

Abstract
Bartonella henselae is a fastidious intraerythrocytic, gram-negative bacteria that causes cat scratch disease in humans. Ixodes ricinus has been confirmed to be a competent vector of B. henselae, and some indirect evidences from clinical cases and epidemiological studies also suggested that some other tick species, including Rhipicephalus sanguineus, may transmit the bacteria. B. henselae has been detected in R. sanguineus but no experimental investigations have been performed to evaluate the vector competency of this tick species regarding B. henselae transmission. To this end, this work aimed to assess the transstadial transmission of B. henselae between larvae and nymphs of R. sanguineus as well as transmission by nymphs infected at the larval stage. Four hundred B. henselae negative larvae were fed with B. henselae-infected blood by using an artificial membrane feeding system. After five days of feeding, B. henselae was detected by PCR in 57.1% (8/14) of engorged larval pools, 66.7% (4/6) of semi-engorged larval pools, and 66.7% (2/3) of larval feces pools. After molting, B. henselae DNA was also detected in 10% (1/10) of nymph pools, but not in tick feces. After a pre-fed step of nymphs infected at the larval stage on non-infected blood meal, B. henselae was detected by PCR in blood sample from the feeder, but no Bartonella colonies could be obtained from culture. These findings showed that B. henselae could be transstadial transmitted from R. sanguineus larvae to nymphs, and also suggest that these nymphs may retransmitted the bacteria through the saliva during their blood meal. This is the first study that validated the artificial membrane feeding system for maintaining R. sanguineus tick colony. It shows the possibility of transstadial transmission of B. henselae from R. sanguineus larvae to nymphs.
Author summary
B. henselae is gram-negative bacteria that infects red blood cells of humans and companion animals and causes cat scratch disease in humans. Ticks were considered to be potential vectors of B. henselae for a long time until it was finally experimentally demonstrated for Ixodes ricinus. Since then, no evidence on B. henselae transmission by other tick species was reported. This study was performed 1) to validate the use of artificial membrane system to feed and infect R. sanguineus ticks and 2) to determine the possibility of B. henselae transmission by R. sanguineus, a world-widely distributed ticks. Our results show that the artificial membrane feeding system can be used to maintain R. sanguineus colony in the laboratory, and that B. henselae can be acquired by R. sanguineus during a blood meal on artificial membrane feeding system and can be transmitted from larvae to nymphs that were able to inject bacterial DNA to blood during a new blood meal. However, further investigations are still needed to confirm the viability of bacteria transmitted to blood by nymphs infected at the larval stage in order to validate B. henselae transmission by R. sanguineus.
Introduction
Bartonella henselae is a fastidious gram-negative bacteria which infects companion animals and is responsible for cat scratch disease (CSD) in humans [1]. Worldwide, B. henselae infection is estimated to be between 5% and 86% in cats [2–4]. B. henselae is normally transmitted from cat to cat by fleas (Ctenocephalides felis felis) and usually transmitted from cat to human by cat scratch due to B. henselae contamination from flea feces [5–9]. Most human CSD cases are self-limited from asymptomatic to skin inflammation, fever and lymphadenopathy [10]. However, B. henselae is becoming increasingly associated with a number of other syndromes including conjunctivitis, encephalopathy and endocarditis [11–13]. In addition, bacillary angiomatosis and peliosis hepatitis are unusual vascular proliferative lesions observed in immunocompromised patients with B. henselae infection [14, 15].
Despite cat flea has been known to be the competent vector of B. henselae, ticks have also been considered as potential vectors for Bartonella transmission for a long time [16–31]. B. henselae DNA has been detected in several species of ticks, including Ixodes pacificus, I. persulcatus, I. ricinus, and R. sanguineus [18, 26–28]. Co-infections of B. henselae with other pathogens known to be transmitted by ticks, such as Anaplasma spp., Borrelia spp. and Rickettsia spp., have been reported in both humans and animals by molecular evidence, suggesting a possible co-transmission of pathogenic agents after a tick bite [19, 20, 22, 25, 30, 32–34]. Moreover, B. henselae has been isolated by blood culture or detected by PCR from human patients with history of tick bites, which emphasized the hypothesis that ticks may serve as vectors for Bartonella spp. [16, 17, 20, 23–25, 29, 31]. In 2008, Cotté and co-workers have demonstrated that B. henselae can survive in I. ricinus during their molting from larvae to nymphs and from nymphs to adults, and can be transmitted to blood through tick saliva in an artificial membrane feeding system [35]. They also showed that the bacteria located into the salivary glands of I. ricinus adults infected at the nymphal stage was alive and infectious as when the corresponding tick salivary glands were injected to cats which then developed a bacteremia. Later, Reis and co-workers also reported that B. birtlesii could be transstadial transmitted from I. ricinus larvae to nymphs and from nymphs to adults [36]. They also demonstrated that when infected on infected mice as larvae, I. ricinus nymphs can retransmit the bacteria to naïve mice, and when infected at the nymphal stage, adults can retransmit the bacteria to blood through the membrane feeding system. All these results proved that I. ricinus can act as a vector for both B. henselae and B. birtlesii [35, 36].
The brown dog tick, R. sanguineus, has the worldwide geographic distribution. This tick plays roles as a vector of several pathogens causing clinical illness, including Babesia vogeli and Ehrlichia canis in dogs, and Rickettsia conorii and R. rickettsii in humans [37]. R. sanguineus has been suspected to be a potential vector of Bartonella spp. since 1992, when two patients developed fever with a relapsing course after tick bites, and B. henselae was isolated by culture from their blood [23]. Several molecular epidemiological surveys were then conducted, and the B. henselae DNA positive rate in R. sanguineus ticks has been reported as being 0.09% (1/209) in Italy, 3.2% (2/62) in California, USA, and 5.3% (15/281) in the central part of Taiwan [28, 30, 38]. In addition, B. vinsonii subsp. berkhoffii DNA has been detected in R. sanguineus adult feces suggesting that tick feces could be a potential source of B. vinsonii subsp. berkhoffii infection [39]. However, no validation of the vector competence of R. sanguineus for Bartonella spp. transmission has been performed until now.
