

Abstract
G protein-coupled receptors (GPCRs) represent the largest class of “druggable” proteins in the human genome. For more than a decade, crystal structures and, more recently, cryoEM structures of GPCR complexes have provided unprecedented insight into GPCR drug binding and cell signaling. Nevertheless, structure determination of receptors in complexes with weakly binding molecules or complex polypeptides remains especially challenging, including for hormones, many of which have so far eluded researchers. Nuclear magnetic resonance (NMR) spectroscopy has emerged as a promising approach to determine structures of ligands bound to their receptors and to provide insights into the dynamics and pharmacokinetics of GPCR-bound drugs. The capability to investigate compounds with weak binding affinities has also been leveraged in NMR applications to identify novel lead compounds in drug screening campaigns. We review recent structural biology studies of GPCR ligands by NMR, highlighting new methodologies enabling studies of GPCRs with native sequences and in native-like membrane environments that provide insights into important drugs and endogenous ligands.
1. Introduction
The human genome encodes 826 G protein-coupled receptors (GPCRs) that recognize and interact with external stimuli spanning a wide range of chemical and physical properties, including ions, photons, small molecules, polypeptides and proteins. GPCRs are also targeted by at least 1/3 of currently FDA-approved drugs and remain top drug targets in academic and industrial laboratories [1]. Knowledge of GPCR ligands and their interaction with receptors informs our understanding of GPCR cellular signaling and drives the development of new drugs [2].
Data from nuclear magnetic resonance (NMR) spectroscopic studies have provided novel mechanistic insights into GPCR function-related dynamics by observing receptors in complexes with various drugs, including with studies of adrenergic receptors [3–9], adenosine receptors [10–15], and opioid receptors [16, 17], as reviewed more extensively elsewhere [18]. In parallel, NMR spectroscopy has also provided insights into GPCR signaling and drug discovery through studies that observe ligands directly. This complementary approach has been leveraged to determine the structures and binding epitopes of GPCR-bound drugs and hormones and, in combination with additional structural data from other techniques, used as input to determine and refine molecular models for GPCR complexes that have not been crystallized or studied with cryoEM. Here we highlight several unique strengths of this approach, including the capability of NMR spectroscopy to study ligands with weak binding affinities or dynamic behavior such as polypeptide hormones. These unique capabilities also enable screening with small molecular fragments by NMR, which provides the opportunity for fragment-based drug discovery approaches on especially challenging GPCR targets.
2. Studies of hormones and polypeptides bound to GPCRs in aqueous solutions and lipid bilayers
Knowledge of GPCR ligand binding modes provides a molecular understanding of the pharmacological activity of drugs and can guide the rational design of investigational and clinical compounds [2]. GPCR crystal structures and, more recently, cryoEM structures have provided the vast majority of information on receptor-ligand interactions. Crystallization of drug-receptor complexes is generally facilitated by the use of ligands with relatively strong affinities (e.g. KD <100 nM) and specific molecular properties [19–21], which can limit the potential chemical space of GPCR drugs for which structural information can be obtained by this method alone. Complementing this approach, NMR spectroscopy in aqueous solutions and solids can provide structural data of bound drugs for compounds with both strong and weak affinities (KD > 100 nM), higher off-rates, or for ligands that exhibit dynamic behavior even while bound to the receptor [18]. NMR spectroscopy has been applied to determine structures of ligands exhibiting a wide range of chemical and physical properties, including small molecule neurotransmitters [22], fatty acids [23–25], proteins [26] and polypeptides [27–37]. NMR methods can be used to determine structures of bound drugs in the absence of a known structure of the receptor, or the respective strengths of NMR spectroscopy and x-ray crystallography can be combined in drug discovery programs that identify lead compound “hits” by NMR that are subsequently optimized and crystallized in complex with the target protein [2, 18, 38–40].
2.1. Determination of GPCR-bound hormone and polypeptide structures by NMR in aqueous solutions
Among the 826 GPCRs encoded by the human genome, over 100 bind endogenous polypeptides [41, 42], including many representatives from Class A and Class B receptors. Numerous endogenous GPCR polypeptides are used in the clinic, including vasopressin, oxytocin, and angiotensin II, and over 26 modified or synthetic polypeptides are approved for clinical applications [43]. For nearly all GPCR polypeptides used in the clinic, there exist no publicly available structural data of the receptor-bound conformation, emphasizing the need for additional techniques to provide such information. Though crystal structures of more than a quarter of polypeptide-binding receptors have been determined for complexes with small molecules, structures of complexes with either endogenous or synthetic polypeptides remain largely underrepresented in the protein data bank [36].
Several general alternative approaches have emerged for determining structures of receptor-bound polypeptides by NMR spectroscopy. NMR methods can be applicable to different regimes of ligand binding affinities and rates of exchange between receptor-bound and free states [23–25, 27, 28, 35, 37] (Figure 1). For ligands exhibiting affinities weaker than 100 nM and relatively fast exchange between bound and free states (koff > R1 longitudinal relaxation rates), 1H–1H distance estimates extracted from transferred nuclear Overhauser effect (trNOE) data can be used to calculate structures of receptor-bound polypeptides [44]. In trNOE experiments1H–1H NOEs develop for the smaller ligand bound to the larger receptor and are observed in NMR spectra of the unbound ligand. In trNOE experiments, samples typically contain an excess of 100 to 1000 ligand molecules per receptor, and by observing the trNOE in spectra of the unbound ligand, the experiment leverages the much higher sensitivity of the unbound ligand than what would be obtained by observing the ligand directly while bound to the receptor. Contributions to the NOESY spectra from the ligand while bound and free can be separated, allowing quantitative analysis of the data to determine cross relaxation rates and 1H–1H distances of the bound polypeptide [45]. Control experiments that confirm NOEs specific to receptor-ligand interactions are carried out by the addition of a higher affinity competing ligand that displaces the more weakly binding compound, or by acquiring data in absence of the receptor [27, 35, 37]. An ensemble of GPCR-bound polypeptide structures is then calculated by simulated annealing restrained by the network of measured 1H–1H distances (Figure 1A) [27, 35, 37]. Significant advantages of this approach are that relatively low concentrations of receptor are required (1 to 10 μM) and stable-isotope labeling of the receptor is not required [27, 35, 37].
Figure 1.
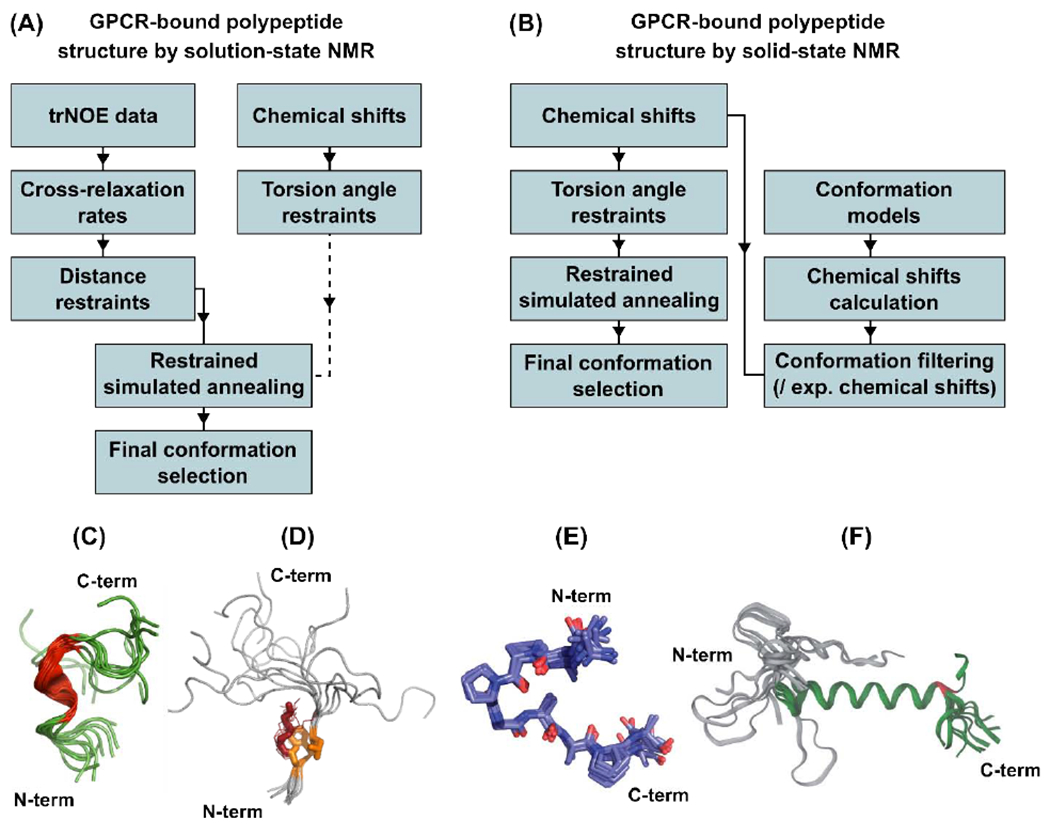
Workflows for determining structures of GPCR bound polypeptides and representative structural ensembles of receptor-bound polypeptides calculated from these approaches. (A) In aqueous solutions, trNOE restraints and chemical shifts are used as input data to calculate receptor-bound polypeptide structures, and (B) in solids, chemical shifts provide experimental restraints and are also used to evaluate molecular models. (C and D) Two examples from the first approach are Dynorphin bound to the κ-opioid receptor (KOP; PDB ID 2N2F) [46] (C) and Ghrelin bound to the growth hormone secretagogue receptor (GHSR; PDB ID 6H3E) [27, 37] shown with ordered side chains in stick representation (one conformer) or lines (other conformers) (D). (E and F) Two examples from the second approach are desArg-kallidin (DAKD) bound to the bradykinin 1 receptor (B1R; PDB ID 6F3Y) [29] (E) and Neuropeptide Y (NPY) bound to the Y2 receptor (Y2R) [33] (F). Panels C, D, E, and F adapted with permission from references [37], [27], [33], and [60].
