

Abstract
Background
Schistosomiasis japonica is an infectious disease caused by Schistosoma japonicum that seriously endangers human health. Domestic animals have important roles in disease transmission and goats are considered a primary reservoir host and source of infection. The prevalence and intensity of schistosomiasis infections have significantly decreased in China, and a more sensitive, specific detection method is urgently needed. The aim of this study was to develop a real-time PCR assay for accurate detection of S. japonicum infection in goats.
Methods
A real-time PCR method for detecting schistosomiasis japonica in goats was developed by amplification of a specific S. japonicum DNA fragment, and validated using a total of 94 negative and 159 positive plasma and serum samples collected in our previous study of S. japonicum infection. Both plasma and serum samples were evaluated by real-time PCR and enzyme-linked immunosorbent assay (ELISA). In addition, 120 goat plasma samples from an S. japonicum-endemic area (Wangjiang) and 33 from a non-endemic region (Weihai) were collected and evaluated using our method.
Results
The sensitivity and specificity of the real-time PCR for detecting infected samples were 98.74% (157/159, 95% CI: 95.53–99.85%) and 100% (94/94, 95% CI: 96.15–100%), respectively. For the ELISA, sensitivity and specificity were 98.11% (156/159, 95% CI: 94.59–99.61%) and 90.43% (85/94, 95% CI: 82.60–95.53%), respectively. Further, we found positivity rates for S. japonicum infection in Wangjiang and Weihai of 8.33% (10/120, 95% CI: 4.07–14.79%) and 0% (0/33, 95% CI: 0–10.58%), respectively.
Conclusions
The results of this study indicate that our real-time PCR method exhibits higher sensitivity and specificity than ELISA and is a useful method for detection of S. japonicum infection in goats.
Keywords: Schistosoma japonicum, Diagnosis, Real-time PCR, ELISA, Goats
Background
Schistosomiasis is a significant infectious disease caused by trematode worms of the genus Schistosoma, which is a global public health concern. Worldwide, schistosomiasis is the second most devastating parasitic disease after malaria, and it is mainly distributed in Asia, South America and Africa [1, 2]. The three major schistosome species that cause infection in humans are Schistosoma japonicum, Schistosoma mansoni and Schistosoma haematobium, among which, S. japonicum is mainly distributed in southeast Asian (China, The Philippines, etc.). According to previous studies, more than a million humans are infected with S. japonicum. Therefore, the disease remains a potential public health issue, despite considerable success in blocking transmission over the past decades [3–5].
Eradicating sources of infection is a vital strategy to eliminate schistosomiasis japonica [6]. Domestic animals are the primary source of infection and play an important role in disease transmission. In addition to bovines, goats are recognized as another major reservoir host in China [7]. Given the significantly decreasing prevalence and intensity of infection in China, development of a diagnostic method with high sensitivity and specificity for testing domestic animals is critical for effective control of schistosomiasis japonica [8].
Pathogenic and immunological diagnosis methods are used commonly for detection of S. japonicum infection. Pathogenic diagnosis primarily involves the Kato-Katz technique and miracidium hatching test (MHT), which have been regarded as the ‘gold standard’ for diagnosis of schistosomiasis [9, 10]. These approaches are used widely because of their low cost and portability; however, their disadvantages include low sensitivity in areas with low-intensity infections and the fact that they are relatively time-consuming [11]. Immunological diagnostic methods include enzyme-linked immunosorbent assay (ELISA) and the indirect hemagglutination (IHA) test, in addition to other methods based on the antibody and antigen reaction [12, 13]. These approaches have higher sensitivity and are simple to apply; however, they have higher cross-reactivity and false positive rates [14]. Given the limitations of pathogenic and immunological diagnosis methods, a more sensitive and specific detection technique is required for schistosomiasis diagnosis [15].
With the development of molecular diagnostic technology, nucleic acid detection of S. japonicum infection has become a research hotspot in recent years. Many studies have demonstrated that nucleic acid detection of S. japonicum infection has higher sensitivity and specificity, and less cross-reactivity than traditional methods [7, 16–18]. Molecular diagnostic techniques used include general PCR, nested-PCR, real-time PCR, loop-mediated isothermal amplification (LAMP), and recombinant enzyme polymerase amplification (RPA), among others [19]. In many previous studies, highly repetitive schistosome gene sequences were used as targets for nucleic acid detection of schistosomiasis. These sequences were mainly cell-free circulating schistosome DNA, derived from the dead worms during migration and the epidermal tissue cells, during their growth process [20–22]. In this study, a real-time PCR method for detecting S. japonicum DNA in goat plasma was developed and used to evaluate S. japonicum infection in goats from endemic and non-endemic regions.
