

Abstract
Diffusion tensor distribution (DTD) imaging builds on principles from diffusion, solid‐state and low‐field NMR spectroscopies, to quantify the contents of heterogeneous voxels as nonparametric distributions, with tensor “size”, “shape” and orientation having direct relations to corresponding microstructural properties of biological tissues. The approach requires the acquisition of multiple images as a function of the magnitude, shape and direction of the diffusion‐encoding gradients, leading to long acquisition times unless fast image read‐out techniques like EPI are employed. While in previous in vivo human brain studies performed at 3 T this proved a viable option, porting these measurements to very high magnetic fields and/or to heterogeneous organs induces B 0‐ and B 1‐inhomogeneity artifacts that challenge the limits of EPI. To overcome such challenges, we demonstrate here that high spatial resolution DTD of mouse brain can be carried out at 15.2 T with a surface‐cryoprobe, by relying on SPatiotemporal ENcoding (SPEN) imaging sequences. These new acquisition and data‐processing protocols are demonstrated with measurements on in vivo mouse brain, and validated with synthetic phantoms designed to mimic the diffusion properties of white matter, gray matter and cerebrospinal fluid. While still in need of full extensions to 3D mappings and of scanning additional animals to extract more general physiological conclusions, this work represents another step towards the model‐free, noninvasive in vivo characterization of tissue microstructure and heterogeneity in animal models, at ≈0.1 mm resolutions.
Keywords: acquisition, diffusion, diffusion MR sequences, high‐order diffusion MR
Diffusion tensor distribution (DTD) imaging allows to quantify the heterogeneity of tissues in terms of local “sizes”, “shapes” and orientations, which are in turn directly related to the microstructural properties of biological tissues. Here, a high‐definition DTD analysis of a mouse brain is presented, carried out at 15.2 T with a surface‐cryoprobe and relying on a SPatiotemporal ENcoding (SPEN) imaging sequence. These results enabled the model‐free, noninvasive characterization of tissue microstructure and heterogeneity in live animals, at ≈0.1 mm resolutions.
Abbreviations used
- ADC
apparent diffusion coefficient
- Aq
sylvius aqueduct
- CB
cerebellum
- cc
corpus callosum
- CSF
cerebrospinal fluid
- D3V
dorsal third ventricle
- DTD
diffusion tensor distribution
- DTI
diffusion tensor imaging
- FA
fractional anisotropy
- FE
fractional eccentricity
- fi
fimbria
- Gl
granular insular cortex
- GM
gray matter
- GrO
granular layer
- LV
lateral ventricles
- MA
microscopic anisotropy index
- MD
mean diffusivity
- SE‐EPI
spin echo‐echo planar imaging
- SPEN
SPatiotemporal ENcoding
- WM
white matter
- μFA
microscopic fractional anisotropy
1. INTRODUCTION
The translational motion of water within brain tissues is restricted by biomembranes forming the outer borders of individual cells as well as of intracellular organelles such as the mitochondria, the endoplasmic reticulum and the cell nucleus. Membranes also constitute the multilayer stacks defining myelin sheaths. The micrometer‐scale organization of these biomembranes imprints itself on the diffusion properties of the water in the tissues, 1 , 2 which can be noninvasively assessed with magnetic resonance imaging (MRI) 3 via diffusion‐weighting gradients. Observables from diffusion‐weighted MRI are conventionally expressed in terms of quantitative metrics such as the apparent diffusion coefficient (ADC), 4 , 5 , 6 mean diffusivity (MD) and fractional anisotropy (FA). 7 This metric calls for the use of diffusion tensor imaging (DTI) 8 modalities, where the water diffusivity properties are approximated as a rank‐two tensor for each imaged voxel. 9 DTI has been widely used to study white matter (WM) diseases, 10 and to track WM pathways in both animal models and humans. 11 , 12 , 13 The orders of magnitude that separate the length scales of water diffusivity (μm) from the imaging resolution (mm) probed by MRI provide these approaches with an intrinsic sensitivity advantage when trying to examine microstructures noninvasively. However, they also introduce ambiguities in the interpretation of conventional DTI data, particularly for heterogeneous voxels containing multiple tissue types 2 , 14 or for WM tracts with complex fiber configurations. 15 Increasing the spatial resolution partially mitigates these heterogeneity effects 16 but remains challenging, especially in vivo, where signal‐to‐noise ratio (SNR) considerations and restrictions on measurement time put practical limits on the achievable imaging resolution. 17 , 18 , 19 , 20
Diffusion tensor distribution (DTD) addresses this challenge by describing the contents of a voxel comprising multiple tissue environments, each possessing a distinct diffusion tensor D, in terms of a P(D) probability function. 21 This P(D) is related to the diffusion‐encoded signal S(b) imaged by DTI as the integral transform
| (1) |
where S 0 is the initial signal amplitude, b is the diffusion‐encoding second‐rank tensor generated by the diffusion‐measuring gradients, 22 and the colon represents the generalized scalar product b : D = ∑i∑jbijDij. Following the treatments in Jian et al 21 and Anderson 23 we constrain the integral in Equation 1 to the space of axisymmetric and semipositive‐definite diffusion tensors; these in turn are fully characterized by four independent parameters: the isotropic diffusivity D iso, the normalized anisotropy D Δ, 24 and the polar and azimuthal angles θ and φ, giving the orientation of the tensor in the lab frame of reference. The two former parameters are related to the axial and radial diffusivities, D || and D ⊥, via D iso = (D || + 2D ⊥)/3 and D Δ = (D ||– D ⊥)/3D iso. When visualizing the tensors as ellipsoid or superquadric glyphs, the values of D iso and D Δ are directly related to the “sizes” and “shapes” of the displayed objects, 25 which are properties inherited from the corresponding diffusion propagators and underlying micrometer‐scale tissue structure. 26 Access to the DTD thus allows the discrimination and structural characterization of various sub‐voxel tissue environments, based on their diffusion properties. Moreover, multiple statistical descriptors carrying different information can be derived from the DTD. The mean “size” E[D iso] is identical to the classical MD. The mean squared “shape” E[D Δ 2] provides similar information as previously introduced anisotropy measures such as the microscopic anisotropy index (MA), 27 the fractional eccentricity (FE) 28 and the microscopic fractional anisotropy (μFA). 29 , 30 All these pure “shape” measures are conceptually different from the traditional FA, 7 which convolves the fundamentally different properties of “shape” and orientational order into a single scalar metric. 29 , 31 The DTD analysis also provides the (co)variances Var[D iso], Var[D Δ 2] and Cov[D iso,D Δ 2], which report on various aspects of intra‐voxel heterogeneity. 32 In particular, the Var[D iso] parameter shows elevated values in voxels containing distinct intra‐ and extracellular water populations, 29 and has been shown to correlate with cell density heterogeneity in brain tumors. 33
Recently, acquisition and analysis protocols allowing unconstrained inversion of Equation 1 and subsequent retrieval of the DTD have been introduced. 32 , 34 , 35 Key to these approaches is the combined use of multidimensional diffusion MRI methods, 36 in which the signal is encoded according to the principles of solid‐state NMR experiments for correlating isotropic and anisotropic chemical shifts, 37 , 38 together with data inversion strategies from low‐field NMR incorporating uncertainty estimation of the metrics derived from the distributions arising from Equation 1. 39 In practice, the measurements involve acquiring data with b ‐tensors of varying magnitude b, normalized anisotropy b ∆ 24 defined by the axial and radial eigenvalues, b || and b ⊥, according to b ∆ = (b ||‐ b ⊥)/b, and orientations (Θ, Φ), in order to establish correlations across the (D iso, D Δ, θ, φ) dimensions of the DTD space. Each tensor is defined by a conventional b‐value and normalized anisotropy b ∆ with b = b || + 2b ⊥ and b ∆ = (b ||‐ b ⊥) /b. The “shape” of the diffusion‐encoding tensor is characterized by b ∆ with special values −1/2, 0 and +1 for planar, spherical and linear diffusion encoding, respectively. 24 While acquiring data with multiple tensorial b‐values provides the increased specificity needed to characterize the heterogeneities associated with diffusivity properties, it also results in long acquisition times; dealing with the latter leads in turn to reduced spatial resolution and/or limited sensitivity. Hence, in order to be compatible with in vivo acquisition times, DTD imaging is usually performed using fast acquisition schemes such as spin echo‐echo planar imaging (SE‐EPI). 16 In murine models, where the sample volume is small, sensitivity usually remains the limiting factor for endowing these diffusion MRI measurements with high spatial resolution. Cryo‐coils 40 and very high magnetic fields 41 can be used to alleviate this limitation; however, increasing the magnetic field B 0 also enhances the susceptibility‐induced magnetic field inhomogeneities, 41 leading to image distortions in SE‐EPI. High magnetic fields are also associated with shortened transverse relaxation times T 2, demanding, in turn, the application of stronger magnetic field gradients, which increase eddy current artifacts. While SE‐EPI can partially overcome some of these drawbacks by relying on segmented acquisitions that yield improved resolution and immunity to magnetic field inhomogeneities, 42 these approaches are notably sensitive to motional and instrumental artifacts.
