

Abstract
Background:
Among urban residents, increased contacts with nature are associated with reduced morbidity and mortality. The concept of allostatic load, a biomarker-based composite measure of physiological dysregulation, can be applied to study subclinical benefits of exposure, and to elucidate pathways leading to improved health.
Objective:
This research explored associations between residential vegetated land cover and an allostatic load index calculated using the statistical distance measure known as Mahalanobis distance.
Methods:
This cross-sectional population-based study involved 186 adult residents of the Durham-Chapel Hill, North Carolina metropolitan area. Measures of tree and grass cover within 500 m of residence were derived from the U.S. Environmental Protection Agency’s EnviroAtlas land cover database. Fifteen biomarkers of immune, neuroendocrine, and metabolic functions were analyzed in serum samples. Regression analysis was conducted using generalized additive models with thin-plate spline functions of geographic coordinates, adjusting for modelled traffic air pollution from local sources and sociodemographic covariates.
Results:
The second and third tertiles of distance-weighted tree cover were associated with 14% (95% Confidence Limits 20%; 8%) and 15% (21%; 8%) reduction in adjusted median allostatic load, respectively, compared to the first tertile. The same tertiles of tree cover were also associated with 0.16 (0.03; 0.76) and 0.04 (0.01; 0.35) adjusted odds ratios of having allostatic load index above the 90th percentile of the sample distribution. Grass cover was inversely correlated with tree cover and was not associated with reduced allostatic load.
Conclusions:
Subclinical beneficial health effects of green spaces demonstrated in this study are consistent with reduced susceptibility to acute environmental and social stressors, and reduced risks of morbidity and mortality.
Keywords: Allostatic load, Vegetated land cover, Health effect biomarkers, Mahalanobis distance
1. Introduction
There is accumulating evidence that contacts with green and natural environments in urban settings are associated with various health benefits (WHO, 2016). These benefits include improved mental health (Alcock et al., 2014, Beyer et al., 2014, Gascon et al., 2015), reduced risks of diabetes (Astell-Burt et al., 2014), cardiovascular disease (Tamosiunas et al., 2014) and mortality (Gascon et al., 2016, James et al., 2016, Kondo et al., 2018). Enhanced mental restoration and alleviation of chronic stress, improved social cohesion, enhanced physical activity, improved immune system functions, and reduced air pollution are some of the hypothesized pathways to better health (Hartig et al., 2014, Kuo, 2015). While relative contributions of these different pathways to specific health outcomes likely depend on the type of urban green space and specific population subgroup (WHO, 2016), restoration and stress alleviation appear to be a major pathway leading to improved mental and physical health (Triguero-Mas et al., 2015, Dadvand et al., 2016).
Previous studies have demonstrated that short-term contacts with the natural environment produced enjoyment, relaxation and reduced stress resulting in short-term beneficial physiological changes and improved levels of certain biomarkers (Haluza et al., 2014, Aspinall et al., 2015). The main objective of our research was to investigate long-term subclinical health benefits of chronic exposure to the natural environment using allostatic load (AL), a biomarker-based measure of physiological dysregulation, as the outcome variable. AL reflects the physiological consequences of neural or endocrine responses resulting from repeated stress (McEwen and Stellar, 1993, Seeman et al., 1997, McEwen, 2002). Social and economic deprivation, and psychological traumas have been linked to increased AL (Clark et al., 2007, Bellingrath et al., 2009, Petrovic et al., 2016, Chandola and Zhang, 2018). Recent studies have also demonstrated inverse associations between residential greenness and allostatic load (Egorov et al., 2017), and between accessibility of urban green spaces and allostatic load (Ribeiro et al., 2019b). In turn, greater AL scores have been linked with adverse health outcomes including reduced cognitive performance, increased risks of cardiovascular diseases, other systemic diseases, and death (Borrell et al., 2010, Juster et al., 2010, Juster et al., 2011, Cohen et al., 2014, Hwang et al., 2014, Karlamangla et al., 2014, Milot et al., 2014, Cohen et al., 2015, McEwen, 2015, Arbeev et al., 2016, Arbeev et al., 2019).
Our previous research has demonstrated that greater vegetated land cover around residence as a marker of chronic exposure to the natural environment was associated with reduced AL indices based on multiple biomarkers of neuroendocrine, immune, cardiovascular, and metabolic functions (Egorov et al., 2017). That analysis utilized biomarkers that are known to be affected by chronic stress and to be predictive of adverse health outcomes. AL was calculated by summing binary biomarker data dichotomized at selected percentiles of sample distributions so that the AL index represented the number of biomarkers with potentially unhealthy values. Analysis of associations between vegetated land cover and individual biomarkers demonstrated that using an AL index was a statistically superior approach producing a much more significant and robust association than any single biomarker (Egorov et al., 2017).
The most common approach to estimating AL is the group AL index, which is based on the sum of biomarkers dichotomized at selected percentiles of a sample distribution (Juster et al., 2010). Variations of this approach involve dichotomizing biomarkers at specific percentiles of a population distribution or at clinical cut-off points when such data are available. A common approach to estimating AL using continuous biomarker data involves calculating a sum of z-scores or standard deviations from the sample mean for each biomarker (Juster et al., 2010). An alternative approach to estimating AL utilizing continuous biomarker data involves calculating statistical distance from the center of the joint distribution of biomarkers taking in account covariance among biomarkers, also known as the Mahalanobis distance(Cohen et al., 2013, Cohen et al., 2014).
The present manuscript describes results of further analysis of the above mentioned study in central North Carolina using a Mahalanobis distance-based AL index hereafter denoted as AL MAHA. In this new analysis, we also used improved exposure data involving separate estimates of grass cover and tree cover, and adjusted effect estimates for traffic air pollution from local sources.
2. Methods
2.1. Study design
This manuscript presents new results of analysis of data from a cross-sectional community study in adult residents of the Durham-Chapel Hill, North Carolina metropolitan area described in our previous publications (Egorov et al., 2017, Egorov et al., 2018). The protocol of this study was approved by the Institutional Review Board at the University of North Carolina in Chapel Hill. Study participants were recruited through advertisement in local newspapers and internet sites. All participants signed informed consent forms prior to data collection. Data collection was conducted during a single visit to the U.S. Environmental Protection Agency (US EPA) Human Studies Facility in Chapel Hill in May-September 2013. Venous blood samples for analysis of biomarkers were drawn by registered nurses. Serum samples were archived at −80 °C until analysis. Physical examinations were limited to measurements of height and weight. Participants completed a questionnaire on socio-demographic and behavioral factors. A total of 351 individuals participated in the parent study. Of these, 186 individuals who resided within the high-resolution urban land cover data area (see below) and had valid laboratory analysis results for all biomarkers were included in the present analysis.
2.2. Land cover data analysis
Residential addresses of study participants were geocoded at the building or property parcel centroid using ArcGIS software (Esri Inc., Redlands, CA, USA). Residential addresses that were not located at the building or property parcel centroid were located manually using a combination of address lookup, satellite imagery, and street-view to identify and confirm the center of the residence. The residential vegetated land cover data for the year 2010 were derived from 1 m resolution urban land cover imagery for the Durham-Chapel Hill, NC area, obtained from the US EPA EnviroAtlas database (https://www.epa.gov/enviroatlas) (Pickard et al., 2015). The high-resolution land cover data included five categories: (i) Water, (ii) Impervious surface, (iii) Soil & barren, (iv) Trees & forest, and (v) Grass & other herbaceous. While our previous analyses (Egorov et al., 2017, Egorov et al., 2018) used total vegetated land cover (trees and grass combined) to assess exposure, the present analysis used separate measures of tree cover and grass cover, as well as total vegetated land cover. Land cover measures were produced for a 500 m radius around each geocoded residence by ten concentric 50 m-wide annuli, from 0-50 m to 450-500 m. Exposure data were produced using four land cover estimation schemes for each land cover measure:
Average proportion of land cover within 150 m of residence
Average proportion of land cover within 500 m of residence
Weighted proportion of land cover within 500 m using equal weights assigned to all ten 50 m annuli
Weighted proportion of land cover within 500 m with annulus-specific weights exponentially declining with distance from the residence.