Because of evidence of R. sanguineus harboring and potentially transmitting Bartonella spp. and limited information on Bartonella transmission by ticks, we performed the present experimental study to investigate the possibility of B. henselae transmission by R. sanguineus using an artificial membrane feeding system. This feeding technique can mimic the natural conditions of tick infection via the digestive tract in controlled condition as validated for I. ricinus with both tick-borne bacteria and parasites and, until now, was never applied to R. sanguineus feeding or infection studies [35, 36, 40–42].
Methods
Tick collection and population maintenance
Engorged R. sanguineus females were collected from dogs in veterinary hospitals in Taiwan and morphological identification was performed by using taxonomic keys [43]. Each engorged female was placed in a container inside a chamber with 80–90% of relative humidity, at room temperature and with a photoperiod of 16:8h (L: D) cycles until finishing her oviposition period [44]. In each larval batch, 10% of larval ticks was randomly selected and tested for B. henselae DNA presence. Only B. henselae negative offspring were then used in all experiments. After feeding and molting from the previous life stage, ticks were starved in the previously mentioned conditions for one-month until next feeding.
B. henselae isolates
B. henselae, isolated from strayed cats in eastern part of Taiwan, was cultured on chocolate agar plates (Taiwan Prepared Media Co., LTD), at 35°C, and in an atmosphere of 5% CO2. Number of Colony-Forming Units (CFU) was evaluated to estimate the amount of viable B. henselae. B. henselae colonies were harvested by using sterilized polypropylene loop (SPL Lifesciences Co., LTD), suspended in sterile 1x of Phosphate-Buffered Saline (1x PBS), and used immediately for tick feeding [35]. DNA of B. henselae used as a positive control for nested-PCR was extracted from a bacterial colony using DNeasy Blood & Tissue kit (Qiagen).
Skin membrane preparation
Outbred ICR (Bltw:CD1) mice (BioLASCO Taiwan Co., LTD), around 10 weeks old, without any treatment from cooperating laboratories in National Pingtung University of Science and Technology, Taiwan were used to obtain skins. Mice skins with the dermal thickness around 300 μm were considered to use in artificial feeding of R. sanguineus larvae and nymphs (hypostome length: 50 μm and 120 μm, respectively) [45, 46]. The mice skins were then processed as previously described [41]. Briefly, dissected skins were first sterilized in 70% ethanol for 5 min, followed by rinsing in sterile distilled water for 5 min and then 1x PBS for 5 min. Those skins were finally aseptically treated in an antimicrobial solution of Gentamicin (10 mg/ml), Amphotericin B (0.25 μg/ml), Penicillin (50 U/ml) and Streptomycin (50 μl/ml) for 10 min. All dissected skins were then stored at -20°C not longer than 3 months until use.
Blood preparation for tick feeding
Goat blood, obtained from a goat farm in Pingtung, Taiwan and confirmed Bartonella DNA negative by PCR, was used in all following experiments (ethical permit IACUC number: NPUST-105-036). After collection, the blood was defibrinated and depleted of functional complement by heat treatment at 56°C for 30 min [35]. To prevent fungal and bacterial contamination during tick feeding, decomplemented blood was supplemented with 20 μl/ml of Fosfomycin, 0.25 μg/ml of Amphotericin B, and 10 KU/ml of Heparin [35].
Feeding larvae with B. henselae-infected blood
The artificial membrane feeding system was adapted from Bonnet et al [41]. This system consists of a glass feeder closed with a mice skin at the bottom, and placed on the top of the tick container. Placing the blood above the skin membrane supports a continuous gravitational pressure on the membrane. The glass feeders were connected to a 37°C water circulation system to mimic host body temperature, attract ticks and preserve Bartonella in the blood (Fig 1). This system was established under room temperature, 80–90% of relative humidity and photoperiod of 16:8h (L: D).
Fig 1. Diagram of an artificial membrane feeding system used to feed R. sanguineus larvae and nymphs.
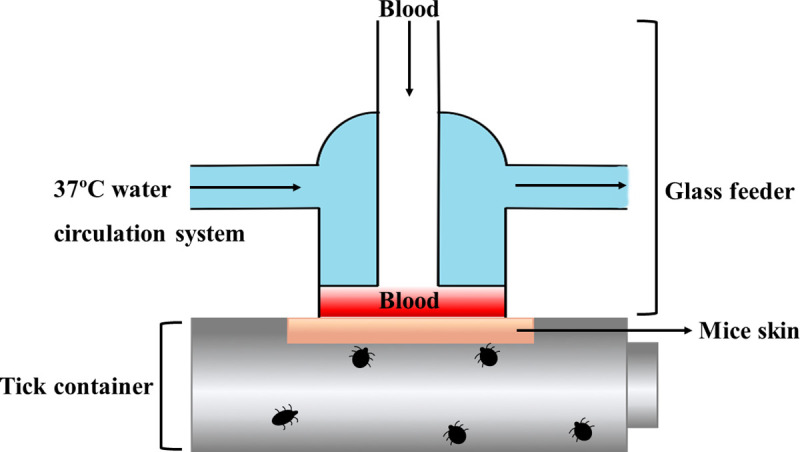
The artificial feeder consists of a glass feeder, animal skin, and tick container. The glass feeder is connected to a 37°C water circulation system to mimic host body temperature.
The general framework of the tick feeding experiment is shown in Fig 2. A total of 400 larvae were fed with B. henselae-infected blood for 5 days by using artificial membrane feeding system under laboratory conditions as described above. The larval feeding duration in the present study was adjusted from the reported time of feeding of R. sanguineus larvae on rabbits under laboratory conditions, i.e. 3 to 6 days [44]. The B. henselae-infected blood was prepared as follow: a total of 40 μl of B. henselae suspension at a concentration of 109 CFU/ml was added to 4 ml of decomplemented blood for feeding. Blood was changed twice a day at 12 h intervals, and the side of mice skin in contact with the blood was washed three times with RPMI 1640 (CORNING) [41]. As a control group, 200 larvae were fed on non-infected blood under the same condition and the blood was changed once a day (Fig 2). At the end of feeding experiment, larvae that spontaneously detached from mice skin were considered as engorged larvae, and the larvae that were still attached on the mice skin and then manually detached were considered as semi-engorged larvae. The attachment rate of R. sanguineus larvae on mice skin was calculated at day 5 after the beginning of the experiment.
