

Abstract
Glaucoma is a progressive neurodegenerative process affecting the retinal ganglion cells (RGCs) and the optic nerve. Oxidative stress has been implicated in glaucoma pathogenesis, and iron is a potent generator of oxidative stress. The oral iron chelator deferiprone (DFP) is protective against retinal degenerations associated with oxidative stress. To test whether DFP could be protective in glaucoma, we used microbead injections to induce elevated intraocular pressure (IOP) in a cohort of 3-month old C57BL/6J mice. One eye of each animal was injected with magnetic microbeads resulting in ocular hypertension for >7 weeks while the fellow eye was injected with saline and served as a normotensive internal control. While half of the cohort received oral DFP (1 mg/ml in the drinking water), the other half did not and served as controls. After 8 weeks, Brn3a immunolabeling of flat-mounted retinas was used for manual RGC quantification. Axon counts were obtained from thin sections of optic nerves using the AxonJ plugin for ImageJ. DFP administration was protective against RGC and optic nerve loss in the setting of elevated IOP. These results suggest that iron chelation by DFP may provide glaucoma neuroprotection.
Although iron plays many crucial roles in cellular metabolism, excess iron can also confer toxicity by generating free radicals responsible for oxidative damage (Aisen et al., 2001; Beard, 2001; Song and Dunaief, 2013). In the eye, iron-induced oxidative stress can cause acute retinal degeneration and is also likely to play a role in the pathogenesis of age-related macular degeneration (AMD) (Hadziahmetovic et al., 2008; Hahn et al., 2003; Hahn et al., 2004). Deferiprone (DFP, Ferriprox), is an orally-administrated iron chelator approved by the FDA for treating patients with iron overload. Oral administration of DFP protects against iron-induced retinal degeneration by reducing retinal iron levels in ceruloplasmin/hephaestin double-knockout and hepcidin knockout mice, both of which exhibit age-related retinal iron accumulation and increased oxidative stress (Hadziahmetovic et al., 2011; Song et al., 2014). Interestingly, DFP administration also prevented retinal degeneration associated with sodium iodate treatment and light exposure in mice, both disease models in which oxidative stress plays a key role (Hadziahmetovic et al., 2012; Song et al., 2012).
Glaucoma is an important cause of irreversible blindness worldwide (Quigley and Broman, 2006). It is characterized by progressive vision loss and visual field defects secondary to retinal ganglion cell (RGC) degeneration and optic atrophy, often in the setting of elevated intraocular pressure (IOP). Oxidative stress has been implicated in glaucomatous optic neuropathies, and antioxidant administration has been shown to rescue rodent RGCs exposed to extended periods of IOP elevation (Benoist d’Azy et al., 2016; Ko et al., 2000; Yang et al., 2016). Further, iron-related genes transferrin, ceruloplasmin and ferritin were shown to be upregulated in a monkey model of glaucoma, and in glaucomatous human post mortem eyes, suggesting a role for iron-induced oxidative stress in glaucoma pathogenesis (Farkas et al., 2004). These associations suggest iron chelation may slow glaucoma progression.
The injection of microbeads into the anterior chamber for drainage angle occlusion has been used to good IOP-elevating effect in mice (Chen et al., 2011; Ito et al., 2016; Sappington et al., 2010). Sappington et al. demonstrated ~30% IOP elevation following a single injection of polystyrene microbeads (Sappington et al., 2010). RGC degeneration was shown to occur in a progressive manner, reaching 20% after 4–5 weeks. Ito et al. 2016 modified the technique by substituting polystyrene microbeads with smaller diameter magnetic beads, which allowed for the injection of a higher concentration of beads as well as targeting of the beads into the angle using magnets (Ito et al., 2016). Compared to laser and vascular cauterization models of induced ocular hypertension, microbeads induce minimal inflammation and allow for repeat treatments, which extend the duration of IOP elevation to 8 weeks (Bouhenni et al., 2012; Chen et al., 2011). The advantage of this technique over anterior chamber cannulation lies in the maintenance of elevated IOP over a prolonged duration (Lu et al., 2017). The purpose of this study is to examine whether oral DFP administration is protective against RGC and optic nerve degeneration in a microbead-induced mouse model of hypertensive glaucoma.
