

Abstract
Endometrial cancer is one of the three major malignant tumors of the female reproductive system. Although cyclin-dependent kinase 9 (CDK9) has a definitive pathogenic role in various types of cancer, little is known concerning its function in endometrial cancer. Our study was conducted to evaluate the expression and therapeutic potential of CDK9 in endometrial cancer. CDK9 expression was determined by immunohistochemistry in endometrial cancer tissues constructed with paired primary, metastatic, and recurrent tumor tissues from 32 endometrial cancer patients. Small interfering RNA (siRNA) and inhibitors of CDK9 were used to evaluate the effect of CDK9 inhibition on the anti-apoptotic activity and proliferation in endometrial cancer cells. Colony formation assay and wound-healing assays were adopted to assess clonal formation and migratory capacity. The results of the immunohistochemistry demonstrated that CDK9 was highly expressed in the human endometrial cancer cell lines; moreover, it was elevated in metastatic and recurrent endometrial tumor tissue compared when compared with that in patient-matched primary endometrial tumor tissue. Knockdown of CDK9 with siRNA and inhibition of CDK9 activity with the inhibitor suppressed cell proliferation and promoted apoptosis in endometrial cancer. In conclusion, our results provide evidence that CDK9 may be a potential prognostic biomarker and a promising therapeutic target for the treatment of endometrial cancer in the future.
Keywords: endometrial cancer, cyclin-dependent kinase 9, siRNA, LDC067, biomarker
Introduction
Endometrial cancer is one of the three commonly diagnosed gynecological malignancies of the genital tract. According to US cancer statistics, endometrial cancer ranks fourth among women's invasive tumors worldwide, and there were an estimated 61,880 new patients and 12,160 deaths in 2019 (1). In China, the incidence of endometrial cancer is increasing and the age of onset is becoming younger (2,3). Contrary to the overall downward trend in the most common cancers, mortality from endometrial cancer increased by about 2% per year between 2011 and 2015 (4). Patients with early endometrial cancer have a relatively good prognosis, but those with advanced endometrial cancer have a poor response to treatment and their prognoses are worse. The prognosis of endometrial cancer is affected by many factors including tumor grade, stage, histologic subtype, lymph node metastasis and myometrial invasion. Lack of effective treatment for patients with advanced endometrial cancer reflects the need to study the molecular mechanisms of endometrial cancer occurrence and development (5).
Cyclin-dependent kinases (CDKs) are a family of serine/threonine kinases whose activity is derived from heterodimeric complexes composed of catalytic kinase subunits and regulatory cyclin subunits (6,7). They play an important role in controlling the cell cycle, transcription, and neuronal function (6). Multiple genetic and epigenetic events lead to a general overactivity of the cell cycle CDK in human cancers, and its inhibition may lead to both cell cycle arrest and apoptosis (8). With regard to the role of CDKs in the cell cycle or transcriptional regulation, CDK1, CDK2, CDK3, CDK4 and CDK6 drive cell cycle, CDK7 regulates cell cycle, CDK7, CDK8, CDK9, CDK10 and CDK11 regulate transcription, and CDK5 regulates neuronal differentiation (6). Of the multiple CDKs involved in transcriptional regulation, CDK9 is considered to be the most important one (9). CDK9 is a Cdc2-like Ser/Thr kinase that mainly binds to cyclin T1 and constitutes the basal transcription factor, positive transcription elongation factor b (p-TEFb), which phosphorylates the carboxy-terminal domain (CTD) of the large subunit of RNA polymerase II (RNAPII) and reaches the RNA transcription elongation, thereby regulating cell proliferation, differentiation, apoptosis, and DNA repair (10,11). CDK9 is involved in the transcription of most eukaryotic cells, including human tissues (12). CDK9 expression is significantly increased in different cellular processes and different tissues, and has an important contribution to the progression of various types of cancer (10). Recently, studies have found that CDK9 is of critical importance in the development of many cancers, including leukemia, cervical cancer, prostate cancer, glioblastoma, breast cancer, melanoma and lung cancer (13–19).
In the present study, in order to investigate the role of CDK9 in the progression of endometrial cancer, we firstly evaluated its expression in endometrial cancer tissue, and then used siRNA and inhibitors to suppress CDK9.
Materials and methods
Human endometrial cancer tissues
The specimens used in this study were collected from 32 patients with endometrial cancer treated at the Second Xiangya Hospital of Central South University (Changsha, Hunan, China) from January 2002 to December 2017. The age of the patients ranged from 39 to 69 years (average, 60.8 years). All patients accepted primary surgery; the primary and metastatic tumor tissues were obtained by the primary surgery. Upon tumor recurrence, 15 patients had pelvic recurrence, 7 patients had vaginal metastasis, 1 patient had vulvar metastasis, 3 patients had inguinal lymph node metastases, 3 patients had bone metastases, 2 patients had lung metastases, and 1 patient had liver metastases. The recurrent tumor tissues were obtained by second surgery or biopsy. The tumor tissues of these 32 patients were collected at three different stages, including: Primary tumor tissue, postoperative metastatic tissue, and recurrent tumor tissue. The tumor tissues were fixed with formaldehyde and embedded in paraffin. We collected data concerning patient age; surgical method; histologic subtype; surgical-pathological stage; pathological grade; recurrence site; recurrence tissue acquisition method; patient survival status at the end of follow-up: Progression-free survival (PFS), defined as the interval between the date of primary surgery and the objective tumor progression or death; overall survival (OS), defined as the interval between the date of primary surgery to last follow-up or death (Table I). The study was approved by the Ethics Committee of The Second Xiangya Hospital of Central South University (IRB protocol number: Study 181). All patients signed a consent form for their tissues and clinical information to be used for this research.
Table I.