In the Scheme 4, annulus-specific weights were derived from a cumulative probability function of an exponential distribution with the parameter λ set equal to 0.0025 as described previously (Egorov et al., 2017). Weight for each annulus was calculated as an interval cumulative density:
where x1 is the inner radius and x2 is the outer radius of each annulus in meters.
To assess the degree of urbanicity, census block-group level data on average number of housing units per acre were abstracted from EPA’s Smart Location Database (https://www.epa.gov/smartgrowth/smart-location-mapping#SLD). Housing unit density data were square root transformed and used as a covariate in regression analysis.
2.3. Health effect biomarkers
The present analysis used 15 serological biomarkers that were available for most study participants as described previously (Egorov et al., 2017). These biomarkers included nine biomarkers of immune function: C-reactive protein (CRP), fibrinogen, interleukin (IL)-1β, IL-6, IL-8, intercellular adhesion molecule 1 (ICAM-1), myeloperoxidase (MPO), tumor necrosis factor (TNF)-α, and vascular cell adhesion molecule 1 (VCAM-1); four biomarkers of neuroendocrine function: dehydroepiandrosterone (DHEA), dopamine, epinephrine, and norepinephrine; and two biomarkers of metabolic function: serum amyloid A (SAA) and uric acid. VCAM-1 and ICAM-1 can also be characterized as biomarkers of endothelial function or vascular injury. All biomarkers were analyzed at EPA laboratories using commercially available assay kits as described previously (Egorov et al., 2017). Concentrations of analytes were estimated in accordance with kit manufacturers’ instructions. Results below the limits of detection (LOD) for dopamine, norepinephrine, MPO, IL-1β, IL-6, and TNF-α were substituted with ½ LOD values. Results for other biomarkers were all above LOD.
2.4. Statistical data analysis
Statistical analysis was conducted using SAS version 9.4 (SAS Institute, Cary, NC) software. Biomarker data were log-transformed for statistical analysis. AL MAHA measures were estimated using the mahalanobis function in the SAS procedure IML using sample-derived mean and covariance values. This function uses the following formula:
where x is the vector of observations for k biomarkers, µ is the vector of mean biomarker values, and S is the covariance matrix.
Regression analysis of associations between vegetated land cover and AL MAHA was conducted using generalized additive models with a two-dimensional spline function (“thin-plate spline”) of geographic coordinates to account for spatial trends (Webster et al., 2006, Ribeiro et al., 2019a). Regression models were fitted using the SAS procedure gampl. Exposure data (tree cover, grass cover or total vegetated land cover) were categorized into tertiles based on the sample distributions. All effect estimates are presented for tertiles 2 and 3 vs. the reference tertile 1.
Two types of regression models were used:
Model 1: Linear regression model for AL MAHA data. As AL MAHA values were approximately log-normally distributed, the outcome was log-transformed for regression analysis. The results are presented as multiplicative changes in median AL MAHA levels.
Model 2: Regression models for binomial outcome using AL MAHA data dichotomized at the 90th percentile of the sample distribution. This analysis was conducted to assess the effect of tree cover on odds of having high AL MAHA level.
Missing data on sociodemographic and behavioral covariates were imputed using the SAS procedure mi for arbitrary missing data applying the multivariate discriminant function method. The initial set of covariates for regression analysis was based on the final model for allostatic load developed previously (Egorov et al., 2017). It included age, gender, race (dichotomized as non-Hispanic Whites vs. all others), education (dichotomized as bachelor’s degree and higher vs. less than bachelor’s degree), BMI, square root transformed housing unit density at the census block group level, and two-dimensional spline of geographic coordinates.
This analysis also involved adjusting for ambient levels of traffic air pollution from local sources. Concentrations of nitrogen oxides (NOx), a representative traffic pollutant were estimated using the C-LINE/C-PORT online dispersion modeling tool for traffic and point sources of emission (Barzyk et al., 2015, Isakov et al., 2017). Model inputs included road network and traffic intensity data from North Carolina Department of Transportation. Annual average concentrations of NOx at residences of study participants were estimated taking in account meteorological data incorporated in the modeling tool.
A final multivariate regression model was developed from this initial model by adding other sociodemographic variables one at a time and removing the least significant variable one at a time (bidirectional elimination). Alternative regression models were compared using the Akaike Information Criterion Corrected (AICc) model fit statistics. Age was included in all models because it a biological factor affecting most biomarkers of health. Housing density, annual average concentration of NOx, and two-dimensional spline of geographic coordinates (“thin plate spline”) were also included in all regression models regardless of their statistical significance in order to demonstrate that effect estimates for land cover variables were adjusted for potential spatially-varying confounders, an important concern in research on green space and health.
To assess the robustness of the effects detected in the full dataset and identify potential effect modifiers, further analysis was conducted using the dataset stratified by binary factors: housing unit density (dichotomized at 1.5 housing units per acre or approximately 3.7 housing units per hectare distribution based cut-off), sex, dichotomized race (White vs. all others), and obesity (obese individuals vs. all others). Due to sample size limitations, only continuous AL MAHA data (Model 1) were used in the stratified analysis. When warranted, potential effect modification was further explored in the full dataset using models with interaction terms.
3. Results
3.1. Descriptive statistics
Age of 186 study participants ranged from 18 to 85 years (mean 37.1 years). Approximately two-thirds (67.2%) of participants were women (Table 1). Just over half of study participants (52.2%) had at least a four-year college degree. This relatively high level of education reflected the composition of source communities. BMI data indicated that 28.5% of study participants were obese according to the World Health Organization’s definition (BMI ≥ 30.0 kg/m2). Current smoking was reported by 26.3% of study participants.
Table 1.
Demographic, socioeconomic and health characteristics of the study population.
| Variable | Category | Number | Percent |
|---|---|---|---|
| All | 186 | 100% | |
| Sex | Female | 125 | 67.2% |
| Male | 61 | 32.8% | |
| Race | White | 106 | 57.0% |
| African American | 66 | 35.5% | |
| American Indian or Alaska Native | 1 | 0.5% | |
| Asian or Pacific Islander | 4 | 2.2% | |
| Other or mixed | 8 | 4.3% | |
| Not reported | 1 | 0.5% | |
| Ethnicity | Hispanic | 13 | 7.0% |
| Not Hispanic | 173 | 93.0% | |
| Education | Did not graduate from high school | 13 | 7.0% |
| High school | 29 | 15.6% | |
| Some college, no degree | 34 | 18.3% | |
| Associate degree | 13 | 7.0% | |
| Bachelor’s degree | 55 | 29.6% | |
| Post baccalaureate degree | 42 | 22.6% | |
| Smoking status | Current smoker | 49 | 26.3% |
| Not a current smoker | 136 | 73.1% | |
| Not reported | 1 | 0.5% | |
| BMI category | Obese | 53 | 28.5% |
| Overweight | 50 | 26.0% | |
| Normal | 80 | 43.0% | |
| Underweight | 3 | 1.6% |
Approximately half of study participants (50%) lived in the city of Durham, 31% lived in Chapel Hill and 19% lived in smaller towns in the Durham-Chapel Hill, NC metropolitan area. Predominant types of housing in the study population were detached single family homes (the most prevalent housing type), townhouses and low-rise apartment buildings.