Twenty 3-month old C57BL/6J mice comprised of 10 males and 10 females were purchased from The Jackson Laboratory (Bar Harbor, ME). The animals were fed ad libitum and maintained on a 12 hour/12 hour light/dark cycle in the University of Pennsylvania vivarium. All procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Pennsylvania. Five males and five females were treated with the iron-chelator DFP (ApoPharma Inc., Toronto, Ontario, Canada) at a concentration of 1 mg/ml in the drinking water throughout the 8-week duration of the study. The other 10 mice received drinking water without DFP and served as controls. Animals in both groups drank approximately 5ml of water per day, equating to 5mg daily dose of DFP for the treatment group.
The Microbead Occlusion Model was used to induce ocular hypertension as previously described (Calkins et al., 2018; Ito et al., 2016). Briefly, all mice received injections of 1.5μL of magnetic microbeads (1.6 × 106 beads/μl of normal saline) in the anterior chamber of their left eye (OS) while the right eye (OD) received concurrent injections of sterile normal saline. Mice were injected on day 0 and again at day 16. IOPs were measured immediately prior to the first injection and then weekly using the Icare® TONOLAB tonometer (Icare TONOVET, Vantaa, Finland). An average of 3 measurements per eye was used.
RGC counting and immunolabeling of flat-mounted retinas were performed as previously described with minor modification noted below (Khan et al., 2017). Briefly, eyes were enucleated immediately after sacrifice and eyecups were created by removing the cornea, iris, and lens. The eyecup was bisected and the temporal half was fixed for 30 minutes in 4% paraformaldehyde. The temporal hemiretinas were then isolated and mounted on glass slides. Following serial washes with 0.5% Triton X-100 in PBS (TPBS) and 15-minute permeabilization through freeze/thaw at −80° C, retinas were incubated overnight at 4°C with anti-Brn3a (Synaptic Systems, Goettingen, Germany) diluted 1:1000 in a blocking buffer (2% bovine serum albumin and 2% Triton X-100 in PBS). The following day, retinas were washed in TPBS and incubated for 3 hours at RT with Alexa Fluor 488 secondary antibody (Invitrogen, Carlsband, CA) diluted 1:1000 in the blocking buffer. Finally, retinas were washed and cover-slipped with Vectashield mounting medium plus DAPI for fluorescence (Vector Laboratories Inc., Burlingame, CA). For each hemiretina, 6 standardized photomicrographs at 1/6, 3/6, and 5/6 distance from the center of the retina were taken in two quadrants at 40X magnification. A masked counter then quantified the number of Brn3a positive RGCs in each image (as shown in Fig. 2A) using Nikon Elements analysis software (Nikon Instruments, Melville, NY, USA).
Fig. 2.

Flat-mounted hemiretina sections were labeled for the RGC marker Brn3a (A). Six 40x images from each hemiretina were analyzed (bottom; white squares). RGCs were counted by a masked observer. Representative images are shown of the DFP-treated and control eyes (top). RGC density was preserved for the microbead-injected eyes of the DFP-treated mice despite elevated IOP (C, top). Error bars = Mean ± SEM; ANOVA with Tukey’s HSD **p < 0.01. The number of RGC axons was counted in optic nerve cross sections 1.5mm posterior to the globe (B). One central and 4 peripheral 75 × 75 μm images were sampled for each cross section (bottom; white squares). Higher magnification representative images (50 × 50 μm) are shown of the DFP-treated and control ONs (top). Axon swelling (arrow), gliosis (asterisk), and astrocyte activation (cross) were prominent in the microbead-injected eyes of the no DFP ONs. The AxonJ Image Analysis Algorithm plugin for ImageJ was used to count the number of axons. Axon density was preserved in the microbead-injected eyes of the DFP-treated mice despite elevated IOP (C, bottom). *p < 0.05.
