

Abstract
The phenomenon of RNA polymerase II (Pol II) pausing at transcription start site (TSS) is one of the key rate-limiting steps in regulating genome-wide gene expression. In Drosophila embryo, Pol II pausing is known to regulate the developmental control genes expression, however, the functional implication of Pol II pausing during later developmental time windows remains largely unknown. A highly conserved zinc finger transcription factor, Motif 1 Binding Protein (M1BP), is known to orchestrate promoter-proximal pausing. We found a new role of M1BP in regulating Drosophila eye development. Downregulation of M1BP function suppresses eye fate resulting in a reduced eye or a “no-eye” phenotype. The eye suppression function of M1BP has no domain constraint in the developing eye. Downregulation of M1BP results in more than two-fold induction of wingless (wg) gene expression along with robust induction of Homothorax (Hth), a negative regulator of eye fate. The loss-of-eye phenotype of M1BP downregulation is dependent on Wg upregulation as downregulation of both M1BP and wg, by using wgRNAi, shows a significant rescue of a reduced eye or a “no-eye” phenotype, which is accompanied by normalizing of wg and hth expression levels in the eye imaginal disc. Ectopic induction of Wg is known to trigger developmental cell death. We found that upregulation of wg as a result of downregulation of M1BP also induces apoptotic cell death, which can be significantly restored by blocking caspase-mediated cell death. Our data strongly imply that transcriptional regulation of wg by Pol II pausing factor M1BP may be one of the important regulatory mechanism(s) during Drosophila eye development.
Subject terms: Developmental biology, Genetics, Neuroscience, Stem cells
Introduction
During organogenesis, the intricate process of gene regulation is facilitated by sequence specific factors and co-regulators, which turns on only a fraction of genes, while the rest of the genes are repressed or turned off. The dynamic temporal and spatial expression patterns of developmental control genes involves several checkpoints, starting from recruiting the general transcription machinery and RNA polymerase II (Pol II) to the gene promoter to initiate transcription. In higher eukaryotes, pausing of Pol II during early elongation phase of transcription serves as one of the regulatory mechanisms1,2. The genome-wide studies have shown that the regulation of Pol II activity near the transcription start site (TSS) is a widespread phenomenon in mammalian embryonic stem cells (ESCs) and Drosophila3,4. During Drosophila embryogenesis, most of the developmental control genes such as Hox genes, including the target gene promoters for various transcription factors and components of signaling pathways are transcriptionally paused. In Drosophila embryo, transcriptional regulation of three critical segmentation genes, sloppy-paired-1 (slp1), wingless (wg) and engrailed (en) by Pol II pausing may play an important role in controlling the gene expression. However, its exact mechanism along later developmental time points is not completely understood2,5–8. We used Drosophila eye model to study the role of Pol II pausing during organogenesis such as development of adult organs/appendages from their imaginal primordium.
Drosophila eye is a highly versatile and tractable model system for understanding the gene regulatory mechanisms underlying complex developmental programs9–13. The adult Drosophila compound eye is a highly organized structure with approximately 600–800 ommatidia or unit eyes arranged in a hexagonal lattice14. Each ommatidium is comprised of approximately 20 cells including 8 photoreceptor (PR) cells, and non-neuronal cells like pigment cells, cone cells and bristles9,14–16. Of these 8 photoreceptor cells, there are outer photoreceptors R1-R6 and inner photoreceptors R7-R8. The adult eye develops from the larval eye-antennal imaginal disc17. The eye imaginal disc is specified during embryonic and early larval development by action of core retinal determination (RD) genes, Pax6 homolog, eyeless (ey) and twin of eyeless (toy) and a network of mainly downstream transcription factors, eyes absent (eya), sine oculis (so), dachshund (dac), eyegone (eyg) and optix (opt)9,12,13,18–27. Of these, ey, a Drosophila homolog of Pax-6, is involved in eye field specification whereas the downstream genes like eya, so and dac are involved in retinal determination and differentiation15,23,27. Loss-of-function of RD genes block the early eye development.
In the developing eye imaginal disc of Drosophila third instar larva, a synchronous wave of retinal differentiation is initiated from the posterior margin of the eye imaginal disc, which moves anteriorly, and is referred to as the Morphogenetic Furrow (MF)14,16. The MF, a transient indentation in the developing eye disc, sweeps progressively across the eye disc towards the anterior margin, resulting in the formation of uniformly spaced photoreceptor clusters behind the MF. This process of differentiation of retinal precursor cells to photoreceptor neurons is driven by combinatorial action of the evolutionarily conserved Hedgehog (Hh) and Decapentaplegic (Dpp) signaling pathways, which plays important role in initiation and progression of the MF14,16,28–30. Within each differentiating photoreceptor cluster, Hh and Dpp signaling activates the expression of proneural genes like atonal (ato) that encodes basic HLH proteins and specifies the R8 photoreceptor31–33. Followed by the R8 selection, sequential recruitment of R2/R5, R3/R4, R1/R6 and R7 occurs9,22,34. Another secreted protein Scabrous (Sca), which is expressed within and near the intermediate clusters in the MF, is required for the correct spacing of photoreceptor clusters35,36.
The differentiation of retinal neurons and MF progression is opposed by the secreted morphogen Wg (a homolog of mouse Wnt-1 gene), which is expressed on the antero-lateral regions of the eye imaginal disc37–39. Wg serves as a ligand for the highly conserved Wg/ WNT signaling pathway. In the developing eye, Wg, a morphogen, is involved in diverse functions like cell proliferation, cell death, and cell-fate specification40–42. Wg expression levels plays important role(s) in determining the eye versus head fate by antagonizing dpp and thereby suppressing retinal determination11,13,15,38,42,43. Wg regulates expression of another negative regulator of eye development, homothorax (hth), a MEIS class gene with a highly conserved Meis-Hth (MH) domain and a homeodomain (HD), which is expressed uniformly anterior to the MF44–48 and suppresses the eye fate. Hence, ectopic upregulation of wg promotes head-specific fate by regulating MF progression during eye development39,44,49.
During eye development, one of the many functions of Wg signaling is to induce programmed cell death by activating the expression of head involution defective (hid), reaper (rpr), and grim in ommatidia at the periphery of the eye during pupal stage50,51. In the developing larval eye field, apoptosis can be induced by a variety of stimuli like inappropriate levels of morphogens or extracellular signaling52,53. Ectopic induction of Wg signaling causes developmental or morphogenetic cell death in the larval eye imaginal disc54. This developmental cell death is caused by activation of caspases and is different from programmed cell death observed in the pupal retina54,55. The baculovirus anti-apoptotic protein P35 upon ectopic expression in the developing field can block caspase-dependent cell death56. P35 acts through inhibition of range of initiator to executioner class of caspases.
In Drosophila, the sequence-specific transcription factors, the GAGA factor (GAF) and the Motif 1 Binding Protein (M1BP) have been implicated in dictating Pol II pausing57. The regulatory mechanism associated with GAF exhibits greater transcriptional plasticity than M1BP. M1BP binds to a core promoter element called Motif 1 and has been shown to orchestrate promoter-proximal pausing in GAF-independent manner57. M1BP is highly conserved across the species and encodes a 55 kDa protein containing zinc-associated domain (ZAD) towards the N-terminus and five C2H2 zinc-fingers domains toward the C-terminus. Drosophila M1BP is functionally homologous to vertebrate zinc finger with a SCAN and a KRAB domain 3 (ZKSCAN3) transcription factors and shows structural similarity for the C-terminal C2H2 zinc-finger domains58,59. The M1BP binding site sequence was reported using bioinformatics and biochemical analysis60. In addition, these studies led to tracking of Motif 1 binding activity, their associated proteins, and a list of 2187 genes whose expression is affected by M1BP57,61.
Here, we demonstrate that wg, which encodes a ligand for the highly conserved Wg/WNT signaling pathway is one of the targets for M1BP mediated transcriptional regulation during Drosophila eye development. We show that downregulation of M1BP ectopically induces wg gene expression in the developing eye disc, which causes suppression of the eye fate and induction of developmental cell death. Our results clearly indicate that M1BP mediated transcriptional regulation of Wg signaling could be a key regulatory mechanism during Drosophila eye development. We found potential M1BP binding sites in regulatory regions of wg gene using bioinformatics. Furthermore, this relation was also observed in the wing imaginal discs.
Materials and methods
Fly stocks
Fly stocks used in this study are described in Flybase (https://flybase.bio.indiana.edu). We used ey-Gal462, eyg-Gal426, bi-Gal4 (BL 58,815)63,64, dpp-Gal465, UAS-M1BPRNAi (BL 41,937)57, UAS-wgRNAi (BL 31,249)66, UAS-hthRNAi, yw, hth1422–4/TM6B, Tb47 dpp-lacZ (BL 5528)67, wg-lacZ62, UAS-P3556. We used the wild-type Canton-S stock of D. melanogaster in this study. Fly stocks were maintained at 25 °C on the regular cornmeal, yeast, molasses food medium.
