

Abstract
Growth hormone-releasing hormone (GHRH) regulates the secretion of growth hormone that virtually controls metabolism and growth of every tissue through its binding to the cognate receptor (GHRHR). Malfunction in GHRHR signaling is associated with abnormal growth, making GHRHR an attractive therapeutic target against dwarfism (e.g., isolated growth hormone deficiency, IGHD), gigantism, lipodystrophy and certain cancers. Here, we report the cryo-electron microscopy (cryo-EM) structure of the human GHRHR bound to its endogenous ligand and the stimulatory G protein at 2.6 Å. This high-resolution structure reveals a characteristic hormone recognition pattern of GHRH by GHRHR, where the α-helical GHRH forms an extensive and continuous network of interactions involving all the extracellular loops (ECLs), all the transmembrane (TM) helices except TM4, and the extracellular domain (ECD) of GHRHR, especially the N-terminus of GHRH that engages a broad set of specific interactions with the receptor. Mutagenesis and molecular dynamics (MD) simulations uncover detailed mechanisms by which IGHD-causing mutations lead to the impairment of GHRHR function. Our findings provide insights into the molecular basis of peptide recognition and receptor activation, thereby facilitating the development of structure-based drug discovery and precision medicine.
Subject terms: G protein-coupled receptors, Cryoelectron microscopy, Cryoelectron microscopy
Growth hormone-releasing hormone (GHRH) controls metabolism and tissue growth through binding to the cognate receptor (GHRHR). Here authors report the structure of the human GHRHR bound to its endogenous ligand and the stimulatory G protein which reveals a characteristic hormone recognition pattern of GHRH by GHRHR.
Introduction
Class B G-protein-coupled receptors (GPCRs) are key players in hormonal homeostasis and important drug targets for endocrinal and neuronal disorders. Growth hormone-releasing hormone receptor (GHRHR), a prototypical class B GPCR, is expressed by somatotropic cells of the pituitary gland. Activation of GHRHR by GHRH, a 44-amino acid peptide released by the hypothalamus1,2, results in the secretion and production of growth hormone (GH) through cyclic adenosine monophosphate (cAMP)-dependent pathways3. Numerous studies demonstrated that GHRH exerts a variety of bioactivities due to its wide distribution and autocrine/paracrine mechanisms4,5. Therefore, GHRH and its analogs, including tesamorelin, MR-409, JI-38, and MIA-690, have been developed as potential therapeutic agents to treat diabetes, cancers, and cardiovascular diseases5–9.
Like other class B GPCRs, GHRHR consists of an extracellular domain (ECD) and a seven-transmembrane helix domain (7-TMD)10,11. Recently published cryo-electron microscopy (cryo-EM) structures of class B GPCRs bound to a Gs heterotrimer protein include parathyroid hormone receptor 1 (PTH1R), glucagon-like peptide-1 receptor (GLP-1R), calcitonin receptor, calcitonin gene-related peptide receptor (CGRPR), two subtypes of corticotrophin-releasing factor (CRF1R and CRF2R), and adrenomedullin receptors (AM1R and AM2R), as well as pituitary adenylate cyclase-activating polypeptide (PACAP) type I receptor (PAC1R) and vasoactive intestinal polypeptide receptor (VIP1R), revealing a common mode of ligand-induced receptor activation10–19. The C-terminal α helix of peptide ligand recognizes and binds to the ECD, thereby allowing its N-terminus to interact with the extracellular TM core. This is followed by a major conformational change that involves a large kink at the TM6 to open the intracellular face for G protein coupling15. However, ligand-binding specificity and roles of ECD in receptor activation vary widely among class B GPCRs due to diverse amino acid sequences of both peptidic ligands and receptors15,20.
Here, we employed the single-particle cryo-EM approach to determine the near-atomic resolution structure of the human GHRHR bound to GHRH in complex with a heterotrimeric Gs protein. Together with functional studies and molecular dynamics (MD) simulations, our results provide key insights into the structural basis of ligand recognition, receptor activation, and isolated growth hormone deficiency (IGHD) causing mechanism related to GHRHR, thereby offering a template for rational design of drugs against this receptor.
Results
Structure determination of the GHRH–GHRHR–Gs complex
We developed a NanoBiT tethering strategy to stabilize the assembly of a GHRH–GHRHR–Gs complex for cryo-EM studies, overcoming the lack of stability of the above complex (Supplementary Fig. 1 and Supplementary Table 1), as it has been used for the VIP1R-Gs complex19. Using this approach, we were able to obtain a GHRH-GHRHR–Gs complex with improved homogeneity and stability (Supplementary Fig. 2). The GHRH–GHRHR–Gs complex was vitrified and cryo-EM images were collected under a Titan Krios microscope equipped with K2 summit direct detector. The structure of GHRH–GHRHR–Gs complex was determined from 307,018 particles to an overall resolution of 2.6 Å (Supplementary Figs. 3 and 4). The resulting model contains 28 residues of GHRH (residues 1–28), Gαβγ subunits except the α-helical domain (AHD) of Gαs, and GHRHR residues from 119 to 394. Besides, the ECD region of GHRHR was not resolvable with this limited dataset, perhaps reflecting its highly dynamic and conformationally flexible property when bound to GHRH. We rigid-body fitted the GHRHR ECD (residues 25–118) crystal structure (PDB accession: 2XDG) to the low-pass filtered map (Fig. 1b). The majority of amino acid side chains were well resolved in the final model (Supplementary Fig. 5), which was refined against the EM density map (Fig. 1a) with excellent geometry (Supplementary Table 2). Owing to the high-resolution map, we identified one water molecule in the orthosteric binding site, and two water molecules in the G protein engaging pocket. Akin to cryo-EM structure of PTH1R–Gs complex, the TMD of GHRHR is surrounded by annular detergent micelle with a diameter of 10 nm, mimicking the natural phospholipid bilayer. Within the micelle, one bound cholesterol and two lipids are also clearly visible in the cryo-EM map.
Fig. 1. The overall cryo-EM structure of GHRH–GHRHR–Gs complex.
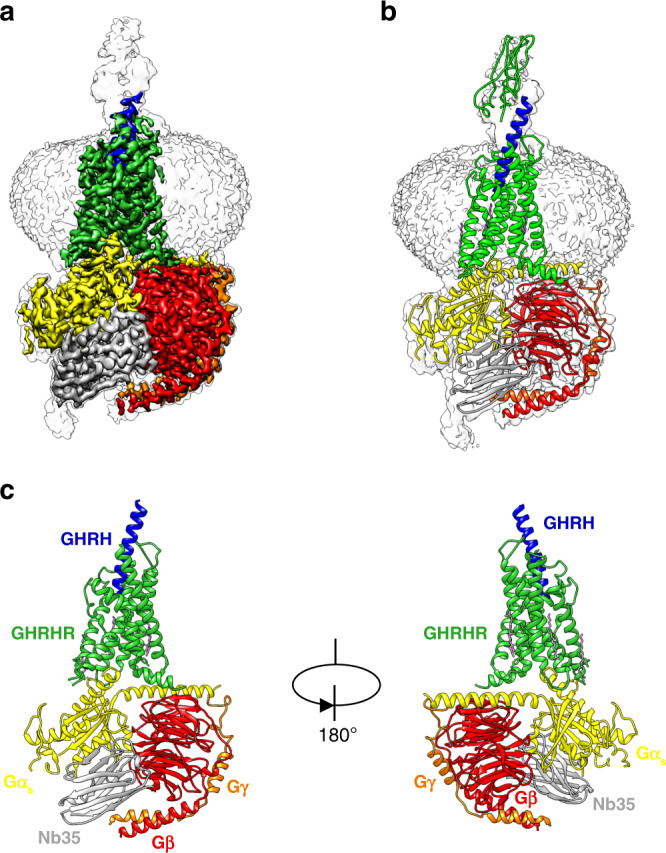
a Cryo-EM density map that illustrates the GHRH–GHRHR–Gs complex and the disc-shaped micelle. The unsharpened cryo-EM density map at the 0.005 threshold shown as light gray surface indicates a micelle diameter of 10 nm. The colored cryo-EM density map is shown at 0.028 threshold. b GHRH–GHRHR–Gs complex model and GHRHR ECD crystal structure model docked into the cryo-EM map. c Cartoon representation of the GHRH–GHRHR–Gs complex is shown with annular lipids in purple stick representation. Lime green, GHRHR; blue, GHRH; yellow, Gs Ras-like domain; red, Gβ; orange, Gγ; gray, Nb35; plum, lipid, and cholesterol.
