

Abstract
Bacteria inhabit a variety of locations and play important roles in the environment and health. Our understanding of bacterial biomechanics has improved markedly in the last decade and has revealed that biomechanics play a significant role in microbial biology. The obtained knowledge has enabled investigation of complex phenomena, such as biofilm formation and the dynamics of the gut flora. A bottom-up strategy, i.e., from the cellular to the macroscale, facilitates understanding of macroscopic bacterial phenomena. In this Review, we first cover the biomechanics of individual bacteria in the bulk liquid and on surfaces as the base of complex phenomena. The collective behaviors of bacteria in simple environments are next introduced. We then introduce recent advances in biofilm biomechanics, in which adhesion force and the flow environment play crucial roles. We also review transport phenomena in the intestine and the dynamics of the gut flora, focusing on that in zebrafish. Finally, we provide an overview of the future prospects for the field.
I. INTRODUCTION
Bacteria appeared on Earth more than 3 billion years ago. They inhabit a wide variety of locations and account for about 13% of the global biomass (Bar-On et al., 2018). Ecologically, bacteria are at the bottom of the food chain and many protists rely on them. Bacteria also play crucial roles in the nitrogen and carbon cycles. In engineering, they are used for water purification in sewage treatment and for extracting metals from ores (biomining). Bacterial biofilms may cause deterioration in the performance of machinery, causing biofouling. The human gut harbors about 1 kg of bacteria, the number of which is about twofold that of human cells. The gut flora plays important roles in health and disease. Therefore, the study of bacteria is an important subject of scientific research.
In the last decade, our understanding of bacterial biomechanics, such as adhesion on surfaces, collective swimming, clustering, and transport phenomena, has improved markedly. Former research revealed that biomechanics play a significant role in microbial biology. The knowledge obtained has been used to investigate more complex phenomena, such as biofilm formation and the dynamics of the gut flora. To understand such macroscopic phenomena, an understanding of the physical properties of the environment, such as nutrient concentration, diffusivity, and viscoelasticity, is needed. These macroscopic properties are governed by mesoscale microbial structures, which comprise intercellular interactions. Therefore, a bottom-up understanding of bacterial phenomena, i.e., from the cellular to the macroscale, is needed (Fig. 1).
FIG. 1.
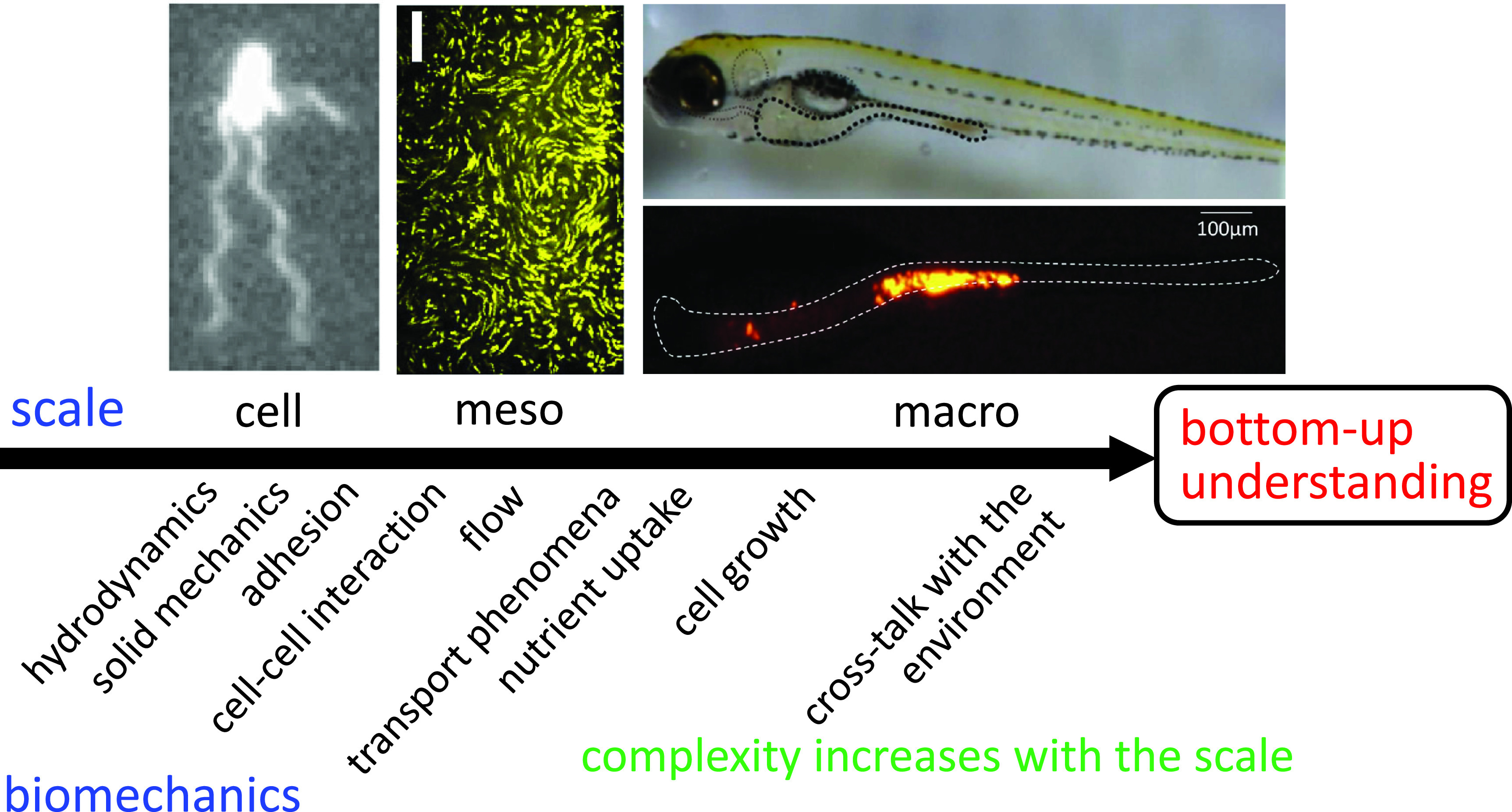