Optic nerves were bisected at the level of the nerve head and fixed in 2.5% glutaraldehyde with 2.0% paraformaldehyde in 0.1M sodium cacodylate buffer, pH 7.4. RGC axons were labeled and counted as described previously (Ito et al., 2016). Briefly, optic nerves were incubated in 2% osmium tetroxide and dehydrated in graded ethanol immersions. Following embedding in epoxy resin Embed 812 (Electron Microscopy Sciences, Hatfield, PA), 0.75 μm thick cross sections were generated from a section of the nerve 1.5mm posterior to the globe and stained with 1% toluidine blue. For each optic nerve cross section, five standardized photomicrographs (75 × 75 μm) were obtained at 100X magnification; one in the center and each of the other four in its own quadrant half-way between the center and edge of the nerve. Axon counts were obtained by a masked operator using the AxonJ Image Analysis Algorithm plugin for ImageJ (http://imagej.nih.gov/ij/plugins/axonj/) (Zarei et al., 2016).
Mean ± standard error of the mean (SEM) was calculated for each group. Statistical analyses for RGC and axon counts were performed using two-way ANOVA followed by Tukey’s post-hoc pairwise comparisons. All statistical analyses were conducted using GraphPad Prism 6.0 (San Diego, CA). P < 0.05 was considered statistically significant.
After sequential injections 2 weeks apart, IOP remained elevated in the microbead-injected eyes for > 7 weeks (Fig. 1). The microbead injected eyes in the DFP group experienced higher IOPs compared to saline-injected eyes throughout the duration of the study, while the microbead injected eyes in the no DFP group experienced higher IOPs compared to saline-injected eyes for 7 weeks post injection. IOP differences between the microbead-injected eyes of the DFP versus no DFP mice were not significant at any point during the study.
Fig. 1.
