

Abstract
Arabidopsis plants in their natural environment are susceptible to infection by oomycete pathogens, in particular to downy mildew and white rust diseases. These naturally occurring infectious agents have imposed evolutionary pressures on Arabidopsis populations and are therefore highly relevant for the study of host-pathogen co-evolution. In addition, the study of oomycete diseases, including infections caused by several Phytophthora species, has led to many scientific discoveries on Arabidopsis immunity and disease. Herein, we describe the major oomycete species used for experiments on Arabidopsis, and how these pathosystems have been used to provide significant insights into mechanistic and evolutionary aspects of plant-oomycete interactions. We also highlight understudied aspects of plant-oomycete interactions, as well as translational approaches, that can be productively addressed using the reference pathosystems described in this article.
INTRODUCTION TO OOMYCETES
Oomycetes are filamentous microorganisms that belong to the Stramenopile kingdom (Dick, 2001). Oomycetes are related to diatoms and brown algae and are proposed to have evolved from a photosynthetic ancestor (Tyler, 2006). Although oomycetes are known as “water molds”, many oomycete species are terrestrial and are pathogenic on animals and plants (Jiang and Tyler, 2012). Several oomycete species are among the most notorious plant pathogens and are known for their huge impact on agriculture and natural ecosystems (Kamoun et al., 2015). The potato late blight pathogen Phytophthora infestans, for instance, is the causal agent of the Irish potato famine, and P. ramorum is known for its devastation of oak trees and other plant species in natural ecosystems (Kamoun et al., 2015).
The phylogenetic relations of different genera of oomycete phytopathogens are shown in Figure 1. In the genus Phytophthora (Greek for plant destroyer) more than 100 species have been described that are grouped in 10 clades based on molecular and phenotypic data (Kroon et al., 2012). These pathogens infect a large range of plants on which they have a hemi-biotrophic life style. Initially the infection is biotrophic, during which the host is kept alive and pathogen and plant live in an intimate association. However, at a given colonization level these pathogens switch to a necrotrophic life style that includes the massive production of hydrolytic enzymes and toxins that kill host cells, leading to the typical blight and rot symptoms. In contrast, the downy mildew species have an obligate biotrophic lifestyle and cannot grow in the absence of a living host. These pathogens have often specialized on a single host plant species. More than 700 downy mildew species are known of which the phylogenetic relationships are still mostly unclear (Thines and Choi, 2016). The relatively close evolutionary relationship between downy mildews and Phytophthora species is reflected in them being present in a single order; the Peronosporales.
Figure 1.
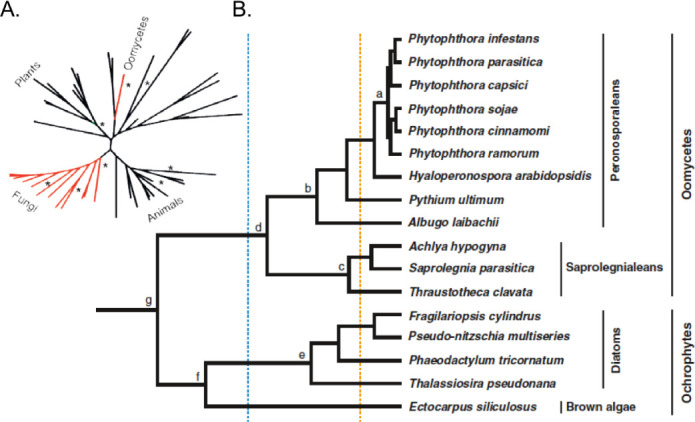
Evolutionary position of oomycetes used in Arabidopsis research.
(A) Phylogeny of oomycetes relative to the major kingdoms of life.
(B) Phylogeny of oomycetes used in Arabidopsis research and their relation to other Stramenopiles. Every species used for Arabidopsis research resides within the Peronosporaleans, within the orders Phytiales (e.g., P. ultimum), Albuginales (e.g., A. laibacii), or Peronosporales ( Phytophthora spp. and H. arabidopsidis).
Two other orders of plant pathogenic oomycetes are outside of the Peronosporales; the Pythiales and Albuginales. Within the Pythiales the genus Pythium contains more than 100 species, most of which inhabit soils or aquatic environments. Pythium species are best known for causing damping-off of seedlings, for example caused by the necrotrophic P. ultimum, while many other Pythium species are considered opportunistic plant pathogens (Lévesque et al., 2010). Albuginales comprise an early diverged lineage of oomycetes that are obligate biotrophs, and contain plant pathogenic Albugo species causing white blister rust, like white rust of Arabidopsis caused by A. laibachii (Thines et al., 2009; Kemen et al., 2011).
OOMYCETES USED IN ARABIDOPSIS RESEARCH
In this section, we summarize experimental attributes of the oomycete species that are utilized for research involving Arabidopsis. We take a comparative approach that emphasizes similarities and differences in the life cycles and infection strategies of each species, along with their relative advantages and challenges as experimental systems. Figure 2 illustrates the life cycles of three species that encompass the diversity of oomycete infection strategies, and Table 1 summarizes key attributes of several long-used oomycete pathogens of Arabidopsis, along with others that have been adopted more recently.
Figure 2.
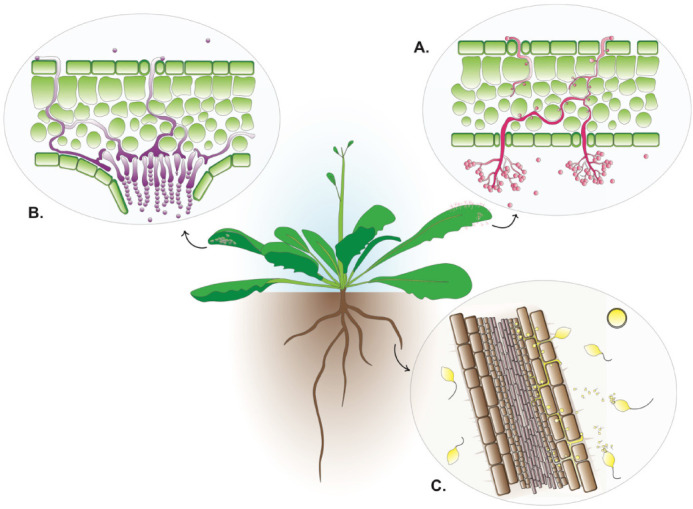
Life cycles of representative species.
(A) Hyaloperonospora arabidopsidis. (B) Albugo candida and A. laibachii. (C) Phytophthora brassicae, P. capsici and P. parasitica.
Table 1.
Attributes of oomycetes used for Arabidopsis research.
| Genus and Species | Natural Host | Lifestyle | Host Organ(s) | Culturable In vitro? | Transformable? | Crossable? | Genome status1,2 |
|---|---|---|---|---|---|---|---|
| Hyaloperonospora arabidopsidis | Arabidopsis thaliana | Obligate biotroph | Leaves | No | No | Yes | 1, 2 |
| Albugo candida | Crucifers | Obligate biotroph | Leaves | No | No | Yes | 1, 2 |
| Albugo laibachii | Arabidopsis thaliana | Obligate biotroph | Leaves | No | No | Yes | 1, 2 |
| Phytophthora brassicae | Brassicaceae | Hemi-biotroph | Leaves and roots | Yes | Yes | Yes | |
| Phytophthora infestans | Potato, tomato | Hemi-biotroph | Leaves and roots | Yes | Yes | Yes | 1, 2 |
| Phytophthora parasitica | Broad range | Hemi-biotroph | Leaves and roots | Yes | Yes | Yes | 1 |
| Phytophthora capsici | Broad range | Hemi-biotroph | Leaves and roots | Yes | Yes | Yes | 1 |
| Pythium ultimum | Broad range | Necrotroph | Roots | Yes | Yes | Yes | 1, 2 |
Assembly can be accessed at 1eumicrobedb.org or 2fungidb.org
Naturally occurring oomycete pathogens of Arabidopsis
In nature, Arabidopsis thaliana is commonly infected by two oomycete pathogen groups; the downy mildew pathogen Hyaloperonospora arabidopsidis (referred to as Hpa hereafter) and two species of the white rust pathogen Albugo (A. laibachii and A. candida). These naturally occurring pathogens have imposed evolutionary pressures on Arabidopsis populations that resulted in a high level of intraspecific variation in resistance (Holub and Beynon, 1996). Many of the underlying resistance genes have been cloned and their downstream signaling pathways have been characterized. Recently, several other eukaryotic microbes have been described as endophytes of Arabidopsis, including representatives of the oomycete genus Pythium (Durán et al., 2018; Sapp et al., 2018). Thus, it is likely that some oomycetes have a close interaction with their Arabidopsis host, but do not cause disease.
Hyaloperonospora arabidopsidis
This species was the first eukaryotic pathogen of Arabidopsis to be documented (Koch and Slusarenko, 1990). It was initially described as Peronospora parasitica (Koch and Slusarenko, 1990), and was later re-named as Hyaloperonospora parasitica (Constantinescu and Fatehi, 2002) and currently Hyaloperonospora arabidopsidis (Voglmayr et al., 2004; Goker et al., 2009). This species is a frequently occurring pathogen in natural Arabidopsis populations (Holub and Beynon, 1996; Holub, 2008). Moreover, the interactions are typified by abundant genetic polymorphism in the host and the pathogen (Holub et al., 1994b). For these reasons, Hpa was adopted as a reference pathogen during the early days of developing Arabidopsis as a system for molecular plant-microbe interactions (Dangl et al., 1992; Crute et al., 1994). These pioneering efforts substantially broadened the impact of Arabidopsis as a model system and are described in detail in several review articles (Holub and Beynon, 1996; Slusarenko and Schlaich, 2003; Coates and Beynon, 2010; McDowell, 2014).
Hpa causes downy mildew disease of Arabidopsis (Koch and Slusarenko, 1990). The disease name comes from the downy appearance of leaves that are covered in sporangiophores (spore-producing structures, Fig. 2A) (Clark and Spencer-Phillips, 2000). Although Hpa does not cause disease on any crop, the closely related species H. parasitica is a serious problem in cabbage and other brassica crops. More distantly related downy mildew species cause important diseases on crops that include grapes, cucurbits, lettuce, spinach, sunflower, and basil (Clark and Spencer-Phillips, 2000). Like other phytopathogenic oomycetes, downy mildew species can quickly overcome host resistance genes and develop resistance to chemical control agents (Lucas et al., 1995). It has been estimated that over 25% of the five billion dollar global fungicide market is spent to control downy mildew diseases (mostly on grape downy mildew) (Gisi, 2002). Thus, Hpa is a reference species for a group of pathogens with significant economic impact.
Like other downy mildew species, Hpa is primarily a foliar pathogen. The asexual (epidemic) phase of the life cycle begins when asexual spores are dispersed by wind or rain to a host plant organ. The spore germinates on the plant surface and produces an infection hypha that typically penetrates the anticlinal wall between epidermal cells (Fig. 2A) (Mims et al., 2004; Soylu et al., 2004). Soon afterwards, the primary hypha establishes lobed feeding structures, called haustoria, that invaginate host cells, most commonly in the mesophyll but also in epidermal cells (Mims et al., 2004; Soylu et al., 2004). Haustoria are of particular interest because they are thought to play a major role in extraction of nutrients from host cells, and in the secretion and translocation of effector proteins into host cells (Spencer-Phillips, 1997; Szabo and Bushnell, 2001; Wang et al., 2017). These aspects of the infection cycle are discussed in greater detail below. Filamentous hyphae grow in the intercellular spaces and branch frequently, forming my-celia that can occupy much of the area within a leaf in only a few days (Soylu and Soylu, 2003). The asexual cycle culminates when hyphae emerge from stomata and differentiate into asexual fruiting bodies, called sporangiophores, on which high numbers of sporangiospores are produced that each typically contain a high number of nuclei (~40/spore) (Koch and Slusarenko, 1990). Hpa can also produce sexual spores, called oospores, when hyphae differentiate into gametogenic antheridia and oogonia. Following meiosis gametes will fuse in the oogonium that will then develop into an oo-spore that contains a single nucleus (Koch and Slusarenko, 1990). Oospores are formed within the plant tissue, are durable and can persist in the soil after the host plant dies and decays. Infection cycles can then be initiated by germinating oospores through infection of an adjacent host plant root, e.g. in the next growing season when the roots of germinated Arabidopsis seeds come into contact with the soil-borne oospores (Holub and Beynon, 1996).
A major aspect of Hpa biology is its strict dependence on a host plant for growth, also known as obligate biotrophy (O’Connell and Panstruga, 2006; McDowell, 2011). Hpa cannot be cultured on synthetic media and appears capable of extracting nutrients only from viable host cells (Lucas et al., 1995). In addition, the host range of Hpa appears to be confined to Arabidopsis. Hpa isolates from Arabidopsis cannot infect related brassica species, and vice versa (Parker et al., 1996). However, some isolates taken from crop brassica species can infect Arabidopsis lines containing mutations that compromise immune responses, indicating that host immunity is an important determinant restricting downy mildew host range (Parker et al., 1996).
Hpa has several advantages and limitations for laboratory experiments (McDowell, 2011). The greatest limitation of this pathogen is its obligate lifestyle: Hpa cannot be propagated apart from the host, which impairs genetic transformation of the pathogen. However, it is relatively straightforward to achieve robust colonization of Arabidopsis by Hpa under laboratory conditions. Key environmental parameters include cool temperatures (~16° C), low light intensity, and high relative humidity (McDowell et al., 2011). Long-term storage of sporangia and oospores at −80 ° C is straightforward and reliable (McDowell et al., 2011). Hpa is homothallic and can self-fertilize, but genetic crosses between isolates can be made, albeit laboriously (Gunn et al., 2002). Genetic mapping populations have been generated in successful support of map-based cloning efforts to identify Hpa avirulence genes, encoding host-recognized effectors (Rehmany et al., 2003; Bailey et al., 2011; Woods-Tor et al., 2018). These and other aspects of lab experiments with Hpa are discussed in detail below and elsewhere (Coates and Beynon, 2010; McDowell, 2011).
A significant advantage of Hpa is that it is a bona fide pathogen of Arabidopsis in the natural world, and has been co-evolving with its host (Holub, 2001). The polymorphisms that have evolved in the host and pathogen were used to molecularly clone Arabidopsis disease resistance genes (“R genes”) and the corresponding effector/avirulence genes from the pathogen (Table 2). Recent studies have demonstrated additional natural variation in effector gene repertoires in Hpa, and in the responses of different Arabidopsis accessions to Hpa effectors, suggesting functional variability in the Arabidopsis proteins that are targeted by Hpa effectors (Fabro et al., 2011; Asai et al., 2014). Thus, natural genetic variation in the Arabidopsis/Hpa interaction can be further exploited for mechanistic insight into plant-oomycete interaction and co-evolution. Furthermore, because of the experimental advantages of Arabidopsis, Hpa is also one of the best reference organisms for investigating obligate biotrophy (McDowell, 2011). A final advantage of Hpa is the availability of a reference genome sequence (Baxter et al., 2010).
Table 2.
Features of cloned Arabidopsis resistance genes encoding NLRs mediating detection of the oomycetes Hpa (RPP) and Albugo (RAC, WRR).
| Gene | Accession | Col-0 gene/allele | Remark | Reference | Recognizing | References |
|---|---|---|---|---|---|---|
| RPP1-WsA | Ws | At3g44480* (rpp1) | Original RPP1 locus | Botella et al., 1998 | unknown | |
| RPP1-WsB | Ws | At3g44480* (rpp1) | Previous RPP10 locus | Botella et al., 1998 | ATR1 alleles | Rehmany et al., 2005 |
| RPP1-WsC | Ws | At3g44480* (rpp1) | Previous RPP14 locus | Botella et al., 1998 | unknown | |
| RPP1-NdA | Nd | At3g44480* (rpp1) | Similar but different specificity as RPP-WsB | Botella et al., 1998 | ATR1 alleles | Rehmany et al., 2005 |
| RPP1-EstA | Est | At3g44480* (rpp1) | Similar but different specificity as RPP-WsB | Goritschnig et al., 2016 | WY motif 2 of ATR1 alleles | Goritschnig et al., 2016 |
| RPP1-ZdrA | Zdr | At3g44480* (rpp1) | Similar but different specificity as RPP-WsB | Goritschnig et al., 2016 | WY motif 2 of ATR1 alleles | Goritschnig et al., 2016 |
| RPP2A | Col-0 | At4g19500 | Requires RPP2B | Sinapidou et al., 2004 | unknown | |
| RPP2B | Col-0 | At4g19510 | Requires RPP2A | Sinapidou et al., 2002 | unknown | |
| RPP4 | Col-0 | At4g16860 | Van der Biezen et al., 2004 | ATR4 | Asai et al., 2018 | |
| RPP5 | Ler | At4g16950, paralog of RPP4 | Parker et al., 1997 | ATR5 | Bailey et al., 2011 | |
| RPP7 | Col-0 | At1g58602 | Eulgem et al., 2007 | unknown | ||
| RPP8-Ler | Ler | At5g43470 (rpp8) | Alleles confer virus resistance | McDowell et al., 1998 | ATR8#, not cloned | Gunn et al. 2002 |
| RPP13-Nd | Nd | At3g46530 | Highly polymorphic inA. thaliana (Rose et al., 2004) | Bittner-Eddy & Beynon, 2001 | ATR13 | Allen et al., 2004 |
| RPP39 | Wei-0 | At1g61180 & At1g61190 | CC-NB-LRR protein signaling through NDR1 | Goritschnig et al., 2012 | ATR39 | Goritschnig et al., 2012 |
| RAC1 | Ksk-1 | At1g31540 | Resistance to Albugo laibachii Nc14 | Borhan et al., 2004 | unknown | |
| WRR4 | Col-0 | At1g56510 | Confers resistance to multiple A. candida races | Borhan et al., 2008 | unknown | |
| WRR4B | Ws-2 | At1g56540 | Resistance to A. candida isolate Ac2V | Cevik et al., 2019 | unknown | |
| WRR8 | Sf-2 | At5g46270 | Resistance to A. candida isolate Ac2V | Cevik et al., 2019 | unknown | |
| WRR9 | Hi-0 | At1g63750 | Resistance to A. candida isolate Ac2V | Cevik et al., 2019 | unknown | |
| WRR12 | Ler | At1g17600 | Resistance to A. candida race AcBoT | Borhan et al., 2008 | unknown |
RPP1 orthologs or family members in Col-0 are: At3g44000, At3g44480, At3g44630, and At3g44670
identified as a segregating locus in Hpa crosses
Albugo candida and Albugo laibachii
White blister pathogens belonging to the genus Albugo are naturally occurring, obligate biotrophic pathogens of Arabidopsis, and their attributes will be summarized in comparison with Hpa. Like Hpa, Albugo is commonly found on Arabidopsis in the wild, and their interactions display natural variability, including gene-for-gene interactions (Holub et al., 1994a; Holub and Beynon, 1996; Borhan et al., 2001; Holub, 2008). In addition, Albugo species cause important crop diseases on cultivated Brassica species (Saharan et al., 2014).
Despite the superficial similarities between Hpa and Albugo species, several interesting differences are apparent: First, the infection cycle of Albugo is very different, as illustrated in Fig. 2. For example, Albugo species produce motile zoospores that deploy germ tubes to enter through the stomata (Soylu et al., 2003). The hallmark of the disease cycle is blisters that are caused by rupture of the plant epidermis to facilitate dissemination of asexual spores (Saharan et al., 2014).
Molecular phylogenies clearly illustrate that the Albugo and downy mildew lineages are distinct (Fig. 1) (Thines, 2014; Ascunce et al., 2017). Thus, Albugo and Hpa lineages have independently evolved to an obligate lifestyle and to compatibility with Arabidopsis (Kemen and Jones, 2012). Interestingly, A. laibachii and A. candida display different host ranges: the former, like Hpa, is restricted to A. thaliana, while host range of A. candida encompasses 63 genera and 241 species within the Brassicaceae (Thines et al., 2009). Thus, these two species provide an opportunity for comparative studies to understand the factors that control host range (McMullan et al., 2015; Jouet et al., 2018). As with Hpa, intraspecific variation for resistance/susceptibility to Albugo is prevalent in Arabidopsis and has been used to clone several Arabidopsis genes for resistance to Albugo (Table 2). Perhaps most interestingly, Albugo species display a remarkable capacity to suppress defense in Arabidopsis (Cooper et al., 2008). Co-infection with Albugo can render Arabidopsis susceptible to otherwise incompatible pathogens, including the late blight pathogen Phytophthora infestans (Belhaj et al., 2017; Prince et al., 2017). The molecular mechanisms by which Albugo strongly suppresses plant immunity are beginning to be revealed (Prince et al., 2017). Further studies are expected to reveal Albugo effectors and their plant targets that are likely to be key components of the plant immune system (Kemen et al., 2011; Links et al., 2011). Albugo-Arabidopsis systems are also being developed to better understand how Albugo influences plant interactions with other microbes (Ruhe et al., 2016; Jouet et al., 2018), as well as the important phenomenon of non-host resistance (Cevik et al., 2019).
The experimental advantages and limitations of the Albugo species are similar to Hpa. Like Hpa, Albugo species cannot be cultured apart from their hosts but they are relatively straightforward to propagate on susceptible host genotypes (Crute et al., 1993). Long-term storage is possible and genetic crosses between isolates can be performed (Adhikari et al., 2003). Importantly, draft genome sequences are available from two different strains of A. laibachii and one of A. candida (Kemen et al., 2011; Links et al., 2011). As with Hpa, the genome data is having a major impact on research with this organism (Kemen and Jones, 2012).