Schematic of a bottom-up understanding of bacterial biomechanics from the cellular to the macroscale. Top left, Escherichia coli cell; top middle, trajectories of tracers in a dense E. coli suspension; top right, zebrafish larva; and bottom right, fluorescent tracer particles in the intestine of zebrafish larva. The complexity of biomechanics increases with scale.
In this Review, we focus on biomechanical aspects of bacterial phenomena, though microbiology has traditionally focused on the influence of chemical environments. Hydrodynamics and solid mechanics are used to discuss bacterial phenomena in all scales. Adhesion force is introduced in discussing the cell-surface interaction. Cell–cell interactions are important at the mesoscale because they determine the collective behaviors of cells. The transport of momentum, cells, and nutrients needs to be clarified at the mesoscale to extract macroscopic properties. At the macroscale, nutrient uptake, which governs the growth rate, must be discussed. Moreover, at the macroscale, cells' activities alter their environment and, in turn, their behaviors change. Thus, crosstalk between cells and their environment has to be discussed. Hence, the complexity in biomechanics necessarily increases as the scale increases (Fig. 1).
In Sec. II, we explain the behaviors of individual bacteria in the bulk liquid and on surfaces as the base of complex phenomena. The collective behaviors of bacteria in simplified environments are introduced in Sec. III. In Sec. IV, we discuss the biomechanics of biofilm formation, in which adhesion forces and the flow environment play crucial roles. In Sec. V, we discuss the gut flora in terms of biomechanics, focusing on that in zebrafish. In each section, we suggest other reviews as further reading. In Sec. VI, we discuss future prospects for this field.
II. BIOMECHANICS OF AN INDIVIDUAL BACTERIUM
The individual behavior of a bacterium is the basis of their collective behaviors. We thus discuss the biomechanics of an individual bacterium in this section. In Sec. II A, we focus on a bacterium swimming in the bulk liquid. The physical environment of the intestine can be complex both rheologically and geometrically; therefore, the effects of solvent rheology and channel geometry on the swimming behaviors are discussed in Sec. II B. In Sec. II C, we focus on a bacterium adhered on a surface, because it is the beginning of biofilm formation. We suggest the following reviews as supplementary reading: Lauga (2016) reviewed bacterial hydrodynamics, Berg (2004) overviewed biological aspects of bacterial swimming, including chemotaxis, and Persat et al. (2015) explained the mechanical environment of surface-associated bacteria.
