent that separates proteins into their most basic forms or subunits, in a 1:10 ratio, to the sample buffer prior to addition of the saliva samples (Beeley et al., 1991). We loaded each gel with 25 μg/μl of protein and ran them at 18 mA per gel until the dye front reached 0.5 cm from the edge of the gel. For each gel, we tested the saliva of each of the 28 herbivore species against a negative control (i.e., lacks proline‐rich proteins; domestic goat saliva) and a positive control (i.e., has proline‐rich proteins; human saliva).
The second approach we used was to run a comparative SDS‐PAGE gel first using the staining and destaining method (Laemmli, 1970), as well as the Beeley et al. (1991) method for staining and destaining to probe for proline‐rich proteins. For Laemmli’s (1970) method, we mixed 45% (v/v) methanol, 10% (v/v) acetic acid, and 0.25% (w/v) Coomassie Brilliant Blue R‐250. We left gels to stain overnight. The gels were then destained with 50% (v/v) methanol and 10% (v/v) acetic acid. Gels in this treatment were compared to gels that were stained in 0.1% w/v CBB R‐250 mixed with 40% v/v ethanol and 10% v/v acetic acid for 3 hr and then destained for 4 d in 10% v/v acetic acid (Beeley et al., 1991). Should any of the herbivore samples contain proline‐rich proteins according to the Beeley et al. (1991) technique, they should stain pink/violet. We were unable to take photographs of the gels because the University of KwaZulu‐Natal did not have adequate color photography. Black‐and‐white images do not show the pink bands effectively, and conventional SLR cameras/flash do not photograph such bands successfully.
2.4. Relative tannin‐binding capacities
To test the tannin‐binding capabilities of herbivore saliva, we altered Hagerman’s (1987) radial diffusion assay to test for relative tannin–protein precipitation (Hagerman, 2011). We used this assay to understand relative tannin precipitation capabilities by herbivore saliva, not to test for specific salivary tannin‐binding proteins or absolute tannin‐binding levels. This inhibition assay tested for a reduction in the size of the ring relative to pure tannic acid. To make the radial diffusion plates, we followed Hagerman’s (2011) procedure that precipitates tannin using bovine serum albumin (BSA). We made a series of 6 μL deep wells in an Ouchterlony double‐immunodiffusion pattern (Ouchterlony, 1953) in place of the traditional 4 wells per plate pattern (Hagerman, 1987). Instead of performing the classic radial diffusion assay, we created an inhibition assay whereby a tannic acid solution was mixed with saliva prior to being pipetted into the wells. For this experiment, we used a concentration of 1 g tannic acid per 100 ml, which was suspended in 70% acetone. The saliva may bind with tannins in the extract prior to contact with the bovine serum albumin mixed into the agar, thus making it unavailable for binding to the BSA. The resulting saliva tannin precipitate rings in the agar could then be compared to rings formed by the tannic acid stock solution alone as an indication of whether there was a difference between the amounts of tannins bound in each treatment (Bryant et al., 1991; Robbins, Hagerman, Austin, McArthur, & Hanley, 1991). For this experiment, we used a concentration of 1 g tannic acid per 100 ml, which was suspended in 70% acetone (Alonso‐Díaz et al., 2008; Ventura‐Cordero et al., 2017). Hagerman (2011) indicates that acetone‐containing extracts do not inhibit the precipitation reaction like other protein‐precipitating methods, so acetone can be used with this method. Because the aim of this assay was to make comparisons across the herbivore species and their relative abilities to precipitate tannins (and not identify absolute tannin‐binding capabilities), we used a standardized concentration of a common type of tannins for this assay. Ultimately, this provided a comparative measure of the tannin‐binding ability of the saliva.
To test for the tannin‐binding capacities of herbivore saliva, we tested 4 μl of the tannic acid solution alone as well as 4 μl of the tannic acid solution combined with 2 μl of herbivore saliva. Because we only aimed to make comparisons of the tannin‐precipitating abilities of the saliva of different herbivore species and were not making absolute estimates of their tannin‐binding abilities, we used a 2:1 ratio of tannic acid solution to saliva because it allowed us to identify any reaction at the scale of the wells. Prior to the addition of the tannic acid solution and saliva mixture to the wells, we mixed the two components in Eppendorf vials, vortexed them for 5 s, and allowed the mixture to react at room temperature (~25°C) in artificial (laboratory) light for 30 min. After reacting for 30 min, we pipetted the solution into the wells. We sealed each petri dish with parafilm and placed them into an incubator at 30°C for four days (Hagerman, 2011). On day four, we measured the perpendicular diameters of the tannin‐binding ring that had formed around each well and took an average (Hagerman, 2011).
2.5. Statistical analyses
We used mean ring diameter (mean of the perpendicular measurements of each tannin‐binding ring) for the tannic acid solution as our dependent factor and treatment (digestive system and feeding niche) as the independent factor. We also tested for an interaction effect between digestive system and feeding niche. We considered the pseudoruminant (hippopotamus—Table 1) as a nonruminant because it is less efficient than a ruminant. To account for normality and homogeneity of variance, we transformed the data, using the reciprocal of the mean saliva‐ring distance, and calculated the log10 of body mass. We ran an ANCOVA with reciprocal of the mean saliva‐ring distance as our dependent variable, digestive system and feeding niche as independent variables, and log10 body mass as a covariate. We also ran a regression between the reciprocal of the mean saliva‐ring distance and the log10 of body mass.
We primarily used a phylogenetically independent contrast (PIC) approach, using the Brownian motion model of evolution that underlies PIC (Garland et al., 1992). We first compiled a phylogeny based on the Tree of Life Web Project (Maddison, Schulz, & Maddison, 2007) and other papers (Bärmann et al., 2013; Buntjer, Otsen, Nijman, Kuiper, & Lenstra, 2002; Gatesy, Amato, Vrba, Schaller, & DeSalle, 1997; Gatesy et al., 1992; Georgiadis et al., 1990; Hedges, 2001; Matthee & Davis, 2001; Matthee & Robinson, 1999; Meredith et al., 2011; Springer et al., 1997; Tabuce et al., 2008) with a focus on the fact that African ungulates (and other African taxa) are now recognized as belonging to the Afrotheria (Figure 2). We excluded species that were not part of the phylogeny. We used fossil ages as calibrations for the phylogeny (Gatesy et al., 1992; Hedges, 2001) and calculated branch lengths (Meredith et al., 2011). We ran the analysis in the modular comparative method program Mesquite (Maddison & Maddison, 2018) using PDAP (Garland et al., 1992). Our categorical independent contrast variables were digestive system and feeding niche and the covariate log10 body mass.
We also calculated phylogenetic signal (Münkemüller et al., 2012). Phylogenetic signal of continuous traits, such as saliva‐ring diameter contrasts, is an important measure of the statistical dependence among species’ trait values due to their phylogenetic relationships (Revell, Harmon, & Collar, 2008). Münkemüller et al. (2012) reviewed several methods of assessing phylogenetic signal and found that Blomberg, Garland, and Ives (2003) to be one of the most useful under a wide range of phylogenetic models. Blomberg et al. (2003) suggested that a reliable way to assess phylogenetic signal is to examine the ratio of the MS Factor (using phylogenetic contrasts) relative to the MS Error. In this case, we analyzed the saliva‐ring‐independent contrasts versus feeding niche‐independent contrasts (browser vs. grazer vs. mixed feeder). A large value for the MS Factor relative to MS Error indicates a significant phylogenetic signal (Münkemüller et al., 2012).