Housing unit density at a census block-group level ranged from 0.2 to 20.0 (median 3.8) housing units per hectare. Descriptive statistics for land cover measures are presented in Table 2. Total average vegetated land cover within 500 m of residence (land cover estimation Scheme 2) ranged from 37% to 95%. Weighted land cover measures, which assigned greater weights to vegetation near residence (Schemes 3 and 4), as well as average vegetated land cover within 150 m (Scheme 1), exhibited greater variability than the average within 500 m. This reflected the greater variability in land cover near residences. At most residences, vegetated land cover was represented mainly by tree cover.
Table 2.
Vegetated land cover around residences, percentage of total area.
| Land cover estimation scheme | Land cover type | Min | 33st pctl | Median | 67th pctl | Max |
|---|---|---|---|---|---|---|
| 1: Average within 150 m | Vegetation | 20 | 59 | 68 | 74 | 95 |
| Trees | 7 | 41 | 49 | 60 | 92 | |
| Grass | 1 | 12 | 14 | 18 | 45 | |
| 2: Average within 500 m | Vegetation | 37 | 66 | 73 | 77 | 95 |
| Trees | 24 | 51 | 57 | 64 | 93 | |
| Grass | 3 | 12 | 14 | 16 | 31 | |
| 3: Weighted average within 500 m with equal weights | Vegetation | 32 | 64 | 69 | 76 | 94 |
| Trees | 21 | 48 | 53 | 60 | 91 | |
| Grass | 3 | 12 | 14 | 17 | 34 | |
| 4: Weighted average within 500 m with exponential decay | Vegetation | 27 | 62 | 68 | 76 | 94 |
| Trees | 17 | 45 | 52 | 59 | 90 | |
| Grass | 3 | 12 | 15 | 17 | 36 |
Tree cover measures were negatively correlated with grass cover measures, with correlation coefficients ranging from −0.46 to −0.58 depending on the land cover estimation scheme (Supplemental Table 1). Total vegetated land cover measures were positively correlated with tree cover (correlation coefficients from 0.75 to 0.94) but weakly negatively correlated with grass cover (correlation coefficients from −0.17 to −0.27). A scatter plot of exponentially weighted grass cover vs. tree cover within 500 m of residence (Scheme 4) is presented in Figure 1.
Fig. 1.
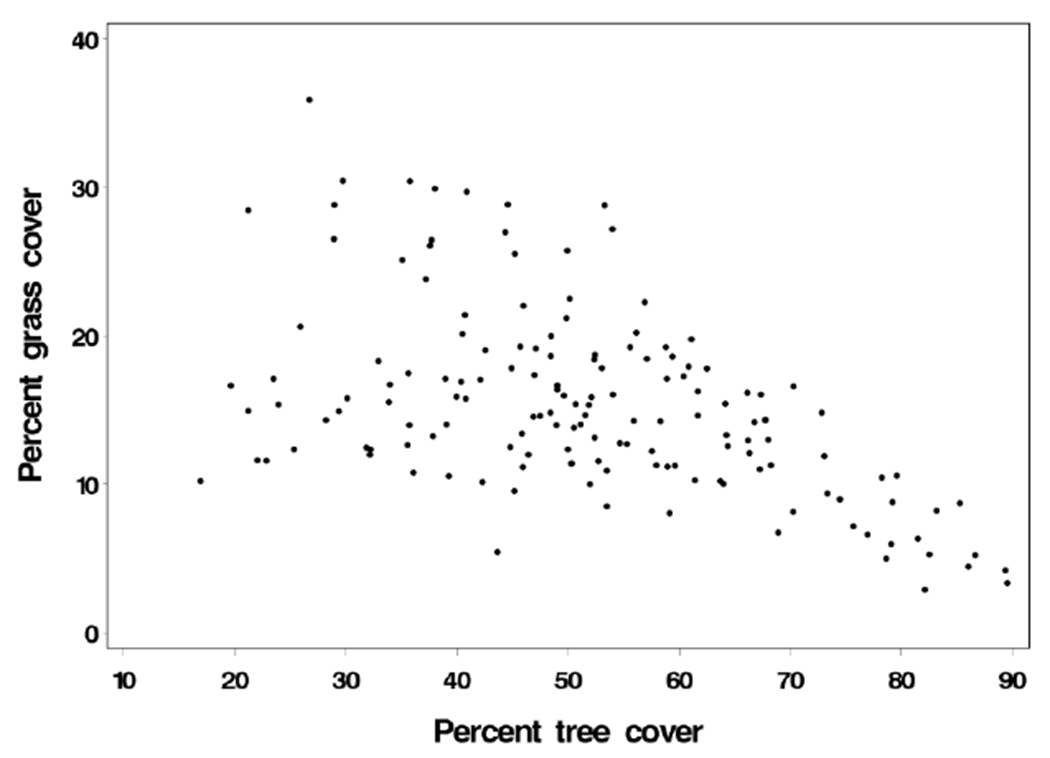
Percent of tree cover vs. percent of grass cover within 500 m of residence, distance-weighted estimates with weights from an exponential decay function (Scheme 4).
Estimated annual average concentrations of NOx from local traffic sources at residences of study participants ranged from 0.5 to 79.5 parts per billion (ppb) with mean 12.4 ppb and interquartile range 10.7 ppb. Average annual background concentration estimated by the modeling software (3.1 ppb) was not included in these estimates. Estimated NOx levels were positively correlated with residential unit density (r = 0.28, p = 0.0001) and weakly negatively correlated with various vegetated land cover measures. The strongest negative correlation was observed for average total vegetated land cover within 500 m of residences (r = −0.19, p = 0.01). For exponentially weighted tree cover within 500 m of residence (land cover estimation Scheme 4), the negative correlation was very weak and not significant (r = −0.07, p = 0.3).
The values of AL MAHA index ranged from 2.0 to 7.2 (median 3.7). Log-transformed approximately normally distributed AL MAHA data were used in further analyses.
3.2. Associations between residential vegetated land cover and allostatic load
In addition to a measure of residential greenness, the final set of covariates for regression analysis included variables for age, education (dichotomized as bachelor’s degree or higher vs. less than bachelor’s degree), smoking status (dichotomized as current smokers vs. non-smokers), BMI (log-transformed), housing unit density (square root transformed), annual average NOx level from local traffic sources, and a two-dimensional spline function of geographic coordinates. Race, ethnicity and sex were not included in regression models as these variables were not significant predictors of AL MAHA; removing them did not result in meaningful changes in parameter estimates for residential greenness variables. Although NOx concentration was positively associated with AL MAHA, its effect was not statistically significant in any model. For example, in the model that included exponentially weighted tree cover data (Scheme 4) as the predictor of interest, an increase of 10 ppb (approximately equal to the interquartile range) in annual average NOx level was associated with 1.01 (0.99; 1.04) multiplicative change in median AL MAHA (p = 0.3), or about 1% increase.
In order to select an optimal land cover estimation scheme, exposure measures produced under the four schemes described above were analyzed using Model 1 for continuous AL MAHA data and Model 2 for dichotomized AL MAHA data. Results were expressed in terms of adjusted multiplicative changes in median AL MAHA for Model 1 and adjusted odds ratios of having AL MAHA above the 90th percentile of the sample distribution for Model 2.