Clinical data for the endometrial cancer patients (N=32).
| Patient | Age (years) | Primary surgery | Histologic subtype | Stage | Grade | Recurrence site | Method of recurrent tissue obtained | PFS (months) | OS (months) | Patient status |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 54 | TH and BSO and lymphadenectomy | Carcinosarcoma | IVB | 3 | Vaginal | Biopsy | 4 | 9 | Deceased |
| 2 | 63 | RH and BSO and lymphadenectomy | Clear cell carcinoma | IVB | 3 | Vaginal | Biopsy | 6 | 11 | Deceased |
| 3 | 64 | RH and BSO and lymphadenectomy | Mixed cell tumors | IVA | 3 | Vaginal | Biopsy | 7 | 14 | Deceased |
| 4 | 58 | RH and BSO and lymphadenectomy | Serous carcinoma | IVB | 3 | Vaginal | Biopsy | 8 | 17 | Deceased |
| 5 | 55 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC2 | 2 | Pelvic | Biopsy | 9 | 20 | Deceased |
| 6 | 66 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 3 | Vaginal | Biopsy | 11 | 25 | Deceased |
| 7 | 68 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIB | 2 | Pelvic | Biopsy | 12 | 26 | Deceased |
| 8 | 53 | RH and BSO and lymphadenectomy | Serous carcinoma | IVB | 3 | Bone | Biopsy | 13 | 27 | Deceased |
| 9 | 67 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC2 | 3 | Pelvic | Biopsy | 13 | 28 | Deceased |
| 10 | 61 | RH and BSO and lymphadenectomy | Mucinous carcinoma | IIIB | 2 | Inguinal lymph node | Biopsy | 16 | 30 | Deceased |
| 11 | 39 | TH and BSO and lymphadenectomy | Carcinosarcoma | IVB | 3 | Lung | Biopsy | 17 | 31 | Deceased |
| 12 | 59 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 2 | Vaginal | Biopsy | 16 | 36 | Deceased |
| 13 | 67 | RH and BSO and lymphadenectomy | Serous carcinoma | IVB | 3 | Liver | Biopsy | 20 | 37 | Deceased |
| 14 | 62 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIA | 2 | Pelvic | Biopsy | 12 | 40 | Deceased |
| 15 | 69 | RH and BSO and lymphadenectomy | Serous carcinoma | IVB | 3 | Inguinal lymph node | Biopsy | 13 | 54 | Deceased |
| 16 | 65 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIA | 2 | Pelvic | Biopsy | 15 | 55 | Deceased |
| 17 | 63 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC2 | 2 | Pelvic | Biopsy | 16 | 57 | Deceased |
| 18 | 67 | RH and BSO and lymphadenectomy | Mixed cell tumors | IVA | 3 | Lung | Biopsy | 16 | 63 | Deceased |
| 19 | 63 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 2 | Pelvic | Biopsy | 18 | 70 | Deceased |
| 20 | 62 | RH and BSO and lymphadenectomy | Clear cell carcinoma | IIIB | 2 | Bone | Biopsy | 20 | 72 | Deceased |
| 21 | 60 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIB | 3 | Pelvic | Biopsy | 25 | 80 | Deceased |
| 22 | 61 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 2 | Pelvic | Biopsy | 26 | 85 | Deceased |
| 23 | 67 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIA | 3 | Pelvic | Biopsy | 30 | 96 | Deceased |
| 24 | 66 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 3 | Bone | Biopsy | 33 | 97 | Deceased |
| 25 | 53 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 2 | Pelvic | Biopsy | 40 | 109 | Deceased |
| 26 | 64 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIA | 3 | Inguinal lymph node | Biopsy | 43 | 120 | Deceased |
| 27 | 69 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIB | 2 | Pelvic | Biopsy | 43 | 127 | Deceased |
| 28 | 60 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIA | 3 | Vaginal | Biopsy | 51 | 139 | Deceased |
| 29 | 59 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 3 | Pelvic | Biopsy | 53 | 140 | Deceased |
| 30 | 58 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIC1 | 3 | Vulvar | Biopsy | 53 | 148 | Deceased |
| 31 | 52 | TH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIB | 1 | Pelvic | Biopsy | 53 | 155 | Alive |
| 32 | 51 | RH and BSO and lymphadenectomy | Endometrioid carcinoma | IIIA | 2 | Pelvic | Biopsy | 57 | 163 | Alive |
TH, total hysterectomy; RH, radical hysterectomy; BSO, bilateral salpingo-oophorectomy; PFS (progression-free survival), interval between the date of primary surgery and the objective tumor progression or death; OS (overall survival), interval between the date of primary surgery to last follow-up or death.
Immunohistochemistry
CDK9 expression was determined by standard immunohistochemical protocol. In short, paraffin-embedded slides (4 µM) were baked at 60°C for 1 h and then dewaxed in xylene and rehydrated by fractionated ethanol (100 and 95%). Antigen was extracted by Dako Target Retrieval Solution (Dako; Agilent Technologies, Inc.), and incubated with 3% H2O2 for 10 min to eliminate endogenous peroxidase activity. Thereafter, the slides were sealed with goat serum for 1 h, and then polyclonal rabbit antibodies against human CDK9 (cat. no. 2316; 1:50 dilution in 1% bovine serum albumin; Cell Signaling Technology, Inc.) were added and incubated overnight. The slides were fully covered with the anti-rabbit SignalStain® Boost Detection Reagent (Cell Signaling Technology) and placed in a humid chamber for 30 min, and then SignalStain® DAB (Cell Signaling Technology) was added to the slides to reveal the staining intensity. Subsequently, the slides were counterstained with Hematoxylin QS (Vector Laboratories, Burlingame, CA, USA) and fixed with VectaMount AQ (Vector Laboratories, Inc.) for long-term storage. Two independent pathologists evaluated the percentage of cells with positive nuclear staining on the immunostained slides. The expression of CDK9 was divided into five levels: i) 1+, <10% positive cells; ii) 2+, 10–25% positive cells; iii) 3+, 26–50% positive cells; iv) 4+, 51–75% positive cells; v) 5+, >75% positive cells. Based on the CDK9 expression scores of the primary endometrial cancer samples, patients were divided into the following two groups: Low CDK9 group (CDK9 staining score ≤2) and high CDK9 group (CDK9 staining score ≥3). CDK9 stained images were captured with an Olympus BX51microscope (original magnification, ×400) (Olympus Corporation of the Americas) and a Spot RT digital camera (Diagnostic Instruments).
Endometrial cancer cell lines and reagents
The human endometrial cancer cell lines HEC-1A, ARK-2H, EC-1B, RL95-2 and SPAC1S were obtained from the American Type Culture Collection (ATCC; Rockville, MD, USA). AN3CA was obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ; Braunschweig, Germany). Ishikawa cells were obtained from the Central Cell Services Facility at the Cancer Research UK (CRUK). All endometrial cancer cell lines were maintained in RPMI-1640 (Invitrogen; Life Technologies) containing 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin. Monoclonal rabbit anti-human CDK9 antibody (cat. no. 2316; Cell Signaling Technology, Inc.) was purchased from Cell Signaling Technology, Inc.. The highly selective CDK9 inhibitor LDC000067 (abbreviated as LDC067) was purchased from Selleck Chemicals. Human non-specific small interfering RNA (siRNA) and CDK9-targeting siRNA (59-GCUGCUAAUGUGCUUAUCA-39) were purchased from Merck KGaA. Lipofectamine RNAiMax was purchased from Thermo Fisher Scientific, Inc. Apoptosis-related antibodies were obtained from Cell Signaling Technology, Inc..