3. RESULTS
3.1. Proline‐rich proteins
Using the assay of Beeley et al. (1991), we found that our positive control (human saliva) yielded several pink/violet bands indicating the presence of proline‐rich proteins (Beeley et al., 1991), whereas our negative control (domestic goat saliva—Lamy et al., 2008) had numerous protein bands but lacked proline‐rich proteins indicated by the absence of a pink/violet band(s). However, all of the 28 herbivore species had proteins in their saliva, but none of the herbivore samples contained proline‐rich proteins (i.e., no pink/violet bands).
3.2. Phylogenetic differences
In these inhibition radial diffusion assays, an effective salivary tannin‐binding protein would reduce the size of the ring (diameter) relative to tannic acid. In our phylogenetic analyses of 28 African ungulate species, there was no significant difference using independent contrasts in digestive system (ruminant vs. nonruminant) for the reciprocal of saliva‐ring size of these species (F = 0.088; p = .769). Similarly, we found no significant difference using independent contrasts between feeding niches (browsers vs. grazers) in terms of the reciprocal of saliva‐ring size in these ungulate species (F = 1.620; p = .214). We found that there was a negative correlation (r = −0.33) between the reciprocal of saliva‐ring diameter and (continuous) log10 body mass‐independent contrasts. However, this relationship was not significant (F = 3.217; p = .085). Consequently, we could not look for outliers that may reflect species that differed significantly from the 95% confidence intervals (Garland & Ives, 2000). There was also no significant relationship between mean saliva‐ring diameter and log10 body mass (r 2 = 0.004, F = 0.072, p = .791) for ruminants only.
There was no significant phylogenetic signal for saliva‐independent contrasts versus feeding niche‐independent contrasts (F = 0.007, p = .933) according to the technique of Blomberg et al. (2003). Similarly, we found no significant phylogenetic signal for digestive system (F = 0.373; p = .547). Thus, because we found no phylogenetic signal, we did not perform further phylogenetically appropriate comparative analyses such as phylogenetic generalized least squares (PGLS—Symonds & Blomberg, 2014). We note that Blomberg, Lefevre, Wells, and Waterhouse (2012) recognize that phylogenetically independent contrasts (PICs) and phylogenetic generalized least squares (PGLS) approaches are functionally similar.
3.3. Ignoring phylogeny
When we ignored the effect of phylogeny on the basis of a nonsignificant phylogenetic signal, we ran an ANCOVA and found that there was no significant difference between digestive system (ruminants vs. hindgut fermenters) (F = 1.139, p = .297) (Figure 4). There was also no significant effect of feeding niche (grazer vs. browser) (F = 0.007, p = .933), nor was there a significant interaction between digestive system and feeding niches (F = 1.910, p = .180). There was also no significant effect of the log10 body mass on mean saliva‐ring diameter (r 2 = 0.003, F = 0.082, p = .777) (Figure 5). There was also no significant relationship between mean saliva‐ring diameter and log10 body mass (r 2 = 0.004, F = 0.072, p = .791) for ruminants only.
Figure 4.
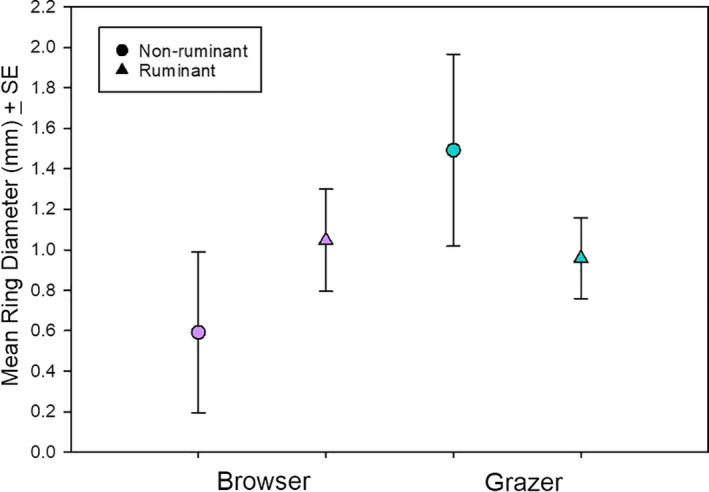
Mean tannin ring diameter ± SE for browsers and grazers, differentiated by digestive system. Nonruminants are hindgut fermenters, with the exception of the hippopotamus, which is considered a pseudoruminant
Figure 5.
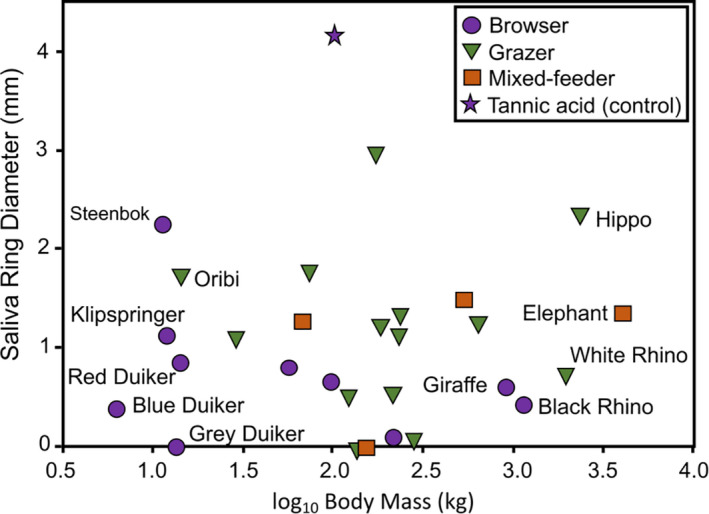
Mean saliva‐ring diameter of 28 African herbivorous mammals. All species bigger than 3 (>1,000 kg) are considered megaherbivores (listed). Small herbivores, most of which are browsers, are also listed. Purple star = tannic acid, the hydrolyzable tannin that acted as a control. Note that all species had saliva that contained proteins that reduced the diameter of the rings, indicating that they were capable of binding tannins. Some species could bind all tannins (mean value = 0 mm)
All species could bind tannins, albeit to differing degrees (Table 1). Of those species that bound tannin completely (saliva‐ring diameter = 0 mm), one was a browser (gray duiker), one was a mixed feeder (bush pig Potamochoerus larvatus), and one was a grazer (red hartebeest Alcelaphus busephalus). Two species had a mean score close to 0 (=0.1), one of which was the greater kudu Tragelaphus strepsiceros (browser) and the other the roan Hippotragus equinus (grazer). The species least able to bind tannins was the gemsbok Oryx gazella (grazer; mean = 3.0 mm) compared to the tannic acid stock (control) solution mean of 4.18 mm (Figure 5). Of the small herbivores, five of the six species were browsers. The only exception was the oribi (grazer). Among the megaherbivores, the black rhinoceros and the giraffe Giraffa camelopardalis (both browsers) were most effective at binding tannins, but the saliva of the white rhinoceros (a grazer) was not that different at binding tannins from the two aforementioned megaherbivore browsers (Figure 5). Among the megaherbivores, the hippopotamus H. amphibius (strictly a grazer) was the least effective at binding tannins.