Laboratory pathosystems
Diseases of Arabidopsis caused by other oomycetes, for instance by Phytophthora or Pythium species, have not been observed under natural conditions, but have been achieved under laboratory conditions. Complete infection cycles have been obtained on Arabidopsis using the Phytophthora species P. brassicae (Roetschi et al., 2001), P. parasitica (Attard et al., 2010; Wang et al., 2011), P. capsici (Wang et al., 2013a), P. palmivora (Daniel and Guest, 2006), and P. cinnamomi (Robinson and Cahill, 2003). Strikingly, different immune signaling pathways seem to be involved in resistance responses to each of these Phytophthora species (Wang et al., 2013). Within Arabidopsis, different accessions show a range of infection phenotypes, from clearly compatible (resulting in disease) to fully incompatible (resistant). Other Phytophthora species tested, e.g., P. infestans (Huitema et al., 2003) and P. sojae (Takemoto et al., 2003), are not able to establish a successful infection on Arabidopsis but are effectively stopped by the plant, likely by early perception by the host immune system. These laboratory pathosystems provide valuable complements to the pathosystems based on Hpa and Albugo and are described in the following sections.
Phytophthora species
P. brassicae (formerly P. porri) is the first representative of the Phytophthora genus to be used in Arabidopsis research (Roetschi et al., 2001). This species parasitizes brassica species closely related to Arabidopsis. Given the close relationship between these hosts and Arabidopsis, it is not surprising that P. brassicae can grow on Arabidopsis. The initial stages of the asexual infection cycle are similar to those of Hpa: spores attach to the organ surface with appressoria and penetrate by growing between epidermal cells (Figure 2C). Hyphae ramify through intercellular spaces and project haustoria into plant cells. After approximately 3 dpi, a necrotrophic program commences in which the pathogen destroys host tissue. This hemi-biotrophic infection strategy is a major difference from obligate biotrophs like Hpa and Albugo species, which cannot thrive on dead plant tissue.
P. brassicae was one of the first Phytophthora species used to test immune responses of various Arabidopsis ecotypes and mutants (Mauch et al., 2008). Arabidopsis researchers have subsequently made use of “non-adapted” oomycete pathogens; that is, species that are incapable of causing disease on wild-type Arabidopsis even under the most permissive conditions. The most productive insights from this approach have involved the infamous species Phytophthora infestans, which causes late blight disease of potato and tomato and is responsible for the famine in Ireland in the 19th century (Fry, 2008). Despite its destructive effects on potato and tomato, P. infestans is stopped quickly in Arabidopsis by multiple, genetically redundant layers of resistance, including so-called “penetration resistance” (Stein et al., 2006). Thus, this pathogen has been used as a probe to dissect the cytological responses and genetic basis of this resistance (see below).
Two recently developed Phytophthora pathosystems are based on P. capsici and P. parasitica. Both of these species exhibit broad host ranges. P. capsici is destructive on cucurbit, legumes, and solanaceous crops (Kamoun et al., 2015; Barchenger et al., 2018). P. parasitica (also called P. nicotianae) causes a variety of diseases on field crops, fruit trees, and ornamental plants (Meng et al., 2014; Kamoun et al., 2015; Meng et al., 2015; Panabieres et al., 2016). Testing of Arabidopsis accessions with different strains of both species revealed natural genetic variation for resistance/susceptibility (Attard et al., 2010; Wang et al., 2011; Wang et al., 2013). P. capsici and P. parasitica can complete their life cycles on leaves or roots of susceptible Arabidopsis accessions. Systems for infection through Arabidopsis roots have been developed and provide a complement to foliar pathogens like Hpa or Albugo (Attard et al., 2010; Wang et al., 2011; Wang et al., 2013; Hou and Ma, 2017). Both species employ a hemi-biotrophic life cycle that is depicted in the root in Figure 2C.
Although only recently developed, these Phytophthora patho-systems hold considerable potential to provide complementary insights to pathosystems based on Hpa or Albugo species. Most importantly, the experimental tools on the pathogen side are relatively advanced. Both species are easy to culture and relatively straightforward to transform, enabling reverse genetics on the pathogen (Le Berre et al., 2008; Wang et al., 2018b). It is also possible to perform genetic crosses and develop segregating populations (Lamour et al., 2007; Lamour et al., 2014). Additionally, genome sequences for both species are available to delineate genes important for pathogenicity (Lamour et al., 2012) (note that the P. parasitica genome data has not been formally published but is publicly available at fungidb.org and eumicrobedb.org). Moreover, successful genome editing by the CRISPR/Cas9 system has now been reported for P. capsici (Wang et al., 2018b). Considering these experimental advantages and the practical value of research involving these emerging, broad host range pathogens, we expect that P. capsici and P. parasitica will become increasingly important for exploiting the experimental advantages of Arabidopsis to understand how diseases are caused by oomycetes.
Pythium species
Members of the Pythium genus cause a number of destructive “damping-off” diseases (Martin and Loper, 1999). Contrasting with downy mildews and Phytophthora, many Pythium species are classical necrotrophs that lack a biotrophic phase during infection (Kamoun et al., 2015). These pathogens exhibit filamentous growth but do not produce haustoria and induce host cell death soon after contact.
Pythium irregulare has been used as a model soilborne pathogen of Arabidopsis since the late 1990s. Bioassays with this pathogen have been useful in mutants with reduced immune responses, particular those in jasmonate- and ethylene-mediated defense (Browse et al., 1998; Staswick et al., 1998; Geraats et al., 2002; Huffaker et al., 2006; Huffaker and Ryan, 2007). Contrastingly, research on Pythium pathogenicity, using Arabidopsis, is practically non-existent. The tools on the pathogen side are reasonably good: Genome sequences from several species have been completed, the pathogen is easy to culture and is transformable (Levesque et al., 2010; Adhikari et al., 2013; Grenville-Briggs et al., 2013). Thus, there is good opportunity to exploit Arabidopsis-Pythium pathosystems to better understand how necrotrophic oomycetes subjugate their hosts. In addition, recent studies of the Arabidopsis root microbiome revealed Pythium species as prominent endophytes, opening a new area of Arabidopsis-oomycete interactions (Durán et al., 2018; Sapp et al., 2018).
PATHOGEN PERCEPTION
Introduction to the plant immune system
Plants have evolved various ways to fend off microbes (Pieterse et al., 2009). A suite of surface-exposed detectors, named pattern recognition receptors (PRRs, Fig. 3), can recognize conserved microbe-associated molecular patterns (MAMPs), such as bacterial flagellin or fungal chitin. Upon detection the PRRs activate a signaling cascade that leads to pattern-triggered immunity (PTI). Most PRRs require a co-receptor to initiate signaling: for example, the receptor FLS2 interacts with the BAK1 co-receptor upon recognition of the MAMP flg22 (see (Macho and Zipfel, 2014) for details on the mechanisms of immune activation by PRRs). BAK1 also interacts with other PRRs (Macho and Zipfel, 2014). PTI signaling via mitogen-activated protein kinase (MAPK) cascades and/or calcium-dependent protein kinases (CDPKs) (Bredow and Monaghan, 2019) activates pathogen-nonspecific immune responses such as the production of reactive oxygen species and nitric oxide, cell wall reinforcement, and induction of defense genes (Hein et al., 2009; Nürnberger and Kemmerling, 2009). Adapted pathogens are able to suppress PTI with effectors that have evolved to interact directly with host defense-associated proteins, resulting in effector-triggered susceptibility (Jones and Dangl, 2006). For example, bacterial effectors can interfere directly with FLS2 or BAK1 (Toruno et al., 2016). Intracellular nucleotide-binding, leucine-rich repeat (NLR, Fig. 3) receptors that recognize these effectors or their activity have evolved in plants as a second layer of pathogen perception. NLR proteins are encoded by plant disease resistance genes and mediate so-called Effector-Triggered Immunity (ETI). This is the molecular basis of the genetic model of “gene-for-gene” resistance, in which pathogen “avirulence (Avr) genes” (now known to encode secreted effector proteins) are recognized inside plant cells by plant “resistance (R) genes” (now known to encode NLR proteins). Resistance to adapted pathogen species is often rapidly broken by loss or mutation of avirulence effector genes or through suppression of ETI by different effectors (Woods-Tor et al., 2018).
Figure 3.
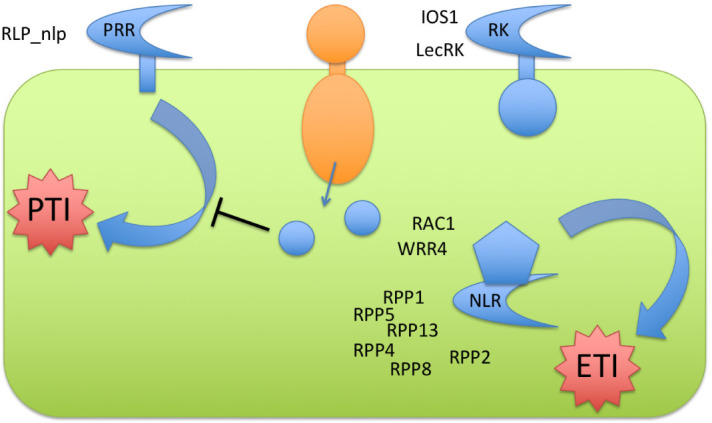
Schematic of PTI and ETI against oomycetes
Pattern recognition receptors
The majority of plant PRRs identified so far belongs to the family of leucine-rich repeat receptor-like kinases (LRR-RLKs), which has more than 600 members in Arabidopsis (Shiu et al., 2004), or receptor-like protein (LRR-RLPs) that lack a C-terminal kinase domain and interact with RLKs for transducing signals (Gust and Felix, 2014). The best-described plant PRR is FLS2 from Arabidopsis that recognizes bacterial flagellin, a major structural protein of bacterial flagella. An N-terminal 22-amino acid residue fragment of this protein (flg22) is sufficient to be recognized by FLS2 and to induce PTI (Gómez-Gómez and Boller, 2000). Naito et al. (2008) showed that these residues are also essential for flagellum function and motility of bacteria, which makes it difficult for bacteria to circumvent this recognition by evolving different amino acid sequences in this region of the protein. A second well-described plant PRR that seems restricted to Brassicaceae is EFR, which recognizes the bacterial elongation factor Tu (EF-Tu) or its shorter N-terminal fragment elf18 (Kunze, 2004). When this PRR is expressed in the solanaceous plants Nicotiana benthamiana and tomato (Solanum lycopersicum), it confers resistance to a broad spectrum of bacteria (Lacombe et al., 2010), illustrating the potential use of PRRs as new sources for genetic resistance to crop diseases.
Fungi can also be recognized by PRRs, e.g. through the Arabidopsis RLK CERK1 that is required for responses to fungal chitin and bacterial peptidoglycan (Miya et al., 2007; Willmann et al., 2011), and the RLP30 PRR that mediates recognition of necrotrophic fungi through a currently unknown proteinaceous MAMP (Zhang et al., 2013). As we will detail in the next section, only a few receptors have been identified so far that mediate the extracellular recognition of oomycete MAMPs.
Oomycete pattern-triggered immunity
The extracellular recognition of oomycetes triggers the first line of defense that plant cells deploy (Raaymakers and Van den Ackerveken, 2016). However, only two PRRs recognizing oomycete MAMPs have been identified to date, the receptor-like proteins ELR of potato and RLP23 of Arabidopsis. ELR mediates the recognition of elicitins, secreted oomycete proteins that have a putative function as extracellular sterol carriers (Derevnina et al., 2016). Elicitins are known to act as MAMPs in certain plant species, e.g. in Solanum microdontum in which it induces a cell death response. ELR was cloned from this wild potato species and found to encode a receptor-like protein that requires the RLK BAK1 to induce elicitin-triggered immunity. The second PRR identified against oomycetes is Arabidopsis RLP23. This protein recognizes Nep1 (Necrosis- and Ethylene-inducing Protein 1)-like proteins (NLPs), secreted proteins that are found in many plant-associated microbes that belong to three different kingdoms of life (Oome et al., 2014). RLP23 was found to require the co-receptors SOBIR1 and BAK1 for NLP-triggered immunity (Albert et al., 2015). NLP genes are found in oomycetes belonging to the Peronosporales and also in fungi and bacteria. In oomycetes both cytotoxic and non-cytotoxic NLPs have been found, but both types act as MAMPs in Arabidopsis (Oome et al., 2014; Böhm et al., 2014). The immunogenic fragment of NLPs could be reduced to a synthetic peptide of <20 amino acids that was shown to bind the RLP23 receptor (Albert et al., 2015).
Other molecules of oomycetes have been proposed to act as MAMPs (Hein et al., 2009): (i) glucans derived from oomycete cell wall polysaccharides that bind to a soybean glucan-binding protein that has glucanase activity, but is insufficient to trigger immunity (Glycine max) (Fliegmann et al., 2004), (ii) the 13-amino acid fragment Pep-13 derived from GP42, a calcium-dependent transglutaminase that is abundant in the cell wall of Phytophthora sojae, and is sufficient to elicit PTI responses in parsley (Brunner et al., 2002), and (iii) the Phytophthora cellulose-binding elicitor lectin (CBEL) that elicits a response in tobacco and Arabidopsis (Hein et al., 2009; Khatib et al., 2004). (iv) The glycoside hydrolase 12 protein XEG1 from Phytophthora sojae, recently shown to be recognized by a receptor complex in N. benthamiana that includes an extracellular LRR receptor, BAK1, and SOBIR1 (Ma et al., 2015; Wang et al., 2018a). Other groups of cell death-inducing or cytotoxic proteins from oomycetes, such as the Crinkler proteins, might also qualify as MAMPs based on their widespread occurrence among different pathogens (Thomma et al., 2011). We only know the tip of the iceberg with respect to oomycete MAMPs and we expect that many new patterns and their cognate receptors will be identified in the coming years. Finally, it is likely that oomycete enzymes release compounds from the host cell wall and membrane that can act as damage-associated molecular patterns (DAMPs). It was proposed that Hpa has evolved to circumvent the release of DAMPs by strong reduction of the number of genes encoding for hydrolytic enzymes compared to Phytophthora species (Baxter et al., 2010). Further exploitation of Arabidopsis to explore these aspects will undoubtedly be productive.
Intracellular perception of oomycetes in Arabidopsis
Plants have evolved intracellular receptors with nucleotide-binding, leucine-rich repeat domains (NLRs) to detect oomycete effectors, or their activity, to trigger immunity (ETI). NLR proteins typically have an N-terminal variable region, either of the coiled-coil type (CC) or Toll/Interleukin-1 receptor-like (TIR), or an RPW8-like domain. The N-terminal domains are followed by a nucleotide binding domain (NB) and C-terminal leucine-rich repeat region (LRR), further detailed in an excellent review (Cui et al., 2015).
The genes encoding these intracellular receptors were first identified in Arabidopsis through genetic mapping of resistance genes. The Arabidopsis genome (Col-0) encodes a total of around 150 NLR proteins, of which only a limited number have an assigned function. The first cloned NLR gene was RPS2 that confers resistance to Pseudomonas bacteria expressing the effector AvrRpt2 (Mindrinos et al., 1994). Soon after that also R genes for resistance to downy mildew and Albugo were cloned (Table 2), based on natural variability of their functions in different accessions of Arabidopsis (Slusarenko and Schlaich, 2003; Holub, 2008; Coates and Beynon, 2010). These included the first R genes to be cloned against oomycetes (Parker et al., 1997; Botella et al., 1998; McDowell et al., 1998). More than 25 R loci conferring isolate-specific resistance to Hpa have been genetically identified in Arabidopsis and are named RPP, for RECOGNITION OF PERONOSPORA PARASITICA, the older name of this pathogen (Slusarenko and Schlaich, 2003). So far, 8 RPP genes have been cloned: RPP1, RPP2, RPP4, RPP5, RPP7, RPP8, RPP13, and RPP39 and all encode cytoplasmic NLRs. Alleles of RPP1 confer different isolate-specific resistances (Botella et al., 1998), and also RPP4 and RPP5 can be considered allelic variants (Van der Biezen et al., 2002). Many of the other RPP loci map to the position of cloned RPP genes and could constitute allelic variants (Nemri et al., 2010). Broad resistance to downy mildew, observed in certain Arabidopsis accessions, appeared to be mediated by combinations of isolate-specific resistance loci (Lapin et al., 2012). Recently, the “helper” NLR proteins ADR1 and NRG1 were identified as signaling components for RPP proteins containing the TIR domain (Bonardi et al., 2011; Castel et al., 2019; Lapin et al., 2019; Wu et al., 2019), It is becoming clear that ETI can be underpinned by networks of NLRs, and deconvolution of these networks is an emerging area of interest (Wu et al., 2018).
At a broad scale, little is known about whether and how the activity of RPP genes is regulated (Lai and Eulgem, 2018). In one case, feedback control involving salicylic acid and WRKY proteins has been implicated (Mohr et al., 2010). In another, gene activity is subject to post-transcriptional control involving alternative polyadenylation, which is regulated by histone marks (Tsuchiya and Eulgem, 2013; Lai et al., 2018). This research has opened up a new avenue towards understanding the mechanisms and evolution of NLR gene regulation (McDowell and Meyers, 2013). Another recent study identified eQTLs providing resistance against Hpa, emphasizing the potential of this pathosystem to understand epigenetic factors that are relevant to plant-oomycete interactions (Furci et al., 2019).
Several avirulence genes from Hpa have also been cloned (Table 2), thanks to the development of genetic and genomic tools for the pathogen (Coates and Beynon, 2010; McDowell, 2014). In fact, the first oomycete Avr gene to be cloned was ATR13 from Hpa (Allen et al., 2004). This gene encodes a small, secreted protein with a signal peptide followed by the conserved protein motif Arg-X-Leu-Arg, first described in (Rehmany et al., 2005). Downy mildew pathogens and Phytophthora species contain hundreds of putative “RXLR genes” in their genomes, and it is clear that they play an important role in pathogen virulence (discussed extensively below). RXLR proteins are secreted from pathogen haustoria and enter the interior of plant cells to reprogram plant regulatory networks and thereby promote virulence. However, RXLR proteins can also be recognized inside plants cells by NLR surveillance proteins. Indeed, every Avr gene cloned from a downy mildew or Phytophthora species to date encodes an RXLR or RXLR-like protein, recognized by the corresponding plant NLR protein. To date, Hpa Avr proteins recognized by RPP1, RPP13, RPP4, RPP5, and RPP39 have been molecularly identified, and the determinants of their virulence and avirulence functions are under investigation (Allen et al., 2004; Rose et al., 2004; Rehmany et al., 2005; Bailey et al., 2011; Chou et al., 2011; Krasileva et al., 2011; Leonelli et al., 2011; Goritschnig et al., 2012; Steinbrenner et al., 2015).
Comparative and molecular studies of RPP genes have provided novel insights into the molecular mechanisms and selective forces that drive NLR gene evolution (McDowell and Simon, 2006). For example, RPP5, RPP1, and RPP8 multigene families are physically linked in clusters and are subject to intra- and intergenic recombination, to produce new NLR gene variants in host-pathogen arms race (Botella et al., 1998; McDowell et al., 1998; Noel et al., 1999). RPP1 homologs have diversified through repeat duplication and sequence divergence, such that they can detect multiple surfaces of the corresponding effector (Goritschnig et al., 2016). ATR1 is a modular protein that can evolve to escape detection by mutations in any of several surfaces that mediate recognition by RPP1 (Chou et al., 2011). RPP13 provides an example of a simple locus, in which a single copy gene displays substantial allelic polymorphism, driven by diversifying selection and intra-allelic recombination (Bittner-Eddy and Beynon, 2001). Interestingly, the Hpa ATR13 locus, encoding the RXLR effector protein recognized by RPP13, displays similar attributes, suggesting that co-evolution has been a major driver for this diversity, along with balancing selection to maintain repertoires of useful alleles in the plant and pathogen populations (Allen et al., 2004). The ATR13 protein displays a novel structure and polymorphisms that mediate recognition specificity by RPP13 map to a single, surface exposed region (Leonelli et al., 2011). Much remains to be learned about plant-oomycete coevolution, and we expect a resurgence of this topic to be driven by our new capacity for cost-effective populations genomics of plants and pathogens sampled from natural populations along with sophisticated genetics (Karasov et al., 2018; Woods-Tor et al., 2018).
In the years following RPP gene cloning, only the RPP1 protein has been investigated in significant mechanistic detail. This protein binds directly to ATR1 via the LRRs. This interaction displays extensive allelic variability (Krasileva et al., 2010; Steinbrenner et al., 2015). In the absence of the ATR1 effector, RPP1 is likely maintained in an inactive state by intramolecular interactions between the N-terminal TIR domain, the NB domain, and the LRRs. Binding of ATR1 via the LRRs disrupts these interactions and permits oligomerization of RPP1, thereby triggering cell death and other immune responses (Krasileva et al., 2010; Steinbrenner et al., 2015). This mode of inactivation/activation appears to be a general aspect of NLR proteins, although details vary between different NLR proteins. As with other NLR proteins, the molecular events between RPP1 activation and deployment of the ultimate cellular immune response remain to be identified. This is a major knowledge gap that extends across all plant-pathogen interactions (Cui et al., 2015) and could be productively addressed by further studies of RPP proteins.