A. Bacteria swimming in bulk liquid
Bacterial flagella, such as those of Escherichia coli (E. coli), are rotated by tiny molecular motors, and the estimated torque of the motors is 2000–5000 pN nm (Chen and Berg, 2000; Sowa et al., 2003; Darnton et al., 2007; Inoue et al., 2008; Shimogonya, 2015). The torque generated by the motor is transmitted to a helical flagellar filament via a hook (Berg, 2003), and the rotation of helical flagella generates the propulsive force. As a result, E. coli (body length ∼2 μm) swims at a velocity of about 20 μm/s. Swimming motion can be broadly characterized by the Reynolds number, which indicates the ratio of inertia force to the viscous force. The Reynolds number of E. coli is on the order of 10−5, and so they live in the viscous dominant world.
Bacteria often have multiple flagella. The multiple flagella of E. coli form a single bundle when all flagella rotate in a counterclockwise direction, resulting in forward motion (Berg, 2004). When some flagella change the direction of rotation, the bundle unravels and the flagella separate. The biomechanics of flagellar bundling has been investigated (Kim et al., 2003; Watari and Larson, 2010; Reigh et al., 2012; Kanehl and Ishikawa, 2014). E. coli can adapt the frequency of alternation between tumbling and running, facilitating chemotaxis (Berg, 2004; Codling et al., 2008; Goto and Nakai, 2016).
B. Behaviors in complex environments
Bacteria can be attracted by a solid surface. Berke et al. (2008) showed that swimming bacteria tend to be attracted to surfaces, because the steady-state density increased 10-fold near surfaces. The attraction mechanism could be explained by the far-field hydrodynamic effect, which requires awareness of the difference between a pusher and a puller. The puller has a flagellum in front of the cell body, while the pusher has a flagellum behind the cell body. Figure 2 shows the flow fields far from the cell body; these are opposite between pullers and pushers and are not affected by cell shape. A pusher-type bacterium such as E. coli tends to orient parallel to and be attracted to surfaces due to the far-field hydrodynamic effect (Berke et al., 2008; Lauga and Powers, 2009).
FIG. 2.

Schematics of a puller and a pusher. The puller has a flagellum in front of the cell body, while the pusher has a flagellum behind the cell body.
A pusher-type bacterium can be entrapped by a surface boundary due to the near-field alignment effect. In the vicinity of a surface boundary, E. coli draws a circular trajectory (Lauga et al., 2006; Frymier et al., 1995; Vigeant and Ford, 1997; Lauga et al., 2006). Similar trajectories were also reported for the singly flagellated bacterium Vibrio alginolyticus (Goto et al., 2005). The minimum separation distance between the cell and the surface has been estimated to be ten to several hundred nanometers (Frymier et al., 1995; Vigeant and Ford, 1997; Vigeant et al., 2002). The mechanism of entrapment by a surface has been explained by hydrodynamics and steric effects (Ramia et al., 1993; Lauga et al., 2006; Giacché et al., 2010, Goto et al., 2010; Giacché et al., 2010). Notably, the circular trajectory found near a solid surface switches the direction near a free surface (Lemelle, 2010). When background shear flow is imposed, E. coli swimming near a wall drift perpendicular to the flow direction (Ishikawa et al., 2014).
Some bacteria live in heterogeneous granular matter, such as soil, and the habitat is influenced by physical parameters including the granular size and shape (Young and Crawford, 2004; Or et al., 2007). Brown et al. (2016) studied E. coli swimming in a monolayer of spheres on a surface. The circular orbits of E. coli on a flat wall were modified to long and straight runs, because the bacteria are unable to turn corners inside the crystal. Swimming in granular matter has also been analyzed using a simple microswimmer model (Volpe et al., 2011; Chamolly et al., 2017), in which straight swimming, random walking, entrapment in a closed orbit, or entrapment of cells were observed. These results illustrate the significant influence of geometric constraints on bacterial behaviors.
The effect of viscosity on bacterial swimming velocity has been investigated. The swimming velocity increases up to about twofold the viscosity of water but decreases monotonically with further increases in viscosity (Schneider and Doetsch, 1974; Greenberg and Canale-Parola, 1977). The decreasing tendency is caused by the constant torque of the molecular motor in the low-speed regime, and the additional viscous drag decreases the rotational velocity. The velocity increase may be related to the non-Newtonian property of fluid induced by the structure of polymer chains (Lauga, 2016).