Western blotting
Total protein lysate was extracted from cells using RIPA lysis buffer (Sigma-Aldrich; Merck KGaA). Thirty micrograms of the protein sample was separated on NuPage 4–12% Bis-Tris gel (Thermo Fisher Scientific, Inc.) and transferred to nitrocellulose membranes (Bio-Rad Laboratories, Inc.), and then incubated with the following primary antibodies in 5% milk with tris-buffered saline Tween 20: CDK9 (cat. no. 2316; 1:1,000 dilution; Cell Signaling Technology, Inc.), Mcl-1 (cat. no. 39224; 1:1,000 dilution; Cell Signaling Technology, Inc.), PARP (cat. no. 9532; 1:1,000 dilution; Cell Signaling Technology, Inc.), Bax (cat. no. 2772; 1:1,000 dilution; Cell Signaling Technology, Inc.) and tubulin (cat. no. 3873; 1:1,000 dilution; Cell Signaling Technology, Inc.). Next, the membranes were further incubated with a goat anti-rabbit IRDye 800CW (926–32,211; 1:5,000 dilution; Li-Cor Biosciences) or goat anti-mouse IRDye 680LT secondary antibody (926–68,020; 1:15,000 dilution; Li-Cor Biosciences) at room temperature for 2 h. The membranes were then scanned by an Odyssey CLx device (Li-Cor Biosciences) to detect bands. Finally, protein bands were quantified by densitometry with Odyssey v. 3.0 software (Li-Cor Biosciences).
Knockout of CDK9 through siRNA transfection and MTT assay
CDK9 in endometrial cancer cells was knocked out by transfection of synthetic CDK9 siRNA. AN3CA and SPAC1S cells were seeded into 12-well plates at a density of 4×104 cells/well or into 96-well plates at a density of 2×103 cells/well. CDK9 siRNA (5′-GCUGCUAAUGUGCUUAUCAUCA-3′) was synthesized with Lipofectamine RNAiMax reagent (Thermo Fisher Scientific, Inc.) at concentrations of 10, 20 and 40 nM, respectively, according to the protocol provided by the manufacturer. Negative control group used non-specific siRNA (40 nM). Five days after CDK9 siRNA transfection, proteins from AN3CA and SPAC1S cells were extracted for analysis of cell proliferation by MTT assay. Then, at the end of cell transfection, 20 µl MTT reagent (5 mg/ml; Merck KGaA) was added to each plate, and then cells were incubated at 37°C for another 4 h. After removing the supernatant, the obtained intracellular formazan crystals were dissolved in 100 µl per well of acid isopropanol. SpectraMax 340PC microplate reader (Molecular Devices, LLC) was used to evaluate the absorbance of the samples.
Suppression of CDK9 expression by LDC067 inhibitor and MTT assay
LDC067 is able to decrease the expression of CDK9 in various cancer cell lines at a concentration of 10 µM (20). AN3CA and SPAC1S cells were plated into a 96-well plate at a density of 4×103 cells/well, or a 6-well plate at a density of 6×105 cells/well. Prior to subsequent experiments, the cells were incubated for 2, 3, or 5 days with increasing LDC067 concentrations (0, 1.25, 2.5, 5.0 and 10 µM). After 5 days of LDC067 treatment, MTT assay was used to study cell proliferation of AN3CA and SPAC1S cells.
Colony formation assay
Colony formation assay is a commonly used method to assess cell viability and proliferation capacity. AN3CA and SPAC1S cells were seeded into 12-well plates at a density of 100 cells/well and treated with different concentrations of CDK9 inhibitor LDC067 (0, 2.5, 5 and 10 µM). Incubation lasted for 14 days, and then methanol was used to fix these colonies for 10 min. After being washed three times with PBS, the cells were stained with 10% Giemsa solution (Merck KGaA) for 20 min. Colonies were rinsed under running water and then dried naturally. Finally, an Olympus digital camera was used to capture images of the (original magnification, ×1) colonies.
Wound-healing assay
Wound healing assay was used to analyze the effect of CDK9 inhibitor LDC067 on the migration activity of endometrial cancer cells. AN3CA and SPAC1S cells were plated into 6-well at a density of 4×105 cells/well plates and incubated overnight. A 30-µl sterile pipette was used to scrape two parallel lines at the adhered cell layer. Next, 5 µM of LDC067 was added to the medium, and incubated in a low serum medium containing 2% fetal calf serum for 72 h (21,22). Although the serum-free medium can prevent cell proliferation, it also increases the number of apoptotic cells, which can affect the conclusion of the wound healing assay. After 0, 24 and 48 h of treatment with LDC067, images of the wounds were obtained respectively, with a Nikon microscope (original magnification, ×200) (diagnostic instrument) equipped with Zen Imaging software (Carl Zeiss). Wound width was assessed by the distance between the two edges of the scratch at 5 sites in each image. The relative cell migration distance was calculated by the following formula: Wound width at the 0 h time point-Wound width at the observed time point/Wound width at the 0 h time point.
Statistical analysis
Statistical analysis was carried out with the assistance of GraphPad Prism 7 (GraphPad Software, Inc.) or SPSS 26.0 (IBM, Corp.). CDK9 scores in primary tumor tissue, recurrent tumor tissue, and metastatic tumor tissue were compared through Friedman's test. PFS and OS were analyzed using Kaplan-Meier survival curves with log-rank tests for significance. The χ2 test was used to evaluate the relationship between CDK9 expression and endometrial cancer clinical-pathological parameters. One-way ANOVA was used for the group comparison, and Tukey's test was performed for pairwise comparison. We considered a difference statistically significant at P-value <0.05.