2.2. Examples with human GPCR hormones and neuropeptides
The approach described in section 2.1 was exemplified in recent studies that determined the structures of two GPCR-bound polypeptides: Dynorphin and Ghrelin. Dynorphin is an endogenous agonist of the κ-opioid receptor (KOR), which regulates important physiological processes including perception of pain, consciousness and moods, and is also a drug target in the effort to mitigate addiction to opioid compounds [46]. Though crystal structures of KOR in complex with an antagonist and agonist are available, currently no crystal structures have been published of KOR in complex with Dynorphin. A collaborative effort of the groups of Milon and Wüthrich used NMR data, including trNOEs and chemical shifts, to calculate an ensemble of structures of Dynorphin bound to KOR in solution [37] (Figure 1, A and C). The NMR structures showed that upon complex formation with KOR, Dynorphin adopts an α-helical turn in the central section of the polypeptide [37]. Intriguingly, both the N-terminus and C-terminus of Dynorphin, which are involved with activation of KOR and selectivity of Dynorphin for KOR, respectively, remain unstructured in the KOR-bound state [37], with implications described below in section 3.1.
In a second example, the structure of the human hormone Ghrelin bound to its receptor, the growth hormone secretagogue receptor (GHSR), was determined using trNOE [27] restraints (Figure 1, A and D). Ghrelin, the “hunger hormone” produced in the stomach and small intestine, stimulates appetite and regulates food intake and growth hormone secretion [47]. Currently no crystal or cryoEM structures are available of GHSR. An NMR structure of Ghrelin was determined using the approach outlined above, which showed a well folded region of the polypeptide that formed a “hydrophobic core” while bound to GHSR [27]. The latter encompasses an octanoyl post-translational modification of Ghrelin, which is essential for its biological activity, and strongly interacts with GHSR. Reminiscent of the flexible ends of Dynorphin, the structure of Ghrelin was also observed to exhibit a large degree of flexibility while bound to its receptor, suggesting that receptor-bound polypeptides may exhibit a large range of dynamics generally [27, 30–32, 35].
2.3. Structure determination of GPCR-bound polypeptides by solid state NMR
NMR approaches are not limited to fast exchange and can also be used to study GPCR ligands that are more tightly bound to their receptors, or ligands where the conformational dynamics has been experimentally reduced, for example by freezing the sample (see below). A second approach to determining structures of receptor-bound polypeptides is based on experimentally measured NMR chemical shifts [26, 29–34, 37] (Figure 1, panels B, E and F). With this approach, 2- or 3-dimensional correlation experiments are recorded for stable-isotope labeled ligands bound to unlabeled receptors, and chemical shifts are determined for assigned signals observed with the receptor-bound ligand.
A pioneering application of this approach was the determination of the structure of neurotensin, a 13 amino acid polypeptide, bound to its receptor NTS1 [31]. Detection of relatively small amounts of neurotensin bound to NTS1 were facilitated by the use of double-quantum filtered correlation experiments that removed contributions from the 13C natural abundance background [31]. The resulting NMR structure was later shown to be consistent with a crystal structure of neurotensin bound to NTS1 [48].
To simplify the NMR spectra of bound polypeptides, spectral editing can be applied and polypeptide ligands can be prepared containing a fraction of amino acid residues that are stable-isotope labeled. An advantage of this approach is that it can be applied to samples in both aqueous solutions and solids and permits studies of receptor-ligand interactions within a wide range of sample environments including detergent micelles, nanodiscs, or in lipid vesicles [29–34]. Stable-isotope labeling of a subset of the polypeptide chain has helped with assigning NMR signals in numerous solid-state studies and facilitated resolving NOESY signals for investigating the structure of GHSR-bound Ghrelin by solution-state experiments using 3D 1H-1H-15N NOESY-HMQC spectra.
Because the chemical shifts of signals from the polypeptide backbone correlate with secondary structure, comparing experimentally measured chemical shifts to values in empirical databases enables prediction of torsion angles for the polypeptide chain [31, 49–51]. These predicted torsion angles can be combined with measured distance restraints from other NMR experiments (Figure 1, A and C) [37] , or used alone as restraints in simulated annealing protocols (Figure 1, B and E) [29, 30]. An alternative strategy consists of using predicted chemical shifts [52, 53] calculated from multiple generated models of the receptor-bound polypeptide conformation and then selecting models with predicted chemical shifts that are most consistent with experimentally determined values (Figure 1, B and F) [29, 32, 33].
2.4. Models of receptor-ligand complexes using experimentally determined structures of GPCR polypeptides
To generate models of GPCR-polypeptide complexes, experimentally-derived bound polypeptide conformations can be used in computational docking protocols with the receptor structure [28, 33, 34, 37, 46], reducing the polypeptide conformational space that must be sampled. Also, experimental restraints can be applied during molecular dynamics (MD) simulations of the polypeptide-GPCR interactions [27, 37, 46] with measured distances or torsion angles or to select models that best fit the experimental data among a larger ensemble with chemical shift information or order parameter profiles [29, 34, 37, 46]. Molecular modeling can thus integrate ligand-detected NMR data with GPCR x-ray or cryo-EM structures or homology models.
2.5. Sensitivity-enhanced solid-state NMR experiments with GPCR polypeptides
NMR spectroscopy has a unique capability to provide atomic resolution structural information on human GPCRs and their ligands with native receptors and native bound hormones, circumventing the need to modify the protein, which can lead to changes in its conformational dynamics and activity [7]. A significant challenge posed by working with native protein sequences is that they generally do not express well, limiting the amount of sample that can be studied. Thus, methods that improve the sensitivity of NMR experiments greatly advance the potential to gain structural insights into GPCRs in their native environments.
One emerging method for improving sensitivity in solid state NMR is the detection of 1H nuclei. While 1H detection has been used routinely for decades in solution NMR experiments, its use in solid state NMR has not yet been as widely implemented due to broad lines that are observed at moderate magic angle spinning (MAS) frequencies (10 – 30 kHz) without substantial deuteration. With the advent of high frequency MAS (60 kHz to over 100 kHz), improved pulse sequences, and selective deuteration methods, the resolution in 1H-detected MAS NMR experiments is significantly improved, permitting its use in structure determination efforts [54–56].
A notable application of this method in studies of GPCR ligands was the determination of the binding mode of the human chemokine interleukin 8 (IL-8) to its chemokine receptor CXCR1 [26] (Figure 2). IL-8 induces chemotaxis and recruits inflammatory cells and thus is a key modulator of immune responses. Opella and colleagues measured 1H-detected NMR data of IL-8 unbound in solution and bound to CXCR1 in solid state preparations that reconstituted the receptor into lipid bilayers (Figure 2). Solid state NMR experiments using cross polarization-based transfers and INEPT-based transfers were used to probe regions of IL-8 that were more rigid or more flexible, respectively. In solid state spectra of IL-8 bound to the full length CXCR1, most signals arose from CP-based experiments, indicating that IL-8 was more rigid in the complex. In contrast to this observation, when the first 38 amino acids of CXCR1 were removed, many more signals of IL-8 were observed in INEPT-based spectra. Comparison of the INEPT and CP-based spectra for the full length and N-terminal truncated proteins thus showed the important interactions of the CXCR1 N-terminus in stabilizing the complex with IL-8 [26]. A recent cryo-EM structure of IL-8 in complex with the related CXCR2 chemokine receptor confirms the important role of the N-terminus of the chemokine receptor in recognition and complex formation with IL-8 [57]. In the cryo-EM structure, residues 30 to 38 of CXCR2 form extensive hydrophobic and electrostatic interactions with IL-8. Notably, residues 1 to 25 are not modeled in the cryo-EM structure due to missing electron density [57]. Arrestin recruitment assays demonstrated that removal of CXCR2 residues 1 to 25 prohibited formation of signaling complexes with IL-8 [57]. Altogether, this suggests that while these residues are important for complex formation they may be dynamic even while interacting with IL-8.
Figure 2.