Fig 2. Duration of tick feeding experiment of R. sanguineus larvae into nymphs.

Larvae were fed for 5 days and were allowed to complete their molting period, which ranged from 15–45 days. After they become nymphs, they were starved around one month and then fed for 4 days. B. henselae were detected by PCR assay from larval samples (at the end of larval feeding), nymphal samples (after molting completed) and blood samples (daily collected during the nymphal feeding). After 4 days of nymphal feeding, blood from each feeder was collected for Bartonella isolation.
Feeding nymphs with non-infected blood
A total of 60 nymphs that molted from experimentally Bartonella-infected larvae were fed in a unique feeder with non-infected blood for 4 days according to the delay of bacteria detection in blood samples reported for I. ricinus [35, 36]. As control, 40 nymphs molted from larvae engorged on non-infected blood were also fed in another feeder with non-infected blood. A total of 100 μl of blood was collected daily from each feeder and used to detect Bartonella DNA by PCR. After 4 days of feeding, 10 μl of blood was spread on chocolate blood agar plates to detect Bartonella colonies [35]. At day 4, nymphs were manually detached on the mice skin and considered as semi-engorged nymphs for calculating the nymph attachment rate (Fig 2).
DNA extraction from tick and blood samples
DNA was extracted using DNeasy Blood & Tissue kit (Qiagen) from 6 kinds of samples, including 100 μl of blood samples, pools of 5 engorged larvae, pools of 10 semi-engorged larvae, pools of larval feces, pools of 3 unfed nymphs (molted from engorged larvae) and pools of nymphal feces from both experimental and control groups. Tick feces were collected from the tick container during the feeding process by sterilized polypropylene loop and needle (SPL Lifesciences Co., LTD), and suspended in 100 μl of sterile 1x PBS. The final elution was 50 μl for all samples. All DNA samples were stored at -20°C until PCR processing for B. henselae DNA detection.
B. henselae PCR amplification
B. henselae DNA was detected by using nested-PCR [47]. The primers forward 5’-CTTCGTTTCTCTTTCTTCA-3’ and reverse 5’-CTTCTCTTCACAATTTCAAT-3’ used for outer reaction, amplified fragments of the 16S-23S rRNA internal transcribed spacer (ITS) region of Bartonella spp. [48]. The primers forward 5’-TTGCTTCTAAAAAGCTTATCAA-3’ and reverse 5’-CAAAAGAGGGATTACAAAATC-3’ used for inner reaction were designed to be specific to B. henselae amplifying a 254-bp fragment [47]. PCR mixtures was set up as follows: 5 μl of DNA template, 1 μl of 10 μM of each primer, 5 μl of 10X Taq buffer (Genomics BioSci & Tech, Taiwan), 4 μl of 2.5 mM of dNTPs Mixture (Genomics BioSci & Tech, Taiwan), 1 μl of 2.5 U/μl of Taq DNA polymerase (Genomics BioSci & Tech, Taiwan), and adjusted to a final volume of 50 μl with distilled water. The PCR conditions for both outer and inner reactions were those described by Sato and co-workers [47].
Sequence and statistical analyses
The B. henselae suspected positive samples were sent for nucleotide sequencing (Genomics BioScience and Technology Co., Ltd., Taiwan). Sequence data was analyzed for genetic relationship in GenBank database by using NCBI nucleotide BLAST tool and validated sequences were aligned and analyzed by using MegAlign (DNASTAR, Inc., WI, USA). The attachment rates of R. sanguineus larvae and nymphs, and the engorgement rates of R. sanguineus larvae in control and experimental groups were calculated, and compared by using the Fisher’s exact test (p<0.05) (GraphPad Prism 8.4.2 software). B. henselae infection rates in tick pools were evaluated by using the bias-corrected maximum likelihood estimation (MLE) method with 95% confidence interval, analyzed per 100 ticks by the PooledInfRate statistical software version 4.0 [49].
Results
High attachment rates of R. sanguineus on artificial membrane feeding system
In the present study, some of R. sanguineus larvae started to attach on mice skin around 6 hours after tick placement, and some engorged larvae from both groups started to detach spontaneously on day 3 (Fig 3A and 3B). After 5 days of feeding on B. henselae-infected blood, a total of 43% (172/400) of larvae were completely engorged, and 41.5% (166/400) were semi-engorged (Table 1 and Fig 3C). For control group, 34% (68/200) and 47.5% (95/200) of larvae were fully engorged and semi-engorged, respectively. The engorgement rate of larvae from experimental group (43%) was significantly higher than in control group (34%) (p = 0.0344). Attachment rates of R. sanguineus larvae on mice skins in experimental and control groups were 84.5% and 81.5%, respectively, with no significant difference (p = 0.3529) (Table 1). Engorged larvae were maintained under rearing conditions until they completed their molting period, which ranged from 15–45 days. Regarding nymphs on day 4 of feeding, 56.7% (34/60) of nymphs from experimental group and 70% (28/40) of nymphs from control group were attached on mice skins and considered as semi-engorged nymphs (Table 1 and Fig 3D). Attachment rates were not significantly different between nymphs from experimental group and that from control group (p = 0.2908).
Fig 3. R. sanguineus larvae and nymphs engorged using goat blood infected or not with B. henselae.

(A) Larvae attached on the mice skin. (B) Engorged larvae detached from the mice skin after 3 days. (C) Engorged larva (left) and semi-engorged larva (right). (D) Semi-engorged nymph (left) and unfed nymph (right).
Table 1. Attachment rates of R. sanguineus larvae and nymphs fed on mice skin by artificial membrane feeding system.
| Ticks fed on mice skin | Larval stage | Nymphal stage | ||
|---|---|---|---|---|
| Experimental group (400) | Control group (200) | Experimental group (60) | Control group (40) | |
| No. of engorged ticks (%) | 172 (43%) * | 68 (34%) * | - | - |
| No. of semi-engorged ticks (%) | 166 (41.5%) | 95 (47.5%) | 34 (56.7%) | 28 (70%) |
| Others (%) | 62 (15.5%) | 37 (18.5%) | 26 (43.3%) | 12 (30%) |
| Attachment rate | 338 (84.5%) | 163 (81.5%) | 34 (56.7%) | 28 (70%) |
Data were analyzed statistically to compare results between ticks fed on B. henselae-infected blood (experimental group) and non-infected blood (control group) by Fisher’s exact test (* p<0.05).