Molecular recognition of GHRH by GHRHR
In the complex structure, GHRH adopts an α-helical configuration when engaged with GHRHR. Compared to GLP-1 (ref. 10) and long-acting parathyroid hormone analog (LA-PTH)15, GHRH binds to the GHRHR TMD through a more extensive and continuous network of interactions involving all the extracellular loops (ECLs), all the TM helices except TM4, and the linker connecting ECD and TMD. The N-terminus of GHRH inserts deeply into the TMD core and engages an extensive set of receptor-specific interactions (Fig. 2a–c and Supplementary Table 3). Interestingly, Tyr1P locates in the equivalent position of the second residue in other class B GPCR peptidic agonists, such as exendin-P5 (ExP5) and LA-PTH, where their first residues of the side chain have different orientations (Fig. 2b). The hydroxyl group of Tyr1P forms hydrogen bonds with H2103.37b (class B GPCR numbering in superscript)21 and van der Waals interactions with T2133.40b and W2825.36b, whereas the main chain NH of Tyr1P forms hydrogen bond with R3577.38b of TM7 and contacts a water molecule that connects with N3466.57b of TM6-ECL3 hinge, possibly stabilizing the ECL3 in an active state. This observation is consistent with our observation that impairing these contacts dramatically decreased the potency of GHRH in stimulating cAMP accumulation (Fig. 2d–f and Supplementary Table 4). The most conserved Asp/Glu at position 3 across peptide hormones of the glucagon receptor (GCGR) subfamily, Asp3P in the case of GHRH, forms salt bridges with K1822.67b, which is further strengthened by a polar network composed of Thr7P, Y1331.43b, and D1832.68b. From evolutional biology perspective, both Asp3P and K1822.67b are fully conserved for GHRH and GHRHR from dozens of species (Supplementary Fig. 6a–b). Indeed, D3PA and K1822.67bA diminished the potency of GHRH by ~4- and 200-fold (Fig. 2e and Supplementary Fig. 6c), respectively. The combined structural, pharmacological, and evolutional investigations point to a crucial role of Asp3P in peptide binding and receptor activation. Phe6P is another conserved residue and contributes extensive contacts with surrounding residues of GHRHR, including Pi–Pi stacking with F1261.36b and Y1331.43b, and hydrophobic contacts with V1291.39b and L3627.43b, where these four residues are either fully conserved or physiochemically similar among GHRHRs of different species. By forming salt bridges with D2745.52b and hydrogen bonds with backbone atoms of H194ECL1 and A271ECL2, fully conserved Arg11P greatly stabilizes the ECL1–ECL2 interface relative to the conserved TM3–ECL2 disulfide bond (Cys3.29b‒CysECL2) of class B GPCRs. This interface is further stabilized by close contacts between ECL1 and GHRH (T-shape Pi stacking between Tyr10P and F187ECL1, and hydrogen bonds between Ser18P and D193ECL1), as well as between ECL2 and GHRH (hydrogen bonds between Asn8P and D274). For residues like Ala2P and Ile5P, they contribute hydrophobic interactions with L3627.43b and D350ECL3, which are conserved among different species. The remaining residues, including Ala4P, Ser9P, and Gly15P, are variable and receptive to substitution by Ala, Gly, Cys, or Ser, indicating their role of structural complementarily. Amino acids of the peptide after 20 may have rich interactions with the ECD as indicated by the model, in which the ECD was docked in the EM map as a rigid body (Fig. 1b). Indeed, cAMP signaling was nearly abolished in HEK 293 T cells expressing a truncated ECD construct, i.e., GHRHR(119–423), suggesting an essential role of ECD in GHRH recognition and receptor activation (Supplementary Fig. 7a). In our 1 μs MD simulation, the ECD of GHRHR was able to twist around the GHRH helix, while the TMD core is quite stable in a single dominant conformational state as seen previously with PTH1R15. Specifically, massive hydrophobic contacts between GHRH (R20, L22, L23, and M27) and GHRHR ECD (L34, L62, F82, Y108, P109, and L118), as well as several hydrogen bonds (Q24 of GHRH and C112 of GHRHR), were observed, which may stabilize the binding of GHRH (Supplementary Fig. 7b). These results are consistent with the findings on the same receptor reported in the literature22,23. In fact, such a dynamic and flexible feature of ECD could also be found among other class B GPCRs, such as PTH1R15, GLP-1R10, and VIP1R19, where the conversion from a closed conformation in the apo-state to an extended open conformation is required for activation (Supplementary Fig. 7c).
Fig. 2. Molecular recognition of GHRH by GHRHR and comparison of that with LA-PTH–PTH1R and GLP-1–GLP-1R.

a Detailed interaction between GHRH and the TMD pocket of GHRHR with hydrogen bonds shown as dotted lines. b, c Pairwise comparison of GHRH bound to GHRHR with GLP-1 and LA-PTH in complex with their corresponding receptors, showing the relative positions of peptide ligands. Lime green, GHRHR; blue, GHRH; red, PTH1R and LA-PTH; purple, GLP-1R and GLP-1. d–f The effects of mutation in the ligand-binding pocket on cAMP accumulation. cAMP levels were measured in wild-type (WT) receptor and alanine mutants in ECL1, 2, and 3 (d), TM1, 2, and 3 (e), and TM5, 6, and 7 (f). cAMP signals were normalized to the maximum response of WT and concentration–response curves were analyzed using a three-parameter logistic equation. Data shown are means ± S.E.M. of at least three independent experiments (n = 3–5), conducted in triplicate. Source data are provided as a Source data file.
Unlike the helical extensions found in ECL1 of GCGR, GLP-1R, CRF1R, CRF2R, AM1R, and AM2R, and the unstructured ECL1 of PTH1R in active state, the ECL1 of GHRHR stretches around GHRH to form broad interactions. Consistently, disruption of GHRH-ECL interaction by F187ECL1A and C195ECL1A reduced GHRH potency by ~5- and 100-fold, respectively (Fig. 2d). Coupled with the conformational change from β-hairpin to α-helix in ECL1 of GCGR upon activation24, and the limited impact on Gs-mediated cAMP signaling by alanine mutation on ECL1 of GLP-1R25, these results demonstrate the dynamic nature and diversified roles of ECL1 in class B GPCRs.