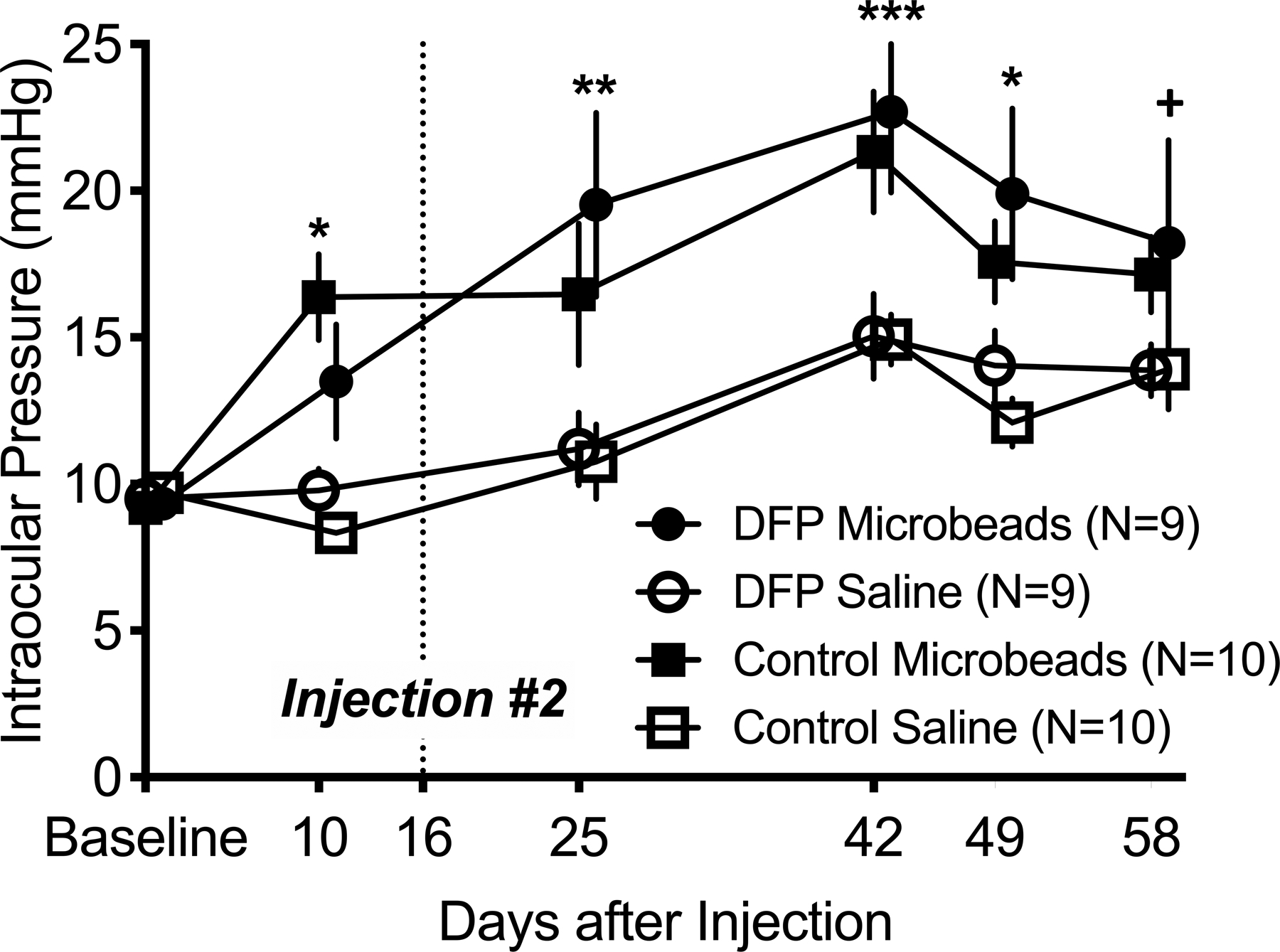
Microbeads were injected into the left eye while the right eye received sterile saline. Mice were injected on day 0 and again at day 16. IOP remained significantly elevated in the left eye for > 7 weeks. Error bars = Mean ± SEM; ANOVA with Tukey’s HSD *p < 0.05; **p < 0.01; ***p < 0.005; +p < 0.05 for the DFP group only. One mouse in the DFP group died and was excluded from the analysis.
To evaluate whether DFP treatment affected RGC survival in the setting of IOP elevation, we labeled hemiretina flat mounts with the RGC marker Brn3a and counted the number of positive RGCs at the end of the experiment (Fig. 2). The no DFP controls were found to have 20% fewer RGCs in the microbead-injected compared to the saline-injected eyes (93.6 ± 4.5 and 117.2 ± 4.7 per 40x HPF, respectively; p < 0.01). In comparison, mice that received oral DFP demonstrated no difference in RGC density between microbead-injected and saline-injected eyes (112.5 ± 5.6 and 118.3 ± 4.6 per 40x HPF, respectively). To evaluate whether DFP administration affected RGC axon survival, the number of axons in optic nerve cross sections was counted using the AxonJ Image Analysis Algorithm (Fig. 2). Without DFP treatment, the microbead-injected eyes demonstrated 13% fewer RGC axons compared to the saline-injected eyes (2543.3 ± 78.5 and 2916.1 ± 92.2 per 75 × 75μm, respectively; p < 0.05). With DFP treatment, in contrast, the number of RGC axons remained similar between the microbead and saline-injected eyes (2876.1 ± 101.6 and 2955 ± 74.6 per 75 × 75μm, respectively).
Overall, this study examined the effects of oral DFP administration on RGC survival in a mouse model of magnetic microbead-induced hypertensive glaucoma. Results showed that DFP improved RGC survival and preserved optic nerve axons during a 2-month period of persistently elevated IOPs.