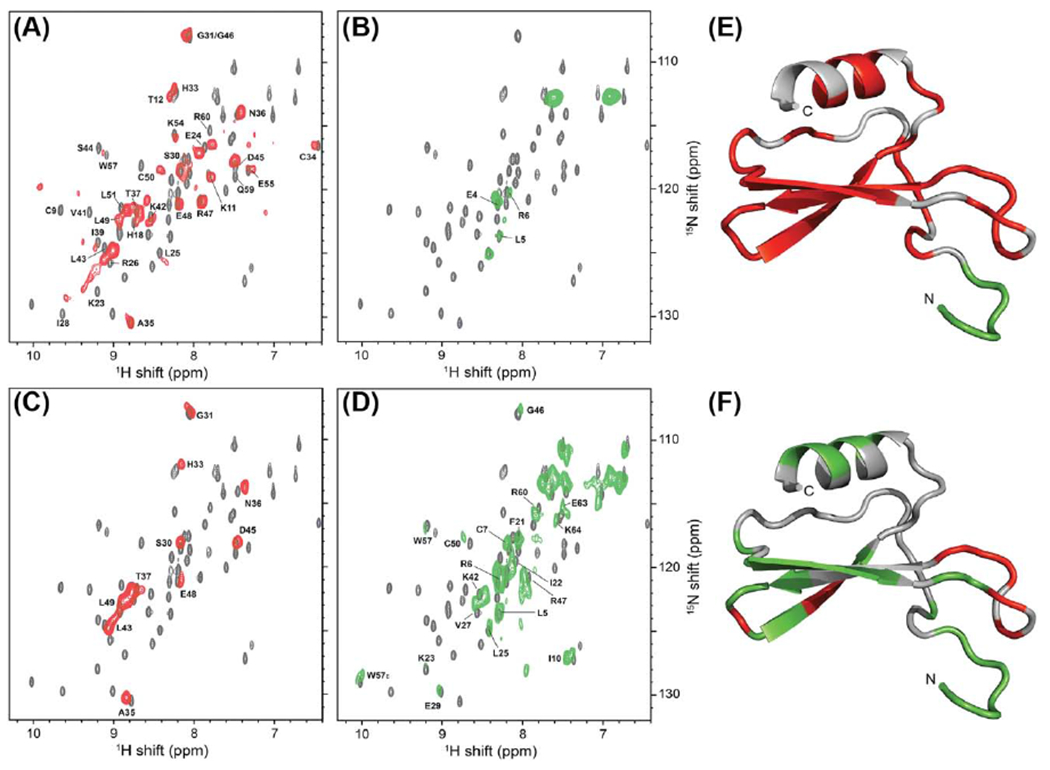
1H-detected solid and solution NMR data of the chemokine IL-8 enable visualization of interactions of the polypeptide with its receptor CXCR1. (A and B) solid state MAS NMR data of [2H, 15N] IL-8 bound to unlabeled full-length CXCR1, and (C and D) bound to an unlabeled CXCR1 variant where the first 38 residues have been removed. 1H-detected solid state NMR data (red and green) are superimposed on solution NMR spectra of unbound IL-8 (grey). The red spectra were measured using cross-polarization (CP) magnetization transfers, which yield spectra containing predominantly signals from more rigid regions of IL-8. The green spectra were measured using INEPT transfers, which yield spectra containing signals from more flexible regions of IL-8. (E and F) Solution NMR structure of IL-8 (PDB ID 5WDZ) where the individual residues have been color coded to indicate if they were observed in CP- or INEPT-based solid-state spectra. Comparison of the two patterns of colors mapped onto IL-8 for complexes with full length CXCR1 (E) and the CXCR1 variant with the first 38 residues removed (F) reveals the role of the interaction between the CXCR1 N-terminus and IL-8. Figure adapted with permission from reference [26].
A second powerful approach to improving the sensitivity of solid-state NMR experiments is with dynamic nuclear polarization (DNP). Sensitivity-enhanced DNP MAS NMR methodology can be used to transfer the inherently higher Boltzmann distribution of electrons to protons and target nuclei such as 13C or 15N. The technique applies microwave irradiation to protein samples containing radicals to provide the increased sensitivity. In pioneering work by Griffin and Herzfeld, DNP MAS NMR was used to probe the mechanistic details of the photoactivation of bacteriorhodopsin, an integral membrane protein sharing the same seven transmembrane topology as GPCRs, revealing the coexistence of multiple function-related conformational states of the bound retinal and photoreceptor [58, 59].
More recently, sensitivity enhancements from DNP NMR have provided insights into the selectivity of polypeptides for bradykinin receptor subtypes [60] (Figure 3). The B1 and B2 bradykinin receptors are involved in distinct physiological responses including inflammation, shock, and response to allergies. While some small molecules and the hormone bradykinin bind both B1 and B2, other small molecules and polypeptides preferentially bind one subtype over the other. Glaubitz and colleagues studied the subtype selectivity for the polypeptide kallidin, which differs from the hormone bradykinin by only one lysine yet shows different affinities for B1 and B2 receptors. Key to this study was the use of native sequences for the B1 and B2 receptors. Because the native proteins express poorly, DNP NMR was critical to obtain the necessary sensitivity that enabled a structural model of receptor-bound kallidin determined by measurement of chemical shifts for the polypeptide [60].
Figure 3.
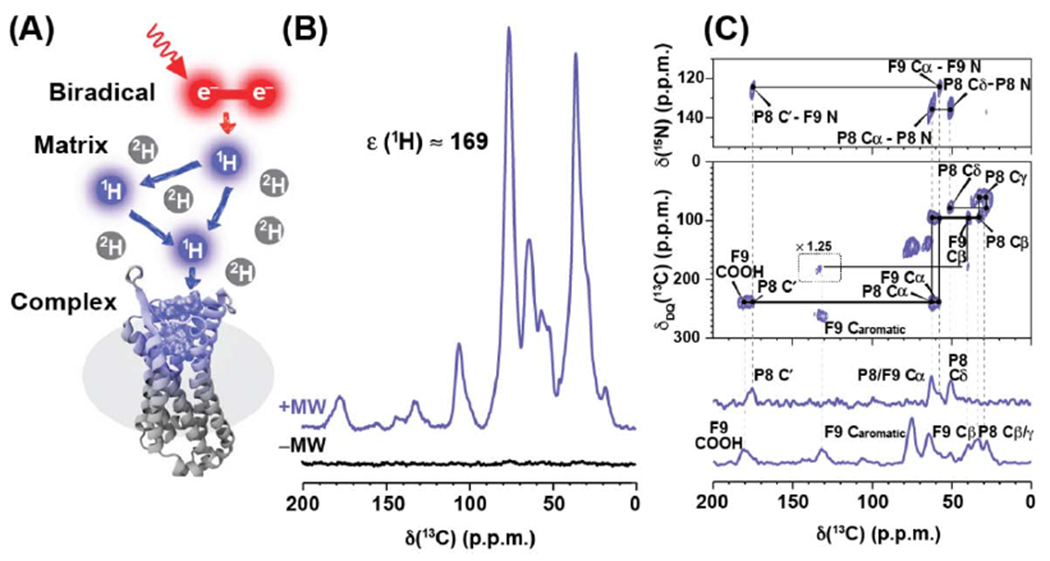
Structure of a polypeptide bound to human bradykinin receptor determined by dynamic nuclear polarization (DNP) NMR. (A) Schematic illustrating the basic principles of DNP NMR. The sample contains the polypeptide-bound receptor in a partially deuterated matrix doped with the radical containing molecule AMUPol[81]. Continuous microwave irradiation is applied to the sample, transferring polarization from electrons in the sample to the bound polypeptide. (B) DNP spectra of stable-isotope labeled DAKD polypeptide bound to B1 with microwaves applied (purple) are enhanced by a factor of 169 in sensitivity compared to spectra when microwaves are turned off (black). (C) Assignments of observed signals were accomplished with DNP MAS NMR 2-dimensional 15N-13C TEDOR (top), 13C-13C DQ-SQ (middle), and 1-dimensional TEDOR and REDOR-filtered 13C experiments (bottom). Figure adapted with permission from reference [30].
3.1. Conformational dynamics of GPCR-bound ligands
Complete characterization of GPCR drugs includes developing an understanding of both the structure and the conformational dynamics underlying molecular recognition and binding of ligands. NMR is uniquely well suited to study both ligand structure and dynamics for the same samples and can, in principle, be applied to the entire known chemical space of GPCR ligands. Some of the foundational observations of receptor-ligand interactions include early NMR studies of the chromophore retinal in complex with its receptor rhodopsin, which established that the confirmation and dynamics of the covalently bound retinal are coupled with structural changes of rhodopsin that alter its functional activity. The first high resolution structures of retinal bound to rhodopsin were determined by solid state NMR [61] and studies from multiple research groups have provided a detailed mechanistic view of how changes in retinal dynamics from photoactiviation underlie rhodopsin function [61–66].