Active structure of GHRHR
The overall arrangement of the GHRH–GHRHR–Gs complex is highly similar to GLP-1–GLP-1R–Gs, glucagon–GCGR–Gs, and LA-PTH–PTH1R–Gs complexes10,14,15. Superimposition of TMs shows that active GHRHR, GLP-1R, and PTH1R share similar folds with respect to the global conformation of the 7-TM bundle, as well as a similar organization of the extracellular end of TM6 and TM7 to accommodate peptide ligand. Except ICL3, other loop regions of the receptor are visible due to the high-quality cryo-EM density map, although these loops are relatively dynamic compared to the TMD bundle (Fig. 1c and Supplementary Fig. 5).
Comparison of the GHRHR complex with inactive class B GPCR structures26,27, such as GLP-1R and GCGR, suggests that the most obvious conformational transformation is located at TM6 (Fig. 3a–c), where there is a large outward movement at the cytoplasmic face in the activated GHRHR structure upon coupling to Gs. The TM6 outward displacement is correlated with the kink at the conserved Pro6.47b-X-X-Gly6.50b motif10,13,16,28.
Fig. 3. Structural comparison of active GHRHR with other class B GRCRs in active, intermediate, and inactive states.

a–c Comparison of active GHRHR with inactive (a), agonist-bound intermediate (b), and both agonist- and G-protein-bound active (c) class B GPCRs. d, e Comparison of ligand conformations in G-protein-bound class B GPCRs; close-up view of the N-terminus (d), middle region (e), and C-terminus (e) of bound peptides in class B GPCRs.
Diversified peptide binding modes
Different from the ligand-binding pockets for small molecules in class A GPCRs, the peptide-binding pockets in class B GPCRs are larger and more extended, involving ECD, ECLs, and TMs. As shown in Fig. 3d, e, class B GPCR agonists adopt diversified conformations and orientations especially in the N- and C-terminals. Contrary to the single continuous helix observed in GHRH, GLP-1, glucagon, ExP5, PACAP38, and LA-PTH, the N-termini of AM, AM2, CGRP, CRF, and UCN1 looping back between TM5 and TM6, while AM, AM2, and CGRP also have a large kink at the C-terminal portion. Comparison of peptide recognition modes in the GCGR subfamily reveals that binding specificity mainly resides in three segments: N-terminus (first three residues in GHRH), middle region (6th to 11th residues), and C-terminal portion (Fig. 3d, e and Supplementary Fig. 8).
The orientation of the residues at N-terminus are different: the side chain of Tyr1P in GHRH and His1P in glucagon directly face TM3 forming hydrogen bonds with H2103.37b and I2353.40b; the side chain of His7P in GLP-1 directly faces TM5 and has cation–Pi interaction from upward R299ECL2; and the first two residues of LA-PTH (Ala1PVal2P) and ExP5 (Glu1PLeu2P) insert into the cleft between TM5 and TM6. Meanwhile, the orientation of highly conserved Glu/Asp3P is significantly different (Supplementary Fig. 8d, e). These observations suggest the complexity of signal initiation.
The middle region of peptides has rich contacts with ECL1 and structurally flexible ECL2 in a peptide-dependent manner (Fig. 3e and Supplementary Fig. 8f). Resulted from longer α-helical extensions of TM2, the elevated ECL1 of GLP-1R and GCGR contribute additional contacts with the peptide C-terminal region. For receptors whose residues at 45.52 (adopted from GPCRdb numbering29, the second residue after the family-wide conserved cysteine, 45.50, in ECL2) are Glu/Asp (e.g., GHRHR and PTH1R), ECL2 shows compact contacts with peptides through direct salt bridge (GHRHR: Arg11P and D27445.52b), or intra-receptor salt bridge (PTH1R: K2402.67b and D35345.52b). In the case of GCGR/GLP-1R whose residues at 45.52 are Thr, ECL2 of GCGR forms multiple hydrogen bonds with glucagon via Gly4P, Ser8P, and Ser11P, and ECL2 of GLP-1R forms polar interaction with multiple serines (Ser14P, Ser17P, and Ser18P) of GLP-1 via T29845.52b and electrostatic interaction with His7P via inward chain of R299ECL2. Remarkably, the binding of ExP5 induces reorganization of ECL2–peptide interface, where R299ECL2 rotates outward and forms salt bridges with Glu16P.
G protein coupling by GHRHR
Like other class B GPCRs, the outward movement of the cytoplasmic end of TM6, and concomitant shift of TM5 and ICL3 of GHRHR form a cavity to accommodate α5 helix of Gαs. This process also involves TM2, TM3, ICL2, and helix 8. The interface residues in this cavity are highly conserved among class B GPCRs, and the arrangement of GHRHR–Gs complex is also similar to other class B GPCR–Gs complexes, and follows a common mechanism of G protein coupling. The high-resolution cryo-EM map allows us unambiguously to assign the water molecules between the interface of GHRHR and Gαs. Comparing with other class B GPCR–Gs complexes in which α5 helix of Gαs loosely interacts with the receptor TM7–H8 hinge (Supplementary Fig. 9a, b), we found two water molecules in the GHRH–GHRHR–Gs complex establishing extensive polar interaction network connecting the C-terminus of Gαs to the TM7–H8 hinge of GHRHR, thereby causing considerable conformational changes compared to GLP-1R–Gs and PTH1R–Gs structures (Supplementary Fig. 9c, d). Specifically, the crevice between the TM7–H8 hinge and TM6 in GHRHR–Gs complex is broader than that in the PTH1R–Gs and GLP-1R–Gs complexes, which may be responsible for G protein moving 3 Å toward the cytoplasmic core of the TMD bundle (Supplementary Fig. 9a, b).
Besides contacting the cytoplasmic cavity of TMs 2, 3, 6, and 7 induced by the opening of the cytoplasmic half of TM6 via Gαs, heterotrimeric Gs protein also establishes an electrostatic interaction network with ICL1 and H8 via Gβ (Supplementary Fig. 9e). Specifically, four residues (D312 of Gβ, R15612.49b of ICL1, and E3868.53b and R3898.56b of H8) are clustered at the interface of GHRHR–Gβ by forming salt bridges. Such an organization is conserved across class B GPCRs, evidenced by the highly conserved residues and previous studies on GLP-1R and PTH1R. Notably, an additional component, Arg/Lys8.60b, from GLP-1R, GCGR, and PTH1R may join and further stabilizes the network, while G3938.60b of GHRHR has neglectable contacts with Gβ. Indeed, G3938.60bR enhances the potency of GHRH, probably by strengthening the electrostatic interactions between GHRHR and Gs, whereas the diminished potency of a double mutant (R15612.49bA/R3898.56bA) is likely resulted from the disruption of the electrostatic interaction network (Supplementary Fig. 9f).
Implication of disease-causing mutations
Based on the structure information, 25 missense mutants (21 were reported to be linked with IGHD previously; Fig. 4a and Supplementary Table 5) were made and assessed for their effects on cAMP signaling and β-arrestin2 recruitment. Four of them were further analyzed by MD simulations. It is established that a common fold of three-layer α-β-β/α architecture30,31 across class B GPCRs is stabilized by three conserved interlayer disulfide bonds (C41‒C64, C55‒C96, and C78‒C112) and the salt bridge/pi stacking within the family-wide conserved D-W-R/K-W motif (D60-W65-R94-W101) in GHRHR, that connect two β-sheets to directly interact with the peptide ligands. Presumably, naturally occurring mutations may disrupt the disulfide bonds (C64G and C112Y) or the conserved D-W-R/K-W motif (R94Q/L/W), thereby destabilizing the stability of ECD and preventing GHRH recognition. Indeed, our MD simulation and functional studies suggest that the IGHD-associated mutation R94Q32 breaks the salt bridge with D60, increases the flexibility of the ECD, decreases the area of GHRH–GHRHR interface, and reduces GHRH-induced cAMP accumulation (Fig. 4b and Supplementary Fig. 10a). We then examined if mutations on D60 could also eliminate the electrostatic interaction between D60 and R94 to exhibit similar functional and phenotypical outcomes. D60G is known for the little mouse phenomenon—the first naturally occurring animal model of inherited autosomal recessive GH-deficient dwarfism33. Like R94Q, it reduces GHRH binding affinity34,35, diminishes its potency on cAMP accumulation (Fig. 4b), and weakens GHRH binding in MD simulations (Supplementary Fig. 10a).