4. DISCUSSION
We found no evidence for proline‐rich proteins, although the inhibition radial diffusion assay that we used allowed us to establish that the saliva of all 28 herbivore species we tested are effective and can bind tannins (they reduced the size of the radial diffusion ring—Table 1). However, we were unable to establish the type/s of salivary proteins that they have. For the herbivores that we tested using the Beeley et al. (1991) technique, we can only say that those species do not have proline‐rich salivary proteins (Schmitt, 2017; Schmitt, Shuttleworth, Shrader, & Ward, 2020), but we do not know whether they are histatins or some as‐yet‐unidentified salivary protein (Lamy et al., 2008; Shimada, 2006).
Among mammals, salivary proline‐rich proteins that bind tannins occur in a number of orders, including the Artiodactyla, Rodentia, and Lagomorpha, as well as some herbivorous Marsupialia, but are absent among the Carnivora (Clausen et al., 1990; McArthur et al., 1991; Robbins et al., 1991). Even humans and other primates have tested positive for proline in their saliva (Bacon & Rhodes, 1998; Bennick & Connell, 1971; Mehansho, Butler, & Carlson, 1987), perhaps because of their historical dependence on tannin‐rich substances (e.g., berries, nuts, and many legumes (Foley & McArthur, 1994; Prinz & Lucas, 2000)). Similarly, domestic rats Rattus norvegicus, mice Mus musculus, and hamsters Mesocricetus auratus may also induce proline (Ann, Clements, Johnstone, & Carlson, 1987; Mehansho et al., 1983, 1987; Skopec, Hagerman, & Karasov, 2004). However, the domestic goat was not found to have proline compounds despite being a mixed feeder (eats both grass and woody plants; Austin et al., 1989; Distel & Provenza, 1991; Makkar, 2003; Schmitt, Ward, & Shrader, 2020; Ventura‐Cordero et al., 2017). Nonetheless, domestic goats are capable of binding tannins in their saliva, suggesting that another protein, as yet undescribed, is employed (Alonso‐Díaz, Torres‐Acosta, Sandoval‐Castro, & Hoste, 2010; Distel & Provenza, 1991; Lamy et al., 2008; Makkar, 2003; Schmitt, Ward, et al., 2020; Vaithiyanathan, Mishra, Sheikh, & Kumar, 2001; Ventura‐Cordero et al., 2017).
Among the Artiodactyla, several authors have found that browsing members of the Cervidae, such as mule deer Odocoileus hemionus, have proline in their saliva (Austin et al., 1989; Robbins et al., 1991). Among the Bovidae, the domestic cow Bos taurus is a grazing bovine and does have tannin‐binding proline proteins in its saliva (although this species is relatively ineffective in blocking tannins; Mole, Butler, et al., 1990). Yet, the closely related fellow bovines that we studied, including the browsers (greater kudu, nyala Tragelaphus angasii, bushbuck T. scriptus, and eland Taurotragus oryx) and grazing buffalos Syncerus caffer (Table 1), do not show evidence of proline. This might reflect a lack of sensitivity in the assay we used, although we think that this is unlikely to be the case. The pink/violet bands in the human saliva samples, highlighting the presence of proline‐rich proteins, were very clearly displayed on every gel, leading us to believe that the Beeley et al. (1991) technique was adequate for our study. Further research will be needed to establish whether these salivary tannin‐binding proteins could have evolved independently on 28 occasions, or whether a smaller number of evolutionary events led to their evolution.
We also predicted that there would be a significant effect of phylogeny on the ability of ruminants and nonruminants (hindgut fermenters) to block tannins. We found no support for this. Our phylogenetic analyses are consistent with earlier conclusions (Pérez‐Barbería, Gordon, & Illius, 2001; Robbins, Spalinger, & Van Hoven, 1995) that there was no support for the anatomical split between browsers and grazers on the basis of their gut morphologies. A continuum rather than a strict browser/grazer dichotomy may exist (Springer et al., 1997), which includes intermediate or mixed feeders between the two categories. The lack of divergence according to feeding niche (browser/grazer) may also apply to salivary proteins. An example that may be particularly pertinent is a study of salivary tannin‐binding proteins in three species of rhinoceros (Clauss et al., 2005). These authors found that the black rhinoceros (browser) had more effective salivary proteins for binding tannins than did the white rhinoceros (grazer) as one might expect. However, the Indian rhinoceros Rhinoceros unicornis, which is also a grazer, had more effective salivary proteins for binding tannins than either the black or white rhinoceros.
We predicted that browsers, which encounter tannins more frequently than grazers, would have a smaller saliva‐ring size, indicating a greater binding of tannins, resulting in greater reduction in their effectiveness (Austin et al., 1989; Hagerman et al., 1992; Robbins, Hanley, et al., 1987; Robbins, Mole, et al., 1987; Schmitt et al., 2016; Windels & Hewitt, 2011; Figure 3). However, we did not find a significant difference between these two groups. We compared nonruminants (hindgut fermenters) and ruminants and found no difference in the tannin‐binding ability that has been suggested to be driven by these anatomical differences between browsers and grazers (Hofmann, 1973, 1989; Hofmann & Stewart, 1972). We also found no correlation between the relative effectiveness of tannin‐binding proteins in the saliva and body mass. This result is inconsistent with data, indicating that there was scaling of salivary gland size and body mass0.75 and thus an assumed greater production of salivary tannin‐binding proteins by larger herbivores (Hofmann et al., 2008). Our results are also inconsistent with those of Hofmann (1973) and Kay, Engelhardt, and White (1980) who found that grazing ruminants had parotid salivary glands that were three times smaller (and thus likely produced less salivary tannin‐binding proteins) than those of browsing ruminants that constantly feed on tannin‐rich food. Yet, there is conflicting evidence, in any case, on the effectiveness of proline contained in binding tannins (Mole, Butler, et al., 1990), which means that scaling of salivary gland size to salivary tannin‐binding protein may not be that meaningful. For example, the domestic cow (a grazer) has high amounts of proline in its salivary glands, yet does not bind tannins any more effectively than another common grazer, the domestic sheep, that has low amounts of proline in its salivary glands (Mole, Butler, et al., 1990). These authors suggested that proline‐rich salivary proteins may not have evolved solely for the purpose of binding tannins but may also be present in the saliva to stabilize minerals in the teeth or to limit tooth decay (Bennick, 1982). However, that does not explain why many herbivores possess proline and carnivores do not (Austin et al., 1989; Harborne, 1993; Hofmann et al., 2008; Robbins et al., 1991).
Of the small‐bodied species we studied, the majority are browsers (red duiker, gray duiker, blue duiker, steenbok, and klipspringer) and often encounter foods that are very tannin‐rich. The only exception among the small antelopes was the oribi, a grazer (Arcese, Jongejan, & Sinclair, 1995; Everett, Perrin, & Rowe‐Rowe, 1992; Stears & Shrader, 2020). These small herbivores are closely related (Figure 2), suggesting some phylogenetic effect. However, the relative abilities of these small herbivores to bind tannins were not necessarily greater than those of several (larger) grazers (Figure 5), substantiating the absence of evidence for phylogenetic effects. Furthermore, the paradox that the oribi is the only small antelope of six species that is a grazer (i.e., eating low‐quality food) is worth re‐examining. We speculate that the oribi is at the body size threshold for being a grazing ruminant (Clauss et al., 2003; Demment & Van Soest, 1985; Gordon & Illius, 1994). Future studies should determine whether the effectiveness of the salivary proteins for binding tannins is integral to the fact that five of the six small herbivores are browsers, or whether behavioral avoidance by virtue of their small mouthparts (Iason & Villalba, 2006; Nobler et al., 2019; Provenza & Balph, 1987) can explain their abilities to access high‐value food items.