SPatiotemporal ENcoding (SPEN) provides a single‐shot acquisition module with potentially higher resilience to magnetic field inhomogeneities and to eddy current artifacts than SE‐EPI. 43 , 44 , 45 Indeed, SPEN can be implemented in a fully T 2*‐refocused manner, where field inhomogeneities are compensated throughout the acquisition instead of at a single echoing time. 44 In addition, SPEN's bandwidth along the blipped dimension—the more artifact‐prone in EPI—is defined at the excitation stage by a chirp pulse and can be set at arbitrary values; this can further decrease sensitivity to magnetic field inhomogeneity, even if at the cost of SNR. Moreover, SPEN records its images directly in real space along the low‐bandwidth dimension; since each signal yields a direct low‐resolution image, this facilitates motion correction and data interleaving between shots, leading to final results that can be freed from motion artifacts in multi‐shot or signal‐averaged acquisitions. 46 , 47 , 48 SPEN's use of a chirped pulse applied in conjunction with an encoding gradient also induces a spatial selectivity, which permits zooming without folding along the low‐bandwidth (blipped) dimension. 49 Finally, as the offset between even and odd echoes that leads to ghost artifacts in EPI can be corrected in SPEN a posteriori via a referenceless method, 46 , 47 , 50 this spares the need for the longer acquisition schemes used in EPI to correct this artifact with double sampling of the readout lines. 51
In this work, we show that DTD imaging can be performed on in vivo mouse brains at ≈100 μm in‐plane spatial resolutions (in this case, with a 700 μm slice thickness) by combining the SNR advantages provided by high magnetic fields (15.2 T) and surface‐cryoprobes, with SPEN methods capable of mitigating the effects of B 0‐ and B 1‐inhomogeneities. The microstructural results provided by the ensuing imaging approach are analyzed, and further understood by ancillary measurements on three synthetic phantoms emulating the diffusion properties of WM, gray matter (GM) and cerebrospinal fluid (CSF).
2. MATERIALS AND METHODS
2.1. Animals
Measurements were performed on healthy adult female C57BL/6 mice aged ~6 months (n = 2, Envigo, Jerusalem). The Institutional Animal Care and Use Committee of the Weizmann Institute of Science, which is fully accredited by the AAALAC, the US NIH Office of Laboratory Animal Welfare, and the Israel Ministry of Health, approved all experiments. The animals were housed in cages in a 12‐hour night/12‐hour daylight cycle, with water and food available ad libitum. Mice were anesthetized with ~3% isoflurane and kept under anesthesia throughout the entire scanning session using ~1.5% isoflurane mixed with 20%/80% oxygen/nitrogen. The animals were kept warm during the experiment by using a water‐heated blanket (positioned under the animals) and setting the external temperature of the cryo‐coil to 37°C. Only respiration was monitored throughout the course of the experiment via a pressure sensor (SA‐II, Stony Brook, NY, USA) and maintained at 30‐60 breaths per minute.
2.2. Phantoms
Validation of the acquisition and analysis pipeline was performed by measurements on three phantoms consisting of 15 mm Eppendorf tubes containing either a lyotropic liquid crystal, n‐dodecane (Sigma‐Aldrich, Israel), or tap water. The phantoms were designed to give diffusion properties similar to the main components of healthy living brains: the liquid crystal phantom captures anisotropic diffusion within WM, the dodecane phantom mimics slow isotropic diffusion in the GM, and the water phantom approximates the fast isotropic diffusion of CSF. The liquid crystal is given the nickname “hex” for its reverse hexagonal phase structure, and was prepared according to the protocol described in detail by Nilsson et al 52 using 41.94 wt% H2O of Milli‐Q quality, 44.12 wt% of the detergent sodium 1,4‐bis(2‐ethylhexoxy)‐1,4‐dioxobutane‐2‐sulfonate (Sigma‐Aldrich, Sweden), and 13.94 wt% of the hydrocarbon 2,2,4‐trimethylpentane (Sigma‐Aldrich, Sweden).
2.3. Image acquisition
All experiments were performed on a horizontal Biospec 15.2 T USR preclinical MRI scanner with an Avance IIIHD console and a B‐GA 6S‐100 three‐axis gradient system with a 60 mm inner diameter, capable of delivering a gradient strength of 1000 mT/m and associated with an integrated first to third order shim set. Data were acquired with a surface 1H quadrature transmit/receive cryoprobe coil with an inner diameter of 20 mm. In vivo measurements were performed without respiratory triggering, but using ear and teeth bars to fix the mouse's head. Prior to all acquisitions, the B 0 homogeneity within the field of view (FoV) was optimized using the field map method 53 , 54 , 55 and the third‐order shims available. For the in vivo experiments, the linewidth at 50% obtained within the targeted FoV by a PRESS sequence was ≈ 60 Hz. Two saturation bands were used to remove the intense signal at the back of the brain and allow better phase adjustment during the SPEN processing. Fat suppression was not used. Additional hardware and imaging parameters used in the in vivo and phantom data acquisitions are reported in Table 1. SPEN data were collected with the sequence shown in Figure 1, which contains a refocusing chirp pulse whose length is half of the acquisition time T a in order to maintain the condition of full T 2*‐refocusing. 44 The rasterized image‐space acquisition inherent to the spatiotemporal encoding induces a gradually increasing echo time from 29 to 52 ms, which in turn leads to a nonuniform T 2‐weighting along the SPEN dimension. The initial encoding and its subsequent step‐by‐step refocusing by the acquisition gradients also leads to a variation in the effective b‐values along the spatiotemporal encoding axis. 56 The spatially incremented echo times and progressive diffusion weighting along the SPEN dimension may lead to a gradual skewing of the estimated DTDs towards components with long T 2 and slow diffusion. On the other hand, the fact that the diffusion gradient waveforms are fully refocused before the application of the SPEN gradients means that the diffusion weighting from the dedicated diffusion encoding and the SPEN imaging gradients can be separated into two independent b‐tensor factors. The latter span relatively small values in the 15‐250 s/mm2 range, which, since constant throughout the DTD waveform variations, could be factored out in the final analysis.
TABLE 1.
Main parameters of the DTD SPEN acquisitions
| Total acquisition time | 2 h 25 min |
|---|---|
| Echo time | 59 +/− 11.5 ms |
| Repetition time | 1 s |
| Interleaved segments | 3 |
| Number of scans | 8 |
| Field of view | 13 × 17 × 0.7 mm3 |
| Matrix size | 180 × 174 × 1 |
| Readout regridded points (effective) | 132 |
| Spatial resolution | 99 × 98 × 700 μm3 |
| Readout bandwidth | 455 kHz |
| Second dimension bandwidth | 10.450 kHz |
| Chirp duration | 11.5 ms |
| Number of images | 364 |
| Number of b 0 images | 4 (one per b shape) |
| Diffusion waveform duration | 18 ms |
| b‐values | 0.042, 0.173, 0.388, 1.080, 2.116 and 3.499 x 109 s/m2 |
| b Δ | −0.5, 0, 0.5 and 1 |
| Number of b‐tensor directions | 15 |
| Linewidth at 50% | 60 Hz |
| Press shim FOV | 5 × 8 × 1 mm3 |
| SNR of the b 0 image | 26 |
FIGURE 1.
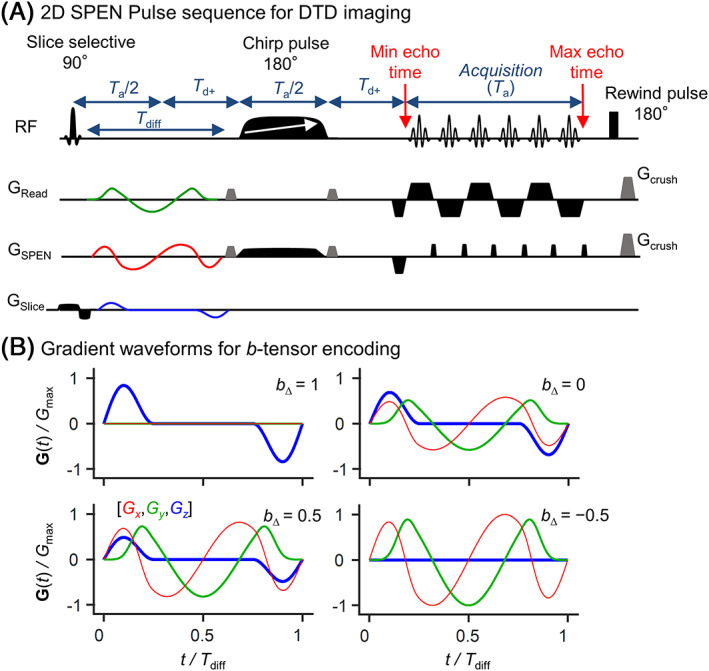
Experimental protocol assayed here for diffusion tensor distribution (DTD) imaging at high spatial resolution. (A) Pulse sequence combining tensor‐valued diffusion encoding with a 2D SPatiotemporal ENcoding (SPEN) acquisition scheme. Diffusion encoding is performed by a set of continuous, self‐refocused gradient waveforms, with duration T diff, allowing for separate control over the magnitude b, normalized anisotropy b ∆ and orientation (Θ, Φ) of the diffusion‐encoding tensor b. 34 The pulse sequence timing is illustrated under conditions that fulfill SPEN's full refocusing condition; to this effect, an additional delay (T d+) was introduced, if/when the length of the diffusion‐encoding gradient waveform exceeded T a/2. (B) Set of gradient waveforms used in this work. Each waveform was designed to give a distinct value of b ∆, as per manipulations of the three orthogonal (color‐coded) gradients
The SPEN readout was performed during the full echo train acquisition, including the gradient ramps. This induced a nonuniform sampling of the k‐space points along the readout direction, which was corrected by regridding 57 prior to Fourier transform. This operation was taken into account when determining the final spatial resolution, which was calculated based on the evenly spaced k‐space points obtained after regridding. Images along the SPEN dimension were reconstructed as described elsewhere 46 , 47 , 48 ; the main elements of this reconstruction are: a phase correction between even and odd segments, a phase correction in between segments which also induce a rigid motion correction, and a super‐resolution operation. The SPEN sequence and SPEN reconstruction pipeline used in this study are available at https://www.weizmann.ac.il/chemphys/Frydman_group/software.