Several Arabidopsis genes for resistance to Albugo have also been cloned (Table 2). The RAC1 gene provides isolate-specific resistance to Albugo laibachii Nc14 (Borhan et al., 2004)(Cevik et al., 2019). WRR4 confers broad spectrum resistance to many Albugo candida isolates and functions as a transgene in Brassica juncea (Borhan et al., 2008 (2010). These R genes encode cytoplasmic NLRs that act similar to many of the RPP proteins, and helper NLRs have also been implicated for WRR-mediated resistance (Castel et al., 2019). Interestingly, the phenomenon of non-host resistance of Arabidopsis to Brassica-infecting races of Albugo candida is mechanistically underpinned by NLR receptors that can be identified by segregation analysis (Cevik et al., 2019). This led to the molecular cloning of WRR4B, WRR8, WRR9 and WRR12. Some of these genes were shown to provide resistance as transgenes in Brassica species (Cevik et al., 2019). Avirulence genes from Albugo remain to be identified, but this task is now feasible thanks to the genomic resources that exist for this pathogen. Considering that RXLR genes do not appear prominently in the Albugo genome (see below) it will be very interesting to compare the structure and function of Avr genes from Albugo with those from Hpa.
IMMUNE SIGNALING AND RESPONSES
Following the perception of pathogens, signaling is initiated that ultimately results in the execution of a broad range of defense responses that stop the invading microorganism. Here, we summarize important signal transduction components and defense responses against oomycete infection of Arabidopsis.
In Arabidopsis, many genetic screens have been performed to identify genes that are important for defense signaling. Some of these screens were focused on signaling downstream of specific R proteins, while others were aimed to identify more general signal transduction components. It is beyond the scope of this chapter to describe the large number of genes involved in immune signaling, which are discussed in a number of excellent reviews (Pieterse et al., 2012; Cui et al., 2015). We focus on important plant proteins, EDS1 and NPR1, that play crucial roles in the interaction with oomycetes and are well studied in Arabidopsis. EDS1 and NPR1 are closely linked to the immunity-related hormone salicylic acid (SA) that is also key for resistance to oomycetes.
EDS1 (ENHANCED DISEASE SUSCEPTIBILITY 1) is a lipase-like protein that is required for the function of a subgroup of NLRs, those with a Toll–interleukin 1 receptor domain called TIR-NLR. In addition, EDS1 has a function in oxidative stress responses and as a positive regulator in basal resistance to virulent pathogens making the eds1 mutant enhanced disease-susceptible (Wiermer et al., 2005). Two sequence-related signaling partners, PAD4 (PHYTOALEXIN-DEFICIENT 4) and SAG101 (SENESCENCE-ASSOCIATED GENE 101), form complexes with EDS1 and have signaling functions in the cytoplasm and nucleus. The NLR helper protein NRG1 also resides in this complex and plays a role in cell death signaling (Lapin et al., 2019). Interestingly, EDS1 was found to associate with AvrRps4, a Pseudomonas effector, and to interact with TNLs, like RPS4 that mediate recognition of AvrRps4. It was hypothesized that EDS1, being important for basal resistance, could be a target for pathogen effectors and thus serve as a molecular bridge to mediate recognition by R proteins that guard the EDS1 protein in addition to its signaling functions (Cui et al., 2015). This hypothesis is currently debated and under further investigation (e.g., (Huh et al., 2017; Halane et al., 2018). The eds1 mutant was originally identified in a screen for loss of resistance to downy mildew (Parker et al., 1996). EDS1 was shown to be required for the function of TIR-NLR genes such as RPP1, RPP4, RPP5, and WRR4. Virulent strains of Hpa and Albugo are also able to cause more severe infections on eds1 mutant plants because of the reduced level of basal resistance (Wiermer et al., 2005). Arabidopsis eds1 mutants are also enhanced susceptible to a number of Phytophthora species, such as P. capsici (Wang et al., 2013a) and P. parasitica (Attard et al., 2010). The lowered basal immunity is associated with reduced production of the defense hormone SA, that is essential for defense against biotrophic pathogens (Vlot et al., 2009). EDS1 positively regulates SA accumulation, acting upstream of SA, but the EDS1 gene is also activated at the transcriptional level by SA. EDS1 thus seems to be part of an SA-associated positive feedback loop of plant defense (Feys et al., 2001).
SA is not only crucial for resistance to downy mildew, but also affects basal and R gene-mediated resistance to many other oomycetes, such as P. capsici (Wang et al., 2013), or P. parasitica (Attard et al., 2010). Exogenous application of the hormone SA is sufficient to trigger efficient defense responses to a broad range of pathogens, including many oomycete pathogens, and in particular to downy mildew and hemi-biotrophic Phytophthora species. SA (2-hydroxybenzoic acid) is synthesized via two main routes, the isochorismate (IC) and phenylalanine ammonia lyase (PAL) route (Dempsey et al., 2011). The Arabidopsis IC route goes via two IC synthases (ICS) and downstream steps that differ from the canonical pathway defined in bacteria (Rekhter et al., 2019b; Torrens-Spence et al., 2019). The ics1 (sid2) mutant has received the majority of experimental attention to date. Ics1 mutants exhibit 90% lower levels of SA (Wildermuth et al., 2001), and levels are ~95% lower in the ics1 ics2 double mutant following powdery mildew infection (Garcion et al., 2008). The ICS1 gene is induced by MAMPs and by NLR-mediated pathogen recognition (Mishina and Zeier, 2007; Cui et al., 2015) leading to high local levels of SA that activate defense. Besides local defenses, SA plays an important role, together with pipecolic acid, in systemic acquired resistance (SAR), that enhances immunity of distant plant tissues to protect against future infections (Bernsdorff et al., 2016; Klessig et al., 2018). Recently the export protein EDS5 was implicated in synthesis of pipecolic acid, in addition to its well-known role in SA biosynthesis, revealing a surprising instance of immune convergence (Rekhter et al., 2019a).
Downstream of SA, the central NPR1 (NONEXPRESSOR of PR GENES 1) protein plays a crucial role in the transcriptional activation of SA-induced defense in Arabidopsis (Pieterse and Van Loon, 2004). Mutation of NPR1 leads to loss of basal resistance to (hemi-)biotrophic pathogens and of SAR (Cao et al., 1997). NPR1 proteins form a multimeric complex in the plant cell cytoplasm. Upon triggering of the plant immune system SA accumulates and subsequently elicits the thioredoxin-mediated reduction of a cysteine residue in NPR1 (Yan and Dong, 2014). This results in the release of monomeric NPR1 that can travel into the nucleus where it interacts with TGA transcription factors to activate transcription of defense genes, like pathogenesis-related (PR) genes (Boyle et al., 2009). NPR1 and the two paralogous Arabidopsis proteins NPR3 and NPR4 have been shown to bind SA and could thus constitute the SA immune receptors (Ding et al., 2018; Innes, 2018).
Interestingly, RPP proteins vary significantly in their requirements for functionality of these signaling components. For example, resistance mediated by the TIR-NLR protein RPP5 is strongly compromised by mutations that nullify EDS1, NPR1, or SA bio-synthesis (van der Biezen et al., 2002). On the other hand, the CC-NLR proteins RPP7 and RPP13 retain almost full functionality in these backgrounds (McDowell et al., 2000; Bittner-Eddy and Beynon, 2001). These differences underscore the complexity of ETI regulation.
Other defense related hormones, in particular jasmonic acid (JA) and ethylene, play important roles in immunity to Pythium infection. Enhanced susceptibility to necrotizing Pythium species was observed in Arabidopsis mutants with reduced JA levels of responses; (i) in the JA nonaccumulating fad3-2_fad7-2_fad8 triple mutant (Browse et al., 1998), and (ii) in the JA-insensitive jar1-1 mutant (Staswick et al., 1998). Also, the ethylene-insensitive ein2-1 mutant, but not the partially-ethylene insensitive mutant etr1-1, showed enhanced susceptibility to Pythium infection (Geraats et al., 2002). Interestingly, small endogenous peptides encoded by the PROPEP1, 2, and 3 genes, function in a feedback loop amplifying JA and ethylene defense signaling pathways initiated by Pythium (Huffaker and Ryan, 2007). Consequently, Arabidopsis transgenics overexpressing PROPEP1 show enhanced resistance to Pythium irregulare (Huffaker et al., 2006).
It is well known that many proteins are specifically produced during plant defense (van Loon et al., 2006). In particular, many secreted and vacuolar proteins have been classified as pathogenesis-related proteins. Also in Arabidopsis many PR proteins have been identified, e.g. PR1, PR2, and PR5 that were found in apoplastic fluid of Arabidopsis treated with 2,6-dichloroisonicotinic acid (INA), which is a mimic of SA (Uknes et al., 1992). Several of these PR proteins have anti-microbial activities, while others contribute to resistance in unknown ways. One of the most used marker genes of SA-induced plant defense is PR1. Plants have multiple PR1 genes encoding different protein variants with different activities (van Loon et al., 2006). Several tobacco PR1 proteins have anti-microbial activity on the oomycete P. infestans. Spore germination and pathogen growth in planta was effectively inhibited by PR1 protein purified from tobacco and other solanaceous plants (Niderman et al., 1995). Overexpression of several PR1 genes in different plant species also increased their resistance to oomycetes (Sarowar et al., 2005; Broekaert et al., 2000), whereas the effect on other pathogen taxa is unclear. Interestingly, PR1 was recently shown to bind sterols, indicative of defensive mode of action that is based on restriction of this important nutrient for oomycetes (Gamir et al., 2017). The PR2 proteins are ß-1,3-beta-glucanases that are thought to attack the cell wall of invading fungi and oomycetes. Several cases of enhanced resistance of plants overexpressing PR2 to oomycetes were reported (Broekaert et al., 2000).
Another important family of PRs are the PR5 proteins, that encompass the osmotins and thaumatin-like proteins (van Loon et al., 2006). A 24-kD protein, named osmotin, isolated from tobacco, was shown to have inhibitory activity to P. infestans in vitro (Woloshuk et al., 1991). Overexpression of osmotin in transgenic potato plants delayed the development of late blight disease caused by P. infestans (Liu et al., 1994). Osmotin was shown to trigger cell death in Saccharomyces cerevisiae through its interaction with a plasma membrane protein (Narasimhan et al., 2005). It is unclear whether a similar activity is also effective against oomycetes.
Other non-proteinaceous compounds that are important for resistance to oomycetes in Arabidopsis are small secondary metabolites. Important anti-microbial compounds produced by Arabidopsis are for instance indole-glucosinolates (iGS) and the phytoalexin camalexin (Glawischnig, 2007; Bednarek, 2012). Mutant plants that no longer produce either iGS or camalexin are only slightly more susceptible to P. brassicae (Schlaeppi et al., 2010; Schlaeppi and Mauch, 2010). The effect appeared stronger when tested with P. capsici (Wang et al., 2013). In both cases, a very strong gain in susceptibility was observed in the double mutant cyp79b2 cyp79b3, that is blocked in the production of indole-3-aldoxime, which is a common precursor for iGS and camalexin. Camalexin appears to contribute to resistance to Arabidopsis downy mildew, as several phytoalexin-deficient (pad) mutants are slightly more susceptible to Hpa. Whether iGS also contributes to downy mildew resistance is not clear, although a proposed role of iGS in innate immunity suggests it would contribute to basal resistance (Clay et al., 2009). Many new compounds with anti-microbial activity are still being discovered in Arabidopsis, such as the recently discovered 4-hydroxyindole-3-carbonyl nitrile (4-OHICN), a previously unknown Arabidopsis metabolite (Rajniak et al., 2015). This is not surprising as each plant species is thought to produce thousands of secondary metabolites, many of which have a presumed role in defense (Dixon, 2001). However, much remains to be learned about the exact mechanisms that inhibit oomycete growth during PTI and ETI.
GENES INVOLVED IN SUSCEPTIBILITY TO OOMYCETES
Many non-immunity related processes are thought to contribute to plant disease susceptibility. Plants often inadvertently facilitate pathogens in establishing disease. Three different levels of susceptibility-enabling processes can be distinguished: (i) attraction and attachment of pathogens to host cells, (ii) accommodation of specialized infection and feeding structures inside plant cells, and (iii) nutrient production and transport from host to pathogen (Lapin and Van den Ackerveken, 2013). One can imagine that mutation of host genes that affect any of these processes leads to plants with reduced disease susceptibility. However, such mutations could also affect basal plant processes that affect the overall physiology of the plant. In genetic terms, susceptibility (S) genes can be defined as genes that are involved in disease susceptibility and contribute positively to the infection process. By this definition, impairment of S genes leads to enhanced disease resistance.
In Arabidopsis several genes for susceptibility to oomycetes have been identified. Several of these are involved in host amino acid metabolism, e.g., the ASPARTATE KINASE2/RAR1 SUPPRESSOR 1 (AK2/RSP1), DIHYDRODIPICOLINATE SYNTHASE2/RAR1 SUPPRESSOR 2 (DHDPS2/RSP2) and DOWNY MILDEW RESISTANT 1 (DMR1). The rsp1 and rsp2 mutants accumulate methionine, threonine and isoleucine, which is linked to increased resistance to Hpa (Stuttmann et al., 2011). Similarly, the Arabidopsis dmr1 mutant, that is mutated in HOMOSERINE KINASE (HSK), has increased levels of homoserine, which is a common precursor of methionine, threonine, and isoleucine. L-homoserine application was sufficient to make Arabidopsis resistant to Hpa (van Damme et al., 2009). The resistance in rsp1, rsp2 and dmr1 mutants is independent of salicylic acid (SA)-induced defense (van Damme et al., 2009; Stuttmann et al., 2011).
A very different group of susceptibility genes are those that encode negative regulators of immunity. When impaired, for instance by mutation, these regulators are no longer able to suppress certain immune responses and therefore show an enhanced disease resistance. Two examples from Arabidopsis are the IMPAIRED OOMYCETE SUSCEPTIBILITY1 (IOS1) and the DOWNY MILDEW RESISTANT 6 (DMR6) proteins. IOS1 is a leucine-rich repeat receptor-like kinase (LRR-RLK) and is required for full susceptibility to Hpa. In addition, the ios1 mutant is also resistant to the oomycete P. parasitica (Hok et al., 2011; Hok et al., 2014). The loss of susceptibility of the ios1 mutant seems linked to ABA hypersensitivity and independent of plant defense mechanisms. Impairment of negative regulation of ABA signaling in ios1 is suggested to lead to impaired susceptibility to Hpa (Hok et al., 2014). Also, Arabidopsis dmr6 mutants are resistant to Hpa, the oomycete P. capsici, and other biotrophic pathogens (van Damme et al., 2009; Zeilmaker et al., 2015). DMR6 and its close paralog DMR6-LIKE OXYGENASE 1 (DLO1/S3H) are needed for hydroxylation of SA to 2,5- and 2,3-dihydroxybenzoic acid, respectively. Overexpression of DMR6 and DLO1 leads to depletion of SA, which renders the plants highly susceptible to Hpa (Zeilmaker et al., 2015).
An important question to keep in mind when analyzing mutants with lesions in genes encoding negative regulators of immunity is whether they truly have a suppressive role in defense. This is particularly important for mutants that show autoimmune phenotypes, also referred to as lesion mimic mutants (Rodriguez et al., 2015). There are several examples of “negative regulator” mutants that can be suppressed by mutations in NLR genes encoding for cytoplasmic immune receptors. For instance, the Arabidopsis mpk4 mutant exhibits constitutive activation of defense and was classified as a negative regulatory mutant. However, it appeared that the mpk4 mutation required the presence of the NLR SUMM2 (Zhang et al., 2012). The idea is that the MPK4 protein, which is a target of the Pseudomonas effector HopAl1, is guarded by the NLR SUMM2. In the mpk4 mutant, SUMM2 would be constitutively triggered resulting in an autoimmune phenotype.
Ongoing and future studies are likely to reveal more plant proteins that have an important role in disease susceptibility to oomycete pathogens. Besides finding additional components of the plant immune system that are targeted by oomycetes, non-immunity-related proteins that function in ‘attracting’, ‘accommodating’, and ‘feeding’ pathogens can be found. The identification and functional analysis of the encoding genes could uncover so far unknown molecular processes in plant disease susceptibility, that will be instrumental in the design of novel strategies for disease resistance breeding. Our understanding of such processes, in the context of Arabidopsis-oomycete interactions, is discussed in the following sections.
SUPPRESSION OF IMMUNITY
The preceding sections of this chapter have established that plants can deploy an effective immune response against oomycete pathogens. However, it is also well-documented that phytopathogenic oomycetes are capable of potent immune suppression (Jiang and Tyler, 2012). This is illustrated in Figure 4A, in which a co-infection experiment demonstrates that Albugo laibachii can suppress Arabidopsis immunity and thereby enables colonization by otherwise incompatible isolate of Hpa (Cooper et al., 2008). As mentioned earlier, A. laibachii can even render Arabidopsis susceptible to Phytophthora infestans, which is nonpathogenic on Arabidopsis due to several layers of genetically redundant resistance mechanisms (Belhaj et al., 2017; Prince et al., 2017). An example of immune suppression at the cellular level is shown in Figure 4B, where activation of PR1 promoter-GUS reporter gene fusion is suppressed specifically in Arabidopsis cells adjacent to Hpa infection structures (Caillaud et al., 2013; Asai et al., 2014). These images graphically illustrate oomycetes’ capacity to tamper with plant immune systems. In the sections below, we will summarize how Arabidopsis is used to dissect the molecular mechanisms that underpin this suppression of an otherwise potent immune response.
Figure 4.
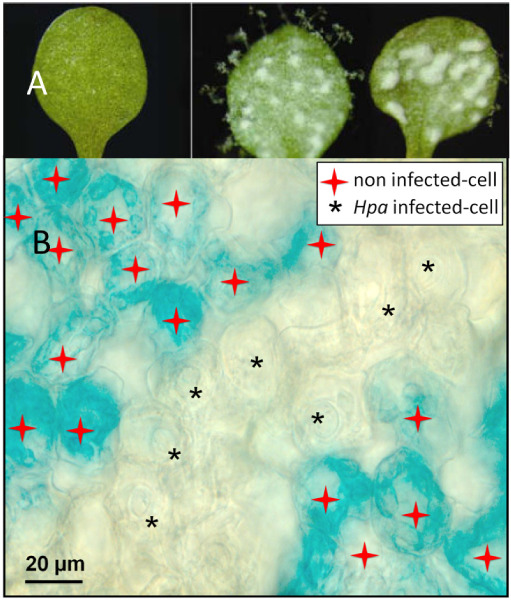
Suppression of host immunity by adapted oomycete pathogens.
(A) Cotyledons of Arabidopsis thaliana Col-5 inoculated solely with aviru-lent H. Arabidopsis Cand5 and showing no symptoms (left) or sequentially inoculated with Albugo candida subsp. arabidopsis isolate Acem1 and Cand5 showing downy mildew sporangiophores (center, upper surface) and white blisters (right, lower surface). Image reproduced from (Cooper et al., 2008).
(B) GUS staining of pro(PR1)::GUS in Arabidopsis leaves 6 DAI Hpa Waco9. No GUS was detected in Hpa-haustoriated mesophyll cell (black stars), while GUS staining was restricted to nonhaustoriated mesophyll cells (red stars). Image reproduced from (Caillaud et al., 2013).
It is becoming increasingly clear that oomycetes’ most important tools for host immune suppression are effector proteins that are secreted from the pathogen to the outside (apoplast) or the interior of plant cells (Stassen and Van den Ackerveken, 2011; Fawke et al., 2015). Genomic analyses, coupled with genetic and molecular experiments, have allowed for discovery of hundreds of putative effector-encoding genes in oomycete genomes (Jiang and Tyler, 2012; Pais et al., 2013). The majority of oomycete effectors contain a classical signal peptide motif directing secretion from the pathogen. C-terminal of the signal peptide are additional known or putative functional motifs that enable classification of the secreted proteins into families (Kamoun, 2006). Such classifications provide a foundation from which to investigate the functions of the genes. Table 3 lists several families of oomycete effectors and estimates of their copy numbers in genomes of oomycetes that infect Arabidopsis. These can be considered only an approximate inventory of effectors in the respective pathogens, and it is likely that these numbers will change as the quality of genome data, experimental evidence, and predictive motifs are iteratively refined (Dalio et al., 2018).
Table 3.
Predicted secreted proteins from oomycete pathogens of Arabidopsis.
| Species | Genome Size | Total Secretome | RxLR | CRN | NEP | Elicitin-like | Reference |
|---|---|---|---|---|---|---|---|
| H. arabidopsidis | 81.6 Mb | 2157 | 134 | 20 | 10 | 15 | (Baxter et al., 2010) |
| A. candida | 45.3 Mb | 929 | 29* | 6 | 0 | 9 | (Links et al., 2011) |
| A. laibachii | 43 Mb | 672 | 49 | 2 | 0 | 1 | (Kemen et al., 2011) |
| P. capsici | 64 Mb | 2911 | 357 | 29 | NP | NP | (Lamour et al., 2012) |
NP-No prediction
The best studied oomycete effector family is defined by the motif RXLR that is found adjacent to the N-terminal signal peptide. Hundreds of putative RXLR effector genes are present in genomes of downy mildew pathogens and species within the Phytophthora genus (Jiang and Tyler, 2012). The RXLR motif is proposed to mediate entry of the protein into plant cells, although this is controversial (Kale et al., 2010; Petre and Kamoun, 2014). Once inside the plant the RXLR motif does not appear to play a functional role. Rather, the function-specific region of the protein is downstream of the signal peptide and RXLR motif (Birch et al., 2006). We previously summarized how some RXLR proteins are recognized as avirulence proteins by NLR immune surveillance proteins. More relevant to this section of the chapter are lines of evidence indicating that RXLR proteins play a major role in immune suppression. These studies are reviewed in detail elsewhere (Birch et al., 2009; Schornack et al., 2009; de Jonge et al., 2011; Kale and Tyler, 2011; Koeck et al., 2011; Stassen and Van den Ackerveken, 2011; Wawra et al., 2012; Pais et al., 2013; Anderson et al., 2015; Fawke et al., 2015). Below, we highlight several examples in which the experimental resources of Arabidopsis have been used to accelerate progress towards understanding how RXLR proteins promote oomycete diseases.