C. Bacteria adhered on surfaces
A single bacterium adhered on a wall can grow and divide and then become sessile and eventually construct multicellular structures, i.e., biofilms (Hall-Stoodley et al., 2004). Thus, it is important to understand the biomechanics of bacterial adhesion (Persat et al. 2015).
Interactions between charged surfaces via a liquid medium can be understood based on the Derjaguin–Landau–Verwey–Overbeek (DLVO) theory of colloid stability (Verwey and Overbeek, 1948). Bacteria have acquired various adhesion strategies to overcome repulsive interactions. Many bacteria have pili, which are hair-like appendages distributed on the cell surface. Pili enhance bacterial adhesion by increasing the probability that a cell makes physical contact with a surface. Bacteria also secrete extracellular polymeric substances (EPSs). The EPS is composed of polysaccharides and other macromolecules and increases the affinity of bacteria to surfaces (Flemming and Wingender, 2010). Microscale surface patterns, such as submicrometer crevices (Friedlander et al., 2013), nanoporous surfaces (Feng et al., 2015), and bump lines (Yang et al., 2019), have the potential to reduce the adhesion of bacteria.
The background flow also influences bacterial adhesion to surfaces. When the wall shear stress is sufficiently strong, the viscous drag acting on the cell body overwhelms the adhesion force and the cell is washed away. Some studies have reported that bacteria can establish a catch bond, in which the dissociation lifetime increases with the tensile force applied to the bond, i.e., the adhesion force increases with the background flow (Thomas, 2008). Nilsson et al. (2006) found that type I fimbriae attachment of E. coli has a shear threshold for binding, below which bacteria do not adhere. This tendency benefits not only biofilm formation but also the gut flora, because bacteria can resist the peristaltic motion of the intestine.
III. COLLECTIVE BEHAVIORS OF BACTERIA
In this section, we explain the collective behaviors of bacteria in simplified settings, to promote understanding of bacterial behaviors in biofilms and the gut. In Sec. III A, collective swimming and coherent structures of bacteria in dense suspensions are introduced. Transport phenomena, such as mass transport and viscosity, are important for macroscopic cell behaviors and growth and are discussed in Sec. III B. In Sec. III C, we discuss colony pattern formation on agar gels, which is important for understanding biofilms. We suggest the following as supplementary reading: Elgeti et al. (2015) reviewed collective behaviors of microswimmers from the physical perspective and Jacob et al. (2004) focused on self-organization of bacterial colonies. In this Review, bioconvection of a bacterial suspension is not discussed. Bioconvection is the biological analogue to a thermal convection and results in geometric patterns. See Hill and Pedley (2005) and Pedley and Kessler (1992).
A. Collective swimming
In dense bacterial suspensions, such as 1010 cell/ml, bacteria tend to swim in a similar direction to their neighbors and generate mesoscale coherent structures (Dombrowski et al., 2004; Ishikawa et al., 2011; Dunkel et al., 2013). The mechanism of this collective motion has been investigated using continuum models, which do not consider near-field cell–cell interactions. By considering the alignment of cells in the stretching direction of background flow, the continuum models reproduced turbulent bacterial spatiotemporal motion (Ramaswamy, 2010; Koch and Subramanian, 2011; Marchetti et al., 2013; Wensink et al., 2012). Therefore, coherent structures can be reproduced by far-field fluid interactions, and near-field cell–cell interactions may not be important.
Discrete models have been employed to investigate the collective motion of individual cells. The hydrodynamic interactions among multiple cells in near- and far-fields have been elucidated (Ishikawa and Pedley, 2008; Ishikawa et al., 2008; Evans et al., 2011; Lambert et al., 2013; Potomkin et al., 2013; Zottl and Stark, 2014). The discrete models also successfully reproduced the turbulent bacterial spatiotemporal motion; however, questions remain regarding the role of near-field fluid mechanics in collective motion.
Kyoya et al. (2015) solved the Stokes flow around hydrodynamically interacting prolate squirmers in a monolayer suspension. Weak pushers, such as E. coli, generated coherent structures [Fig. 3(a)]. To separate the effects of near- and far-field hydrodynamic interactions, they performed physically inconsistent but sophisticated trial simulations. The flow field of Stokes flow can be expressed by a boundary integral equation, in which the integral of traction forces over all squirmer surfaces appears. Instead of performing it rigorously, they performed the integral for cells only in the near field, although it was not physically consistent. The flow generated by far-field cells disappeared, and the resultant structure was generated solely by near-field cell–cell interactions. Kyoya et al. showed that the coherent structures found in bacterial baths are generated mainly by near-field cell–cell interactions.