The protective effects of iron chelators on neuro- and retinal degeneration have been demonstrated in various conditions associated with iron dysregulation. Compared to other iron chelators, DFP exhibits advantages such as ease of systemic administration in the oral form, an ability to readily cross the blood-brain barrier, and lack of an association with retinal toxicity (Cappellini and Pattoneri, 2009; Fredenburg et al., 1996). In the eye, DFP has been shown to decrease brain and retinal iron levels and protect against neuro- and retinal degeneration in ceruloplasmin/hephaestin double-knockout mice (Hadziahmetovic et al., 2011; Zhao et al., 2015). DFP was also protective against retinal degeneration in hepcidin knockout mice (Song et al., 2014).
Neurodegenerative conditions such as AMD, aceruloplasminemia, Friedreich’s ataxia, Alzheimer’s disease, and Parkinson’s disease have all been linked to iron excess and/or toxicity, thus supporting the exploration of iron chelators for treating certain types of neurodegenerative processes (Babady et al., 2007; Boddaert et al., 2007; Dunaief et al., 2005; Hahn et al., 2003; Lv et al., 2011; Richardson, 2004; Smith et al., 1997; Zheng et al., 2005). DFP has also been shown to be protective in ROS-associated models of retinal degeneration, and glaucoma is a group of neurodegenerative diseases affecting the RGC and the optic nerve where oxidative damage plays a pathogenic role. (Hadziahmetovic et al., 2012; Song et al., 2012; Ung et al., 2017). In particular, iron chelators zinc-deferoxamine and deferasirox have been shown to reduce oxidative stress and protect against excitoneurotoxicity following intravitreal NMDA injections, further supporting a role for iron-chelation as treatment for oxidative stress-associated neurodegeneration (Sakamoto et al., 2018).
While our study demonstrated 20% reduction in RGC loss following DFP treatment, reduction in loss of optic nerve axons was found to be closer to 13%. This difference may be related to our choice of Brn3a to label RGCs. While widely used, Brn3a has been shown to be an imperfect marker as it is only expressed in 80–90% of RGCs in mouse retinas and does not appear to label certain subpopulations of RGCs (Nadal-Nicolas et al., 2012; Rodriguez et al., 2014). Evidence also exists to suggest that Brn3a is down-regulated prior to RGC death resulting in decreased labeling of ailing RGCs (Nadal-Nicolas et al., 2012; Nadal-Nicolas et al., 2009). Both factors could have contributed to the difference we observed between RGC and axon survival.
In this study, we chose to use magnetic as opposed to polystyrene microbeads due to their ability to consistently elicit prolonged IOP elevation in our hands. These microbeads do contain iron and could conceivably induce retinal and/or RGC degeneration due to iron toxicity. However, retinal iron exposure was unlikely in this model as injections were carefully targeted into the anterior chamber. In addition, in our model, we did not observe any loss of photoreceptors, the retinal cell type most sensitive to iron toxicity (Rogers et al., 2007).
A downside of DFP treatment lies in its link to reversible agranulocytosis, which occurs in about 1% of patients with thalassemia who are treated with this drug, necessitating weekly blood draws for monitoring (Cappellini and Pattoneri, 2009; Cappellini and Piga, 2008). As such, the utility of DFP as long-term treatment for a slowly progressive neurodegenerative condition like glaucoma remains to be seen. Alternatives to systemic DFP administration may lie in local, targeted ocular administration and/or other iron chelators. The findings of our study can best be thought of as evidence of an association between iron chelation and neuroprotection in glaucoma and possibly as the genesis of additional experiments where iron chelation may be examined as treatment for both acute and chronic forms of glaucoma.
Highlights.
Oral DFP improved RGC survival in a microbead-induced mouse model of glaucoma.
DFP administration preserved retinal ganglion cells and optic nerve axons in hypertensive glaucomatous eyes.