Genetic crosses
We used Gal4/UAS TARGET system to misexpress the gene of interest68. All Gal4/UAS crosses were maintained at 18 °C, 25 °C and 29 °C, unless specified, to sample different induction levels10. The ey-Gal4 driver used in this study targets misexpression of inducible transgene M1BPRNAi in the entire developing eye domain (ey > M1BPRNAi) of larval eye imaginal disc. To misexpress M1BPRNAi in specific domains of the eye disc, different Gal4 drivers were used: eyg-Gal4 targets misexpression of transgene at the equator, bi-Gal4 selectively targets the expression of the transgene at the dorso-ventral (DV) eye margin64, dpp-Gal4 drives the expression of the transgene at the posterior margin of the eye disc30.
We also tested the gain-of-function of M1BP using the CRISPR/Cas9- based transcriptional activation approach69 to overexpress TRiP-CRISPR Overexpression (TRiP-OE) M1BP (BL 80231) in the dpp domain of the developing eye by crossing the TOE M1BP flies with dpp-Gal4; dcas9-VPR (BL 67045) flies, in which the tissue-specific Gal4 directs expression of a catalytically inactive dead Cas9 (dCas9) fused to a tripartite transcriptional activator domain, VP64-p65-Rta (VPR).
Immunohistochemistry
Eye-antennal discs of wandering third instar larvae were dissected in 1 × phosphate buffered saline (PBS), fixed in 4% paraformaldehyde in PBS (fixative) for 20 min and washed in PBST (three times). The tissues were stained with a combination of antibodies following the standard protocol39. Primary antibodies used were rabbit anti-β-GAL (1:100; Cappel); rat anti-Elav (1:100), mouse anti-Wg (1:100; Developmental Studies Hybridoma Bank, DSHB), mouse anti- Dlg (1:100); mouse anti-Eya (1:100; DSHB), mouse anti-Dac (1:100; DSHB), goat anti-Hth (1:200; Santa Cruz), mouse anti-Sca (1:100), goat anti-Ato (1:50), rabbit anti-Dcp1 (1:150, Santa Cruz), mouse anti-pH3 (1:300, Cell Signaling). The discs were washed in PBST thrice for 10 min. Secondary antibodies used were donkey anti-rat IgG conjugated to Cy5 (1:250), donkey anti-rabbit IgG conjugated to Cy3 (1:300) or goat anti-mouse IgG conjugated to FITC (1:200) (Jackson Laboratories). The discs were mounted in Vectashield and photo-documented on a Fluoview 3000 Laser Scanning Confocal Microscope70. We took the images at 20 × magnification unless stated otherwise. We analyzed and prepared the final figures with images using Adobe Photoshop CS6 software.
Adult eye imaging
Adult eye images were captured after freezing flies at − 20 °C for ~ 4 h. Images were taken on a MrC5 color camera mounted on an Axioimager.Z1 Zeiss Apotome using a Z-sectioning function of Axiovision software 4.6.371. The final images were prepared using Adobe Photoshop CS6 software.
Real time quantitative polymerase chain reaction (RT-qPCR)
Tissue was collected and homogenized in TRIzol Reagent (Invitrogen, Cat# 15596026). Total RNA was extracted following TRIzol protocol. Aqueous phase was transferred to RNA Clean & Concentrator-5 (Zymo research, Cat# R1013) columns and eluted in DNase/RNase-free water. Quality and quantity of isolated RNA was determined by Nanodrop 2000 spectrophotometer (Thermo Scientific). cDNA was synthesized from 1 µg of total RNA through Reverse Transcription reaction (RT) using first-strand cDNA synthesis kit (GE healthcare, Cat# 27926101). RT-qPCR was performed using BioRad iQ SYBR Green Supermix (Bio-Rad, Cat# 1708860) according to the standard protocol72–74 . Fold change was calculated using comparative CT method (2−ΔΔCT method). The primers used are:
GAPDHFw5′-GGCGGATAAAGTAAATGTGTGC-3′.
GAPDHRev5′-AGCTCCTCGTAGACGAACAT-3′.
wgFw5′-TCAGGGACGCAAGCATAATAG-3′.
wgRev5′-CGAAGGCTCCAGATAGACAAG-3′.
Statistics
Statistical analysis was performed using Microsoft excel software. The P-values were calculated using student’s t-test and the error bars represent Standard deviation from Mean. Statistical significance in each graph is shown by P-value: ***P < 0.001; **P < 0.01; *P < 0.0575–78.
Results
Downregulation of M1BP function suppresses the eye fate
The larval eye imaginal disc (Fig. 1A) develops into the adult compound eye comprising of 600–800 ommatidia or unit eyes (Fig. 1B). Targeted misexpression of UAS-GFP reporter transgene under ey-Gal4 driver (ey > GFP, shown in green) marks the entire eye imaginal disc (Fig. 1A). The eye discs were stained with a membrane-specific marker Dlg and pan-neuronal marker Elav (red), which marks the nuclei of the photoreceptor neurons. Targeted misexpression of inducible UAS-M1BPRNAi transgene using ey-Gal4 driver (ey > M1BPRNAi), which downregulates M1BP function in the developing eye imaginal disc, results in the suppression of eye fate (Fig. 1C,E). The eye suppression phenotype of ey > M1BPRNAi is evident from pan-neuronal marker Elav expression, which results in either highly reduced eye field (Fig. 1C) or a “no-eye” phenotype (Fig. 1E). The adult flies of ey > M1BPRNAi genotype also exhibits reduced eye phenotype (Fig. 1D). The penetrance of eye phenotype(s) in the adult ranges from “small-eye” (Fig. 1D, 5%, n = 100) to a “no-eye” (Fig. 1F, 95%, n = 100). Further, quantification of the adult eye area shows that the eye size significantly reduces in case of both small-eye as well as “no-eye” phenotype in ey > M1BPRNAi flies, when compared with the control (ey > GFP) flies (p < 0.001, Fig. 1G). We also studied the M1BP gain-of-function phenotype using the CRISPR/Cas9- based transcriptional activation approach69. We did not see any eye phenotypes in terms of change in size or fate. Although we found higher levels of M1BP protein expressed in the dpp-Gal4 driver expression domain (Fig. S1). These results suggest that M1BP function is required for Drosophila eye development.
Figure 1.

Downregulation of M1BP suppresses the eye fate during Drosophila eye development. (A) ey-Gal4 driven expression of UAS-GFP transgene in the eye. Note that ey > GFP (green) expressed in the entire eye field of third instar larval eye disc. Note that eye imaginal disc is stained with pan neuronal marker Elav (red) which marks the nuclei of retinal neurons and Dlg, a membrane specific marker, to mark outline of the tissue. (B) Adult eye. (C–F) Downregulation of M1BP in the eye by driving expression of UAS-M1BPRNAi (ey > M1BPRNAi) suppresses the eye fate as seen in (C,E) the eye imaginal disc and the (D,F) adult eye. ey > M1BPRNAi exhibits a range of eye suppression phenotype ranging from a (C,D) small-eye to a "no-eye". (G) The area of adult eye was quantified using Image J software (NIH). The p values for the eye size (μm2) were calculated in a set of five (n = 5) using Student’s t-test in MS Excel Software. ey-Gal4 was found to be statistically significant from ey > M1BPRNAi in case of both small-eye (p < 0.001, ***) and no-eye phenotype (p < 0.001, ***). The orientation of all imaginal discs is identical with posterior to the left and dorsal up. The magnification of all eye-antennal imaginal disc is 20 × and the adult eye is 10 ×. A total of five eye-antennal imaginal discs (n = 5) for each genotype were analyzed for respective immunohistochemistry staining.
Figure 5.

Downregulation of hth rescues M1BP loss-of-function phenotype of reduced eye. The reduced eye phenotype of (B) ey > M1BPRNAi is rescued when hth levels are downregulated using (D) hthRNAi (ey > M1BPRNAi + hthRNAi) or heterozygous combination of (F) hth1422–4 (ey > M1BPRNAi + hth122–4 / +), a null allele of hth. Eye-antennal disc of (A) ey-Gal4, (B) ey > M1BPRNAi, (C) ey > hthRNAi, (D) ey > M1BPRNAi + hthRNAi. (E) ey > hthRNAi, (F) ey > M1BPRNAi + hthRNAi .