FIG. 3.

Collective motion of bacteria in a dense suspension. (a) Numerical simulation of collective swimming of prolate squirmers in a monolayer suspension. Yellow arrows, velocity vectors; and gray lines, orientations. The areal fraction of cells is 0.3, and the cells are pushers (β = −0.3). (Problem settings are identical to Kyoya et al., 2015.). (b) Trajectories of tracer particles in a dense E. coli suspension. Scale bar, 50 μm. (Settings are identical to Ishikawa et al., 2011.)
B. Transport phenomena in bacterial suspensions
We discuss here the transport of momentum, cells, and substances in a bacterial suspension (Ishikawa, 2009). The transport of momentum is closely related to the rheological properties of the suspension and so affects the flow field. The transport of cells governs their distribution in the suspension. Transport of substances, such as oxygen and nutrients, determines the growth rate of cells. Thus, transport phenomena are important for understanding short- and long-term bacterial behaviors.
The bulk stress tensor in a Newtonian fluid can be given by Newton's law of viscosity. When small particles are suspended in a Newtonian fluid, however, an additional contribution to the bulk stress tensor is generated. The additional stress is termed a particle stress tensor and can be calculated from the traction forces and velocities on the particle surface (Batchelor, 1970). The particle stress tensor governs the rheological properties of the suspension, such as its viscosity and normal stress differences. Ishikawa and Pedley (2007) investigated the shear viscosity in a semi-dilute suspension of active cells. They showed that, when the active cells are aligned in a certain direction, the shear viscosity is altered from that of non-motile cells. For example, when a pusher is placed relative to the shear flow as shown in Fig. 4(a), it helps the wall movement and the shear viscosity is decreased. The change in the shear viscosity is opposite between the pullers and pushers (cf. Fig. 2). Also, when the cells are aligned, normal stress differences appeared in the suspension, as shown in Fig. 4(b). If forming or destroying the cells' alignment requires some time, the stress tensor relaxes with time (Ishikawa et al., 2007). Thus, bacterial suspensions have non-Newtonian rheological properties.
FIG. 4.

Schematics of the stress field generated by pushers. (a) A pusher with the given orientation helps the wall movement, and thus the shear viscosity is decreased. (b) Normal stress appears when orientation of pushers is anisotropic.
The shear viscosity in a bacterial suspension can be decreased below the viscosity of solvent fluid (Hatwalne et al., 2004; Haines et al., 2009; Ryan et al., 2011). Giomi et al. (2010) predicted a superfluid phase with vanishing viscosity for strongly active swimmers. López et al. (2015) showed, using E. coli, that a bacterial suspension displays a superfluid-like transition at which viscous resistance to shear vanishes. Therefore, the activity of pusher bacteria organized by shear can overcome viscous dissipation.
Macroscopic transport of cells can be modeled by a continuum model that incorporates advection, diffusion, and growth effects (Pedley and Kessler, 1992). The advection effect can be expressed by the drift velocity, which is a function of background flow, surface boundaries, and chemotaxis (Berg, 2004; Ishikawa, 2012; Lauga, 2016). The diffusion effect may be expressed using a diffusion tensor, which can be caused by the intrinsic randomness of biological processes and cell–cell hydrodynamic interactions (Cates, 2012). Ishikawa and Pedley (2007) investigated the hydrodynamic effect in a semi-dilute regime. The translational diffusivity was inversely proportional to the cell-number density, while the rotational diffusivity was proportional to the number density.
In a dense bacterial suspension, transport of tracer particles is significantly increased. Wu and Libchaber (2000) showed, using E. coli, that the diffusivity of 10-μm-diameter tracer particles is three orders of magnitude larger than Brownian diffusivity. This marked increase in diffusivity is caused by the turbulent-like flow structure [Fig. 3(b)]. A similar increase in tracer diffusivity has been reported by others (Sokolov et al., 2009; Ishikawa et al., 2011; Jepson et al., 2013).