A multitude of experimental approaches are available to quantitatively measure the dynamics of bound ligands for a range of exchange rates between bound and unbound ligand. For ligands that exhibit fast exchange between bound and free states, 15N transverse relaxation rates (R2) are related to the 15N–1H vector order parameters (S2) of the ligand bound to the receptor. These order parameters describe amplitudes of motions faster than the microscopic tumbling of the receptor-bound ligand in solution [37]. Thus 15N relaxation of a labelled polypeptide ligand can be analyzed together with trNOE data and an ensemble of structures to describe the dynamics of the receptor-bound ligand [27, 37]. In one example of this approach, the conformational dynamics of the polypeptide dynorphin bound to KOR were quantitatively measured from 15N–1H vector order parameters (see Figure 4 and section 2.2). Analysis of the order parameters confirmed a rigid core of the polypeptide comprising residues L5 to R9 that formed an alpha helical turn. The C-terminus of dynorphin showed significantly reduced order parameters (Figure 4B), measured at residue L12, indicating the presence of fast dynamics [37]. More surprisingly, order parameters measured for the first three residues were significantly decreased and indicated the presence of fast motion. Thus while the core of the polypeptide was rigid in the complex with KOR, the first three residues were dynamic even though they were deep in the orthosteric binding pocket [37]. This observation provides an intriguing potential rationalization for the difficulty in crystallizing a KOR complex with dynorphin and future studies may investigate the extent of dynorphin conformational dynamics in the orthosteric pocket for tertiary complexes with partner signaling proteins.
Figure 4.
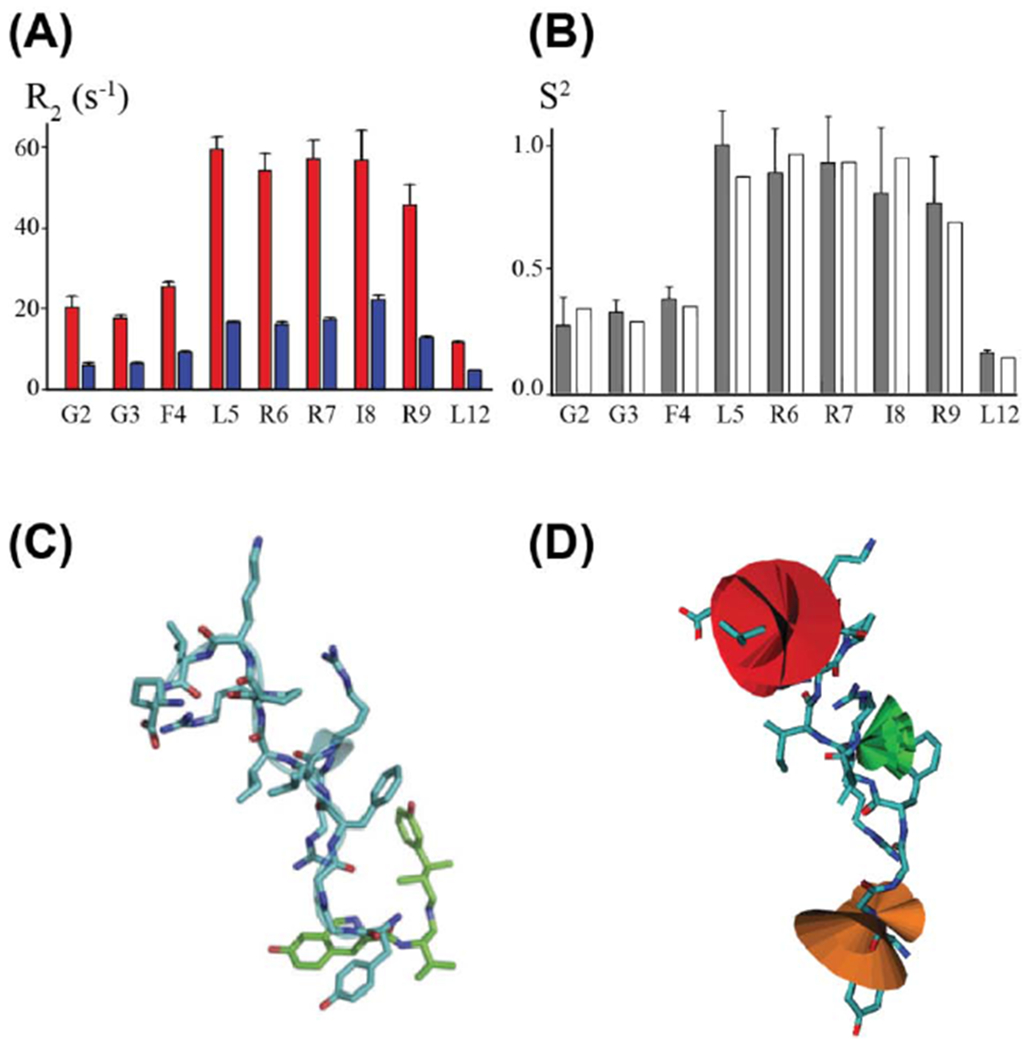
Conformational dynamics of a GPCR neuropeptide in complex with an opioid receptor. (A) 15N R2 relaxation rate constants were measured for stable-isotope labeled dynorphin bound to the KOR (red) and upon addition of the high affinity antagonist JDTic (blue), which displaces the polypeptide from the receptor. (B) Order parameter profile, S2/S2max, derived from experimentally measured R2 relaxation rate constants (grey bars) and MD simulations with the NMR structure of dynorphin docked into a crystal structure of KOR (white bars). (C) NMR structure of dynorphin (blue) superimposed on the crystal structure of JDTic (green) bound to KOR (PDB ID 4DJH). (D) Visualization of order parameters from NMR data displayed on the NMR structure of dynorphin. The relative width of the superimposed cones indicates the magnitude of dynamics. Figure adapted with permission from reference [37].
The dynamic behavior of receptor-bound ligands can also be modulated in variable temperature NMR experiments. This allows researchers to study dynamics both at physiologically relevant temperatures and at very cold temperatures that reduce dynamics to allow observation of more dynamic regions of bound drugs. For example, low temperatures were used in the MAS NMR structure determination of the hormone bradykinin bound to the B2 receptor [30].
NMR studies have provided a more complex view of ligand-receptor interactions and demonstrated that multiple interconverting conformations can be observed for receptor-bound ligands. A recent example is highlighted in ligand-observed 19F NMR studies of the anti-nausea drug aprepitant bound to the neurokinin 1 receptor (NK1R) in aqueous solution revealed multiple conformations of aprepitant bound to NK1R that exchanged on a slow NMR time scale [67]. Ligand-observed NMR data of aprepitant bound to NK1R variants containing amino acid substitutions at position E2.50 showed larger perturbations in the 19F NMR data for substitutions that impacted receptor signaling, relating changes in the conformation of the ligand binding pocket [67] to changes in function. As a significant number of pharmaceutical compounds contain 19F, this approach is likely widely applicable to many future studies of receptor-drug complexes.
3.2. Sample preparation requirements for ligand-observed NMR studies
Preparation of ligand-receptor complexes suitable for NMR studies is a prerequisite for characterization of their structures and dynamics. Several advantages in the preparation of samples for ligand-observed NMR studies make them an appealing option for structural investigations. Nearly all GPCR ligands, including polypeptides or small molecules, can be produced synthetically, or biosynthetically through recombinant expression techniques, often in quantities sufficient for NMR studies. Stable-isotope labels can be incorporated into the ligand uniformly or selectively, permitting observation of individual amino acids within a polypeptide chain or specific chemical moieties in small molecule ligands. Several studies have utilized this approach to simplify NMR spectra and allow for unambiguous assignment of detected signals, including for polypeptides bound to KOR [37], the bradykinin receptor 1 [60], and to GSHR [27].
Given the capability of producing stable-isotope labeled polypeptides or small molecules, the limitation in NMR studies of receptor-ligand complexes remains the production and isolation of the target receptor protein. There are now multiple approaches to producing human GPCRs for NMR studies, as reviewed elsewhere [18] [68]. With respect to protein production requirements, ligand-observed NMR studies offer two advantages over protein observed NMR experiments. First, stable-isotope labels do not need to be incorporated into proteins, which allows production of receptors in expression hosts that are more challenging to incorporate stable-isotopes (i.e. mammalian or insect cells). Second, ligand-observed NMR experiments are typically carried out with samples that contain an excess of ligand, thus requiring lower concentration receptors than protein-observed NMR. With the advent of sensitivity-enhanced DNP NMR (see section 3.5) for solids, sub-milligram quantities of both ligand and receptor can be sufficient for structural investigations [60].
4. Discovery of novel GPCR-targeted small molecules by ligand-observed NMR
In addition to providing structural information of bound polypeptides, ligand-observed NMR spectroscopy has also been implemented in drug discovery programs to identify new small molecules that bind to GPCRs [19, 69–71]. The capability of NMR to detect interactions with affinities as weak as KD ~1 mM has made it an important tool in identification of molecular fragment “hits” that can be subsequently optimized by linking or merging fragments into compounds with improved affinities or potencies [40, 72]. The versatility of ligand-detected NMR experiments is documented by its application for GPCRs reconstituted into a wide range of environments, including detergent micelles, bicelles, nanodiscs or lipid vesicles [19, 27, 32, 35, 37, 69–71, 73–75]. Because relatively small quantities of unlabeled target protein are required, ligand detected NMR holds great promise for studies with difficult-to-express GPCRs [72].