Although small browsers might be expected to have evolved salivary tannin‐binding proteins because of their high metabolic rates, we have shown elsewhere (Schmitt et al., 2016) that even animals as large as elephants have saliva that can bind tannins to maintain their large body masses (see also Dierenfeld, Du Toit, & Braselton, 1995; Furstenburg & Van Hoven, 1994; Muller, 2013; Owen‐Smith & Chafota, 2012; Schmitt et al., 2018; Shaw, 2011; Ward, Muller, & Shrader, 2017). All of the five African megaherbivores that we studied have protein(s) in their saliva that bind to tannins, regardless of whether they are browsers (giraffe and black rhinoceros), mixed feeders (elephants), or grazers (hippopotamus and white rhinoceros; Figure 5). Only two species of megaherbivores are closely related (black and white rhinoceros; Figure 2). Thus, there is no phylogenetic effect. This suggests that the relationships between body size and the quality of food (as determined by plant secondary metabolites) required by large herbivores should be re‐examined.
CONFLICT OF INTEREST
The authors declare no competing interests.
AUTHOR CONTRIBUTION
David Ward: Conceptualization (equal); Data curation (lead); Formal analysis (lead); Funding acquisition (lead); Investigation (equal); Methodology (equal); Project administration (equal); Writing‐original draft (lead); Writing‐review & editing (lead). Melissa Schmitt: Conceptualization (equal); Data curation (equal); Methodology (equal); Resources (equal); Validation (equal); Visualization (equal); Writing‐original draft (supporting); Writing‐review & editing (supporting). Adrian Shrader: Conceptualization (equal); Investigation (equal); Project administration (equal); Resources (equal); Supervision (equal); Validation (equal); Visualization (equal); Writing‐original draft (supporting); Writing‐review & editing (supporting).
ACKNOWLEDGMENTS
We thank the anonymous reviewers for their useful comments on this manuscript. We thank Conrad Matthee and Tony Verboom for their advice on phylogenies. This research was funded in part by the National Research Foundation of South Africa and the Herrick Trust of Kent State University. We thank S. Hensman, M. Hensman, J. Crosby, and the staff at Adventures with Elephants and the Rory Hensman Conservation and Research Unit for allowing us to collect samples from the elephants on their premises. Furthermore, we would like to thank Dr. S. Pfitzer (WildVET), I. Colenbrander (Chui Wildlife Services), Dr. G. Bauer (Gondwana Wildlife Services), Dr. R. van Deventer (Wildlife Solutions), Dr. D. Grobler, and Dr. K. Stears for their assistance with sample collection. We would also like to thank Dr. R. Krause for laboratory support and guidance. This work was supported by the National Research Foundation of South Africa (grant numbers: 90448, 97262, 77582), the Gay Langmuir Bursary from the University of KwaZulu‐Natal (School of Life Sciences) awarded to MH Schmitt, and the Herrick Trust (to DW).
Ward D, Schmitt MH, Shrader AM. Are there phylogenetic differences in salivary tannin‐binding proteins between browsers and grazers, and ruminants and hindgut fermenters?. Ecol Evol. 2020;10:10426–10439. 10.1002/ece3.6698
Contributor Information
David Ward, Email: dward21@kent.edu.
Melissa H. Schmitt, Email: melissa.h.schmitt@gmail.com.
Adrian M. Shrader, Email: adrian.shrader@up.ac.za.
DATA AVAILABILITY STATEMENT
All information is present here (Table 1; Figure 2).
REFERENCES
- Alonso‐Díaz, M. A. , Torres‐Acosta, J. F. J. , Sandoval‐Castro, C. A. , Capetillo‐Leal, C. , Brunet, S. , & Hoste, H. (2008). Effects of four tropical tanniniferous plant extracts on the inhibition of larval migration and the exsheathment process of Trichostrongylus colubriformis infective stage. Veterinary Parasitology, 153, 187–192. 10.1016/j.vetpar.2008.01.011 [DOI] [PubMed] [Google Scholar]
- Alonso‐Díaz, M. , Torres‐Acosta, J. , Sandoval‐Castro, C. , & Hoste, H. (2010). Tannins in tropical tree fodders fed to small ruminants: A friendly foe? Small Ruminant Research, 89, 164–173. 10.1016/j.smallrumres.2009.12.040 [DOI] [Google Scholar]
- Ann, D. K. , Clements, S. , Johnstone, E. M. , & Carlson, D. M. (1987). Induction of tissue‐specific proline‐rich protein multigene families in rat and mouse parotid glands by isoproterenol. Journal of Biological Chemistry, 262, 889–904. [PubMed] [Google Scholar]
- Arcese, P. , Jongejan, G. , & Sinclair, A. R. E. (1995). Behavioural flexibility in a small African antelope: Group size and composition in the Oribi (Ourebia ourebi, Bovidae). Ethology, 99, 1–23. 10.1111/j.1439-0310.1995.tb01085.x [DOI] [Google Scholar]
- Austin, P. J. , Suchar, L. A. , Robbins, C. T. , & Hagerman, A. E. (1989). Tannin‐binding proteins in saliva of deer and their absence in saliva of sheep and cattle. Journal of Chemical Ecology, 15, 1335–1347. 10.1007/BF01014834 [DOI] [PubMed] [Google Scholar]
- Bacon, J. R. , & Rhodes, M. J. C. (1998). Development of a competition assay for the evaluation of the binding of human parotid salivary proteins to dietary complex phenols and tannins using a peroxidase‐labeled tannin. Journal of Agricultural & Food Chemistry, 46, 5083–5088. 10.1021/jf980791c [DOI] [Google Scholar]
- Bärmann, E. V. , Rössner, G. E. , & Wörheide, G. (2013). A revised phylogeny of Antilopini (Bovidae, Artiodactyla) using combined mitochondrial and nuclear genes. Molecular Phylogenetics & Evolution, 67, 484–493. 10.1016/j.ympev.2013.02.015 [DOI] [PubMed] [Google Scholar]
- Beeley, J. A. , Khoo, K. S. , & Lamey, P. J. (1991). Two‐dimensional electrophoresis of human parotid salivary proteins from normal and connective tissue disorder subjects using immobilised pH gradients. Electrophoresis, 12, 493–499. 10.1002/elps.1150120707 [DOI] [PubMed] [Google Scholar]
- Bell, R. H. V. (1971). A grazing ecosystem in the Serengeti. Scientific American, 25, 86–93. 10.1038/scientificamerican0771-86 [DOI] [Google Scholar]
- Bennick, A. (1982). Salivary proline‐rich proteins. Molecular & Cellular Biochemistry, 45, 83–99. 10.1007/BF00223503 [DOI] [PubMed] [Google Scholar]
- Bennick, A. (2002). Interaction of plant polyphenols with salivary proteins. Critical Reviews in Oral Biology and Medicine, 13, 184–196. [DOI] [PubMed] [Google Scholar]