Others: unfed, inactive, or died ticks.
B. henselae DNA detection in engorged R. sanguineus larvae and transstadial transmission to nymphs
Pools of engorged larvae (N = 5), pools of semi-engorged larvae (N = 10), and pools of larval feces were collected and tested for B. henselae DNA presence after 5 days of feeding on infected blood. PCR result showed that 57.1% (8/14) of engorged larval pools presented the expected B. henselae-specific 254-bp DNA fragment. 66.7% (4/6) of semi-engorged larval pools and 66.7% (2/3) of larval feces pools from the experimental group were also positive for B. henselae DNA. The MLE of B. henselae infection rate among pools of engorged and semi-engorged larvae were then estimated as 14.94% (95% CI, 7.30–27.59%) and 9.06% (95% CI, 3.15–23.79%), respectively. Samples from the control group were all negative for B. henselae detection (Table 2 and Fig 4).
Table 2. Detection of B. henselae DNA in R. sanguineus larvae engorged on infected blood and in nymphs after molting and maximum likelihood estimation (MLE) of tick infection.
| Pooled samples | No. of individuals | No. of positive pools/no. of pools tested (%) | Maximum likelihood estimation (MLE) | 95% CI |
|---|---|---|---|---|
| Experimental group | ||||
| Engorged larvae | 70 | 8/14 (57.1) | 14.94% | 7.30–27.59% |
| Semi-engorged larvae | 60 | 4/6 (66.7) | 9.06% | 3.15–23.79% |
| Larval feces | 2/3 (66.7) | - | - | |
| Unfed nymphs | 30 | 1/10 (10) | 3.33% | 0.20–15.30% |
| Nymphal feces | 0/3 (0) | - | - | |
| Control group | ||||
| Engorged larvae | 10 | 0/2 (0) | ||
| Semi-engorged larvae | 20 | 0/2 (0) | ||
| Larval feces | 0/2 (0) | |||
| Unfed nymphs | 12 | 0/4 (0) | ||
| Nymphal feces | 0/2 (0) |
95% CI: 95% confidence interval.
Fig 4. B. henselae DNA detection in R. sanguineus larvae and nymphs.

Detection of B. henselae DNA by nested-PCR in representative samples: M, DNA marker; EL1 and EL2, pooled engorged larvae (Experimental group); SEL, pooled semi-engorged larvae (Experimental group); LF, pooled larval feces (Experimental group); N, pooled unfed nymphs (Experimental group); NF, pooled nymphal feces (Experimental group); EL-, pooled engorged larvae (Control group); SEL-, pooled semi-engorged larvae (Control group); LF-, pooled larval feces (Control group); N-, pooled unfed nymphs (Control group); NF-, pooled nymphal feces (Control group); +, positive control (B. henselae DNA); -, negative control (distilled water).
After larvae molted to nymphs, some of the unfed nymphs and their feces were tested for B. henselae DNA detection by PCR. One (10%) pooled nymph sample was detected positive for B. henselae DNA, when pooled nymphal feces samples from the experimental group were all negative. The MLE of B. henselae infection rate of pooled nymphs was then evaluated as 3.33% (95% CI, 0.20–15.30%). B. henselae DNA was not detected in samples from the control group (Table 2 and Fig 4). All obtained sequences showed 100% identity to each other as well as with the partial sequence of 16S-23S ribosomal RNA intergenic spacer B. henselae (Accession number MN170540.1).
Bartonella DNA detection in blood during feeding of nymphs infected at the larval stage
Sixty nymphs from the experimental group infected as larvae and forty nymphs from the control group were fed separately on non-infected blood for 4 days. To determine whether B. henselae was transmitted by nymphs through blood sucking, blood samples were collected daily and tested for B. henselae DNA. The PCR result showed that B. henselae DNA was detected in blood 4 days after nymph attachment to the skin for those molted from larvae engorged on infected blood, when PCR were all negative for the control group (Fig 5). Unfortunately, no colonies were obtained after 14 days of culture of the blood sample taken on day 4, as for control.
Fig 5. Detection by nested-PCR of B. henselae DNA in non-infected blood after feeding of nymphs infected at the larval stage.
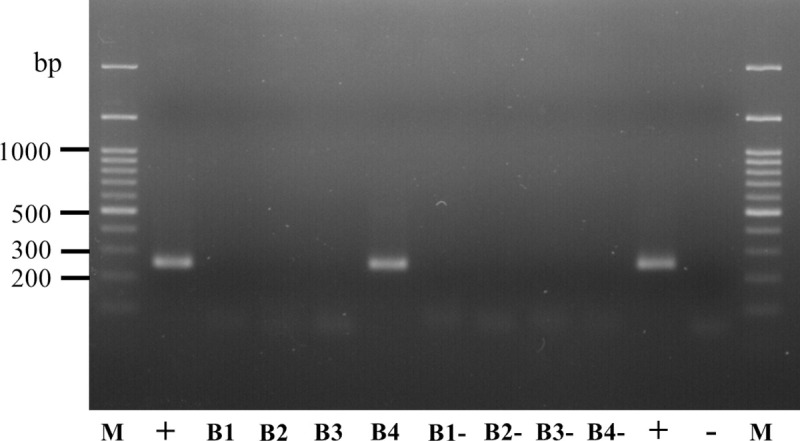
M, DNA marker; B1—B4, blood samples collected from day 1 to 4, respectively (Experimental group); B1- - B4-, blood samples collected from day 1 to 4, respectively (Control group); +, positive control (B. henselae DNA); -, negative control (distilled water).