4.1. Small molecule drug screening by NMR with immobilized receptor targets
High throughput methods are clearly desirable in drug discovery, but scaling the throughput of NMR experiments to be practical for screening sufficiently large numbers of compounds can be challenging. One notable application that was developed to increase the throughput of NMR screening methods is target-immobilized NMR screening (TINS) [76]. In this application, the receptor target is fixed to a solid support and solutions containing mixtures of small molecules are flowed over the receptor. Interactions between one or more of the small molecules are then detected by the observed diminishment or absence of signals compared with reference data of the same compounds in the absence of the receptor (Figure 5A). TINS experiments have been used to identify small molecules for both the β1 adrenergic (β1AR) and the A2A adenosine (A2AAR) receptors [70, 71]. For both β1R and A2AAR, variants of the native protein were employed containing multiple stabilizing point mutations that increased the ability of the GPCRs to remain folded and functional for longer periods of time near room temperature [70, 71], a requirement to efficiently screen numerous compounds.
Figure 5.
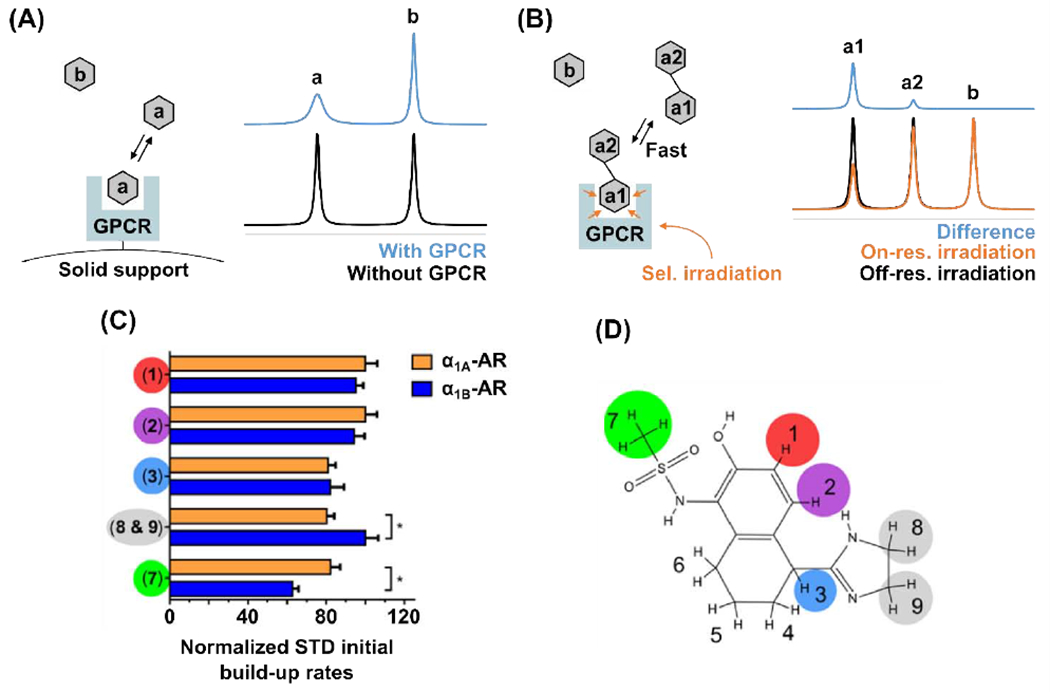
(A and B) Schematic representation of ligand-observed NMR experiments for drug screening: target-immobilized NMR screening (TINS) (A) and saturation transfer difference (STD) (B). In the TINS experiment, detection of the binding of molecule “a” to the receptor is observed by a decrease in signal intensity in the NMR spectrum. In the STD NMR experiment, detection of the binding of molecule “a1-a2” to the receptor is observed as a positive difference between spectra measured with and without RF irradiation applied to the protein target. The difference in signal intensities observed for the two chemical moieties “a1” and “a2” of the compound are due to different spatial proximities of each chemical group to the ligand binding pocket and provide information on the binding mode of the ligand. (C and D) STD NMR enabled epitope mapping of the compound A-61603 to the α1A- and α1B-adrenergic receptors. (C) A histogram of normalized STD NMR build-up rates observed upon binding of A-61603 to α1Aor to α1B. Differences in the normalized STD rates between the different chemical groups indicate which chemical groups of the molecule interact more closely with the receptor, and differences observed between the α1A or to α1B receptors indicates selectivity of the drug for a specific receptor subtype. (D) Chemical structure of A-61603 with numbered chemical groups color coded to correspond with the observed STD NMR signals. Panels C and D adapted from reference [19]
4.2. NMR identification of lead compounds by saturation transfer methods
A second general approach to identification of GPCR-binding compounds is the application of saturation transfer methods, which benefit from detection of ligand binding through observing changes in signals of small molecules that bind a target protein. One of the more popular approaches is saturation transfer difference (STD) NMR. In STD NMR experiments, 1H detected spectra are recorded with samples containing unlabeled receptor and mixtures of compounds. NMR spectra are then recorded with and without radio frequency (RF) irradiation applied to the protein. If a compound binds to the protein while RF irradiation is applied, magnetization is transferred from the protein to the small molecule, resulting in a decrease of signals from the interaction. Positive “hits” are detected by subtracting the two spectra with and without applied irradiation, resulting in a positive difference for NMR signals from compounds that bind to the protein [77, 78] (Figure 5B). Typically applied RF irradiation frequencies are selected to correspond with the chemical shifts of buried methyl groups of the protein, which are well separated from chemical shifts of the small molecules. A particular strength of STD NMR is the ability to detect binding of small molecules that interact weakly with their target protein and are generally applied to samples with ligands in molar excess of target proteins, requiring relatively little amounts of receptor. Thus, STD NMR is a powerful technique for fragment-based small molecule screening campaigns [72].
In screening campaigns, STD NMR data can be combined with water-ligand observed via gradient spectroscopy (Water-LOGSY) experiments [72, 79]. In the Water-LOGSY experiment, magnetization from bulk water is transferred to the protein-ligand complex. In spectra of the unbound ligand, signals from compounds that interact with the protein have an opposite sign than signals that did not interact with the protein. Water-LOGSY has been applied to identify ligands of the chemokine receptor CXCR4 [28].
Saturation transfer methods can also be used to quantify ligand binding affinities [75] and to map points of interaction between ligands and receptors [19, 77, 78, 80] (Figure 5 B, and D). The magnitude of the STD NMR signal depends on the proximity of the local chemical moiety to the receptor binding pocket. 1H nuclei from regions of a molecule bound deep in a binding pocket will show a larger STD signal than 1H nuclei from regions that only partially interact with the receptor. Thus, quantification of relative STD NMR signals can provide information on ligand binding epitopes. In this way, STD NMR has been applied to complete structural descriptions of ligand-GPCR interactions [19, 69, 74], such as in the case of polypeptide-GPCR complexes [27, 28, 32]. Data combined from epitope mapping, NMR structures of the bound ligand, and structure-activity-relationship (SAR) studies can inform the design criteria to develop new drugs by modifying fragment-ligands identified by NMR screening or other methods [2, 18, 39, 40, 70, 71].
A recent and notable application of STD NMR by Jawhari, Krimm, and colleagues highlights the potential strength of NMR as a tool for small molecule drug discovery with native receptors [69]. Applying STD NMR to the full length human A2A adenosine receptor (Figure 6), the authors identify novel A2AAR small molecule antagonists and validate their function with orthogonal techniques. An especially novel aspect of this approach was the use of full length A2AAR, enabled by solubilizing the receptor in an optimized mixture of detergents (Figure 6) [69]. This approach established a proof-of-principle method for applying NMR techniques to native receptors in a drug discovery pipeline.
Figure 6.