Diffusion‐encoded images were collected for a full mouse brain coronal slice, with an in‐plane spatial resolution of 100 μm and a slice thickness of 700 μm. A coronal orientation was chosen so as to cover the largest spatial dimensions of the mouse brain entirely. The choice of this challenging orientation, less common than the axial orientation usually used in EPI‐based DWI studies, is possible due to the increased robustness of the SPEN acquisition to B 0 inhomogeneities, and of its ability to suitably scan its spatial axis while avoiding signal folding. The repetition time was 1 second, and three segments were interleaved in each image acquisition. The diffusion‐encoding protocol comprised 364 such image acquisitions, decomposed into four b Δ‐values (−0.5, 0, 0.5 and 1), 24 15 directions, and six b‐values ranging from 0.042 to 3.5 x 109 s/m2. Gradients were calculated using analytical waveforms 58 derived on the basis of the variable‐angle spinning technique 38 in solid‐state NMR. The maximum b‐value 3.5 x 109 s/m2 was reached at 90% of the maximum available gradient strength (1000 mT/m) using a waveform of duration Τ diff = 18 ms. Together with a chirp duration T a/2 = 11.5 ms, this constrained the SPEN echo times to the 47.5 to 70.5 ms range, which was difficult to reduce by increasing segmentation. To mitigate the low SNR arising as a result of long echo times and relatively short T 2s (~ 25 ms for GM at 15.2 T 59 ), a set of eight averages per scan was acquired, leading to a total experiment time of 2 hours and 25 minutes. This scanning time, long but still compatible with in vivo preclinical investigations, requires the use of fast acquisition schemes such as EPI or SPEN, both of which provide similar acceleration factors. While these choices allowed us to acquire 2D images with high spatial resolution (100 μm in‐plane) using a limited number of interleaved segments (three), the SNR demands of DTD imaging also required us to average eight scans. Coupling these factors to the large number of images required for DTD data inversion motivates our acquisition of just a single slice.
2.4. DTD estimation
All the acquired images were denoised using random matrix theory, 60 with an optimized 3 × 3 bidimensional kernel. The SNRs of the various images were computed prior to any denoising; SNR values were estimated as the mean value of the GM signal divided by the standard deviation of the noise in an area without signal.
For each voxel, DTDs were estimated from the acquired signal via a Monte Carlo inversion 39 , 61 of Equation 1, using an algorithm that has been described in detail previously 32 , 35 and that is freely available online (https://github.com/daniel-topgaard/md-dmri). The algorithm approximates the DTD as a discrete set of N components {wn, D || ,n, D ⊥,n, θ n, φ n}1 ≤ n ≤ N, where wn is the weight of component n. Uncertainty estimations were performed using bootstrapping with replacement, to generate 96 DTDs consistent with the input signal data. For the purpose of calculating the parameter maps, the DTDs were converted to statistical descriptors quantifying the total weight/initial signal S 0, as well as means E[x], variances Var[x] and covariances Cov[x,y] of the various dimensions of the distribution space. 32 These statistical descriptors were derived for both the entirety and from smaller divisions (“bins”) of the DTD space, and are reported using a (D iso, D Δ 2, θ, φ) parametrization rather than the original (D ||, D ⊥, θ, φ) representation: E[D iso], E[D Δ 2], Var[D iso], Var[D Δ 2] and Cov[D iso,D Δ 2]. Here, we defined three bins meant to capture the contributions of WM, GM and CSF: WM, 0.005 < D iso / 10–9 m2s–1 < 2 and 0.25 < D Δ 2 < 1; GM, 0.005 < D iso/10–9 m2s–1 < 2 and 0 < D Δ 2 < 0.25; CSF, 2 < D iso/10–9 m2s–1 < 5 and 0 < D Δ 2 < 1. The bootstrap ensembles of scalar descriptors were condensed to a set of averages by computing the medians 〈·〉 of the 96 plausible values of S 0, E[x], Var[x], and Cov[x,y]. Parameter maps of 〈S 0〉, 〈E[D iso]〉, 〈E[D Δ 2]〉, 〈Var[D iso]〉, 〈Var[D Δ 2]〉 and 〈Cov[D iso,D Δ 2]〉 were then used to visualize the main features of the estimated DTDs.
3. RESULTS AND DISCUSSION
Even when relying on segmented acquisitions combined with animal immobilization using ear and teeth bars, careful optimization of the B 0 homogeneity methods 53 , 54 , 55 and brand new hardware, retrieving full‐brain mouse coronal SE‐EPI images at 15.2 T remains challenging. Figure 2 panel A and B compares one such representative image against a SPEN coronal in vivo counterpart, both recorded with high SNR (48 averages), in three segments at a 100 μm in‐plane resolution, under equivalent shimming conditions and similar parameters, except for the echo time and the phase dimension bandwidth. Although the two images show similar anatomical details, contrast differences between CSF and GM are noticeable between the images; these arise mainly due to the longer echo times of SPEN (ranging spatially from 29 to 52 ms) compared with the SE‐EPI acquisition (30 ms). A decay in overall signal intensities along the (antero‐posterior) SPEN dimension is also noticeable; this originates from the progressively stronger T 2 and diffusion weightings induced by the SPEN readout over the course of the image acquisition. 56 Still, as these weights are constant throughout the various b‐encoded acquisitions, their effects factor out from the processed DTD images. On the other hand, the SE‐EPI image evidences a number of ghost artifacts (Figure 2, orange arrow) and of susceptibility‐induced B 0 inhomogeneity artifacts (Figure 2, green arrows). These ghost artifacts could not be eliminated with classical one‐dimensional phase correction via polynomial fitting to reference data scans, 57 and although they could have been eliminated with a double‐sampling approach, 51 this would double the EPI acquisition time, which would in turn increase the B 0 inhomogeneity artifacts. It follows that under these good shimming conditions and for the same number of segments, SPEN allows one to obtain these data without ghosts and with much reduced susceptibility‐induced B 0 artifacts, even if subtle stripe‐like artifacts were also evident in the high SNR SPEN image. The absence of significant ghost artifacts in the SPEN image is made possible by its referenceless reconstruction algorithm, including a bidimensional phase correction which minimizes the phase difference between even and odd readout segments, and between shots. 46 , 62 This optimization is easier in SPEN due to the direct image space acquisition, which allows the phase optimization to be performed directly in the image space where the phase is smoother than in k‐space. The reduction of the B 0 inhomogeneity artifacts is aided by SPEN's increased bandwidth in the second dimension (10.45 vs. 7.57 kHz for SE‐EPI) and by its fully T 2*‐refocused acquisition. The extent of the artifacts reduction is more clearly evidenced in Figure 2C, which compares (in red and blue) normalized 1D profiles extracted from the high SNR SE‐EPI and SPEN images, respectively. Notice in these the reduction in the intensities of the B 0 inhomogeneity artefacts like those evidenced by the green arrows, and the elimination of ghost artefacts of the kind highlighted by the orange arrow. These and other similar artifact attenuations explain our use of SPEN in this kind of study. Figure 2D exhibits one of the SPEN b 0 images within the DTD dataset acquired with the same parameters as the higher SNR SPEN image in panel (B), except for a number of averages reduced to eight and an echo time increased to 59 ± 11.5 ms due to the inclusion of the diffusion delay (T diff) of 18 ms. This image was denoised using random matrix theory 60 and a 3 × 3 bidimensional kernel. Notice the difference in contrast between CSF and GM in the two SPEN images arising due to the changing echo times.
FIGURE 2.