Fig. 4. Disease-causing missense mutations of GHRHR.

IGHD refers to conditions of growth hormone deficiency that are not necessarily associated with other pituitary hormone deficiencies or with an organic lesion. While there is no single cause identified for IGHD, defects in growth hormone, GHRHR and GH1 genes are implicated as causative factors55. Of which, naturally occurring missense mutations of GHRHR have been studied extensively56. a Naturally occurring mutations in the ECD, ligand-binding pocket, and G-protein-coupling region are colored cyan, marine, and magenta, respectively. The central region connecting the ligand-binding pocket and G-protein-coupling region is colored orange. b, c Functional assessment of effects of disease-causing GHRHR mutations on Gs-mediated cAMP accumulation (b) and β-arrestin2 recruitment (c), cAMP accumulation assays and β-arrestin2 recruitment were performed in HEK 293 T cells and data shown are means ± S.E.M. of at least three independent experiments (n = 3–8), performed in quadruplicate or duplicate, respectively. Statistically significant differences were determined with a two-tailed Student’s t test. *P < 0.05, **P < 0.01 vs. wild type (WT); AUC area-under-the-curve. Source data are provided as a Source data file.
In the peptide-binding pocket, R357C was shown to loosen the compact GHRHR contacts in MD simulation (Supplementary Fig. 10b) and reduce GHRH potency by 1000-fold (Fig. 4b), consistent with the decreased potency of R357A and D350A mutants (Fig. 2d). Different from Ala/Val/Val in GLP-1R/PTH1R/GCGR, the residue at 3.41 of GHRHR located at the bottom of the ligand-binding pocket is M214, whose long side chain points to a hydrophobic cleft between TM4 and TM5. Another mutation associated with IGHD, M214V, was found to decrease GHRH potency by tenfold and selectively reduce β-arrestin2 recruitment (Fig. 4b).
The connector region is known for the sharp kink in the middle of TM6 around Pro6.47b-X-X-Gly6.50b. It appears that P3366.47bL hampered the sharp kink upon receptor activation evidenced by a reduced EC50 value for cAMP signaling (Fig. 4b). As a comparison, S1401.50bP eliminated the hydrogen bonds between TM1 and TM7, increased the flexibility of TMD region and the bound GHRH, and reduced cAMP accumulation by over 7000-fold (Fig. 4b and Supplementary Fig. 10b). A1762.61b oriented toward TM1, and contributed close hydrophobic contacts with H1371.47b, I1411.51b, and I1732.58b. Substitution of A1762.61b with a larger hydrophobic valine at this position reduced GHRH potency (tenfold) and β-arrestin2 recruitment (39%).
Within the G-protein-coupling region, N1622.47bI, N1622.47bD, and H1652.50bQ were previously proposed to be deleterious36—a view that was verified experimentally in this study (Fig. 4b) As shown in the structure (Fig. 4a), N162 formed a hydrogen bond with the backbone of L157ICL1, H1652.50b from HETX motif10,14 played a crucial role in receptor activation, while H165Q might have directly altered receptor–G protein interface and abolished G protein coupling.
Discussion
In summary, the high-resolution structure of the GHRHR–Gs complex, and mutational studies provide a basis for GHRH recognition and receptor activation. These results also unveiled three distinct malfunctioning mechanisms of GHRHR signaling: (i) impaired peptide binding in the ECD (D60G and R94Q); (ii) reduced ligand recognition (R357C)/G protein coupling (N162I/D and H165Q via functional validation); and (iii) disrupted signal propagation in the connector region (S140P). Together, our work solved a longstanding puzzle of IGHD-causing mutations, which result in the impairment of GHRHR-mediated signaling at the ECD or TMD, singly or in combination.
Methods
Cell culture
Spodoptera frugiperda (Sf9) insect cells (Expression Systems) were grown in ESF 921 serum-free medium (Expression Systems) at 27 °C and 120 r.p.m. HEK 293 T cells were purchased from the Cell Bank at the Chinese Academy of Sciences, cultured in Dulbecco’s modified Eagle’s medium (DMEM; Life Technologies) supplemented with 10% fetal bovine serum (Gibco) and maintained in a humidified chamber with 5% CO2 at 37 °C.
Constructs of GHRHR and Gs heterotrimer
For structural studies, wild-type (WT) human GHRHR DNA (Genewiz) was cloned into pFastBac vector (Invitrogen) with its native signal sequence (M1-G22) replaced by the hemagglutinin (HA) signal peptide. Eighteen amino acids (A406–C423) were truncated at the C-terminus and LgBiT subunit (Promega) was fused with a 15-amino acid polypeptide (GSSGGGGSGGGGSSG) linker at the C-terminus followed by a Tev protease cleavage site and a double maltose-binding protein (MBP) tag to facilitate expression and purification. A dominant-negative human Gαs (DNGαs) was generated by site-directed mutagenesis as previously described to stabilize the interaction with the βγ subunits11,37. Rat Gβ1 with an N-terminal 10× His-tag was fused with a SmBiT38 (peptide 86, Promega) subunit by the 15-amino acid polypeptide linker at its C-terminus. Human DNGαs, rat Gβ1, and bovine Gγ2 were cloned into pFastBac vector, respectively. In addition to clone the constructs into the pBiT vector (Promega) for NanoBiT assay, they all contained an N-terminal Flag tag (DYKDDDD) proceeded by a HA signal sequence, and were cloned into the pcDNA3.1 vector (Invitrogen) for functional studies.
To obtain a GHRHR–Gs complex with good homogeneity and stability, the human GHRHR was modified by replacing the N-terminal native signal peptide with HA sequence. In addition, 18 residues (A406–C423) were truncated at the C-terminus, followed by an LgBiT subunit and a double MBP affinity tag to improve protein yield and stability (Supplementary Fig. 2a). These modifications did not affect ligand binding and receptor activation (Supplementary Fig. 2b, c and Supplementary Table 1). In addition, a DNGαs11,37, His10-Gβ1-peptide 86 and Gγ2 were co-expressed with GHRHR(23-405)-15AA-LgBiT-2MBP in insect cells. Formation of GHRHR–Gs complex on the membrane was stimulated with an excess amount of GHRH, and in the presence of Gα- and Gβ-binding nanobody 35 (Nb35)39 (Supplementary Fig. 2d–e).
Expression and purification of GHRHR–Gs complex
GHRHR-15AA-LgBiT-2MBP or GHRHR-2MBP, DNGαs, His10-Gβ1-peptide 86, and Gγ2 recombinant baculoviruses were prepared using Bac-to-Bac Baculovirus Expression System (Invitrogen) severally. Sf9 insect cells were grown to a density of 3 × 106 cells per mL and then coinfected with four separate viruses at a ratio of 1:3:3:3 for GHRHR, DNGαs, Gβ1, and Gγ2. Cells were harvested by centrifugation 48 h post infection and pellets were stored at −80 °C until use.
