

Fingolimod (FTY720), an FDA-approved immunomodulatory drug for treating multiple sclerosis, is an agonist of sphingosine-1-phosphate receptor (S1PR), which has been used as a research tool for inhibiting immune cell trafficking. FTY720 was recently reported to inhibit Chlamydia dissemination. Since genital Chlamydia spreading to the gastrointestinal tract correlated with its pathogenicity in the upper genital tract, we evaluated the effect of FTY720 on chlamydial pathogenicity in the current study.
KEYWORDS: FTY720, fingolimod, chlamydia, dissemination, hydrosalpinx
ABSTRACT
Fingolimod (FTY720), an FDA-approved immunomodulatory drug for treating multiple sclerosis, is an agonist of sphingosine-1-phosphate receptor (S1PR), which has been used as a research tool for inhibiting immune cell trafficking. FTY720 was recently reported to inhibit Chlamydia dissemination. Since genital Chlamydia spreading to the gastrointestinal tract correlated with its pathogenicity in the upper genital tract, we evaluated the effect of FTY720 on chlamydial pathogenicity in the current study. Following an intravaginal inoculation, live chlamydial organisms were detected in mouse rectal swabs. FTY720 treatment significantly delayed live organism shedding in the rectal swabs. However, FTY720 failed to block chlamydial spreading to the gastrointestinal tract. The live chlamydial organisms recovered from rectal swabs reached similar levels between mice with or without FTY720 treatment by day 42 in C57BL/6J and day 28 in CBA/J mice, respectively. Thus, genital Chlamydia is able to launch a 2nd wave of spreading via an FTY720-resistant pathway after the 1st wave of spreading is inhibited by FTY720. As a result, all mice developed significant hydrosalpinx. The FTY720-resistant spreading led to stable colonization of chlamydial organisms in the colon. Consistently, FTY720 did not alter the colonization of intracolonically inoculated Chlamydia. Thus, we have demonstrated that, following a delay in chlamydial spreading caused by FTY720, genital Chlamydia is able to both spread to the gastrointestinal tract via an FTY720-resistant pathway and maintain its pathogenicity in the upper genital tract. Further characterization of the FTY720-resistant pathway(s) explored by Chlamydia for spreading to the gastrointestinal tract may promote our understanding of Chlamydia pathogenic mechanisms.
INTRODUCTION
Chlamydia trachomatis infection is a leading bacterial cause of tubal infertility due to its ability to induce tubal inflammation, leading to sequelae that include adhesion, fibrosis, hydrosalpinx (1–3; https://www.cdc.gov/std/stats16/default.htm). Chlamydia muridarum induction of long-lasting tubal fibrosis and hydrosalpinx in mice (4–8) has been used to investigate the pathological mechanisms of C. trachomatis. Although the C. muridarum mouse model has been useful in identifying pathogenesis determinants from both Chlamydia (9–14) and host (8, 15–18), it remains unknown how the long-lasting tubal fibrosis is maintained after the genital tract infection is cleared. When we used an in vivo imaging approach to monitor chlamydial ascending infection in mice (19), we accidentally found genital Chlamydia spreading to and colonizing in the gastrointestinal (GI) tract for long periods of time (20). This finding has not only logically linked prior observations on chlamydial spreading following mucosal inoculations (21) and persisting in the gut (22) but also suggested a new dimension for understanding chlamydial pathogenic mechanisms in the upper genital tract (23).
Genital chlamydial spreading to the GI tract may promote chlamydial pathogenicity in the upper genital tract. First, the long-lasting hydrosalpinx induced by the self-limited Chlamydia infection in the mouse genital tract correlates with chlamydial long-lasting colonization in the GI tract (8, 20, 24). Second, attenuated Chlamydia mutants are defective in both inducing hydrosalpinx and colonizing the GI tract (25–27). Finally, an intragastric coinoculation with wild-type Chlamydia can successfully rescue an attenuated Chlamydia to induce hydrosalpinx (28). Although it is not known whether gastrointestinal Chlamydia trachomatis may also contribute to chlamydial pathogenicity in women’s upper genital tracts, C. trachomatis has been frequently detected in human GI tracts (29–33). Furthermore, systemic distribution of Chlamydia has also been reported in women (34, 35).
Given the potential medical importance of genital Chlamydia spreading to the GI tract, efforts have been made to reveal the mechanisms of chlamydial spreading. We have previously shown that genital Chlamydia spreading to the GI tract is not caused by vagina-anal contamination or coprophagy but likely dependent on blood circulation (20, 36). Recently, fingolimod (FTY720), an FDA-approved immunomodulatory drug for treating multiple sclerosis (37), was found to inhibit genital Chlamydia spreading to the GI tract (38). FTY720 is a selective agonist of sphingosine-1-phosphate receptor S1PR that down-modulates S1PR (39, 40) and inhibits trafficking of both dendritic cells (41) and lymphocytes (42). The observation of FTY720 inhibition of chlamydial spreading suggests that chlamydial spreading may be dependent on host cell trafficking.
The current study is designed to evaluate whether FTY720 can also impact chlamydial pathogenicity in the upper genital tract since genital Chlamydia spreading to the GI tract promotes chlamydial pathogenicity in the genital tract (23, 28). We found that FTY720 treatment significantly delayed the appearance of genital chlamydial organisms in the GI tracts of both C57BL/6J (C57) and CBA/J (CBA) mice. Since FTY720 did not alter colonization of intracolonically inoculated Chlamydia in the GI tract, the FTY720-mediated delay in chlamydial appearance in the GI tract was probably due to the inhibition of chlamydial spreading to the GI tract. Nevertheless, genital Chlamydia successfully entered the GI tracts of both mouse strains at a later time despite the FTY720 inhibition, suggesting that Chlamydia is able to use an FTY720-resistant pathway for spreading. As a result, both C57BL/6J and CBA/J mice developed significant hydrosalpinx even in the presence of FTY720 inhibition. Thus, we have demonstrated that following a transient delay caused by FTY720, genital Chlamydia can still spread to the GI tract via an FTY720-resistant pathway(s) and maintain its pathogenicity in the upper genital tract. It will be worth further revealing the FTY720-resistant pathway(s) Chlamydia has explored for both spreading to the GI tract and maintaining pathogenicity in the upper genital tract.
RESULTS
FTY720 treatment delays but does not completely block genital chlamydial spreading to the GI tract.