RXLR effectors are thought to enter plant cells and target specific plant proteins or protein complexes for destruction or modification to benefit the pathogen. Thus, the experimental tools of Arabidopsis are well-suited to identify RXLR protein targets, to understand the functions of these targets in plant regulatory networks and define the mechanisms through which the target is engaged by the corresponding effector and the resultant effect on plant cellular functions. Arabidopsis has been used to particularly good advantage in studies aimed at high- to medium-throughput examination of effectors to provide first steps toward mechanistic understanding (Anderson et al., 2015). One such approach utilized a phytopathogenic Pseudomonas bacterial strain as a surrogate to deliver Hpa effectors to Arabidopsis cells via Type III secretion (Fabro et al., 2011). This system was based on two clever features: The first was an “effector-detector vector” in which the effector of interest is fused to a leader from a bacterial Type III effector that directs the oomycete effector through the Type III system and thus to the interior of plant cells (Sohn et al., 2007). The second feature was to engineer the bacteria with a constitutively active luciferase gene, by which in planta bacterial growth could be easily quantified by light emission (Fabro et al., 2011). This assay provided an estimation of whether the secreted effector promoted or retarded bacterial growth, indicative of virulence or avirulence activity, respectively. This system enabled facile examination of 64 RXLR genes, providing evidence for virulence-promoting activity for 42 effectors (Fabro et al., 2011).
Another approach is to exploit the ease of Arabidopsis transformation to generate stably transformed plants that express effector protein genes, controlled by plant regulatory sequences. This approach has been very productive for bacterial Type III effectors and is now being used to characterize RXLR proteins. For example, Pel and colleagues created transgenic lines expressing several RXLR proteins from Hpa and used these lines to establish virulence-promoting activities using pathogen disease phenotypes, cytological assays, and marker genes that are well-established in Arabidopsis (Pel et al., 2014). Arabidopsis transgenic plants have also been used in genetic screens to identify effectors that target specific signaling pathways/sectors, based on suppression of a promoter-reporter gene fusion that provides a readout for the pathway of interest. This approach has implicated RXLR effectors that suppress SA-mediated signaling based on reduction of SA-induced PR1-GUS activity (Asai et al., 2014). Another approach was to use Arabidopsis expressing RXLR effector-GFP protein fusions to estimate subcellular localization of 49 Hpa RXLR effectors (Caillaud et al., 2012). This approach revealed the nucleus and plasma membrane as predominant destinations. These clever screens represent only the beginning of what might be achieved using different Arabidopsis genetic resources. For example, it would be of great interest to exploit Arabidopsis lines that could report reprogramming of relevant host processes other than immune signaling (e.g., host cell structure, trafficking, or metabolism) (Lapin and Van den Ackerveken, 2013).
The most dramatic application of Arabidopsis tools to understand RXLR effector biology was described in two studies that exploited the Arabidopsis ORFeome to conduct a protein-protein interaction screen with effectors from Hpa, the bacterium Pseudomonas syringae, and the obligate biotrophic fungus Golovinomyces orontii (Mukhtar et al., 2011; Wessling et al., 2014). Together, these studies revealed nine Arabidopsis proteins that interact with effectors from all three pathogens, along with another 23 that interact with effectors from two of the three pathogens. The biological relevance of these interactions could then be tested by testing loss of function mutations, easily obtained from the Arabidopsis T-DNA knockout collection, for effects on susceptibility to pathogen infection. The targeting of plant proteins by pathogens from different biological kingdoms validates the prediction that pathogens might evolve convergently to target points of vulnerability in host signaling networks. This reaffirms that Arabidopsis studies will yield important knowledge of conserved immune strategies in crop species. In addition to these points of convergence, the screen also revealed proteins that bind to effectors from only one of the three species, providing leads into virulence strategies that are specific to oomycete, bacterial, or fungal lineages (Mukhtar et al., 2011; Wessling et al., 2014).
These screens provide an important basis for prioritizing hundreds of RXLR effector genes for the painstaking studies to reveal their modes of action. The experimental tools of Arabidopsis provide obvious advantages for studies aimed at deep mechanistic understanding. This is exemplified by examination of the Hpa effector RXLR44, through which a subunit of the mediator transcriptional complex was revealed as the target (Caillaud et al., 2013). RXLR44 targets the Med19 subunit for proteosomal degradation. Nullification of Med19 affects hormone signaling sectors that control immunity: JA-responsive genes are de-repressed, which activates antagonistic cross-talk through which SA signaling is repressed and the plant immune response is compromised (Caillaud et al., 2013). In this way, it was revealed that Hpa exploits JA-SA interplay to promote virulence, thereby joining bacteria and fungi in convergent exploitation of this malleable node in the immune signaling network (Gimenez-Ibanez et al., 2016; Zhang et al., 2017).
Although most of the studies discussed so far focus on effectors from the natural Arabidopsis pathogen Hpa, it is important to emphasize that Arabidopsis is also an informative system for effectors from Phytophthora species. For example, the Penetration-Specific Effector 1 (PSE1) effector from P. parasitica was shown to affect auxin signaling in Arabidopsis (Evangelisti et al., 2013). Arabidopsis can also be exploited to identify functions for effectors from nonpathogens of Arabidopsis that presumably target plant proteins which are broadly conserved and are targeted by diverse oomycetes. This is illustrated by a study in which Phytophthora infestans RXLR proteins that affect PTI were identified from screens of Arabidopsis expressing the PAMP-inducible FRK9 promoter fused to a Luciferase reporter (Zheng et al., 2014). The Arabidopsis extracellular ATP receptor kinase LeRK-I9 is targeted by the Phytophthora RXLR effector IPI-O, perhaps to destabilize the plasma membrane-cell wall interface and/or interfere with perception of eATP (Bouwmeester et al., 2011b; Bouwmeester et al., 2011a; Bouwmeester et al., 2014; Balague et al., 2017). Another example is the Phytophthora suppressor of RNA silencing (PSR) effectors from the soybean pathogen Phytophthora sojae that suppress plant RNA interference to manipulate immune signaling (Qiao et al., 2013; Xiong et al., 2014). In this case, Arabidopsis was used to identify one target of these proteins: an RNA-binding protein that had not been previously linked with immune system regulation (Qiao et al., 2015). These and other studies illustrate how Arabidopsis could be used to identify effector target proteins that are “conserved” in the sense that they are targeted in diverse plant species, perhaps by diverse pathogens (Anderson et al., 2012; Deb et al., 2018b; Deb et al., 2018a; Tomczynska et al., 2018). Such proteins could be attractive targets for breeding/bioengineering.
The above examples also illustrate a major benefit of effector-driven research: the effectors often “lead” us to novel insights into the composition and function of plant regulatory networks (Win et al., 2012). In this context, we draw attention back to Table 3, which illustrates that oomycetes contain several effector families with important functions in disease. The Nep1 (Necrosis and Ethylene-inducing 1)-like proteins (NLPs) were introduced previously in the section on PTI. These proteins are thought to be utilized by necrotrophic and hemi biotrophic pathogens to induce plant cell death; however, the obligate biotroph Hpa expresses a number of NLP proteins that are not capable of inducing necrosis (Baxter et al., 2010). Their virulence function awaits definition. The “crinkler” family of effectors was functionally defined by plant cell necrosis activity and also contains a definitive protein motif (Stam et al., 2013). Another broadly conserved class of effectors are elicitin-like proteins, which are thought to bind sterols in the apoplast (Derevnina et al., 2016). This is important because phytopathogenic oomycetes do not synthesize sterols. These and other effector families have received little attention, compared to RXLR proteins, and represent good opportunities for productive experimentation, using the Arabidopsis tools described above. It will be particularly intriguing to explore Pythium or Albugo effector families that might be analogous to RXLR effectors in these important but poorly studied oomycete lineages. The recently described CHXC family from Albugo is particularly intriguing and might provide a mechanistic link to the recent report that Albugo can alter Arabidopsis tryptophan metabolism and interfere with responses to salicylic acid (Kemen et al., 2011; Prince et al., 2017). As mentioned earlier, Albugo can also act as a hub to affect the microbiome of infected plants and appears highly tolerant to host immune responses (McMullan et al., 2015; Ruhe et al., 2016). These traits are theorized to provide a competitive advantage for niche colonization in the wild. Much more remains to be learned about these and other fascinating aspects of Arabidopsis-oomycete interactions.
ACCOMMODATION OF OOMYCETE INFECTION STRUCTURES
The preceding section established that secretion of effectors is a major mechanism through which oomycetes can suppress host immunity. Another critical process in which the pathogen likely manipulates the host is in accommodation of infection structures, particularly the haustorium which penetrates the plant cell wall to create intimate associations with individual host cells (Fig. 2, Fig. 5) (Lapin and Van den Ackerveken, 2013). As mentioned previously, the haustorium is a specialized feeding structure that is formed by biotrophic and hemi-biotrophic oomycetes during plant colonization (Spencer-Phillips, 1997; Szabo and Bushnell, 2001; Wang et al., 2017). Effectors are secreted from haustoria to the exterior and interior of host cells (Wang et al., 2017). Haustorium formation is initiated when oomycetes secrete hydrolases to locally degrade the plant cell wall and establish an entry point. Microscopy of infected Arabidopsis leaves has illustrated the ultrastructure of oomycete haustoria (Mims et al., 2004; Soylu et al., 2004; Lu et al., 2012) (Fig. 5). Following penetration, haustoria grow into the host cell by invagination of the plant cell membrane. The pathogen haustorial membrane is distinct from the host extrahaustorial membrane and the two membranes are separated by a glycan-rich extrahaustorial matrix. The haustorium itself contains numerous large organelles including vacuoles, mitochondria, and lipid bodies (Fig. 5A). These structures suggest the haustoria maintains an active metabolism and reinforces the hypothesis that it is the primary site of pathogen feeding (Mims et al., 2004). The extrahaustorial membrane displays a highly convoluted topology, perhaps resulting from vesicle traffic to and from the haustoria (Fig. 5A).
Figure 5.
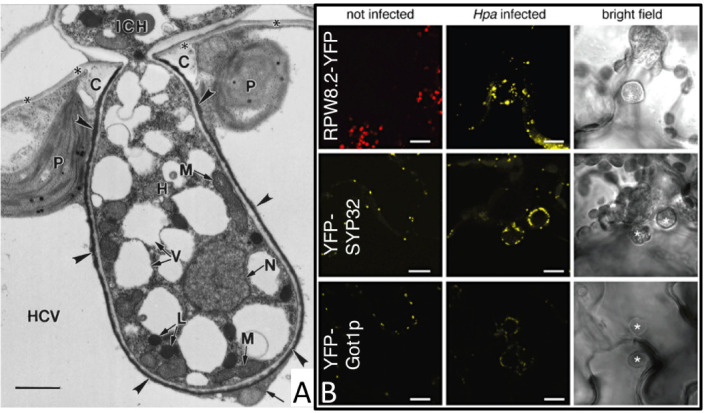
Manipulation of host cell structure and metabolism.
(A) Transmission electron micrograph of fixed Hpa haustoria in leaf cells of At. Near-median longitudinal section of a haustorium (H) showing its continuity with an intercellular hypha (ICH). collar of host cell wall material (C). Host-cell structures: the cell wall (asterisks), plastids (P), the central vacuole (HCV), and cytoplasm (arrow). Pathogen structures: nucleus (N), mitochondria (M), lipid bodies (L), vacuoles (V), and extrahaustorial matrix (arrowheads). Scale bar = 1 μm. Image reproduced from (Mims et al., 2004).
(B) Secretory vesicles differentially localize around Hpa. Confocal micrographs of At transgenic lines expressing the indicated fluorophore fusions show cross-sections of non-infected and Hpa-infected leaves at 3 dpi. Hpa haustoria are shown in bright field images indicated by asterisks. RPW8.2–YFP, YFP–SYP32 and YFP–Got1p are detected in vesicles around Hpa haustoria. Red signals seen in uninfected RPW8.2–YFP leaves are chlorophyll autofluorescence. Bar = 10 μm. Image reproduced from (Lu et al., 2012).
The signaling and trafficking differences that constitute host resistance versus successful haustorial formation are not well understood. It is clear that many responses originate from the plant, but the pathogen likely coopts some of these functions for its own purposes. The extrahaustorial membrane is synthesized de novo and lacks many integral proteins typically found in the plant PM (Lu et al., 2012). Some plant defensive proteins localize to the extrahaustorial membrane, including immune receptors such as FLS2 and trafficking proteins such as PENETRATION1 (PEN1, Fig. 5B). Studies of early Hpa infection reveals haustorial penetration is accompanied by reorganization of GFP-tagged cytoskeleton and organelles in Arabidopsis mesophyll cells (Takemoto et al., 2003; Mims et al., 2004; Lu et al., 2012). This reorganization is observed in compatible and incompatible interactions. Some of these responses are stimulated by plant immune signaling as well as mechanical disturbance (Hardham et al., 2008). However, a successful infection would likely benefit from plant organelles close to haustorium for access to plant cell trafficking and metabolism. Host cell rearrangements are accompanied by accumulation of secretory vesicles, flush with lipids for the new membrane (Takemoto et al., 2003). Other cargo contained in these vesicles remains to be identified. The purpose of these vesicles as defense mechanism or transporter of nutrients to the pathogen is unclear.
A signature of successful Arabidopsis resistance to Hpa and Albugo invasion is robust production of cell wall appositions, or encapsulation of haustorium in callose (Donofrio and Delaney, 2001). These callose-containing structures are hypothesized to provide physical protection against pathogen feeding structures (Allen and Friend, 1983). Arabidopsis PLASMODESMATA-LOCATED PROTEIN 1 moves to haustorial sites and stimulates callose production to impede haustorial entry. Suppression of immunity likely includes reduction in PDLP1 expression and activity, as the gene is not expressed in fully colonized cells (Caillaud et al., 2014). Plants deficient in salicylic acid signaling fail to produce robust callose depositions and often produce callose bands at the neck of haustorium (Donofrio and Delaney, 2001). While abundant callose appears to stymie haustorium formation, cal-lose neck bands at the site of cell wall penetration of Hpa and Albugo haustorium may promote haustorial accommodation.
Although studies with Arabidopsis have provided some initial insights, the biogenesis of haustoria and cellular aspects of interaction with plant cells are poorly understood and represent a very fruitful area for inquiry that encompasses cell biology of the plant and the oomycete. Recently, Arabidopsis orthologs of symbiosis genes of legumes were shown to also play a role during downy mildew infection. Mutants were affected in haustorial maintenance, suggesting overlap in accommodation of microbes between the different plant species (Ried et al., 2018). Arabidopsis will be particularly useful for identifying host factors that are manipulated during haustorial accommodation.
NUTRIENT ACQUISITION
Another challenge faced by oomycetes is the need to obtain nutrients from the host (Lapin and Van den Ackerveken, 2013). This challenge is compounded in obligate oomycetes by the loss of conserved metabolic pathways (McDowell, 2011; Kemen and Jones, 2012). For example, genome analysis of the obligate biotroph Hpa revealed loss of the machinery to reduce inorganic nitrate and sulfite to their essential organic forms (Baxter et al., 2010). The Albugo genome reveals similar deficiencies along with the loss of the thymine synthetic pathway (Links et al., 2011). Thus, these pathogens must have a way to compensate for the lack of metabolic machinery in their own genomes, and it will be interesting to compare their nutrient acquisition strategies with those employed by Phytophthora pathogens in which these pathways are intact. Unfortunately, similar to accommodation of haustoria, nutrient acquisition by oomycetes is poorly understood and Arabidopsis has not been fully exploited. Only one publication reports physiological aspects of an Arabidopsis-oomycete interaction: Arabidopsis invertase activity is induced during Albugo infection, while chlorophyll production and photosynthesis are down-regulated (Chou et al., 2000). These changes occurred only in regions directly associated with the pathogen, suggestive of pathogen manipulations or a localized defense response.
Much potential exists to exploit the tools of Arabidopsis to better understand the metabolic interplay between plants and oomycetes. Vesicle traffic through the extrahaustorial matrix is hypothesized to contain effectors and virulence factors from the pathogen, and sugars, proteins, and lipids from the host. Bacterial plant pathogens are now known to induce expression of sugar transporters in their hosts at the site of infection (Chen et al., 2010), and research is ongoing to identify if this phenomenon occurs during oomycete infections. Indeed, a mutant in the Arabidopsis SWEET2 gene reduces susceptibility to Pythium in roots (Chen et al., 2015). However, initial clues are evident in recent gene expression profiling studies: A survey of gene expression in Phytophthora parasitica during Arabidopsis root colonization suggested that uptake of host amino acids is prevalent during early infection (Attard et al., 2014). A survey of Arabidopsis gene expression during infection of leaves by Hpa identified an up-regulated cluster of genes involved in nitrate transport (Asai et al., 2014). Detailed experimental follow-up of these observations would likely provide important insights into the relationship between host and pathogen metabolism.
In this context, the accessible genetic tools of Arabidopsis and capacity for gene editing provide a splendid resource to identify genes related to metabolism that affect the interaction with the pathogen (Sonawala et al., 2018). If a pathogen requires specific metabolites as a carbon or nitrogen source, pathogen starvation may be caused by loss of a gene responsible for that metabolite’s synthesis or transport. Such genes might fulfill the genetic definition of a susceptibility gene (S-gene) as discussed in the immune signaling section. Importantly, such genetic knockouts could be resistant because of deficiencies in nutrients critical to pathogen growth, rather than activation/priming of an immune response. One such example might be the aforementioned mutants rsp1 and rsp2, which accumulate methionine, threonine and isoleucine, without induction of a defense response (Stuttmann et al., 2011). These alterations in metabolism are linked to increased resistance to Hpa and a biotrophic fungal pathogen (Stuttmann et al., 2011). Similarly, the Arabidopsis dmr1 mutant, that is mutated in HOMOSERINE KINASE (HSK), has increased levels of homoserine, which is a common precursor of methionine, threonine, and isoleucine. L-homoserine application was sufficient to make Arabidopsis resistant to Hpa (van Damme et al., 2009). The resistance in rsp1, rsp2 and dmr1 mutants is independent of salicylic acid (SA)-induced defense (van Damme et al., 2009; Stuttmann et al., 2011). This separation of metabolic incompatibility from canonical immunity demonstrates that pathogen virulence depends on more than host immune suppression. Pathogens have adapted to use metabolites present in their host, and perturbations of these host nutrient pools apparently leads to host resistance, particularly to biotrophic species. Certain metabolic genes may be the direct or downstream targets of effectors, as is the case for bacteria. These targets might be more difficult to identify than well conserved immune hubs. Even so, Arabidopsis provides an ideal system for continued study in this area. The well-studied metabolism, and availability of T-DNA insertion mutants for most genes will allow for screens and analysis of additional S-genes and sources of metabolic incompatibility.
Although much remains to be discovered, it is clear that oomycete plant pathogens are adapted to exploit their host to their own ends. These pathogens turn their hosts’ cell biology against them to achieve full virulence. Host cells respond differently in compatible and incompatible interactions giving indication of pathogen manipulations. Structures are rebuilt to physically accommodate the parasite. The pathogen metabolism is streamlined to utilize molecules found in the host. Understanding how plants accommodate pathogens has potential to establish new mechanisms of resistance. Crop species can be bred or engineered as inhospitable hosts, by removing S-genes or establishing metabolic incompatibility. This may provide novel forms of protection from dangerous and economically important plant diseases. Work in Arabidopsis-oomycete pathosystems will yield further insights into these interactions and open these translational opportunities.
SUMMARY AND OPPORTUNITIES FOR FUTURE RESEARCH
Much progress has been made over the three decades in which Arabidopsis has been used to understand plant-oomycete interactions. Indeed, research with Arabidopsis has been at the fore-front of some of the most important insights in this area, which are summarized in Table 4.
Table 4.