Eye suppression phenotype due to downregulation of M1BP function has no domain constraint
In order to understand if the M1BP function has any domain constraint, we downregulated M1BP functions in different domains of the developing eye by misexpressing inducible UAS-M1BPRNAi transgene using various Gal4 drivers. The bi-Gal4 drives expression of UAS-GFP reporter (bi > GFP, shown in green) along the dorso-ventral (DV) margin of larval eye imaginal disc (Fig. 2A) as well as the adult eye (Fig. 2B)64,79. Downregulation of M1BP function in the bi expression domain (bi > M1BPRNAi) results in eye suppression along the DV margin in the eye imaginal disc (Fig. 2C). About 56% (n = 100) of the bi > M1BPRNAi adult flies showed reduced eye phenotype (Fig. 2D). We employed dpp-Gal4 driver, which drives expression of UAS-GFP reporter (dpp > GFP, shown in green) along the posterior margin of the developing eye imaginal disc (Fig. 2E)30, and on its own it does not affect the phenotype of adult eye (Fig. 2F). Downregulation of M1BP function in the dpp expression domain (dpp > M1BPRNAi) results in reduced eye field as seen in the eye imaginal disc (Fig. 2G) and in 19% (n = 100) of the adult flies observed (Fig. 2H). Further, we used eyg-Gal4, which drives the expression of UAS-GFP reporter (eyg > GFP, shown in green) at the equator of the developing eye disc (Fig. 2I)26, and does not affect the eye size on its own (Fig. 2J). Downregulation of M1BP function in eyg expression domain results in headless phenotype (Fig. 2K,L). The frequency of headless flies were around 42% (n = 100) of the adults screened. These results clearly demonstrated that there is no domain constraint in M1BP function to promote eye development.
Figure 2.

Eye suppression phenotype due to downregulation of M1BP has no domain constraint in the developing eye. Downregulation of M1BP in different domains of the developing eye using (A–D) bi-Gal4 driver (dorso-ventral margin, green), (E–H) dpp-Gal4 driver (posterior margin, green), (I–L) eyg-Gal4 driver (equatorial domain, green). Note that pan-neuronal marker Elav (red) marks the retinal neuron fate and GFP (green) marks the expression domain of the driver. Downregulation of M1BP using (UAS-M1BPRNAi) (C,D) in bi-Gal4 (bi > M1BPRNAi) domain results in suppression of eye fate, (G,H) in dpp-Gal4 (dpp > M1BPRNAi) domain results in eye suppression, and (K,L) in eyg-Gal4 domain (eyg > M1BPRNAi) results in complete loss of eye and head field as seen in the (K) eye imaginal disc and (L) the adult eye. The orientation of all imaginal discs is identical with posterior to the left and dorsal up. The magnification of all eye-antennal imaginal disc is 20 × and adult eye is 10 ×. A total of five eye-antennal imaginal discs (n = 5) for each genotype were analyzed for respective immunohistochemistry staining.
Downregulation of M1BP function blocks the eye fate and MF progression
Since downregulation of M1BP causes eye suppression, we studied Retinal Determination (RD) gene expression levels as a read-out to study retinal determination and differentiation, the fundamental processes in the developing eye. A RD gene, eya, which acts downstream to ey, is expressed in a broader stripe in the differentiated cells posterior to the MF (Fig. 3A)80 whereas dac is expressed as two stripes directly anterior and posterior to the MF (Fig. 3C)81. We found that the downregulation of M1BP in the entire eye disc using ey-Gal4 (ey > M1BPRNAi) significantly reduces the size of the eye field as evident from the pan-neuronal marker Elav expression, which was accompanied by strong suppression of Eya and Dac expression levels (Fig. 3B,D, arrows).
Figure 3.

Downregulation of M1BP affects retinal determination, retinal differentiation and Morphogenetic furrow (MF) progression. Eye-antennal imaginal disc of third instar larva stained for pan-neuronal marker Elav (red), which marks the photoreceptors and retinal determination markers (A,B) Eya (green), (C,D) Dac (green), Retinal differentiation markers (E,F) Ato, (G,H) Sca, and MF marker (I,J) dpp-lacZ reporter. Note that wild-type expression of (A) Eya (green), (C) Dac (green), (E) Ato (green), (G) Sca (green) and (I) dpp-lacZ (green) is downregulated in (B,D,F,H,J) ey > M1BPRNAi (arrow). Downregulation of M1BP function in the eye disc (ey > M1BP RNAi) suppresses expression of MF specific marker dpp-lacZ (arrow) as the eye field is reduced. The orientation of all imaginal discs is identical with posterior to the left and dorsal up. The magnification of all eye-antennal imaginal disc is 20 ×. A total of five eye-antennal imaginal discs (n = 5) for each genotype were analyzed for respective immunohistochemistry staining.
Expression of Atonal (Ato) and Scabrous (Sca) serves as early markers for retinal differentiation and are employed for R8 specification (Fig. 3E,G)32,82,83. Based on Elav and RD gene expression, we found that misexpression of UAS-M1BPRNAi in the developing eye (ey > M1BPRNAi) suppresses retinal neuron(s) differentiation as evident from significantly reduced expression levels of Ato and Sca (Fig. 3F,H, arrows). Our data suggests that downregulation of M1BP function not only affects the retinal determination but also suppresses the markers for R8 photoreceptor differentiation. It is known that R8 specification and differentiation is associated with MF progression. We therefore tested the requirement of M1BP function in MF progression.
In the developing eye imaginal disc, Hh and Dpp signaling is required for normal initiation and progression of MF14,29,84,85. We used dpp-lacZ, a transcriptional reporter for dpp gene, which also marks the progression of MF in the third instar eye imaginal disc. dpp-lacZ is expressed in a thin stripe that overlays the apical constrictions caused by the MF cells and marks the anterior boundary of Elav positive differentiated retinal neurons (Fig. 3I). dpp-lacZ expression in ey > M1BPRNAi discs shows that MF fails to progress from the posterior margin of the eye disc towards the anterior side (Fig. 3J, arrow) and hence, downregulation of M1BP in the developing eye represses differentiation, resulting in eye suppression. To discern the mechanism behind eye suppression phenotypes of ey > M1BPRNAi, we looked for the putative target(s) of M1BP.
Downregulation of M1BP function induces wg and Hth expression
In the developing eye imaginal disc, Wg serve as a negative regulator of eye fate, and blocks the progression of MF16,37,38,85,86. We tested if downregulation of M1BP affects the wg gene expression. Since M1BP is a transcriptional pausing factor, we studied the wg gene transcription quantitatively by using qPCR approach and qualitatively by using wg-lacZ reporter. The qPCR data showed that there is 2.2 fold increase in wg mRNA levels in ey > M1BPRNAi discs as compared to the controls (Fig. 4A).
Figure 4.

Downregulation of M1BP induces ectopic expression wg and hth expression in the developing eye disc. (A) Relative expression of wg at the transcriptional level using quantitative PCR (q-PCR) in ey-Gal4, and ey > M1BPRNAi background. (B,E,H) Eye antennal imaginal disc stained for (B) wg-lacZ reporter (green) (E) Wg protein exhibits antero-lateral expression, (H) Hth (Green), a negative regulator of eye, is expressed anterior to the MF in the eye disc. (C,F,I) Downregulation of M1BP (ey > M1BPRNAi) suppresses the eye fate accompanied with ectopic induction of (B) wg transcription as evident from lacZ reporter (arrow), (E) Wg protein (arrow), and (I) Hth, (arrow) in the developing eye field. (D,G,J) Downregulation of wg by using ey > wgRNAi along with M1BPRNAi (ey > M1BPRNAi + wgRNAi) restores reduced eye size and (D) wg-lacZ, (G) Wg protein, and (J) Hth expression in the eye imaginal disc. The orientation of all imaginal discs is identical with posterior to the left and dorsal up. The magnification of all eye-antennal imaginal disc is 20 ×. A total of five eye-antennal imaginal discs (n = 5) for each genotype were analyzed for respective immunohistochemistry staining.
The wg-lacZ reporter is expressed at antero-lateral margins of the developing third instar larval eye imaginal disc (Fig. 4B, shown in green). The ey > M1BPRNAi eye imaginal discs are significantly reduced in size and exhibits a robust ectopic induction of wg-lacZ reporter (arrow, Fig. 4C). To test if wg upregulation in ey > M1BPRNAi discs is responsible for eye suppression phenotype, we downregulated wg gene expression levels using wgRNAi in the background of ey > M1BPRNAi (ey > M1BPRNAi + wgRNAi), which resulted in significant reduction in eye suppression phenotype and restoration of size of eye field to near wild-type (Fig. 4D). In addition wg-lacZ reporter expression is restored (Fig. 4D). In comparison to the wild-type Wg expression in eye disc (Fig. 4E), we found robust induction and ectopic localization of Wg protein in reduced eye disc of ey > M1BPRNAi (arrow, Fig. 4F) whereas Wg protein levels are restored to wild-type levels in ey > M1BPRNAi + wgRNAi background (Fig. 4G).