Iron chelation may be neuroprotective in glaucoma.
Acknowledgements
This work was supported by R01EY015240; K12EY015398; Research to Prevent Blindness; the F.M. Kirby Foundation; A gift in memory of Lee F. Mauger, MD; The Paul and Evanina Bell Mackall Foundation Trust; American Glaucoma Society.
The authors would like to thank Dr. David Calkins’ laboratory for microbead injection training and Dr. Kenneth Shindler’s laboratory for training in retina flat-mount.
References
- Aisen P, Enns C, Wessling-Resnick M, 2001. Chemistry and biology of eukaryotic iron metabolism. Int J Biochem Cell Biol 33, 940–959. [DOI] [PubMed] [Google Scholar]
- Babady NE, Carelle N, Wells RD, Rouault TA, Hirano M, Lynch DR, Delatycki MB, Wilson RB, Isaya G, Puccio H, 2007. Advancements in the pathophysiology of Friedreich’s Ataxia and new prospects for treatments. Mol Genet Metab 92, 23–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beard JL, 2001. Iron biology in immune function, muscle metabolism and neuronal functioning. J Nutr 131, 568S–579S; discussion 580S. [DOI] [PubMed] [Google Scholar]
- Benoist d’Azy C, Pereira B, Chiambaretta F, Dutheil F, 2016. Oxidative and Anti-Oxidative Stress Markers in Chronic Glaucoma: A Systematic Review and Meta-Analysis. PLoS One 11, e0166915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boddaert N, Le Quan Sang KH, Rotig A, Leroy-Willig A, Gallet S, Brunelle F, Sidi D, Thalabard JC, Munnich A, Cabantchik ZI, 2007. Selective iron chelation in Friedreich ataxia: biologic and clinical implications. Blood 110, 401–408. [DOI] [PubMed] [Google Scholar]
- Bouhenni RA, Dunmire J, Sewell A, Edward DP, 2012. Animal models of glaucoma. J Biomed Biotechnol 2012, 692609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calkins DJ, Lambert WS, Formichella CR, McLaughlin WM, Sappington RM, 2018. The Microbead Occlusion Model of Ocular Hypertension in Mice. Methods Mol Biol 1695, 23–39. [DOI] [PubMed] [Google Scholar]
- Cappellini MD, Pattoneri P, 2009. Oral iron chelators. Annu Rev Med 60, 25–38. [DOI] [PubMed] [Google Scholar]
- Cappellini MD, Piga A, 2008. Current status in iron chelation in hemoglobinopathies. Curr Mol Med 8, 663–674. [DOI] [PubMed] [Google Scholar]
- Chen H, Wei X, Cho KS, Chen G, Sappington R, Calkins DJ, Chen DF, 2011. Optic neuropathy due to microbead-induced elevated intraocular pressure in the mouse. Invest Ophthalmol Vis Sci 52, 36–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunaief JL, Richa C, Franks EP, Schultze RL, Aleman TS, Schenck JF, Zimmerman EA, Brooks DG, 2005. Macular degeneration in a patient with aceruloplasminemia, a disease associated with retinal iron overload. Ophthalmology 112, 1062–1065. [DOI] [PubMed] [Google Scholar]
- Farkas RH, Chowers I, Hackam AS, Kageyama M, Nickells RW, Otteson DC, Duh EJ, Wang C, Valenta DF, Gunatilaka TL, Pease ME, Quigley HA, Zack DJ, 2004. Increased expression of iron-regulating genes in monkey and human glaucoma. Invest Ophthalmol Vis Sci 45, 1410–1417. [DOI] [PubMed] [Google Scholar]
- Fredenburg AM, Sethi RK, Allen DD, Yokel RA, 1996. The pharmacokinetics and blood-brain barrier permeation of the chelators 1,2 dimethly-, 1,2 diethyl-, and 1-[ethan-1’ol]-2-methyl-3-hydroxypyridin-4-one in the rat. Toxicology 108, 191–199. [DOI] [PubMed] [Google Scholar]