Major insights into plant-oomycete interactions from Arabidopsis research.
| Insight | Key References |
|---|---|
| Cloning of the first R genes against oomycetes | Parker et al., 1997; Botella et al., 1998; McDowell et al., 1998 |
| First insights into evolutionary dynamics of R genes against oomycetes | Botella et al., 1998; McDowell et al., 1998; Bittner-Eddy et al., 2000 |
| Definition of important immune system regulators | Parker et al., 1996 |
| Identification and map-based cloning of the first mutants exhibiting gain of resistance to oomycetes | Van Damme et al., 2005; van Damme et al., 2008 |
| Genetic definition of the complexity of the immune signaling network against oomycetes | Aarts et al., 1998; McDowell et al., 2000; Bittner-Eddy and Beynon, 2001 |
| Definition of penetration resistance | Lipka et al., 2005 |
| Molecular cloning of the first oomycete Avr gene | Allen et al., 2004 |
| First insights into evolutionary dynamics of Avr gene loci | Allen et al., 2004; Armstrong et al., 2005; Rehmany et al., 2005 |
| Identification of the RxLR motif | Rehmany et al., 2005 |
| Test bed for new technologies to understand effector functions | Fabro et al., 2011; Caillaud et al., 2012 |
| Discovery that oomycete, fungal, and bacterial effectors can target the same plant signaling hubs | Mukhtar et al., 2011; Wessling et al., 2014 |
| First insights into genomic basis and evolution of obligate biotrophy | Baxter et al., 2010; Kemem et al., 2011 |
However, there are many aspects of plant-oomycete interactions that are poorly understood. Table 5 outlines a number of general challenges and questions that remain open: For example, despite the impressive steps forward in understanding how plants perceive oomycete PAMPs and effectors, we lack a comprehensive map of downstream immune signaling pathways and we have limited direct proof about the causal mechanisms through which oomycete growth is impaired during a successful immune response. Linkages of the immune system to other plant physiological process also remain to be revealed. In addition, we have only scratched the surface of mechanistic comprehension of how oomycetes target key nodes in the immune system. The interactome studies referenced above provide an inspiring example of how these questions can be addressed productively. Furthermore, we emphasize strongly that suppression of immunity represents only one of several tasks that an adapted pathogen must accomplish to complete its life cycle on the host. Very little is known about how oomycetes interpret physical and/or chemical cues to navigate through host tissues, nor how they manipulate host cell structure establish feeding conduits, nor how they manipulate host metabolism to serve their own needs (Lapin and Van den Ackerveken, 2013). All of these processes comprise major gaps in understanding of how plants and oomycetes interact. In addition, huge potential exists to exploit natural variation in Arabidopsis-oomycete interactions to consider host-microbe ecology and evolution, along with the role of the microbiome in plant-oomycete interactions and the impacts of oomycetes on plant-microbiota interactions (Holub, 2008; Kemen and Jones, 2012). In summary, there is extensive space for innovative, impactful research within Arabidopsis-oomycete pathosystems.
Table 5.
Major knowledge gaps in understanding of plant-oomycete interactions that can be efficiently addressed with Arabidopsis research.
| Challenge |
|---|
| Comprehensive map of the plant immune system, including connections to other processes |
| Understand how points of vulnerability in the immune network are targeted by oomycetes and other pathogens |
| Functional studies of unknown effector proteins |
| Explore how oomycetes obtain nutrients from plant hosts |
| Understand how oomycetes manipulate plant cell structure and physiology |
| Exploring the evolution and ecology of plant-oomycete interactions |
| Defining how the plant microbiome influences interactions with oomycetes |
| Leverage Arabidopsis resources to test innovative strategies for disease control |
For these reasons, we strongly recommend that the molecular plant-microbe research community continue to exploit the well-developed experimental advantages of Arabidopsis as a reference system. In this way, we can move efficiently towards a systems-level understanding of plant-oomycete interactions, which will undoubtedly inspire new approaches towards low-input mitigation of crop diseases (Michelmore et al., 2017). Indeed, leads for translational research are already emerging directly from basic research on Arabidopsis-oomycete interactions. For example, cloned pattern-recognition receptors (e.g., RLP23) (Albert et al., 2015) and NLR proteins could be moved to crop species, as already demonstrated for WRR proteins (Borhan et al., 2010; Cevik et al., 2019). Another approach is to use the power of Arabidopsis genetics to identify plant genes in which loss-of-function mutants lead to reduced susceptibility (i.e., mutants of oomycete disease susceptibility genes or negative regulators of immunity) (Boevink et al., 2016). Such genes can then be mutated in crops (e.g., by TILLING or by editing) and tested for pathogen resistance. This approach is currently being pursued for the DMR6 gene (Zeilmaker et al., 2015). Finally, Arabidopsis can serve as a test bed for high-risk approaches, such as host-induced gene silencing, for which a genetically facile host could speed optimization (Govindarajulu et al., 2015).
Acknowledgments
We gratefully acknowledge the craftpersonship of Kasia Dinkeloo on Figure 2. This work was made possible by support from the National Science Foundation Grant nos. IOS-0744875 and IOS-1353366, by Agriculture and Food Research Initiative Competitive Grant no. 224426 from the USDA National Institute of Food and Agriculture, and a Less is More grant no. 847.13.006 of the Netherlands Organization of Scientific Research (NWO).
REFERENCES
- Adhikari B.N, Hamilton J.P, Zerillo M.M, Tisserat N, Levesque C.A, Buell C.R. Comparative genomics reveals insight into virulence strategies of plant pathogenic oomycetes. PloS One. 2013 doi: 10.1371/journal.pone.0075072. e75072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adhikari T.B, Liu J.Q, Mathur S, Wu C.X, Rismmer S.R. Genetic and molecular analyses in crosses of Race 2 and Race 7 of Albugo candida. Phytopathology. 2003:959–965. doi: 10.1094/PHYTO.2003.93.8.959. [DOI] [PubMed] [Google Scholar]
- Albert I, Bohm H, Albert M, Feiler C.E, Imkampe J, Wallmeroth N, Brancato C, Raaymakers T.M, Oome S, Zhang H, Krol E, Grefen C, Gust A.A, Chai J, Hedrich R, Van den Ackerveken G, Nurnberger T. An RLP23-SOBIR1-BAK1 complex mediates NLP-triggered immunity. Nat Plants. 2015:15140. doi: 10.1038/nplants.2015.140. [DOI] [PubMed] [Google Scholar]
- Allen F.H.E, Friend J. Resistance of potato tubers to infection by Phytophthora infestans a structural study of haustorial encasement. Physiological Plant Pathology. 1983:285-IN284. [Google Scholar]
- Allen R.L, Bittner-Eddy P.D, Grenvitte-Briggs L.J, Meitz J.C, Rehmany A.P, Rose L.E, Beynon J.L. Host-parasite coevolutionary conflict between Arabidopsis and downy mildew. Science. 2004:1957–1960. doi: 10.1126/science.1104022. [DOI] [PubMed] [Google Scholar]
- Anderson R.G, Deb D, Fedkenheuer K, McDowell J.M. Recent progress in RXLR effector research. Molecular Plant-Microbe Interactions. 2015:1063–1072. doi: 10.1094/MPMI-01-15-0022-CR. [DOI] [PubMed] [Google Scholar]
- Anderson R.G, Casady M.S, Fee R.A, Vaughan M.M, Deb D, Fedkenheuer K, Huffaker A, Schmelz E.A, Tyler B.M, McDowell J.M. Homologous RXLR effectors from Hyaloperonospora arabidopsidis and Phytophthora sojae suppress immunity in distantly related plants. Plant J. 2012:882–893. doi: 10.1111/j.1365-313X.2012.05079.x. [DOI] [PubMed] [Google Scholar]
- Asai S, Furzer O.J, Cevik V, Kim D.S, Ishaque N, Goritschnig S, Staskawicz B.J, Shirasu K, Jones J.D.G. A downy mildew effector evades recognition by polymorphism of expression and subcellular localization. Nature Communications. 2018:5192. doi: 10.1038/s41467-018-07469-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asai S, Rallapalli G, Piquerez S.J, Caillaud M.C, Furzer O.J, Ishaque N, Wirthmueller L, Fabro G, Shirasu K, Jones J.D. Expression profiling during Arabidopsis/downy mildew interaction reveals a highly-expressed effector that attenuates responses to salicylic acid. PLoS Pathogens. 2014 doi: 10.1371/journal.ppat.1004443. e1004443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ascunce M.S, Huguet-Tapia J.C, Ortiz-Urquiza A, Keyhani N.O, Braun E.L, Goss E.M. Phylogenomic analysis supports multiple instances of polyphyly in the oomycete Peronosporalean lineage. Mol Phylogenet Evol. 2017:199–211. doi: 10.1016/j.ympev.2017.06.013. [DOI] [PubMed] [Google Scholar]
- Attard A, Gourgues M, Callemeyn-Torre N, Keller H. The immediate activation of defense responses in Arabidopsis roots is not sufficient to prevent Phytophthora parasitica infection. New Phytol. 2010:449–460. doi: 10.1111/j.1469-8137.2010.03272.x. [DOI] [PubMed] [Google Scholar]
- Attard A, Evangelisti E, Kebdani-Minet N, Panabieres F, Deleury E, Maggio C, Ponchet M, Gourgues M. Transcriptome dynamics of Arabidopsis thaliana root penetration by the oomycete pathogen Phytophthora parasitica. BMC Genomics. 2014:538. doi: 10.1186/1471-2164-15-538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey K, Cevik V, Holton N, Byrne-Richardson J, Sohn K.H, Coates M, Woods-Tor A, Aksoy H.M, Hughes L, Baxter L, Jones J.D, Beynon J, Holub E.B, Tor M. Molecular cloning of ATR5(Emoy2) from Hyaloperonospora arabidopsidis an avirulence determinant that triggers RPP5mediated defense in Arabidopsis. Mol Plant Microbe Interact. 2011:827–838. doi: 10.1094/MPMI-12-10-0278. [DOI] [PubMed] [Google Scholar]
- Balague C, Gouget A, Bouchez O, Souriac C, Haget N, Boutet-Mercey S, Govers F, Roby D, Canut H. The Arabidopsis thaliana lectin receptor kinase LecRK-I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling. Mol Plant Pathol. 2017:937–948. doi: 10.1111/mpp.12457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barchenger D.W, Lamour K.H, Bosland P.W. Challenges and strategies for breeding resistance in Capsicum annuum to the multifarious pathogen, Phytophthora capsici. Frontiers in Plant Science. 2018:628. doi: 10.3389/fpls.2018.00628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baxter L, Tripathy S, Ishaque N, Boot N, Cabral A, Kemen E, Thines M, Ah-Fong A, Anderson R, Badejoko W, Bittner-Eddy P, Boore J.L, Chibucos M.C, Coates M, Dehal P, Delehaunty K, Dong S, Downton P, Dumas B, Fabro G, Fronick C, Fuerstenberg S.I, Fulton L, Gaulin E, Govers F, Hughes L, Humphray S, Jiang R.H.Y, Judelson H, Kamoun S, Kyung K, Meijer H, Minx P, Morris P, Nelson J, Phuntumart V, Qutob D, Rehmany A, Rougon-Cardoso A, Ryden P, Torto-Alalibo T, Studholme D, Wang Y, Win J, Wood J, Clifton S.W, Rogers J, Van den Ackerveken G, Jones J.D.G, McDowell J.M, Beynon J, Tyler B.M. Signatures of adaptation to obligate biotrophy in the Hyaloperonospora arabidopsidis genome. Science. 2010:1549–1551. doi: 10.1126/science.1195203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bednarek P. Sulfur-containing secondary metabolites from Arabidopsis thaliana and other Brassicaceae with function in plant immunity. Chembiochem. 2012:1846–1859. doi: 10.1002/cbic.201200086. [DOI] [PubMed] [Google Scholar]
- Belhaj K, Cano L.M, Prince D.C, Kemen A, Yoshida K, Dagdas Y.F, Etherington G.J, Schoonbeek H.J, van Esse H.P, Jones J.D, Kamoun S, Schornack S. Arabidopsis late blight: infection of a nonhost plant by Albugo laibachii enables full colonization by Phytophthora infestans. Cell Microbiol. 2017 doi: 10.1111/cmi.12628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernsdorff F, Doring A.C, Gruner K, Schuck S, Brautigam A, Zeier J. Pipecolic acid orchestrates plant systemic acquired resistance and defense priming via salicylic acid-dependent and -independent pathways. Plant Cell. 2016:102–129. doi: 10.1105/tpc.15.00496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bigeard J, Colcombet J, Hirt H. Signaling mechanisms in pattern-triggered immunity (PTI) Mol Plant. 2015:521–539. doi: 10.1016/j.molp.2014.12.022. [DOI] [PubMed] [Google Scholar]
- Birch P.R, Rehmany A.P, Pritchard L, Kamoun S, Beynon J.L. Trafficking arms: oomycete effectors enter host plant cells. Trends Microbiol. 2006:8–11. doi: 10.1016/j.tim.2005.11.007. [DOI] [PubMed] [Google Scholar]
- Birch P.R, Armstrong M, Bos J, Boevink P, Gilroy E.M, Taylor R.M, Wawra S, Pritchard L, Conti L, Ewan R, Whisson S.C, van West P, Sadanandom A, Kamoun S. Towards understanding the virulence functions of RXLR effectors of the oomycete plant pathogen Phytophthora infestans. J Exp Bot. 2009:1133–1140. doi: 10.1093/jxb/ern353. [DOI] [PubMed] [Google Scholar]
- Bittner-Eddy P.D, Beynon J.L. The Arabidopsis downy mildew resistance gene, RPP13-Nd, functions independently of NDR1 and EDS1 and does not require the accumulation of salicylic acid. Mol Plant Microbe Interact. 2001:416–421. doi: 10.1094/MPMI.2001.14.3.416. [DOI] [PubMed] [Google Scholar]
- Boevink P.C, McLellan H, Gilroy E.M, Naqvi S, He Q, Yang L, Wang X, Turnbull D, Armstrong M.R, Tian Z. Oomycetes seek help from the plant: Phytophthora infestans effectors target host susceptibility factors. Molecular Plant. 2016:636–638. doi: 10.1016/j.molp.2016.04.005. [DOI] [PubMed] [Google Scholar]
- Bohm H, Albert I, Oome S, Raaymakers T.M, Van den Ackerveken G, Nurnberger T. A conserved peptide pattern from a widespread microbial virulence factor triggers pattern-induced immunity in Arabidopsis. PLoS Pathogens. 2014 doi: 10.1371/journal.ppat.1004491. e1004491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonardi V, Tang S, Stallmann A, Roberts M, Cherkis K, Dangl J.L. Expanded functions for a family of plant intracellular immune receptors beyond specific recognition of pathogen effectors. Proceedings of the National Academy of Sciences. 2011:16463. doi: 10.1073/pnas.1113726108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borhan M.H, Brose E, Beynon J.L, Holub E.B. White rust (Albugo candida) resistance loci on three Arabidopsis chromosomes are closely linked to downy mildew (Peronospora parasitica) resistance loci. Mol Plant Pathol. 2001:87–95. doi: 10.1046/j.1364-3703.2001.00056.x. [DOI] [PubMed] [Google Scholar]
- Borhan M.H, Holub E.B, Beynon J.L, Rozwadowski K, Rimmer S.R. The Arabidopsis TIR-NB-LRR gene RAC1 confers resistance to Albugo candida (white rust) and is dependent on EDS1 but not PAD4. Mol Plant Microbe Interact. 2004:711–719. doi: 10.1094/MPMI.2004.17.7.711. [DOI] [PubMed] [Google Scholar]
- Borhan M.H, Holub E.B, Kindrachuk C, Omidi M, Bozorgmanesh-Frad G, Rimmer S.R. WRR4 a broad-spectrum TIR-NBLRR gene from Arabidopsis thaliana that confers white rust resistance in transgenic oilseed brassica crops. Mol Plant Pathol. 2010:283–291. doi: 10.1111/j.1364-3703.2009.00599.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borhan M.H, Gunn N, Cooper A, Gulden S, Tor M, Rimmer S.R, Holub E.B. WRR4 encodes a TIR-NB-LRR protein that confers broad-spectrum white rust resistance in Arabidopsis thaliana to four physiological races of Albugo candida. Mol Plant Microbe Interact. 2008:757–768. doi: 10.1094/MPMI-21-6-0757. [DOI] [PubMed] [Google Scholar]
- Botella M.A, Parker J.E, Frost L.N, Bittner-Eddy P.D, Beynon J.L, Daniels M.J, Holub E.B, Jones J.D. Three genes of the Arabidopsis RPP1 complex resistance locus recognize distinct Peronospora parasitica avirulence determinants. Plant Cell. 1998:1847–1860. doi: 10.1105/tpc.10.11.1847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouwmeester K, Meijer H.J, Govers F. At the frontier; RXLR effectors crossing the Phytophthorahost interface. Front Plant Sci. 2011a:75. doi: 10.3389/fpls.2011.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouwmeester K, de Sain M, Weide R, Gouget A, Klamer S, Canut H, Govers F. The lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential host target for a RXLR effector. PLoS Pathogens. 2011b doi: 10.1371/journal.ppat.1001327. e1001327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouwmeester K, Han M, Blanco-Portales R, Song W, Weide R, Guo L.Y, van der Vossen E.A, Govers F. The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal. 2014:10–16. doi: 10.1111/pbi.12111. [DOI] [PubMed] [Google Scholar]
- Boyle P, Le Su E, Rochon A, Shearer H.L, Murmu J, Chu J.Y, Fobert P.R, Despres C. The BTB/POZ domain of the Arabidopsis disease resistance protein NPR1 interacts with the repression domain of TGA2 to negate its function. Plant Cell. 2009:3700–3713. doi: 10.1105/tpc.109.069971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bredow M, Monaghan J. Regulation of plant immune signaling by calcium-dependent protein kinases. Mol Plant Microbe Interact. 2019:6–19. doi: 10.1094/MPMI-09-18-0267-FI. [DOI] [PubMed] [Google Scholar]
- Browse J, Vijayan P, Shockey J, Levesque C.A, Cook R.J. A role for jasmonate in pathogen defense of Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America. 1998:7209–7214. doi: 10.1073/pnas.95.12.7209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunner F, Rosahl S, Lee J, Rudd J.J, Geiler C, Kauppinen S, Rasmussen G, Scheel D, Nurnberger T. Pep-13, a plant defense-inducing pathogen-associated pattern from Phytophthora transglutaminases. Embo J. 2002:6681–6688. doi: 10.1093/emboj/cdf667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caillaud M.-C, Wirthmueller L, Sklenar J, Findlay K, Piquerez S.J, Jones A.M, Robatzek S, Jones J.D, Faulkner C. The plasmodesmal protein PDLP1 localises to haustoria-associated membranes during downy mildew infection and regulates callose deposition. PLoS Pathogens. 2014 doi: 10.1371/journal.ppat.1004496. e1004496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caillaud M.C, Asai S, Rallapalli G, Piquerez S, Fabro G, Jones J.D. A downy mildew effector attenuates salicylic acid-triggered immunity in Arabidopsis by interacting with the host mediator complex. PLoS Biology. 2013 doi: 10.1371/journal.pbio.1001732. e1001732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caillaud M.C, Piquerez S.J, Fabro G, Steinbrenner J, Ishaque N, Beynon J, Jones J.D. Subcellular localization of the Hpa RxLR effector repertoire identifies a tonoplast-associated protein HaRxL17 that confers enhanced plant susceptibility. Plant J. 2012:252–265. doi: 10.1111/j.1365-313X.2011.04787.x. [DOI] [PubMed] [Google Scholar]
- Cao H, Glazebrook J, Clarke J.D, Volko S, Dong X.N. The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell. 1997:57–63. doi: 10.1016/s0092-8674(00)81858-9. [DOI] [PubMed] [Google Scholar]
- Castel B, Ngou P.-M, Cevik V, Redkar A, Kim D.-S, Yang Y, Ding P, Jones J.D.G. Diverse NLR immune receptors activate defence via the RPW8-NLR NRG1. New Phytologist. 2019:966–980. doi: 10.1111/nph.15659. [DOI] [PubMed] [Google Scholar]
- Cevik V, Boutrot F, Apel W, Robert-Seilaniantz A, Furzer O.J, Redkar A, Castel B, Kover P.X, Prince D.C, Holub E.B, Jones J.D.G. Transgressive segregation reveals mechanisms of Arabidopsis immunity to Brassica-infecting races of white rust Albugo candida. Proceedings of the National Academy of Sciences. 2019:2767. doi: 10.1073/pnas.1812911116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen H.Y, Huh J.H, Yu Y.C, Ho L.H, Chen L.Q, Tholl D, Frommer W.B, Guo W.J. The Arabidopsis vacuolar sugar transporter SWEET2 limits carbon sequestration from roots and restricts Pythium infection. Plant J. 2015:1046–1058. doi: 10.1111/tpj.12948. [DOI] [PubMed] [Google Scholar]