Since FTY720 was reported to prevent genital Chlamydia from spreading to the GI tracts of C57BL/6J mice (38), we evaluated the effect of FYT720 on chlamydial organism distribution in both genital (via vaginal swabs) and GI (rectal swabs) tracts of C57BL/6J mice following an intravaginal inoculation (Fig. 1). The intravaginally inoculated chlamydial organisms appeared in rectal swabs as early as day 7 and plateaued on day 14. The live organism level in the GI tract was maintained throughout the experiment up to day 56, while live organisms were completely cleared from the genital tract by day 35. FTY720 treatment significantly delayed live organism appearance in rectal swabs. Only by day 21 were live organisms detected in rectal swabs. The levels of live organisms were kept significantly lower in rectal swabs of the FTY720-treated mice than those from control mice up to day 35. Nevertheless, starting from day 42, the levels of rectal swab live organisms were no longer different between mice with or without FTY720 treatment. Interestingly, the same FTY720 treatment did not significantly affect chlamydial colonization in the genital tract. It is clear that, despite the significant delay in the appearance of live chlamydial organisms in the GI tract during the first 5 weeks, the intravaginally inoculated chlamydial organisms gradually reached the GI tract and maintained their colonization in the GI tract. To test whether this delay and recovery of spreading the phenotype are unique to C57BL/6J mice, we further evaluated the FTY720 treatment effect using CBA/J mice (Fig. 2). This is because CBA/J mice are far more susceptible to chlamydial colonization than C57BL/6J mice (8). We found that significant spreading of genital Chlamydia to the gastrointestinal tract was detected on day 7 regardless of FTY720 treatment. By day 14, the rectal swab live organisms plateaued in the absence of FTY720 treatment. FTY720 treatment significantly reduced live organism levels in rectal swabs on both days 14 and 21. By day 28, the levels of live chlamydial organisms became similar between mice with or without FTY720 treatment. Again, FTY720 did not significantly alter chlamydial colonization in the genital tract. Thus, we have demonstrated in two different mouse strains that, following a delay caused by FTY720 treatment, genital Chlamydia is able to spread to the GI tract using an FTY720-resistant pathway and maintain its colonization in the GI tract.
FIG 1.
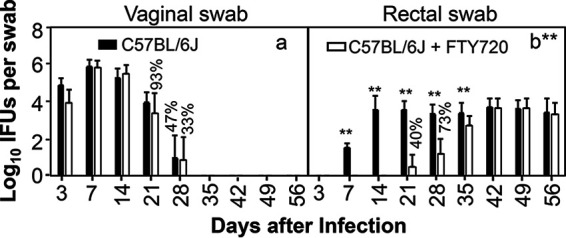
FTY720 significantly delays genital Chlamydia spreading to the gastrointestinal tract in C57BL/6J mice. Groups of female C57BL/6J mice (n = 15) infected intravaginally with Chlamydia were treated without (solid bar) or with (open bar) FTY720 throughout the experiment. All mice were monitored for live chlamydial organism shedding from both vaginal (a) and rectal (b) swabs, and the titers are expressed as log10 inclusion-forming units (IFUs) per swab (y axis) on different days after intravaginal infection (x axis). The percentages of mice with positive detection of chlamydial organisms in swab samples are indicated on top of the bars when the percentages in the corresponding groups are less than 100%. Note that FTY720 treatment significantly reduced spreading of genital Chlamydia to gastrointestinal tract. n = 15; data are from two independent experiments. **P < 0.01 (Wilcoxon rank sum).
FIG 2.
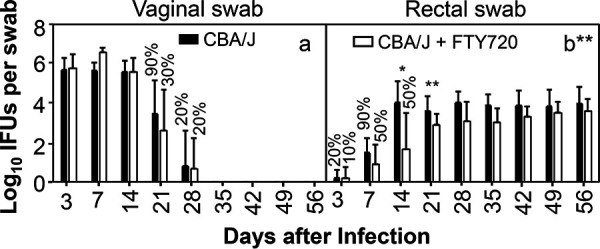
FTY720 significantly delays genital Chlamydia spreading to the gastrointestinal tract in CBA/J mice. Groups of female CBA/J mice (n = 10) infected intravaginally with Chlamydia were treated without (solid bar) or with (open bar) FTY720 throughout the experiment. All mice were monitored for live chlamydial organism shedding from both vaginal (a) and rectal (b) swabs, and the titers were expressed as log10 inclusion-forming units (IFUs) per swab (y axis) on different days after intravaginal infection (x axis). The percentages of mice with positive detection of chlamydial organisms in swab samples are indicated on top of the bars when the percentages in the corresponding groups are less than 100%. Note that FTY720 treatment significantly reduced spreading of genital Chlamydia to gastrointestinal tract. n = 10; data are from two independent experiments. *, P < 0.05; **, P < 0.01 (Wilcoxon rank sum).
FTY720-delayed chlamydial spreading does not affect chlamydial pathogenicity in the upper genital tract.
Since FTY720 only delayed but did not completely block genital Chlamydia spreading to the gastrointestinal tracts of both C57 and CBA mice (see above), we further evaluated the effect of FTY720 treatment on chlamydial induction of pathology in the upper genital tracts of the same two strains of mice (Fig. 3 and 4). We found that 53% of C57BL/6J mice developed significant hydrosalpinx on day 56 following an intravaginal inoculation. After FTY720 treatment, significant hydrosalpinx was still induced in 40% of mice (Fig. 3). The reduction in both hydrosalpinx rate and severity was not statistically significant. We also evaluated the impact of FTY720 on chlamydial pathogenicity in CBA/J mice (Fig. 4). Significant hydrosalpinx was induced in 60% of CBA mice. In the presence of FTY720 treatment, hydrosalpinx was induced in 80% of mice. Neither the hydrosalpinx incidence nor severity was significantly different between mice with or without FTY720 treatment. Thus, we have demonstrated that although FTY720 treatment significantly delayed genital Chlamydia spreading to the GI tract, FTY720 did not significantly impact chlamydial pathogenicity in the upper genital tract.
FIG 3.
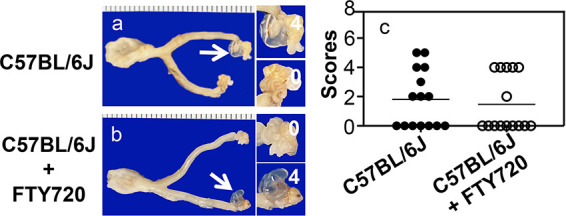
FTY720 fails to alter Chlamydia induction of hydrosalpinx in upper genital tracts of C57BL/6J. (a and b) The same two groups of C57BL/6J mice (n = 15) as described in the Fig. 1 legend were sacrificed for observing genital pathology of the hydrosalpinx on day 56 after intravaginal infection. One gross image of the genital tract tissue is presented from each group, with vagina on the left and oviduct/ovary on the right. The hydrosalpinx is marked with a white arrow. The magnified oviduct/ovary images on the right are marked with the corresponding hydrosalpinx scores. (c) Hydrosalpinx severity scores are plotted. n = 15; data are from two independent experiments.