Methods
Worm and egg collection
Adult S. japonicum worms (42 days post-infection) were collected from the portal vein systems of five BALB/c mice artificially infected with S. japonicum cercariae [23]. Male and female S. japonicum worms were manually separated and S. japonicum eggs collected from infected BALB/c mouse livers [24]. All S. japonicum worms and eggs were stored in RNAlater (Sigma-Aldrich, St. Louise, USA) at − 20 °C until use.
Collection of plasma and serum samples
A total of 159 positive plasma and serum samples were simultaneously collected from artificially infected goats. The goat was restrained by a special restraint device, which could make it supine lying, preventing the animal from struggling. The abdominal wool was shaved off and an area of skin of approximately 10 × 10 cm was exposed. Then, goats were percutaneously infected with 300 cercariae for 20 min after counting the cercariae under a microscope. In addition, a total of 94 plasma and serum samples were collected from 94 goats without schistosome infection. Plasma and serum samples were collected respectively in EDTA-K2 vacuum blood collection tubes and a vacuum blood collection tubes with no additive. The blood was collected from jugular vein of each goat, and the supernatant was separated after centrifugation at 3000× rpm for 10 min at 25 °C. Plasma samples from 120 goats which grazed freely on marshlands were collected in Wangjiang county, which is a region endemic for schistosomiasis japonica in Anhui Province, China. Further, plasma samples from 33 goats which grazed freely on farms were collected in Weihai city, which is a non-endemic area for schistosomiasis japonica in Shandong Province, China. All plasma and serum samples were stored at − 20 °C until use. In addition, 32 positive plasma samples and their corresponding serum samples, as well as 10 positive plasma samples, were selected randomly for storage and use in later experiments.
Plasma storage
The 10 positive plasma samples were randomly selected, divided into 4 aliquots of 6 ml each, and stored at 4 °C, − 20 °C, − 40 °C and − 80 °C, respectively. Samples were analyzed after 14 days, 28 days, 42 days, 3 months, 6 months, and 1 year of storage.
Extraction of genomic and circulating DNA
Genomic DNA was extracted from the tissues of an uninfected goat, and from male and female S. japonicum worms and eggs using a TIANamp Genomic DNA Kit (Tiangen Biotech, Beijng, China), according to the manufacturer’s instructions. Genomic DNA samples from Haemonchus contortus, Fasciola gigantica, Toxoplasma gondii, Sarcocystis sp., Trichinella spiralis, Paramphistomum, Babesia, and Spirometra were provided by our institute and stored at − 20 °C.
DNA was extracted from goat plasma/serum samples using a Magnetic Serum/Plasma Circulating DNA Maxi Kit (Tiangen Biotech), according to the manufacturer’s instructions. Extracted DNA samples were stored at − 20 °C.
Establishment and optimization of the real-time PCR detection system
PCR primers were designed according to a DNA fragment containing S. japonicum target sequence (GenBank: FN356222.1), which was verified through bioinformatics analysis and some screening experiments in our laboratory: forward primer 1 (FP1: 5ʹ-GCA GCG GCT TTA GGC AAC AC-3ʹ) and reverse primer 1 (RP1: 5ʹ-TCA AAC TAA TCC CTC TAT GGT TAT CAC AAG-3ʹ) (Fig. 1). PCR was performed using LA Taq DNA polymerase (Takara, Kyoto, Japan). The total reaction volume was 50 μl and included 5 μl 10× LA PCR Buffer II (Mg2+ plus), 8 μl dNTP mixture, 0.5 μl TaKaRa LA Taq, 1 μl each primer (FP1 and RP1), 32 μl ddH2O, and 2.5 μl DNA template. The reaction procedure comprised an initial denaturing step at 94 °C for 30 s, followed by 40 cycles at 94 °C for 15 s, 58 °C for 34 s, and 72 °C for 10 s.
Fig. 1.
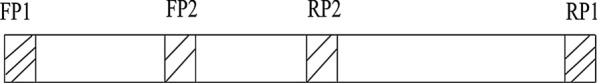
Relationship between two pairs of primers (FP1/RP1 and FP2/RP2)
PCR products were purified using a Universal DNA Purification Kit (Tiangen Biotech), according to the manufacturer’s instructions. The purified products were then subcloned into the pMD-19T vector overnight at 4 °C and recombinant plasmids transferred into Escherichia coli DH5α [25]. Recombinant plasmids were extracted using a Mini Plasmid Kit (Tiangen Biotech) and confirmed by DNA sequencing. Plasmid concentration was detected using an ultra-micro nucleic acid protein instrument (Thermo Fisher Scientific, Waltham, USA), plasmids were diluted to 2.6 ng/μl, and a 10-fold gradient dilution, up to 10−9, generated using sterile double distilled water for use in real-time PCR detection.