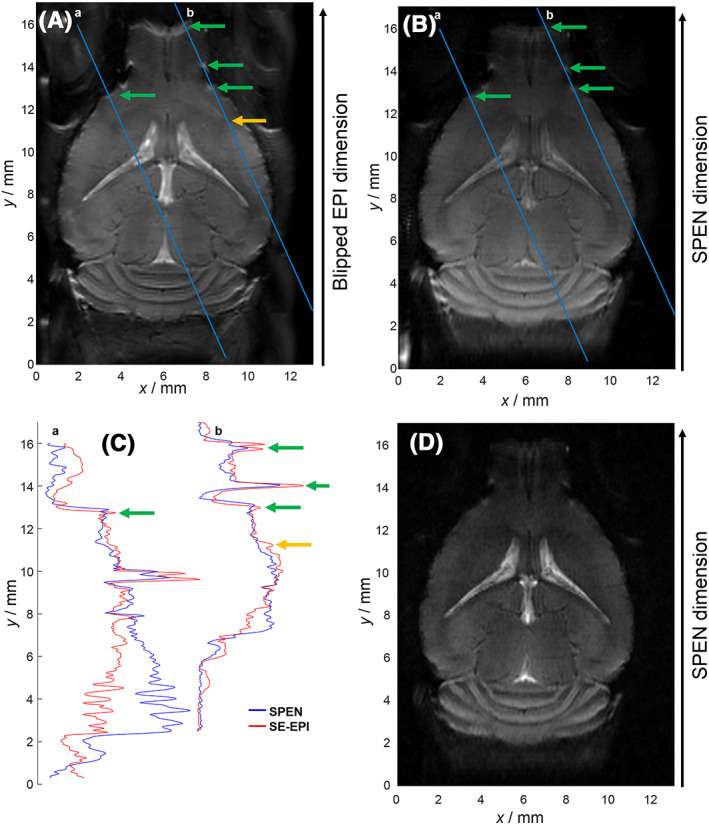
Comparison between images resulting from SE‐EPI and SPEN experiments on an in vivo mouse brain coronal slice at 15.2 T using identical shim conditions. All images were acquired with a three‐segment acquisition and had in‐plane resolutions of 100 × 100 µm² and a slice thickness of 700 µm. A, High‐SNR SE‐EPI image recorded with 48 averages leading to an acquisition time of 2 min and 24 seconds. The SE‐EPI sequence had an echo time of 30 ms and an effective bandwidth in the blipped dimension of 7.57 kHz. This image exhibits ghost artifacts like the one shown by the orange arrow and B 0 inhomogeneity artifacts like those shown by the green arrows. B, High‐SNR fully T 2*‐refocused SPEN image recorded with 48 averages leading to an acquisition time of 2 min and 24 seconds. The fully T 2*‐refocused SPEN image was acquired with echo times of 40.5 ± 11.5 ms and an effective SPEN‐dimension bandwidth of 10.45 kHz. The increased bandwidth combined with the full T 2*‐refocusing lead to the higher robustness to B 0 inhomogeneity artifacts compared to the SE‐EPI image, while SPEN’s referenceless reconstruction algorithm 47 allows one to minimize ghost artifacts. C, Comparison of normalized profiles extracted from the aforementioned SE‐EPI and SPEN images along the blue lines. Arrows indicate the above‐mentioned artefacts. D, b 0 SPEN image acquired with eight averages leading to an acquisition time of 24 seconds and an echo time increased to 59 ± 11.5 ms due to the inclusion of a diffusion delay (T diff) of 18 ms. This image was denoised using random matrix theory 60 and a 3 × 3 bidimensional kernel, and is representative of the sets employed in the subsequent DTD calculations
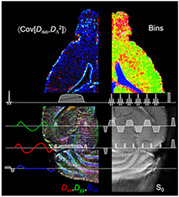
Figure 3 displays some aspects of the DTD experiments and the ensuing SPEN data for three representative voxels containing WM, GM and CSF. For simplicity these results are shown as a function of an acquisition point index (Figure 3A), characterized by independent b‐tensor parameters. Each of the voxels yields a distinct signal pattern as a function of this index, highlighting the contrast provided by DTD in relation to the underlying microstructure. The WM voxel is characterized by a signal pattern that is highly influenced by both the anisotropy (b Δ) and the orientation (Θ, Φ) of the b‐tensor, a behavior that is consistent with anisotropic diffusion within and along orientationally ordered axons that dominate WM. CSF and GM voxels yield signal data that are insensitive to both b Δ and (Θ, Φ), indicative of isotropic media. Yet the CSF signal differs from GM signals by its sharper decay with increasing b‐values, reflecting a higher diffusivity. These various signal patterns can be processed as described above, to retrieve distinct parameters quantifying the main diffusivity features of the underlying microscopic environments. As done in previous studies, 26 , 32 , 34 , 35 Figure 4A reports these distributions in a logarithmic space based on the diffusion tensor's “size” log10(D iso), and “shape” log10(D ||/D ⊥). Within this representation we observe that CSF is characterized by large (log10(D iso/m2s−1) ≈ −8.5) and isotropic (log10(D ||/D ⊥) ≈ 0) diffusion tensors, that WM and GM both present a lower MD compared with CSF (−9.2 < log10(D iso/m2s−1) < −8.8), and that the DTDs from WM comprise highly anisotropic diffusion components (log10(D ||/D ⊥) ≈ 1). While the displayed DTDs capture these gross properties even from a simple visual inspection of the signal patterns, the accuracy of the data inversion process is challenged by the images' limited SNR. Noise scatters the signal profiles (see, for instance, Figure 3C), giving rise to a general broadening of the distributions as well as to the emergence of spurious components that are not biologically plausible; eg, the planar components with log10(D ||/D ⊥) ≈ −2 for WM and GM in Figure 4A. A more subtle effect also noticed for the noise, is to bias the inversion algorithm towards finding combinations of anisotropic components, instead of a single isotropic one; this gives rise to the nearly symmetric “butterfly” spread of components centered about the log10(D ||/D ⊥) = 0 line in the “Thick” DTD. This effect is analogous to the well‐known positive bias in the anisotropy metrics of conventional DTI at low SNR. 31 A map of the ratio between the initial signal amplitude S 0 and the square root of the residual sum of squares (RSS), S 0/(RSS)1/2, is included in Figure S1 panel A, and provides an assessment of the SNR levels throughout the whole brain images. A second map, describing the ratio between the mean of S(b,b Δ = 0) data points and their standard deviation at the highest b‐value (3.499 x 109 s/m2), is also presented in Figure S1 panel B, and shows that signal still remains at high T 2 and b‐value weightings. SNR‐limited scattering notwithstanding, we find it convenient to divide the experimental DTD SPEN results into bins that we refer to as “Thin”, “Thick” and “Big”, to describe the visual appearance of the corresponding tensor glyphs; this facilitates the separation of signal contributions into components with distinct diffusion properties. Slowly diffusing anisotropic and isotropic components appear in the bins labeled Thin and Thick, respectively, while fast‐diffusing components are captured by the Big bin. Figure 4B shows the boundaries that were assumed for these bins. Figure 4C maps how the sampled signals fractionate into these bins, with the aid of an additive color map display. CSF‐rich areas such as the lateral ventricles (LV), the dorsal third ventricle (D3V) and the sylvius aqueduct (Aq), are all well captured by the Big bin (blue voxels). WM‐rich areas such as (from bottom to top) the WM in the cerebellum (CB, which correspond to the intense red voxels), the fimbria (fi, which correspond to the red voxels below the LV) and the corpus callosum (cc, which correspond to the dispersed red voxels above and in between the LV), appear primarily as the Thin bin. So do the granular layer (GrO) as well as the granular insular cortex (Gl) of the olfactory bulb, which are, respectively, the central and lateral structures of this organ. While literature reports and preliminary data (not shown) validate the main features of high FA in the GrO and GI structures, as reflected in Figure 4C for the olfactory bulb, SNR limitations in this region probably exacerbate spurious components with high anisotropy. This may lead to an overestimation of E[D Δ 2], as well as to an overestimation of the diffusion anisotropy and of the Thin bin fractions observed in this region. The separation of Thin and Thick signal populations also identifies WM/GM partial volume effects in the vicinity of the cc (yellow voxels above the ventricles), and in the CB at the edges of the WM areas. The mixture of red and yellow voxels characterizing the cc in Figure 4 is in part reflecting partial volume effects arising mainly due to the relatively large slice thickness chosen (0.7 mm), but is also reinforced by the human brain‐based boundary values chosen for the various bins. Indeed, the mouse cc is intrinsically not as anisotropic as the human one: the conventional FA in cc is ~ 0.5‐0.6 for mouse, 19 and ≈0.8‐0.9 for human. Hence, values of ≈0.5 fall on the edge between the Thin and Thick bins optimized for human brain segmentations, 63 and result in the admixing of a yellow contribution to this representation.
FIGURE 3.
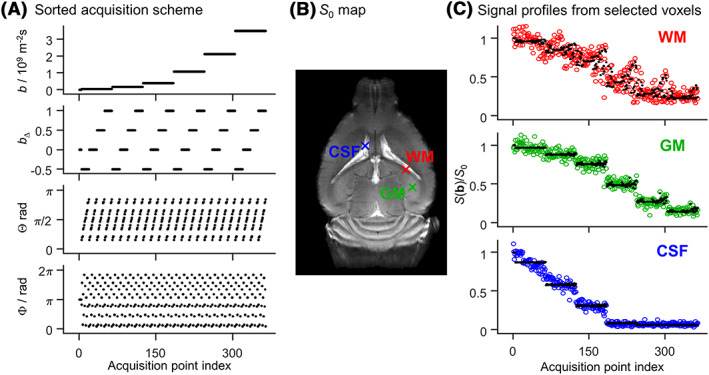
Acquisition protocol and signal data from three representative voxels collected from an in vivo mouse brain. (A) Acquisition protocol comprising 364 diffusion encoding tensors b 34 with six different magnitudes b, four different values of normalized anisotropy b Δ, and 15 orientations (Θ, Φ). The (b, b Δ, Θ, Φ) coordinates are displayed as a function of acquisition point index. (B) Initial signal amplitude S 0, with crosses indicating three voxels containing white matter (WM, red), gray matter (GM, green) and cerebrospinal fluid (CSF, blue). (C) Experimental (colored circles) and fitted (black dots) signal profiles from the three selected voxels. The normalized signals S(b)/S 0 are sorted according to the acquisition scheme shown in (A)
FIGURE 4.