FIG 4.
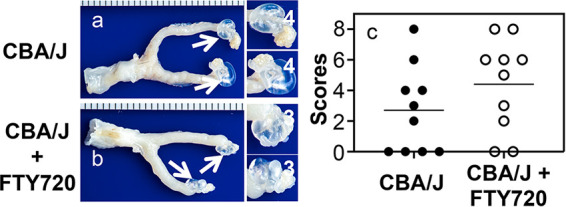
FTY720 fails to alter Chlamydia induction of hydrosalpinx in the upper genital tracts of CBA/J mice. (a and b) The same two groups of CBA/J mice (n = 10) as described in the Fig. 1 legend were sacrificed for observing the hydrosalpinx on day 56 after intravaginal infection. One gross image of the genital tract tissue is presented from each group, with vagina on the left and oviduct/ovary on the right. Hydrosalpinx was marked with a white arrow. The magnified oviduct/ovary images on the right are marked with the corresponding hydrosalpinx scores. (c) Hydrosalpinx severity scores are plotted. n = 10; data are from two independent experiments.
FTY720 delays genital Chlamydia spreading to GI tract tissues without affecting chlamydial colonization in large intestine.
We further monitored tissue distribution of Chlamydia following an intravaginal inoculation in mice with or without FTY720 treatment (Fig. 5). Although intravaginally inoculated Chlamydia spread to spleen and both small and large intestinal tissues on day 4 after inoculation, FTY720 treatment completely blocked the spreading. By day 7, 1 of 4 FTY720-treated mice allowed spreading of genital Chlamydia to both the spleen and GI tract. By day 56, all mice were equally susceptible to chlamydial colonization in large intestinal tissues regardless of FTY720 treatment. Thus, genital Chlamydia is able to overcome FTY720 inhibition to eventually regain spreading to the GI tract and to establish stable colonization in the colon. We further tested whether FTY720-induced delay in genital Chlamydia spreading to the large intestine is due to its direct inhibition of chlamydial replication in the large intestine (Fig. 6). We found that FTY720 treatment did not significantly affect chlamydial colonization in the GI tract following an intracolonic inoculation.
FIG 5.
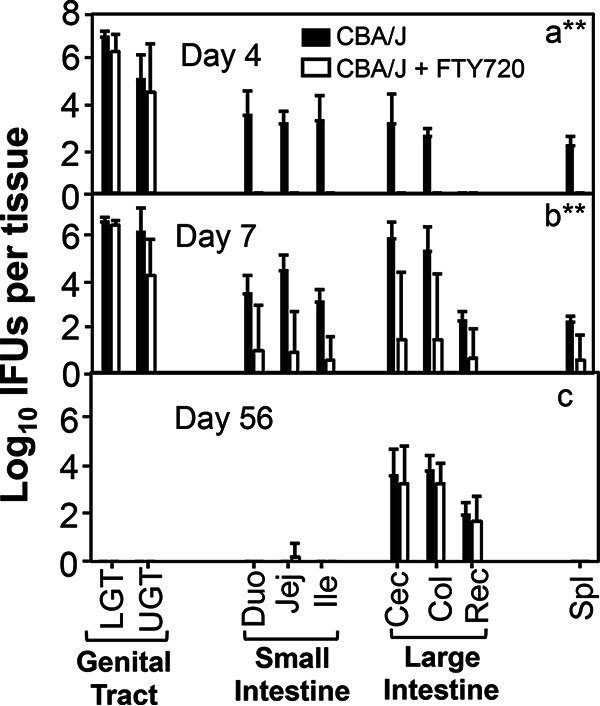
FTY720 significantly delayed genital Chlamydia spreading to the gastrointestinal tract and systemically in CBA/J mice. Groups of female CBA/J mice (n = 4) infected intravaginally with Chlamydia were treated without (solid bar) or with (open bar) FTY720. On days 4 (a), 7 (b), and 56 (c), groups of mice (n = 4) were sacrificed for measuring live chlamydial organisms from different tissues as shown along the x axis. The tissues include lower genital (LGT) and upper genital (UGT) tracts, small intestine duodenum (Duo), Jejunum (Jej), ilium (Ili), and large intestine tissues cecum (Cec), colon (Col), rectum (Rec), as well as spleen tissue (Spl). The live chlamydial organism titers were expressed as log10 IFUs per tissue (y axis). Note that FTY720 treatment significantly reduced spreading of genital Chlamydia to the gastrointestinal tract tissue on days 4 and 7 but not 56. n = 4; **P < 0.01 (Wilcoxon rank sum).
FIG 6.
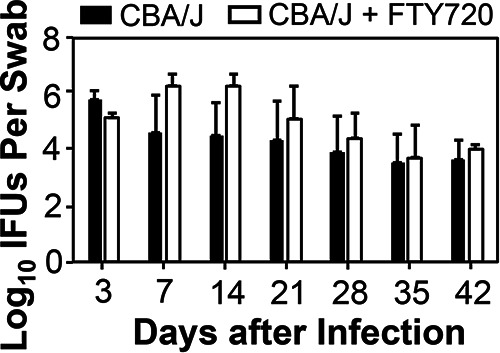
FTY720 treatment fails to impact Chlamydia colonization in the large intestine. Groups of female CBA/J mice (n = 4) infected via an intracolonic inoculation with Chlamydia were treated without (solid bar) or with (open bar) FTY720 on days 3 and 7 and weekly thereafter postinfection, as listed along the x axis. All mice were monitored for live chlamydial organism shedding from swabs. The live chlamydial organism titers were expressed as log10 IFUs per swab (y axis). n = 4; P > 0.05 (Wilcoxon rank sum).