Real-time PCR primers were designed according to a specific DNA fragment used as target sequence (GenBank: FN356222.1), which was identified in our previous study (our unpublished data). The forward primer 2 (FP2: 5ʹ-CCG AAC ACT TCA AGG AAC AGT TTA G-3ʹ) and the reverse primer 2 (RP2: 5ʹ-CTT CCT CGT TTC AGG TTA GAT ATA GC-3ʹ) were used (Fig. 1). The total reaction system volume was 20 μl, including 10 μl 2× ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech, Nanjing, China), 0.4 μl (10 μmol/l) of each primer (FP2 and RP2), 5.2 μl ddH2O, and 4 μl (~20 ng) DNA template. The reaction procedure comprised an initial denaturing step at 94 °C for 30 s, followed by 40 cycles at 94 °C for 15 s, 58 °C for 34 s, and 72 °C for 10 s, and a melting curve step, comprising denaturation at 95 °C for 15 s, annealing at 60 °C for 15 s, and extension at 72 °C for 15 s using real-time PCR.
Optimization of annealing temperature was conducted using the S. japonicum target gene plasmid as a template, with annealing temperatures of 57 °C, 58 °C and 59 °C. To optimize plasma volume, 400, 500, 600 and 750 μl plasma from the same 10 positive goat samples were used. Furthermore, the amount of template DNA was optimized by extracting 10 nucleic acid samples from 600 μl plasma from infection-positive goats, using an annealing temperature of 58 °C and either 2 μl (approximately 10 ng) or 4 μl (approximately 20 ng) template.
Detection of schistosomiasis japonica by real-time PCR
The plasmid containing the target sequence was used as a positive control. DNA from uninfected negative goat plasma was used as a negative control, and sterile double distilled water was used as a blank control in each assay. In each PCR detection, the positive control, the negative control and the blank control were included. All samples, positive control, negative control and blank control were detected in duplicate. Assays were performed using a qTOWER 3G instrument (Analytik Jena AG, Jena, Germany), and results determined using melting curve analysis. Samples were judged positive when both duplicate reactions were positive; suspicious when one reaction was positive, and sample nucleic acid re-extracted for a second detection, following which samples were judged as positive if one reaction was positive; and a sample was considered negative when two reactions were negative.
Detection of schistosomiasis japonica by ELISA
Schistosoma japonicum soluble egg antigens (SEA) were prepared according to previously described methods [26]. SEA were the detection antigen and all serum samples were tested by ELISA. First, SEA were diluted appropriately with coating solution, used to coat 96-well microtiter plates (100 μg per well; Corning, Corning, USA) overnight at 4 °C, then washed three times with 0.05% Tween 20 in PBS (PBST). Next, plates were blocked with 1% BSA (100 μl/well), incubated for 1 h at 37 °C, then washed three times. Serum samples were diluted 1:100 with PBS, added (100 μl/well), incubated for 2 h at 37 °C, and washed three times. HRP-labeled donkey anti-goat IgG (H+L) (Beyotime, Shanghai, China) diluted 1:3000 in PBST was added (100 μl/well), incubated for 1 h at 37 °C, and then washed three times. Finally, 100 μl/well soluble TMB substrate solution (Tiangen Biotech) was added, plates incubated in the dark for 5–10 min, and reactions stopped by adding 2 M sulfuric acid (50 μl/well). Optical density (OD) was measured at 450 nm on a microplate reader (BioTek, Vermont, USA). In each ELISA detection, a positive serum control and a negative serum control were included. All samples, positive serum control and negative serum control were detected in triplicate. The mean absorbance value of the reference negative serum multiplied by 2.1 was set as the cutoff value. A sample was judged positive when its mean absorbance value was higher than the cutoff value.
Statistical analysis
Sensitivity and specificity were calculated as follows: sensitivity = no. of true positives/(no. of true positives + false negatives); specificity = no. of true negatives/(no. of true negatives + false positives). Stata/SE 12.0 was used to calculate 95% confidence intervals (CI) for the sensitivity and specificity of each detection. The mean and standard deviation (SD) were calculated. Comparisons among methods of analysis were conducted using a Chi-square test in SPSS Statistics (version 20). A value of P < 0.05 was considered statistically significant.
Results
Establishment of a real-time PCR detection system to detect S. japonicum
The real-time PCR results were determined using melting curve analysis (Fig. 2). The results showed that the positive detection rates were 100% (10/10) at annealing temperatures of 57 °C and 58 °C; however, the melting curve for the reaction had more than one peak at 57 °C. At an annealing temperature of 59 °C, the detection positive rate was 90% (9/10) and the melting curve had single peak. Therefore, we concluded that 58 °C was the optimum annealing temperature.
Fig. 2.