Since Wg is a negative regulator of eye development and it promotes head fate by inducing downstream hth expression, we further analyzed Hth protein localization in the eye discs of ey-Gal4 (Fig. 4H), ey > M1BPRNAi (Fig. 4I) and ey > M1BPRNAi + wgRNAi background(s) (Fig. 4J). Hth, which is predominantly expressed anterior to the MF (Fig. 4H)39,47,48, exhibits robust induction in the reduced eye field of ey > M1BPRNAi (arrow, Fig. 4I). We found that downregulation of M1BP in the eye disc induces robust Hth expression. These results strongly imply that M1BP plays an important role in promoting eye development by negatively regulating Wg and downstream Hth levels in the developing eye.
Reducing hth function rescues M1BP loss-of-function phenotype in developing eye
We wanted to determine, if reduced eye or no-eye phenotype observed in ey > M1BPRNAi background, is due to induction of hth or if there is/are other downstream target(s) of Wg signaling. Therefore, we reduced hth levels using inducible UAS-hthRNAi or a heterozygous background of hth null allele, hth1422–4/TM6BTb47. In comparison to the control eye imaginal disc (Fig. 5A), downregulation of M1BP (ey > M1BPRNAi) results in a highly reduced “no-eye” phenotype (Fig. 5B), whereas downregulation of hth levels using UAS-hthRNAi in ey > M1BPRNAi background (ey > M1BPRNAi + hthRNAi) results in a significant rescue of ey > M1BPRNAi phenotype (Fig. 5D). In a heterozygous combination, the null allele of hth, hth1422–4/TM6B, exhibits a normal eye phenotype (Fig. 5E). However, reduction of hth function in ey > M1BPRNAi background (ey > M1BPRNAi, hth1422–4/TM6B) exhibits significant rescue of “no-eye” phenotype (Fig. 5F). However, there is no complete rescue to a wild-type eye and the frequency is low.
Downregulation of M1BP triggers developmental cell death
It has been shown that ectopic induction of Wg signaling in the eye disc induces developmental cell death, which results in reduced eye phenotypes54. To understand the genetic mechanism responsible for the reduced eye phenotype, manifested by flies where M1BP function was downregulated, we tested the role of cell death. It is known that ectopic expression of baculovirus P35 blocks caspase-dependent cell death56. In comparison to the wild-type eye-antennal imaginal disc and the adult eye (Fig. 6A,B), downregulation of M1BP function in ey > M1BPRNAi results in reduced eye as seen in the eye imaginal disc and the adult eye (Fig. 6C,D). Blocking caspase-dependent cell death by ectopic expression of UAS-P35 transgene in ey > M1BPRNAi (ey > M1BPRNAi + P35) background can restore the eye suppression phenotype as observed in the eye disc and the adult eye (Fig. 6E,F). Further, quantification of the area of the adult eyes of ey-Gal4 (Fig. 6B,G), ey > M1BPRNAi (Fig. 6D,G) and ey > M1BPRNAi + wgRNAi background(s) (Fig. 6F,G) shows that the eye size significantly reduces in ey > M1BPRNAi flies, when compared with ey-Gal4 (p < 0.001, Fig. 6G). However, blocking caspase-dependent cell death significantly restores the eye size in ey > M1BPRNAi + P35 background, when compared with the ey > M1BPRNAi flies (p < 0.001, Fig. 6G), and less significant than control ey-Gal4 flies (p < 0.01, Fig. 6G).
Figure 6.

Blocking cell death can rescue eye suppression phenotype of M1BP loss-of-function. (A, B) Wild-type eye imaginal disc and the adult eye. Note that eye imaginal disc is stained for Elav (red) and Wg (green). (C) Loss-of-function of M1BP (ey > M1BPRNAi) results in suppression of eye fate as seen in the eye disc (D) adult eye. Blocking cell death by ectopic expression of P35 in ey > M1BPRNAi background can rescue the eye suppression phenotype as seen in the eye disc and the adult eye. (G) The area of the adult eye was quantified using ImageJ software (NIH). The p values for the eye size (μm2) were calculated in a set of five (n = 5) using Student’s t-test in MS Excel Software. The eye size in ey-Gal4 flies were found to be highly significant from ey > M1BPRNAi (p < 0.001, ***) than ey > M1BPRNAi + P35 flies (p < 0.01, **). Blocking cell death in ey > M1BPRNAi + P35 flies restores the eye size, when compared with the ey-Gal4 flies (p < 0.05, *). (H–J) Eye imaginal disc stained for Elav (red) and Dcp-1 (green). Note that Dcp-1 marks the dying cells in the disc. (H’–J’) Eye antennal imaginal disc showing split channel for Dcp-1 staining. (K) Quantification of the Dcp-1 positive cells (green) shows that the average number of dying cells were significantly higher in ey > M1BPRNAi (p < 0.001, ***) than ey-Gal4 discs, however, when compared with the ey > M1BPRNAi + P35 discs, the number of dying cells were found to be comparable (non-significant) with the ey-Gal4 discs. The orientation of all imaginal discs is identical with posterior to the left and dorsal up. The magnification of all eye-antennal imaginal disc is 20 × and the adult eye is 10 ×. A total of five eye-antennal imaginal discs (n = 5) for each genotype were analyzed for respective immunohistochemistry staining.
To validate our hypothesis that ectopic upregulation of wg induces developmental cell death, which results in reduced eye phenotype seen in ey > M1BPRNAi eye disc, we used the antibody against Drosophila effector caspase, death caspase-1 (Dcp-1). Dcp-1, a critical executioner of apoptosis, serves as an excellent marker for cell death87. In the control ey-Gal4 eye disc, we found a few Dcp-1 positive dying cells/ retinal neurons (Fig. 6H,H’). The number of Dcp-1 positive dying cells gets almost doubled in ey > M1BPRNAi eye disc, which is highly reduced in size (Fig. 6I,I’,K). The number of Dcp-1positive dying cells is restored to the control ey-Gal4 (Fig. 6K) when P35 levels are upregulated in ey > M1BPRNAi (ey > M1BPRNAi + P35) background (Fig. 6J,J’). Further, quantification of the Dcp-1 positive nuclei shows that downregulation of M1BP in the eye disc (ey > M1BPRNAi) induces apoptotic cell death as the average number of dying cells were significantly higher in ey > M1BPRNAi (p < 0.001, Fig. 6K) as compared to the control ey-Gal4 eye discs, however, when compared with the ey > M1BPRNAi + P35 discs, the number of Dcp-1 positive dying cells were non-significant with respect to the ey-Gal4 eye discs (ns, Fig. 6K). These results suggest that overexpressing P35 where M1BP levels are downregulated restores the size of the eye field by reducing the average number of dying cells.
Downregulation of M1BP is independent of cell proliferation function
Since downregulation of M1BP function results in the small-eye phenotype, it is possible that the reduced number of Elav positive cells (red; Fig. 7B), which marks the photoreceptor neurons in the eye disc, is due to reduced cell proliferation. To test the role of cell proliferation in reduced eye phenotype, we stained the eye imaginal discs with phospho-histone 3 (pH3) that marks the proliferating cells (Fig. 7). Quantification of the pH3 positive cells show that the proliferating cells are significantly reduced in ey > M1BPRNAi discs (Fig. 7B,B’,D; p < 0.001) when compared with ey-Gal4 discs (Fig. 7A,A’,D). The reduction in number of pH3 positive cells in ey > M1BPRNAi discs does not clearly address if both cell death and cell-proliferation are involved. Although we have seen earlier that reduced size of the ey > M1BPRNAi eye disc is due to developmental cell death. In order to test the role of cell proliferation, we counted the pH3 positive cells in the ey > M1BPRNAi + P35 eye disc (where caspase-dependent cell death is blocked). Overexpression of UAS-P35 transgene along with downregulation of M1BP (ey > M1BPRNAi + P35) results in significant increase in the number of proliferating cells ey > M1BPRNAi + P35 discs, when compared with ey > M1BPRNAi discs (p < 0.001, Fig. 7C,C’, D). Interestingly, the number of pH3 positive nuclei are restored to the control (Fig. 7A,D). This data suggests that cell proliferation function is not the major contributing factor in reduced eye phenotype in ey > M1BPRNAi.
Figure 7.

Overexpression of P35 in loss-of-function background of M1BP promotes proliferation in the developing eye field. (A–C) Eye imaginal disc stained for Elav (red) and pH3 (green). Note that pH3 marks the proliferating cells in the disc. (A’–C’) Eye antennal imaginal disc showing split channel for pH3 staining. (D) Quantification of the pH3 positive cells shows (green) that the proliferating cells are significantly reduced in ey > M1BPRNAi (p < 0.001, ***) than eyGal4 discs, however, when compared with the ey > M1BPRNAi + P35 discs, the number of proliferating cells were found to be comparable (non-significant) with the ey-Gal4 discs. Overexpression of P35 in M1BP loss-of-function background promotes significantly higher rate of proliferation (p < 0.001, ***), when compared with the ey > M1BPRNAi discs. The orientation of all imaginal discs is identical with posterior to the left and dorsal up. The magnification of all eye-antennal imaginal disc is 20 ×. A total of five eye-antennal imaginal discs (n = 5) for each genotype were analyzed for respective immunohistochemistry staining.