DISCUSSION
Despite extensive efforts, the precise mechanisms by which Chlamydia induces long-lasting pathology in the upper genital tracts remain unclear (1–4, 6, 8). Recent studies have led to the proposal of a 2-hit model for chlamydial pathogenicity in the upper genital tract (23). Lower genital Chlamydia may both ascend to infect tubal epithelial cells, causing the 1st hit, and spread to GI tract to induce a pathogenic response as a 2nd hit. Thus, efforts have been made to investigate the mechanisms by which genital Chlamydia spreads to the GI tract. A recent study has revealed that FTY720 is able to inhibit genital Chlamydia from spreading to the GI tract (38), suggesting that chlamydial spreading may be dependent on host immune cell trafficking. This is because FTY720 is a selective agonist of sphingosine-1-phosphate receptor (S1PR) and can inhibit trafficking of both dendritic cells (41) and lymphocytes (42) via down-modulating S1PR (39, 40).
In the current study, we evaluated the effect of FTY720 on chlamydial pathogenicity. First, we reproduced a previous observation that FTY720 treatment inhibited genital Chlamydia spreading to the GI tracts of C57BL/6J mice within 2 weeks after intravaginal infection (38). However, after we extended the observation beyond 2 weeks, we found that this inhibition was not sustainable. Instead, FTY720 only delayed the appearance of live chlamydial organisms in the gastrointestinal tracts. For example, by day 21, live Chlamydia still reached the GI tract despite the FTY720 treatment. By day 42, the levels of live chlamydial organisms recovered from the GI tract became similar between C57BL/6J mice with or without FTY720 treatment. Thus, we have demonstrated for the first time that Chlamydia can spread using an FTY720-resistant pathway. Second, chlamydial spreading to the GI tract via both FTY720-susceptible and FTY720-resistant pathways was reproduced in CBA/J mice. Interestingly, FTY720 inhibition of chlamydial spreading was less effective in CBA/J mice, suggesting that this mouse strain might permit more extensive chlamydial spreading via the FTY720-resistant pathway. Nevertheless, observations from both strains of mice have consistently shown that genital Chlamydia spread to the GI tract using an FTY720-sensitive or -susceptible pathway first, which then was followed by the FTY720-resistant pathway(s). Third, FTY720 did not significantly impact chlamydial pathogenicity in the upper genital tract. Both C57BL/6J and CBA/J mice developed significant hydrosalpinx regardless of FTY720 inhibition. These observations suggest that chlamydial spreading to the GI tract via the FTY720-resistant pathway is sufficient for exacerbating genital tract pathology. Finally, we further monitored chlamydial organism distribution at tissue levels and confirmed that genital Chlamydia was able to spread to the GI tract via an FTY720-resistant pathway following the delay caused by FTY720. Despite continuous FTY720 treatment, live chlamydial organisms recovered from large intestinal tissues on day 56 after the initial intravaginal infection reached a level similar to that of mice without FTY720 treatment. Thus, we have demonstrated that following a transient delay caused by FTY720, genital Chlamydia can still spread to the gastrointestinal tract via an FTY720-resistant pathway and maintain its pathogenicity in the upper genital tract.
There are obvious limitations to using FTY720 to probe pathogen dissemination pathways. FTY720 is an FDA-approved immunomodulatory drug that functions as a selective agonist of S1PR (39, 40). The therapeutic effect of FYT720 is attributed to its ability to internalize the Gi-coupled S1PR on lymphocytes. As a result, these cells become unresponsive to the natural ligand S1P and thus cannot exit from regional lymph nodes (42, 43). That is why FTY720 is also called a functional antagonist of S1PR. In addition to inhibition of lymphocyte trafficking, FTY720 can also inhibit trafficking of dendritic cells (41). It has been used to investigate the role of dendritic cells (DCs) in intracellular bacterium Francisella tularensis dissemination to draining mediastinal lymph nodes following lung infection (44). FTY720 treatment significantly reduced Francisella-laden DCs and Francisella colonization in draining lymph nodes 48 h after infection. FTY720 has also been shown to reduce HIV systemic dissemination in humanized mice 14 days after intravaginal inoculation, which corelated with inhibition of DC trafficking (45). The FTY720 inhibition of genital Chlamydia spreading to the GI tract was also most obvious during the first 2 weeks (38; this study). However, due to FTY720’s multiple effects on different types of cell trafficking, it is difficult to pinpoint the contribution of a specific cell type. Furthermore, FTY720 has been shown to inhibit T cell activation in an S1PR-independent manner (46). Thus, many more experiments are required for revealing the precise mechanism of any FTY720-inhibitable phenotypes. Finally, most FTY720 inhibition effects were measured within 2 weeks; the long-term effect of FTY720 on pathogen spreading and pathogenicity remained unknown prior to the current study.
In the current study, we extended our observation beyond 2 weeks after infection and found that FTY720 inhibition of genital Chlamydia spreading to the GI tract gradually declined and disappeared as the observation time prolonged. The time-dependent loss of the FTY720 inhibition of chlamydial spreading may not be due to the decline in FTY720 efficacy in inhibiting host immune cell trafficking. This is because FTY720 has been shown to be able to maintain its inhibition on lymphocyte trafficking for more than 100 days (47). FTY720 completely blocked chlamydial organisms from spreading to the GI tract for the 1st 3 weeks in C57 mice but failed to completely block the spreading at any time points in CBA mice. These observations suggest that Chlamydia may use multiple pathways for spreading, including those that are inhibitable by FTY720 and those that are resistant to FTY720 inhibition. CBA mice may possess more of the latter. We propose to define the FTY720-inhibitable spreading as the 1st wave of spreading, while the FTY720-resistant spreading is the 2nd wave. The 1st wave of spreading may be mediated by myeloid/lymphoid cells, the migration of which is susceptible to FTY720 inhibition. The 2nd wave of spreading must be mediated by FTY720-resistant cells/mechanisms. Monitoring chlamydial organism spreading into the GI tract of FTY720-treated mice following an intravaginal inoculation may be a most suitable model for further characterizing the 2nd wave. Besides hiding inside host cells, bacteria have also evolved other strategies for survival in the bloodstream (48). For example, Borrelia burgdorferi and Borrelia garinii may use their outer surface protein C (OspC) to interact with the complement component C4b to achieve bloodstream survival in animals (49). By simply following the blood flow, bacteria can conveniently reach their target organs. The question is whether mucosally inoculated chlamydial organisms can enter and survive the blood circulation in the form of free elementary bodies for reaching the GI tract lumen. It has been shown that following a genital tract inoculation, Chlamydia can survive in the blood circulation for ∼2 weeks, which led to the proposal that genital chlamydial organisms might be able to spread via a host cell-free pathway (20, 36). The current study has laid a foundation for further characterizing the mechanism of FTY720-resistant pathway(s).