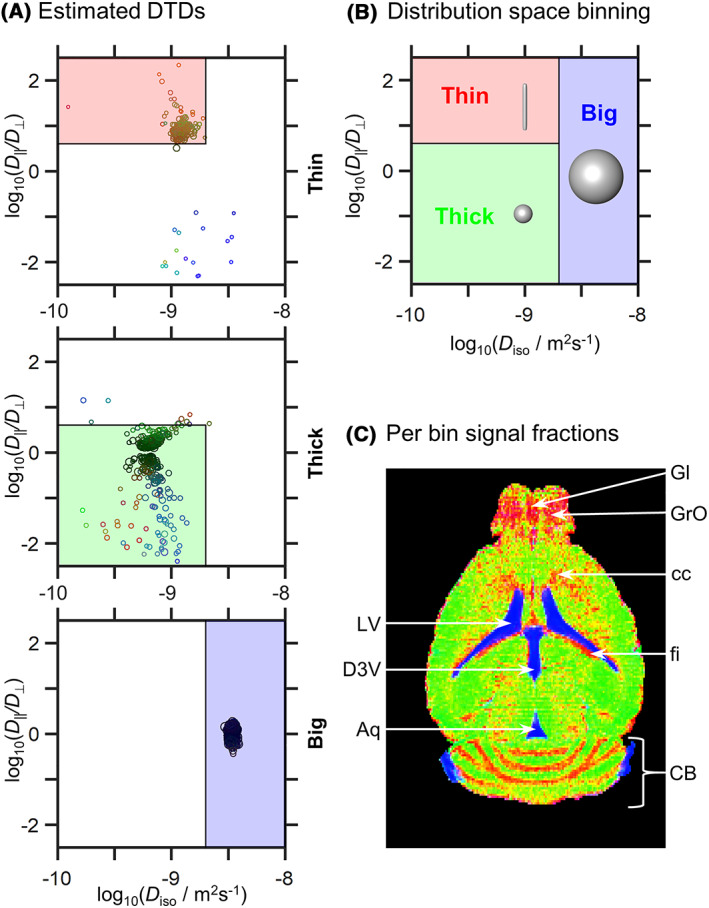
Distribution space and parameter maps derived from subsets (“bins”) of the 4D DTD space. (A) DTDs obtained by nonparametric inversion of the signal data displayed as 2D logarithmic bubble charts of isotropic diffusivities D iso and axial‐radial diffusivity ratios D ∥/D ⊥. The component weights and orientations are indicated with the circle areas and colors [R, G, B] = [cosø sinθ, sinø sinθ, cosθ]. (B) Division of the DTD space into three different bins—Thin, Thick and Big—that loosely capture the diffusion features of white matter, gray matter and cerebrospinal fluid, respectively. (C) Signal contributions from the fractional populations in the different bins are visualized as an additive color map ([R, G, B] = [Thin, Thick, Big]). Voxels comprising multiple fractions are visible as a weighted superposition of the three primary colors, eg, equal proportions of Thin and Thick populations yield a yellow voxel. Legend abbreviations 64 : LV, lateral ventricles; D3V, dorsal third ventricle; Aq, sylvius aqueduct; Gl, granular insular cortex of the olfactory bulb; GrO, granular layer of the olfactory bulb; cc, corpus callosum; fi: fimbria; CB, cerebellum
Analyses of the mean parameter maps enable further characterization of the bin‐resolved signal fractions (Figure 5). For example, inspection of the 〈E[Orientation]〉 maps from the Thin bin reveals the alignment direction of the anisotropic tissues found in the CB and the fi. It should be noted that the Thin 〈E[x]〉 maps possess a noisy appearance that is characterized by the existence of multiple voxels with low brightness. While such voxels hamper the visual inspection of the Thin maps, their reduced brightness indicates that they have a small contribution to the retrieved DTDs (this is consistent with the overall “green” appearance of the composite map in Figure 4C). For simplifying the analysis of these distributions, the spatially resolved DTDs were converted into 16 individual parameter maps, following the procedure detailed in the Materials and Methods (section 2). Selected parameter maps derived in this manner from the full DTD space are shown in Figure 5. Conventional diffusion protocols can be used to estimate S 0 and E[D iso], whereas the E[D Δ 2], Var[D iso], Var[D Δ 2] and Cov[D iso,D Δ 2] metrics can only be retrieved with tensor‐valued diffusion protocols. 25 , 28 , 31 The E[D iso] and E[D Δ 2] metrics provide measures of MD and diffusion anisotropy, respectively, and can be used to obtain a rough characterization of the spatial distribution of CSF (high E[D iso], low E[D Δ 2]), WM (low E[D iso], high E[D Δ 2]) and GM (low E[D iso], low E[D Δ 2]). It is important to highlight that the E[D Δ 2] metric quantifies diffusion anisotropy independently from the underlying degree of orientational order; this is in contrast to common anisotropy metrics such as the FA, 7 which convolve the fundamentally different properties of diffusion anisotropy and orientational order into a single scalar value. 29 Notice that, according to the 〈E[D Δ 2]〉 map, this mean anisotropy metric is for the most part characterized by small but nonzero values. Whether such small anisotropy reflects the tissue properties of the mouse brain or originates from the bias discussed in the previous paragraph is a point that merits further investigation. Intra‐voxel heterogeneities can be further studied through inspection of (co)variance maps of the type shown in Figure 6. Notice, for instance, how voxels comprising mixtures of CSF and GM (in red) display intermediate values of E[D iso] and are characterized by elevated Var[D iso], nonnegligible Var[D Δ 2] and negative Cov[D iso,D Δ 2]. The high Var[D iso] indicates a mixture of slow‐ and fast‐diffusing components, 29 and the negative Cov[D iso,D Δ 2] suggests that the slower diffusing components possess a higher diffusion anisotropy. 32 These results are consistent with a linear combination of the diffusion properties measured in pure CSF (〈E[D iso]〉 ≈ 3.1 x10–9 m2s−1, 〈E[D Δ 2]〉 ≈ 0) and GM (〈E[D iso]〉 ≈ 0.7 x 10−9 m2s−1, 〈E[D Δ 2]〉 ≈ 0.2) voxels. The Var[D Δ 2] metric has been shown 32 to correctly identify voxels containing both isotropic and anisotropic diffusion components in a synthetic phantom. The nonzero 〈Var[D Δ 2]〉 values may then be interpreted as indicating mixtures of anisotropic and isotropic tissues.
FIGURE 5.
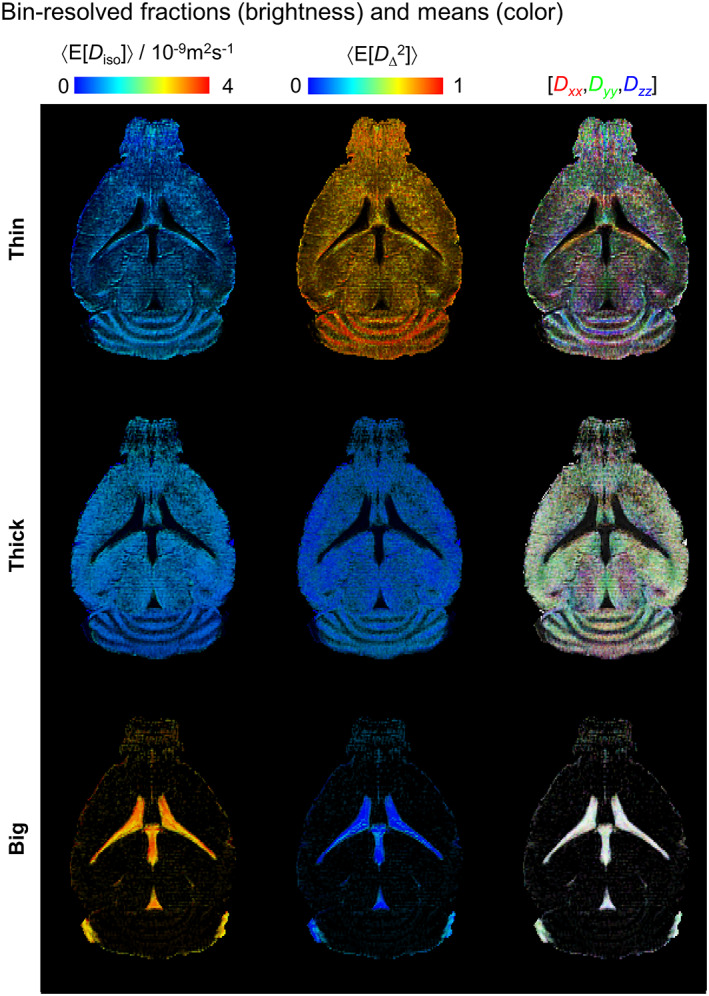
Parameter maps derived from subsets (“bins”) of the 4D DTD space. Per‐bin signal fractions (brightness) and per‐bin average mean values (color) are shown for the isotropic diffusivity 〈E[D iso]〉, the squared normalized diffusion anisotropy 〈E[D Δ 2]〉, and the diffusion tensor orientation 〈E[Orientation]〉. The values of 〈E[D iso]〉 and 〈E[D Δ 2]〉 are indicated by the corresponding linear color scales, while the 〈E[Orientation]〉 maps are color‐coded as [R, G, B] = [Dxx,Dyy,Dzz]/max(Dxx,Dyy,Dzz), where Dii are the i‐th diagonal elements of the diffusion tensor as measured in the laboratory frame of reference
FIGURE 6.