- Bennick, A. , & Connell, G. E. (1971). Purification and partial characterization of four proteins from human parotid saliva. Biochemical Journal, 123, 455–464. 10.1042/bj1230455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blomberg, S. P. , Garland, T. , & Ives, A. R. (2003). Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution, 57, 717–745. 10.1111/j.0014-3820.2003.tb00285.x [DOI] [PubMed] [Google Scholar]
- Blomberg, S. P. , Lefevre, J. G. , Wells, J. A. , & Waterhouse, M. (2012). Independent contrasts and PGLS regression estimators are equivalent. Systematic Biology, 61, 382–391. 10.1093/sysbio/syr118 [DOI] [PubMed] [Google Scholar]
- Bradford, M. M. (1976). Rapid and sensitive method for quantitation of microgram quantities of protein utilization principle of protein‐dye binding. Analytical Biochemistry, 72, 248–254. [DOI] [PubMed] [Google Scholar]
- Bryant, J. P. , Provenza, F. D. , Pastor, J. , Reichardt, P. B. , Clausen, T. P. , & Du Toit, J. T. (1991). Interactions between woody plants and browsing mammals mediated by secondary metabolites. Annual Review of Ecology and Systematics, 22, 431–446. 10.1146/annurev.es.22.110191.002243 [DOI] [Google Scholar]
- Buntjer, J. B. , Otsen, M. , Nijman, I. J. , Kuiper, M. T. R. , & Lenstra, J. A. (2002). Phylogeny of bovine species based on AFLP fingerprinting. Heredity, 88, 46–51. 10.1038/sj.hdy.6800007 [DOI] [PubMed] [Google Scholar]
- Chesselet, P. , Wolfson, M. M. , & Ellis, R. P. (1992). A comparative histochemical study of plant polyphenols in southern African grasses. Journal of the Grassland Society of Southern Africa, 9, 119–126. 10.1080/02566702.1992.9648311 [DOI] [Google Scholar]
- Clausen, T. P. , Provenza, F. D. , Burritt, E. A. , Reichardt, P. B. , & Bryant, J. P. (1990). Ecological implications of condensed tannin structure: A case study. Journal of Chemical Ecology, 16, 2381–2392. 10.1007/BF01017463 [DOI] [PubMed] [Google Scholar]
- Clauss, M. , Frey, R. , Kiefer, B. , Lechner‐Doll, M. , Loehlein, W. , Polster, C. , … Streich, W. J. (2003). The maximum attainable body size of herbivorous mammals: Morphophysiological constraints on foregut, and adaptations of hindgut fermenters. Oecologia, 136, 14–27. 10.1007/s00442-003-1254-z [DOI] [PubMed] [Google Scholar]
- Clauss, M. , Gehrke, J. , Hatt, J.‐M. , Dierenfeld, E. S. , Flach, E. J. , Hermes, R. , … Fickel, J. (2005). Tannin‐binding salivary proteins in three captive rhinoceros species. Comparative Biochemistry & Physiology, Part A, 140, 67–72. 10.1016/j.cbpb.2004.11.005 [DOI] [PubMed] [Google Scholar]
- Clauss, M. , Hume, I. D. , & Hummel, J. (2010). Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal, 4, 979–992. 10.1017/S1751731110000388 [DOI] [PubMed] [Google Scholar]
- Clauss, M. , & Lechner‐Doll, M. (2001). Differences in selective reticulo‐ruminal particle retention as a key factor in ruminant diversification. Oecologia, 129, 321–327. 10.1007/s004420100735 [DOI] [PubMed] [Google Scholar]
- Codron, J. , Codron, D. , Sponheimer, M. , Kirkman, K. , Duffy, K. J. , Raubenheimer, E. J. , … Lee‐Thorp, J. A. (2012). Stable isotope series from elephant ivory reveal lifetime histories of a true dietary generalist. Proceedings of the Royal Society B: Biological Sciences, 279(1737), 2433–2441. 10.1098/rspb.2011.2472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper, S. M. , & Owen‐Smith, N. (1985). Condensed tannins deter feeding by browsing ruminants in a South African savanna. Oecologia, 67, 142–146. 10.1007/BF00378466 [DOI] [PubMed] [Google Scholar]
- Demment, M. W. , & Van Soest, P. J. (1985). A nutritional explanation for body‐size patterns of ruminant and non‐ruminant herbivores. American Naturalist, 125, 641–672. 10.1086/284369 [DOI] [Google Scholar]
- Dierenfeld, E. , Du Toit, R. , & Braselton, W. E. (1995). Nutrient composition of selected browses consumed by black rhinoceros (Diceros bicornis) in the Zambezi Valley, Zimbabwe. Journal of Zoo & Wildlife Medicine, 26, 220–230. [Google Scholar]
- Distel, R. A. , & Provenza, F. D. (1991). Experience early in life affects voluntary intake of blackbrush by goats. Journal of Chemical Ecology, 17, 431–450. 10.1007/BF00994343 [DOI] [PubMed] [Google Scholar]
- Duncan, P. , Foose, T. J. , Gordon, I. J. , Gakahu, C. G. , & Lloyd, M. (1990). Comparative nutrient extraction from forages by bovids and equids: A test of the nutritional model of bovid/equid competition and coexistence. Oecologia, 84, 411–418. [DOI] [PubMed] [Google Scholar]
- Ellis, C. J. , Foo, L. Y. , & Porter, L. J. (1983). Enantiomerism: A characteristic of the proanthocyanidin chemistry of the Monocotyledonae. Phytochemistry, 22, 483–487. 10.1016/0031-9422(83)83030-1 [DOI] [Google Scholar]
- Ellis, R. P. (1990). Tannin‐like substances in grass leaves. Memoirs of the Botanical Survey of South Africa, 59, 1–80. [Google Scholar]
- Estes, R. D. (1999). The Safari companion: A guide to watching African mammals. White Rivers Junction, VT: Chelsea Green Publishing Company. [Google Scholar]
- Everett, P. S. , Perrin, M. R. , & Rowe‐Rowe, D. T. (1992). Diet of oribi on farmland in Natal. South African Journal of Wildlife Research, 22, 7–10. [Google Scholar]
- Felsenstein, J. (1985). Phylogenies and the comparative method. American Naturalist, 125, 1–15. 10.1086/284325 [DOI] [PubMed] [Google Scholar]
- Foley, W. J. , & McArthur, C. (1994). The effects and costs of allelochemicals for mammalian herbivores: An ecological perspective In Chivers D. J., & Langer P. (Eds.), The digestive system in mammals (pp. 370–391). Cambridge, UK: Cambridge University Press. [Google Scholar]
- Freeland, W. J. (1991). Plant secondary metabolites: Biochemical coevolution with herbivores In Palo R. T., & Robbins C. T. (Eds.), Plant defenses against mammalian herbivory (pp. 62–79). Boca Raton, FL: CRC Press. [Google Scholar]
- Freeland, W. J. , & Janzen, D. H. (1974). Strategies in herbivory by mammals: The role of plant secondary compounds. American Naturalist, 108, 269–289. 10.1086/282907 [DOI] [Google Scholar]
- Furstenburg, D. , & van Hoven, W. (1994). Condensed tannin as anti‐defoliate agent against browsing by giraffe (Giraffa camelopardalis) in the Kruger National Park. Comparative Biochemistry & Physiology, Series B, 107, 425–431. 10.1016/0300-9629(94)90402-2 [DOI] [Google Scholar]
- Garland, T. , Harvey, P. H. , & Ives, A. R. (1992). Procedures for the analysis of comparative data using phylogenetically independent contrasts. Systematic Biology, 41, 18–32. 10.1093/sysbio/41.1.18 [DOI] [Google Scholar]