- Hadziahmetovic M, Dentchev T, Song Y, Haddad N, He X, Hahn P, Pratico D, Wen R, Harris ZL, Lambris JD, Beard J, Dunaief JL, 2008. Ceruloplasmin/hephaestin knockout mice model morphologic and molecular features of AMD. Invest Ophthalmol Vis Sci 49, 2728–2736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadziahmetovic M, Pajic M, Grieco S, Song Y, Song D, Li Y, Cwanger A, Iacovelli J, Chu S, Ying GS, Connelly J, Spino M, Dunaief JL, 2012. The Oral Iron Chelator Deferiprone Protects Against Retinal Degeneration Induced through Diverse Mechanisms. Transl Vis Sci Technol 1, 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadziahmetovic M, Song Y, Wolkow N, Iacovelli J, Grieco S, Lee J, Lyubarsky A, Pratico D, Connelly J, Spino M, Harris ZL, Dunaief JL, 2011. The oral iron chelator deferiprone protects against iron overload-induced retinal degeneration. Invest Ophthalmol Vis Sci 52, 959–968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn P, Milam AH, Dunaief JL, 2003. Maculas affected by age-related macular degeneration contain increased chelatable iron in the retinal pigment epithelium and Bruch’s membrane. Arch Ophthalmol 121, 1099–1105. [DOI] [PubMed] [Google Scholar]
- Hahn P, Qian Y, Dentchev T, Chen L, Beard J, Harris ZL, Dunaief JL, 2004. Disruption of ceruloplasmin and hephaestin in mice causes retinal iron overload and retinal degeneration with features of age-related macular degeneration. Proc Natl Acad Sci U S A 101, 13850–13855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito YA, Belforte N, Cueva Vargas JL, Di Polo A, 2016. A Magnetic Microbead Occlusion Model to Induce Ocular Hypertension-Dependent Glaucoma in Mice. J Vis Exp, e53731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan RS, Dine K, Bauman B, Lorentsen M, Lin L, Brown H, Hanson LR, Svitak AL, Wessel H, Brown L, Shindler KS, 2017. Intranasal Delivery of A Novel Amnion Cell Secretome Prevents Neuronal Damage and Preserves Function In A Mouse Multiple Sclerosis Model. Sci Rep 7, 41768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko ML, Hu DN, Ritch R, Sharma SC, 2000. The combined effect of brain-derived neurotrophic factor and a free radical scavenger in experimental glaucoma. Invest Ophthalmol Vis Sci 41, 2967–2971. [PubMed] [Google Scholar]
- Lu W, Albalawi F, Beckel JM, Lim JC, Laties AM, Mitchell CH, 2017. The P2X7 receptor links mechanical strain to cytokine IL-6 up-regulation and release in neurons and astrocytes. J Neurochem 141, 436–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lv Z, Jiang H, Xu H, Song N, Xie J, 2011. Increased iron levels correlate with the selective nigral dopaminergic neuron degeneration in Parkinson’s disease. J Neural Transm (Vienna) 118, 361–369. [DOI] [PubMed] [Google Scholar]
- Nadal-Nicolas FM, Jimenez-Lopez M, Salinas-Navarro M, Sobrado-Calvo P, Alburquerque-Bejar JJ, Vidal-Sanz M, Agudo-Barriuso M, 2012. Whole number, distribution and co-expression of brn3 transcription factors in retinal ganglion cells of adult albino and pigmented rats. PLoS One 7, e49830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadal-Nicolas FM, Jimenez-Lopez M, Sobrado-Calvo P, Nieto-Lopez L, Canovas-Martinez I, Salinas-Navarro M, Vidal-Sanz M, Agudo M, 2009. Brn3a as a marker of retinal ganglion cells: qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Invest Ophthalmol Vis Sci 50, 3860–3868. [DOI] [PubMed] [Google Scholar]