Real-time PCR melting curves. Key: 1, positive control; 2–4, positive samples; 5–6, negative samples; 7, negative control; 8, blank control
The positive detection rates using 400, 500, 600 and 750 μl goat plasma samples were 60% (6/10), 80% (8/10), 100% (10/10) and 100% (10/10) by real-time PCR; hence, the optimal plasma volume was determined as 600 μl. Real-time PCR was performed using 2 μl (c.10 ng) and 4 μl (c.20 ng) template, and the detection positive rates were 80% (8/10) and 100% (10/10), respectively; hence, the final optimal template amount was determined as 4 μl.
Detection using plasma and serum samples
Nucleic acids samples were extracted from 32 positive goat plasma and corresponding serum samples and analyzed by real-time PCR detection using the same conditions. The results showed that all 32 plasma samples were positive (32/32, 95% CI: 89.11–100%) and 27 serum samples were positive (84.38%, 27/32, 95% CI: 67.21–94.72%). These data indicate that plasma samples were more suitable for detection than serum samples.
Effects of plasma storage
Next, real-time PCR was performed using samples stored in different environments and for various lengths of time. After storage of plasma samples for 14 days at 4 °C, − 20 °C, − 40 °C and − 80 °C, detection rates were 100% (10/10). Nine samples were positive (90%, 9/10) following storage at 4 °C after 28 days, while detection rates of samples stored at the three other temperatures were 100% (10/10). Three samples were positive (30%, 3/10) after storage at 4 °C for 42 days, while detection rates for samples stored at the other three temperatures were 100% (10/10); the detection rates for all samples stored at − 20 °C, − 40 °C, and − 80 °C were 100% (10/10) after 3 months, 6 months, and 1 year, respectively (Table 1).
Table 1.
Results of real-time PCR analysis of plasma following storage for various time periods and at different temperatures
| Storage time | Temperature | |||
|---|---|---|---|---|
| 4 °C | − 20 °C | − 40 °C | − 80 °C | |
| 14 days | 100% | 100% | 100% | 100% |
| 28 days | 90% | 100% | 100% | 100% |
| 42 days | 30% | 100% | 100% | 100% |
| 3 months | na | 100% | 100% | 100% |
| 6 months | na | 100% | 100% | 100% |
| 12 months | na | 100% | 100% | 100% |
Abbreviation: na, no detection
Sensitivity and specificity of real-time PCR and ELISA
The results of real-time PCR showed that there was no target gene amplification from goat genomic DNA, while the assay could be used to detect schistosomiasis japonica in goats. Sample concentration was calculated using a standard curve based on a 10-fold gradient dilution of the positive control plasmid. The amplification curve (Fig. 3) indicated that the lowest detection level was 10−7 dilution, with a corresponding copy number of 74.9. The copy numbers of the target gene in one egg, a single male, and a single female of S. japonicum were (8.73 ± 0.03) × 106, (8.61 ± 0.03) × 109, and (5.51 ± 0.03) × 109, respectively.
Fig. 3.

Real-time PCR amplification curves for the positive control plasmid at different dilutions. Key: 1–8, plasmid diluted at 1 (1 ng/μl); 10−1 (100 pg/μl); 10−2 (10 pg/μl); 10−3 (1 pg/μl); 10−4 (100 fg/μl); 10−5 (10 fg/μl); 10−6 (1 fg/μl); and 10−7 (0.1 fg/μl), respectively
The sensitivity and specificity of real-time PCR were 98.74% (157/159, 95% CI: 95.53–99.85%) and 100% (94/94, 95% CI: 96.15–100%), respectively. Meanwhile, the sensitivity and specificity of ELISA were 98.11% (156/159, 95% CI: 94.59–99.61%) and 90.43% (85/94, 95% CI: 82.60–95.53%), respectively. There was no significant difference in sensitivity between real-time PCR and ELISA for detecting goat schistosomiasis japonica (P = 0.652); however, there was a significant difference in the specificity of the two methods in goat samples (P = 0.002).
Cross-reactivity of real-time PCR
PCR assays using genomic DNA of eight parasite species (H. contortus, F. gigantica, T. gondii, Sarcocystis sp., T. spiralis, Paramphistomum, Babesia and Spirometra) as template were negative, with no amplification of the target gene detected. Thus, our real-time PCR method exhibited no cross-reactivity with these organisms.
Real-time PCR detection using samples from goats from endemic and non-endemic areas
Our real-time PCR method was used to analyze 120 plasma samples from goats from an endemic area (Wangjiang) and 33 from goats from a non-endemic area (Weihai). The results showed that the positive rate of samples from Wangjiang was 8.33% (10/120, 95% CI: 4.07–14.79%), while that in Weihai was 0% (0/33, 95% CI: 0–10.58%).