Interestingly, we also found that FTY720 treatment slightly increased Chlamydia colonization in the GI tract following an intracolon inoculation throughout the experiment. This observation is consistent with a recent report that treatment with FTY720 delayed the clearance of Citrobacter rodentium infection in mice by blocking the migration of T cells and other immune cells to the inflamed colon (50). The FTY720 impact on chlamydial colonization following intracolon inoculation was not statistically significant. This is probably due to the fact that Chlamydia muridarum behaves as a normal commensal species in the mouse colon (20, 51, 52), and it is able to cooperate with the colonic defense system to achieve long-lasting colonization (53). However, C. rodentium is still a pathogen and is eventually cleared from the mouse GI tract. Thus, FTY720 inhibition of myeloid and lymphoid cell recruitment to the colon is supposed to be more impactful on C. rodentium than C. muridarum.
MATERIALS AND METHODS
Chlamydial organism growth.
Chlamydia muridarum clone CMpmCherry (13) used in the current study was derived from strain Nigg3 (GenBank accession no. CP009760.1). CMpmCherry retains the wild-type Nigg3 genome sequence and is thus designated wild-type Chlamydia. Chlamydial organisms were propagated in HeLa cells (human cervical carcinoma epithelial cells; ATCC CCL-2.1) and purified as elementary bodies (EBs) as reported previously (54). Aliquots of purified EBs were stored at −80°C until use. The storage buffer was SPG (sucrose, phosphate, and glutamate) consisting of 220 mM sucrose, 12.5 mM phosphate, and 4 mM l-glutamic acid, pH 7.5.
Mouse infection.
Mouse experiments were carried out in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Committee on the Ethics of Laboratory Animal Experiments of the University of Texas Health Science Center at San Antonio.
Chlamydia EBs were used to inoculate 6- to 7-week-old female mice (Jackson Laboratories, Inc., Bar Harbor, ME) intravaginally or intracolonically as described previously (20, 52). The mouse strains used in the current study include C57BL/6J (stock no. 0006640) and CBA/J (stock no. 000656). For intravaginal inoculation, stock EBs diluted in 10 μl SPG that contains 2 × 105 inclusion-forming units (IFUs) were delivered to the ectocervix area by use of a 20-μl micropipette tip. Five days prior to inoculation, each mouse was injected subcutaneously with 2.5 mg Depo-Provera (Pharmacia & Upjohn, Kalamazoo, MI) suspended in sterile phosphate-buffered saline (PBS). For intracolon inoculation, EBs were diluted in 50 μl of SPG buffer containing 2 × 105 IFUs and delivered to the colon using a straight ball-tipped needle designed for mouse oral gavage (N-PK 020; Braintree Scientific, Inc., Braintree, MA). Following the inoculation, both vaginal and rectal swabs were taken periodically, or organs/tissues were harvested (after mice were sacrificed) for titrating viable organisms as described previously (20, 36). On day 56 after intravaginal inoculation, all mice were sacrificed for observing genital tract pathology as described below.
In some experiments, mice were treated with FTY720 (catalog no. A10408; AdooQ Bioscience, USA) as follows. FTY720 was applied to mice 4 days prior to chlamydial infection via two consecutive daily gavages, each with 20 μg in 100 μl sterile PBS, and maintained in drinking water throughout the experiment. FTY720 concentrations in drinking water were 1.85 μg/ml for C57BL/6J mice and 2.5 μg/ml for CBA/J mice.
Titrating live chlamydial organisms recovered from swabs and tissue homogenates.
To quantitate live chlamydial organisms in vaginal or rectal swabs, each swab was soaked in 0.5 ml of SPG and vortexed with glass beads, and the chlamydial organisms released into the supernatants were titrated on HeLa cell monolayers in duplicate. The infected cultures were processed for immunofluorescence assay as described previously (55) and below. Inclusions were counted in five random fields per coverslip under a fluorescence microscope. For coverslips with less than one IFU per field, entire coverslips were counted. Coverslips showing obvious cytotoxicity of HeLa cells were excluded. The total number of IFUs per swab was calculated based on the mean IFUs per view and the ratio of the view area to that of the well, dilution factor, and inoculation volumes. Where possible, a mean IFU/swab was derived from the serially diluted and duplicate samples for any given swab. The total number of IFUs/swab was converted into log10, which was used to calculate the mean and standard deviation across mice of the same group at each time point.
For quantitating live organisms from mouse organs and tissue segments, each organ or tissue segment was transferred to a tube containing 0.5 to 5 ml SPG, depending the sizes of the organs. GI tract tissues included small intestine duodenum; jejunum, ileum, and large intestine cecum; colon; and anorectum (rectum), while each genital tract was divided into upper (ovary, oviduct, uterine horn, uterus, and endocervix) and lower (ectocervix and vagina) genital tract tissues. The organs and tissue segments were homogenized in cold SPG using a 2-ml tissue grinder (catalog no. K885300-0002; Fisher Scientific, Pittsburg, PA) or an automatic homogenizer (Omni tissue homogenizer; catalog no. TH115; Omni International, Kennesaw, GA). The homogenates were briefly sonicated and spun at 3,000 rpm for 5 min to pellet the remaining large debris. The supernatants were titrated for live C. muridarum organisms on HeLa cells as described above. The results were expressed as log10 IFUs per organ or tissue segment.
Immunofluorescence assay.
The immunofluorescence assay used for titration of live organisms was carried out as described previously (56). A rabbit antibody (designated R1604, raised with purified C. muridarum EBs) was used as a primary antibody to label all C. muridarum in HeLa cells and was visualized with a goat anti-rabbit IgG conjugated with Cy2 (green; catalog no. 111-225-144, Jackson ImmunoResearch Laboratories, Inc., West Grove PA). The DNA dye Hoechst 3328 (blue; Sigma-Aldrich, St. Louis, MO) was used to visualize nuclei. The double-labeled samples were used for counting for C. muridarum under a fluorescence microscope (AX70, Olympus) equipped with a charge-coupled-device (CCD) camera (Hamamatsu).
Evaluating genital tract pathology macroscopically and microscopically.