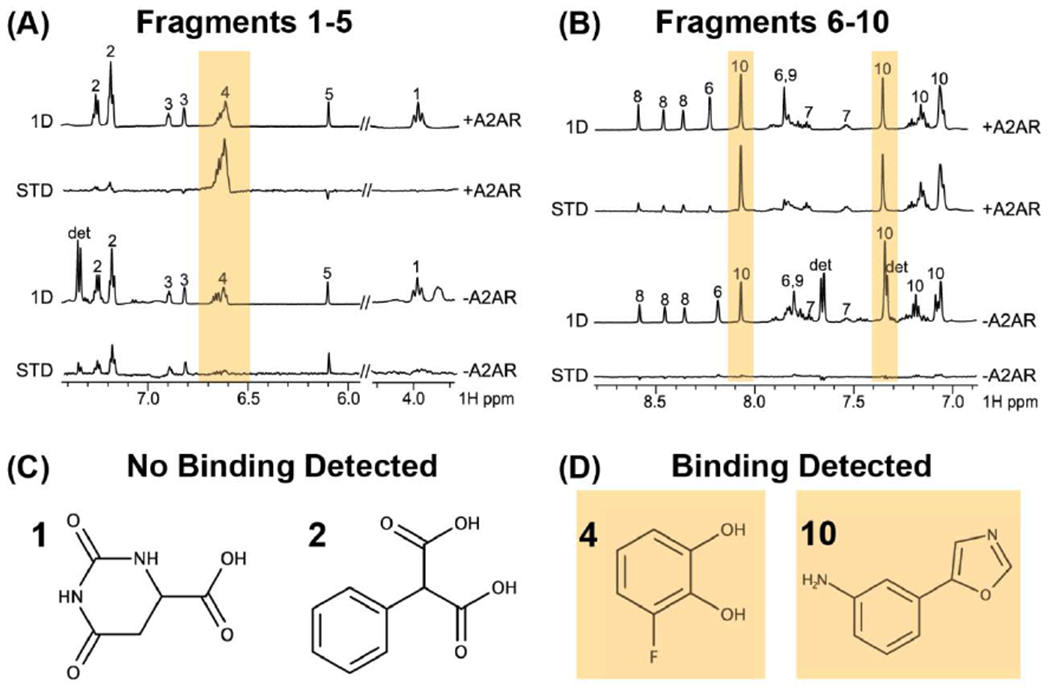
NMR identification of small molecule fragments that bind the full length human GPCR A2AAR. (A and B) 1-dimensional 1H NMR spectra of solutions containing mixtures of small molecules recorded in the presence and absence of the receptor. Saturation transfer different (STD) data were obtained from subtracting spectra recorded with and without RF irradiation applied to the target receptor. Yellow bars highlight signals detected in the STD NMR data that indicate the observation of the small molecule binding to the receptor. (C and D) Chemical structures of molecules that were observed to not bind A2AAR (C) and to bind A2AAR (D) identified from the STD NMR experiments. Figure was adapted with permission from reference [69] under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/)
5. Conclusions and future outlook
NMR studies in solution will continue to contribute valuable information on GPCR drugs, complementing structure determination of GPCR complexes by x-ray crystallography and cryoEM. Integration of structural data from NMR, x-ray crystallography and cryoEM will enable a more expanded view of the chemical space of GPCR ligands than with any one technique alone. NMR spectroscopic studies of GPCR ligands may open new lines of research and allow scientists to gain much needed initial insight into receptors that have been especially challenging targets of structural biologists, including orphan receptors. Emerging solid-state NMR technologies that enable detection of small quantities of GPCR ligands, including 1H detection and DNP NMR continue to evolve, and it is exciting to anticipate future in situ applications of these techniques with studies of human GPCRs in intact cells or even tissues derived from patients. It is a source of enthusiasm to forecast that the combination of novel NMR, x-ray crystallography and cryoEM methods will provide unprecedented insight into the dynamic interplay between the interactions of GPCRs with orthosteric ligands, allosteric modulators, signaling protein partners and all the constituents of the complex cellular environment.
Structure determination of hormones and polypeptides bound to human GPCRs by NMR spectroscopy
New methods enable studies of native receptors in endogenous environments
Methods for GPCR lead compound identification by NMR spectroscopy in solution
6. Acknowledgements
The authors wish to acknowledge funding from the National Institutes of Health (Maximizing Investigators’ Research Award [MIRA] R35, 1R35GM138291) and from the UF College of Liberal Arts and Sciences and the University of Florida.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References.
- [1].Hauser AS, Attwood MM, Rask-Andersen M, Schiöth HB, Gloriam DE, Trends in GPCR drug discovery: new agents, targets and indications, Nat. Rev. Drug Discov 16(12) (2017) 829–842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Wacker D, Stevens RC, Roth BL, How ligands illuminate GPCR molecular pharmacology, Cell 170(3) (2017)414–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Manglik A, Kim TH, Masureel M, Altenbach C, Yang Z, Hilger D, Lerch MT, Kobilka TS, Thian FS, Hubbell WL, Prosser RS, Kobilka BK, Structural insights into the dynamic process of β2-adrenergic receptor signaling, Cell 161(5) (2015) 1101–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Nygaard R, Zou Y, Dror RO, Mildorf TJ, Arlow DH, Manglik A, Pan AC, Liu CW, Fung JJ, Bokoch MP, Thian FS, Kobilka TS, Shaw DE, Mueller L, Prosser RS, Kobilka BK, The dynamic process of β2-adrenergic receptor activation, Cell 152(3) (2013) 532–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Shiraishi Y, Natsume M, Kofuku Y, Imai S, Nakata K, Mizukoshi T, Ueda T, Iwaï H, Shimada I, Phosphorylation-induced conformation of β2-adrenoceptor related to arrestin recruitment revealed by NMR, Nat. Commun 9(1) (2018) 194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Kofuku Y, Ueda T, Okude J, Shiraishi Y, Kondo K, Mizumura T, Suzuki S, Shimada I, Functional dynamics of deuterated β2-adrenergic receptor in lipid bilayers revealed by NMR spectroscopy, Angew. Chem. Int. Ed 53(49) (2014) 13376–9. [DOI] [PubMed] [Google Scholar]
- [7].Eddy MT, Didenko T, Stevens RC, Wüthrich K, β2-adrenergic receptor conformational response to fusion protein in the third intracellular loop, Structure 24(12) (2016) 2190–2197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Horst R, Liu JJ, Stevens RC, Wüthrich K, β2-adrenergic receptor activation by agonists studied with 19F NMR spectroscopy, Angew. Chem. Int. Ed. Engl 52(41) (2013) 10762–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Liu JJ, Horst R, Katritch V, Stevens RC, Wüthrich K, Biased signaling pathways in β2-adrenergic receptor characterized by 19F-NMR, Science 335(6072) (2012) 1106–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Eddy MT, Lee MY, Gao ZG, White KL, Didenko T, Horst R, Audet M, Stanczak P, McClary KM, Han GW, Jacobson KA, Stevens RC, Wüthrich K, Allosteric coupling of drug binding and intracellulars signaling in the A2A adenosine receptor, Cell 172(1-2) (2018) 68–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Ye L, Neale C, Sljoka A, Lyda B, Pichugin D, Tsuchimura N, Larda ST, Pomès R, García AE, Ernst OP, Sunahara RK, Prosser RS, Mechanistic insights into allosteric regulation of the A2A adenosine G protein-coupled receptor by physiological cations., Nature Comm. 9(1) (2018)1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Ye L, Van Eps N, Zimmer M, Ernst OP, Prosser RS, Activation of the A2A adenosine G-protein-coupled receptor by conformational selection, Nature 533(7602) (2016) 265–8. [DOI] [PubMed] [Google Scholar]
- [13].Clark LD, Dikiy I, Chapman K, Rödström KE, Aramini J, LeVine MV, Khelashvili G, Rasmussen SG, Gardner KH, Rosenbaum DM, Ligand modulation of sidechain dynamics in a wild-type human GPCR, Elife 6 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Sušac L, Eddy MT, Didenko T, Stevens RC, Wüthrich K, A2A adenosine receptor functional states characterized by 19F-NMR., Proc. Natl. Acad. Sci. USA 115(50) (2018) 12733–12738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Eddy MT, Gao Z-G, Mannes P, Patel N, Jacobson KA, Katritch V, Stevens RC, Wüthrich K, Extrinsic tryptophans as NMR probes of allosteric coupling in membrane proteins: application to the A2A adenosine receptor, J. Am. Chem. Soc 140(26) (2018) 8228–8235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Sounier R, Mas C, Steyaert J, Laeremans T, Manglik A, Huang W, Kobilka BK, Déméné H, Granier S, Propagation of conformational changes during μ-opioid receptor activation, Nature 524(7565) (2015) 375–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Okude J, Ueda T, Kofuku Y, Sato M, Nobuyama N, Kondo K, Shiraishi Y, Mizumura T, Onishi K, Natsume M, Maeda M, Tsujishita H, Kuranaga T, Inoue M, Shimada I, Identification of a conformational equilibrium that determines the efficacy and functional selectivity of the μ-opioid receptor, Angew. Chem. Int. Ed. Engl 54(52) (2015) 15771–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Shimada I, Ueda T, Kofuku Y, Eddy MT, Wüthrich K, GPCR drug discovery: integrating solution NMR data with crystal and cryo-EM structures., Nat. Rev. Drug Disc 18(1) (2019) 59–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Yong KJ, Vaid TM, Shilling PJ, Wu FJ, Williams LM, Deluigi M, Pluckthun A, Bathgate RAD, Gooley PR, Scott DJ, Determinants of ligand subtype-selectivity at alpha1A-adrenoceptor revealed using saturation transfer difference (STD) NMR, ACS Chem Biol 13(4) (2018) 1090–1102. [DOI] [PubMed] [Google Scholar]
- [20].Miller RL, Thompson AA, Trapella C, Guerrini R, Malfacini D, Patel N, Han GW, Cherezov V, Caló G, Katritch V, Stevens RC, The ilmportance of ligand-receptor conformational pairs in stabilization: spotlight on the N/OFQ G protein-coupled receptor, Structure 23(12) (2015) 2291–2299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Zhang X, Stevens R, X. F, The importance of ligands for G protein-coupled receptor stability., Trends Biochem. Sci 40 (2015) 79–87. [DOI] [PubMed] [Google Scholar]