Results
Expression of CDK9 in primary endometrial cancer tissue and its matched metastasis and recurrent endometrial cancer tissue
We used immunohistochemistry to compare the expression of CDK9 in different endometrial cancer tissues. Based on CDK9 expression scores in primary endometrial cancer samples, patients were divided into the following two groups: Low CDK9 group (CDK9 staining score ≤2; 59.4%) and high CDK9 group (CDK9 staining score ≥3; 40.6%) (Fig. 1). There was no statistical difference in the surgical-pathological stage (P=0.5993, based on the χ2 test), pathological grade (P=0.8206, based on the χ2 test), and histologic subtype (P=0.7224, based on the χ2 test) between the patients with low CDK9 expression and those with high CDK9 expression (Table II). We examined the expression of CDK9 in primary, metastatic, and recurrent tumor specimens from 32 patients with endometrial cancer. CDK9 is mainly located in the nucleus. Although the expression level of CDK9 in metastatic and recurrent endometrial cancer tissues was not statistically different (P>0.999, based on the Friedman's test), the CDK9 expression level in primary endometrial cancer tissues was significantly lower than that in metastatic and recurrent endometrial cancer tissues. These differences were statistically significant (metastatic vs. primary, P<0.001; recurrent vs. primary, P<0.001, based on Friedman's test) (Fig. 2A).
Figure 1.

Different CDK9 staining intensities and H&E staining of endometrial cancer tissues. According to the CDK9 staining in the tumor samples, the staining patterns were divided into 5 groups: i) l<10% positive cells (1+); ii) 10–25% positive cells (2+); iii) 26–50% positive cells (3+); iv) 51–75% positive cells (4+); v) >75% positive cells (5+). (Original magnification, ×400). CDK9, cyclin-dependent kinase 9; H&E, hematoxylin and eosin.
Table II.
Association between CDK9 expression and clinicopathological features of the endometrial cancer cases.
| CDK9 expression | ||||
|---|---|---|---|---|
| Clinicopathological features | No. of cases | Low | High | P-value |
| All patients | 32 | 19 | 13 | |
| Stage | ||||
| IV | 9 | 6 | 3 | 0.5993 |
| III | 23 | 13 | 10 | |
| Grade | ||||
| 3 | 18 | 11 | 7 | 0.8206 |
| ≤2 | 14 | 8 | 6 | |
| Histologic subtype | ||||
| Endometrioid carcinoma | 21 | 12 | 9 | 0.7224 |
| Mucinous carcinoma | 1 | 1 | ||
| Serous carcinoma | 4 | 3 | 1 | |
| Clear cell carcinoma | 2 | 2 | 0 | |
| Mixed cell tumors | 2 | 1 | 1 | |
| Carcinosarcoma | 2 | 0 | 2 | |
CDK9, cyclin-dependent kinase 9.
Figure 2.

Higher expression of CDK9 is present in metastatic and recurrent endometrial cancer tissues compared with that found in the patient matched primary tumors and CDK9 is correlated with poor patient prognosis. (A) Distribution of CDK9 immunohistochemical staining scores among primary, metastatic, and recurrent endometrial cancer tissues. (B and C) Correlation between expression of CDK9 in the primary endometrial cancer tissues (Low, CDK9 staining ≤2+; High, CDK9 staining ≥3+) and PFS (B) or OS (C) in endometrial cancer patients by Kaplan-Meier survival curve analysis. CDK9, cyclin-dependent kinase 9; PFS, progression-free survival; OS, overall survival.
Relationship between CDK9 expression and patient prognosis
Kaplan-Meier survival curve showed that compared with patients with endometrial cancer in the CDK9 low expression group (CDK9 staining score ≤2), those in the CDK9 high expression group (CDK9 staining score ≥3) had significantly shorter PFS and OS, and the differences were statistically significant (P<0.0001, based on the log-rank test) (Fig. 2B and C). Specifically, the median PFS and OS of the CDK9 low-expression group were 39 and 96 months, while the median PFS and OS of the CDK9 high-expression group were 14 and 26 months. These results suggest that elevated CDK9 expression is closely associated with poor prognosis in patients with endometrial cancer.
Expression of CDK9 protein in various endometrial cancer cell lines
We detected the expression of CDK9 protein in different endometrial cancer cell lines (AN3CA, ARK-2, HEC-1A, HEC-1B, lshikawa, RL95-2 and SPAC1S) by western blot analysis. The results showed that CDK9 was expressed in all endometrial cancer cell lines, as shown in Fig. 3.
Figure 3.
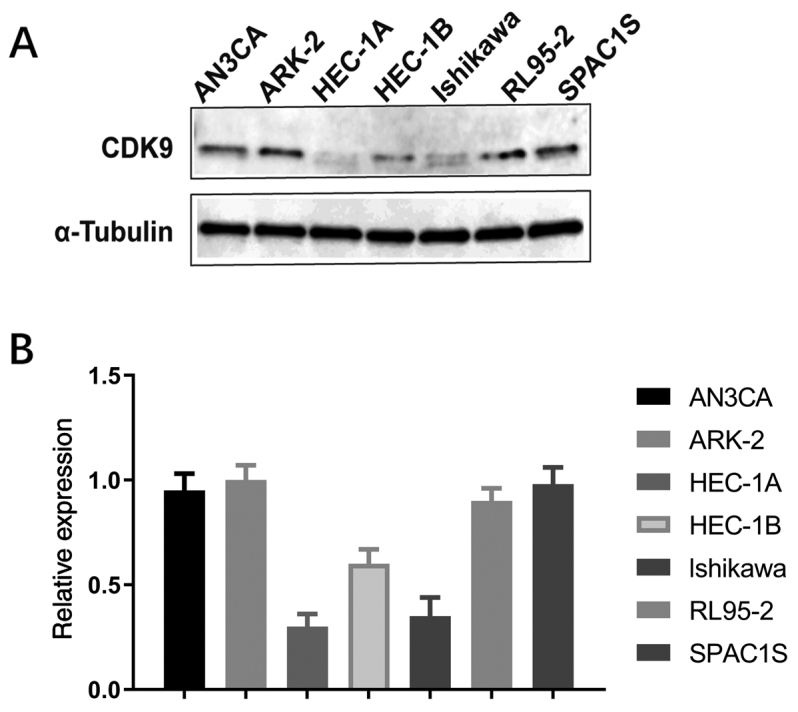
CDK9 expression in endometrial cancer cell lines. (A) Expression levels of CDK9 in endometrial cancer cell lines (AN3CA, ARK-2, HEC-1A, HEC-1B, lshikawa, RL95-2 and SPAC1S) as determined by western blotting. (B) Relative expression of CDK9 and α-tubulin in the endometrial cancer cell lines. CDK9, cyclin-dependent kinase 9.