C. Colony pattern formation
The study of bacterial colonies is important for understanding biofilms (Jacob et al., 2004). Bacteria show adaptable collective responses and form colonies with geometric patterns. Colony dynamics comprise the trajectories of aggregates that grow, move, and reproduce simultaneously. Wakita et al. (1997) classified the colony patterns of Bacillus subtilis into five types: (i) diffusion-limited aggregation-like, (ii) Eden-like, (iii) concentric ring-like, (iv) disk-like, and (v) dense branching morphology-like. The morphology was dependent on the substrate softness and nutrient concentration. Therefore, transport phenomena in bacterial colonies strongly affect their morphology.
Bacteria in colonies can migrate collectively on surfaces and generate large swirls and jets. Such swarming involves several cellular processes, such as changes in the levels of key proteins, intercellular chemical communication, and mechanical factors (Beer and Ariel, 2019). Therefore, swarming can be distinguished from collective swimming (Sec. III A), in which cellular processes do not play a major role. Ariel et al. (2015) investigated the motion of bacteria within a swarm and found that bacterial spreading was not diffusive but superdiffusive.
To describe colony dynamics, several mathematical models have been proposed. Mimura et al. (2000) proposed a model by combining a reaction diffusion system of nutrient dynamics, the nucleation theory of aggregate generation, and individual-based dynamics of the motion and growth of aggregates. The model consistently reproduced the experimentally observed branching patterns and effects of the initial nutrient concentration. The model was enhanced to take into account active and inactive bacterial cells (Matsushita et al., 2004). Gerlee and Anderson (2007) proposed a hybrid cellular automaton model of colony growth, in which colony growth is limited by the concentration of a nutrient, which inhibits cell division if it falls below a threshold. Dong and Klumpp (2018) used a cellular automaton model to investigate colony pattern formation under the differential adhesion hypothesis and in the presence of cell proliferation.
Farrell et al. (2013) assumed that bacteria interact purely mechanically, by pushing each other away as they grow and consume a diffusing nutrient. They showed that mechanical interactions can explain the transition between circular and branching colonies. The importance of mechanics is also emphasized by the study of Ghosh et al. (2015), in which repulsive forces generated phase-separated patterns in the growing colony. Although these mathematical models clarified aspects of colony growth, our understanding remains limited. For example, the effect of swarming on transport phenomena in the colony is unclear. Future studies are expected to provide a more quantitative description of colony growth.
IV. BIOFILM FORMATION
Biofilms are surface-associated microbial communities encased in a self-secreted matrix of extracellular polymeric substances (EPSs) (Jang et al., 2017). Bacteria typically live in biofilms as their natural habitat (Drescher et al., 2013), and biofilms account for a large proportion of bacterial biomass. For example, biofilms are the sources of infections associated with catheters and implanted devices (Brooks and Flint, 2008). In industry, biofouling and biocorrosion by biofilms are responsible for pipe-clogging and product contamination (Campoccia et al., 2013; Jang et al., 2017). Biofilms are different from planktonic bacteria, exhibiting markedly greater resistance to antibiotics and external stressors (Nguyen et al., 2011).
Both the chemical and biomechanical environments play significant roles in biofilm formation, as explained in Sec. IV A. The surrounding hydrodynamics can generate flow-shaped biofilms, called streamers (Rusconi et al., 2010) (Sec. IV B). The deformation of biofilms depends on their physical properties, which are reviewed in Sec. IV C. To understand the physical mechanism of biofilm formation several mathematical models have been developed. We review them in Sec. IV D. We suggest the following as supplementary reading: Dufrêne and Persat (2020) reviewed bacterial mechanobiology, Persat et al. (2015) focused on mechanical aspects of biofilm formation, and Even et al. (2017) reviewed the mechanical properties of biofilms.
A. Biofilm formation
Biofilm formation is influenced by many factors, such as species, genotype, pH, nutrient, water content, and the presence of other microbial species (Albareda et al., 2006). Bacteria within biofilms can communicate to control their growth rate, i.e., quorum sensing. Shrout et al. (2006) reported that quorum sensing in P. aeruginosa exerted nutritionally conditional control of biofilm development by regulating swarming motility. Also, distant bacterial biofilms can coordinate their growth dynamics to resolve competition for limited resources (Liu et al., 2017). Biofilm structures can thus result from the interaction between the environment and individual cells.