The cell pellets were thawed on ice and lysed in a buffer containing 20 mM HEPES, pH 7.5, 100 mM NaCl, 10% (v/v) glycerol, 10 mM MgCl2, 5 mM CaCl2, 1 mM MnCl2, 100 μM TCEP, and supplemented with EDTA-free protease inhibitor cocktail (Bimake) by dounce homogenization. The complex formation was initiated by addition of 10 μM GHRH (GL Biochem), 10 μg/mL Nb35, 25 mU/mL apyrase (NEB), and the lysate was incubated for 1.5 h at room temperature (RT). The membrane was further solubilized by 0.5% (w/v) lauryl maltose neopentyl glycol (LMNG; Anatrace) and 0.1% (w/v) cholesterol hemisuccinate (CHS; Anatrace) for 2 h at 4 °C. After centrifugation at 65,000 × g for 30 min, the supernatant was isolated and incubated with amylose resin (NEB) for 2 h at 4 °C. The resin was then collected by centrifugation at 600 × g for 10 min, loaded into a gravity flow column (Sangon Biotech), and first washed with five column volumes of buffer containing 20 mM HEPES, pH 7.5, 100 mM NaCl, 10% (v/v) glycerol, 5 mM MgCl2, 1 mM MnCl2, 25 μM TCEP, 1 μM GHRH, 0.1% (w/v) LMNG, and 0.02% (w/v) CHS, followed by washing with 15 column volumes of buffer containing 20 mM HEPES, pH 7.5, 100 mM NaCl, 10% (v/v) glycerol, 5 mM MgCl2, 1 mM MnCl2, 25 μM TCEP, 1 μM GHRH, 0.03% (w/v) LMNG, 0.01% (w/v) glyco-diosgenin (Anatrace), and 0.008% (w/v) CHS. The protein was then incubated overnight with His-tagged Tev protease (customer-made) on the column to remove the C-terminal 2MBP-tag in the buffer above. The flow through was collected next day and concentrated with a 100 kDa molecular weight cut-off concentrator (Millipore). Concentrated GHRH–GHRHR–Gs–Nb35 complex was loaded onto a Superdex 200 increase 10/300 GL column (GE Healthcare) with running buffer containing 20 mM HEPES, pH 7.5, 100 mM NaCl, 10 mM MgCl2, 100 μM TCEP, 5 μM GHRH, and 0.001% digitonin (Anatrace). The fractions for monomeric complex were collected and concentrated to 20–30 mg/mL for EM examination.
Expression and purification of Nb35
Nb35 with a C-terminal 6× His-tag was expressed in the periplasm of Escherichia coli BL21 (DE3) cells, extracted and purified by nickel affinity chromatography as previously described36. Eluted protein was concentrated using a 10 kDa molecular weight cut-off concentrator (Millipore) and loaded onto a HiLoad 16/600 Superdex 75 column (GE Healthcare), with running buffer containing 20 mM HEPES, pH 7.5, and 100 mM NaCl. The monomeric fractions were pooled and supplemented with 30% (v/v) glycerol. Purified Nb35 was finally flash frozen in liquid nitrogen and stored in −80 °C.
Cryo-EM data acquisition and image processing
The purified GHRH–GHRHR–Gs complex (3.5 μL) at a concentration of 22 mg/mL was applied to glow-discharged holey carbon grids (Quantifoil R1.2/1.3, 200 mesh), and subsequently vitrified using a Vitrobot Mark IV (ThermoFisher Scientific). Cryo-EM images were collected on a Titan Krios equipped with a Gatan K2 Summit direct electron. The microscope was operated at 300 kV accelerating voltage, at a nominal magnification of 29,000× in counting mode, corresponding to a pixel size of 1.014 Å. In total, 3813 image stacks were obtained at the dose rate of ~7.8 electrons per Å2 per second with a defocus range of −0.5 to −2.5 μm. The total exposure time was set to 8 s with intermediate frames recorded every 0.2 s, resulting in an accumulated of dose of 62 electrons per Å2.
Dose-fractionated image stacks were subjected to beam-induced motion correction and dose-weighting using MotionCor2.1 (ref. 40). A sum of all frames, filtered according to the exposure dose, in each image stack was used for further processing. Contrast transfer function parameters for each micrograph were determined by Gctf v1.06 (ref. 41). Further data processing was performed in RELION-3.0-beta2 (ref. 42). Particle selection, two-dimensional classification, and the first round of three-dimensional classification were performed on a binned dataset with a pixel size of 2.028 Å.
Auto-picking yielded 2,586,606 particle projections that were subjected to reference-free two-dimensional classification to discard false-positive particles or particles categorized in poorly defined classes, producing 1,456,108 projections for further processing. This subset of particle projections was subjected to consecutive rounds of three-dimensional classification with a pixel size of 2.028 Å. The map of PTH1R–Gs complex (EMDB-0410) low-pass filtered to 40 Å was used as an initial reference model for two rounds of three-dimensional classification, resulting in two subsets accounting for 481,220 projections that showed better EM densities. Further three-dimensional classifications focusing on the complex, with the exception of AHD of Gαs, produced one good subset with higher resolution, which was subsequently subjected to three-dimensional refinement and Bayesian polishing. The final refinement with frames 1–25 generated a map with an indicated global resolution of 2.6 Å, with 307,018 projections at a Fourier shell correlation of 0.143. Local resolution was determined using the Bsoft package with half maps as input maps43.
Model building and refinement
The structure of the LA-PTH–PTH1R–Gs complex was used as an initial template for model building. Lipid coordinates and geometry restraints were generated using phenix.elbow. Models were docked into the EM density map using UCSF Chimera44. This starting model was then subjected to iterative rounds of manual adjustment and automated refinement in Coot45 and Phenix46, respectively. The final refinement statistics were validated using the module “comprehensive validation (cryo-EM)” in PHENIX. Structural figures were prepared in Chimera, Chimera X, and PyMOL (https://pymol.org/2/). The final refinement statistics are provided in Supplementary Table 2.
NanoBiT assay
HEK 293 T cells were transiently transfected with GHRHR-LgBiT, Gαs, Gβ1-SmBiT (peptide 86 or 114), and Gγ2 48 h before assaying in a mass ratio of 1:1:1:1. Twenty-four hour post transfection, cells were digested and seeded into 96-well white plate (PerkinElmer) at optimal density (30,000 cells/well) and the medium was replaced with Opti-MEM (Gibco) 3 h prior to assaying. After equilibration to ambient temperature, the Nano-Glo Live Cell Substrate (Promega) was 20-fold diluted to create a 5× stock and mixed with cell culture medium to a final 1× concentration, then luminescence signal was measured using an EnVision Multimode Plate Reader (PerkinElmer). Following the baseline reading at 10 s interval for 2.5 min, GHRH was diluted with Opti-MEM and added to a final concentration of 10 nM, and reading continued at 10 s intervals for 10 min. RLU time-course response curves were normalized to that of background and calculated as percentage of the baseline value. Interaction intensity of the complex was expressed as area-under-the-curve (AUC) across the full kinetic trace (0–10 min).