Effect of CDK9 siRNA on the proliferation of endometrial cancer cells
We transfected AN3CA and SPAC1S cell lines with CDK9 siRNA to knock down CDK9 expression to confirm the function of CDK9 in the proliferation and progression of endometrial cancer cells. The cell lines were transfected with increasing concentrations of CDK9 siRNA (10–40 nM) for 5 days, and we discovered that cell viability of both cell lines was significantly and dose-dependently inhibited. This was absent in the non-specific siRNA transfected cells (40 nM NC siRNA) (Fig. 4A and B). Western blot analysis confirmed that non-specific siRNA (NC siRNA) had no effect on CDK9 expression, while CDK9 siRNA significantly reduced CDK9 expression during a 2-day observation period. Knockdown of CDK9 reduced the level of anti-apolipoprotein myeloid cell leukemia-1 (Mcl-1) and increased the level of the proapoptotic protein BCL2 associated X, apoptosis regulator (Bax) (Fig. 4C and D). These data illustrate the key role of CDK9 in the growth and proliferation of endometrial cancer cells.
Figure 4.

CDK9 knockdown by siRNA transfection suppresses endometrial cancer cell proliferation. (A and B) MTT assay revealed significant dose-dependent inhibition of cell proliferation after CDK9 siRNA treatment. **P<0.01 compared with the cell only control group. (C and D) Expression levels of CDK9 and related signaling pathway proteins involved in transcription and apoptosis after transfection of CDK9 siRNA and nonspecific siRNA (NC siRNA) in AN3CA and SPAC1S cell lines by western blot analysis. CDK9, cyclin-dependent kinase 9; Mcl-1, myeloid cell leukemia-1; Bax, proapoptotic protein BCL2 associated X, apoptosis regulator.
Effect of CDK9 inhibitor on endometrial cancer cell proliferation
Subsequently, we examined the effects of a novel CDK9-specific inhibitor LDC067 on the proliferation of endometrial cancer cells. Similar to CDK9 siRNA transfection, CDK9 selective inhibitor LDC067 decreased AN3CA and SPAC1S cell viability in a dose-dependent manner during a specified 5-day observation period (Fig. 5A and B). To further verify the function of LDC067 on the transcriptional regulation of endometrial cancer cells, we investigated the expression of apoptosis-related proteins. AN3CA and SPAC1S cell lines were incubated with 1.25, 2.5, 5.0 and 10 µM LDC067 for 48 h. The results showed that LDC067 could increase the expression of pro-apoptotic proteins cleaved poly(ADP-ribose) polymerase (PARP) and Bax, while inhibiting the expression of anti-apoptotic protein Mcl-1 in a concentration-dependent manner. Importantly, LDC067 only inhibited the activity of CDK9, and there was no significant difference in protein expression (Fig. 5C and D).
Figure 5.

CDK9 inhibitor reduces endometrial cancer cell proliferation by suppressing transcription elongation and inducing apoptosis in endometrial cancer cells. (A and B) Relative cell viability of AN3CA and SPAC1S cells after exposure to different concentrations of the CDK9 inhibitor LDC067 for 5 days. **P<0.01 compared with the lowest concentration group (1×10−3 µM). (C and D) Expression levels of CDK9 and related signaling pathway proteins involved in transcription and apoptosis after treatment with LDC067 in cells by western blot analysis. CDK9, cyclin-dependent kinase 9; Mcl-1, myeloid cell leukemia-1; Bax, proapoptotic protein BCL2 associated X, apoptosis regulator; PARP, poly(ADP-ribose) polymerase.
Effect of CDK9 inhibitor on endometrial cancer cell colony formation
Colony formation assay is a commonly used method to assess cell viability and proliferation capacity. Endometrial cancer AN3CA and SPAC1S cells were exposed to 2.5, 5.0 and 10.0 µM of LDC067 for 14 days. Colony formation assay showed that compared with the untreated cells, both the numbers and sizes of colonies formed in the LDC067 treatment group were significantly decreased in a dose-dependent manner (P<0.01). These results suggest that LDC067 can inhibit the ability of endometrial cancer to form colonies in a concentration-dependent manner (Fig. 6).
Figure 6.

Inhibition of CDK9 suppresses endometrial cancer cell colony formation. (A) Representative images of endometrial cancer cell colony formation after incubation with different concentrations of LDC067 (0, 2.5, 5.0, and 10 µM) for 14 days. (B and C) Quantification of clonogenicity formation of AN3CA (B) and SPAC1S (C) cells after LDC067 treatment. **P<0.01 compared with the Cell only group. CDK9, cyclin-dependent kinase 9.
Effect of CDK9 inhibitor on the migration ability of endometrial cancer cells
Tumor cell migration plays a significant role in cancer metastasis and recurrence. We analyzed the effect of LDC067 on inhibiting the migration activity of endometrial cancer cells through wound-healing assay. As shown in Fig. 7, after 24 and 48 h, the addition of 5 µM LDC067 brought about a time-dependent decrease in the migration capacity of the two cell lines compared to the untreated cells (P<0.05). In summary, the results indicate that LDC067 inhibits the migration capacity of endometrial cancer cells in a time-dependent manner.
Figure 7.

Inhibition of CDK9 reduces endometrial cancer cell migration. (A and B) Representative images of AN3CA and SPAC1S cell migration after CDK9 inhibitor LDC067 treatment for 0, 24, and 48 h. (C and D) Quantification of cell migration distance of AN3CA and SPAC1S cells after LDC067 treatment. **P<0.01 compared with the Cell only group. CDK9, cyclin-dependent kinase 9.