Greater tree cover was consistently associated with reduced median AL MAHA and reduced odds of having AL MAHA above the 90th percentile (Table 3). For both models, Scheme 2 (simple average within 500 m) consistently produced the poorest model fit with the biggest AICc values and the smallest effect estimates while Scheme 4 (exponentially weighted mean within 500 m) produced the best model fit with smallest AICc values and the greatest adjusted effect estimates. In Model 1 for Scheme 4, the 3rd tertile of tree cover compared to the 1st tertile was associated with a multiplicative change of 0.85 (0.79, 0.92), p <0.0001 in AL MAHA. In other words, the median adjusted AL MAHA was reduced by 15 % (21%; 8%). In Model 2 for Scheme 4, the adjusted odds ratio of having AL MAHA above the 90th percentile of the sample distribution was 0.04 (0.01; 0.35) - an approximately 25-fold reduction in the odds of having a high level of physiological dysregulation.
Table 3.
Adjusted estimates of the effects of tree cover on AL MAHA by alternative land cover estimation schemes. Model 1: Multiplicative changes in continuous log-transformed AL data; Model 2: Odds ratios for AL above the 90th percentile of the sample distribution. Effect estimates were adjusted for age, smoking status, education, BMI, density of residential units per census block groups, concentration of NOx from local traffic, and thin plate spline of geographic coordinates.
| Tree cover estimation scheme | Tertile of tree cover | Model 1: multiplicative change in continuous AL MAHA | AICc for model 1a | Model 2: aOR of AL MAHA above 90th pctl | AICc for model 2a |
|---|---|---|---|---|---|
| 1: Average within 150 m | 1st | Reference | −338 | Reference | 125 |
| 2nd | 0.92 (0.85, 0.99)* | 0.29 (0.08, 1.07) | |||
| 3rd | 0.87 (0.81, 0.95)** | 0.11 (0.02, 0.59)* | |||
| 2: Average within 500 m | 1st | Reference | −335 | Reference | 128 |
| 2nd | 0.95 (0.88, 1.02) | 0.59 (0.18, 1.90) | |||
| 3rd | 0.89 (0.82, 0.97)* | 0.14 (0.02, 0.76)* | |||
| 3: Average weighted with 500 m with equal weights | 1st | Reference | −345 | Reference | 121 |
| 2nd | 0.86 (0.80, 0.93)** | 0.15 (0.03, 0.73)* | |||
| 3rd | 0.87 (0.80, 0.94)** | 0.06 (0.01, 0.39)* | |||
| 4: Average weighted within 500 m with exponential decay | 1st | Reference | −348 | Reference | 120 |
| 2nd | 0.86 (0.80, 0.92)*** | 0.16 (0.03, 0.76)* | |||
| 3rd | 0.85 (0.79, 0.92)*** | 0.04 (0.01, 0.35)* |
p < 0.05
p < 0.001
p < 0.0001
Lower values indicate better model fit.
Further analyses comparing various exposure characterization approaches based on different land cover measures were conducted using Scheme 4 only. Both total vegetated land cover (exposure characterization approach A) and tree cover (exposure characterization approach B) were significantly associated with reduced AL MAHA (Table 4). However, both regression Models 1 and 2 for tree cover produced better fit, and stronger effect estimates with smaller p-values than corresponding models for total vegetated land cover. Grass cover, when analyzed without adjusting for tree cover (exposure characterization approach C), appeared to have a detrimental effect on AL MAHA. This effect was apparently due to confounding by tree cover which was negatively correlated with grass cover. When grass cover and tree cover variables were included in the same regression model (exposure characterization approach D), the effect of grass cover on AL MAHA was no longer significant. In contrast, adjusting for grass cover did not result in meaningful changes in effect estimates for tree cover.
Table 4.
The estimated effects of land cover types on AL. Model 1: Multiplicative changes in continuous log-transformed AL MAHA data; Model 2: Odds ratios for AL above the 90th percentile of the sample distribution. Effect estimates were adjusted for age, smoking status, education, BMI, density of residential units per census block group, concentration of NOx from local traffic, and thin plate spline of geographic coordinates.
| Exposure characterization approach | Type of land cover | Tertile of land cover | Model 1: adjusted multiplicative change in continuous AL MAHA | AICc for model 1 | Model 2: aOR of AL MAHA above 90th percentile | AICc for model 2 |
|---|---|---|---|---|---|---|
| A | Total vegetated cover | 1st | Reference | −333 | Reference | 129 |
| 2nd | 0.94 (0.87, 1.02) | 0.32 (0.09, 1.20) | ||||
| 3rd | 0.91 (0.84, 0.99)* | 0.13 (0.02, 0.78)* | ||||
| B | Tree cover | 1st | Reference | −348 | Reference | 120 |
| 2nd | 0.86 (0.80, 0.92)*** | 0.16 (0.03, 0.76)* | ||||
| 3rd | 0.85 (0.79, 0.92)*** | 0.04 (0.01, 0.35)* | ||||
| C | Grass cover | 1st | Reference | −334 | Reference | 133 |
| 2nd | 1.00 (0.96, 1.12) | 2.06 (0.55, 7.72) | ||||
| 3rd | 1.10 (1.02, 1.19) | 1.69 (0.43, 6.61) | ||||
| D | Tree cover | 1st | Reference | −338 | Reference | 130 |
| 2nd | 0.85 (0.79, 0.92)*** | 0.20 (0.05, 0.82)* | ||||
| 3rd | 0.86 (0.78, 0.93)** | 0.07 (0.01, 0.51)* | ||||
| Grass cover | 1st | Reference | Reference | |||
| 2nd | 1.00 (0.93, 1.09) | 0.69 (0.15, 3.18) | ||||
| 3rd | 1.04 (0.96, 1.14) | 0.78 (0.17, 3.62) |
p < 0.05
p < 0.001
p < 0.0001
Regression analysis stratified by four binary factors was conducted using the same regression models as described above, except analysis stratified by obesity excluded BMI as a predictor. The results (Figure 2) show that the inverse associations between tree cover and AL MAHA were consistent in all strata. Specifically, the estimated adjusted effect of the 3rd tertile of tree cover was statistically significant in seven out of eight stratified tests: in low housing density (N = 92) and high housing density (N = 94) strata, in males and females, in obese and non-obese individuals, as well as in non-Hispanic Whites (N = 100), but not in non-White or Hispanic individuals (N = 86). For example, estimated multiplicative effect on AL MAHA for the 3rd tertile of tree cover vs. 1st tertile were 0.86 (0.76; 0.97) in men and 0.84 (0.77; 0.93) in women. Further regression analysis of an interaction effect of dichotomized race/ethnicity and tree cover did not produce evidence of significant modification (p > 0.5, data not shown). Overall, stratified analyses produced no evidence of effect modification by housing unit density, sex, race, or obesity.
Figure 2.