Negative-stain EM
Uranyl formate (0.75% (w/v), Electron Microscopy Sciences) solution was prepared as previously described47. Copper grids (300 mesh) coated with carbon film (Electron Microscopy Sciences) were glow charged, using PELCO easiGlowTM Glow Discharge Cleaning System (Ted Pella Inc.) for 25 s at 25 mA. Purified protein samples (4 μL, 0.01 mg/mL) were applied to glow-charged holey grids for 1 min and then blotted off using filter paper. Uranyl formate solution (0.75%, 5 μL) was added to the grid surface twice: the first application was blotted off immediately and the second was retained for 1 min before blotting with filter paper. The stained girds were loaded into a Tecnai G2 Spirit transmission electron microscope (Thermo FEI) operated at 120 kV. Negative staining images were acquired at a magnification of 57,000× within a −1.5 to −2.0 μm defocus range.
Dynamic light scattering
Dynamic light scattering determines size and size distribution by measuring the rapid changes in laser light intensity of molecules or particles in solution. Briefly, freshly purified protein samples (15 μL each) were concentrated to 1 mg/mL and loaded into DynaPro NanoStar (Wyatt Technology) to measure time-dependent fluctuations of scattered intensity at 30 °C. Data were analyzed using the dynamics software supplied with the instrument.
cAMP accumulation assay
GHRH-stimulated cAMP accumulation was measured by a LANCE Ultra cAMP kit (PerkinElmer). Briefly, 24 h after transfection with GHRHR(23-423), GHRHR(23-405), GHRHR(23-405)-LgBiT-2MBP, or HA-Flag-GHRHR (WT and mutants), HEK 293 T cells (3000/well in 384-well white plates, PerkinElmer) were digested by 0.2% (w/v) EDTA and 5 μL stimulation buffer (HBSS supplemented with 5 mM HEPES, 0.5 mM IBMX, and 0.1% (w/v) BSA, pH 7.4). Different concentrations (5 μL) of GHRH were then added and the stimulation lasted for 30 min at RT. The reaction was stopped by adding 5 μL Eu-cAMP tracer and 5 μL ULight-anti-cAMP. After 1 h RT incubation, TR-FRET signals (excitation: 320 nm, emission: 615 and 665 nm) were measured by an EnVision (PerkinElmer).
Whole cell binding assay
This assay was conducted in 96-well Iso-plates (PerkinElmer) coated with poly-D-lysine hydrobromide (Sigma-Aldrich). Twenty-four hour after transfection with GHRHR(23-423) or GHRHR(23-405)-LgBiT-2MBP, HEK 293 T cells were washed twice, and incubated with blocking buffer (DMEM medium supplemented with 33 mM HEPES, and 0.1% (w/v) BSA, pH 7.4) for 2 h at 37 °C. Then, radiolabeled 125I-GHRH (80 pM, Phoenix Biotech) and seven decreasing concentrations of unlabeled peptide (10 μM, five serial gradient dilutions) were added and competitively reacted with the cells in binding buffer (PBS supplemented with 10% (w/v) BSA, pH 7.4) at RT for 3 h. Cells were washed with ice-cold PBS and lysed by 50 μL lysis buffer (PBS supplemented with 20 mM Tris-HCl and 1% (v/v) Triton X-100, pH 7.4). Finally, 150 μL of scintillation cocktail (OptiPhase SuperMix, PerkinElmer) was employed and radioactivity (counts per minute) read in a scintillation counter (MicroBeta2 plate counter, PerkinElmer).
β-arrestin2 recruitment assay
HEK 293 T cells were seeded at a density of 30,000 cells per well into 96-well culture plates pretreated with poly-D-lysine hydrobromide. After incubation for 24 h to reach 70–80% confluence, the cells were transiently transfected with HA-Flag-GHRHR-Rluc8 (WT and mutants) and β-arrestin2-Venus at a 1:9 mass ratio, using Lipofectamine 3000 reagent (Invitrogen) and cultured for another 24 h. Thereafter, cells were washed once and incubated for 30 min at 37 °C with HBSS buffer (pH 7.4) supplemented with 0.1% (m/v) BSA and 10 mM HEPES. A total of 5 μM coelenterazine h (YEASEN Biotechnology) was then added and incubated for 5 min in the dark. The bioluminescence resonance energy transfer (BRET) signals were detected with an EnVision by calculating the ratio of emission at 535 nm over emission at 470 nm. A 1.5 min baseline of BRET measurements was taken before the addition of 250 μM GHRH or vehicle and BRET signal was measured at 10 s intervals for further 9 min. After removing baseline and background readings by subtracting average values of the baseline measurement and average values of vehicle-treated samples, respectively, the AUC across the time-course response curve was determined and normalized to the WT, which was set to 100%.
Molecular dynamics simulation
All-atom MD simulations of the GHRH–GHRHR were performed by Gromacs 2018.5. After removing all G protein subunits and heteroatoms, the receptor and its disease-causing mutants (D60G, R94Q, S140P, and R357C) were prepared and capped by the Protein Preparation Wizard (Schrodinger 2017-4). Titratable residues were left in their dominant state at pH 7.0. The complexes were embedded in a bilayer composed of 225 POPC lipids and solvated with 0.15 M NaCl in explicitly represented waters, using CHARMM-GUI Membrane Builder48. The CHARMM36-CAMP force filed49 was adopted for protein, peptides, lipids, and salt ions, while the CHARMM TIP3P model was chosen for water. The particle mesh Ewald method50 was used to treat all electrostatic interactions beyond a cutoff of 10 Å and the bonds involving hydrogen atoms were constrained using LINCS algorithm51. The constructed system was firstly relaxed using the steepest descent energy minimization, followed with slow heating of the system to 310 K with restraints. The restraints were reduced gradually over 50 ns, with a simulation step of 1 fs. Finally, a 1 μs production run without restraints was carried out for each simulation, with a time step of 2 fs in the NPT ensemble at 310 K and 1 bar using the v-rescale thermostat52 and the semiisotropic Parrinello–Rahman barostat53, respectively. The last 500 ns trajectory was used for analysis, and the GHRH–GHRHR interface area was calculated by the program FreeSASA54, using the Sharke-Rupley algorithm with a probe radius of 1.2 Å.
Statistical analysis
All functional study data were analyzed using Prism 7 (GraphPad) and presented as means ± S.E.M. from at least three independent experiments. Concentration–response curves were evaluated with a three-parameter logistic equation. The significance was determined with either two-tailed Student’s t test or one-way ANOVA, and P < 0.05 was considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Supplementary information
Acknowledgements
We thank Chenyao Li for technical assistance. The cryo-EM data were collected at the Center of Cryo-Electron Microscopy, Zhejiang University. This work was partially supported by National Natural Science Foundation of China 81872915 (M.-W.W.), 81922071 (Yan Zhang), 81773792 (D.Y.), 81973373 (D.Y.), 21704064 (Q.Z.), 31770796 (Y.J.), and 31971178 (S.Z.); National Science and Technology Major Project of China—Key New Drug Creation and Manufacturing Program 2018ZX09735–001 (M.-W.W.), 2018ZX09711002–002–005 (D.Y.), and 2018ZX09711002–002–002 (Y.J.); Shanghai Municipal Science and Technology Major Project 2019SHZDZX02 (H.E.X.); Ministry of Science and Technology of China Major Project XDB08020303 (H.E.X.); National Key R&D Program of China 2018YFA0507000 (M.-W.W. and S.Z.); Zhejiang Province Science Fund for Distinguished Young Scholars LR19H310001 (Yan Zhang); Shanghai Science and Technology Development Fund 18ZR1447800 (L.-H.Z.); Young Innovator Association of CAS (2018325 to L.-H.Z.); SA-SIBS Scholarship Program (L.-H.Z.), and Novo Nordisk-CAS Research Fund grant NNCAS-2017–1-CC (D.Y.).