Hydrodynamics also influence biofilm formation, because they govern drag force and mass transport (Liu and Tay, 2002). Strong shear stress may lead to breakage of a biofilm, which induces dispersion of the biofilm to new territories. Stoodley et al. (1999) observed biofilm growth of gram-negative species under turbulent and laminar flows and found patchy biofilms in laminar flow and patches of ripples and elongated streamers in turbulent flow. Simoes et al. (2007) investigated biofilm formation by Pseudomonas under turbulent and laminar flows and found that turbulent flow-generated biofilms were metabolically more active and had higher cell densities than laminar flow-generated biofilms. Vieira et al. (1993) measured the mass transfer coefficients in Pseudomonas biofilms at different fluid velocities; the mass transfer rates decreased more quickly as the fluid velocities increased, and lower internal diffusivities resulted in lower final thicknesses of the biofilms.
Pathogens also take advantage of biofilm formation. For example, plant-associated bacteria can sense plant exudates released from specific sites and aggregate via chemotaxis. Yao and Allen (2006) showed that directed motility, not simply random motion, is required for full virulence and that chemotaxis is an important trait for virulence and pathogenic fitness in plant pathogens. Fujishige et al. (2006) used flagella-less Sinorhizobium mutants and found that nodule formation was significantly delayed without cell motility. These results show that cell motility plays important roles in the initiation, maturation, and spread of biofilms and colonization of new habitats. Moreover, the surrounding flow field influences biofilm formation by pathogens. Kostenko et al. (2010) investigated the biofilm morphology and tolerance of Staphylococcus to antibiotics under oscillating shear stress as a model of bacterial infections in the blood system. They found that pulsatile flow promotes the formation of more tolerant biofilms and engenders difficult-to-treat infection sites.
B. Streamers—biofilm formation under flow
In the last decade, the link between hydrodynamics and complex biofilm structures has been investigated experimentally and theoretically. A remarkable example is filamentous flow-shaped biofilms, i.e., streamers (Rusconi et al., 2010). Streamers are frequently observed in soil-like porous environments and industrial filters. Biofilm streamers may cause rapid clogging in porous materials (Drescher et al., 2013). Flow-induced shedding of the extracellular matrix from surface-attached biofilms generates a sieve-like network that exponentially accelerates clogging.
The impact of fluid flow has been studied using microfluidic devices (Biswas et al., 2016). Rusconi et al. (2010) investigated streamer formation in curved microchannels under laminar flow. The streamers were spatially localized to the middle plane of the curved channel and connected only at the inner curve during each turn, where a secondary vortical flow was observed. The effect of secondary flow on biofilm formation was evaluated by Marty et al. (2014). They discussed the role of secondary flow in streamer formation in three steps. The first step was shear-enhanced adhesion following the streamlines toward the surfaces at the pore entrance. The second step was formation of filaments. EPSs form filaments and, as the third step, the filaments form a net, which captures floating bacteria. Hydrodynamics profoundly affects how bacteria compete and evolve in biofilms (Coyte et al., 2017). Thus, hydrodynamical modeling of biofilm formation is important for predicting and controlling their development and detachment.
C. Physical properties of biofilms
Even et al. (2017) reviewed the mechanical properties of biofilms, which affect the deformation and breakage of biofilms. Several measurement techniques, such as rheometry, uniaxial compression, atomic force microscopy, and hydrodynamic load, have been used to assay the macroscopic mechanical properties of biofilms. These techniques revealed that biofilms are viscoelastic. However, the elastic modulus and viscosity values of the biofilm strongly depend on the experimental settings. The effective shear modulus ranges widely from 10−2 to 106 Pa (Even et al., 2017). The storage and loss moduli G′ and G″ were also measured (Lieleg et al., 2011). The results suggested that biofilms behave as viscoelastic solids rather than viscoelastic liquids, because G′ was about tenfold larger than G″. However, this was dependent on the time scale of the experiment (Galy et al., 2012). Strain-hardening and viscoplastic effects have also been reported (Hollenbeck et al., 2016).