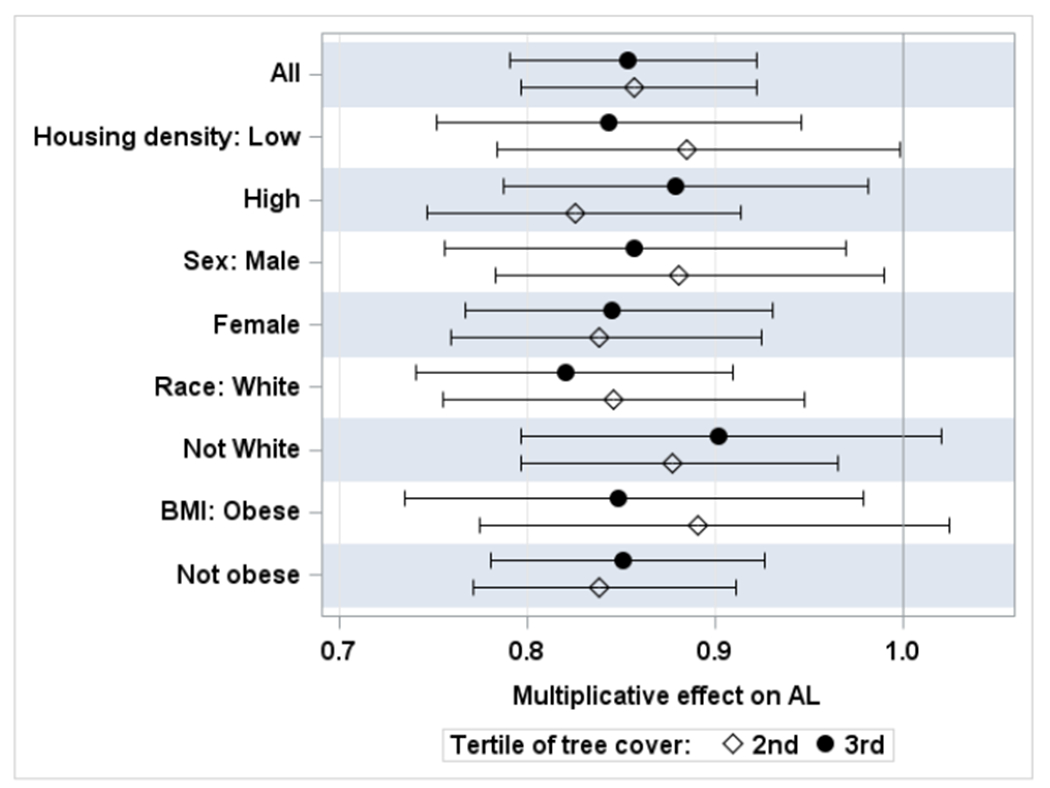
Results of stratified analysis of continuous AL MAHA data (Model 1) vs. tertiles of tree cover (Scheme 4) using the 1st tertile as a reference category.
4. Discussion
4.1. Summary of findings and interpretation
This research applied a composite biomarker-based measure of physiological dysregulation known as allostatic load to demonstrate subclinical health benefits of residential tree cover and total vegetated land cover. The present analysis employed the Mahalanobis distance approach to estimate an allostatic load index denoted in this manuscript as AL MAHA. This approach utilizes continuous biomarker data and takes in account covariance. The results demonstrated a strong beneficial association between greater distance-weighted proportion of tree cover within 500 m of residence and reduced AL MAHA in adult residents of central North Carolina.
These new results corroborate our previous analyses which demonstrated the effects of total residential vegetated land cover on the AL index calculated using a different approach – a sum of dichotomized biomarker values reflecting the count of biomarkers with potentially unhealthy values (Egorov et al., 2017, Egorov et al., 2018).
The main findings showing beneficial effects of tree cover on AL MAHA were highly significant (p < 0.0001) despite the rather small study population of 186 adults. The observed effects were also consistent in regression analyses stratified by various sociodemographic and geographic factors, such as gender, race, obesity status, and housing unit density, suggesting that subclinical health benefits of urban greenery were similarly expressed in various sub-populations, and in urban and suburban areas.
Previous prospective cohort studies using AL MAHA indices comprised of various sets of neuroendocrine, metabolic and immune biomarkers demonstrated that greater AL MAHA was strongly associated with increased risks of cardiovascular and other systemic diseases and mortality, and that these associations were robust to changes in the composition of biomarkers (Cohen et al., 2014, Milot et al., 2014, Cohen et al., 2015, Arbeev et al., 2016, Arbeev et al., 2019).
The results of our study suggest that a mechanism linking exposure to urban green spaces with reduced morbidity and mortality effects demonstrated in previous studies (van den Bosch and Ode Sang, 2017, Kondo et al., 2018) involves a reduced stress-related physiological dysregulation. Our previous analysis also demonstrated significant beneficial effects of residential greenery on individual biomarkers of immune function and inflammation (Egorov et al., 2017). Chronic stress is known to be associated with increased chronic inflammation while chronic inflammation plays an important role in the etiology of cardiovascular and other systemic diseases (Wirtz and von Kanel, 2017, Furman et al., 2019). Alleviating chronic stress leads to reduced chronic inflammation and physiological dysregulation, and reduced susceptibility to systemic diseases.
This analysis involved adjusting effect estimates for vegetated land cover by estimated annual average exposure to NOx from local traffic sources. We modeled only NOx air pollution from local sources because background levels of nitrogen oxides change slowly over distance. Gradual spatial effects on the outcome would be accounted for by the two-dimensional spline function of geographic coordinates that was included in all regression models.
As developed urban areas with higher levels of traffic air pollution tend to have less vegetation, air pollution could be a positive confounder inflating apparent beneficial effects of tree cover on AL. However, the results of this analysis show that traffic air pollution did not confound the observed beneficial effects of tree cover in the study area. This finding may not be generalizable to other urban areas, especially those with higher air pollution levels or with stronger negative associations between air pollution and greenness.
Reduction of air pollution exposure by urban greenery, such as green belts along roads providing barriers to the spread of air pollution to near-road residences, can potentially mediate health benefits of green spaces. It has been shown, however, that only properly designed, dense, uninterrupted green belts or barriers along roads can reduce behind-barrier levels of traffic air pollution (Abhijith et al., 2017, Baldauf, 2017). This may explain why some studies failed to show a reduction in traffic air pollution behind green belts (Yli-Pelkonen et al., 2017). As C-LINE/C-PORT models do not take in account vegetation, we conducted additional exploratory analysis using Google street view (not shown in the Results section), and found that very few of the study participants lived in residences behind dense green barriers that could substantially reduce exposure to traffic air pollution from local sources.
4.2. Limitations
This cross-sectional observational study had certain limitations. Its findings could be affected by unmeasured confounding factors, such as health-oriented behaviors that might be correlated with living in greener areas. While the study involved collecting standard individual level socioeconomic and demographic data, behavioral data were limited to smoking and handling soil with bare hands – a risk factor for soil-borne infections (Egorov et al., 2018). Data on physical activity, spending time outdoors, gardening, and dietary habits were not collected. Further research on green space and physiological dysregulation should involve collecting more extensive behavioral data to address these concerns.
This study used a convenience sampling approach. Women were overrepresented relative to the general population due to their greater willingness to participate in the study. The level of education and racial composition were generally representative of the source population as discussed previously (Egorov et al., 2017). As this study involved adults only, these findings are not applicable to children. The findings may not be generalizable beyond the study area in central North Carolina. This research will need to be replicated in areas with different types of vegetation, climate and land use practices, and in populations of different compositions.
This study aimed to assess beneficial subclinical effects of greenery in the vicinity of residences. Assessing potential effects of city parks, forests, green corridors, and other public green spaces was beyond the scope of this study. While potentially increased physical activity in residents of greener areas and greater benefits of physical activity in the natural environment (“green exercise”) could also be important pathways to health, this study was not designed to assess their benefits. Our previous analysis of the same dataset produced no evidence of an association between residential vegetated land cover and BMI (Egorov et al., 2017). This finding supports the hypothesis that for the greenery around residences, relaxation, stress alleviation and improved immune system functioning are the most important pathways to improved health.
4.3. Strengths
The statistical method for estimating AL MAHA has several apparent advantages (Cohen et al., 2013). Compared to the AL estimation method based on dichotomized biomarkers, AL MAHA uses greater amount of the available information on individual biomarkers and their interrelationships. AL MAHA is also an improvement compared to another commonly used approach to estimating AL involving a sum of z-scores for individual biomarkers, which does not account for covariance (Ribeiro et al., 2019b).