- Chen L.-Q, Hou B.-H, Lalonde S, Takanaga H, Hartung M.L, Qu X.-Q, Guo W.-J, Kim J.-G, Underwood W, Chaudhuri B. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature. 2010:527–532. doi: 10.1038/nature09606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou H.M, Bundock N, Rolfe S.A, Scholes J.D. Infection of Arabidopsis thaliana leaves with Albugo candida (white blister rust) causes a reprogramming of host metabolism. Molecular Plant Pathology. 2000:99–113. doi: 10.1046/j.1364-3703.2000.00013.x. [DOI] [PubMed] [Google Scholar]
- Chou S, Krasileva K.V, Holton J.M, Steinbrenner A.D, Alber T, Staskawicz B.J. Hyaloperonospora arabidopsidis ATR1 effector is a repeat protein with distributed recognition surfaces. Proceedings of the National Academy of Sciences. 2011:13323–13328. doi: 10.1073/pnas.1109791108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark J, Spencer-Phillips P. In Encyclopedia of Microbiology. Academic Press, Inc.; 2000. Downy Mildews; pp. 117–129. [Google Scholar]
- Clay N.K, Adio A.M, Denoux C, Jander G, Ausubel F.M. Glucosinolate metabolites required for an Arabidopsis innate immune response. Science. 2009:95–101. doi: 10.1126/science.1164627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coates M.E, Beynon J.L. Hyaloperonospora arabidopsidis as a pathogen model. Annu Rev Phytopathol. 2010:329–345. doi: 10.1146/annurev-phyto-080508-094422. [DOI] [PubMed] [Google Scholar]
- Constantinescu O, Fatehi J. Peronosporalike fungi (Chromista, Peronosporales) parasitic on Brassicaceae and related hosts. Nova Hedwigia. 2002:291–338. [Google Scholar]
- Cooper A.J, Latunde-Dada A.O, Woods-Tor A, Lynn J, Lucas J.A, Crute I.R, Holub E.B. Basic compatibility of Albugo candida in Arabidopsis thaliana and Brassica juncea causes broad-spectrum suppression of innate immunity. Mol Plant Microbe Interact. 2008:745–756. doi: 10.1094/MPMI-21-6-0745. [DOI] [PubMed] [Google Scholar]
- Crute I.R, Holub E.B, Tor M, Brose E, Beynon J.L. The identification and mapping of loci in Arabidopsis thaliana for recognition of the fungal pathogens Peronospora parasitica (downy mildew) and Albugo candida (white blister) In: Nester E.W, Verma D.P.S, editors. In Advances in Molecular Genetics of Plant-Microbe Interactions. Dordrecht: Kluwer Academic; 1993. pp. 437–444. [Google Scholar]
- Crute I.R, Beynon J, Dangl J.L, Holub E.B, Mauch-Mani B, Slusarenko A, Staskawicz B.J, Ausubel F.M. Microbial pathogenesis of Arabidopsis. In: Meyerowitz E.M, Somerville C.R, editors. In Arabidopsis. Cold Spring Harbor: Cold Spring harbor Laboratory Press; 1994. pp. 705–748. [Google Scholar]
- Cui H, Tsuda K, Parker J.E. Effector-triggered immunity: from pathogen perception to robust defense. Annu Rev Plant Biol. 2015:487–511. doi: 10.1146/annurev-arplant-050213-040012. [DOI] [PubMed] [Google Scholar]
- Dalio R.J.D, Herlihy J, Oliveira T.S, McDowell J.M, Machado M.A.A. Effector biology in focus: a primer for computational prediction and functional characterization. Mol Plant Microbe Interact. 2018:22–33. doi: 10.1094/MPMI-07-17-0174-FI. [DOI] [PubMed] [Google Scholar]
- Dangl J.L, Holub E.B, Debener T, Lehnackers H, Ritter C, Crute I.R. Genetic definition of loci involved in Arabidopsis-pathogen interactions. In: Koncz C, Chua N.-H, Schell J, editors. In Methods in Arabidopsis Research. London: World Scientific Publishing Co.; 1992. pp. 393–418. [Google Scholar]
- Daniel R, Guest D. Defence responses induced by potassium phosphonate in Phytophthora palmivorachallenged Arabidopsis thaliana. Physiological and Molecular Plant Pathology. 2005:194–201. [Google Scholar]
- de Jonge R, Bolton M.D, Thomma B.P.H.J. How filamentous pathogens co-opt plants: the ins and outs of fungal effectors. Current Opinion in Plant Biology. 2011:400–406. doi: 10.1016/j.pbi.2011.03.005. [DOI] [PubMed] [Google Scholar]
- Deb D, Mackey D, Opiyo S.O, McDowell J.M. Application of alignment-free bioinformatics methods to identify an oomycete protein with structural and functional similarity to the bacterial AvrE effector protein. PloS One. 2018a doi: 10.1371/journal.pone.0195559. e0195559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deb D, Anderson R, Kin T.H.Y, Tyler B.M, McDowell J.M. Conserved RxLR effectors from oomycetes Hyaloperonospora arabidopsidis and Phytophthora sojae suppress PAMP- and effector-triggered immunity in diverse plants. Mol Plant Microbe Interact. 2018b:374–385. doi: 10.1094/MPMI-07-17-0169-FI. [DOI] [PubMed] [Google Scholar]
- Dempsey D.A, Vlot A.C, Wildermuth M.C, Klessig D.F. Salicylic Acid biosynthesis and metabolism. Arabidopsis Book. 2011 doi: 10.1199/tab.0156. e0156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derevnina L, Dagdas Y.F, De la Concepcion J.C, Bialas A, Kellner R, Petre B, Domazakis E, Du J, Wu C.-H, Lin X, Aguilera-Galvez C, Cruz-Mireles N, Vleeshouwers V.G.A.A, Kamoun S. Nine things to know about elicitins. New Phytologist. 2016:888–895. doi: 10.1111/nph.14137. [DOI] [PubMed] [Google Scholar]
- Dick M. Straminipilous Fungi: Systematics of the Peronosporomycetes Including Accounts of the Marine Straminipilous Protists, the Plasmodiophorids and Similar Organisms. Dordrecht / Boston / London: Kluwer Academic Publishers; 2001. [Google Scholar]
- Ding Y, Sun T, Ao K, Peng Y, Zhang Y, Li X, Zhang Y. Opposite Roles of Salicylic Acid Receptors NPR1 and NPR3/NPR4 in Transcriptional Regulation of Plant Immunity. Cell. 2018:1454–1467.e1415. doi: 10.1016/j.cell.2018.03.044. [DOI] [PubMed] [Google Scholar]
- Dixon R.A. Natural products and plant disease resistance. Nature. 2001:843–847. doi: 10.1038/35081178. [DOI] [PubMed] [Google Scholar]
- Donofrio N.M, Delaney T.P. Abnormal callose response phenotype and hypersusceptibility to Peronospora parasitica in defense-compromised Arabidopsis nim1-1 and salicylate hydroxylase-expressing plants. Molecular Plant-Microbe Interactions. 2001:439–450. doi: 10.1094/MPMI.2001.14.4.439. [DOI] [PubMed] [Google Scholar]
- Durán P, Thiergart T, Garrido-Oter R, Agler M, Kemen E, Schulze-Lefert P, Hacquard S. Microbial interkingdom interactions in roots promote Arabidopsis survival. Cell. 2018:973–983.e914. doi: 10.1016/j.cell.2018.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evangelisti E, Govetto B, Minet-Kebdani N, Kuhn M.L, Attard A, Ponchet M, Panabieres F, Gourgues M. The Phytophthora parasitica RXLR effector penetration-specific effector 1 favours Arabidopsis thaliana infection by interfering with auxin physiology. New Phytol. 2013:476–489. doi: 10.1111/nph.12270. [DOI] [PubMed] [Google Scholar]
- Fabro G, Steinbrenner J, Coates M, Ishaque N, Baxter L, Studholme D.J, Korner E, Allen R.L, Piquerez S.J, Rougon-Cardoso A, Greenshields D, Lei R, Badel J.L, Caillaud M.C, Sohn K.H, Van den Ackerveken G, Parker J.E, Beynon J, Jones J.D. Multiple candidate effectors from the oomycete pathogen Hyaloperonospora arabidopsidis suppress host plant immunity. PLoS Pathogens. 2011 doi: 10.1371/journal.ppat.1002348. e1002348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fawke S, Doumane M, Schornack S. Oomycete interactions with plants: infection strategies and resistance principles. Microbiology and Molecular Biology Reviews : MMBR. 2015:263–280. doi: 10.1128/MMBR.00010-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feys B.J, Parker J.E. Interplay of signaling pathways in plant disease resistance. Trends Genet. 2000:449–455. doi: 10.1016/s0168-9525(00)02107-7. [DOI] [PubMed] [Google Scholar]
- Fliegmann J, Mithofer A, Wanner G, Ebel J. An ancient enzyme domain hidden in the putative beta-glucan elicitor receptor of soybean may play an active part in the perception of pathogen-associated molecular patterns during broad host resistance. J Biol Chem. 2004:1132–1140. doi: 10.1074/jbc.M308552200. [DOI] [PubMed] [Google Scholar]
- Fry W. Phytophthora infestans the plant (and R gene) destroyer. Molecular Plant Pathology. 2008:385–402. doi: 10.1111/j.1364-3703.2007.00465.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furci L, Jain R, Stassen J, Berkowitz O, Whelan J, Roquis D, Baillet V, Colot V, Johannes F, Ton J. Identification and characterisation of hypomethylated DNA loci controlling quantitative resistance in Arabidopsis. Elife. 2019 doi: 10.7554/eLife.40655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamir J, Darwiche R, Van’t Hof P, Choudhary V, Stumpe M, Schneiter R, Mauch F. The sterol-binding activity of PATHOGENESIS-RELATED PROTEIN 1 reveals the mode of action of an antimicrobial protein. Plant J. 2017:502–509. doi: 10.1111/tpj.13398. [DOI] [PubMed] [Google Scholar]
- Garcion C, Lohmann A, Lamodiere E, Catinot J, Buchala A, Doermann P, Metraux J.P. Characterization and biological function of the ISOCHORISMATE SYNTHASE2 gene of Arabidopsis. Plant Physiol. 2008:1279–1287. doi: 10.1104/pp.108.119420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geraats B.P, Bakker P.A, van Loon L.C. Ethylene insensitivity impairs resistance to soilborne pathogens in tobacco and Arabidopsis thaliana. Mol Plant Microbe Interact. 2002:1078–1085. doi: 10.1094/MPMI.2002.15.10.1078. [DOI] [PubMed] [Google Scholar]
- Gimenez-Ibanez S, Chini A, Solano R. How microbes twist jasmonate signaling around their little fingers. Plants (Basel) 2016 doi: 10.3390/plants5010009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gisi U. Chemical Control of Downy Mildews. In: Spencer-Phillips P, Gisi U, Lebeda A, editors. In Advances in Downy Mildew Research. Dordrecht, NE: Kluwer Academic Publishers; 2002. pp. 119–160. [Google Scholar]
- Glawischnig E. Camalexin. Phytochemistry. 2007:401–406. doi: 10.1016/j.phytochem.2006.12.005. [DOI] [PubMed] [Google Scholar]
- Goker M, Voglmayr H, Blazquez G.G, Oberwinkler F. Species delimitation in downy mildews: the case of Hyaloperonospora in the light of nuclear ribosomal ITS and LSU sequences. Mycol Res. 2009:308–325. doi: 10.1016/j.mycres.2008.11.006. [DOI] [PubMed] [Google Scholar]
- Gomez-Gomez L, Boller T. FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Molecular Cell. 2000:1003–1011. doi: 10.1016/s1097-2765(00)80265-8. [DOI] [PubMed] [Google Scholar]
- Goritschnig S, Krasileva K.V, Dahlbeck D, Staskawicz B.J. Computational prediction and molecular characterization of an oomycete effector and the cognate Arabidopsis resistance gene. PLoS Genet. 2012 doi: 10.1371/journal.pgen.1002502. e1002502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goritschnig S, Steinbrenner A.D, Grunwald D.J, Staskawicz B.J. Structurally distinct Arabidopsis thaliana NLR immune receptors recognize tandem WY domains of an oomycete effector. New Phytol. 2016:984–996. doi: 10.1111/nph.13823. [DOI] [PubMed] [Google Scholar]
- Govindarajulu M, Epstein L, Wroblewski T, Michelmore R.W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. 2015:875–883. doi: 10.1111/pbi.12307. [DOI] [PubMed] [Google Scholar]
- Grenville-Briggs L.J, Horner N.R, Phillips A.J, Beakes G.W, van West P. A family of small tyrosine rich proteins is essential for oogonial and oospore cell wall development of the mycoparasitic oomycete Pythium oligandrum. Fungal Biol. 2013:163–172. doi: 10.1016/j.funbio.2013.01.001. [DOI] [PubMed] [Google Scholar]
- Gunn N.D, Byrne J, Holub E.B. Outcrossing of Two Homothallic Isolates of Peronospora parasitica and Segregation of Avirulence Matching Six Resistance Loci in Arabidopsis thaliana. In: Spencer-Phillips P, Gisi U, Lebeda A, editors. In Advances in Downy Mildew Research. Dordrecht, NE: Kluwer Academic Publishers; 2002. pp. 185–188. [Google Scholar]
- Gust A.A, Felix G. Receptor like proteins associate with SOBIR1-type of adaptors to form bimolecular receptor kinases. Current Opinion In Plant Biology. 2014:104–111. doi: 10.1016/j.pbi.2014.07.007. [DOI] [PubMed] [Google Scholar]
- Halane M.K, Kim S.H, Spears B.J, Garner C.M, Rogan C.J, Okafor E.C, Su J, Bhattacharjee S, Gassmann W. The bacterial type III-secreted protein AvrRps4 is a bipartite effector. PLoS Pathogens. 2018 doi: 10.1371/journal.ppat.1006984. e1006984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardham A.R, Takemoto D, White R.G. Rapid and dynamic subcellular reorganization following mechanical stimulation of Arabidopsis epidermal cells mimics responses to fungal and oomycete attack. BMC Plant Biology. 2008:63. doi: 10.1186/1471-2229-8-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hein I, Gilroy E.M, Armstrong M.R, Birch P.R.J. The zig-zag-zig in oomycete-plant interactions. Molecular Plant Pathology. 2009:547–562. doi: 10.1111/j.1364-3703.2009.00547.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermanns M, Slusarenko A.J, Schlaich N.L. Organ-specificity in a plant disease is determined independently of R gene signaling. Molecular Plant-Microbe Interactions. 2003:752–759. doi: 10.1094/MPMI.2003.16.9.752. [DOI] [PubMed] [Google Scholar]
- Hok S, Danchin E.G, Allasia V, Panabieres F, Attard A, Keller H. An Arabidopsis (malectin-like) leucine-rich repeat receptor-like kinase contributes to downy mildew disease. Plant, Cell & Environment. 2011:1944–1957. doi: 10.1111/j.1365-3040.2011.02390.x. [DOI] [PubMed] [Google Scholar]
- Hok S, Allasia V, Andrio E, Naessens E, Ribes E, Panabieres F, Attard A, Ris N, Clement M, Barlet X, Marco Y, Grill E, Eichmann R, Weis C, Huckelhoven R, Ammon A, Ludwig-Muller J, Voll L.M, Keller H. The receptor kinase IMPAIRED OOMYCETE SUSCEPTIBILITY1 attenuates abscisic acid responses in Arabidopsis. Plant Physiol. 2014:1506–1518. doi: 10.1104/pp.114.248518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holub E, Brose E, Tör M, Clay C, Crute I.R, Beynon J.L. Phenotypic and genotypic variation in the interaction between Arabidopsis thaliana and Albugo candida. Molec. Plant-Microbe Interact. 1994a:916–928. doi: 10.1094/mpmi-8-0916. [DOI] [PubMed] [Google Scholar]
- Holub E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001:516–527. doi: 10.1038/35080508. [DOI] [PubMed] [Google Scholar]
- Holub E.B. Natural history of Arabidopsis thaliana and oomycete symbioses. Eur J Plant Pathol. 2008:91–109. [Google Scholar]
- Holub E.B, Beynon J.L. Symbiology of mouse ear cress (Arabidopsis thaliana) and oomycetes. Adv. Bot. Res. 1996:228–273. [Google Scholar]
- Holub E.B, Beynon J.L, Crute I.R. Phenotypic and genotypic characterization of interactions between isolates of Peronospora parasitica and accessions of Arabidopsis thaliana. Mol Plant-Microbe Interact. 1994b:223–239. [Google Scholar]
- Hou Y, Ma W. Small RNA and mRNA profiling of Arabidopsis in response to Phytophthora infection and PAMP treatment. Methods Mol Biol. 2017:273–283. doi: 10.1007/978-1-4939-6859-6_23. [DOI] [PubMed] [Google Scholar]
- Huffaker A, Ryan C.A. Endogenous peptide defense signals in Arabidopsis differentially amplify signaling for the innate immune response. Proceedings of the National Academy of Sciences of the United States of America. 2007:10732–10736. doi: 10.1073/pnas.0703343104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huffaker A, Pearce G, Ryan C.A. An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proceedings of the National Academy of Sciences of the United States of America. 2006:10098–10103. doi: 10.1073/pnas.0603727103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huh S.U, Cevik V, Ding P, Duxbury Z, Ma Y, Tomlinson L, Sarris P.F, Jones J.D.G. Protein-protein interactions in the RPS4/RRS1 immune receptor complex. PLoS pathogens. 2017 doi: 10.1371/journal.ppat.1006376. e1006376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huitema E, Vleeshouwers V.G, Francis D.M, Kamoun S. Active defence responses associated with non-host resistance of Arabidopsis thaliana to the oomycete pathogen Phytophthora infestans. Mol Plant Pathol. 2003:487–500. doi: 10.1046/j.1364-3703.2003.00195.x. [DOI] [PubMed] [Google Scholar]
- Innes R. The positives and negatives of NPR: A unifying model for salicylic acid signaling in plants. Cell. 2018:1314–1315. doi: 10.1016/j.cell.2018.05.034. [DOI] [PubMed] [Google Scholar]
- Jiang R.H, Tyler B.M. Mechanisms and evolution of virulence in oomycetes. Annu Rev Phytopathol. 2012:295–318. doi: 10.1146/annurev-phyto-081211-172912. [DOI] [PubMed] [Google Scholar]
- Jones J.D.G, Dangl J.L. The plant immune system. Nature. 2006:323–329. doi: 10.1038/nature05286. [DOI] [PubMed] [Google Scholar]
- Jouet A, Saunders D.G.O, McMullan M, Ward B, Furzer O, Jupe F, Cevik V, Hein I, Thilliez G.J.A, Holub E, van Oosterhout C, Jones J.D.G. Albugo candida race diversity, ploidy and host-associated microbes revealed using DNA sequence capture on diseased plants in the field. New Phytol. 2018:1529–1543. doi: 10.1111/nph.15417. [DOI] [PubMed] [Google Scholar]
- Kale S.D, Tyler B.M. Entry of oomycete and fungal effectors into plant and animal host cells. Cell Microbiol. 2011:1839–1848. doi: 10.1111/j.1462-5822.2011.01659.x. [DOI] [PubMed] [Google Scholar]
- Kale S.D, Gu B, Capelluto D.G, Dou D, Feldman E, Rumore A, Arredondo F.D, Hanlon R, Fudal I, Rouxel T, Lawrence C.B, Shan W, Tyler B.M. External lipid PI3P mediates entry of eukaryotic pathogen effectors into plant and animal host cells. Cell. 2010:284–295. doi: 10.1016/j.cell.2010.06.008. [DOI] [PubMed] [Google Scholar]
- Kamoun S. A catalogue of the effector secretome of plant pathogenic oomycetes. Annu Rev Phytopathol. 2006:41–60. doi: 10.1146/annurev.phyto.44.070505.143436. [DOI] [PubMed] [Google Scholar]
- Kamoun S, Furzer O, Jones J.D, Judelson H.S, Ali G.S, Dalio R.J, Roy S.G, Schena L, Zambounis A, Panabieres F, Cahill D, Ruocco M, Figueiredo A, Chen X.R, Hulvey J, Stam R, Lamour K, Gijzen M, Tyler B.M, Grunwald N.J, Mukhtar M.S, Tome D.F, Tor M, Van Den Ackerveken G, McDowell J, Daayf F, Fry W.E, Lindqvist-Kreuze H, Meijer H.J, Petre B, Ristaino J, Yoshida K, Birch P.R, Govers F. The Top 10 oomycete pathogens in molecular plant pathology. Mol Plant Pathol. 2015:413–434. doi: 10.1111/mpp.12190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karasov T.L, Almario J, Friedemann C, Ding W, Giolai M, Heavens D, Kersten S, Lundberg D.S, Neumann M, Regalado J, Neher R.A, Kemen E, Weigel D. Arabidopsis thaliana and Pseudomonas pathogens exhibit stable associations over evolutionary timescales. Cell Host & Microbe. 2018:168–179.e164. doi: 10.1016/j.chom.2018.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemen E, Jones J.D. Obligate biotroph parasitism: can we link genomes to lifestyles? Trends Plant Sci. 2012:448–457. doi: 10.1016/j.tplants.2012.04.005. [DOI] [PubMed] [Google Scholar]
- Kemen E, Gardiner A, Schultz-Larsen T, Kemen A.C, Balmuth A.L, Robert-Seilaniantz A, Bailey K, Holub E, Studholme D.J, Maclean D, Jones J.D. Gene gain and loss during evolution of obligate parasitism in the white rust pathogen of Arabidopsis thaliana. PLoS biology. 2011 doi: 10.1371/journal.pbio.1001094. e1001094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khatib M, Lafitte C, Esquerre-Tugaye M.T, Bottin A, Rickauer M. The CBEL elicitor of Phytophthora parasitica var. nicotianae activates defence in Arabidopsis thaliana via three different signalling pathways. New Phytologist. 2004:501–510. [Google Scholar]