Discussion
In the present study, an artificial membrane feeding system using mice skin and initially designed for I. ricinus was successfully set up for R. sanguineus [41]. Our results show high larval attachment rate of 81.5% in the control group and 84.5% in the experimental group, as well as 70% in the control group and 56.7% in the experimental group for nymphs. Under similar feeding conditions and treatments, the engorgement rate of R. sanguineus larvae (43%; 172/400) in experimental group shown in the present study were lower than those (81% and 84.5%) reported in previous studies for I. ricinus [35, 41]. For this last tick species, it has been demonstrated that blood origin (chicken or sheep) does not impact the proportion of engorged ticks, the weight of engorged ticks or the duration of feeding [50]. It is therefore conceivable that, despite we used different blood origin, the difference observed here is due to the tick species, including for example differences in appetite for mice skin, or into the length of tick mouthparts, which are 50 μm and 90 μm in the larvae of R. sanguineus and I. ricinus, respectively [45].
B. henselae DNA was detected in pooled engorged and semi-engorged larvae fed on B. henselae-infected blood, which supports the hypothesis that Rhipicephalus ticks could acquire Bartonella or at least Bartonella DNA during their feeding, as already suggested in some epidemiological studies [28, 30, 38]. In this study, the engorgement performance of larvae fed on B. henselae-infected blood (43%) was higher than on non-infected blood (34%). This finding has also been found for some other vector-borne pathogens, such as Plasmodium spp. and Trypanosoma spp., suggesting that the infectious status of the hosts can enhance feeding behavior of their vectors [51–54]. However, to the contrary, it has been reported that for I. ricinus, feeding on a B. henselae-infected blood meal through membrane feeding or on infected mice decreased the proportion and the weight of engorged ticks, but did not affect tick feeding duration [50]. The same trend is found here for the nymphs with a higher attachment rate for the control group, which might suggest that tick infection perturbs their feeding. However, experiments involving a higher number of ticks with sufficient time of feeding for their completely engorgements would be necessary to confirm this hypothesis. In addition, obtaining the weight of ticks by measuring the amount of ingested hemoglobin using spectrophotometry in future studies can provide a quantitative evaluation of the ticks’ fully engorged and semi-engorged status and the pathogen uptake by ticks [55, 56].
The presence of B. henselae DNA in nymphs molted from larvae engorged on infected blood suggests that B. henselae could be transstadially transmitted from larvae to nymphs during the molting process. This possibility is supported by our results showing that nymphs infected at the larval stage are able to inject the bacterial DNA into blood during their blood meal. Indeed, such a result suggests that DNA is injected through the saliva, and that the bacteria is then present into the salivary glands of the tick infected as the previous life stage, as demonstrated for both of B. henselae and B. birtlesii in I. ricinus ticks [35, 36]. It is therefore more than likely that it is viable bacteria, which escaped from the tick digestive tract and then invaded the salivary glands of nymphs. Transmitted pathogens need to effectively to hide or defeat on defense mechanisms in tick midgut, which is the first tick-pathogen interaction [57, 58]. The pathogens eventually migrate to tick ovaries for promoting transovarial transmission, or migrate to salivary glands for being transmitted to a host through tick saliva [57, 58]. Thus, although the bacteria present in the blood from the feeders could not be cultivated, our results provide strong assumptions about the ability of R. sanguineus to transmit B. henselae during the bite.
One possible reason for failure to obtain bacterial colonies can be that the number of viable bacteria from the collected sample was too low as only 10 μl out of the 4 ml of blood from each feeder was used for culture. Once ticks are feeding on an artificial membrane, a few microliters of tick saliva are mixed with the blood in the feeder, which possibly reduce the bacterial concentration below the detectable level [36]. In the study of Cotté and co-workers, some B. henselae colonies were obtained from 10 μl of blood taken from the feeder 84 hours after I. ricinus attachment, but that study concerned females infected at the nymphal stage, which inject more saliva during their feeding than nymphs and then possibly more bacteria [35]. To validate or not the viability of the bacteria presents into the blood of the feeder after R. sanguineus nymphal feeding, a higher volume of blood should be then tested in culture. Another hypothesis is that the bacteria, although alive to join the salivary glands, was however destroyed by the tick immune system in the salivary glands, as for example, by the salivary 5.3kDa antimicrobial protein (ISAMP), which has the ability to kill gram-positive and gram-negative bacteria [59]. In this case, this explanation echoed the hypothesis of selective adaptation of Bartonella spp. In fact, several arthropod vectors may harbor a wide range of Bartonella spp. but may not be efficient vectors for transferring all these Bartonella spp. to their hosts [60].
The presence of B. henselae DNA in larval feces samples also suggest that B. henselae-contaminated tick feces could be a potential source of Bartonella infection for humans or animals as it is the case for fleas [5–9]. However, no B. henselae DNA was detected in nymphal feces suggesting that bacteria may not stay in tick digestive system during the tick molting period. However, further investigations, confirming the viability of B. henselae from tick feces by culture, are required to understand the role of tick feces in Bartonella transmission. In a previous study, B. vinsonii subsp. berkhoffii DNA has been detected in R. sanguineus tick feces after post-capillary tube feeding, but no viable Bartonella could be isolated because of bacterial and fungal contamination [39]. Even if in vivo experiment has also showed that no dogs became bacteremia nor seroconverted when inoculated with tick feces collected from Bartonella-fed ticks, tick feces could not be ruled out as a potential source of Bartonella infection [39].
In conclusion, our findings demonstrated that the membrane feeding system could be successfully used to engorge and maintain R. sanguineus colony in the laboratory. Such a development now opens up real perspectives in the study of R. sanguineus, its biology, its vector competence, as well as in the study of molecular interactions with the transmitted pathogens. Our results also support that B. henselae could be transstadial transmitted from R. sanguineus larvae to nymphs. However, future both in vitro and in vivo studies would need to be performed to clarify more evidence of the vector competence of R. sanguineus larvae, nymphs, and adults for B. henselae. Despite having probably low epidemiological importance, the demonstration of such competence would first emphasize, another time, broad arthropod host range for bartonellae. Secondly, it would explain some cases of bartonellosis in patients, especially to include the diagnosis of bartonellosis following R. sanguineus tick bites, which is the most widespread tick in the world.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This study was supported by the Ministry of Science and Technology, Taiwan ROC (grant number MOST 105-2311-B-020-005 and MOST 107-2313-B-020-013).