A potential disadvantage of the AL MAHA method (as well as the AL estimation approach based on the sum of z scores) is that the statistical center of the sample distribution may not represent individuals in optimal health. Typical study participants may have a substantial amount of physiological dysregulation. As a result, a perfectly healthy, non-stressed individual may have a substantial estimated AL MAHA value. To assess the magnitude of this potential problem, we re-analyzed data using an alternative AL MAHA index which was calculated using the means of log-transformed biomarker values derived from a subset of 67 healthiest study participants (36% of the study population). This subset excluded individuals with self-reported diabetes, arthritis, heart, kidney and liver diseases, ulcers, dyspepsia, inflammatory bowel disease, HIV infection, history of depression, and self-reported health status worse than “very good”, as well as individuals with positive results of serological IgG test for Toxoplasma gondii, which was linked with increased AL in our previous analysis (Egorov et al., 2018). Pearson correlation between the two log-transformed alternative AL MAHA indices was 0.94. The results of regression analysis using the alternative AL MAHA index were within 2 % of the results reported above. For example, the multiplicative effect estimates for the 3rd tertile vs. 1st tertile of distance-weighted average tree cover changed from 0.85 (0.79; 0.92) to 0.84 (0.78; 0.91). This suggests that the study findings were not affected by a high proportion of individuals with chronic diseases in the study sample.
Associations between residential vegetation and AL MAHA were analyzed using two alternative models: (1) linear regression for log-transformed AL MAHA to assess multiplicative effects of tree cover on median AL MAHA; and (2) regression models using dichotomized AL MAHA data (a binomial outcome) to assess the effect of tree cover on the odds of having AL MAHA above the 90th percentile of the sample distribution. The results consistently demonstrated significant associations suggesting beneficial subclinical effects of exposure to greenery. The third tertile of tree cover compared to the first tertile was associated with an approximately 15% reduction in median AL MAHA (or approximately 0.7 standard deviations reduction in log-transformed AL MAHA) and approximately 25-fold smaller odds of having AL MAHA above the 90th percentile of the sample distribution. The apparent discrepancy between the relatively small effect on the median AL MAHA and the pronounced effect on the odds of having high AL MAHA could be due to a combination of two mechanisms: (i) the entire distribution of AL MAHA values being shifted to the left in individuals exposed to greater residential tree cover; and (ii) stronger beneficial effects in highly stressed individuals on the right tail of the AL MAHA distribution. Future research using larger sample sizes should attempt to characterize effects of urban green spaces in highly stressed individuals.
The use of 1-meter resolution land cover data from US EPA’s EnviroAtlas enabled detailed analysis of vegetated land cover around each residence using concentric annuli. Accounting for distance-dependent decline in the effect of greenery was a strength of this analysis. Another strength was a regression modeling approach that involved fitting a two-dimensional spline function of geographic coordinates to adjust for potential confounding by unknown geographically-varying factors associated with greenness and health.
In our previous analysis, an interquartile increase in total exponentially-weighted vegetated land cover was associated with an approximately 37% reduction in the mean Poisson-distributed AL, which was calculated using dichotomized data on the same 15 biomarkers (Egorov et al., 2017). Re-analysis of the same Poisson-distributed AL index against newly generated tree cover data produced a comparable effect estimate: 57% (43%, 68%) reduction in AL at the third tertile of tree cover compared to the first tertile (not shown in the Results section). The corresponding effect estimate on AL MAHA was a relatively modest 15% (21%, 8%) reduction in the median AL MAHA value (Model 1, exposure characterization approach B in Table 4). It should be noted that the effects of greenery on AL index based on dichotomized biomarkers and AL MAHA based on continuous biomarkers are not directly comparable. For the former, the effect is a percent reduction in the count of biomarkers with potentially unhealthy levels; for the latter, the effect means a percent reduction in the statistical distance from the center of a multivariate distribution. The results of these two analyses support each other and demonstrate that the beneficial effects of residential tree cover are robust to variations in statistical approaches to estimating AL.
In this analysis the distance-weighted measure of tree cover within 500 m of residence with weights derived from an exponential distribution provided the best model fit and the biggest effect estimates. This finding is consistent with our previous analysis (Egorov et al., 2017). While the 500 m buffer size is common in epidemiological research on residential greenness, previous studies have used various buffer sizes from 100 m to 2 km (Dadvand et al., 2015, Browning and Lee, 2017, Dadvand et al., 2017). The strength of our approach was in accounting for the decline of effects with increasing distance from the residence. The results suggest that trees in proximity to the residence are associated with substantially stronger beneficial effects on health than trees located at a greater distance.
Using a greater buffer size, such as 1,000 m, would result in a reduced sample size as some residences were located near the boundary of the high resolution EnviroAtlas land cover area. Results of an exploratory analysis using the 1,000 m buffer (not shown) suggested that the effects of trees located farther than 500 m from residence on AL MAHA were very small and, therefore, could be ignored. A large nationwide prospective cohort study also found a stronger protective effect on mortality of residential greenness within a 250 m buffer compared to 1,250 m buffer (James et al., 2016).
In the present study, tree cover was associated with reduced AL MAHA while grass cover was not linked with health benefits. In fact, grass cover was associated with a significantly increased AL MAHA in a regression model not adjusting for tree cover. As tree cover and grass cover were strongly negatively correlated in the study area, the observed negative effect of grass cover appeared to be due to confounding by tree cover. The effect of grass cover practically disappeared after adjusting for tree cover (Table 4, Approach D) while the effect of tree cover was not substantially affected by adjusting for grass cover. These findings may not be generalizable outside the study area. Further research in various geographic and climatic areas is warranted to elucidate the types of vegetation and its characteristics that are responsible for subclinical and clinical beneficial health effects.
To summarize, main strengths of this study include: (1) applying the statistically powerful concept of AL as a composite biomarker-based measure of subtle, subclinical health benefits of urban green spaces in a community-based setting; (2) utilizing the available information on biomarkers to the fullest extent and accounting for covariance; (3) accounting for the decline of effects of greenery with increasing distance from the residence; and (4) analyzing effects of different vegetation types and demonstrating that health benefits were associated with greater tree cover near the residence.
In supporting our previous findings (Egorov et al., 2017), this new analysis again demonstrated that summarizing data on multiple biomarkers of health in a single AL index is a very statistically powerful approach that can facilitate studies of subtle subclinical effects of environmental and social factors in small populations. Application of biomarker-based AL indices may enable assessment of subclinical health benefits in community studies and provide opportunities to demonstrate beneficial impacts of local policy interventions aiming to improve the urban environment. Examples of such studies may include urban greening and brownfield revitalization initiatives, and interventions aiming to improve lifestyle. Further research characterizing temporal changes in AL in response to changing environmental or social conditions is warranted in order to support the application of this approach in prospective studies assessing effects of policy measures.
5. Conclusions
This study demonstrated that greater tree cover near residence was associated with reduced Mahalanobis distance-based allostatic load. As allostatic load has been shown to be predictive of morbidity and mortality these results suggest that individuals living in greener environments have reduced risks of adverse health outcomes.
Supplementary Material
Acknowledgments
Funding sources
This article presents results of intramural research at U.S. Environmental Protection Agency (US EPA). Jennifer Styles was funded by the Cooperative Training Agreements CR-83591401-0 and CR-8357850-1 between US EPA and University of North Carolina at Chapel Hill.