- Quigley HA, Broman AT, 2006. The number of people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol 90, 262–267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson DR, 2004. Novel chelators for central nervous system disorders that involve alterations in the metabolism of iron and other metal ions. Ann N Y Acad Sci 1012, 326–341. [DOI] [PubMed] [Google Scholar]
- Rodriguez AR, de Sevilla Muller LP, Brecha NC, 2014. The RNA binding protein RBPMS is a selective marker of ganglion cells in the mammalian retina. J Comp Neurol 522, 1411–1443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers BS, Symons RC, Komeima K, Shen J, Xiao W, Swaim ME, Gong YY, Kachi S, Campochiaro PA, 2007. Differential sensitivity of cones to iron-mediated oxidative damage. Invest Ophthalmol Vis Sci 48, 438–445. [DOI] [PubMed] [Google Scholar]
- Sakamoto K, Suzuki T, Takahashi K, Koguchi T, Hirayama T, Mori A, Nakahara T, Nagasawa H, Ishii K, 2018. Iron-chelating agents attenuate NMDA-Induced neuronal injury via reduction of oxidative stress in the rat retina. Exp Eye Res 171, 30–36. [DOI] [PubMed] [Google Scholar]
- Sappington RM, Carlson BJ, Crish SD, Calkins DJ, 2010. The microbead occlusion model: a paradigm for induced ocular hypertension in rats and mice. Invest Ophthalmol Vis Sci 51, 207–216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith MA, Harris PL, Sayre LM, Perry G, 1997. Iron accumulation in Alzheimer disease is a source of redox-generated free radicals. Proc Natl Acad Sci U S A 94, 9866–9868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song D, Dunaief JL, 2013. Retinal iron homeostasis in health and disease. Front Aging Neurosci 5, 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song D, Song Y, Hadziahmetovic M, Zhong Y, Dunaief JL, 2012. Systemic administration of the iron chelator deferiprone protects against light-induced photoreceptor degeneration in the mouse retina. Free Radic Biol Med 53, 64–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song D, Zhao L, Li Y, Hadziahmetovic M, Song Y, Connelly J, Spino M, Dunaief JL, 2014. The oral iron chelator deferiprone protects against systemic iron overload-induced retinal degeneration in hepcidin knockout mice. Invest Ophthalmol Vis Sci 55, 4525–4532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ung L, Pattamatta U, Carnt N, Wilkinson-Berka JL, Liew G, White AJR, 2017. Oxidative stress and reactive oxygen species: a review of their role in ocular disease. Clin Sci (Lond) 131, 2865–2883. [DOI] [PubMed] [Google Scholar]
- Yang X, Hondur G, Tezel G, 2016. Antioxidant Treatment Limits Neuroinflammation in Experimental Glaucoma. Invest Ophthalmol Vis Sci 57, 2344–2354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarei K, Scheetz TE, Christopher M, Miller K, Hedberg-Buenz A, Tandon A, Anderson MG, Fingert JH, Abramoff MD, 2016. Automated Axon Counting in Rodent Optic Nerve Sections with AxonJ. Sci Rep 6, 26559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao L, Hadziahmetovic M, Wang C, Xu X, Song Y, Jinnah HA, Wodzinska J, Iacovelli J, Wolkow N, Krajacic P, Weissberger AC, Connelly J, Spino M, Lee MK, Connor J, Giasson B, Harris ZL, Dunaief JL, 2015. Cp/Heph mutant mice have iron-induced neurodegeneration diminished by deferiprone. J Neurochem 135, 958–974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng H, Youdim MB, Weiner LM, Fridkin M, 2005. Novel potential neuroprotective agents with both iron chelating and amino acid-based derivatives targeting central nervous system neurons. Biochem Pharmacol 70, 1642–1652. [DOI] [PubMed] [Google Scholar]