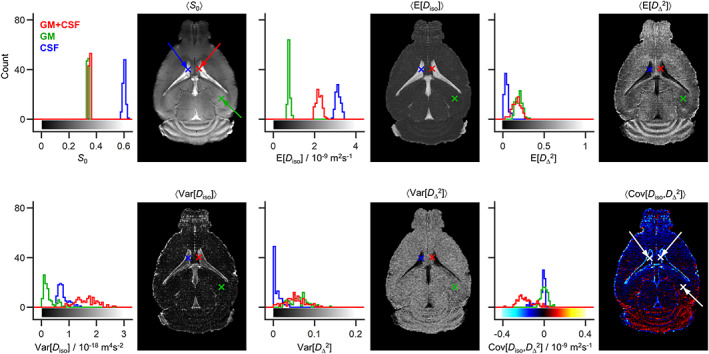
Statistical descriptors derived from the 4D diffusion tensor distributions (DTDs) introduced in Figures 4 and 5. For each voxel, the bootstrapping yields 96 different DTDs consistent with the signal data. The sets of DTDs were converted to sets of the following statistical descriptors: total signal amplitude S 0, mean E[x], variance Var[x] and covariance Cov[x,y], for the isotropic diffusivity D iso and the squared normalized diffusion anisotropy D Δ 2. The parameter maps show 〈S 0〉, 〈E[x]〉, 〈Var[x]〉 and 〈Cov[x,y]〉, where the angular brackets indicate averages over these 96 bootstrap realizations. The results from all 96 bootstrap realizations are also shown as histograms for three selected voxels containing gray matter (GM, in green), cerebrospinal fluid (CSF in blue), and a binary mixture of GM and CSF (GM + CSF, in red). The linear color scales of the various maps are indicated by the bars along the horizontal axes of the corresponding histograms
The borders of the three bins introduced in Figure 4 were selected to demarcate the DTD components observed in pure WM, GM and CSF voxels. In order to more thoroughly test this analysis strategy, synthetic phantoms emulating the diffusion properties of the tissue components were also investigated. This set of synthetic samples included: a “hex” phantom, where the confinement of water into nanometer‐diameter channels in a matrix of detergent and oil leads to slow anisotropic diffusion as in WM 52 ; a neat liquid dodecane sample with a slow isotropic diffusion akin to that of GM 65 ; and pure water, where fast isotropic diffusion happens, as in CSF. These phantoms were subjected to the same DTD SPEN analyses as the living mice brains; Figure 7 shows their results. Notice how all their DTD components fall cleanly within the designated bins: hex – “Thin”, dodecane – “Thick” and water – “Big”. For all phantoms, the 〈E[D iso]〉 and 〈E[D Δ 2]〉 maps are also constant throughout the imaged volumes, having values consistent with expected diffusion properties. Additionally, the maps with the intra‐voxel heterogeneity measures, 〈Var[D iso]〉, 〈Var[D Δ 2]〉 and 〈Cov[D iso,D Δ 2]〉, show vanishingly low values. The speckled appearance of the 〈S 0〉 map for the hex phantom results from variations in the local B 0 field induced by the pronounced magnetic susceptibility anisotropy of the liquid crystalline domains, the orientations of which are illustrated in the 〈E[Orientation]〉 map. To facilitate comparison with the phantom data, the in vivo maps from Figure 6 and Figure 4 are reproduced in Figure 7. These phantom results—with their clean separation into the expected bins, constant 〈E[D iso]〉 and 〈E[D Δ 2]〉 maps, and nearly null values of 〈Var[D iso]〉, 〈Var[D Δ 2]〉 and 〈Cov[D iso,D Δ 2]〉—verify that our acquisition and analysis pipeline has the capability to characterize components with the diffusion hallmarks of WM, GM and CSF. A second set of in vivo mouse brain DTD parameter maps and statistical descriptors is also presented in Figure S2. It has been acquired on the second mouse (n = 2) with identical parameters as previously described.
FIGURE 7.
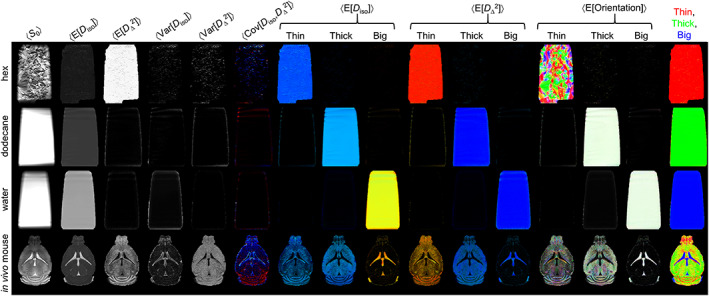
SPEN DTD parameter maps of phantoms designed to give diffusion properties similar to the white matter, gray matter and cerebrospinal fluid components in in vivo mouse brain. Phantom and in vivo data were acquired with the same hardware, imaging parameters and DTD acquisition protocol as reported in Figure 3 and Table 1. Row 1: hex 51 (anisotropic) phantom, row 2: dodecane (slow isotropic) phantom, row 3: water (fast isotropic) phantom, row 4: in vivo mouse brain. The parameter maps are as explained in Figures 4 and 5. Note that for the phantoms, all heterogeneity measures Var[D iso], Var[D Δ 2] and Cov[D iso,D Δ 2] are zero and that all DTD components fall within the intended bins: Thin: hex phantom, Thick: dodecane, and Big: water
4. CONCLUSIONS
Combining the SNR advantages of very high magnetic fields and surface cryocoils with the robustness of spatiotemporal encoding towards B 0‐ and B 1‐inhomogeneity enabled us to image DTDs for in vivo mouse brains at ≈0.1 mm in‐plane spatial resolutions. As mentioned, the high in‐plane spatial resolution targeted in this study was associated with sensitivity penalties, and hence with a need for performing multi‐scan, multi‐shot interleaved averages, to obtain the kind of SNR required for nonparametric DTD inversion. SPEN MRI, which provides its data directly in image space, is well endowed to perform such joint multi‐scan/multi‐shot processing with minimal interferences from potential motional artifacts. Still, the long acquisition times required by these procedures led us to their realization in solely a single slice. Multi‐slice or 3D acquisitions would also have been possible, but keeping their overall scanning times within our self‐imposed 2.5‐hour limit would have required a substantial lowering of the spatial resolution. We estimate, for instance, that increasing the in‐plane voxel size from 100 to 170 μm would have allowed us to obtain the required SNR while reducing the per‐slice scanning time by an order of magnitude, leading to durations that would be compatible with volumetric acquisitions. This approach possessed distinct limitations including relatively long effective echo times and a limited range of b‐values, which may have skewed the DTDs and their derived statistical descriptors towards components with longer T 2 and slower diffusivities. However, apart from a general decrease in S 0 along the SPEN dimension (Figure 7), these features did not appear to introduce an evident bias in the DTD metrics, even for voxels containing multiple components with distinctly different relaxation or diffusion properties. Rigorously taking care of these effects would require including an additional acquisition dimension probing a range of echo times 63 ; such investigation is onerous in terms of sensitivity and acquisition time, but it is in progress. Overall, these developments pave the way for investigating in further detail the cell densities, shapes and orientations in animal models at very high spatial resolution, thereby leading to new insights into tissue microstructure in health and disease. Further improvements of the methods presented here would enable translation into the realm of human MRI.
FUNDING INFORMATION
The authors acknowledge support from the Israel Science Foundation (grants 2508/17 and 965/18), the Swedish Foundation for Strategic Research (AM13‐0090, ITM17‐0267), the Swedish Research Council (2018‐03697), the Kimmel Institute for Magnetic Resonance (Weizmann Institute) and the generosity of the Perlman Family Foundation.
Supporting information
Figure S1: A) Map of the SNR calculated by with S 0 the initial signal amplitude and RSS the square‐root of the residual sum of squares (RSS). B) Map of the ratio between the mean of S(b,b Δ = 0) data points and their standard deviation, Mean[S(b,b Δ = 0)]/SD[S(b,b Δ = 0)] for b = 3.499·109 s/m2. Notice that even at the maximum acquired b‐value some signal is remaining.