Source data
Author contributions
F.Z., Z.C., and L.-H.Z. designed the expression constructs, purified the GHRH–GHRHR–Gs complex, prepared the final samples for negative-stain/data collection toward the structure, and participated in manuscript preparation; H.Z. and C.Y.M. performed specimen screening, cryo-EM grid preparation/data collection, map calculation, and the bulk figures preparation; Q.Z. performed structural analysis, MD simulations on GHRHR, mutation studies, and manuscript preparation; X.C. conducted MD simulations on GHRHR and participated in manuscript preparation; D.-D.S. and C.M. assisted in the specimen screening by negative-stain EM; X.Q.C., Y.W., A.D., and Yan Zhou performed ligand binding and signaling experiments; W.S. and F.H.Z. helped construct preparation, protein expression, and purification; S.Z. and H.J. supervised MD simulations; Y.J. participated in data analysis and manuscript editing; D.Y. supervised ligand binding and signaling experiments, participated in data analysis, and manuscript preparation; H.E.X., Yan Zhang, and M.-W.W. initiated the project, supervised the studies, analyzed the data, and wrote the manuscript with inputs from all co-authors.
Data availability
Data supporting the findings of this manuscript are available from the corresponding authors upon reasonable request. A reporting summary for this article is available as a Supplementary information file. The atomic coordinates and the electron microscopy maps have been deposited in the Protein Data Bank (PDB) under accession number PDB 7CZ5 and Electron Microscopy Data Bank (EMDB) accession number EMD-30505. Source data are provided with this paper.
Competing interests
The authors declare no competing interests.
Footnotes
Peer review information Nature Communications thanks Javier Garcia-Nafria, Asuka Inoue, and Zhi-Jie Liu for their contribtions to the peer review fof this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Fulai Zhou, Huibing Zhang, Zhaotong Cong, Li-Hua Zhao, Qingtong Zhou.
Contributor Information
Dehua Yang, Email: dhyang@simm.ac.cn.
H. Eric Xu, Email: eric.xu@simm.ac.cn
Yan Zhang, Email: zhang_yan@zju.edu.cn.
Ming-Wei Wang, Email: mwwang@simm.ac.cn.
Supplementary information
Supplementary information is available for this paper at 10.1038/s41467-020-18945-0.
References
- 1.Pandol SJ, Seifert H, Thomas MW, Rivier J, Vale W. Growth hormone-releasing factor stimulates pancreatic enzyme secretion. Science. 1984;225:326–328. doi: 10.1126/science.6204379. [DOI] [PubMed] [Google Scholar]
- 2.Guillemin R, et al. Growth hormone-releasing factor from a human pancreatic tumor that caused acromegaly. Science. 1982;218:585–587. doi: 10.1126/science.6812220. [DOI] [PubMed] [Google Scholar]
- 3.Cohen LE, Hashimoto Y, Zanger K, Wondisford F, Radovick S. CREB-independent regulation by CBP is a novel mechanism of human growth hormone gene expression. J. Clin. Investig. 1999;104:1123–1130. doi: 10.1172/JCI7308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Granata R. Peripheral activities of growth hormone-releasing hormone. J. Endocrinol. Investig. 2016;39:721–727. doi: 10.1007/s40618-016-0440-x. [DOI] [PubMed] [Google Scholar]
- 5.Cai R, et al. Synthesis of new potent agonistic analogs of growth hormone-releasing hormone (GHRH) and evaluation of their endocrine and cardiac activities. Peptides. 2014;52:104–112. doi: 10.1016/j.peptides.2013.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fridlyand LE, Tamarina NA, Schally AV, Philipson LH. Growth hormone-releasing hormone in diabetes. Front. Endocrinol. 2016;7:129. doi: 10.3389/fendo.2016.00129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Shen J, et al. Regulation of vascular calcification by growth hormone-releasing hormone and its agonists. Circ. Res. 2018;122:1395–1408. doi: 10.1161/CIRCRESAHA.117.312418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Villanova T, et al. Antagonists of growth hormone-releasing hormone (GHRH) inhibit the growth of human malignant pleural mesothelioma. Proc. Natl Acad. Sci. USA. 2019;116:2226–2231. doi: 10.1073/pnas.1818865116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schally AV, Varga JL, Engel JB. Antagonists of growth-hormone-releasing hormone: an emerging new therapy for cancer. Nat. Clin. Pr. Endocrinol. Metab. 2008;4:33–43. doi: 10.1038/ncpendmet0677. [DOI] [PubMed] [Google Scholar]
- 10.Zhang Y, et al. Cryo-EM structure of the activated GLP-1 receptor in complex with a G protein. Nature. 2017;546:248–253. doi: 10.1038/nature22394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Liang YL, et al. Phase-plate cryo-EM structure of a biased agonist-bound human GLP-1 receptor-Gs complex. Nature. 2018;555:121–125. doi: 10.1038/nature25773. [DOI] [PubMed] [Google Scholar]
- 12.Liang YL, et al. Cryo-EM structure of the active, Gs-protein complexed, human CGRP receptor. Nature. 2018;561:492–497. doi: 10.1038/s41586-018-0535-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liang YL, et al. Phase-plate cryo-EM structure of a class B GPCR-G-protein complex. Nature. 2017;546:118–123. doi: 10.1038/nature22327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Qiao A, et al. Structural basis of Gs and Gi recognition by the human glucagon receptor. Science. 2020;367:1346–1352. doi: 10.1126/science.aaz5346. [DOI] [PubMed] [Google Scholar]
- 15.Zhao LH, et al. Structure and dynamics of the active human parathyroid hormone receptor-1. Science. 2019;364:148–153. doi: 10.1126/science.aav7942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ma S, et al. Molecular basis for hormone recognition and activation of corticotropin-releasing factor receptors. Mol. Cell. 2020;77:669–680 e664. doi: 10.1016/j.molcel.2020.01.013. [DOI] [PubMed] [Google Scholar]
- 17.Liang YL, et al. Toward a structural understanding of class B GPCR peptide binding and activation. Mol. Cell. 2020;77:656–668 e655. doi: 10.1016/j.molcel.2020.01.012. [DOI] [PubMed] [Google Scholar]
- 18.Liang YL, et al. Structure and dynamics of adrenomedullin receptors AM1 and AM2 reveal key mechanisms in the control of receptor phenotype by receptor activity-modifying proteins. ACS Pharm. Transl. Sci. 2020;3:263–284. doi: 10.1021/acsptsci.9b00080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Duan J, et al. Cryo-EM structure of an activated VIP1 receptor-G protein complex revealed by a NanoBiT tethering strategy. Nat. Commun. 2020;11:4121. doi: 10.1038/s41467-020-17933-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhao LH, et al. Differential requirement of the extracellular domain in activation of class B G protein-coupled receptors. J. Biol. Chem. 2016;291:15119–15130. doi: 10.1074/jbc.M116.726620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wootten D, Simms J, Miller LJ, Christopoulos A, Sexton PM. Polar transmembrane interactions drive formation of ligand-specific and signal pathway-biased family B G protein-coupled receptor conformations. Proc. Natl Acad. Sci. USA. 2013;110:5211–5216. doi: 10.1073/pnas.1221585110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cervini LA, Donaldson CJ, Koerber SC, Vale WW, Rivier JE. Human growth hormone-releasing hormone hGHRH(1-29)-NH2: systematic structure-activity relationship studies. J. Med. Chem. 1998;41:717–727. doi: 10.1021/jm970618s. [DOI] [PubMed] [Google Scholar]