Discussion
Pol II pausing near the transcription start site has been identified as a key step in optimizing transcription of many genes in metazoans. It has been proposed that pausing allows the coupling of transcription and RNA processing88. Pausing can contribute to dynamic regulation of gene expression in response to developmental and environmental signals7,89, and can function to repress transcription90. The genome-wide studies have revealed that ~ 10–40% of all genes in mammalian embryonic stem cells and Drosophila have paused promoters2,91–93. In Drosophila, while the phenomenon of promoter proximal pausing has been well studied in regulation of genes encoding the heat shock proteins (Hsp) and different components involved in immune response pathways6,90,94, it is also proposed to play important role in regulating the gene expression during early developmental events such as patterning, sex determination etc.2,5,7,95. So far, the sequence-specific transcription factors such as GAGA factor and M1BP, and other regulators HEXIM, LARP7 (La Ribonucleoprotein 7, Transcriptional Regulator) have been implicated in dictating Pol II pausing in Drosophila57,96. However, the biological relevance of transcriptional pausing and the exact mechanism by which the regulatory factors may contribute in pausing of Pol II is not fully understood.
M1BP regulates retinal determination and MF progression in developing eye
We tested for the first time the role of transcription pausing factor, M1BP during Drosophila eye development. We found that downregulation of M1BP levels in the developing eye results in strong suppression of eye fate (Fig. 1C–F), however, gain-of-function of M1BP did not affect the eye fate (Fig. S1) suggesting that optimum levels of M1BP are required for Drosophila eye development. Furthermore, we did not find any domain constraint in eye suppression function when M1BP levels were downregulated (Fig. 2C,D,G,H,K,L). In addition, when M1BP levels were downregulated (ey > M1BPRNAi) the expression of retinal determination and differentiation genes were strongly downregulated (Fig. 3B,D,F,H). Interestingly, we found that protein encoded by RD genes were downregulated in ey > M1BPRNAi background. Therefore, M1BP may not be affecting RD gene expression directly.
During eye development, a wave of differentiation, emanates from the posterior margin of the developing eye imaginal disc, which sweeps anteriorly across the retinal primordium. The crest of this wave is referred to as the MF, which results in retinal differentiation behind it14,85. The two signals dpp and hh plays an important role in initiation and progression of MF. We found that downregulation of M1BP affects retinal differentiation as well as progression of MF (Fig. 3). It suggests that M1BP role is to promote retinal differentiation as well as MF progression. Also, M1BP downregulates the level of negative regulator(s) of the eye fate. We screened for the genes, which may serve as target for M1BP mediated transcriptional pausing mechanism in Drosophila eye imaginal disc.
M1BP regulates wg gene expression in the developing eye
The protein encoded by Drosophila wg gene, a member of Wg/WNT signaling pathway, act short range inducer, which organizes the pattern of cells at a distance in the embryo. Since M1BP downregulation resulted in blocking retinal differentiation and MF progression, we looked for the targets of M1BP transcriptional pausing function using the candidate gene approach. We found that wg-lacZ reporter, which serves as a transcriptional read out for Wg, exhibits robust induction in eye imaginal discs where M1BP levels were downregulated (Fig. 4C). This observation was further validated by qPCR approach which showed that there is a 2.2-fold increase in wg gene expression (Figs. 4A, 8A). Furthermore, in high throughput microarray screen carried out in S2R + cells, wg was also identified as a target whose expression is downregulated by M1BP using M1BP RNAi. According to microarray analysis, wg shows a 5.5-fold change (raw value against wg gene ID) when cells are treated with M1BPRNAi 57.
Figure 8.
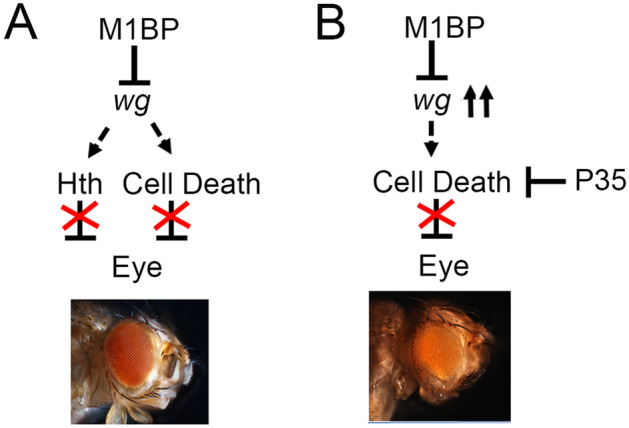
Models for M1BP function during Drosophila eye development. (A) M1BP suppresses the head fate by downregulating Wg and downstream Hth in the developing eye. Note that Wg and Hth are negative regulators of the eye development. (B) Blocking caspase-dependent cell death by overexpressing anti-apoptotic P35 transgene in ey > M1BPRNAi (ey > M1BPRNAi + P35) background significantly rescues the reduced eye phenotype.
To validate the results from qPCR approach as well induction of wg-lacZ reporter expression in ey > M1BPRNAi eye imaginal disc (Fig. 4), we also employed bioinformatics analysis to determine if there are M1BP binding sites in the wingless (wg) gene. The M1BP binding sequence (YGGTCACACTR) has been reported earlier57,61. We used this sequence for MEME analysis to screen for M1BP binding sites in wg gene and regulatory region60. We found 36 potential binding sites for M1BP in wingless gene and regulatory regions as shown in (S. Fig. 2B, Supplementary Table S1). Using these all 36 potential binding sites web logo was generated from weblogo.berkeley.edu/logo.cgi (Fig. S2B).
Wg, a ligand for evolutionarily conserved Wg/WNT signaling pathway, is known to act as a negative regulator of eye development13,37–39. During Drosophila eye development, Wg activity promotes head specific fate by negatively regulating MF progression in the differentiating eye imaginal disc22,37,38. Wg regulates expression of downstream gene hth, which encodes a MEIS class of transcription factor, and act as a negative regulator of eye development (Figs. 4, 8A)12,47–49. We found that in ey > M1BPRNAi background, robust induction of wg transcription also accompanies ectopic induction of hth along with the suppression of the eye fate (Figs. 4I, 8). Further, downregulation of wg levels, using wgRNAi, in ey > M1BPRNAi background rescued the eye suppression phenotype (Figs. 4, 8). This data clearly suggested that M1BP downregulates levels of wg, which in turn regulate expression of hth in the developing Drosophila eye (Fig. 5).
M1BP blocks Wg upregulation mediated developmental cell death
Higher levels of Wg are known to trigger developmental cell death in the developing eye field54. Interestingly, in ey > M1BPRNAi eye discs, the eye field was significantly reduced. Since, majority of the cell death is triggered by the activation of caspase-dependent cell death, blocking caspase-dependent cell death by ectopic expression of anti-apoptotic P35 transgene56 in ey > M1BPRNAi background showed rescue of eye suppression phenotype (Fig. 6E,F). However, these P35 mediated rescues of ey > M1BPRNAi were not as significant as seen with wgRNAi (Fig. 3). This suggests that Wg might be regulating eye fate through hth induction (Figs. 4I, 8A) and eye field size by triggering caspase mediated cell death (Figs. 6, 8B). In order to rule out that these in ey > M1BPRNAi phenotypes are not affected by reduced cell proliferation rates, we also tested levels of pH3 in these developing eye fields (Fig. 7). We found that cell proliferation rates were not affected by this transcriptional pausing mechanism in the developing eye.
Our results imply that the transcription pausing function of M1BP in regulating Wg signaling may play a critical role in Drosophila eye development (Fig. 8). However, other factors and signaling pathways involved in regulating the M1BP function at the mechanistic level is yet to be determined. In order to further understand, if M1BP mediated transcriptional regulation is also implicated during development of other imaginal discs in Drosophila, we studied the downregulation of M1BP function in bi-Gal4 domains of wing imaginal disc (Fig. S3). We wanted to test if this role of M1BP in regulating wg gene expression is exclusive to developing eye disc or it extends to other larval imaginal disc. We employed a bi-GA4 driver which drives the expression of a transgene in wing imaginal disc (Fig. S2A, A” shown in green)63,64. Downregulation of M1BP in bi-Gal4 expression domains of wing (bi > M1BPRNAi, Fig. S2B, B”) exhibits ectopic upregulation wg expression in the pouch region of the wing imaginal disc (Fig. S2B’, arrowhead). Furthermore, M1BP expression levels are downregulated in the wing pouch region, which corresponds to the bi-Gal4 expression domain. These results suggested that the transcription pausing function of M1BP may have similar target in the eye and wing imaginal disc. Recently, HEXIM, another transcriptional regulator associated with pol II pausing, has been reported to affect wing development in Drosophila by regulating Hh signaling97. In Drosophila wing imaginal disc, HEXIM knockdown causes developmental defects by inducing ectopic expression of hh and its transcriptional effector cubitus interuptus (ci), which triggers apoptosis. This suggests that the regulatory factors involved in Pol II pausing are important in maintaining the expression levels of different signaling pathways during development in Drosophila.