Discussion
Surgery is the main treatment for endometrial cancer. Most patients in the early stage can be cured by surgery, but treatment is limited for patients with metastasis and recurrence (23). It is widely recognized that laparoscopy is superior to laparotomy in most patients with endometrial cancer. Laparoscopy can reduce invasiveness, but still maintains safety and surgical effectiveness, optimizes surgical outcomes, and improves the cosmetic effect. The endoscopic single-site (LESS) surgery, 3 mm laparoscopy (MiniLPS) and percutaneous system (PSS) include the principal innovations in ultra-minimally invasive surgery introduced during the last few years. Even when reducing the port number or instrument size to reduce invasiveness, these improvements still maintain the same surgical technique, efficacy and safety of standard laparoscopy (LPS) (24–27). Although there exists continuous research concerning the tumorigenesis and development of endometrial cancer, it still remains unclear which signaling pathways or related molecules promote the occurrence and development of endometrial cancer. Many studies confirm that overexpression of cyclin-dependent kinase 9 (CDK9) is directly related to tumor development (13–19). A recent study found that CDK9 inhibition dephosphorylates the SWI/SNF protein BRG1 (ATP-dependent helicase) and reactivates epigenetic silenced genes in cancer, thereby restoring tumor-suppressor gene expression (28). In the present study, we found that CDK9 was overexpressed in metastatic and recurrent tissue samples from patients with endometrial cancer, and survival analysis showed that CDK9 can be used as a prognostic indicator for endometrial cancer patients. This finding is consistent with that of Wang et al who revealed that CDK9 expression is elevated in human ovarian cancer cell lines and is also increased in metastatic and recurrent ovarian tumor tissue compared with patient-matched primary ovarian tumor tissue. In addition, increased CDK9 was significantly associated with poor patient prognosis (29). This also is in accordance with earlier observations, which showed that during the progression of cervical cancer from pre-invasive lesions to squamous cell carcinoma, the intracellular concentration of CDK9 increases (30). Our results provide evidence that CDK9 has the opportunity to become a potential prognostic biomarker for endometrial cancer.
Research has shown that the CDK9 signaling system may have important significance in the development of malignant tumors and/or maintaining the cell phenotype (31). Inhibition of CDK9 kinase activity significantly affects the gene transcription regulation, which could be used as a strategy for cancer treatment interventions (29). The antitumor effect of CDK9 inhibition has been attributed to MYC proto-oncogene, BHLH transcription factor (MYC) and/or Mcl-1) inhibition (28). CDK9 is involved in many types of cancer by recruiting p-TEFb to transcribe the downstream proto-oncogene MYC involved in cell growth and cell cycle progression (32). In addition, over-stimulation of CDK9 increases the inhibitory effect of Mcl-1 on apoptosis, which will lead to cell survival and subsequent cancer progression (33).
It has also been reported that in ovarian cancer, leukemia, breast cancer, prostate cancer and liver cancer, reducing the expression of CDK9 by siRNA can inhibit cell proliferation and induce apoptosis (28,34–37). We used liposome-mediated transfection of synthetic CDK9 siRNA to silence or reduce CDK9 gene expression. The results demonstrated that CDK9 siRNA transfection significantly reduced the expression of CDK9 in a concentration-dependent manner. CDK9 siRNA can inhibit the phosphorylation of RNAPII, resulting in the overall decrease in mRNA levels, thereby reducing the level of anti-apoptotic protein Mcl-1 and increasing the level of pro-apoptotic protein Bax. With the increase in CDK9 siRNA transfection concentration, the proliferation of endometrial cancer cells was significantly inhibited.
CDK4/6 is part of the core cell cycle mechanism that drives cell proliferation. In many types of human tumors, CDK4 and CDK6 are abnormally activated through different mechanisms. The chemical inhibitors of CDK4/6, palbociclib, ribociclib and abemaciclib, have been used in multiple clinical trials worldwide to treat more than 30 different types of tumors, including endometrial cancer (38). A recent study demonstrated that increased expression of phosphorylated NPM/B23 (at Thr199) in cancer cells promotes its interaction with CDK6 and plays a key role in endometrial tumorigenesis (39). By inhibiting CDK4/6 kinase, palbociclib can inhibit the phosphorylation of NPM/B23 (Thr199) and promote the expression of estrogen receptor (ER)α, ultimately making hormone-refractory endometrial cancer sensitive to endocrine therapy (39). Yet, clinical trials of treatment with CDK9 inhibitors remain unsuccessful and involve many adverse effects (40). LDC067 is a novel CDK9-specific inhibitor that has been used in different types of cancer (20). In the present study, LDC067 reduced the phosphorylation of RNAPII, induced apoptosis in endometrial cancer cells, and inhibited cell proliferation activity in a dose-dependent manner. These results are consistent with those of Albert et al who found LDC067 targeted the ser2 of the RNAPII CTD, prevented phosphorylation and induced apoptosis (20). Unlike CDK9 siRNA, LDC067 only inhibits the activity of CDK9, while the protein expression of CDK9 is not significantly reduced. It was reported that gene expression profiling of tumor cells treated with CDK9 inhibitor LDC067 showed a selective reduction of short-lived mRNAs, including the MYC and Mcl-1 genes that regulate proliferation and apoptosis (20).
In the clone formation and wound healing assays of two endometrial cancer cell lines, the average number and size of colonies formed after LDC067 treatment were significantly inhibited, and the cell migration ability was also restrained in a time-dependent manner. In a recent study, CDK9 knockout in osteosarcoma cell lines and ovarian cancer cell lines triggered an effective antitumor response (29,41). These results are consistent with what we have observed. Collectively, these results support CDK9 as a potential factor for the progression of endometrial cancer.
In conclusion, our research demonstrated that high expression of CDK9 is closely connected with the poor prognosis of human endometrial cancer. In addition, inhibition of CDK9 significantly decreased endometrial cancer cell proliferation by suppressing the phosphorylation of RNAPII. Our research indicates that CDK9 may be a potential marker for the prognosis of endometrial cancer and may become a new direction for tumor-targeted therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science Foundation of China (nos. 81801425, 81671437, 81771558 and 81702582) and the Natural Science Foundation of Hunan Province in China (2018JJ3739).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
SH wrote the manuscript and substantially contributed to the design of the study and the interpretation of the data. XF performed the literature search for this article. XX and TH revised and corrected the manuscript critically for important intellectual content. XX and TH assisted in analyzing the data for the study and also contributed to the conception of the study. TZ conceived and designed the study. All authors read and approved the final manuscript and agree to be accountable for all aspects of the research in ensuring that the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Ethics approval and consent to participate
The study was approved by the Ethics Committee of The Second Xiangya Hospital, Central South University (Changsha, China) (IRB protocol number: Study 181), and written informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing interests.