Discussion
More than 40 mammalian species can be reservoir hosts of S. japonicum, and goats are an important source of human infection. Due to large-scale treatment with praziquantel, safety education, and extensive comprehensive control strategies, the prevalence and infection intensity of schistosomiasis japonica have been reduced successfully and maintained at a very low level [27, 28]; however, the disease continues to spread, although at a very low-intensity of infection, and eggs in the feces of goats are one of the main sources of transmission [29].
Currently, no S. japonicum diagnosis is available with easy field access and high sensitivity and specificity in low-intensity infection areas [30]. Thus, schistosomiasis diagnosis remains challenging in areas of low infection intensity [31, 32]. The real-time PCR method has numerous advantages for diagnosis, including its high sensitivity, high specificity, low cross-reactivity with other parasites, and the fact that the results can be evaluated directly, by the melting curve. Schistosoma japonicum can infect many mammals; however, not all have corresponding commercial secondary antibodies. Thus, the use of immunological methods to diagnose schistosomiasis japonica is limited in different animals. In contrast, the real-time PCR method detects S. japonicum nucleic acid, providing the potential advantage for schistosomiasis diagnosis in a range of animals.
In this study, we developed a real-time PCR diagnostic method to detect schistosomiasis japonica in goat plasma samples. The real-time PCR target gene was a specific DNA fragment, previously screened in our laboratory (data not shown). This gene could be detected in schistosomiasis japonica-positive plasma, and there was no cross-reaction with goat genomic DNA. Therefore, we determined that it was suitable for use in detecting schistosomiasis japonica in goats.
In previous studies, host serum was generally used for extraction of nucleic acids and PCR detection [33–35]. In the present study, plasma and serum samples from schistosomiasis-positive goats were analyzed. The detection was higher in goat plasma than that in serum. This may be because cell-free DNA is more stable in plasma than serum, and DNase activity is very high in serum, leading to rapid degradation of cell-free DNA. Further, the EDTA used in the preparation of plasma is a DNase inhibitor, and can inhibit cell-free DNA degradation in plasma. Moreover, plasma contains more fibrinogen and clotting factors than serum, and it is possible that serum can drag down a proportion of cell-free DNA during the clotting process, leading to a loss of cell-free DNA in serum compared with plasma [36, 37].
The influences of storage temperature and time on plasma samples were also evaluated in this study. Positive plasma samples were stable at least for one year when stored at − 20 °C, − 40 °C and − 80 °C. The detection rate was 100% within two weeks when samples were stored at 4 °C; however, after four weeks at 4 °C, detection rates declined. Therefore, plasma samples should not be stored for over two weeks at 4 °C, and can be used for at least for one year if stored at − 20 °C, − 40 °C, or − 80 °C.
The lower limit of detection for the real-time PCR method was 0.26 fg target gene, with a corresponding copy number of 74.9. Moreover, in single male and female mean S. japonicum genome copy numbers were (8.61 ± 0.03) × 109 and (5.51 ± 0.03) × 109, respectively, while a single egg contained only (8.73 ± 0.03) × 106 copies. In previous study, we reported a nested-PCR method that can detect target DNA from a single egg; however, it required two-rounds of reactions, which could increase the risk of contamination [7]. In contrast, the real-time PCR assay developed here only requires a single-round reaction and the results can be judged directly by melting curve analysis, which is a potentially significant advantage.
In this study, the real-time PCR exhibited slightly better performance than ELISA for detection of S. japonicum-positive samples. There was no significant difference in sensitivity between real-time PCR and ELISA for detection of schistosomiasis japonica-positive samples from goats (P = 0.652); however, there was a significant difference in specificity between the two methods (P = 0.002). Hence, real-time PCR may improve the specificity of schistosomiasis diagnosis in goats, with superior specificity compared with ELISA.
In addition, the real-time PCR method did not cross-react with H. contortus, F. gigantica, T. gondii, Sarcocystis sp., T. spiralis, Paramphistomum, Babesia and Spirometra. Nevertheless, in some previous studies, ELISA methods cross-reacted with H. contortus in goats and Paramphistomum in water buffaloes [13, 38]. Thus, our real-time PCR method has the potential to improve the detection accuracy compared with that achieved using an ELISA approach.
Finally, this method was used to evaluate the goats from endemic and non-endemic regions. No samples from Weihai tested positive, while ten samples from Wangjiang did. In a previous study, a nested-PCR method was also used to evaluate the schistosomiasis japonica infection in goats from Wangjiang, and the positive rate was 16.67% [7]. Although real-time PCR may be a more sensitive method than nested-PCR, the positive rate using real-time PCR was slightly lower (8.33%) than that using nested-PCR. We speculate that this may be due to the considerable efforts made to control and prevent schistosomiasis japonica in goats in Wangjiang; however, our data demonstrate that schistosomiasis japonica infection in goats in endemic regions remains at a high level and warrants further attention. Plasma samples from goats from Weihai were all negative, confirming that the real-time PCR method has excellent specificity. In this study, DNA extracted from the plasma of goats was finally selected for detection after comparing the diagnostic effect of plasma and serum from goats. Meanwhile, to the best of our knowledge, this is the first time schistosomiasis japonica has been diagnosed in goats by real-time PCR. The sensitivity and specificity of this method was not worse than other PCR methods that have been used to diagnose schistosomiasis in human or other animals for detecting DNA from different samples. However, real-time PCR also has some shortcomings, such as reliance on expensive instruments and its higher cost, which would be resolved with continuous development.