Figure S2: Parameter maps and statistical descriptors derived from 4D DTD acquisition of the second mouse (n = 2). As a reminder: total signal amplitude S 0, mean E[x], variance Var[x], and covariance Cov[x,y], for the isotropic diffusivity D iso and the squared normalized diffusion anisotropy . The parameter maps show 〈S 0〉, 〈E[x]〉, 〈Var[x]〉, and 〈Cov[x,y]〉 where the angular bracket indicate average over these 96 bootstrap realization. Per‐bin signal fractions (brightness) and per‐bin average mean values (color) are shown for the isotropic diffusivity 〈E[D iso]〉, the squared normalized diffusion anisotropy 〈E[ ]〉, and the diffusion tensor orientation 〈E[Orientation]〉. The values of 〈E[D iso]〉 and 〈E[ ]〉 are indicated by the corresponding linear color scales, while the 〈E[Orientation]〉 maps are color‐coded as [R,G,B] = [D xx,D yy,D zz]/max (D xx,D yy,D zz), where D ii are the i‐th diagonal elements of the diffusion tensor as measured in the laboratory frame of reference.
ACKNOWLEDGEMENTS
We are grateful to Dr. Sascha Koehler (Bruker BioSpin) for his help with the programming of the SPEN DTD pulse sequences. The authors also acknowledge the support from the Israel Science Foundation (grants 2508/17 and 965/18), the Swedish Foundation for Strategic Research (AM13‐0090, ITM17‐0267), the Swedish Research Council (2018‐03697), the Kimmel Institute for Magnetic Resonance (Weizmann Institute) and the generosity of the Perlman Family Foundation.
Yon M, de Almeida Martins JP, Bao Q, Budde MD, Frydman L, Topgaard D. Diffusion tensor distribution imaging of an in vivo mouse brain at ultrahigh magnetic field by spatiotemporal encoding. NMR in Biomedicine. 2020;33:e4355 10.1002/nbm.4355
REFERENCES
- 1. Le Bihan D. Molecular diffusion, tissue microdynamics and microstructure. NMR Biomed. 1995;8(7):375‐386. [DOI] [PubMed] [Google Scholar]
- 2. Beaulieu C. The basis of anisotropic water diffusion in the nervous system ‐ A technical review. NMR Biomed. 2002;15(7–8):435‐455. [DOI] [PubMed] [Google Scholar]
- 3. Le Bihan D. Looking into the functional architecture of the brain with diffusion MRI. Nat Rev Neurosci. 2003;4(6):469‐480. [DOI] [PubMed] [Google Scholar]
- 4. Le Bihan D, Breton E, Lallemand D, Grenier P, Cabanis E, Laval‐Jeantet M. MR imaging of intravoxel incoherent motions: Application to diffusion and perfusion in neurologic disorders. Radiology. 1986;161(2):401‐407. [DOI] [PubMed] [Google Scholar]
- 5. Moseley ME, Cohen Y, Mintorovitch J, et al. Early detection of regional cerebral ischemia in cats: Comparison of diffusion‐ and T2‐weighted MRI and spectroscopy. Magn Reson Med. 1990;14(2):330‐346. [DOI] [PubMed] [Google Scholar]
- 6. Moseley ME, Cohen Y, Kucharczyk J, et al. Diffusion‐weighted MR imaging of anisotropic water diffusion in cat central nervous system. Radiology. 1990;176(2):439‐445. [DOI] [PubMed] [Google Scholar]
- 7. Basser PJ, Pierpaoli C. Microstructural and physiological features of tissues elucidated by quantitative‐diffusion‐tensor MRI. J Magn Reson ‐ Ser B. 1996;111(3):209‐219. [DOI] [PubMed] [Google Scholar]
- 8. Le Bihan D, Mangin J‐F, Poupon C, et al. Diffusion tensor imaging: Concepts and applications. J Magn Reson Imaging. 2001;13(4):534‐546. [DOI] [PubMed] [Google Scholar]
- 9. Basser PJ, Mattiello J, LeBihan D. MR diffusion tensor spectroscopy and imaging. Biophys J. 1994;66(1):259‐267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Horsfield MA, Jones DK. Applications of diffusion‐weighted and diffusion tensor MRI to white matter diseases ‐ A review. NMR Biomed. 2002;15(7–8):570‐577. [DOI] [PubMed] [Google Scholar]
- 11. Basser PJ, Pajevic S, Pierpaoli C, Duda J, Aldroubi A. In vivo fiber tractography using DT‐MRI data. Magn Reson Med. 2000;44(4):625‐632. [DOI] [PubMed] [Google Scholar]
- 12. Conturo TE, Lori NF, Cull TS, et al. Tracking neuronal fiber pathways in the living human brain. Proc Natl Acad Sci. 1999;96(18):10422‐10427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Mori S, Crain BJ, Chacko VP, Van Zijl PCM. Three‐dimensional tracking of axonal projections in the brain by magnetic resonance imaging. Ann Neurol. 1999;45(2):265‐269. [DOI] [PubMed] [Google Scholar]
- 14. Jones DK, Knösche TR, Turner R. White matter integrity, fiber count, and other fallacies: the do's and don'ts of diffusion MRI. Neuroimage. 2013;73:239‐254. [DOI] [PubMed] [Google Scholar]
- 15. Jeurissen B, Leemans A, Tournier JD, Jones DK, Sijbers J. Investigating the prevalence of complex fiber configurations in white matter tissue with diffusion magnetic resonance imaging. Hum Brain Mapp. 2013;34(11):2747‐2766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Szczepankiewicz F, Lasič S, van Westen D, et al. Quantification of microscopic diffusion anisotropy disentangles effects of orientation dispersion from microstructure: Applications in healthy volunteers and in brain tumors. Neuroimage. 2015;104:241‐252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Jaermann T, De Zanche N, Staempfli P, et al. Preliminary experience with visualization of intracortical fibers by focused high‐resolution diffusion tensor imaging. Am J Neuroradiol. 2008;29(1):146‐150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Yassa MA, Muftuler LT, Stark CEL. Ultrahigh‐resolution microstructural diffusion tensor imaging reveals perforant path degradation in aged humans in vivo. Proc Natl Acad Sci. 2010;107(28):12687‐12691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Wu D, Xu J, McMahon MT, et al. In vivo high‐resolution diffusion tensor imaging of the mouse brain. Neuroimage. 2013;83:18‐26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wu D, Reisinger D, Xu J, et al. Localized diffusion magnetic resonance micro‐imaging of the live mouse brain. Neuroimage. 2014;91:12‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Jian B, Vemuri BC, Özarslan E, Carney PR, Mareci TH. A novel tensor distribution model for the diffusion‐weighted MR signal. Neuroimage. 2007;37(1):164‐176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Basser PJ, Mattiello J, Lebihan D. Estimation of the effective self‐diffusion tensor from the NMR spin echo. J Magn Reson Ser B. 1994;103(3):247‐254. [DOI] [PubMed] [Google Scholar]
- 23. Anderson AW. Measurement of fiber orientation distributions using high angular resolution diffusion imaging. Magn Reson Med.. 2005;54(5):1194‐1206. [DOI] [PubMed] [Google Scholar]
- 24. Eriksson S, Lasič S, Nilsson M, Westin C‐F, Topgaard D. NMR diffusion‐encoding with axial symmetry and variable anisotropy: Distinguishing between prolate and oblate microscopic diffusion tensors with unknown orientation distribution. J Chem Phys. 2015;142(10):104201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Topgaard D. Chapter 7. NMR Methods for Studying Microscopic Diffusion Anisotropy In: Diffusion NMR of Confined Systems: Fluid Transport in Porous Solids and Heterogeneous Materials. The Royal Society of Chemistry; 2016:226‐259. [Google Scholar]
- 26. Topgaard D. Multiple dimensions for random walks. J Magn Reson. 2019;306:1‐5. [DOI] [PubMed] [Google Scholar]
- 27. Lawrenz M, Koch MA, Finsterbusch J. A tensor model and measures of microscopic anisotropy for double‐wave‐vector diffusion‐weighting experiments with long mixing times. J Magn Reson. 2010;202(1):43‐56. [DOI] [PubMed] [Google Scholar]
- 28. Jespersen SN, Lundell H, Sønderby CK, Dyrby TB. Orientationally invariant metrics of apparent compartment eccentricity from double pulsed field gradient diffusion experiments. NMR Biomed. 2013;26(12):1647‐1662. [DOI] [PubMed] [Google Scholar]
- 29. Lasič S, Szczepankiewicz F, Eriksson S, Nilsson M, Topgaard D. Microanisotropy imaging: quantification of microscopic diffusion anisotropy and orientational order parameter by diffusion MRI with magic‐angle spinning of the q‐vector. Front Physiol. 2014;2(February):1‐14. [Google Scholar]
- 30. Shemesh N, Jespersen SN, Alexander DC, et al. Conventions and nomenclature for double diffusion encoding NMR and MRI. Magn Reson Med. 2016;75(1):82‐87. [DOI] [PubMed] [Google Scholar]
- 31. Pierpaoli C, Basser PJ. Toward a quantitative assessment of diffusion anisotropy. Magn Reson Med. 1996;36(6):893‐906. [DOI] [PubMed] [Google Scholar]