Abbreviations
- aOR
adjusted odds ratio
- AL
allostatic load
- BMI
body mass index
- CL
confidence limits
- CRP
C-reactive protein
- DHEA
dehydroepiandrosterone
- ICAM-1
intercellular adhesion molecule 1
- IL
interleukin
- IQR
interquartile range
- pctl
percentile
- SAA
serum amyloid A
- TNF
tumor necrosis factor
- US EPA
United States Environmental Protection Agency
- VCAM-1
vascular cell adhesion molecule 1
Footnotes
Publisher's Disclaimer: Disclaimer
The views expressed in this article are those of the authors and do not necessarily represent the views or policies of US EPA. Mention of trade names, products, or services does not convey, and should not be interpreted as conveying official US EPA approval, endorsement or recommendation.
Human subjects
The observational study described in this article involved human subjects. It has been carried out in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki). The study protocol involving the use of human subjects was approved by the Institutional Review Board at the University of North Carolina in Chapel Hill. Informed consent was obtained from all study participants prior to data collection.
References
- ABHIJITH K, KUMAR P, GALLAGHER J, MCNABOLA A, BALDAUF R, PILLA F, BRODERICK B, DI SABATINO S & PULVIRENTI B 2017. Air pollution abatement performances of green infrastructure in open road and built-up street canyon environments–A review. Atmospheric Environment, 162, 71–86. [Google Scholar]
- ALCOCK I, WHITE MP, WHEELER BW, FLEMING LE & DEPLEDGE MH 2014. Longitudinal effects on mental health of moving to greener and less green urban areas. Environmental science & technology, 48, 1247–1255. [DOI] [PubMed] [Google Scholar]
- ARBEEV KG, COHEN AA, ARBEEVA LS, MILOT E, STALLARD E, KULMINSKI AM, AKUSHEVICH I, UKRAINTSEVA SV, CHRISTENSEN K & YASHIN AI 2016. Optimal Versus Realized Trajectories of Physiological Dysregulation in Aging and Their Relation to Sex-Specific Mortality Risk. Front Public Health, 4, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ARBEEV KG, UKRAINTSEVA SV, BAGLEY O, ZHBANNIKOV IY, COHEN AA, KULMINSKI AM & YASHIN AI 2019. “Physiological Dysregulation” as a Promising Measure of Robustness and Resilience in Studies of Aging and a New Indicator of Preclinical Disease. J Gerontol A Biol Sci Med Sci, 74, 462–468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ASPINALL P, MAVROS P, COYNE R & ROE J 2015. The urban brain: analysing outdoor physical activity with mobile EEG. Br J Sports Med, 49, 272–6. [DOI] [PubMed] [Google Scholar]
- ASTELL-BURT T, FENG X & KOLT GS 2014. Is neighborhood green space associated with a lower risk of type 2 diabetes? Evidence from 267,072 Australians. Diabetes Care, 37, 197–201. [DOI] [PubMed] [Google Scholar]
- BALDAUF R 2017. Roadside vegetation design characteristics that can improve local, near-road air quality. Transportation Research Part D: Transport and Environment, 52, 354–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BARZYK TM, ISAKOV V, ARUNACHALAM S, VENKATRAM A, COOK R & NAESS B 2015. A near-road modeling system for community-scale assessments of traffic-related air pollution in the United States. Environmental Modelling & Software, 66, 46–56. [Google Scholar]
- BELLINGRATH S, WEIGL T & KUDIELKA BM 2009. Chronic work stress and exhaustion is associated with higher allostastic load in female school teachers. Stress, 12, 37–48. [DOI] [PubMed] [Google Scholar]
- BEYER KM, KALTENBACH A, SZABO A, BOGAR S, NIETO FJ & MALECKI KM 2014. Exposure to neighborhood green space and mental health: evidence from the survey of the health of Wisconsin. Int J Environ Res Public Health, 11, 3453–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BORRELL LN, DALLO FJ & NGUYEN N 2010. Racial/ethnic disparities in all-cause mortality in U.S. adults: the effect of allostatic load. Public Health Rep, 125, 810–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BROWNING M & LEE K 2017. Within What Distance Does “Greenness” Best Predict Physical Health? A Systematic Review of Articles with GIS Buffer Analyses across the Lifespan. Int J Environ Res Public Health, 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHANDOLA T & ZHANG N 2018. Re-employment, job quality, health and allostatic load biomarkers: prospective evidence from the UK Household Longitudinal Study. Int J Epidemiol, 47, 47–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CLARK MS, BOND MJ & HECKER JR 2007. Environmental stress, psychological stress and allostatic load. Psychol Health Med, 12, 18–30. [DOI] [PubMed] [Google Scholar]
- COHEN AA, LI Q, MILOT E, LEROUX M, FAUCHER S, MORISSETTE-THOMAS V, LEGAULT V, FRIED LP & FERRUCCI L 2015. Statistical distance as a measure of physiological dysregulation is largely robust to variation in its biomarker composition. PLoS One, 10, e0122541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- COHEN AA, MILOT E, LI Q, LEGAULT V, FRIED LP & FERRUCCI L 2014. Cross-population validation of statistical distance as a measure of physiological dysregulation during aging. Exp Gerontol, 57, 203–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- COHEN AA, MILOT E, YONG J, SEPLAKI CL, FÜLÖP T, BANDEEN-ROCHE K & FRIED LP 2013. A novel statistical approach shows evidence for multi-system physiological dysregulation during aging. Mechanisms of ageing and development, 134, 110–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DADVAND P, BARTOLL X, BASAGANA X, DALMAU-BUENO A, MARTINEZ D, AMBROS A, CIRACH M, TRIGUERO-MAS M, GASCON M, BORRELL C & NIEUWENHUIJSEN MJ 2016. Green spaces and General Health: Roles of mental health status, social support, and physical activity. Environ Int, 91, 161–7. [DOI] [PubMed] [Google Scholar]
- DADVAND P, NIEUWENHUIJSEN MJ, ESNAOLA M, FORNS J, BASAGANA X, ALVAREZ-PEDREROL M, RIVAS I, LOPEZ-VICENTE M, DE CASTRO PASCUAL M, SU J, JERRETT M, QUEROL X & SUNYER J 2015. Green spaces and cognitive development in primary schoolchildren. Proc Natl Acad Sci U S A, 112, 7937–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DADVAND P, TISCHER C, ESTARLICH M, LLOP S, DALMAU-BUENO A, LOPEZ-VICENTE M, VALENTIN A, DE KEIJZER C, FERNANDEZ-SOMOANO A, LERTXUNDI N, RODRIGUEZ-DEHLI C, GASCON M, GUXENS M, ZUGNA D, BASAGANA X, NIEUWENHUIJSEN MJ, IBARLUZEA J, BALLESTER F & SUNYER J 2017. Lifelong Residential Exposure to Green Space and Attention: A Population-based Prospective Study. Environ Health Perspect, 125, 097016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EGOROV AI, CONVERSE R, GRIFFIN SM, STYLES J, KLEIN E, SAMS E, HUDGENS E & WADE TJ 2018. Environmental risk factors for Toxoplasma gondii infections and the impact of latent infections on allostatic load in residents of Central North Carolina. BMC Infect Dis, 18, 421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EGOROV AI, GRIFFIN SM, CONVERSE RR, STYLES JN, SAMS EA, WILSON A, JACKSON LE & WADE TJ 2017. Vegetated land cover near residence is associated with reduced allostatic load and improved biomarkers of neuroendocrine, metabolic and immune functions. Environ Res, 158, 508–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FURMAN D, CAMPISI J, VERDIN E, CARRERA-BASTOS P, TARG S, FRANCESCHI C, FERRUCCI L, GILROY DW, FASANO A, MILLER GW, MILLER AH, MANTOVANI A, WEYAND CM, BARZILAI N, GORONZY JJ, RANDO TA, EFFROS RB, LUCIA A, KLEINSTREUER N & SLAVICH GM 2019. Chronic inflammation in the etiology of disease across the life span. Nat Med, 25, 1822–1832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GASCON M, TRIGUERO-MAS M, MARTINEZ D, DADVAND P, FORNS J, PLASENCIA A & NIEUWENHUIJSEN MJ 2015. Mental health benefits of long-term exposure to residential green and blue spaces: a systematic review. Int J Environ Res Public Health, 12, 4354–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GASCON M, TRIGUERO-MAS M, MARTINEZ D, DADVAND P, ROJAS-RUEDA D, PLASENCIA A & NIEUWENHUIJSEN MJ 2016. Residential green spaces and mortality: A systematic review. Environ Int, 86, 60–7. [DOI] [PubMed] [Google Scholar]