- [22].Ratnala VRP, Kiihne SR, Buda F, Leurs R, de Groot HJM, DeGrip WJ, Solid-State NMR evidence for a protonation switch in the binding pocket of the HI receptor upon binding of the agonist histamine, J. Am. Chem. Soc 129 (2007) 867–872. [DOI] [PubMed] [Google Scholar]
- [23].Catoire LJ, Damian M, Giusti F, Martin A, van Heijenoort C, Popot JL, Guittet E, Baneres JL, Structure of a GPCR ligand in its receptor-bound state: leukotriene B4 adopts a highly constrained conformation when associated to human BLT2, J. Am. Chem. Soc 132(26) (2010) 9049–57. [DOI] [PubMed] [Google Scholar]
- [24].Giusti F, Casiraghi M, Point E, Damian M, Rieger J, Bon CL, Pozza A, Moncoq K, Banères J-L, Catoire LJ, Structure of the agonist 12–HHT in its BLT2 receptor-bound state, Sci. Rep 10(1) (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Catoire LJ, Damian M, Baaden M, Guittet É, Banères J-L, Electrostatically-driven fast association and perdeuteration allow detection of transferred cross-relaxation for G protein-coupled receptor ligands with equilibrium dissociation constants in the high-to-low nanomolar range, J. Biomol. NMR 50(3) (2011) 191–195. [DOI] [PubMed] [Google Scholar]
- [26].Park SH, Berkamp S, Radoicic J, De Angelis AA, Opella SJ, Interaction of Monomeric Interleukin-8 with CXCR1 Mapped by Proton-Detected Fast MAS Solid-State NMR, Biophys. J 113(12) (2017) 2695–2705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Ferré G, Louet M, Saurel O, Delort B, Czaplicki G, M’Kadmi C, Damian M, Renault P, Cantel S, Gavara L, Demange P, Marie J, Fehrentz JA, Floquet N, Milon A, Baneres JL, Structure and dynamics of G protein-coupled receptor-bound ghrelin reveal the critical role of the octanoyl chain, Proc Natl Acad Sci USA 116(35) (2019) 17525–17530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Brancaccio D, Diana D, Di Maro S, Di Leva FS, Tomassi S, Fattorusso R, Russo L, Scala S, Trotta AM, Portella L, Novellino E, Marinelli L, Carotenuto A, Ligand-based NMR study of C-X-C chemokine receptor type 4 (CXCR4)-ligand interactions on living cancer cells, J. Med. Chem 61(7) (2018) 2910–2923. [DOI] [PubMed] [Google Scholar]
- [29].Joedicke L, Mao J, Kuenze G, Reinhart C, Kalavacherla T, Jonker HRA, Richter C, Schwalbe H, Meiler J, Preu J, Michel H, Glaubitz C, The molecular basis of subtype selectivity of human kinin G-protein-coupled receptors, Nat Chem Biol 14(3) (2018) 284–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Lopez JJ, Shukla AK, Reinhart C, Schwalbe H, Michel H, Glaubitz C, The structure of the neuropeptide bradykinin bound to the human G-protein coupled receptor bradykinin B2 as determined by solid-state NMR spectroscopy, Angew. Chem. Int. Ed. Engl 47(9) (2008) 1668–71. [DOI] [PubMed] [Google Scholar]
- [31].Luca S, White JF, Sohal AK, Filippov DV, van Boom JH, Grisshammer R, Baldus M, The conformation of neurotensin bound to its G protein-coupled receptor., Proc Natl Acad Sci U S A 100 (2003) 10706–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Bender BJ, Vortmeier G, Ernicke S, Bosse M, Kaiser A, Els-Heindl S, Krug U, Beck-Sickinger A, Meiler J, Huster D, Structural model of ghrelin bound to its G protein-coupled receptor, Structure 27(3) (2019) 537–544 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Kaiser A, Muller P, Zellmann T, Scheidt HA, Thomas L, Bosse M, Meier R, Meiler J, Huster D, Beck-Sickinger AG, Schmidt P, Unwinding of the C-Terminal Residues of Neuropeptide Y is critical for Y(2) Receptor Binding and Activation, Angew. Chem. Int. Ed. Engl 54(25) (2015) 7446–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Yang Z, Han S, Keller M, Kaiser A, Bender BJ, Bosse M, Burkert K, Kogler LM, Wifling D, Bernhardt G, Plank N, Littmann T, Schmidt P, Yi C, Li B, Ye S, Zhang R, Xu B, Larhammar D, Stevens RC, Huster D, Meiler J, Zhao Q, Beck-Sickinger AG, Buschauer A, Wu B, Structural basis of ligand binding modes at the neuropeptide Y Y1 receptor, Nature 556(7702) (2018) 520–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Inooka H, Ohtaki T, Kitahara O, Ikegami T, Endo S, Kitada C, Ogi K, Onda H, Fujino M, Shirakawa M, Conformation of a peptide ligand bound to its G-protein coupled receptor, Nat. Struct. Mol. Biol 8 (2001) 161–165. [DOI] [PubMed] [Google Scholar]
- [36].Wu F, Song G, de Graaf C, Stevens RC, Structure and Function of Peptide-Binding G Protein-Coupled Receptors., J. Mol. Biol 429 (2017) 2726–2745. [DOI] [PubMed] [Google Scholar]
- [37].O’Connor C, White KL, Doncescu N, Didenko T, Roth BL, Czaplicki G, Stevens RC, Wdthrich K, Milon A, NMR structure and dynamics of the agonist dynorphin peptide bound to the human kappa opioid receptor, Proc. Natl. Acad. Sci. U. S. A 112(38) (2015) 11852–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Congreve M, Andrews SP, Dore AS, Hollenstein K, Hurrell E, Langmead CJ, Mason JS, Ng IW, Tehan B, Zhukov A, Weir M, Marshall FH, Discovery of 1,2,4-triazine derivatives as adenosine A2A antagonists using structure based drug design, J. Med. Chem 55(5) (2012) 1898–903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Christopher JA, Aves SJ, Bennett KA, Dore AS, Errey JC, Jazayeri A, Marshall FH, Okrasa K, Serrano-Vega MJ, Tehan BG, Wiggin GR, Congreve M, Fragment and Structure-Based Drug Discovery for a Class C GPCR: Discovery of the mGlu5 Negative Allosteric Modulator HTL14242 (3-Chloro-5-[6-(5-fluoropyridin-2-yl)pyrimidin-4-yl]benzonitrile), J. Med. Chem 58(16) (2015) 6653–64. [DOI] [PubMed] [Google Scholar]
- [40].Andrews SP, Brown GA, Christopher JA, Structure-based and fragment-based GPCR drug discovery, ChemMedChem 9(2) (2014) 256–75. [DOI] [PubMed] [Google Scholar]
- [41].Pandy-Szekeres G, Munk C, Tsonkov TM, Mordalski S, Harpsoe K, Hauser AS, Bojarski AJ, Gloriam DE, GPCRdb in 2018: adding GPCR structure models and ligands, Nucleic Acids Res. 46(D1) (2018) D440–D446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Wu F, Song G, de Graaf C, Stevens RC, Structure and function of peptide-binding G protein-coupled receptors, J. Mol. Biol 429(17) (2017) 2726–2745. [DOI] [PubMed] [Google Scholar]
- [43].Davenport AP, Scully CCG, Graaf C, Brown AJH, Maguire JJ, Advances in therapeutic peptides targeting G protein-coupled receptors, Nat. Rev. Drug Disc (2020) 1–25. [DOI] [PubMed] [Google Scholar]
- [44].Clore GM, Gronenborn AM, Theory and applications of the transferred nuclear Overhauser effect to the study of the conformations of small ligands bound to proteins, J. Mag. Res 48(3) (1982) 402–417. [Google Scholar]
- [45].Vogeli B, The nuclear Overhauser effect from a quantitative perspective, Prog. Nucl. Magn. Reson. Spectrosc 78 (2014) 1–46. [DOI] [PubMed] [Google Scholar]
- [46].Ferré G, Czaplicki G, Demange P, Milon A, Structure and dynamics of dynorphin peptide and its receptor, Vitam Horm 111 (2019) 17–47. [DOI] [PubMed] [Google Scholar]
- [47].Müller TD, Nogueiras R, Andermann ML, Andrews ZB, Anker SD, Argente J, Batterham RL, Benoit SC, Bowers CY, Broglio F, Casanueva FF, D'Alessio D, Depoortere I, Geliebter A, Ghigo E, Cole PA, Cowley M, Cummings DE, Dagher A, Diano S, Dickson SL, Diéguez C, Granata R, Grill HJ, Grove K, Habegger KM, Heppner K, Heiman ML, Holsen L, Holst B, Inui A, Jansson JO, Kirchner H, Korbonits M, Laferrère B, LeRoux CW, Lopez M, Morin S, Nakazato M, Nass R, Perez-Tive D, Pfluger PT, Schwartz TW, Seeley RJ, Sleeman M, Sun Y, Sussel L, Tong J, Thorner MO, van der Lely AJ, van der Ploeg LHT, Zigman JM, Kojima M, Kangawa K, Smith RG, Horvath T, Tschop MH, Ghrelin., Molecular metabolism 4(6) (2015) 437–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].White JF, Noinaj N, Shibata Y, Love J, Kloss B, Xu F, Gvozdenovic-Jeremic J, Shah P, Shiloach J, Tate CG, Grisshammer R, Structure of the agonist-bound neurotensin receptor, Nature 490(7421) (2012) 508–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Shen Y, Delaglio F, Cornilescu G, Bax A, TALOS+: a hybrid method for predicting protein backbone torsion angles from NMR chemical shifts, J. Biomol. NMR 44(4) (2009) 213–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Shen Y, Bax A, Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks, J. Biomol. NMR 56(3) (2013) 227–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Berjanskii MV, Neal S, Wishart DS, PREDITOR: a web server for predicting protein torsion angle restraints, Nucleic Acids Res. 34(2) (2006) W63–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Neal S, Nip AM, Zhang H, Wishart DS, Rapid and accurate calculation of protein 1H, 13C and 15N chemical shifts, J. Biomol. NMR 26 (2003) 215–240. [DOI] [PubMed] [Google Scholar]