On day 56 after intravaginal infection, mice were euthanized for evaluating genital tract pathology. The focus was on the upper genital tract hydrosalpinx. Before the tissues were removed, an in situ gross examination was performed for evidence of oviduct hydrosalpinx or any other related abnormalities of oviducts. The severity of oviduct hydrosalpinx was scored based on the following criteria: no hydrosalpinx, 0; hydrosalpinx detectable only under microscopic examination, 1; hydrosalpinx clearly visible with naked eyes but the size was smaller than the ovary on the same side, 2; equal to the ovary on the same side, 3; or larger than the ovary on the same side, 4. The excised tissues, after photographing, were fixed in 10% neutral formalin, embedded in paraffin, and serially sectioned longitudinally (with 5 μm/each section). Efforts were made to include the cervix, both uterine horns and oviducts, as well as luminal structures of each tissue in each section. The sections were stained with hematoxylin and eosin (H&E) as described elsewhere (55). The H&E-stained sections were assessed by a pathologist blinded to mouse treatment and scored for severity of inflammation and pathologies based on the modified schemes established previously (13). The oviducts were scored for both luminal dilation (0, no significant dilatation; 1, mild dilation of a single cross section; 2, one to three dilated cross sections; 3, more than three dilated cross sections; and 4, confluent pronounced dilation) and inflammatory cell infiltration (0, no significant infiltration; 1, infiltration at a single focus; 2, infiltration at two to four foci; 3, infiltration at more than four foci; and 4, confluent infiltration). Scores assigned to individual mice were calculated into means ± standard errors for each group of animals.
Statistical analyses.
All data, including the time courses of live organism shedding (IFUs) and pathology scores, were compared using the area under the curve (AUC) between two groups using Wilcoxon rank sum test (an in-house Excel sheet), while category data, including number of mice positive for live organism shedding or hydrosalpinx, were analyzed using Fisher's exact test (http://vassarstats.net/tab2x2.html).
ACKNOWLEDGMENT
This study was supported in part by NIH grants (R01AI047997 and R01AI121989 to G.Z.).
REFERENCES
- 1.Budrys NM, Gong S, Rodgers AK, Wang J, Louden C, Shain R, Schenken RS, Zhong G. 2012. Chlamydia trachomatis antigens recognized in women with tubal factor infertility, normal fertility, and acute infection. Obstet Gynecol 119:1009–1016. doi: 10.1097/AOG.0b013e3182519326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rodgers AK, Budrys NM, Gong S, Wang J, Holden A, Schenken RS, Zhong G. 2011. Genome-wide identification of Chlamydia trachomatis antigens associated with tubal factor infertility. Fertil Steril 96:715–721. doi: 10.1016/j.fertnstert.2011.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rodgers AK, Wang J, Zhang Y, Holden A, Berryhill B, Budrys NM, Schenken RS, Zhong G. 2010. Association of tubal factor infertility with elevated antibodies to Chlamydia trachomatis caseinolytic protease P. Am J Obstet Gynecol 203:494.e7–494.e14. doi: 10.1016/j.ajog.2010.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.de la Maza LM, Pal S, Khamesipour A, Peterson EM. 1994. Intravaginal inoculation of mice with the Chlamydia trachomatis mouse pneumonitis biovar results in infertility. Infect Immun 62:2094–2097. doi: 10.1128/IAI.62.5.2094-2097.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Morrison RP, Feilzer K, Tumas DB. 1995. Gene knockout mice establish a primary protective role for major histocompatibility complex class II-restricted responses in Chlamydia trachomatis genital tract infection. Infect Immun 63:4661–4668. doi: 10.1128/IAI.63.12.4661-4668.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shah AA, Schripsema JH, Imtiaz MT, Sigar IM, Kasimos J, Matos PG, Inouye S, Ramsey KH. 2005. Histopathologic changes related to fibrotic oviduct occlusion after genital tract infection of mice with Chlamydia muridarum. Sex Transm Dis 32:49–56. doi: 10.1097/01.olq.0000148299.14513.11. [DOI] [PubMed] [Google Scholar]
- 7.Murthy AK, Li W, Guentzel MN, Zhong G, Arulanandam BP. 2011. Vaccination with the defined chlamydial secreted protein CPAF induces robust protection against female infertility following repeated genital chlamydial challenge. Vaccine 29:2519–2522. doi: 10.1016/j.vaccine.2011.01.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chen J, Zhang H, Zhou Z, Yang Z, Ding Y, Zhou Z, Zhong E, Arulanandam B, Baseman J, Zhong G. 2014. Chlamydial induction of hydrosalpinx in 11 strains of mice reveals multiple host mechanisms for preventing upper genital tract pathology. PLoS One 9:e95076. doi: 10.1371/journal.pone.0095076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lei L, Chen J, Hou S, Ding Y, Yang Z, Zeng H, Baseman J, Zhong G. 2014. Reduced live organism recovery and lack of hydrosalpinx in mice infected with plasmid-free Chlamydia muridarum. Infect Immun 82:983–992. doi: 10.1128/IAI.01543-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.O'Connell CM, Ingalls RR, Andrews CW Jr, Scurlock AM, Darville T. 2007. Plasmid-deficient Chlamydia muridarum fail to induce immune pathology and protect against oviduct disease. J Immunol 179:4027–4034. doi: 10.4049/jimmunol.179.6.4027. [DOI] [PubMed] [Google Scholar]