Conclusions
The real-time PCR method developed in this study has higher sensitivity and specificity than previously published approaches and has potential for application for detection of S. japonicum infection in domestic animals, particularly goats.
Acknowledgements
The authors wish to thank Hao Li and Ke Lu from the National Reference Laboratory of Animal Schistosomiasis, Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences, for technical assistance with parasite collection.
Abbreviations
- ELISA
enzyme-linked immunosorbent assay
- MHT
miracidium hatching test
- IHA
indirect hemagglutination assay
- LAMP
loop-mediated isothermal amplification
- RPA
recombinant enzyme polymerase amplification
- SEA
soluble egg antigen
Authors’ contributions
QG, CC, KZ and YY performed the experiments. QG drafted the manuscript. YH and JL designed the study and critically revised the manuscript. QG, JL and ZF performed the statistical analysis. YL, LT, JZ, PS and KZ, provided most of materials needed. All authors read and approved the final manuscript.
Funding
This study was supported by the Shanghai Science and Technology Promotion Agriculture Innovation Program (2019No.3-3), the National Key Research and Development Program of China (2017YFD0501306), the National Natural Science Foundation of China (31402192, 31872256) and the Scientific and Technical Innovation Project of the Chinese Academy of Agricultural Sciences.
Availability of data and materials
Data supporting the conclusions of this article are included within the article.
Ethics approval and consent to participate
All animals were approved by the Animal Care and Use Committee of the Shanghai Veterinary Research Institute.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Qinghong Guo, Email: 1909106236@qq.com.
Cheng Chen, Email: 1571854290@qq.com.
Keke Zhou, Email: 1693069845@qq.com.
Yugang Li, Email: hcqykzx@163.com.
Laibao Tong, Email: tlbwjx@163.com.
Yongcheng Yue, Email: 1628853935@qq.com.
Kerou Zhou, Email: 956792656@qq.com.
Jinming Liu, Email: jimyliu@shvri.ac.cn.
Zhiqiang Fu, Email: fuzhiqiang@shvri.ac.cn.
Jiaojiao Lin, Email: jjlin@shvri.ac.cn.
Jiaxi Zhao, Email: wjzhaojiaxi@163.com.
Pengxiang Sun, Email: 834253908@qq.com.
Yang Hong, Email: hongyang_7@126.com.
References
- 1.Gryseels B, Polman K, Clerinx J, Kestens L. Human schistosomiasis. Lancet. 2006;368:1106–1118. doi: 10.1016/S0140-6736(06)69440-3. [DOI] [PubMed] [Google Scholar]
- 2.LoVerde PT. Schistosomiasis. Adv Exp Med Biol. 2019;1154:45–70. doi: 10.1007/978-3-030-18616-6_3. [DOI] [PubMed] [Google Scholar]
- 3.Hotez PJ, Alvarado M, Basanez MG, Bolliger I, Bourne R, Boussinesq M, et al. The global burden of disease study 2010: interpretation and implications for the neglected tropical diseases. PLoS Negl Trop Dis. 2014;8:e2865. doi: 10.1371/journal.pntd.0002865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Utzinger J, Zhou XN, Chen MG, Bergquist R. Conquering schistosomiasis in China: the long march. Acta Trop. 2005;96:69–96. doi: 10.1016/j.actatropica.2005.08.004. [DOI] [PubMed] [Google Scholar]
- 5.Zhou XN, Wang LY, Chen MG, Wu XH, Jiang QW, Chen XY, et al. The public health significance and control of schistosomiasis in China - then and now. Acta Trop. 2005;96:97–105. doi: 10.1016/j.actatropica.2005.07.005. [DOI] [PubMed] [Google Scholar]
- 6.Qian C, Zhang Y, Zhang X, Yuan C, Gao Z, Yuan H, et al. Effectiveness of the new integrated strategy to control the transmission of Schistosoma japonicum in China: a systematic review and meta-analysis. Parasite. 2018;25:54. doi: 10.1051/parasite/2018058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhang X, He CC, Liu JM, Li H, Lu K, Fu ZQ, et al. Nested-PCR assay for detection of Schistosoma japonicum infection in domestic animals. Infect Dis Poverty. 2017;6:86. doi: 10.1186/s40249-017-0298-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang LJ, Xu ZM, Guo JY, Dai SM, Dang H, Lu S, et al. Endemic status of schistosomiasis in People’s Republic of China in 2018. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2019;31:576–582. doi: 10.16250/j.32.1374.2019247. [DOI] [PubMed] [Google Scholar]