- Klessig D.F, Choi H.W, Dempsey D.A. Systemic acquired resistance and salicylic acid: past, present and future. Mol Plant Microbe Interact. 2018 doi: 10.1094/MPMI-03-18-0067-CR. [DOI] [PubMed] [Google Scholar]
- Koch E, Slusarenko A. Arabidopsis is susceptible to infection by a downy mildew fungus. Plant Cell. 1990:437–445. doi: 10.1105/tpc.2.5.437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koeck M, Hardham A.R, Dodds P.N. The role of effectors of biotrophic and hemibiotrophic fungi in infection. Cell Microbiol. 2011:1849–1857. doi: 10.1111/j.1462-5822.2011.01665.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krasileva K.V, Dahlbeck D, Staskawicz B.J. Activation of an Arabidopsis resistance protein is specified by the in planta association of its leucine-rich repeat domain with the cognate oomycete effector. Plant Cell. 2010:2444–2458. doi: 10.1105/tpc.110.075358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krasileva K.V, Zheng C, Leonelli L, Goritschnig S, Dahlbeck D, Staskawicz B.J. Global analysis of Arabidopsis/downy mildew interactions reveals prevalence of incomplete resistance and rapid evolution of pathogen recognition. PloS One. 2011 doi: 10.1371/journal.pone.0028765. e28765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroon L.P, Brouwer H, de Cock A.W, Govers F. The genus Phytophthora anno 2012. Phytopathology. 2012:348–364. doi: 10.1094/PHYTO-01-11-0025. [DOI] [PubMed] [Google Scholar]
- Kunze G, Zipfel C, Robatzek S, Niehaus K, Boller T, Felix G. The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant Cell. 2004:3496–3507. doi: 10.1105/tpc.104.026765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacombe S, Rougon-Cardoso A, Sherwood E, Peeters N, Dahlbeck D, van Esse H.P, Smoker M, Rallapalli G, Thomma B.P.H.J, Staskawicz B, Jones J.D.G, Zipfel C. Inter-family transfer of a plant pattern-recognition receptor confers broad-spectrum bacterial resistance. Nat Biotechnol. 2010:365-U394. doi: 10.1038/nbt.1613. [DOI] [PubMed] [Google Scholar]
- Lai Y, Eulgem T. Transcript-level expression control of plant NLR genes. Molecular Plant Pathology. 2018:1267–1281. doi: 10.1111/mpp.12607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai Y, Cuzick A, Lu X.M, Wang J, Katiyar N, Tsuchiya T, Le Roch K, McDowell J.M, Holub E, Eulgem T. The Arabidopsis RRM domain protein EDM3 mediates race-specific disease resistance by controlling H3K9me2-dependent alternative polyadenylation of RPP7 immune receptor transcripts. Plant J. 2019:646–660. doi: 10.1111/tpj.14148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamour K, Hu J, Lefebvre V, Mudge J, Howden A, Huitema E. Illuminating the Phytophthora capsici Genome. In: Dean R.A, Lichens-Park A, Kole C, editors. In Genomics of Plant-Associated Fungi and Oomycetes: Dicot Pathogens. Springer Berlin Heidelberg; 2014. pp. 121–132. [Google Scholar]
- Lamour K.H, Win J, Kamoun S. Oomycete genomics: new insights and future directions. FEMS Microbiol Lett. 2007:1–8. doi: 10.1111/j.1574-6968.2007.00786.x. [DOI] [PubMed] [Google Scholar]
- Lamour K.H, Mudge J, Gobena D, Hurtado-Gonzales O.P, Schmutz J, Kuo A, Miller N.A, Rice B.J, Raffaele S, Cano L.M, Bharti A.K, Donahoo R.S, Finley S, Huitema E, Hulvey J, Platt D, Salamov A, Savidor A, Sharma R, Stam R, Storey D, Thines M, Win J, Haas B.J, Dinwiddie D.L, Jenkins J, Knight J.R, Affourtit J.P, Han C.S, Chertkov O, Lindquist E.A, Detter C, Grigoriev I.V, Kamoun S, Kingsmore S.F. Genome sequencing and mapping reveal loss of heterozygosity as a mechanism for rapid adaptation in the vegetable pathogen Phytophthora capsici. Mol Plant Microbe Interact. 2012:1350–1360. doi: 10.1094/MPMI-02-12-0028-R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapin D, Van den Ackerveken G. Susceptibility to plant disease: more than a failure of host immunity. Trends Plant Sci. 2013:546–554. doi: 10.1016/j.tplants.2013.05.005. [DOI] [PubMed] [Google Scholar]
- Lapin D, Meyer R.C, Takahashi H, Bechtold U, Van den Ackerveken G. Broad-spectrum resistance of Arabidopsis C24 to downy mildew is mediated by different combinations of isolate-specific loci. New Phytol. 2012:1171–1181. doi: 10.1111/j.1469-8137.2012.04344.x. [DOI] [PubMed] [Google Scholar]
- Lapin D, Kovacova V, Sun X, Dongus J, Bhandari D.D, Born P.v., Bautor J, Guarneri N, Stuttmann J, Beyer A, Parker J.E. A coevolved EDS1-SAG101-NRG1 module mediates cell death signaling by TIR-domain immune receptors. bioRxiv. 2019:572826. doi: 10.1105/tpc.19.00118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Berre J.Y, Engler G, Panabieres F. Exploration of the late stages of the tomato-Phytophthora parasitica interactions through histological analysis and generation of expressed sequence tags. New Phytol. 2008:480–492. doi: 10.1111/j.1469-8137.2007.02269.x. [DOI] [PubMed] [Google Scholar]
- Leonelli L, Pelton J, Schoeffler A, Dahlbeck D, Berger J, Wemmer D.E, Staskawicz B. Structural elucidation and functional characterization of the Hyaloperonospora arabidopsidis effector protein ATR13. PLoS Pathogens. 2011 doi: 10.1371/journal.ppat.1002428. e1002428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levesque C.A, Brouwer H, Cano L, Hamilton J.P, Holt C, Huitema E, Raffaele S, Robideau G.P, Thines M, Win J, Zerillo M.M, Beakes G.W, Boore J.L, Busam D, Dumas B, Ferriera S, Fuerstenberg S.I, Gachon C.M, Gaulin E, Govers F, Grenville-Briggs L, Horner N, Hostetler J, Jiang R.H, Johnson J, Krajaejun T, Lin H, Meijer H.J, Moore B, Morris P, Phuntmart V, Puiu D, Shetty J, Stajich J.E, Tripathy S, Wawra S, van West P, Whitty B.R, Coutinho P.M, Henrissat B, Martin F, Thomas P.D, Tyler B.M, De Vries R.P, Kamoun S, Yandell M, Tisserat N, Buell C.R. Genome sequence of the necrotrophic plant pathogen Pythium ultimum reveals original pathogenicity mechanisms and effector repertoire. Genome Biol. 2010:R73. doi: 10.1186/gb-2010-11-7-r73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Links M.G, Holub E, Jiang R.H, Sharpe A.G, Hegedus D, Beynon E, Sillito D, Clarke W.E, Uzuhashi S, Borhan M.H. De novo sequence assembly of Albugo candida reveals a small genome relative to other biotrophic oomycetes. BMC Genomics. 2011:503. doi: 10.1186/1471-2164-12-503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu D, Raghothama K.G, Hasegawa P.M, Bressan R.A. Osmotin overexpression in potato delays development of disease symptoms. Proceedings of the National Academy of Sciences of the United States of America. 1994:1888–1892. doi: 10.1073/pnas.91.5.1888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Y.J, Schornack S, Spallek T, Geldner N, Chory J, Schellmann S, Schumacher K, Kamoun S, Robatzek S. Patterns of plant subcellular responses to successful oomycete infections reveal differences in host cell reprogramming and endocytic trafficking. Cellular Microbiology. 2012:682–697. doi: 10.1111/j.1462-5822.2012.01751.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucas J, Hayter J, Crute I. The Downy Mildews: Host Specificity and Pathogenesis. In: Singh U, Singh R, editors. In Pathogenesis and Host Specificity in Plant Diseases. UK: Permagon; 1995. pp. 217–234. [Google Scholar]
- Ma Z, Song T, Zhu L, Ye W, Wang Y, Shao Y, Dong S, Zhang Z, Dou D, Zheng X, Tyler B.M, Wang Y. A Phytophthora sojae Glycoside Hydrolase 12 Protein Is a Major Virulence Factor during Soybean Infection and Is Recognized as a PAMP. Plant Cell. 2015:2057–2072. doi: 10.1105/tpc.15.00390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macho A.P, Zipfel C. Plant PRRs and the activation of innate immune signaling. Molecular Cell. 2014:263–272. doi: 10.1016/j.molcel.2014.03.028. [DOI] [PubMed] [Google Scholar]
- Martin F.N, Loper J.E. Soilborne Plant Diseases Caused by Pythium spp.: Ecology, Epidemiology, and Prospects for Biological Control. Critical Reviews in Plant Sciences. 1999:111–181. [Google Scholar]
- Mauch F, Torche S, Schläppi K, Branciard L, Belhaj K, Parisy V, Si-Ammour A. Phytophthora brassicae as a pathogen of Arabidopsis. Oomycete Genetics and Genomics. John Wiley & Sons, Inc.; 2008. [Google Scholar]
- McDowell J, Simon S. Recent insights into R gene evolution. Molecular Plant Pathology. 2006:437–448. doi: 10.1111/j.1364-3703.2006.00342.x. [DOI] [PubMed] [Google Scholar]
- McDowell J.M. Genomes of obligate plant pathogens reveal adaptations for obligate parasitism. Proceedings of the National Academy of Sciences of the United States of America. 2011:8921–8922. doi: 10.1073/pnas.1105802108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.M. Hyaloperonospora arabidopsidis A Model Pathogen of Arabidopsis. In: Dean R.A, Lichens-Park A, Kole C, editors. In Genomics of Plant-Associated Fungi and Oomycetes: Dicot Pathogens. Springer Berlin Heidelberg; 2014. pp. 209–234. [Google Scholar]
- McDowell J.M, Meyers B.C. A transposable element is domesticated for service in the plant immune system. Proceedings of the National Academy of Sciences of the United States of America. 2013:14821–14822. doi: 10.1073/pnas.1314089110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.M, Hoff T, Anderson R.G, Deegan D. Propagation, storage, and assays with Hyaloperonospora arabidopsidis A model oomycete pathogen of Arabidopsis. Methods Mol Biol. 2011:137–151. doi: 10.1007/978-1-61737-998-7_12. [DOI] [PubMed] [Google Scholar]
- McDowell J.M, Cuzick A, Can C, Beynon J, Dangl J.L, Holub E.B. Downy mildew (Peronospora parasitica) resistance genes in Arabidopsis vary in functional requirements for NDR1 EDS1 NPR1 and salicylic acid accumulation. Plant J. 2000:523–529. doi: 10.1046/j.1365-313x.2000.00771.x. [DOI] [PubMed] [Google Scholar]
- McDowell J.M, Dhandaydham M, Long T.A, Aarts M.G, Goff S, Holub E.B, Dangl J.L. Intragenic recombination and diversifying selection contribute to the evolution of downy mildew resistance at the RPP8 locus of Arabidopsis. Plant Cell. 1998:1861–1874. doi: 10.1105/tpc.10.11.1861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McMullan M, Gardiner A, Bailey K, Kemen E, Ward B.J, Cevik V, Robert-Seilaniantz A, Schultz-Larsen T, Balmuth A, Holub E, van Oosterhout C, Jones J.D. Evidence for suppression of immunity as a driver for genomic introgressions and host range expansion in races of Albugo candida a generalist parasite. Elife. 2015 doi: 10.7554/eLife.04550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng Y, Zhang Q, Ding W, Shan W. Phytophthora parasitica a model oomycete plant pathogen. Mycology. 2014:43–51. doi: 10.1080/21501203.2014.917734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng Y, Huang Y, Wang Q, Wen Q, Jia J, Zhang Q, Huang G, Quan J, Shan W. Phenotypic and genetic characterization of resistance in Arabidopsis thaliana to the oomycete pathogen Phytophthora parasitica. Front Plant Sci. 2015:378. doi: 10.3389/fpls.2015.00378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michelmore R, Coaker G, Bart R, Beattie G, Bent A, Bruce T, Cameron D, Dangl J, Dinesh-Kumar S, Edwards R, Evesvan den Akker S, Gassmann W, Greenberg J.T, Hanley-Bowdoin L, Harrison R.J, Harvey J, He P, Huffaker A, Hulbert S, Innes R, Jones J.D.G, Kaloshian I, Kamoun S, Katagiri F, Leach J, Ma W, McDowell J, Medford J, Meyers B, Nelson R, Oliver R, Qi Y, Saunders D, Shaw M, Smart C, Subudhi P, Torrance L, Tyler B, Valent B, Walsh J. Foundational and translational research opportunities to improve plant health. Mol Plant Microbe Interact. 2017:515–516. doi: 10.1094/MPMI-01-17-0010-CR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mims C.W, Richardson E.A, Holt Iii B.F, Dangl J.L. Ultrastructure of the host–pathogen interface in Arabidopsis thaliana leaves infected by the downy mildew Hyaloperonospora parasitica. Canadian Journal of Botany. 2004:1001–1008. [Google Scholar]
- Mindrinos M, Katagiri F, Yu G.L, Ausubel F.M. The A. thaliana disease resistance gene RPS2 encodes a protein containing a nucleotide-binding site and leucine-rich repeats. Cell. 1994:1089–1099. doi: 10.1016/0092-8674(94)90282-8. [DOI] [PubMed] [Google Scholar]
- Mishina T.E, Zeier J. Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in Arabidopsis. Plant J. 2007:500–513. doi: 10.1111/j.1365-313X.2007.03067.x. [DOI] [PubMed] [Google Scholar]
- Miya A, Albert P, Shinya T, Desaki Y, Ichimura K, Shirasu K, Narusaka Y, Kawakami N, Kaku H, Shibuya N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America. 2007:19613–19618. doi: 10.1073/pnas.0705147104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohr T.J, Mammarella N.D, Hoff T, Woffenden B.J, Jelesko J.G, McDowell J.M. The Arabidopsis downy mildew resistance gene RPP8 is induced by pathogens and salicylic acid and is regulated by W box cis elements. Mol Plant Microbe Interact. 2010:1303–1315. doi: 10.1094/MPMI-01-10-0022. [DOI] [PubMed] [Google Scholar]
- Mukhtar M.S, Carvunis A.R, Dreze M, Epple P, Steinbrenner J, Moore J, Tasan M, Galli M, Hao T, Nishimura M.T, Pevzner S.J, Donovan S.E, Ghamsari L, Santhanam B, Romero V, Poulin M.M, Gebreab F, Gutierrez B.J, Tam S, Monachello D, Boxem M, Harbort C.J, McDonald N, Gai L, Chen H, He Y, Vandenhaute J, Roth F.P, Hill D.E, Ecker J.R, Vidal M, Beynon J, Braun P, Dangl J.L. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science. 2011:596–601. doi: 10.1126/science.1203659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naito K, Taguchi F, Suzuki T, Inagaki Y, Toyoda K, Shiraishi T, Ichinose Y. Amino acid sequence of bacterial microbe-associated molecular pattern flg22 is required for virulence. Molecular Plant-Microbe Interactions. 2008:1165–1174. doi: 10.1094/MPMI-21-9-1165. [DOI] [PubMed] [Google Scholar]
- Narasimhan M.L, Coca M.A, Jin J, Yamauchi T, Ito Y, Kadowaki T, Kim K.K, Pardo J.M, Damsz B, Hasegawa P.M, Yun D.J, Bressan R.A. Osmotin is a homolog of mammalian adiponectin and controls apoptosis in yeast through a homolog of mammalian adiponectin receptor. Molecular Cell. 2005:171–180. doi: 10.1016/j.molcel.2004.11.050. [DOI] [PubMed] [Google Scholar]
- Nemri A, Atwell S, Tarone A.M, Huang Y.S, Zhao K, Studholme D.J, Nordborg M, Jones J.D. Genome-wide survey of Arabidopsis natural variation in downy mildew resistance using combined association and linkage mapping. Proceedings of the National Academy of Sciences of the United States of America. 2010:10302–10307. doi: 10.1073/pnas.0913160107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niderman T, Genetet I, Bruyere T, Gees R, Stintzi A, Legrand M, Fritig B, Mosinger E. Pathogenesis-related PR-1 proteins are antifungal. Isolation and characterization of three 14-kilo-dalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans. Plant Physiol. 1995:17–27. doi: 10.1104/pp.108.1.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noel L, Moores T.L, van Der Biezen E.A, Parniske M, Daniels M.J, Parker J.E, Jones J.D. Pronounced intraspecific haplotype divergence at the RPP5 complex disease resistance locus of Arabidopsis. Plant Cell. 1999:2099–2112. [PMC free article] [PubMed] [Google Scholar]
- Nurnberger T, Kemmerling B. PAMP-triggered basal immunity in plants. Adv Bot Res. 2009:1–38. [Google Scholar]
- O’Connell R.J, Panstruga R. Tete a tete inside a plant cell: establishing compatibility between plants and biotrophic fungi and oomycetes. New Phytol. 2006:699–718. doi: 10.1111/j.1469-8137.2006.01829.x. [DOI] [PubMed] [Google Scholar]
- Oome S, Van den Ackerveken G. Comparative and functional analysis of the widely occurring family of Nep1-like proteins. Mol Plant Microbe Interact. 2014:1081–1094. doi: 10.1094/MPMI-04-14-0118-R. [DOI] [PubMed] [Google Scholar]
- Oome S, Raaymakers T.M, Cabral A, Samwel S, Bohm H, Albert I, Nurnberger T, Van den Ackerveken G. Nep1-like proteins from three kingdoms of life act as a microbe-associated molecular pattern in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America. 2014:16955–16960. doi: 10.1073/pnas.1410031111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pais M, Win J, Yoshida K, Etherington G.J, Cano L.M, Raffaele S, Banfield M.J, Jones A, Kamoun S, Saunders D.G. From pathogen genomes to host plant processes: the power of plant parasitic oomycetes. Genome Biol. 2013:211. doi: 10.1186/gb-2013-14-6-211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panabieres F, Ali G.S, Allagui M.B, Dalio R.J.D, Gudmestad N.C, Kuhn M.-L, Guha Roy S, Schena L, Zampounis A. Phytophthora nicotianae diseases worldwide: new knowledge of a long-recognised pathogen. Phytopathologia Mediterranea. 2016;Vol 55(No 1) (2016) [Google Scholar]
- Parker J.E, Holub E.B, Frost L.N, Falk A, Gunn N.D, Daniels M.J. Characterization of eds1, a mutation in Arabidopsis suppressing resistance to Peronospora parasitica specified by several different RPP genes. Plant Cell. 1996:2033–2046. doi: 10.1105/tpc.8.11.2033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker J.E, Coleman M.J, Szabo V, Frost L.N, Schmidt R, van der Biezen E, Moores T, Dean C, Daniels M.J, Jones J.D.G. The Arabidopsis downy mildew resistance gene Rpp5 shares similarity to the Toll and interleukin-1 receptors with N and L6. Plant Cell. 1997:879–894. doi: 10.1105/tpc.9.6.879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pel M.J, Wintermans P.C, Cabral A, Robroek B.J, Seidl M.F, Bautor J, Parker J.E, Van den Ackerveken G, Pieterse C.M. Functional Analysis of Hyaloperonospora arabidopsidis RXLR Effectors. PloS one. 2014 doi: 10.1371/journal.pone.0110624. e110624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petre B, Kamoun S. How do filamentous pathogens deliver effector proteins into plant cells? PLoS Biology. 2014 doi: 10.1371/journal.pbio.1001801. e1001801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pieterse C.M, Van Loon L.C. NPR1: the spider in the web of induced resistance signaling pathways. Current Opinion In Plant Biology. 2004:456–464. doi: 10.1016/j.pbi.2004.05.006. [DOI] [PubMed] [Google Scholar]
- Pieterse C.M, Van der Does D, Zamioudis C, Leon-Reyes A, Van Wees S.C. Hormonal modulation of plant immunity. Annu Rev Cell Dev Biol. 2012:489–521. doi: 10.1146/annurev-cellbio-092910-154055. [DOI] [PubMed] [Google Scholar]
- Pieterse C.M.J, Leon-Reyes A, Van der Ent S, Van Wees S.C.M. Networking by small-molecule hormones in plant immunity. Nat Chem Biol. 2009:308–316. doi: 10.1038/nchembio.164. [DOI] [PubMed] [Google Scholar]
- Prince D.C, Rallapalli G, Xu D, Schoonbeek H.J, Cevik V, Asai S, Kemen E, Cruz-Mireles N, Kemen A, Belhaj K, Schornack S, Kamoun S, Holub E.B, Halkier B.A, Jones J.D. Albugoimposed changes to tryptophan-derived antimicrobial metabolite biosynthesis may contribute to suppression of non-host resistance to Phytophthora infestans in Arabidopsis thaliana. BMC Biol. 2017:20. doi: 10.1186/s12915-017-0360-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiao Y, Shi J, Zhai Y, Hou Y, Ma W. Phytophthora effector targets a novel component of small RNA pathway in plants to promote infection. Proceedings of the National Academy of Sciences of the United States of America. 2015:5850–5855. doi: 10.1073/pnas.1421475112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiao Y, Liu L, Xiong Q, Flores C, Wong J, Shi J, Wang X, Liu X, Xiang Q, Jiang S, Zhang F, Wang Y, Judelson H.S, Chen X, Ma W. Oomycete pathogens encode RNA silencing suppressors. Nature Genetics. 2013:330–333. doi: 10.1038/ng.2525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raaymakers T.M, Van den Ackerveken G. Extracellular recognition of oomycetes during biotrophic infection of plants. Front Plant Sci. 2016:906. doi: 10.3389/fpls.2016.00906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajniak J, Barco B, Clay N.K, Sattely E.S. A new cyanogenic metabolite in Arabidopsis required for inducible pathogen defence. Nature. 2015:376–379. doi: 10.1038/nature14907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rehmany A.P, Grenville L.J, Gunn N.D, Allen R.L, Paniwnyk Z, Byrne J, Whisson S.C, Birch P.R, Beynon J.L. A genetic interval and physical contig spanning the Peronospora parasitica (At) avirulence gene locus ATR1Nd. Fungal Genet Biol. 2003:33–42. doi: 10.1016/s1087-1845(02)00515-7. [DOI] [PubMed] [Google Scholar]