References
- 1.Kordick DL, Wilson KH, Sexton DJ, Hadfield TL, Berkhoff HA, Breitschwerdt EB. Prolonged Bartonella bacteremia in cats associated with cat-scratch disease patients. J Clin Microbiol. 1995;33(12):3245–51. 10.1128/JCM.33.12.3245-3251.1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Boulouis HJ, Chang CC, Henn JB, Kasten RW, Chomel BB. Factors associated with the rapid emergence of zoonotic Bartonella infections. Vet Res. 2005;36(3):383–410. 10.1051/vetres:2005009 . [DOI] [PubMed] [Google Scholar]
- 3.Celebi B, Kilic S, Aydin N, Tarhan G, Carhan A, Babur C. Investigation of Bartonella henselae in cats in Ankara, Turkey. Zoonoses Public Health. 2009;56(4):169–75. 10.1111/j.1863-2378.2008.01170.x . [DOI] [PubMed] [Google Scholar]
- 4.Fabbi M, De Giuli L, Tranquillo M, Bragoni R, Casiraghi M, Genchi C. Prevalence of Bartonella henselae in Italian stray cats: evaluation of serology to assess the risk of transmission of Bartonella to humans. J Clin Microbiol. 2004;42(1):264–8. 10.1128/jcm.42.1.264-268.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Abbott RC, Chomel BB, Kasten RW, Floyd-Hawkins KA, Kikuchi Y, Koehler JE, et al. Experimental and natural infection with Bartonella henselae in domestic cats. Comp Immunol Microbiol Infect Dis. 1997;20(1):41–51. Epub 1997/01/01. 10.1016/s0147-9571(96)00025-2 . [DOI] [PubMed] [Google Scholar]
- 6.Chomel BB, Kasten RW, Floyd-Hawkins K, Chi B, Yamamoto K, Roberts-Wilson J, et al. Experimental transmission of Bartonella henselae by the cat flea. J Clin Microbiol. 1996;34(8):1952–6. 10.1128/JCM.34.8.1952-1956.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Foil L, Andress E, Freeland RL, Roy AF, Rutledge R, Triche PC, et al. Experimental infection of domestic cats with Bartonella henselae by inoculation of Ctenocephalides felis (Siphonaptera: Pulicidae) feces. J Med Entomol. 1998;35(5):625–8. 10.1093/jmedent/35.5.625 . [DOI] [PubMed] [Google Scholar]
- 8.Gurfield AN, Boulouis HJ, Chomel BB, Heller R, Kasten RW, Yamamoto K, et al. Coinfection with Bartonella clarridgeiae and Bartonella henselae and with different Bartonella henselae strains in domestic cats. J Clin Microbiol. 1997;35(8):2120–3. Epub 1997/08/01. 10.1128/JCM.35.8.2120-2123.1997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Iannino F, Salucci S, Di Provvido A, Paolini A, Ruggieri E. Bartonella infections in humans dogs and cats. Vet Ital. 2018;54(1):63–72. Epub 2018/04/10. 10.12834/VetIt.398.1883.2 . [DOI] [PubMed] [Google Scholar]
- 10.Carithers HA. Cat-scratch disease. An overview based on a study of 1,200 patients. Am J Dis Child. 1985;139(11):1124–33. Epub 1985/11/01. 10.1001/archpedi.1985.02140130062031 . [DOI] [PubMed] [Google Scholar]
- 11.Brouqui P, Raoult D. Endocarditis due to rare and fastidious bacteria. Clin Microbiol Rev. 2001;14(1):177–207. Epub 2001/01/09. 10.1128/CMR.14.1.177-207.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Malatack JJ, Jaffe R. Granulomatous hepatitis in three children due to cat-scratch disease without peripheral adenopathy. An unrecognized cause of fever of unknown origin. Am J Dis Child. 1993;147(9):949–53. Epub 1993/09/01. 10.1001/archpedi.1993.02160330039014 . [DOI] [PubMed] [Google Scholar]
- 13.Mazur-Melewska K, Mania A, Kemnitz P, Figlerowicz M, Sluzewski W. Cat-scratch disease: a wide spectrum of clinical pictures. Postepy Dermatol Alergol. 2015;32(3):216–20. Epub 2015/07/15. 10.5114/pdia.2014.44014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Arisoy ES, Correa AG, Wagner ML, Kaplan SL. Hepatosplenic cat-scratch disease in children: selected clinical features and treatment. Clinical Infectious Diseases. 1999;28(4):778–84. 10.1086/515197 [DOI] [PubMed] [Google Scholar]
- 15.Chian CA, Arrese Estrada J, Pierard G. Skin manifestations of Bartonella infections. International journal of dermatology. 2002;41(8):461–6. 10.1046/j.1365-4362.2002.01489.x [DOI] [PubMed] [Google Scholar]
- 16.Breitschwerdt EB, Maggi RG, Duncan AW, Nicholson WL, Hegarty BC, Woods CW. Bartonella species in blood of immunocompetent persons with animal and arthropod contact. Emerg Infect Dis. 2007;13(6):938–41. 10.3201/eid1306.061337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Breitschwerdt EB, Maggi RG, Nicholson WL, Cherry NA, Woods CW. Bartonella sp. bacteremia in patients with neurological and neurocognitive dysfunction. J Clin Microbiol. 2008;46(9):2856–61. 10.1128/JCM.00832-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chang CC, Chomel BB, Kasten RW, Romano V, Tietze N. Molecular evidence of Bartonella spp. in questing adult Ixodes pacificus ticks in California. J Clin Microbiol. 2001;39(4):1221–6. 10.1128/JCM.39.4.1221-1226.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Duplan F, Davies S, Filler S, Abdullah S, Keyte S, Newbury H, et al. Anaplasma phagocytophilum, Bartonella spp., haemoplasma species and Hepatozoon spp. in ticks infesting cats: a large-scale survey. Parasit Vectors. 2018;11(1):201 10.1186/s13071-018-2789-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Eskow E, Rao RV, Mordechai E. Concurrent infection of the central nervous system by Borrelia burgdorferi and Bartonella henselae: evidence for a novel tick-borne disease complex. Arch Neurol. 2001;58(9):1357–63. 10.1001/archneur.58.9.1357 . [DOI] [PubMed] [Google Scholar]