- Garland, T. , & Ives, A. R. (2000). Using the past to predict the present: Confidence intervals for regression equations in phylogenetic comparative methods. American Naturalist, 155, 346–364. 10.1086/303327 [DOI] [PubMed] [Google Scholar]
- Gatesy, J. , Amato, G. , Vrba, E. S. , Schaller, G. , & DeSalle, R. (1997). A cladistic analysis of mitochondrial ribosomal DNA from the Bovidae. Molecular Phylogenetics & Evolution, 7, 303–319. 10.1006/mpev.1997.0402 [DOI] [PubMed] [Google Scholar]
- Gatesy, J. , Yelon, D. , DeSalle, R. , & Vrba, E. S. (1992). Phylogeny of the Bovidae (Artiodactyla, Mammalia), based on mitochondrial ribosomal DNA sequences. Molecular Biology & Evolution, 9, 433–446. [DOI] [PubMed] [Google Scholar]
- Geist, V. (1974). On the relationship of social evolution and ecology in ungulates. American Zoologist, 14, 205–220. 10.1093/icb/14.1.205 [DOI] [Google Scholar]
- Georgiadis, N. J. , Kat, P. W. , Oketch, H. , & Patton, J. (1990). Allozyme divergence within the Bovidae. Evolution, 44, 2135–2149. 10.1111/j.1558-5646.1990.tb04317.x [DOI] [PubMed] [Google Scholar]
- Gordon, I. J. , & Illius, A. W. (1994). The functional significance of the browser‐grazer dichotomy in African ruminants. Oecologia, 98, 167–175. 10.1007/BF00341469 [DOI] [PubMed] [Google Scholar]
- Hagerman, A. E. (1987). Radial diffusion method for determining tannin in plant extracts. Journal of Chemical Ecology, 13, 437–449. 10.1007/BF01880091 [DOI] [PubMed] [Google Scholar]
- Hagerman, A. E. (2011). The Tannin Handbook. Retrieved from http://www.users.miamioh.edu/hagermae/ [Google Scholar]
- Hagerman, A. E. , Robbins, C. T. , Weerasuriya, Y. , Wilson, T. C. , & McArthur, C. (1992). Tannin chemistry in relation to digestion. Journal of Range Management, 45, 57–62. 10.2307/4002526 [DOI] [Google Scholar]
- Harborne, J. B. (1993). Introduction to ecological biochemistry, 4th ed New York: Academic Press. [Google Scholar]
- Hassanin, A. , & Douzery, E. J. P. (1999). The tribal radiation of the family Bovidae (Artiodactyla) and the evolution of the mitochondrial cytochrome b gene. Molecular Phylogenetics & Evolution, 13, 227–243. 10.1006/mpev.1999.0619 [DOI] [PubMed] [Google Scholar]
- Hedges, S. B. (2001). Afrotheria: Plate tectonics meets genomics. Proceedings of the National Academy of Sciences, 98(1), 1–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann, R. R. (1973). The ruminant stomach (stomach structure and feeding habits of East African game ruminants). East African Monographs in Biology, vol. 2 Nairobi, Kenya: East African Literature Bureau. [Google Scholar]
- Hofmann, R. R. (1989). Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system. Oecologia, 78, 443–457. 10.1007/BF00378733 [DOI] [PubMed] [Google Scholar]
- Hofmann, R. R. , & Stewart, D. R. M. (1972). Grazer or browser: A classification based on the stomach structure and feeding habits of East African ruminants. Mammalia, 36, 226–240. 10.1515/mamm.1972.36.2.226 [DOI] [Google Scholar]
- Hofmann, R. R. , Streich, W. J. , Fickel, J. , Hummel, J. , & Clauss, M. (2008). Convergent evolution in feeding types: Salivary gland mass differences in wild ruminant species. Journal of Morphology, 269, 240–257. 10.1002/jmor.10580 [DOI] [PubMed] [Google Scholar]
- Holz, P. , Holz, R. M. , & Barnett, J. E. F. (1994). Effects of atropine on medetomidine/ ketamine immobilization in the gray wolf (Canis lupus). Journal of Zoo & Wildlife Medicine, 25, 209–213. [Google Scholar]
- Huey, R. B. , Garland, T. , & Turelli, M. (2019). Revisiting a key innovation in evolutionary biology: Felsenstein’s “Phylogenies and the comparative method”. American Naturalist, 193, 755–772. 10.1086/703055 [DOI] [PubMed] [Google Scholar]
- Iason, G. R. , Hodgson, J. , & Barry, T. N. (1995). Variation in condensed tannin concentration of a temperate grass (Holcus lanatus) in relation to season and reproductive development. Journal of Chemical Ecology, 21, 1103–1112. 10.1007/BF02228314 [DOI] [PubMed] [Google Scholar]
- Iason, G. R. , & Villalba, J. J. (2006). Behavioral strategies of mammal herbivores against plant secondary metabolites: The avoidance–tolerance continuum. Journal of Chemical Ecology, 32, 1115–1132. 10.1007/s10886-006-9075-2 [DOI] [PubMed] [Google Scholar]
- Jarman, P. J. (1974). The social organization of antelope in relation to their ecology. Behaviour, 48, 215–267. [Google Scholar]
- Juntheikki, M.‐R. (1996). Comparison of tannin‐binding proteins in saliva of Scandinavian and North America moose (Alces alces). Biochemical Systematics & Ecology, 24, 595–601. [Google Scholar]
- Kay, R. N. B. , Engelhardt, W. V. , & White, R. G. (1980). The digestive physiology of wild ruminants In Ruckebusch Y., & Thivend P. (Eds.), Digestive physiology and metabolism in ruminants (pp. 743–761). Lancaster, UK: MTP Press. [Google Scholar]
- Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680–685. 10.1038/227680a0 [DOI] [PubMed] [Google Scholar]
- Lamy, E. , da Costa, G. , e Silva, F. C. , Potes, J. , Coelho, A. V. , & Baptista, E. S. (2008). Comparison of electrophoretic protein profiles from sheep and goat parotid saliva. Journal of Chemical Ecology, 34, 388–397. 10.1007/s10886-008-9442-2 [DOI] [PubMed] [Google Scholar]
- Lundberg, P. , & Palo, T. (1993). Resource use, plant defenses, and optimal digestion in ruminants. Oikos, 68, 224–228. 10.2307/3544834 [DOI] [Google Scholar]
- Maddison, D. R. , Schulz, K.‐S. , & Maddison, W. R. (2007). The Tree of Life Web Project. Retrieved from http://tolweb.org [Google Scholar]
- Maddison, W. P. , & Maddison, D. R. (2018). Mesquite: A modular system for evolutionary analysis. Version 3.51 Retrieved from http://www.mesquiteproject.org
- Makkar, H. P. S. (2003). Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin‐rich feeds. Small Ruminant Research, 49, 241–256. 10.1016/S0921-4488(03)00142-1 [DOI] [Google Scholar]
- Manser, M. B. , & Brotherton, P. N. M. (1995). Environmental constraints on the foraging behaviour of a dwarf antelope (Madoqua kirkii). Oecologia, 102, 404–412. 10.1007/BF00341352 [DOI] [PubMed] [Google Scholar]
- Matthee, C. A. , & Davis, S. K. (2001). Molecular insights into the evolution of the family Bovidae: A nuclear DNA perspective. Molecular Biology & Evolution, 18, 1220–1230. 10.1093/oxfordjournals.molbev.a003908 [DOI] [PubMed] [Google Scholar]