- 11.Zhong G. 2017. Chlamydial plasmid-dependent pathogenicity. Trends Microbiol 25:141–152. doi: 10.1016/j.tim.2016.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu Y, Huang Y, Yang Z, Sun Y, Gong S, Hou S, Chen C, Li Z, Liu Q, Wu Y, Baseman J, Zhong G. 2014. Plasmid-encoded Pgp3 is a major virulence factor for Chlamydia muridarum to induce hydrosalpinx in mice. Infect Immun 82:5327–5335. doi: 10.1128/IAI.02576-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Conrad TA, Gong S, Yang Z, Matulich P, Keck J, Beltrami N, Chen C, Zhou Z, Dai J, Zhong G. 2016. The chromosome-encoded hypothetical protein TC0668 is an upper genital tract pathogenicity factor of Chlamydia muridarum. Infect Immun 84:467–479. doi: 10.1128/IAI.01171-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chen C, Zhou Z, Conrad T, Yang Z, Dai J, Li Z, Wu Y, Zhong G. 2015. In vitro passage selects for Chlamydia muridarum with enhanced infectivity in cultured cells but attenuated pathogenicity in mouse upper genital tract. Infect Immun 83:1881–1892. doi: 10.1128/IAI.03158-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yang Z, Conrad T, Zhou Z, Chen J, Dutow P, Klos A, Zhong G. 2014. Complement factor C5 but not C3 contributes significantly to hydrosalpinx development in mice infected with Chlamydia muridarum. Infect Immun 82:3154–3163. doi: 10.1128/IAI.01833-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sun X, Tian Q, Wang L, Xue M, Zhong G. 2017. IL-6-mediated signaling pathways limit Chlamydia muridarum infection and exacerbate its pathogenicity in the mouse genital tract. Microbes Infect 19:536–545. doi: 10.1016/j.micinf.2017.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cheng W, Shivshankar P, Li Z, Chen L, Yeh IT, Zhong G. 2008. Caspase-1 contributes to Chlamydia trachomatis-induced upper urogenital tract inflammatory pathologies without affecting the course of infection. Infect Immun 76:515–522. doi: 10.1128/IAI.01064-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Igietseme JU, Omosun Y, Partin J, Goldstein J, He Q, Joseph K, Ellerson D, Ansari U, Eko FO, Bandea C, Zhong G, Black CM. 2013. Prevention of Chlamydia-induced infertility by inhibition of local caspase activity. J Infect Dis 207:1095–1104. doi: 10.1093/infdis/jit009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Campbell J, Huang Y, Liu Y, Schenken R, Arulanandam B, Zhong G. 2014. Bioluminescence imaging of Chlamydia muridarum ascending infection in mice. PLoS One 9:e101634. doi: 10.1371/journal.pone.0101634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang Q, Huang Y, Gong S, Yang Z, Sun X, Schenken R, Zhong G. 2015. In vivo and ex vivo imaging reveals a long-lasting chlamydial infection in the mouse gastrointestinal tract following genital tract inoculation. Infect Immun 83:3568–3577. doi: 10.1128/IAI.00673-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Perry LL, Hughes S. 1999. Chlamydial colonization of multiple mucosae following infection by any mucosal route. Infect Immun 67:3686–3689. doi: 10.1128/IAI.67.7.3686-3689.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yeruva L, Spencer N, Bowlin AK, Wang Y, Rank RG. 2013. Chlamydial infection of the gastrointestinal tract: a reservoir for persistent infection. Pathog Dis 68:88–95. doi: 10.1111/2049-632X.12052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhong G. 2018. Chlamydia spreading from the genital tract to the gastrointestinal tract - a two-hit hypothesis. Trends Microbiol 26:611–623. doi: 10.1016/j.tim.2017.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhang H, Zhou Z, Chen J, Wu G, Yang Z, Zhou Z, Baseman J, Zhang J, Reddick RL, Zhong G. 2014. Lack of long lasting hydrosalpinx in A/J mice correlates with rapid but transient chlamydial ascension and neutrophil recruitment in the oviduct following intravaginal inoculation with Chlamydia muridarum. Infect Immun 82:2688–2696. doi: 10.1128/IAI.00055-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shao L, Melero J, Zhang N, Arulanandam B, Baseman J, Liu Q, Zhong G. 2017. The cryptic plasmid is more important for Chlamydia muridarum to colonize the mouse gastrointestinal tract than to infect the genital tract. PLoS One 12:e0177691. doi: 10.1371/journal.pone.0177691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Shao L, Zhang T, Liu Q, Wang J, Zhong G. 2017. Chlamydia muridarum with mutations in chromosomal genes tc0237 and/or tc0668 is deficient in colonizing the mouse gastrointestinal tract. Infect Immun 85:e00321-17. doi: 10.1128/IAI.00321-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shao L, Zhang T, Melero J, Huang Y, Liu Y, Liu Q, He C, Nelson DE, Zhong G. 2017. The genital tract virulence factor pGP3 Is essential for Chlamydia muridarum colonization in the gastrointestinal tract. Infect Immun 86:e00429-17. doi: 10.1128/IAI.00429-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tian Q, Zhou Z, Wang L, Abu-Khdeir AH, Huo Z, Sun X, Zhang N, Schenken R, Wang Y, Xue M, Zhong G. 2020. Gastrointestinal coinfection promotes chlamydial pathogenicity in the genital tract. Infect Immun 88:e00905-19. doi: 10.1128/IAI.00905-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Craig AP, Kong FY, Yeruva L, Hocking JS, Rank RG, Wilson DP, Donovan B. 2015. Is it time to switch to doxycycline from azithromycin for treating genital chlamydial infections in women? Modelling the impact of autoinoculation from the gastrointestinal tract to the genital tract. BMC Infect Dis 15:200. doi: 10.1186/s12879-015-0939-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Peters RP, Dubbink JH, van der Eem L, Verweij SP, Bos ML, Ouburg S, Lewis DA, Struthers H, McIntyre JA, Morre SA. 2014. Cross-sectional study of genital, rectal, and pharyngeal chlamydia and gonorrhea in women in rural South Africa. Sex Transm Dis 41:564–569. doi: 10.1097/OLQ.0000000000000175. [DOI] [PubMed] [Google Scholar]
- 31.Gratrix J, Singh AE, Bergman J, Egan C, Plitt SS, McGinnis J, Bell CA, Drews SJ, Read R. 2015. Evidence for increased chlamydia case finding after the introduction of rectal screening among women attending 2 Canadian sexually transmitted infection clinics. Clin Infect Dis 60:398–404. doi: 10.1093/cid/ciu831. [DOI] [PubMed] [Google Scholar]
- 32.Musil K, Currie M, Sherley M, Martin S. 2016. Rectal chlamydia infection in women at high risk of chlamydia attending Canberra Sexual Health Centre. Int J STD AIDS 27:526–530. doi: 10.1177/0956462415586317. [DOI] [PubMed] [Google Scholar]
- 33.Gratrix J, Singh AE, Bergman J, Egan C, McGinnis J, Drews SJ, Read R. 2014. Prevalence and characteristics of rectal chlamydia and gonorrhea cases among men who have sex with men after the introduction of nucleic acid amplification test screening at 2 Canadian sexually transmitted infection clinics. Sex Transm Dis 41:589–591. doi: 10.1097/OLQ.0000000000000176. [DOI] [PubMed] [Google Scholar]