A number of highly conserved transcriptional pausing and elongation factors such as Spt5 precisely regulate transcription during Drosophila embryogenesis. The Spt5W049 missense mutation causes defects in the anterior–posterior patterning and segmental patterning during embryogenesis98. Interestingly, the mutant allele of Spt5 (foggym806) in Zebrafish also causes multiple developmental defects such as discrete problems with pigmentation, tail outgrowth, ear formation and cardiac differentiation. These studies suggest that the regulatory mechanism in Pol II pausing during fly development are also conserved in higher organisms. The Drosophila compound eye shares similarities with the vertebrate eye at the level of genetic machinery as well as the processes of differentiation99,100. Therefore, the information generated in Drosophila can be extrapolated to higher organisms11,100,101. Since Wnt signaling is known to induce programmed cell death in patterning the vasculature of the vertebrate eye102, it will be important to study what molecules other than M1BP can prevent Wg signaling from inducing cell death during early eye development.
Supplementary information
Acknowledgements
We thank Bloomington Drosophila Stock Center (BDSC) for Drosophila strains, and the Developmental Studies Hybridoma Bank (DSHB) for antibodies. We would like to thank Catherine Yeates, Ankita Sarkar, Abijeet Singh Mehta, and Evan J Wypasek for their help. We also thank Justin Kumar, David Gilmour, Y. Henry Sun, and Kyung Ok Cho for gift of fly strains and antibodies; and members of Singh and Kango-Singh lab for critical comments on the manuscript. Confocal microscopy was supported by core facility at University of Dayton. A.S. is supported by NIH1R15GM124654-01 from NIH, Schuellein Chair Endowment Fund and STEM Catalyst Grant from the University of Dayton.
Author contributions
A.S.: developed the concept. A.R. and A.C.V.: performed experiments. A.R., A.C.V., and A.S.: Analyzed the data. A.R. and A.S.: wrote the main manuscript text. A.R., A.C.V., and A.S.: Prepared figures. A.R., A.C.V., and A.S.: Reviewed the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-020-73891-7.
References
- 1.Mayer A, Landry HM, Churchman LS. Pause & go: from the discovery of RNA polymerase pausing to its functional implications. Curr. Opin. Cell Biol. 2017;46:72–80. doi: 10.1016/j.ceb.2017.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zeitlinger J, et al. RNA polymerase stalling at developmental control genes in the Drosophila melanogaster embryo. Nat. Genet. 2008;39:1512–1516. doi: 10.1038/ng.2007.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Adelman K, Lis JT. Promoter-proximal pausing of RNA polymerase II: emerging roles in metazoans. Nat. Rev. Genet. 2012;13:720–731. doi: 10.1038/nrg3293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Larson M, et al. A pause sequence enriched at translation start sites drives transcription dynamics in vivo. Science. 2014;344:1. doi: 10.1126/science.1251871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gaertner B, Zeitlinger J. RNA polymerase II pausing during development. Development. 2014;141:1179–1183. doi: 10.1242/dev.088492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gilchrist DA, et al. Regulating the regulators: the pervasive effects of Pol II pausing on stimulus-responsive gene networks. Genes Dev. 2012;26:933–944. doi: 10.1101/gad.187781.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Levine M. Paused RNA polymerase II as a developmental checkpoint. Cell. 2011;145:502–511. doi: 10.1016/j.cell.2011.04.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zouaz A, et al. The Hox proteins Ubx and AbdA collaborate with the transcription pausing factor M1BP to regulate gene transcription. EMBO J. 2017;36:2887–2906. doi: 10.15252/embj.201695751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kumar JP. My what big eyes you have: how the Drosophila retina grows. Dev. Neurobiol. 2011;71:1133–1152. doi: 10.1002/dneu.20921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Singh A, Chan J, Chern JJ, Choi KW. Genetic interaction of Lobe with its modifiers in dorsoventral patterning and growth of the Drosophila eye. Genetics. 2005;171:169–183. doi: 10.1534/genetics.105.044180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Singh A, Irvine KD. Drosophila as a model for understanding development and disease. Dev. Dyn. 2012;241:1–2. doi: 10.1002/dvdy.23712. [DOI] [PubMed] [Google Scholar]
- 12.Singh A, Tare M, Puli OR, Kango-Singh M. A glimpse into dorso-ventral patterning of the Drosophila eye. Dev. Dyn. 2012;241:69–84. doi: 10.1002/dvdy.22764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.13Tare, M., Puli, O. R., and & Singh, A. in Molecular Genetics of Axial Patterning, Growth and Disease in the Drosophila Eye(eds Amit Singh, and, & Madhuri Kango-Singh) Ch. 2, 37–75 (Springer, 2013).
- 14.Ready DF, Hanson TE, Benzer S. Development of the Drosophila retina, a neurocrystalline lattice. Dev. Biol. 1976;53:217–240. doi: 10.1016/0012-1606(76)90225-6. [DOI] [PubMed] [Google Scholar]
- 15.15Gogia, N., Puli, O. R., Raj, A., and & Singh, A. in Molecular Genetics of Axial Patterning, Growth and Disease in the Drosophila Eye Vol. II (eds Amit Singh, and, & Madhuri Kango-Singh) Ch. 2, 53–96 (Springer International Publishing, 2020).
- 16.16Kumar, J. P. in Molecular Genetics of Axial Patterning, Growth and Disease in the Drosophila Eye Vol. II (eds Amit Singh, and, & Madhuri Kango-Singh) Ch. 3, 97–120 (Springer International Publishing, 2020).
- 17.17Poulson, D. F. in Biology of Drosophila (ed M Demerec) 168–274 (Wiley, 1950).
- 18.Halder G, et al. Eyeless initiates the expression of both sine oculis and eyes absent during Drosophila compound eye development. Development. 1998;125:2181–2191. doi: 10.1242/dev.125.12.2181. [DOI] [PubMed] [Google Scholar]
- 19.Halder G, Callaerts P, Gehring WJ. Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science. 1995;267:1788–1792. doi: 10.1126/science.7892602. [DOI] [PubMed] [Google Scholar]
- 20.Quiring R, Walldorf U, Kloter U, Gehring WJ. Homology of the eyeless gene of Drosophila to the Small eye gene in mice and Aniridia in humans. Science. 1994;265:785–789. doi: 10.1126/science.7914031. [DOI] [PubMed] [Google Scholar]
- 21.Singh A, Tsonis PA. Focus on molecules: Six3–master or apprentice? Exp. Eye Res. 2010;90:535–536. doi: 10.1016/j.exer.2010.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Treisman JE. Retinal differentiation in Drosophila. Wiley Interdiscip. Rev. Dev. Biol. 2013;2:545–557. doi: 10.1002/wdev.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mishra, A. K. & Sprecher, S. G. in Molecular Genetics of Axial Patterning, Growth and Disease in the Drosophila Eye(eds Amit Singh, and, & Madhuri Kango-Singh) Ch. 1, 1–52 (Springer, 2020).
- 24.Czerny T, et al. twin of eyeless, a second Pax-6 gene of Drosophila, acts upstream of eyeless in the control of eye development. Mol. Cell. 1999;3:297–307. doi: 10.1016/S1097-2765(00)80457-8. [DOI] [PubMed] [Google Scholar]
- 25.Pignoni F, et al. The eye-specification proteins So and Eya form a complex and regulate multiple steps in Drosophila eye development. Cell. 1997;91:881–891. doi: 10.1016/s0092-8674(00)80480-8. [DOI] [PubMed] [Google Scholar]
- 26.Jang CC, et al. Two Pax genes, eye gone and eyeless, act cooperatively in promoting Drosophila eye development. Development. 2003;130:2939–2951. doi: 10.1242/dev.00522. [DOI] [PubMed] [Google Scholar]
- 27.Silver SJ, Rebay I. Signaling circuitries in development: insights from the retinal determination gene network. Development. 2005;132:3–13. doi: 10.1242/dev.01539. [DOI] [PubMed] [Google Scholar]
- 28.Wolff T, Ready DF. The beginning of pattern formation in the Drosophila compound eye: the morphogenetic furrow and the second mitotic wave. Development. 1991;113:841–850. doi: 10.1242/dev.113.3.841. [DOI] [PubMed] [Google Scholar]
- 29.Wolff, T. & Ready, D. F. in Bate, Martinez Arias, 1993.1277--1325.