- 32. Topgaard D. Diffusion tensor distribution imaging. NMR Biomed. 2019;32(November 2018):1‐12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Szczepankiewicz F, van Westen D, Englund E, et al. The link between diffusion MRI and tumor heterogeneity: Mapping cell eccentricity and density by diffusional variance decomposition (DIVIDE). Neuroimage. 2016;142:522‐532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. De Almeida Martins JP, Topgaard D. Two‐dimensional correlation of isotropic and directional diffusion using NMR. Phys Rev Lett. 2016;116(8):1‐6. [DOI] [PubMed] [Google Scholar]
- 35. De Almeida Martins JP, Topgaard D. Multidimensional correlation of nuclear relaxation rates and diffusion tensors for model‐free investigations of heterogeneous anisotropic porous materials. Sci Rep. 2018;8(1):1‐12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Topgaard D. Multidimensional diffusion MRI. J Magn Reson. 2017;275:98‐113. [DOI] [PubMed] [Google Scholar]
- 37. Bax A, Szeverenyi NM, Maciel GE. Chemical shift anisotropy in powdered solids studied by 2D FT NMR with flipping of the spinning axis. J Magn Reson. 1983;55(3):494‐497. [Google Scholar]
- 38. Frydman L, Chingas GC, Lee YK, et al. Variable‐angle correlation spectroscopy in solid‐state nuclear magnetic resonance. J Chem Phys. 1992;97(7):4800‐4808. [Google Scholar]
- 39. Prange M, Song YQ. Quantifying uncertainty in NMR T2 spectra using Monte Carlo inversion. J Magn Reson. 2009;196(1):54‐60. [DOI] [PubMed] [Google Scholar]
- 40. Darrasse L, Ginefri J‐C. Perspectives with cryogenic RF probes in biomedical MRI. Biochimie. 2003;85(9):915‐937. [DOI] [PubMed] [Google Scholar]
- 41. Ladd ME, Bachert P, Meyerspeer M, et al. Pros and cons of ultra‐high‐field MRI/MRS for human application. Prog Nucl Magn Reson Spectrosc. 2018;109:1‐50. [DOI] [PubMed] [Google Scholar]
- 42. Butts K, Riederer SJ, Ehman RL, Thompson RM, Jack CR. Interleaved echo planar imaging on a standard MRI system. Magn Reson Med. 1994;31(1):67‐72. [DOI] [PubMed] [Google Scholar]
- 43. Ben‐Eliezer N, Frydman L. Spatiotemporal encoding as a robust basis for fast three‐dimensional in vivo MRI. NMR Biomed. 2011;24(10):1191‐1201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Schmidt R, Frydman L. New spatiotemporal approaches for fully refocused, multislice ultrafast 2D MRI. Magn Reson Med. 2014;71(2):711‐722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Solomon E, Liberman G, Nissan N, Frydman L. Robust diffusion tensor imaging by spatiotemporal encoding: Principles and in vivo demonstrations. Magn Reson Med. 2017;77(3):1124‐1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Seginer A, Schmidt R, Leftin A, Solomon E, Frydman L. Referenceless reconstruction of spatiotemporally encoded imaging data: Principles and applications to real‐time MRI. Magn Reson Med. 2014;72(6):1687‐1695. [DOI] [PubMed] [Google Scholar]
- 47. Schmidt R, Seginer A, Frydman L. Interleaved multishot imaging by spatiotemporal encoding: A fast, self‐referenced method for high‐definition diffusion and functional MRI. Magn Reson Med. 2016;75(5):1935‐1948. [DOI] [PubMed] [Google Scholar]
- 48. Bao Q, Liberman G, Solomon E, Frydman L. High‐resolution diffusion MRI studies of development in pregnant mice visualized by novel spatiotemporal encoding schemes. NMR Biomed. 2020;33(3):e4208. [DOI] [PubMed] [Google Scholar]
- 49. Li J, Zhang M, Chen L, Cai C, Sun H, Cai S. Reduced field‐of‐view imaging for single‐shot MRI with an amplitude‐modulated chirp pulse excitation and Fourier transform reconstruction. Magn Reson Imaging. 2015;33(5):503‐515. [DOI] [PubMed] [Google Scholar]
- 50. Cousin SF, Liberman G, Solomon E, Otikovs M, Frydman L. A regularized reconstruction pipeline for high‐definition diffusion MRI in challenging regions incorporating a per‐shot image correction. Magn Reson Med. 2019;82(4):1322‐1330. [DOI] [PubMed] [Google Scholar]
- 51. Yang QX, Posse S, Bihan DLE, Smith MB. Double‐sampled echo‐planar imaging at 3 tesla. J Magn Reson ‐ Ser B. 1996;113(2):145‐150. [DOI] [PubMed] [Google Scholar]
- 52. Nilsson M, Larsson J, Lundberg D, et al. Liquid crystal phantom for validation of microscopic diffusion anisotropy measurements on clinical MRI systems. Magn Reson Med. 2018;79(3):1817‐1828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Vanzijl PCM, Sukumar S, Johnson MO, Webb P, Hurd RE. Optimized shimming for high‐resolution NMR using three‐dimensional image‐based field mapping. J Magn Reson Ser A. 1994;111(2):203‐207. [Google Scholar]
- 54. Schneider E, Glover G. Rapid in vivo proton shimming. Magn Reson Med. 1991;18(2):335‐347. [DOI] [PubMed] [Google Scholar]
- 55. Wen H, Jaffer FA. An in vivo automated shimming method taking into account shim current constraints. Magn Reson Med. 1995;34(6):898‐904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Solomon E, Shemesh N, Frydman L. Diffusion weighted MRI by spatiotemporal encoding: Analytical description and in vivo validations. J Magn Reson. 2013;232:76‐86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Bruder H, Fischer H, Reinfelder HE, Schmitt F. Image reconstruction for echo planar imaging with nonequidistant k‐space sampling. Magn Reson Med. 1992;23(2):311‐323. [DOI] [PubMed] [Google Scholar]
- 58. Topgaard D. Director orientations in lyotropic liquid crystals: Diffusion MRI mapping of the Saupe order tensor. Phys Chem Chem Phys. 2016;18(12):8545‐8553. [DOI] [PubMed] [Google Scholar]
- 59. Han SH, Son JP, Cho HJ, Park JY, Kim SG. Gradient‐echo and spin‐echo blood oxygenation level–dependent functional MRI at ultrahigh fields of 9.4 and 15.2 Tesla. Magn Reson Med. 2019;81(2):1237‐1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Veraart J, Novikov DS, Christiaens D, Ades‐aron B, Sijbers J, Fieremans E. Denoising of diffusion MRI using random matrix theory. Neuroimage. 2016;142:394‐406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. De Kort DW, Van Duynhoven JPM, Hoeben FJM, Janssen HM, Van As H. NMR nanoparticle diffusometry in hydrogels: Enhancing sensitivity and selectivity. Anal Chem. 2014;86(18):9229‐9235. [DOI] [PubMed] [Google Scholar]
- 62. Bao Q, Liberman G, Solomon E, Lustig M, Frydman L. Diffusion‐weighted in vivo imaging with =100 um resolution: principles and applications to ADC mapping of pregnant mice. Proc Intl Soc Mag Res Med. 2018:1021. [Google Scholar]
- 63. de Almeida Martins JP, Tax CMW, Szczepankiewicz F, Jones DK, Westin C‐F, Topgaard D. Transferring principles of solid‐state and Laplace NMR to the field of in vivo brain MRI. Magn Reson. 2020;1(1):27‐43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Franklin KBJ, Paxinos G. The mouse brain in stereotaxic coordinates. 3rd ed. New York: Academic press; 2008. [Google Scholar]
- 65. Holz M, Heil SR, Sacco A. Temperature‐dependent self‐diffusion coefficients of water and six selected molecular liquids for calibration in accurate 1H NMR PFG measurements. Phys Chem Chem Phys. 2000;2(20):4740‐4742. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1: A) Map of the SNR calculated by with S 0 the initial signal amplitude and RSS the square‐root of the residual sum of squares (RSS). B) Map of the ratio between the mean of S(b,b Δ = 0) data points and their standard deviation, Mean[S(b,b Δ = 0)]/SD[S(b,b Δ = 0)] for b = 3.499·109 s/m2. Notice that even at the maximum acquired b‐value some signal is remaining.
Figure S2: Parameter maps and statistical descriptors derived from 4D DTD acquisition of the second mouse (n = 2). As a reminder: total signal amplitude S 0, mean E[x], variance Var[x], and covariance Cov[x,y], for the isotropic diffusivity D iso and the squared normalized diffusion anisotropy . The parameter maps show 〈S 0〉, 〈E[x]〉, 〈Var[x]〉, and 〈Cov[x,y]〉 where the angular bracket indicate average over these 96 bootstrap realization. Per‐bin signal fractions (brightness) and per‐bin average mean values (color) are shown for the isotropic diffusivity 〈E[D iso]〉, the squared normalized diffusion anisotropy 〈E[ ]〉, and the diffusion tensor orientation 〈E[Orientation]〉. The values of 〈E[D iso]〉 and 〈E[ ]〉 are indicated by the corresponding linear color scales, while the 〈E[Orientation]〉 maps are color‐coded as [R,G,B] = [D xx,D yy,D zz]/max (D xx,D yy,D zz), where D ii are the i‐th diagonal elements of the diffusion tensor as measured in the laboratory frame of reference.