References
- 1.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69:7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- 2.Wong YF, Cheung TH, Lo KW, Yim SF, Siu NS, Chan SC, Ho TW, Wong KW, Yu MY, Wang VW, et al. Identification of molecular markers and signaling pathway in endometrial cancer in Hong Kong Chinese women by genome-wide gene expression profiling. Oncogene. 2007;26:1971–1982. doi: 10.1038/sj.onc.1209986. [DOI] [PubMed] [Google Scholar]
- 3.Zhang Y, Liu Z, Yu X, Zhang X, Lü S, Chen X, Lü B. The association between metabolic abnormality and endometrial cancer: A large case-control study in China. Gynecol Oncol. 2010;117:41–46. doi: 10.1016/j.ygyno.2009.12.029. [DOI] [PubMed] [Google Scholar]
- 4.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68:7–30. doi: 10.3322/caac.21442. [DOI] [PubMed] [Google Scholar]
- 5.Boren T, Xiong Y, Hakam A, Wenham R, Apte S, Wei Z, Kamath S, Chen DT, Dressman H, Lancaster JM. MicroRNAs and their target messenger RNAs associated with endometrial carcinogenesis. Gynecol Oncol. 2008;110:206–215. doi: 10.1016/j.ygyno.2008.03.023. [DOI] [PubMed] [Google Scholar]
- 6.Malumbres M, Barbacid M. Mammalian cyclin-dependent kinases. Trends Biochem Sci. 2005;30:630–641. doi: 10.1016/j.tibs.2005.09.005. [DOI] [PubMed] [Google Scholar]
- 7.Malumbres M. Cyclin-dependent kinases. Genome Biol. 2014;15:122. doi: 10.1186/gb4184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shapiro GI. Cyclin-dependent kinase pathways as targets for cancer treatment. J Clin Oncol. 2006;24:1770–1783. doi: 10.1200/JCO.2005.03.7689. [DOI] [PubMed] [Google Scholar]
- 9.Wang S, Fischer PM. Cyclin-dependent kinase 9: A key transcriptional regulator and potential drug target in oncology, virology and cardiology. Trends Pharmacol Sci. 2008;29:302–313. doi: 10.1016/j.tips.2008.03.003. [DOI] [PubMed] [Google Scholar]
- 10.Franco LC, Morales F, Boffo S, Giordano A. CDK9: A key player in cancer and other diseases. J Cell Biochem. 2018;119:1273–1284. doi: 10.1002/jcb.26293. [DOI] [PubMed] [Google Scholar]
- 11.Kohoutek J. P-TEFb- the final frontier. Cell Div. 2009;4:19. doi: 10.1186/1747-1028-4-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bagella L, MacLachlan TK, Buono RJ, Pisano MM, Giordano A, De Luca A. Cloning of murine CDK9/PITALRE and its tissue-specific expression in development. J Cell Physiol. 1998;177:206–213. doi: 10.1002/(SICI)1097-4652(199811)177:2<206::AID-JCP2>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- 13.Brägelmann J, Dammert MA, Dietlein F, Heuckmann JM, Choidas A, Böhm S, Richters A, Basu D, Tischler V, Lorenz C, et al. Systematic kinase inhibitor profiling identifies CDK9 as a synthetic lethal target in NUT midline carcinoma. Cell Rep. 2017;20:2833–2845. doi: 10.1016/j.celrep.2017.08.082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rahaman MH, Kumarasiri M, Mekonnen LB, Yu M, Diab S, Albrecht H, Milne RW, Wang S. Targeting CDK9: A promising therapeutic opportunity in prostate cancer. Endocr Relat Cancer. 2016;23:T211–T226. doi: 10.1530/ERC-16-0299. [DOI] [PubMed] [Google Scholar]
- 15.Mitra P, Yang RM, Sutton J, Ramsay RG, Gonda TJ. CDK9 inhibitors selectively target estrogen receptor-positive breast cancer cells through combined inhibition of MYB and MCL-1 expression. Oncotarget. 2016;7:9069–9083. doi: 10.18632/oncotarget.6997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Whittaker SR, Barlow C, Martin MP, Mancusi C, Wagner S, Self A, Barrie E, Te Poele R, Sharp S, Brown N, et al. Molecular profiling and combinatorial activity of CCT068127: A potent CDK2 and CDK9 inhibitor. Mol Oncol. 2018;12:287–304. doi: 10.1002/1878-0261.12148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Baker A, Gregory GP, Verbrugge I, Kats L, Hilton JJ, Vidacs E, Lee EM, Lock RB, Zuber J, Shortt J, et al. The CDK9 inhibitor dinaciclib exerts potent apoptotic and antitumor effects in preclinical models of MLL-rearranged acute myeloid leukemia. Cancer Res. 2016;76:1158–1169. doi: 10.1158/0008-5472.CAN-15-1070. [DOI] [PubMed] [Google Scholar]
- 18.Su YT, Chen R, Wang H, Song H, Zhang Q, Chen LY, Lappin H, Vasconcelos G, Lita A, Maric D, et al. Novel targeting of transcription and metabolism in glioblastoma. Clin Cancer Res. 2018;24:1124–1137. doi: 10.1158/1078-0432.CCR-17-2032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ajiro M, Sakai H, Onogi H, Yamamoto M, Sumi E, Sawada T, Nomura T, Kabashima K, Hosoya T, Hagiwara M. CDK9 inhibitor FIT-039 suppresses viral oncogenes E6 and E7 and has a therapeutic effect on HPV-induced neoplasia. Clin Cancer Res. 2018;24:4518–4528. doi: 10.1158/1078-0432.CCR-17-3119. [DOI] [PubMed] [Google Scholar]
- 20.Albert TK, Rigault C, Eickhoff J, Baumgart K, Antrecht C, Klebl B, Mittler G, Meisterernst M. Characterization of molecular and cellular functions of the cyclin-dependent kinase CDK9 using a novel specific inhibitor. Br J Pharmacol. 2014;171:55–68. doi: 10.1111/bph.12408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Baranyi U, Winter B, Gugerell A, Hegedus B, Brostjan C, Laufer G, Messner B. Primary human fibroblasts in culture switch to a myofibroblast-like phenotype independently of TGF beta. Cells. 2019;8:721. doi: 10.3390/cells8070721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lay V, Yap J, Sonderegger S, Dimitriadis E. Interleukin 11 regulates endometrial cancer cell adhesion and migration via STAT3. Int J Oncol. 2012;41:759–764. doi: 10.3892/ijo.2012.1486. [DOI] [PubMed] [Google Scholar]