- 30.Sarkar A, Gogia N, Farley K, Payton L, Singh A. Characterization of a morphogenetic furrow specific Gal4 driver in the developing Drosophila eye. PLoS ONE. 2018;13:e0196365. doi: 10.1371/journal.pone.0196365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bertrand N, Castro DS, Guillemot F. Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 2002;3:517–530. doi: 10.1038/nrn874. [DOI] [PubMed] [Google Scholar]
- 32.Jarman AP, Grell EH, Ackerman L, Jan LY, Jan YN. Atonal is the proneural gene for Drosophila photoreceptors. Nature. 1994;369:398–400. doi: 10.1038/369398a0. [DOI] [PubMed] [Google Scholar]
- 33.Tanaka-Matakatsu M, Du W. Direct control of the proneural gene atonal by retinal determination factors during Drosophila eye development. Dev. Biol. 2008;313:787–801. doi: 10.1016/j.ydbio.2007.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Treisman JE, Heberlein U. Eye development in Drosophila: formation of the eye field and control of differentiation. Curr. Top. Dev. Biol. 1998;39:119–158. doi: 10.1016/s0070-2153(08)60454-8. [DOI] [PubMed] [Google Scholar]
- 35.Baker NE, Yu S, Han D. Evolution of proneural atonal expression during distinct regulatory phases in the developing Drosophila eye. Curr. Biol. 1996;6:1290–1301. doi: 10.1016/s0960-9822(02)70715-x. [DOI] [PubMed] [Google Scholar]
- 36.Lee EC, Hu X, Yu SY, Baker NE. The scabrous gene encodes a secreted glycoprotein dimer and regulates proneural development in Drosophila eyes. Mol. Cell Biol. 1996;16:1179–1188. doi: 10.1128/mcb.16.3.1179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ma C, Moses K. Wingless and patched are negative regulators of the morphogenetic furrow and can affect tissue polarity in the developing Drosophila compound eye. Development. 1995;121:2279–2289. doi: 10.1242/dev.121.8.2279. [DOI] [PubMed] [Google Scholar]
- 38.Treisman JE, Rubin GM. wingless inhibits morphogenetic furrow movement in the Drosophila eye disc. Development. 1995;121:3519–3527. doi: 10.1242/dev.121.11.3519. [DOI] [PubMed] [Google Scholar]
- 39.Singh A, Kango-Singh M, Sun YH. Eye suppression, a novel function of teashirt, requires Wingless signaling. Development. 2002;129:4271–4280. doi: 10.1242/dev.129.18.4271. [DOI] [PubMed] [Google Scholar]
- 40.Seto ES, Bellen HJ. Internalization is required for proper Wingless signaling in Drosophila melanogaster. J. Cell Biol. 2006;173:95–106. doi: 10.1083/jcb.200510123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Swarup S, Verheyen EM. Wnt/Wingless signaling in Drosophila. Cold Spring Harb. Perspect. Biol. 2012;4:1. doi: 10.1101/cshperspect.a007930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Roignant JY, Treisman JE. Pattern formation in the Drosophila eye disc. Int. J. Dev. Biol. 2009;53:795–804. doi: 10.1387/ijdb.072483jr. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Royet J, Finkelstein R. Establishing primordia in the Drosophila eye-antennal imaginal disc: the roles of decapentaplegic, wingless and hedgehog. Development. 1997;124:4793–4800. doi: 10.1242/dev.124.23.4793. [DOI] [PubMed] [Google Scholar]
- 44.Bessa J, Gebelein B, Pichaud F, Casares F, Mann RS. Combinatorial control of Drosophila eye development by eyeless, homothorax, and teashirt. Genes Dev. 2002;16:2415–2427. doi: 10.1101/gad.1009002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kurant E, et al. Dorsotonals/homothorax, the Drosophila homologue of meis1, interacts with extradenticle in patterning of the embryonic PNS. Development. 1998;125:1037–1048. doi: 10.1242/dev.125.6.1037. [DOI] [PubMed] [Google Scholar]
- 46.Moskow JJ, Bullrich F, Huebner K, Daar IO, Buchberg AM. Meis1, a PBX1-related homeobox gene involved in myeloid leukemia in BXH-2 mice. Mol. Cell Biol. 1995;15:5434–5443. doi: 10.1128/MCB.15.10.5434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pai CY, et al. The Homothorax homeoprotein activates the nuclear localization of another homeoprotein, extradenticle, and suppresses eye development in Drosophila. Genes Dev. 1998;12:435–446. doi: 10.1101/gad.12.3.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rieckhof GE, Casares F, Ryoo HD, Abu-Shaar M, Mann RS. Nuclear translocation of extradenticle requires homothorax, which encodes an extradenticle-related homeodomain protein. Cell. 1997;91:171–183. doi: 10.1016/S0092-8674(00)80400-6. [DOI] [PubMed] [Google Scholar]
- 49.Singh A, et al. Opposing interactions between homothorax and Lobe define the ventral eye margin of Drosophila eye. Dev. Biol. 2011;359:199–208. doi: 10.1016/j.ydbio.2011.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cordero J, Jassim O, Bao S, Cagan R. A role for wingless in an early pupal cell death event that contributes to patterning the Drosophila eye. Mech. Dev. 2004;121:1523–1530. doi: 10.1016/j.mod.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 51.Lin HV, Rogulja A, Cadigan KM. Wingless eliminates ommatidia from the edge of the developing eye through activation of apoptosis. Development. 2004;131:2409–2418. doi: 10.1242/dev.01104. [DOI] [PubMed] [Google Scholar]
- 52.Mehlen P, Mille F, Thibert C. Morphogens and cell survival during development. J. Neurobiol. 2005;64:357–366. doi: 10.1002/neu.20167. [DOI] [PubMed] [Google Scholar]
- 53.53Yeates, C. J., Sarkar, A., Kango-Singh, M. & Singh, A. in Insights into Human Neurodegeneration: Lessons Learnt from Drosophila(eds Mousumi Mutsuddi, and, & Ashim Mukherjee) Ch. 2, 251–277 (Springer, 2019).
- 54.Singh A, Shi X, Choi K-W. Lobe and Serrate are required for cell survival during early eye development in Drosophila. Development. 2006;133:4771. doi: 10.1242/dev.02686. [DOI] [PubMed] [Google Scholar]
- 55.Singh A. Neurodegeneration: a means to an end. J. Cell Sci. Ther. 2012;3:1. doi: 10.4172/2157-7013.1000e107. [DOI] [Google Scholar]
- 56.Hay BA, Wolff T, Rubin GM. Expression of baculovirus P35 prevents cell death in Drosophila. Development. 1994;120:2121–2129. doi: 10.1242/dev.120.8.2121. [DOI] [PubMed] [Google Scholar]
- 57.Li J, Gilmour DS. Distinct mechanisms of transcriptional pausing orchestrated by GAGA factor and M1BP, a novel transcription factor. EMBO J. 2013;32:1829–1841. doi: 10.1038/emboj.2013.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Barthez, M. et al. Human ZKSCAN3 and Drosophila M1BP are functionally homologous transcription factors in autophagy regulation. doi:10.1101/747824 (2019). [DOI] [PMC free article] [PubMed]
- 59.Chung H-R, Schäfer U, Jäckle H, Böhm S. Genomic expansion and clustering of ZAD-containing C2H2 zinc-finger genes in Drosophila. EMBO Rep. 2003;3:1158–1162. doi: 10.1093/embo-reports/kvf243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Bailey TL, et al. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009;37:W202–208. doi: 10.1093/nar/gkp335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ohler U, Liao GC, Niemann H, Rubin GM. Computational analysis of core promoters in the Drosophila genome. Genome Biol. 2002;3:1. doi: 10.1186/gb-2002-3-12-research0087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hazelett DJ, Bourouis M, Walldorf U, Treisman JE. decapentaplegic and wingless are regulated by eyes absent and eyegone and interact to direct the pattern of retinal differentiation in the eye disc. Development. 1998;125:3741–3751. doi: 10.1242/dev.125.18.3741. [DOI] [PubMed] [Google Scholar]
- 63.Calleja M, Moreno E, Pelaz S, Morata G. Visualization of gene expression in living adult Drosophila. Science. 1996;274:252–255. doi: 10.1126/science.274.5285.252. [DOI] [PubMed] [Google Scholar]
- 64.Tare M, Puli OR, Moran MT, Kango-Singh M, Singh A. Domain specific genetic mosaic system in the Drosophila eye. Genesis. 2013;51:68–74. doi: 10.1002/dvg.22355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Staehling-Hampton K, Jackson PD, Clark MJ, Brand AH, Hoffmann FM. Specificity of bone morphogenetic protein-related factors: cell fate and gene expression changes in Drosophila embryos induced by decapentaplegic but not 60A. Cell Growth Differ. 1994;5:585–593. [PubMed] [Google Scholar]
- 66.Kabashi E, et al. TARDBP mutations in individuals with sporadic and familial amyotrophic lateral sclerosis. Nat. Genet. 2008;40:572–574. doi: 10.1038/ng.132. [DOI] [PubMed] [Google Scholar]