- 34.Dan M, Tyrrell LD, Goldsand G. 1987. Isolation of Chlamydia trachomatis from the liver of a patient with prolonged fever. Gut 28:1514–1516. doi: 10.1136/gut.28.11.1514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Carter JD, Hudson AP. 2017. Recent advances and future directions in understanding and treating Chlamydia-induced reactive arthritis. Expert Rev Clin Immunol 13:197–206. doi: 10.1080/1744666X.2017.1233816. [DOI] [PubMed] [Google Scholar]
- 36.Dai J, Zhang T, Wang L, Shao L, Zhu C, Zhang Y, Failor C, Schenken R, Baseman J, He C, Zhong G. 2016. Intravenous inoculation with Chlamydia muridarum leads to a long-lasting infection restricted to the gastrointestinal tract. Infect Immun 84:2382–2388. doi: 10.1128/IAI.00432-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Volpi C, Orabona C, Macchiarulo A, Bianchi R, Puccetti P, Grohmann U. 2019. Preclinical discovery and development of fingolimod for the treatment of multiple sclerosis. Expert Opin Drug Discov 14:1199–1212. doi: 10.1080/17460441.2019.1646244. [DOI] [PubMed] [Google Scholar]
- 38.Howe SE, Shillova N, Konjufca V. 2019. Dissemination of Chlamydia from the reproductive tract to the gastro-intestinal tract occurs in stages and relies on Chlamydia transport by host cells. PLoS Pathog 15:e1008207. doi: 10.1371/journal.ppat.1008207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Huwiler A, Zangemeister-Wittke U. 2018. The sphingosine 1-phosphate receptor modulator fingolimod as a therapeutic agent: recent findings and new perspectives. Pharmacol Ther 185:34–49. doi: 10.1016/j.pharmthera.2017.11.001. [DOI] [PubMed] [Google Scholar]
- 40.Stepanovska B, Huwiler A. 2020. Targeting the S1P receptor signaling pathways as a promising approach for treatment of autoimmune and inflammatory diseases. Pharmacol Res 154:104170. doi: 10.1016/j.phrs.2019.02.009. [DOI] [PubMed] [Google Scholar]
- 41.Lan YY, De Creus A, Colvin BL, Abe M, Brinkmann V, Coates PT, Thomson AW. 2005. The sphingosine-1-phosphate receptor agonist FTY720 modulates dendritic cell trafficking in vivo. Am J Transplant 5:2649–2659. doi: 10.1111/j.1600-6143.2005.01085.x. [DOI] [PubMed] [Google Scholar]
- 42.Chiba K. 2005. FTY720, a new class of immunomodulator, inhibits lymphocyte egress from secondary lymphoid tissues and thymus by agonistic activity at sphingosine 1-phosphate receptors. Pharmacol Ther 108:308–319. doi: 10.1016/j.pharmthera.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 43.Matloubian M, Lo CG, Cinamon G, Lesneski MJ, Xu Y, Brinkmann V, Allende ML, Proia RL, Cyster JG. 2004. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 427:355–360. doi: 10.1038/nature02284. [DOI] [PubMed] [Google Scholar]
- 44.Bar-Haim E, Gat O, Markel G, Cohen H, Shafferman A, Velan B. 2008. Interrelationship between dendritic cell trafficking and Francisella tularensis dissemination following airway infection. PLoS Pathog 4:e1000211. doi: 10.1371/journal.ppat.1000211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Deruaz M, Murooka TT, Ji S, Gavin MA, Vrbanac VD, Lieberman J, Tager AM, Mempel TR, Luster AD. 2017. Chemoattractant-mediated leukocyte trafficking enables HIV dissemination from the genital mucosa. JCI Insight 2:e88533. doi: 10.1172/jci.insight.88533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Baer A, Colon-Moran W, Bhattarai N. 2018. Characterization of the effects of immunomodulatory drug fingolimod (FTY720) on human T cell receptor signaling pathways. Sci Rep 8:10910. doi: 10.1038/s41598-018-29355-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liu L, Wang C, He X, Shang W, Bi Y, Wang D. 2007. Long-term effect of FTY720 on lymphocyte count and islet allograft survival in mice. Microsurgery 27:300–304. doi: 10.1002/micr.20360. [DOI] [PubMed] [Google Scholar]
- 48.Minasyan H. 2019. Sepsis: mechanisms of bacterial injury to the patient. Scand J Trauma Resusc Emerg Med 27:19. doi: 10.1186/s13049-019-0596-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Caine JA, Lin YP, Kessler JR, Sato H, Leong JM, Coburn J. 2017. Borrelia burgdorferi outer surface protein C (OspC) binds complement component C4b and confers bloodstream survival. Cell Microbiol 19. doi: 10.1111/cmi.12786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Murphy CT, Hall LJ, Hurley G, Quinlan A, MacSharry J, Shanahan F, Nally K, Melgar S. 2012. The sphingosine-1-phosphate analogue FTY720 impairs mucosal immunity and clearance of the enteric pathogen Citrobacter rodentium. Infect Immun 80:2712–2723. doi: 10.1128/IAI.06319-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lin H, He C, Koprivsek JJ, Chen J, Zhou Z, Arulanandam B, Xu Z, Tang L, Zhong G. 2019. Antigen-specific CD4(+) T cell-derived gamma interferon is both necessary and sufficient for clearing chlamydia from the small intestine but not the large intestine. Infect Immun 87:e00055-19. doi: 10.1128/IAI.00055-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Koprivsek JJ, Zhang T, Tian Q, He Y, Xu H, Xu Z, Zhong G. 2019. Distinct roles of chromosome- versus plasmid-encoded genital tract virulence factors in promoting Chlamydia muridarum colonization in the gastrointestinal tract. Infect Immun 87:e00265-19. doi: 10.1128/IAI.00265-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Koprivsek JJ, He Y, Song C, Zhang N, Tumanov A, Zhong G. 2019. Evasion of innate lymphoid cell-regulated gamma interferon responses by Chlamydia muridarum to achieve long-lasting colonization in mouse colon. Infect Immun 88:e00798-19. doi: 10.1128/IAI.00798-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Mukhopadhyay S, Clark AP, Sullivan ED, Miller RD, Summersgill JT. 2004. Detailed protocol for purification of Chlamydia pneumoniae elementary bodies. J Clin Microbiol 42:3288–3290. doi: 10.1128/JCM.42.7.3288-3290.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tang L, Zhang H, Lei L, Gong S, Zhou Z, Baseman J, Zhong G. 2013. Oviduct infection and hydrosalpinx in DBA1/j mice is induced by intracervical but not intravaginal inoculation with Chlamydia muridarum. PLoS One 8:e71649. doi: 10.1371/journal.pone.0071649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fan T, Lu H, Hu H, Shi L, McClarty GA, Nance DM, Greenberg AH, Zhong G. 1998. Inhibition of apoptosis in chlamydia-infected cells: blockade of mitochondrial cytochrome c release and caspase activation. J Exp Med 187:487–496. doi: 10.1084/jem.187.4.487. [DOI] [PMC free article] [PubMed] [Google Scholar]