- 21.Holden K, Boothby JT, Kasten RW, Chomel BB. Co-detection of Bartonella henselae, Borrelia burgdorferi, and Anaplasma phagocytophilum in Ixodes pacificus ticks from California, USA. Vector Borne Zoonotic Dis. 2006;6(1):99–102. 10.1089/vbz.2006.6.99 . [DOI] [PubMed] [Google Scholar]
- 22.Kang JG, Ko S, Smith WB, Kim HC, Lee IY, Chae JS. Prevalence of Anaplasma, Bartonella and Borrelia Species in Haemaphysalis longicornis collected from goats in North Korea. J Vet Sci. 2016;17(2):207–16. 10.4142/jvs.2016.17.2.207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lucey D, Dolan MJ, Moss CW, Garcia M, Hollis DG, Wegner S, et al. Relapsing illness due to Rochalimaea henselae in immunocompetent hosts: implication for therapy and new epidemiological associations. Clin Infect Dis. 1992;14(3):683–8. 10.1093/clinids/14.3.683 . [DOI] [PubMed] [Google Scholar]
- 24.Maggi RG, Ericson M, Mascarelli PE, Bradley JM, Breitschwerdt EB. Bartonella henselae bacteremia in a mother and son potentially associated with tick exposure. Parasit Vectors. 2013;6:101 10.1186/1756-3305-6-101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Podsiadly E, Chmielewski T, Tylewska-Wierzbanowska S. Bartonella henselae and Borrelia burgdorferi infections of the central nervous system. Ann N Y Acad Sci. 2003;990:404–6. 10.1111/j.1749-6632.2003.tb07400.x . [DOI] [PubMed] [Google Scholar]
- 26.Rar VA, Fomenko NV, Dobrotvorsky AK, Livanova NN, Rudakova SA, Fedorov EG, et al. Tickborne pathogen detection, Western Siberia, Russia. Emerg Infect Dis. 2005;11(11):1708–15. 10.3201/eid1111.041195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Regier Y, Ballhorn W, Kempf VA. Molecular detection of Bartonella henselae in 11 Ixodes ricinus ticks extracted from a single cat. Parasit Vectors. 2017;10(1):105 10.1186/s13071-017-2042-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tsai YL, Lin CC, Chomel BB, Chuang ST, Tsai KH, Wu WJ, et al. Bartonella infection in shelter cats and dogs and their ectoparasites. Vector Borne Zoonotic Dis. 2011;11(8):1023–30. 10.1089/vbz.2010.0085 . [DOI] [PubMed] [Google Scholar]
- 29.Vayssier-Taussat M, Moutailler S, Femenia F, Raymond P, Croce O, La Scola B, et al. Identification of Novel Zoonotic Activity of Bartonella spp., France. Emerg Infect Dis. 2016;22(3):457–62. 10.3201/eid2203.150269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wikswo ME, Hu R, Metzger ME, Eremeeva ME. Detection of Rickettsia rickettsii and Bartonella henselae in Rhipicephalus sanguineus ticks from California. J Med Entomol. 2007;44(1):158–62. 10.1603/0022-2585(2007)44[158:dorrab]2.0.co;2 . [DOI] [PubMed] [Google Scholar]
- 31.Zangwill KM, Hamilton DH, Perkins BA, Regnery RL, Plikaytis BD, Hadler JL, et al. Cat scratch disease in Connecticut. Epidemiology, risk factors, and evaluation of a new diagnostic test. N Engl J Med. 1993;329(1):8–13. 10.1056/NEJM199307013290102 . [DOI] [PubMed] [Google Scholar]
- 32.Halos L, Jamal T, Maillard R, Beugnet F, Le Menach A, Boulouis HJ, et al. Evidence of Bartonella sp. in questing adult and nymphal Ixodes ricinus ticks from France and co-infection with Borrelia burgdorferi sensu lato and Babesia sp. Vet Res. 2005;36(1):79–87. 10.1051/vetres:2004052 . [DOI] [PubMed] [Google Scholar]
- 33.Maggi RG, Mascarelli PE, Havenga LN, Naidoo V, Breitschwerdt EB. Co-infection with Anaplasma platys, Bartonella henselae and Candidatus Mycoplasma haematoparvum in a veterinarian. Parasit Vectors. 2013;6:103 Epub 2013/04/17. 10.1186/1756-3305-6-103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sykes JE, Lindsay LL, Maggi RG, Breitschwerdt EB. Human coinfection with Bartonella henselae and two hemotropic mycoplasma variants resembling Mycoplasma ovis. J Clin Microbiol. 2010;48(10):3782–5. Epub 2010/08/13. 10.1128/JCM.01029-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cotte V, Bonnet S, Le Rhun D, Le Naour E, Chauvin A, Boulouis HJ, et al. Transmission of Bartonella henselae by Ixodes ricinus. Emerg Infect Dis. 2008;14(7):1074–80. 10.3201/eid1407.071110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Reis C, Cote M, Le Rhun D, Lecuelle B, Levin ML, Vayssier-Taussat M, et al. Vector competence of the tick Ixodes ricinus for transmission of Bartonella birtlesii. PLoS Negl Trop Dis. 2011;5(5):e1186 10.1371/journal.pntd.0001186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Dantas-Torres F, Chomel BB, Otranto D. Ticks and tick-borne diseases: a One Health perspective. Trends Parasitol. 2012;28(10):437–46. 10.1016/j.pt.2012.07.003 . [DOI] [PubMed] [Google Scholar]
- 38.Satta G, Chisu V, Cabras P, Fois F, Masala G. Pathogens and symbionts in ticks: a survey on tick species distribution and presence of tick-transmitted micro-organisms in Sardinia, Italy. J Med Microbiol. 2011;60(Pt 1):63–8. 10.1099/jmm.0.021543-0 . [DOI] [PubMed] [Google Scholar]
- 39.Billeter SA, Kasten RW, Killmaster LF, Breitschwerdt EB, Levin ML, Levy MG, et al. Experimental infection by capillary tube feeding of Rhipicephalus sanguineus with Bartonella vinsonii subspecies berkhoffii. Comp Immunol Microbiol Infect Dis. 2012;35(1):9–15. Epub 2011/11/09. 10.1016/j.cimid.2011.09.004 . [DOI] [PubMed] [Google Scholar]