- [53].Meiler J, PROSHIFT: Protein chemical shift prediction using artificial neural networks, J. Biomol. NMR 26 (2003) 25–37. [DOI] [PubMed] [Google Scholar]
- [54].Penzel S, Oss A, Org M-L, Samoson A, Böckmann A, Ernst M, Meier BH, Spinning faster: protein NMR at MAS frequencies up to 126 kHz., J. Biomol. NMR 73(1-2) (2019) 19–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Struppe J, Quinn CM, Lu M, Wang M, Hou G, Lu X, Kraus J, Andreas LB, Stanek J, Lalli D, Lesage A, Pintacuda G, Maas W, Gronenborn AM, Polenova T, Expanding the horizons for structural analysis of fully protonated protein assemblies by NMR spectroscopy at MAS frequencies above 100&nbsp;kHz, Solid State Nucl. Magn. Reson 87 (2017) 117–125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Schubeis T, Le Marchand T, Andreas LB, Pintacuda G, 1H magic-angle spinning NMR evolves as a powerful new tool for membrane proteins, J. Mag. Res 287 (2018) 140–152. [DOI] [PubMed] [Google Scholar]
- [57].Liu K, Wu L, Yuan S, Wu M, Xu Y, Sun Q, Li S, Zhao S, Hua T, Liu Z.-j, Structural basis of CXC chemokine receptor 2 activation and signalling., Nature 1 (2020) 95–6. [DOI] [PubMed] [Google Scholar]
- [58].Bajaj VS, Mak-Jurkauskas ML, Belenky M, Herzfeld J, Griffin RG, Functional and shunt states of bacteriorhodopsin resolved by 250 GHz dynamic nuclear polarization-enhanced solid-state NMR., Proc. Natl. Acad. Sci. USA 106(23) (2009) 9244–9249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Mak-Jurkauskas ML, Bajaj VS, Hornstein MK, Belenky M, Griffin RG, Herzfeld J, Energy transformations early in the bacteriorhodopsin photocycle revealed by DNP-enhanced solid-state NMR., Proc. Natl. Acad. Sci. USA 105(3) (2008) 883–888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Joedicke L, Mao J, Kuenze G, Reinhart C, Kalavacherla T, Jonker HRA, Richter C, Schwalbe H, Meiler J, Preu J, Michel H, Glaubitz C, The molecular basis of subtype selectivity of human kinin G-protein-coupled receptors, Nat. Chem. Biol 14(3) (2018) 284–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Gröbner G, Burnett IJ, Glaubitz C, Choi G, Mason AJ, Watts A, Observations of light-induced structural changes of retinal within rhodopsin, Nature 405(6788) (2000) 810–813. [DOI] [PubMed] [Google Scholar]
- [62].Kimata N, Pope A, Eilers M, Opefi CA, Ziliox M, Hirshfeld A, Zaitseva E, Vogel R, Sheves M, Reeves PJ, Smith SO, Retinal orientation and interactions in rhodopsin reveal a two-stage trigger mechanism for activation., Nature Comm. 7(1) (2016) 12683–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Struts AV, Salgado GFJ, Martínez-Mayorga K, Brown MF, Retinal dynamics underlie its switch from inverse agonist to agonist during rhodopsin activation., Nature Structural and Molecular Biology 18(3) (2011) 392–394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Struts AV, Salgado GFJ, Brown MF, Solid-state 2H NMR relaxation illuminates functional dynamics of retinal cofactor in membrane activation of rhodopsin., Proc. Natl. Acad. Sci. USA 108(20) (2011) 8263–8268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Patel AB, Crocker E, Eilers M, Hirshfeld A, Sheves M, Smith SO, Coupling of retinal isomerization to the activation of rhodopsin., Proc. Natl. Acad. Sci. USA 101(27) (2004) 10048–10053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Carravetta M, Zhao X, Johannessen OG, Lai WC, Verhoeven MA, Bovee-Geurts PHM, Verdegem PJE, Kiihne S, Luthman H, de Groot HJM, DeGrip WJ, Lugtenburg J, Levitt MH, Protein-induced bonding perturbation of the rhodopsin chromophore detected by double-quantum solid-state NMR., J. Am. Chem. Soc 126(12) (2004) 3948–3953. [DOI] [PubMed] [Google Scholar]
- [67].Chen S, Lu M, Liu D, Yang L, Yi C, Ma L, Zhang H, Liu Q, Frimurer TM, Wang M-W, Schwartz TW, Stevens RC, Wu B, Wüthrich K, Zhao Q, Human substance P receptor binding mode of the antagonist drug aprepitant by NMR and crystallography, Nature Comm. (2019) 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Casiraghi M, Damian M, Lescop E, Banères J-L, Catoire LJ, Illuminating the energy landscape of GPCRs: The key contribution of solution-state NMR associated with escherichia colias an expression host, Biochemistry 57(16) (2018) 2297–2307. [DOI] [PubMed] [Google Scholar]
- [69].Igonet S, Raingeval C, Cecon E, Pucic-Bakovic M, Lauc G, Cala O, Baranowski M, Perez J, Jockers R, Krimm I, Jawhari A, Enabling STD-NMR fragment screening using stabilized native GPCR: A case study of adenosine receptor, Sci Rep 8(1) (2018) 8142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Congreve M, Rich RL, Myszka DG, Figaroa F, Siegal G, Marshall FH, Fragment screening of stabilized G protein-coupled receptors using biophysical methods, Methods Enzymol. 493 (2011) 115–36. [DOI] [PubMed] [Google Scholar]
- [71].Chen D, Errey JC, Heitman LH, Marshall FH, Ijzerman AP, Siegal GD, Fragment screening of GPCRs using biophysical methods: identification of ligands of the adenosine A2A receptor with novel biological activity, ACS Chem. Biol 12 (2012) 2064–2073. [DOI] [PubMed] [Google Scholar]
- [72].Erlanson DA, Fesik SW, Hubbard RE, Jahnke W, Jhoti H, Twenty years on: the impact of fragments on drug discovery, Nat Rev Drug Discov 15(9) (2016) 605–619. [DOI] [PubMed] [Google Scholar]
- [73].Assadi-Porter FM , Tonelli M, Maillet E, Hallenga K, Benard O, Max M, J. L M., Direct NMR detection of the binding of functional ligands to the human sweet receptor, a heterodimeric family 3 GPCR, J. Am. Chem. Soc 130 (2008) 7212–7213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Cox BD, Mehta AK, DiRaddo JO, Liotta DC, Wilson LJ, Snyder JP, Structural analysis of CXCR4 - Antagonist interactions using saturation-transfer double-difference NMR, Biochem. Biophys. Res. Commun 466(1) (2015) 28–32. [DOI] [PubMed] [Google Scholar]
- [75].Bartoschek S, Klabunde T, Defossa E, Dietrich V, Stengelin S, Griesinger C, Carlomagno T, Focken I, Wendt KU, Drug design for G-protein-coupled receptors by a ligand-based NMR method, Angew. Chem. Int. Ed. Engl 49(8) (2010) 1426–9. [DOI] [PubMed] [Google Scholar]
- [76].Vanwetswinkel S, Heetebrij R, van Duynhoven J, Hollander J, Filippov D, Hajduk P, Siegal GD, TINS, target immobilized NMR screening: an efficient and sensitive method for ligand discovery., Chem Biol. 12 (2005) 207–16. [DOI] [PubMed] [Google Scholar]
- [77].Mayer M, Meyer B, Group epitope mapping by saturation transfer difference NMR to identify segments of a ligand in direct contact with a protein receptor, J. Am. Chem. Soc 123(25) (2001) 6108–17. [DOI] [PubMed] [Google Scholar]
- [78].Mayer M, Meyer B, Characterization of ligand binding by saturation transfer difference NMR spectroscopy, Angew. Chem. Int. Ed 38 (1999) 1784–1788. [DOI] [PubMed] [Google Scholar]
- [79].Dalvit C, Pevarello P, Tatò M, Veronesi M, Vulpetti A, Sundström M, Identification of compounds with binding affinity to proteins via magnetization transfer from bulk water., J. Biomol. NMR 18 (2000) 65–68. [DOI] [PubMed] [Google Scholar]
- [80].Cala O, Krimm I, Ligand-orientation based fragment selection in STD NMR screening, J. Med. Chem 58(21) (2015) 8739–42. [DOI] [PubMed] [Google Scholar]
- [81].Sauvée C, Rosay M, Casano G, Aussenac F, Weber RT, Ouari O, Tordo P, Highly efficient, water-soluble polarizing agents for dynamic nuclear polarization at high frequency, Angew. Chem. Int. Ed 52(41) (2013) 10858–10861. [DOI] [PubMed] [Google Scholar]