- 9.de Noya B, Ruiz R, Losada S, Colmenares C, Contreras R, Cesari IM, et al. Detection of schistosomiasis cases in low-transmission areas based on coprologic and serologic criteria: the Venezuelan experience. Acta Trop. 2007;103:41–9. [DOI] [PubMed]
- 10.Yu JM, de Vlas SJ, Jiang QW, Gryseels B. Comparison of the Kato-Katz technique, hatching test and indirect hemagglutination assay (IHA) for the diagnosis of Schistosoma japonicum infection in China. Parasitol Int. 2007;56:45–49. doi: 10.1016/j.parint.2006.11.002. [DOI] [PubMed] [Google Scholar]
- 11.Barenbold O, Raso G, Coulibaly JT, NʼGoran EK, Utzinger J, Vounatsou P. Estimating sensitivity of the Kato-Katz technique for the diagnosis of Schistosoma mansoni and hookworm in relation to infection intensity. PLoS Negl Trop Dis. 2017;11:e0005953. [DOI] [PMC free article] [PubMed]
- 12.Wang XY, Xu J, Zhao S, Li W, Zhang JF, He J, et al. Estimating the prevalence of schistosomiasis japonica in China: a serological approach. Infect Dis Poverty. 2018;7:62. doi: 10.1186/s40249-018-0443-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lv C, Hong Y, Fu Z, Lu K, Cao X, Wang T, et al. Evaluation of recombinant multi-epitope proteins for diagnosis of goat schistosomiasis by enzyme-linked immunosorbent assay. Parasit Vectors. 2016;9:135. doi: 10.1186/s13071-016-1418-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang W, Li Y, Li H, Xing Y, Qu G, Dai J, et al. Immunodiagnostic efficacy of detection of Schistosoma japonicum human infections in China: a meta analysis. Asian Pac J Trop Med. 2012;5:15–23. doi: 10.1016/S1995-7645(11)60238-1. [DOI] [PubMed] [Google Scholar]
- 15.Weerakoon KG, Gobert GN, Cai P, McManus DP. Advances in the diagnosis of human schistosomiasis. Clin Microbiol Rev. 2015;28:939–967. doi: 10.1128/CMR.00137-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.He P, Gordon CA, Williams GM, Li Y, Wang Y, Hu J, et al. Real-time PCR diagnosis of Schistosoma japonicum in low transmission areas of China. Infect Dis Poverty. 2018;7:8. doi: 10.1186/s40249-018-0390-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mwangi IN, Agola EL, Mugambi RM, Shiraho EA, Mkoji GM. Development and evaluation of a loop-mediated isothermal amplification assay for diagnosis of Schistosoma mansoni infection in faecal samples. J Parasitol Res. 2018;2018:7. doi: 10.1155/2018/1267826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.ten Hove RJ, Verweij JJ, Vereecken K, Polman K, Dieye L, van Lieshout L. Multiplex real-time PCR for the detection and quantification of Schistosoma mansoni and S. haematobium infection in stool samples collected in northern Senegal. Trans R Soc Trop Med Hyg. 2008;102:179–85. [DOI] [PubMed]
- 19.Weerakoon KG, Gordon CA, McManus DP. DNA Diagnostics for schistosomiasis control. Trop Med Infect Dis. 2018;3:81. doi: 10.3390/tropicalmed3030081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bischoff FZ, Lewis DE, Simpson JL. Cell-free fetal DNA in maternal blood: kinetics, source and structure. Hum Reprod Update. 2005;11:59–67. doi: 10.1093/humupd/dmh053. [DOI] [PubMed] [Google Scholar]
- 21.Xu J, Liu AP, Guo JJ, Wang B, Qiu SJ, Sun H, et al. The sources and metabolic dynamics of Schistosoma japonicum DNA in serum of the host. Parasitol Res. 2013;112:129–133. doi: 10.1007/s00436-012-3115-3. [DOI] [PubMed] [Google Scholar]
- 22.Wichmann D, Panning M, Quack T, Kramme S, Burchard GD, Grevelding C, et al. Diagnosing schistosomiasis by detection of cell-free parasite DNA in human plasma. PLoS Negl Trop Dis. 2009;3:e422. doi: 10.1371/journal.pntd.0000422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gu K, Li Y, Driguez P, Zeng Q, Yu X, Sun H, et al. Clinical diagnostic value of viable Schistosoma japonicum eggs detected in host tissues. BMC Infect Dis. 2017;17:244. doi: 10.1186/s12879-017-2362-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Osada Y, Kumagai T, Masuda K, Suzuki T, Kanazawa T. Mutagenicity evaluation of Schistosoma spp. extracts by the umu-test and V79/HGPRT gene mutation assay. Parasitol Int. 2005;54:29–34. [DOI] [PubMed]