- 67.Blackman RK, Sanicola M, Raftery LA, Gillevet T, Gelbart WM. An extensive 3' cis-regulatory region directs the imaginal disk expression of decapentaplegic, a member of the TGF-beta family in Drosophila. Development. 1991;111:657–666. doi: 10.1242/dev.111.3.657. [DOI] [PubMed] [Google Scholar]
- 68.Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- 69.Ewen-Campen B, et al. Optimized strategy for in vivo Cas9-activation in Drosophila. Proc. Natl. Acad. Sci. U S A. 2017;114:9409–9414. doi: 10.1073/pnas.1707635114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Tare M, Sarkar A, Bedi S, Kango-Singh M, Singh A. Cullin-4 regulates Wingless and JNK signaling-mediated cell death in the Drosophila eye. Cell Death Dis. 2016;7:e2566. doi: 10.1038/cddis.2016.338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Irwin M, et al. A positive feedback loop of Hippo- and c-Jun-Amino-terminal kinase signaling pathways regulates amyloid-beta-mediated neurodegeneration. Front. Cell Dev. Biol. 2020;8:117. doi: 10.3389/fcell.2020.00117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Mehta AS, Luz-Madrigal A, Li JL, Tsonis PA, Singh A. Comparative transcriptomic analysis and structure prediction of novel Newt proteins. PLoS ONE. 2019;14:e0220416. doi: 10.1371/journal.pone.0220416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Mehta A, Singh A. Real time quantitative PCR to demonstrate gene expression in an undergraduate lab. Dros. Inf. Serv. 2017;100:5. [Google Scholar]
- 74.Mehta AS, Luz-Madrigal A, Li JL, Panagiotis TA, Singh A. Total RNA extraction from transgenic flies misexpressing foreign genes to perform Next generation RNA sequencing. Protocols. 2019;1:1–3. doi: 10.17504/protocols.io.5bng2me. [DOI] [Google Scholar]
- 75.Cutler T, et al. Drosophila eye model to study neuroprotective role of CREB binding protein (CBP) in Alzheimer's disease. PLoS ONE. 2015;10:e0137691–e0137691. doi: 10.1371/journal.pone.0137691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Steffensmeier AM, et al. Novel neuroprotective function of apical-basal polarity gene crumbs in amyloid beta 42 (Aβ42) mediated neurodegeneration. PLoS ONE. 2013;8:e78717. doi: 10.1371/journal.pone.0078717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Tare M, et al. Activation of JNK signaling mediates amyloid-ß-dependent cell death. PLoS ONE. 2011;6:e24361. doi: 10.1371/journal.pone.0024361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gogia N, et al. Inactivation of Hippo and cJun-N-terminal Kinase (JNK) signaling mitigate FUS mediated neurodegeneration in vivo. Neurobiol. Dis. 2020;140:104837. doi: 10.1016/j.nbd.2020.104837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Singh A, Kango-Singh M, Choi KW, Sun YH. Dorso-ventral asymmetric functions of teashirt in Drosophila eye development depend on spatial cues provided by early DV patterning genes. Mech. Dev. 2004;121:365–370. doi: 10.1016/j.mod.2004.02.005. [DOI] [PubMed] [Google Scholar]
- 80.Bonini NM, Leiserson WM, Benzer S. The eyes absent gene: genetic control of cell survival and differentiation in the developing Drosophila eye. Cell. 1993;72:379–395. doi: 10.1016/0092-8674(93)90115-7. [DOI] [PubMed] [Google Scholar]
- 81.Mardon G, Solomon NM, Rubin GM. dachshund encodes a nuclear protein required for normal eye and leg development in Drosophila. Development. 1994;120:3473–3486. doi: 10.1242/dev.120.12.3473. [DOI] [PubMed] [Google Scholar]
- 82.Baker NE, Mlodzik M, Rubin GM. Spacing differentiation in the developing Drosophila eye: a fibrinogen-related lateral inhibitor encoded by scabrous. Science. 1990;250:1370–1377. doi: 10.1126/science.2175046. [DOI] [PubMed] [Google Scholar]
- 83.Mlodzik M, Baker NE, Rubin GM. Isolation and expression of scabrous, a gene regulating neurogenesis in Drosophila. Genes Dev. 1990;4:1848–1861. doi: 10.1101/gad.4.11.1848. [DOI] [PubMed] [Google Scholar]
- 84.Schlichting K, Dahmann C. Hedgehog and Dpp signaling induce cadherin Cad86C expression in the morphogenetic furrow during Drosophila eye development. Mech. Dev. 2008;125:712–728. doi: 10.1016/j.mod.2008.04.005. [DOI] [PubMed] [Google Scholar]
- 85.Kumar, J. P. in In Molecular genetics of axial patterning, growth and disease in the Drosophila eye. (ed Amit and Kango-Singh Singh, Madhuri) 75–97 (Springer, 2013).
- 86.Wittkorn E, Sarkar A, Garcia K, Kango-Singh M, Singh A. The Hippo pathway effector Yki downregulates Wg signaling to promote retinal differentiation in the Drosophila eye. Development. 2015;142:2002–2013. doi: 10.1242/dev.117358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Song Z, McCall K, Steller H. DCP-1, a Drosophila cell death protease essential for development. Science. 1997;275:536–540. doi: 10.1126/science.275.5299.536. [DOI] [PubMed] [Google Scholar]
- 88.Sims RJ, 3rd, Belotserkovskaya R, Reinberg D. Elongation by RNA polymerase II: the short and long of it. Genes Dev. 2004;18:2437–2468. doi: 10.1101/gad.1235904. [DOI] [PubMed] [Google Scholar]
- 89.Boettiger AN, Levine M. Synchronous and stochastic patterns of gene activation in the Drosophila embryo. Science. 2009;325:471–473. doi: 10.1126/science.1173976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ghosh SK, Missra A, Gilmour DS. Negative elongation factor accelerates the rate at which heat shock genes are shut off by facilitating dissociation of heat shock factor. Mol. Cell. Biol. 2011;31:4232–4243. doi: 10.1128/MCB.05930-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Core LJ, Waterfall JJ, Lis JT. Nascent RNA sequencing reveals widespread pausing and divergent initiation at human promoters. Science. 2008;322:1845–1848. doi: 10.1126/science.1162228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Guenther MG, Levine SS, Boyer LA, Jaenisch R, Young RA. A chromatin landmark and transcription initiation at most promoters in human cells. Cell. 2007;130:77–88. doi: 10.1016/j.cell.2007.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Muse GW, et al. RNA polymerase is poised for activation across the genome. Nat. Genet. 2007;39:1507–1511. doi: 10.1038/ng.2007.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Gilmour DS, Lis JT. RNA polymerase II interacts with the promoter region of the noninduced hsp70 gene in Drosophila melanogaster cells. Mol. Cell Biol. 1986;6:3984–3989. doi: 10.1128/mcb.6.11.3984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Chen K, et al. A global change in RNA polymerase II pausing during the Drosophila midblastula transition. Elife. 2013;2:e00861. doi: 10.7554/eLife.00861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Jennings BH. Pausing for thought: disrupting the early transcription elongation checkpoint leads to developmental defects and tumourigenesis. BioEssays. 2013;35:553–560. doi: 10.1002/bies.201200179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Nguyen D, Fayol O, Buisine N, Lecorre P, Uguen P. Functional Interaction between HEXIM and Hedgehog signaling during drosophila wing development. PLoS ONE. 2016;11:e0155438. doi: 10.1371/journal.pone.0155438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Jennings BH, et al. Locus-specific requirements for Spt5 in transcriptional activation and repression in Drosophila. Curr. Biol. 2004;14:1680–1684. doi: 10.1016/j.cub.2004.08.066. [DOI] [PubMed] [Google Scholar]
- 99.Peters MA. Patterning the neural retina. Curr. Opin. Neurobiol. 2002;12:43–48. doi: 10.1016/s0959-4388(02)00288-x. [DOI] [PubMed] [Google Scholar]
- 100.Rai, A., Narwal, S., Kanodia, H. & Tare, M. in Molecular Genetics of Axial Patterning, Growth and Disease in the Drosophila Eye Vol. II (eds Amit Singh, and, & Madhuri Kango-Singh) Ch. 2, 343–358 (Springer International Publishing, 2020).
- 101.Bier E. Drosophila, the golden bug, emerges as a tool for human genetics. Nat. Rev. Genet. 2005;6:9–23. doi: 10.1038/nrg1503. [DOI] [PubMed] [Google Scholar]
- 102.Lobov IB, et al. WNT7b mediates macrophage-induced programmed cell death in patterning of the vasculature. Nature. 2005;437:417–421. doi: 10.1038/nature03928. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.