- 23.De U, Son JY, Sachan R, Park YJ, Kang D, Yoon K, Lee BM, Kim IS, Moon HR, Kim HS. A new synthetic histone deacetylase inhibitor, MHY2256, induces apoptosis and autophagy cell death in endometrial cancer cells via p53 acetylation. Int J Mol Sci. 2018;19:2743. doi: 10.3390/ijms19092743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Espedal H, Fonnes T, Fasmer KE, Krakstad C, Haldorsen IS. Imaging of preclinical endometrial cancer models for monitoring tumor progression and response to targeted therapy. Cancers (Basel) 2019;11:1885. doi: 10.3390/cancers11121885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gueli Alletti S, Rossitto C, Cianci S, Restaino S, Costantini B, Fanfani F, Fagotti A, Cosentino F, Scambia G. Telelap ALF-X vs standard laparoscopy for the treatment of early-stage endometrial cancer: A single-institution retrospective cohort study. J Minim Invasive Gynecol. 2016;23:378–383. doi: 10.1016/j.jmig.2015.11.006. [DOI] [PubMed] [Google Scholar]
- 26.Scaletta G, Dinoi G, Capozzi V, Cianci S, Pelligra S, Ergasti R, Fagotti A, Scambia G, Fanfani F. Comparison of minimally invasive surgery with laparotomic approach in the treatment of high risk endometrial cancer: A systematic review. Eur J Surg Oncol. 2020;46:782–788. doi: 10.1016/j.ejso.2019.11.519. [DOI] [PubMed] [Google Scholar]
- 27.Rossitto C, Cianci S, Gueli Alletti S, Perrone E, Pizzacalla S, Scambia G. Laparoscopic, minilaparoscopic, single-port and percutaneous hysterectomy: Comparison of perioperative outcomes of minimally invasive approaches in gynecologic surgery. Eur J Obstet Gynecol Reprod Biol. 2017;216:125–129. doi: 10.1016/j.ejogrb.2017.07.026. [DOI] [PubMed] [Google Scholar]
- 28.Zhang H, Pandey S, Travers M, Sun H, Morton G, Madzo J, Chung W, Khowsathit J, Perez-Leal O, Barrero CA, et al. Targeting CDK9 reactivates epigenetically silenced genes in cancer. Cell. 2018;175:1244–1258.e26. doi: 10.1016/j.cell.2018.09.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang J, Dean DC, Hornicek FJ, Shi H, Duan Z. Cyclin- dependent kinase 9 (CDK9) is a novel prognostic marker and therapeutic target in ovarian cancer. FASEB J. 2019;33:5990–6000. doi: 10.1096/fj.201801789RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ramdass B, Maliekal TT, Lakshmi S, Rehman M, Rema P, Nair P, Mukherjee G, Reddy BK, Krishna S, Radhakrishna Pillai M. Coexpression of Notch1 and NF-kappaB signaling pathway components in human cervical cancer progression. Gynecol Oncol. 2007;104:352–361. doi: 10.1016/j.ygyno.2006.08.054. [DOI] [PubMed] [Google Scholar]
- 31.Romano G. Deregulations in the cyclin-dependent kinase-9- related pathway in cancer: Implications for drug discovery and development. ISRN Oncol. 2013;2013:305371. doi: 10.1155/2013/305371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lu H, Xue Y, Yu GK, Arias C, Lin J, Fong S, Faure M, Weisburd B, Ji X, Mercier A, et al. Compensatory induction of MYC expression by sustained CDK9 inhibition via a BRD4-dependent mechanism. Elife. 2015;4:e06535. doi: 10.7554/eLife.09993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yin T, Lallena MJ, Kreklau EL, Fales KR, Carballares S, Torrres R, Wishart GN, Ajamie RT, Cronier DM, Iversen PW, et al. A novel CDK9 inhibitor shows potent antitumor efficacy in preclinical hematologic tumor models. Mol Cancer Ther. 2014;13:1442–1456. doi: 10.1158/1535-7163.MCT-13-0849. [DOI] [PubMed] [Google Scholar]
- 34.Caracciolo V, Laurenti G, Romano G, Carnevale V, Cimini AM, Crozier-Fitzgerald C, Gentile Warschauer E, Russo G, Giordano A. Flavopiridol induces phosphorylation of AKT in a human glioblastoma cell line, in contrast to siRNA-mediated silencing of Cdk9: Implications for drug design and development. Cell Cycle. 2012;11:1202–1216. doi: 10.4161/cc.11.6.19663. [DOI] [PubMed] [Google Scholar]
- 35.Walsby E, Pratt G, Shao H, Abbas AY, Fischer PM, Bradshaw TD, Brennan P, Fegan C, Wang S, Pepper C. A novel Cdk9 inhibitor preferentially targets tumor cells and synergizes with fludarabine. Oncotarget. 2014;5:375–385. doi: 10.18632/oncotarget.1568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rajput S, Khera N, Guo Z, Hoog J, Li S, Ma CX. Inhibition of cyclin dependent kinase 9 by dinaciclib suppresses cyclin B1 expression and tumor growth in triple negative breast cancer. Oncotarget. 2016;7:56864–56875. doi: 10.18632/oncotarget.10870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Huang CH, Lujambio A, Zuber J, Tschaharganeh DF, Doran MG, Evans MJ, Kitzing T, Zhu N, de Stanchina E, Sawyers CL, et al. CDK9-mediated transcription elongation is required for MYC addiction in hepatocellular carcinoma. Genes Dev. 2014;28:1800–1814. doi: 10.1101/gad.244368.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fassl A, Sicinski P. Chemotherapy and CDK4/6 inhibition in cancer treatment: Timing is everything. Cancer Cell. 2020;37:265–267. doi: 10.1016/j.ccell.2020.02.007. [DOI] [PubMed] [Google Scholar]
- 39.Lin CY, Lee LY, Wang TH, Hsu CL, Tsai CL, Chao A, Lai CH. Palbociclib promotes dephosphorylation of NPM/B23 at threonine 199 and inhibits endometrial cancer cell growth. Cancers (Basel) 2019;11:1025. doi: 10.3390/cancers11071025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Morales F, Giordano A. Overview of CDK9 as a target in cancer research. Cell Cycle. 2016;15:519–527. doi: 10.1080/15384101.2016.1138186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ma H, Seebacher NA, Hornicek FJ, Duan Z. Cyclin-dependent kinase 9 (CDK9) is a novel prognostic marker and therapeutic target in osteosarcoma. EBioMedicine. 2019;39:182–193. doi: 10.1016/j.ebiom.2018.12.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.