- Matthee, C. A. , & Robinson, T. J. (1999). Cytochrome b phylogeny of the family Bovidae: Resolution within the Alcelaphini, Antilopini, Neotragini, and Tragelaphini. Molecular Phylogenetics & Evolution, 12, 31–46. 10.1006/mpev.1998.0573 [DOI] [PubMed] [Google Scholar]
- McArthur, C. , Hagerman, A. E. , & Robbins, C. T. (1991). Physiological strategies of mammalian herbivores against plant defenses In Palo R. T., & Robbins C. T. (Eds.), Plant defenses against mammalian herbivory (pp. 103–114). Boca Raton, FL: CRC Press. [Google Scholar]
- McNaughton, S. J. , Tarrants, J. L. , McNaughton, M. M. , & Davis, R. D. (1985). Silica as a defense against herbivory and a growth promotor in African grasses. Ecology, 66, 528–535. 10.2307/1940401 [DOI] [Google Scholar]
- Mehansho, H. , Butler, L. G. , & Carlson, D. M. (1987). Dietary tannins and salivary proline‐rich proteins: Interaction, induction, and defense mechanisms. Annual Review of Nutrition, 7, 423–440. [DOI] [PubMed] [Google Scholar]
- Mehansho, H. , Hagerman, A. , Clements, S. , Butler, L. , Rogler, J. , & Carlson, D. M. (1983). Modulation of proline‐rich protein biosynthesis in rat parotid glands by sorghums with high tannin levels. Proceedings of the National Academy of Sciences, 80(13), 3948–3952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meredith, R. W. , Janecka, J. E. , Gatesy, J. , Ryder, O. A. , Fisher, C. A. , Teeling, E. C. , … Murphy, W. J. (2011). Impacts of the Cretaceous terrestrial revolution and KPg extinction on mammal diversification. Science, 334, 521–524. 10.1126/science.1211028 [DOI] [PubMed] [Google Scholar]
- Mole, S. (1993). The systematic distribution of tannins in the leaves of angiosperms: A tool for ecological studies. Biochemical Systematics & Ecology, 21, 833–846. 10.1016/0305-1978(93)90096-A [DOI] [Google Scholar]
- Mole, S. , Butler, L. G. , & Iason, G. (1990). Defense against dietary tannin in herbivores: A survey for proline‐rich salivary proteins in mammals. Biochemical Systematics & Ecology, 18, 287–293. 10.1016/0305-1978(90)90073-O [DOI] [Google Scholar]
- Mole, S. , Rogler, J. , Morell, C. J. , & Butler, L. G. (1990). Herbivore growth reduction by tannins: Use of Waldbauer ratio techniques and manipulation of salivary protein production to elucidate mechanisms of action. Biochemical Systematics & Ecology, 18, 183–197. 10.1016/0305-1978(90)90056-L [DOI] [Google Scholar]
- Muller, K. (2013). What drives the seasonal movements of African Elephants (Loxodonta africana) in Ithala Game Reserve? Unpubl. MSc thesis. Pietermaritzburg, South Africa: University of KwaZulu‐Natal. [Google Scholar]
- Münkemüller, T. , Lavergne, S. , Bzeznik, B. , Dray, S. , Jombart, T. , Schiffers, K. , & Thuiller, W. (2012). How to measure and test phylogenetic signal. Methods in Ecology & Evolution, 3, 743–756. 10.1111/j.2041-210X.2012.00196.x [DOI] [Google Scholar]
- Nobler, J. D. , Camp, M. J. , Crowell, M. M. , Shipley, L. A. , Dadabay, C. , Rachlow, J. L. , … Forbey, J. S. (2019). Preferences of specialist and generalist mammalian herbivores for mixtures versus individual plant secondary metabolites. Journal of Chemical Ecology, 45, 74–85. 10.1007/s10886-018-1030-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orians, C. M. , & Ward, D. (2010). Evolution of plant defenses in nonindigenous environments. Annual Review of Entomology, 55, 439–459. 10.1146/annurev-ento-112408-085333 [DOI] [PubMed] [Google Scholar]
- Ouchterlony, Ö. (1953). Antigen‐antibody reactions in gels. Acta Pathologica Microbiologia Scandinavica, 32, 231–240. 10.1111/j.1699-0463.1953.tb00247.x [DOI] [PubMed] [Google Scholar]
- Owen‐Smith, N. (1982). Factors influencing the consumption of plant products by large herbivores In Huntley B. J., & Walker B. H. (Eds.), Ecology of Tropical Savannas (pp. 359–404). Berlin, Germany: Springer. [Google Scholar]
- Owen‐Smith, N. , & Chafota, J. (2012). Selective feeding by a megaherbivore, the African elephant (Loxodonta africana). Journal of Mammalogy, 93, 698–705. [Google Scholar]
- Owen‐Smith, R. N. (1988). Megaherbivores: The influence of very large body size on ecology. Cambridge, UK: Cambridge University Press. [Google Scholar]
- Pérez‐Barberìa, F. J. , Elston, D. A. , Gordon, I. J. , & Illius, A. W. (2004). The evolution of phylogenetic differences in the efficiency of digestion of ruminants. Proceedings of the Royal Society, Series B, 271, 1081–1090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pérez‐Barbería, F. J. , Gordon, I. J. , & Illius, A. W. (2001). Phylogenetic analysis of stomach adaptation in digestive strategies in African ruminants. Oecologia, 129, 498–508. 10.1007/s004420100768 [DOI] [PubMed] [Google Scholar]
- Pérez‐Barberìa, F. J. , Pérez‐Fernandez, E. , Robertson, E. , & Alvarez‐Enriquez, B. (2008). Does the Jarman‐Bell principle at intra‐specific level explain sexual segregation in polygynous ungulates? Sex differences in forage digestibility in Soay sheep. Oecologia, 157, 21–30. 10.1007/s00442-008-1056-4 [DOI] [PubMed] [Google Scholar]
- Prinz, J. F. , & Lucas, P. W. (2000). Saliva tannin interactions. Journal of Oral Rehabilitation, 27, 991–994. 10.1046/j.1365-2842.2000.00578.x [DOI] [PubMed] [Google Scholar]
- Provenza, F. D. , & Balph, D. F. (1987). Diet learning by domestic ruminants: Theory, evidence and practical implications. Applied Animal Behaviour Science, 18, 211–232. 10.1016/0168-1591(87)90218-8 [DOI] [Google Scholar]
- Revell, L. J. , Harmon, L. J. , & Collar, D. C. (2008). Phylogenetic signal, evolutionary process, and rate. Systematic Biology, 57, 591–601. 10.1080/10635150802302427 [DOI] [PubMed] [Google Scholar]
- Rhoades, D. F. , & Cates, R. G. (1976). Toward a general theory of plant antiherbivore chemistry In Wallace J. W., & Mansell R. L. (eds) Biochemical interaction between plants and insects. Recent advances in phytochemistry, vol. 10, pp. 168–2213. Boston, MA: Springer. [Google Scholar]
- Robbins, C. T. , Hagerman, A. E. , Austin, P. J. , McArthur, C. , & Hanley, T. (1991). Variation in mammalian physiological responses to a condensed tannin and its ecological implications. Journal of Mammalogy, 72, 481–486. 10.2307/1382130 [DOI] [Google Scholar]