- Rehmany A.P, Gordon A, Rose L.E, Allen R.L, Armstrong M.R, Whisson S.C, Kamoun S, Tyler B.M, Birch P.R, Beynon J.L. Differential recognition of highly divergent downy mildew avirulence gene alleles by RPP1 resistance genes from two Arabidopsis lines. Plant Cell. 2005:1839–1850. doi: 10.1105/tpc.105.031807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rekhter D, Mohnike L, Feussner K, Zienkiewicz K, Zhang Y, Feussner I. Enhanced Disease Susceptibility 5 (EDS5) is required for N-hydroxy pipecolic acid formation. bioRxiv. 2019a:630723. [Google Scholar]
- Rekhter D, Lüdke D, Ding Y, Feussner K, Zienkiewicz K, Lipka V, Wiermer M, Zhang Y, Feussner I. PBS3 is the missing link in plant-specific isochorismate-derived salicylic acid bio-synthesis. bioRxiv. 2019b:600692. doi: 10.1126/science.aaw1720. [DOI] [PubMed] [Google Scholar]
- Ried M.K, Banhara A, Binder A, Hwu F.-H, Gust A.A, Höfle C, Hückelhoven R, Nürnberger T, Parniske M. Symbiosis-related genes sustain the development of a downy mildew pathogen on Arabidopsis thaliana. bioRxiv. 2018:286872. [Google Scholar]
- Robinson L.H, Cahill D.M. Ecotypic variation in the response of Arabidopsis thaliana to Phytophthora cinnamomi. Australas Plant Path. 2003:53–64. [Google Scholar]
- Rodriguez E, El Ghoul H, Mundy J, Petersen M. Making sense of plant autoimmunity and ‘negative regulators’. FEBS J. 2016:1385–1391. doi: 10.1111/febs.13613. [DOI] [PubMed] [Google Scholar]
- Roetschi A, Si-Ammour A, Belbahri L, Mauch F, Mauch-Mani B. Characterization of an Arabidopsis–Phytophthora pathosystem: resistance requires a functional PAD2 gene and is independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant Journal. 2001:293–305. doi: 10.1046/j.1365-313x.2001.01148.x. [DOI] [PubMed] [Google Scholar]
- Rose L.E, Bittner-Eddy P.D, Langley C.H, Holub E.B, Michelmore R.W, Beynon J.L. The maintenance of extreme amino acid diversity at the disease resistance gene, RPP13, in Arabidopsis thaliana. Genetics. 2004:1517–1527. doi: 10.1534/genetics.166.3.1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruhe J, Agler M.T, Placzek A, Kramer K, Finkemeier I, Kemen E.M. Obligate biotroph pathogens of the genus Albugo are better adapted to active host defense compared to niche competitors. Front Plant Sci. 2016:820. doi: 10.3389/fpls.2016.00820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saharan G.S, Verma P.R, Meena P.D, Kumar A. White Rust of Crucifers: Biology, Ecology and Management. Springer India; 2014. [Google Scholar]
- Sapp M, Ploch S, Fiore-Donno A.M, Bonkowski M, Rose L.E. Protists are an integral part of the Arabidopsis thaliana microbiome. Environ Microbiol. 2018:30–43. doi: 10.1111/1462-2920.13941. [DOI] [PubMed] [Google Scholar]
- Sarowar S, Kim Y.J, Kim E.N, Kim K.D, Hwang B.K, Islam R, Shin J.S. Overexpression of a pepper basic pathogenesis-related protein 1 gene in tobacco plants enhances resistance to heavy metal and pathogen stresses. Plant Cell Rep. 2005:216–224. doi: 10.1007/s00299-005-0928-x. [DOI] [PubMed] [Google Scholar]
- Schlaeppi K, Mauch F. Indolic secondary metabolites protect Arabidopsis from the oomycete pathogen Phytophthora brassicae. Plant Signal Behav. 2010:1099–1101. doi: 10.4161/psb.5.9.12410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlaeppi K, Abou-Mansour E, Buchala A, Mauch F. Disease resistance of Arabidopsis to Phytophthora brassicae is established by the sequential action of indole glucosinolates and camalexin. Plant J. 2010:840–851. doi: 10.1111/j.1365-313X.2010.04197.x. [DOI] [PubMed] [Google Scholar]
- Schornack S, Huitema E, Cano L.M, Bozkurt T.O, Oliva R, Van Damme M, Schwizer S, Raffaele S, Chaparro-Garcia A, Farrer R, Segretin M.E, Bos J, Haas B.J, Zody M.C, Nusbaum C, Win J, Thines M, Kamoun S. Ten things to know about oomycete effectors. Mol Plant Pathol. 2009:795–803. doi: 10.1111/j.1364-3703.2009.00593.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiu S.H, Karlowski W.M, Pan R.S, Tzeng Y.H, Mayer K.F.X, Li W.H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell. 2004:1220–1234. doi: 10.1105/tpc.020834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slusarenko A.J, Schlaich N.L. Downy mildew of Arabidopsis thaliana caused by Hyaloperonospora parasitica (formerly Peronospora parasitica) Mol Plant Pathol. 2003:159–170. doi: 10.1046/j.1364-3703.2003.00166.x. [DOI] [PubMed] [Google Scholar]
- Sohn K.H, Lei R, Nemri A, Jones J.D. The downy mildew effector proteins ATR1 and ATR13 promote disease susceptibility in Arabidopsis thaliana. Plant Cell. 2007:4077–4090. doi: 10.1105/tpc.107.054262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonawala U, Dinkeloo K, Danna C.H, McDowell J.M, Pilot G. Review: Functional linkages between amino acid transporters and plant responses to pathogens. Plant Sci. 2018:79–88. doi: 10.1016/j.plantsci.2018.09.009. [DOI] [PubMed] [Google Scholar]
- Soylu E, Soylu S. Light and electron microscopy of the compatible interaction between Arabidopsis and the downy mildew pathogen Peronospora parasitica. Journal of Phytopathology. 2003:300–306. [Google Scholar]
- Soylu E.M, Soylu S, Mansfield J.W. Ultrastructural characterisation of pathogen development and host responses during compatible and incompatible interactions between Arabidopsis thaliana and Peronospora parasitica. Physiological and Molecular Plant Pathology. 2004:67–78. [Google Scholar]
- Soylu S, Keshavarzi M, Brown I, Mansfield J.W. Ultrastructural characterisation of interactions between Arabidopsis thaliana and Albugo candida. Physiological and molecular plant pathology. 2003:201–211. [Google Scholar]
- Spencer-Phillips P. Function of fungal haustoria in epiphytic and endophytic infections. Advances in Botanical Research. 1997:309–333. [Google Scholar]
- Stam R, Jupe J, Howden A.J, Morris J.A, Boevink P.C, Hedley P.E, Huitema E. Identification and characterisation of CRN Effectors in Phytophthora capsici shows modularity and functional diversity. PloS One. 2013 doi: 10.1371/journal.pone.0059517. e59517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stassen J.H, Van den Ackerveken G. How do oomycete effectors interfere with plant life? Current Opinion In Plant Biology. 2011:407–414. doi: 10.1016/j.pbi.2011.05.002. [DOI] [PubMed] [Google Scholar]
- Staswick P.E, Yuen G.Y, Lehman C.C. Jasmonate signaling mutants of Arabidopsis are susceptible to the soil fungus Pythium irregulare. The Plant Journal. 1998:747–754. doi: 10.1046/j.1365-313x.1998.00265.x. [DOI] [PubMed] [Google Scholar]
- Stein M, Dittgen J, Sanchez-Rodriguez C, Hou B.H, Molina A, Schulze-Lefert P, Lipka V, Somerville S. Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to non-host resistance to inappropriate pathogens that enter by direct penetration. Plant Cell. 2006:731–746. doi: 10.1105/tpc.105.038372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinbrenner A.D, Goritschnig S, Staskawicz B.J. Recognition and activation domains contribute to allele-specific responses of an Arabidopsis NLR receptor to an oomycete effector protein. PLoS Pathogens. 2015 doi: 10.1371/journal.ppat.1004665. e1004665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stuttmann J, Hubberten H.-M, Rietz S, Kaur J, Muskett P, Guerois R, Bednarek P, Hoefgen R, Parker J.E. Perturbation of Arabidopsis amino acid metabolism causes incompatibility with the adapted biotrophic pathogen Hyaloperonospora arabidopsidis. Plant Cell. 2011:2788–2803. doi: 10.1105/tpc.111.087684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabo L.J, Bushnell W.R. Hidden robbers: the role of fungal haustoria in parasitism of plants. Proceedings of the National Academy of Sciences of the United States of America. 2001:7654–7655. doi: 10.1073/pnas.151262398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takemoto D, Jones D.A, Hardham A.R. GFP-tagging of cell components reveals the dynamics of subcellular re-organization in response to infection of Arabidopsis by oomycete pathogens. The Plant Journal. 2003:775–792. doi: 10.1046/j.1365-313x.2003.01673.x. [DOI] [PubMed] [Google Scholar]
- Thines M. Phylogeny and evolution of plant pathogenic oomycetes‚ a global overview. Eur J Plant Pathol. 2014:1–17. [Google Scholar]
- Thines M, Choi Y.J. Evolution, diversity, and taxonomy of the Peronosporaceae, with focus on the genus Peronospora. Phytopathology. 2016:6–18. doi: 10.1094/PHYTO-05-15-0127-RVW. [DOI] [PubMed] [Google Scholar]
- Thines M, Choi Y.J, Kemen E, Ploch S, Holub E.B, Shin H.D, Jones J.D. A new species of Albugo parasitic to Arabidopsis thaliana reveals new evolutionary patterns in white blister rusts (Albuginaceae) Persoonia. 2009:123–128. doi: 10.3767/003158509X457931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomma B.P.H.J, Nurnberger T, Joosten M.H.A.J. Of PAMPs and effectors: The blurred PTI-ETI dichotomy. Plant Cell. 2011:4–15. doi: 10.1105/tpc.110.082602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomczynska I, Stumpe M, Mauch F. A conserved RxLR effector interacts with host RABA-type GTPases to inhibit vesicle-mediated secretion of antimicrobial proteins. Plant J. 2018:187–203. doi: 10.1111/tpj.13928. [DOI] [PubMed] [Google Scholar]
- Torrens-Spence M.P, Bobokalonova A, Carballo V, Glinkerman C.M, Pluskal T, Shen A, Weng J.-K. PBS3 and EPS1 complete salicylic acid biosynthesis from isochorismate in Arabidopsis. bioRxiv. 2019:601948. doi: 10.1016/j.molp.2019.11.005. [DOI] [PubMed] [Google Scholar]
- Toruno T.Y, Stergiopoulos I, Coaker G. Plant-pathogen effectors: cellular probes interfering with plant defenses in spatial and temporal manners. Annu Rev Phytopathol. 2016:419–441. doi: 10.1146/annurev-phyto-080615-100204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuchiya T, Eulgem T. An alternative polyadenylation mechanism coopted to the Arabidopsis RPP7 gene through intronic retrotransposon domestication. Proceedings of the National Academy of Sciences of the United States of America. 2013:E3535–3543. doi: 10.1073/pnas.1312545110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyler B.M, Tripathy S, Zhang X, Dehal P, Jiang R.H, Aerts A, Arredondo F.D, Baxter L, Bensasson D, Beynon J.L, Chapman J, Damasceno C.M, Dorrance A.E, Dou D, Dickerman A.W, Dubchak I.L, Garbelotto M, Gijzen M, Gordon S.G, Govers F, Grunwald N.J, Huang W, Ivors K.L, Jones R.W, Kamoun S, Krampis K, Lamour K.H, Lee M.K, McDonald W.H, Medina M, Meijer H.J, Nordberg E.K, Maclean D.J, Ospina-Giraldo M.D, Morris P.F, Phuntumart V, Putnam N.H, Rash S, Rose J.K, Sakihama Y, Salamov A.A, Savidor A, Scheuring C.F, Smith B.M, Sobral B.W, Terry A, Torto-Alalibo T.A, Win J, Xu Z, Zhang H, Grigoriev I.V, Rokhsar D.S, Boore J.L. Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science. 2006:1261–1266. doi: 10.1126/science.1128796. [DOI] [PubMed] [Google Scholar]
- Uknes S, Mauch-Mani B, Moyer M, Potter S, Williams S, Dincher S, Chandler D, Slusarenko A, Ward E, Ryals J. Acquired resistance in Arabidopsis. Plant Cell. 1992:645–656. doi: 10.1105/tpc.4.6.645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Damme M, Zeilmaker T, Elberse J, Andel A, de Sain-van der Velden M, van den Ackerveken G. Downy Mildew Resistance in Arabidopsis by Mutation of HOMOSERINE KINASE. Plant Cell. 2009:2179. doi: 10.1105/tpc.109.066811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Biezen E.A, Freddie C.T, Kahn K, Parker J.E, Jones J.D. Arabidopsis RPP4 is a member of the RPP5 multigene family of TIR-NB-LRR genes and confers downy mildew resistance through multiple signalling components. Plant J. 2002:439–451. doi: 10.1046/j.0960-7412.2001.01229.x. [DOI] [PubMed] [Google Scholar]
- van Loon L.C, Rep M, Pieterse C.M. Significance of inducible defense-related proteins in infected plants. Annu Rev Phytopathol. 2006:135–162. doi: 10.1146/annurev.phyto.44.070505.143425. [DOI] [PubMed] [Google Scholar]
- Vlot A.C, Dempsey D.A, Klessig D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu Rev Phytopathol. 2009:177–206. doi: 10.1146/annurev.phyto.050908.135202. [DOI] [PubMed] [Google Scholar]
- Voglmayr H, Riethmuller A, Goker M, Weiss M, Oberwinkler F. Phylogenetic relationships of Plasmopara, Bremia and other genera of downy mildew pathogens with pyriform haustoria based on Bayesian analysis of partial LSU rDNA sequence data. Mycol Res. 2004:1011–1024. doi: 10.1017/s0953756204000954. [DOI] [PubMed] [Google Scholar]
- Wang S, Boevink P.C, Welsh L, Zhang R, Whisson S.C, Birch P.R.J. Delivery of cytoplasmic and apoplastic effectors from Phytophthora infestans haustoria by distinct secretion pathways. New Phytologist. 2017:205–215. doi: 10.1111/nph.14696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Bouwmeester K, van de Mortel J.E, Shan W, Govers F. Induced expression of defense-related genes in Arabidopsis upon infection with Phytophthora capsici. Plant Signal Behav. 2013 doi: 10.4161/psb.24618. e24618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Meng Y, Zhang M, Tong X, Wang Q, Sun Y, Quan J, Govers F, Shan W. Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host specificity. Mol Plant Pathol. 2011:187–201. doi: 10.1111/j.1364-3703.2010.00659.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Xu Y, Sun Y, Wang H, Qi J, Wan B, Ye W, Lin Y, Shao Y, Dong S, Tyler B.M, Wang Y. Leucine-rich repeat receptor-like gene screen reveals that Nicotiana RXEG1 regulates glycoside hydrolase 12 MAMP detection. Nat Commun. 2018a:594. doi: 10.1038/s41467-018-03010-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z, Tyler B.M, Liu X. Protocol of Phytophthora capsici transformation using the CRISPR-Cas9 system. Methods Mol Biol. 2018b:265–274. doi: 10.1007/978-1-4939-8724-5_17. [DOI] [PubMed] [Google Scholar]
- Wawra S, Belmonte R, Lobach L, Saraiva M, Willems A, van West P. Secretion, delivery and function of oomycete effector proteins. Current Opinion In Microbiology. 2012:685–691. doi: 10.1016/j.mib.2012.10.008. [DOI] [PubMed] [Google Scholar]
- Wessling R, Epple P, Altmann S, He Y, Yang L, Henz S.R, Mc-Donald N, Wiley K, Bader K.C, Glasser C, Mukhtar M.S, Haigis S, Ghamsari L, Stephens A.E, Ecker J.R, Vidal M, Jones J.D, Mayer K.F, Ver Loren van Themaat E, Weigel D, Schulze-Lefert P, Dangl J.L, Panstruga R, Braun P. Convergent targeting of a common host protein-network by pathogen effectors from three kingdoms of life. Cell Host & Microbe. 2014:364–375. doi: 10.1016/j.chom.2014.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiermer M, Feys B.J, Parker J.E. Plant immunity: the EDS1 regulatory node. Current opinion in plant biology. 2005:383–389. doi: 10.1016/j.pbi.2005.05.010. [DOI] [PubMed] [Google Scholar]
- Wildermuth M.C, Dewdney J, Wu G, Ausubel F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature. 2001:562–565. doi: 10.1038/35107108. [DOI] [PubMed] [Google Scholar]
- Willmann R, Lajunen H.M, Erbs G, Newman M.A, Kolb D, Tsuda K, Katagiri F, Fliegmann J, Bono J.J, Cullimore J.V, Jehle A.K, Gotz F, Kulik A, Molinaro A, Lipka V, Gust A.A, Nurnberger T. Arabidopsis lysin-motif proteins LYM1 LYM3 CERK1 mediate bacterial peptidoglycan sensing and immunity to bacterial infection. Proceedings of the National Academy of Sciences of the United States of America. 2011:19824–19829. doi: 10.1073/pnas.1112862108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Win J, Chaparro-Garcia A, Belhaj K, Saunders D.G, Yoshida K, Dong S, Schornack S, Zipfel C, Robatzek S, Hogenhout S.A, Kamoun S. Effector biology of plant-associated organisms: concepts and perspectives. Cold Spring Harbor symposia on quantitative biology. 2012:235–247. doi: 10.1101/sqb.2012.77.015933. [DOI] [PubMed] [Google Scholar]
- Woloshuk C.P, Meulenhoff J.S, Selabuurlage M, Vandenelzen P.J.M, Cornelissen B.J.C. Pathogen-induced proteins with inhibitory activity toward Phytophthora infestans. Plant Cell. 1991:619–628. doi: 10.1105/tpc.3.6.619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods-Tor A, Studholme D.J, Cevik V, Telli O, Holub E.B, Tor M. A suppressor/avirulence gene combination in Hyaloperonospora arabidopsidis determines race specificity in Arabidopsis thaliana. Front Plant Sci. 2018:265. doi: 10.3389/fpls.2018.00265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu C.-H, Derevnina L, Kamoun S. Receptor networks underpin plant immunity. Science. 2018:1300. doi: 10.1126/science.aat2623. [DOI] [PubMed] [Google Scholar]
- Wu Z, Li M, Dong O.X, Xia S, Liang W, Bao Y, Wasteneys G, Li X. Differential regulation of TNL-mediated immune signaling by redundant helper CNLs. New Phytologist. 2019:938–953. doi: 10.1111/nph.15665. [DOI] [PubMed] [Google Scholar]
- Xiong Q, Ye W, Choi D, Wong J, Qiao Y, Tao K, Wang Y, Ma W. Phytophthora Suppressor of RNA Silencing 2 is a conserved RxLR effector that promotes infection in soybean and Arabidopsis thaliana. Mol Plant Microbe Interact. 2014:1379–1389. doi: 10.1094/MPMI-06-14-0190-R. [DOI] [PubMed] [Google Scholar]
- Yan S.P, Dong X.N. Perception of the plant immune signal salicylic acid. Current Opinion In Plant Biology. 2014:64–68. doi: 10.1016/j.pbi.2014.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeilmaker T, Ludwig N.R, Elberse J, Seidl M.F, Berke L, Van Doorn A, Schuurink R.C, Snel B, Van den Ackerveken G. DOWNY MILDEW RESISTANT 6 and DMR6-LIKE OXYGEN-ASE 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2015:210–222. doi: 10.1111/tpj.12719. [DOI] [PubMed] [Google Scholar]
- Zhang L, Zhang F, Melotto M, Yao J, He S.Y. Jasmonate signaling and manipulation by pathogens and insects. J Exp Bot. 2017:1371–1385. doi: 10.1093/jxb/erw478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W, Fraiture M, Kolb D, Loffelhardt B, Desaki Y, Boutrot F.F, Tor M, Zipfel C, Gust A.A, Brunner F. Arabidopsis receptor-like protein30 and receptor-like kinase suppressor of BIR1-1/EVERSHED mediate innate immunity to necrotrophic fungi. Plant Cell. 2013:4227–4241. doi: 10.1105/tpc.113.117010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z, Wu Y, Gao M, Zhang J, Kong Q, Liu Y, Ba H, Zhou J, Zhang Y. Disruption of PAMP-induced MAP kinase cascade by a Pseudomonas syringae effector activates plant immunity mediated by the NB-LRR protein SUMM2. Cell Host & Microbe. 2012:253–263. doi: 10.1016/j.chom.2012.01.015. [DOI] [PubMed] [Google Scholar]
- Zheng X, McLellan H, Fraiture M, Liu X, Boevink P.C, Gilroy E.M, Chen Y, Kandel K, Sessa G, Birch P.R, Brunner F. Functionally redundant RXLR effectors from Phytophthora infestans act at different steps to suppress early flg22-triggered immunity. PLoS Pathogens. 2014 doi: 10.1371/journal.ppat.1004057. e1004057. [DOI] [PMC free article] [PubMed] [Google Scholar]