- 25.Kostylev M, Otwell AE, Richardson RE, Suzuki Y. Cloning Should Be Simple: Escherichia coli DH5alpha-mediated assembly of multiple DNA fragments with short end homologies. PLoS One. 2015;10:e0137466. doi: 10.1371/journal.pone.0137466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hong Y, Sun A, Zhang M, Gao F, Han Y, Fu Z, et al. Proteomics analysis of differentially expressed proteins in schistosomula and adult worms of Schistosoma japonicum. Acta Trop. 2013;126:1–10. doi: 10.1016/j.actatropica.2012.12.009. [DOI] [PubMed] [Google Scholar]
- 27.Spear RC, Seto EY, Carlton EJ, Liang S, Remais JV, Zhong B, et al. The challenge of effective surveillance in moving from low transmission to elimination of schistosomiasis in China. Int J Parasitol. 2011;41:1243–1247. doi: 10.1016/j.ijpara.2011.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Song LG, Wu XY, Sacko M, Wu ZD. History of schistosomiasis epidemiology, current status, and challenges in China: on the road to schistosomiasis elimination. Parasitol Res. 2016;115:4071–4081. doi: 10.1007/s00436-016-5253-5. [DOI] [PubMed] [Google Scholar]
- 29.Jiao-Jiao L. Strengthening the control of goat schistosomiasis to facilitate the progress towards elimination of schistosomiasis in China. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2016;28:481–484. doi: 10.16250/j.32.1374.2016213. [DOI] [PubMed] [Google Scholar]
- 30.Sun LP, Wang W, Hong QB, Li SZ, Liang YS, Yang HT, et al. Approaches being used in the national schistosomiasis elimination programme in China: a review. Infect Dis Poverty. 2017;6:55. doi: 10.1186/s40249-017-0271-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cao Z, Huang Y, Wang T. Schistosomiasis japonica control in domestic animals: progress and experiences in China. Front Microbiol. 2017;8:2464. doi: 10.3389/fmicb.2017.02464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cai P, Weerakoon KG, Mu Y, Olveda RM, Ross AG, Olveda DU, et al. Comparison of Kato Katz, antibody-based ELISA and droplet digital PCR diagnosis of schistosomiasis japonica: lessons learnt from a setting of low infection intensity. PLoS Negl Trop Dis. 2019;13:e0007228. doi: 10.1371/journal.pntd.0007228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Diab RG, Mady RF, Tolba MM, Ghazala RA. Urinary circulating DNA and circulating antigen for diagnosis of schistosomiasis mansoni: a field study. Trop Med Int Health. 2019;24:371–378. doi: 10.1111/tmi.13193. [DOI] [PubMed] [Google Scholar]
- 34.Kato-Hayashi N, Leonardo LR, Arevalo NL, Tagum MN, Apin J, Agsolid LM, et al. Detection of active schistosome infection by cell-free circulating DNA of Schistosoma japonicum in highly endemic areas in Sorsogon Province, the Philippines. Acta Trop. 2015;141:178–183. doi: 10.1016/j.actatropica.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 35.Weerakoon KG, Gordon CA, Williams GM, Cai P, Gobert GN, Olveda RM, et al. Droplet digital PCR diagnosis of human schistosomiasis: parasite cell-free DNA detection in diverse clinical samples. J Infect Dis. 2017;216:1611–1622. doi: 10.1093/infdis/jix521. [DOI] [PubMed] [Google Scholar]
- 36.Wong FC, Sun K, Jiang P, Cheng YK, Chan KC, Leung TY, et al. Cell-free DNA in maternal plasma and serum: a comparison of quantity, quality and tissue origin using genomic and epigenomic approaches. Clin Biochem. 2016;49:1379–1386. doi: 10.1016/j.clinbiochem.2016.09.009. [DOI] [PubMed] [Google Scholar]
- 37.Barra GB, Santa Rita TH, de Almeida Vasques J, Chianca CF, Nery LF, Santana Soares Costa S. EDTA-mediated inhibition of DNases protects circulating cell-free DNA from ex vivo degradation in blood samples. Clin Biochem. 2015;48:976–81. [DOI] [PubMed]
- 38.Lv C, Fu Z, Lu K, Yue R, Wang T, Cao X, et al. A perspective for improving the sensitivity of detection: the application of multi-epitope recombinant antigen in serological analysis of buffalo schistosomiasis. Acta Trop. 2018;183:14–18. doi: 10.1016/j.actatropica.2018.03.025. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data supporting the conclusions of this article are included within the article.