- HALUZA D, SCHONBAUER R & CERVINKA R 2014. Green perspectives for public health: a narrative review on the physiological effects of experiencing outdoor nature. Int J Environ Res Public Health, 11, 5445–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HARTIG T, MITCHELL R, DE VRIES S & FRUMKIN H 2014. Nature and health. Annual Review of Public Health, 35, 207–228. [DOI] [PubMed] [Google Scholar]
- HWANG AC, PENG LN, WEN YW, TSAI YW, CHANG LC, CHIOU ST & CHEN LK 2014. Predicting all-cause and cause-specific mortality by static and dynamic measurements of allostatic load: a 10-year population-based cohort study in Taiwan. J Am Med Dir Assoc, 15, 490–6. [DOI] [PubMed] [Google Scholar]
- ISAKOV V, BARZYK TM, SMITH ER, ARUNACHALAM S, NAESS B & VENKATRAM A 2017. A web-based screening tool for near-port air quality assessments. Environmental modelling & software, 98, 21–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JAMES P, HART JE, BANAY RF & LADEN F 2016. Exposure to Greenness and Mortality in a Nationwide Prospective Cohort Study of Women. Environ Health Perspect, 124, 1344–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JUSTER RP, MCEWEN BS & LUPIEN SJ 2010. Allostatic load biomarkers of chronic stress and impact on health and cognition. Neurosci Biobehav Rev, 35, 2–16. [DOI] [PubMed] [Google Scholar]
- JUSTER RP, SINDI S, MARIN MF, PERNA A, HASHEMI A, PRUESSNER JC & LUPIEN SJ 2011. A clinical allostatic load index is associated with burnout symptoms and hypocortisolemic profiles in healthy workers. Psychoneuroendocrinology, 36, 797–805. [DOI] [PubMed] [Google Scholar]
- KARLAMANGLA AS, MILLER-MARTINEZ D, LACHMAN ME, TUN PA, KORETZ BK & SEEMAN TE 2014. Biological correlates of adult cognition: midlife in the United States (MIDUS). Neurobiol Aging, 35, 387–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KONDO MC, FLUEHR JM, MCKEON T & BRANAS CC 2018. Urban Green Space and Its Impact on Human Health. Int J Environ Res Public Health, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KUO M 2015. How might contact with nature promote human health? Promising mechanisms and a possible central pathway. Frontiers in psychology, 6, 1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MCEWEN BS 2002. Sex, stress and the hippocampus: allostasis, allostatic load and the aging process. Neurobiol Aging, 23, 921–39. [DOI] [PubMed] [Google Scholar]
- MCEWEN BS 2015. Biomarkers for assessing population and individual health and disease related to stress and adaptation. Metabolism, 64, S2–S10. [DOI] [PubMed] [Google Scholar]
- MCEWEN BS & STELLAR E 1993. Stress and the individual. Mechanisms leading to disease. Arch Intern Med, 153, 2093–101. [PubMed] [Google Scholar]
- MILOT E, MORISSETTE-THOMAS V, LI Q, FRIED LP, FERRUCCI L & COHEN AA 2014. Trajectories of physiological dysregulation predicts mortality and health outcomes in a consistent manner across three populations. Mech Ageing Dev, 141-142, 56–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PETROVIC D, PIVIN E, PONTE B, DHAYAT N, PRUIJM M, EHRET G, ACKERMANN D, GUESSOUS I, YOUNES SE, PECHERE-BERTSCHI A, VOGT B, MOHAUPT M, MARTIN PY, PACCAUD F, BURNIER M, BOCHUD M & STRINGHINI S 2016. Sociodemographic, behavioral and genetic determinants of allostatic load in a Swiss population-based study. Psychoneuroendocrinology, 67, 76–85. [DOI] [PubMed] [Google Scholar]
- PICKARD BR, DANIEL J, MEHAFFEY M, JACKSON LE & NEALE A 2015. EnviroAtlas: A new geospatial tool to foster ecosystem services science and resource management. Ecosystem Services, 14, 45–55. [Google Scholar]
- RIBEIRO AI, SANTOS AC, VIEIRA VM & BARROS H 2019a. Hotspots of childhood obesity in a large metropolitan area: does neighbourhood social and built environment play a part? Int J Epidemiol. 1–10. 10.1093/ije/dyz205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- RIBEIRO AI, TAVARES C, GUTTENTAG A & BARROS H 2019b. Association between neighbourhood green space and biological markers in school-aged children. Findings from the Generation XXI birth cohort. Environ Int, 132, 105070. [DOI] [PubMed] [Google Scholar]
- SEEMAN TE, SINGER BH, ROWE JW, HORWITZ RI & MCEWEN BS 1997. Price of adaptation--allostatic load and its health consequences. MacArthur studies of successful aging. Arch Intern Med, 157, 2259–68. [PubMed] [Google Scholar]
- TAMOSIUNAS A, GRAZULEVICIENE R, LUKSIENE D, DEDELE A, REKLAITIENE R, BACEVICIENE M, VENCLOVIENE J, BERNOTIENE G, RADISAUSKAS R, MALINAUSKIENE V, MILINAVICIENE E, BOBAK M, PEASEY A & NIEUWENHUIJSEN MJ 2014. Accessibility and use of urban green spaces, and cardiovascular health: findings from a Kaunas cohort study. Environ Health, 13, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- TRIGUERO-MAS M, DADVAND P, CIRACH M, MARTINEZ D, MEDINA A, MOMPART A, BASAGANA X, GRAZULEVICIENE R & NIEUWENHUIJSEN MJ 2015. Natural outdoor environments and mental and physical health: relationships and mechanisms. Environ Int, 77, 35–41. [DOI] [PubMed] [Google Scholar]
- VAN DEN BOSCH M & ODE SANG A 2017. Urban natural environments as nature-based solutions for improved public health - A systematic review of reviews. Environ Res, 158, 373–384. [DOI] [PubMed] [Google Scholar]
- WEBSTER T, VIEIRA V, WEINBERG J & ASCHENGRAU A 2006. Method for mapping population-based case-control studies: an application using generalized additive models. Int J Health Geogr, 5, 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO 2016. Urban green spaces and health - a review of evidence. In: EGOROV AI MP, BRAUBACH M, MARTUZZI M (ed.). Copenhagen: World Health Organization. [Google Scholar]
- WIRTZ PH & VON KANEL R 2017. Psychological Stress, Inflammation, and Coronary Heart Disease. Curr Cardiol Rep, 19, 111. [DOI] [PubMed] [Google Scholar]
- YLI-PELKONEN V, VIIPPOLA V, KOTZE DJ & SETÄLÄ H 2017. Greenbelts do not reduce NO2 concentrations in near-road environments. Urban Climate, 21, 306–317. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.