- 40.Bonnet S, Brisseau N, Hermouet A, Jouglin M, Chauvin A. Experimental in vitro transmission of Babesia sp. (EU1) by Ixodes ricinus. Vet Res. 2009;40(3):21 Epub 2009/02/13. 10.1051/vetres/2009004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bonnet S, Jouglin M, Malandrin L, Becker C, Agoulon A, L'Hostis M, et al. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology. 2007;134(Pt 2):197–207. 10.1017/S0031182006001545 . [DOI] [PubMed] [Google Scholar]
- 42.Bonnet S, Liu X. Laboratory artificial infection of hard ticks: a tool for the analysis of tick-borne pathogen transmission. Acarologia. 2012;52(4):453–64. [Google Scholar]
- 43.Teng K, Jiang Z. Economic Insect Fauna of China: Fasc 39: Acari: Ixodidae Beijing, China: Science Press; 1991. [Google Scholar]
- 44.Troughton DR, Levin ML. Life cycles of seven ixodid tick species (Acari: Ixodidae) under standardized laboratory conditions. J Med Entomol. 2007;44(5):732–40. 10.1603/0022-2585(2007)44[732:lcosit]2.0.co;2 . [DOI] [PubMed] [Google Scholar]
- 45.Krober T, Guerin PM. In vitro feeding assays for hard ticks. Trends Parasitol. 2007;23(9):445–9. Epub 2007/08/08. 10.1016/j.pt.2007.07.010 . [DOI] [PubMed] [Google Scholar]
- 46.Wei JCJ, Edwards GA, Martin DJ, Huang H, Crichton ML, Kendall MAF. Allometric scaling of skin thickness, elasticity, viscoelasticity to mass for micro-medical device translation: from mice, rats, rabbits, pigs to humans. Sci Rep. 2017;7(1):15885 Epub 2017/11/23. 10.1038/s41598-017-15830-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sato S, Kabeya H, Negishi A, Tsujimoto H, Nishigaki K, Endo Y, et al. Molecular survey of Bartonella henselae and Bartonella clarridgeiae in pet cats across Japan by species-specific nested-PCR. Epidemiol Infect. 2017;145(13):2694–700. Epub 2017/08/07. 10.1017/S0950268817001601 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rolain JM, Franc M, Davoust B, Raoult D. Molecular detection of Bartonella quintana, B. koehlerae, B. henselae, B. clarridgeiae, Rickettsia felis, and Wolbachia pipientis in cat fleas, France. Emerg Infect Dis. 2003;9(3):338–42. 10.3201/eid0903.020278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Biggerstaff B. PooledInfRate, version 4.0: A microsoft office add-in to compute prevalence estimates from pooled samples. 2009.
- 50.Liu XY, Cote M, Paul RE, Bonnet SI. Impact of feeding system and infection status of the blood meal on Ixodes ricinus feeding. Ticks Tick Borne Dis. 2014;5(3):323–8. Epub 2014/03/07. 10.1016/j.ttbdis.2013.12.008 . [DOI] [PubMed] [Google Scholar]
- 51.Cornet S, Nicot A, Rivero A, Gandon S. Malaria infection increases bird attractiveness to uninfected mosquitoes. Ecol Lett. 2013;16(3):323–9. Epub 2012/12/05. 10.1111/ele.12041 . [DOI] [PubMed] [Google Scholar]
- 52.Koella JC, Sorensen FL, Anderson RA. The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc Biol Sci. 1998;265(1398):763–8. Epub 1998/06/17. 10.1098/rspb.1998.0358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lacroix R, Mukabana WR, Gouagna LC, Koella JC. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol. 2005;3(9):e298 Epub 2005/08/04. 10.1371/journal.pbio.0030298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Van Den Abbeele J, Caljon G, De Ridder K, De Baetselier P, Coosemans M. Trypanosoma brucei modifies the tsetse salivary composition, altering the fly feeding behavior that favors parasite transmission. PLoS Pathog. 2010;6(6):e1000926 Epub 2010/06/10. 10.1371/journal.ppat.1000926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Campbell EM, Burdin M, Hoppler S, Bowman AS. Role of an aquaporin in the sheep tick Ixodes ricinus: assessment as a potential control target. Int J Parasitol. 2010;40(1):15–23. Epub 2009/07/29. 10.1016/j.ijpara.2009.06.010 . [DOI] [PubMed] [Google Scholar]
- 56.Koch HG, Sauer JR. Quantity of blood ingested by four species of hard ticks (Acari: Ixodidae) fed on domestic dogs. Annals of the Entomological Society of America. 1984;77(2):142–6. [Google Scholar]
- 57.de la Fuente J, Antunes S, Bonnet S, Cabezas-Cruz A, Domingos AG, Estrada-Pena A, et al. Tick-Pathogen Interactions and Vector Competence: Identification of Molecular Drivers for Tick-Borne Diseases. Front Cell Infect Microbiol. 2017;7:114 Epub 2017/04/26. 10.3389/fcimb.2017.00114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hajdusek O, Sima R, Ayllon N, Jalovecka M, Perner J, de la Fuente J, et al. Interaction of the tick immune system with transmitted pathogens. Front Cell Infect Microbiol. 2013;3:26 Epub 2013/07/23. 10.3389/fcimb.2013.00026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pichu S, Ribeiro JM, Mather TN. Purification and characterization of a novel salivary antimicrobial peptide from the tick, Ixodes scapularis. Biochem Biophys Res Commun. 2009;390(3):511–5. Epub 2009/10/27. 10.1016/j.bbrc.2009.09.127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tsai YL, Chang CC, Chuang ST, Chomel BB. Bartonella species and their ectoparasites: selective host adaptation or strain selection between the vector and the mammalian host? Comp Immunol Microbiol Infect Dis. 2011;34(4):299–314. Epub 2011/05/28. 10.1016/j.cimid.2011.04.005 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.