- 23.Matsoukas MT, Spyroulias GA. Dynamic properties of the growth hormone releasing hormone receptor (GHRHR) and molecular determinants of GHRH binding. Mol. Biosyst. 2017;13:1313–1322. doi: 10.1039/c7mb00130d. [DOI] [PubMed] [Google Scholar]
- 24.Zhang H, et al. Structure of the glucagon receptor in complex with a glucagon analogue. Nature. 2018;553:106–110. doi: 10.1038/nature25153. [DOI] [PubMed] [Google Scholar]
- 25.Wootten D, et al. The extracellular surface of the GLP-1 receptor is a molecular trigger for biased agonism. Cell. 2016;165:1632–1643. doi: 10.1016/j.cell.2016.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Song G, et al. Human GLP-1 receptor transmembrane domain structure in complex with allosteric modulators. Nature. 2017;546:312–315. doi: 10.1038/nature22378. [DOI] [PubMed] [Google Scholar]
- 27.Siu FY, et al. Structure of the human glucagon class B G-protein-coupled receptor. Nature. 2013;499:444–449. doi: 10.1038/nature12393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Thal DM, Glukhova A, Sexton PM, Christopoulos A. Structural insights into G-protein-coupled receptor allostery. Nature. 2018;559:45–53. doi: 10.1038/s41586-018-0259-z. [DOI] [PubMed] [Google Scholar]
- 29.Isberg V, et al. Generic GPCR residue numbers—aligning topology maps while minding the gaps. Trends Pharm. Sci. 2015;36:22–31. doi: 10.1016/j.tips.2014.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hollenstein K, et al. Insights into the structure of class B GPCRs. Trends Pharm. Sci. 2014;35:12–22. doi: 10.1016/j.tips.2013.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pal K, Melcher K, Xu HE. Structure and mechanism for recognition of peptide hormones by class B G-protein-coupled receptors. Acta Pharm. Sin. 2012;33:300–311. doi: 10.1038/aps.2011.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cohen E, et al. Contribution of functionally assessed GHRHR mutations to idiopathic isolated growth hormone deficiency in patients without GH1 mutations. Hum. Mutat. 2019;40:2033–2043. doi: 10.1002/humu.23847. [DOI] [PubMed] [Google Scholar]
- 33.Eicher EM, Beamer WG. Inherited ateliotic dwarfism in mice. Characteristics of the mutation, little, on chromosome6. J. Hered. 1976;67:87–91. doi: 10.1093/oxfordjournals.jhered.a108682. [DOI] [PubMed] [Google Scholar]
- 34.Lin SC, et al. Molecular basis of the little mouse phenotype and implications for cell type-specific growth. Nature. 1993;364:208–213. doi: 10.1038/364208a0. [DOI] [PubMed] [Google Scholar]
- 35.Corazzini V, Salvatori R. Molecular and clinical aspects of GHRH receptor mutations. Endocr. Dev. 2013;24:106–117. doi: 10.1159/000342575. [DOI] [PubMed] [Google Scholar]
- 36.Landrum MJ, et al. ClinVar: improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 2018;46:D1062–D1067. doi: 10.1093/nar/gkx1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Liang YL, et al. Dominant negative G proteins enhance formation and purification of agonist-GPCR-G protein complexes for structure determination. ACS Pharm. Transl. Sci. 2018;1:12–20. doi: 10.1021/acsptsci.8b00017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dixon AS, et al. NanoLuc complementation reporter optimized for accurate measurement of protein interactions in cells. ACS Chem. Biol. 2016;11:400–408. doi: 10.1021/acschembio.5b00753. [DOI] [PubMed] [Google Scholar]
- 39.Rasmussen SG, et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature. 2011;477:549–555. doi: 10.1038/nature10361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zheng SQ, et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods. 2017;14:331–332. doi: 10.1038/nmeth.4193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang K. Gctf: real-time CTF determination and correction. J. Struct. Biol. 2016;193:1–12. doi: 10.1016/j.jsb.2015.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Scheres SH. Processing of structurally heterogeneous cryo-EM data in RELION. Methods Enzymol. 2016;579:125–157. doi: 10.1016/bs.mie.2016.04.012. [DOI] [PubMed] [Google Scholar]
- 43.Heymann JB. Single particle reconstruction and validation using Bsoft for the map challenge. J. Struct. Biol. 2018;204:90–95. doi: 10.1016/j.jsb.2018.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pettersen EF, et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004;25:1605–1612. doi: 10.1002/jcc.20084. [DOI] [PubMed] [Google Scholar]
- 45.Emsley P, Cowtan K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D. Biol. Crystallogr. 2004;60:2126–2132. doi: 10.1107/S0907444904019158. [DOI] [PubMed] [Google Scholar]
- 46.Adams PD, et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D. Biol. Crystallogr. 2010;66:213–221. doi: 10.1107/S0907444909052925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Booth DS, Avila-Sakar A, Cheng Y. Visualizing proteins and macromolecular complexes by negative stain EM: from grid preparation to image acquisition. J. Vis. Exp. 2011;58:3227. doi: 10.3791/3227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wu EL, et al. CHARMM-GUI membrane builder toward realistic biological membrane simulations. J. Comput. Chem. 2014;35:1997–2004. doi: 10.1002/jcc.23702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guvench O, et al. CHARMM additive all-atom force field for carbohydrate derivatives and its utility in polysaccharide and carbohydrate-protein modeling. J. Chem. Theory Comput. 2011;7:3162–3180. doi: 10.1021/ct200328p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Darden T, York D, Pedersen L. Particle mesh Ewald—an N.Log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993;98:10089–10092. [Google Scholar]
- 51.Hess B. P-LINCS: a parallel linear constraint solver for molecular simulation. J. Chem. Theory Comput. 2008;4:116–122. doi: 10.1021/ct700200b. [DOI] [PubMed] [Google Scholar]
- 52.Bussi G, Donadio D, Parrinello M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007;126:014101. doi: 10.1063/1.2408420. [DOI] [PubMed] [Google Scholar]
- 53.Aoki KM, Yonezawa F. Constant-pressure molecular-dynamics simulations of the crystal-smectic transition in systems of soft parallel spherocylinders. Phys. Rev. A. 1992;46:6541–6549. doi: 10.1103/physreva.46.6541. [DOI] [PubMed] [Google Scholar]
- 54.Mitternacht S. FreeSASA: an open source C library for solvent accessible surface area calculations. F1000Res. 2016;5:189. doi: 10.12688/f1000research.7931.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hernandez LM, Lee PD, Camacho-Hubner C. Isolated growth hormone deficiency. Pituitary. 2007;10:351–357. doi: 10.1007/s11102-007-0073-3. [DOI] [PubMed] [Google Scholar]
- 56.Alba M, Salvatori R. Familial growth hormone deficiency and mutations in the GHRH receptor gene. Vitam. Horm. 2004;69:209–220. doi: 10.1016/S0083-6729(04)69007-8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data supporting the findings of this manuscript are available from the corresponding authors upon reasonable request. A reporting summary for this article is available as a Supplementary information file. The atomic coordinates and the electron microscopy maps have been deposited in the Protein Data Bank (PDB) under accession number PDB 7CZ5 and Electron Microscopy Data Bank (EMDB) accession number EMD-30505. Source data are provided with this paper.