- Robbins, C. T. , Hanley, T. A. , Hagerman, A. E. , Hjeljord, O. , Baker, D. L. , Schwartz, C. C. , & Mautz, W. W. (1987). Role of tannins in defending plants against ruminants: Reduction in protein availability. Ecology, 68, 98–107. 10.2307/1938809 [DOI] [PubMed] [Google Scholar]
- Robbins, C. T. , Mole, S. , Hagerman, A. E. , & Hanley, T. A. (1987). Role of tannins in defending plants against ruminants: Reduction in dry matter digestion? Ecology, 68, 1606–1615. 10.2307/1939852 [DOI] [PubMed] [Google Scholar]
- Robbins, C. T. , Spalinger, D. E. , & Van Hoven, W. (1995). Adaptation of ruminants to browse and grass diets: Are anatomical‐based browser‐grazer interpretations valid? Oecologia, 103, 208–213. 10.1007/BF00329082 [DOI] [PubMed] [Google Scholar]
- Rohner, C. , & Ward, D. (1997). Chemical and mechanical defense against herbivory in two sympatric species of desert Acacia . Journal of Vegetation Science, 8, 717–726. [Google Scholar]
- Schmitt, M. H. (2017). The influences of plant secondary metabolites on the foraging behaviour and carrying capacities of the African Elephant, Loxodonta africana. Unpubl. PhD thesis. Pietermaritzburg, South Africa: University of KwaZulu‐Natal. [Google Scholar]
- Schmitt, M. H. , Shuttleworth, A. , Shrader, A. M. , & Ward, D. (2020). The role of volatile plant secondary metabolites as pre‐ingestive cues and potential toxins dictating diet selection by African elephants. Oikos, 129, 24–34. 10.1111/oik.06665 [DOI] [Google Scholar]
- Schmitt, M. H. , Shuttleworth, A. , Ward, D. , & Shrader, A. M. (2018). African elephants use plant odours to make foraging decisions across multiple spatial scales. Animal Behaviour, 141, 17–27. 10.1016/j.anbehav.2018.04.016 [DOI] [Google Scholar]
- Schmitt, M. H. , Ward, D. , & Shrader, A. M. (2016). Incorporating secondary metabolites, tannin‐binding proteins, and diet breadth into carrying‐capacity models for African Elephants. Ecological Modelling, 332, 8–18. 10.1016/j.ecolmodel.2016.03.016 [DOI] [Google Scholar]
- Schmitt, M. H. , Ward, D. , & Shrader, A. M. (2020). Salivary tannin‐binding proteins: A foraging advantage for goats? Livestock Science, 234, e103974 10.1016/j.livsci.2020.103974 [DOI] [Google Scholar]
- Scully, C. (2003). Drug effects on salivary glands: Dry mouth. Oral Diseases, 9, 165–176. 10.1034/j.1601-0825.2003.03967.x [DOI] [PubMed] [Google Scholar]
- Searle, K. R. , & Shipley, L. A. (2008). The comparative feeding behaviour of large browsing and grazing herbivores In Gordon I. J., & Prins H. H. T. (Eds.), The ecology of browsing and grazing (pp. 117–148). Dordrecht, The Netherlands: Springer. [Google Scholar]
- Shaw, J. A. (2011). Adaptive resource use in a re‐introduced Black Rhinoceros population. Unpubl. PhD thesis Johannesburg, South Africa: University of the Witwatersrand. [Google Scholar]
- Shimada, T. (2006). Salivary proteins as a defense against dietary tannins. Journal of Chemical Ecology, 32, 1149–1163. 10.1007/s10886-006-9077-0 [DOI] [PubMed] [Google Scholar]
- Shrader, A. M. , Bell, C. , Bertolli, L. , & Ward, D. (2012). Forest or the trees: At what scale do elephants make foraging decisions? Acta Oecologica, 42, 3–12. 10.1016/j.actao.2011.09.009 [DOI] [Google Scholar]
- Skopec, M. M. , Hagerman, A. E. , & Karasov, W. H. (2004). Do salivary proline‐rich proteins counteract dietary hydrolyzable tannin in laboratory rats? Journal of Chemical Ecology, 30, 1679–1692. 10.1023/B:JOEC.0000042395.31307.be [DOI] [PubMed] [Google Scholar]
- Springer, M. S. , Cleven, G. C. , Madsen, O. , De Jong, W. W. , Waddell, V. G. , Amrine, H. M. , & Stanhope, M. J. (1997). Endemic African mammals shake the phylogenetic tree. Nature, 388, 61–64. 10.1038/40386 [DOI] [PubMed] [Google Scholar]
- Stears, K. , & Shrader, A. M. (2020). Coexistence between wildlife and livestock is contingent on cattle density and season but not differences in body size. PLoS ONE, 15, e0236895 10.1016/j.anbehav.2015.07.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steuer, P. , Südekum, K.‐H. , Müller, D. W. H. , Kaandorp, J. , Clauss, M. , & Hummel, J. (2013). Fibre digestibility in large herbivores as related to digestion type and body mass — an in vitro approach. Comparative Biochemistry and Physiology A, 164, 319–326. 10.1016/j.cbpa.2012.10.026 [DOI] [PubMed] [Google Scholar]
- Symonds, M. R. E. , & Blomberg, S. P. (2014). A primer on phylogenetic generalised least squares In Garamszegi L. Z. (Eds.), Modern phylogenetic comparative methods and their application in evolutionary biology (Ch. 5, pp. 105–130). Dordrecht, The Netherlands: Springer. [Google Scholar]
- Tabuce, R. , Asher, R. J. , & Lehmann, T. (2008). Afrotherian mammals: A review of current data. Mammalia, 72, 2–14. 10.1515/MAMM.2008.004 [DOI] [Google Scholar]
- Vaithiyanathan, S. , Mishra, J. , Sheikh, Q. , & Kumar, R. (2001). Salivary gland tannins binding proteins of sheep and goat. Indian Journal of Animal Science, 71, 1131–1134. [Google Scholar]
- Van Soest, P. J. (1994). Nutritional ecology of the ruminant, 2nd ed. Ithaca, NY: Cornell University Press. [Google Scholar]
- Ventura‐Cordero, J. , Sandoval‐Castro, C. A. , Torres‐Acosta, J. F. J. , & Capetillo‐Leal, C. M. (2017). Do goats have a salivary constitutive response to tannins? Journal of Applied Animal Research, 45, 29–34. 10.1080/09712119.2015.1102728 [DOI] [Google Scholar]
- Ward, D. (2000). Do polyandrous shorebirds trade off egg size with egg number? Journal of Avian Biology, 31, 473–478. 10.1034/j.1600-048X.2000.310406.x [DOI] [Google Scholar]
- Ward, D. , Muller, K. , & Shrader, A. M. (2017). Soil fertility on granite and sedimentary soils is associated with seasonal differences in foraging by elephants. Plant & Soil, 413, 73–81. 10.1007/s11104-016-3067-y [DOI] [Google Scholar]
- Ward, D. , & Seely, M. K. (1996). Adaptation and constraint in the evolution of the physiology and behavior of the Namib desert tenebrionid beetle genus Onymacris . Evolution, 50, 1231–1240. [DOI] [PubMed] [Google Scholar]
- Westoby, M. (1978). What are the biological bases of varied diets? American Naturalist, 112, 627–631. 10.1086/283303 [DOI] [Google Scholar]
- Windels, S. K. , & Hewitt, D. G. (2011). Effects of plant secondary compounds on nutritional carrying capacity estimates of a browsing ungulate. Rangeland Ecology & Management, 64, 264–275. 10.2111/REM-D-10-00078.1 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All information is present here (Table 1; Figure 2).