The diffusivity inside biofilms is important because it modulates mass transport. Stewart (2003) discussed the influence of diffusion on the chemistry and biology of biofilms. He measured the effective diffusion coefficient in the biofilm De and showed that De is reduced compared to that in water (Daq) because of the presence of microbial cells and extracellular polymers. The reduction ratio De/Daq was estimated to be 0.6 for light gases (e.g., oxygen, nitrous oxide, carbon dioxide, or methane) and 0.25 for most organic solutes. For light gases, we have the scaling De L2/T 0.6Daq, where L is the length scale, T is the time scale, and Daq 10−9 m2/s. The timescale for diffusive equilibration can be scaled as T L2/(0.6Daq), which ranges from one second to tens of minutes when the length scale is tens to thousands of micrometers. Also, topological heterogeneity does not alter the fundamental phenomena of diffusive transport in a biofilm.
Physical properties of biofilms composed of multi-species communities have been investigated. A dual-species biofilm, composed of Vibrio and E. coli, showed competition at the interface between the two species (Abriat et al., 2020). The viscoelastic properties of the dual-species biofilm were dominated by Vibrio, because Vibrio could form a mature biofilm faster than E. coli. A dual-species biofilm of Bacillus and Pseudomonas was investigated by Abriat et al. (2019). Mechanical properties of the biofilm were dominated by Pseudomonas, and the study of the planktonic and biofilm growths for each species revealed that Pseudomonas grew faster than Bacillus. These studies illustrate the importance of growth kinetics in the bacteria competition for the interface in a dual-species biofilm. Mixed-species cultures grow biofilms that are more robust and protective to bacteria than biofilms grown from single-species cultures (Burmolle et al., 2006; Wilking et al., 2011; Gordon et al., 2017). Paramonova et al. (2009) investigated the influence of hydrodynamics on oral biofilms composed of single- and multi-species, because mechanical removal of oral biofilms is accepted as the best way to prevent caries and periodontal diseases. They found that multi-species biofilms were stronger than single-species biofilms. In response to increased hydrodynamic shear, biofilm strength decreased and architecture changed from uniform carpet-like to fluffy with a higher thickness.
D. Modeling of biofilm formation
To simulate biofilm formation, several mechanical models have been developed. There are two major types of biofilm models; individual particle-based and continuum multiphase flow models. These are described in the following.
A particle-based biofilm model is focused on individual cell behaviors in an extracellular matrix. Cells are typically represented by Lagrangian spheres in an Eulerian liquid domain. Biomass accumulation and growth are simulated as new cells are added to the biofilm. Picioreanu et al. (2004) developed a particle-based biofilm model coupled to a reaction-diffusion equation. The growth of biomass was given by the reaction rate, which was described by the Monod equation. In order to express the boundary condition of mass flux through the biofilm surface, a concentration boundary layer can be introduced (Xavier et al., 2005). Dillon et al. (1996) developed a fluid-structure interaction model by the immersed boundary method. Cell–cell and cell-wall adhesions were modeled by elastic springs between points on each adherent entity. The total force acting on a particle is then transmitted to the fluid using an immersed boundary method.
These particle-based models reproduced three-dimensional two-species biofilm formation and predicted the spatiotemporal distributions of bacteria and substrates (Picioreanu et al., 2004; Xavier et al., 2005). For example, Xavier et al. (2005) simulated the formation of biofilms by polyhydroxybutyrate (PHB)- and EPSs-producing organisms. The PHB-producing organisms achieved dominance by supplying substrate intermittently in feast/famine cycles, as observed experimentally. Kitamura et al. (2019) reproduced streamers formed in a microchannel with blocks (Fig. 5). Particle-based modeling is a bottom-up approach and can be extended to multispecies biofilms.
FIG. 5.

Particle-based simulation of the biofilm. A bacterial suspension flows through blocks, and streamers are formed on the downstream side. Spheres represent individual bacteria that have adhered to each other and the blocks.
Biofilms can also be simulated by continuum multiphase flow modeling (Albero et al., 2014; Batchelor et al., 2018; Cogan and Keener, 2004; Horn and Lackner, 2014; Picioreanu et al., 2004; Rittmann, 1982; Taylor and Jaffe, 1990; Winstanley et al., 2011; Zhang et al., 2008). Bacterial biofilms may be modeled as a polymer solution immersed in a Newtonian liquid. Biofilm motion and growth are described by mass conservation of the polymer phase, solvent phase, and nutrients, as well as by momentum conservation in both the polymer and solvent phases. Time-dependent growth of biofilms is then simulated by coupling these equations. The models provide insight into biofilm development, such as biofilm dynamics in shear flow (Zhang et al., 2008) and the relationship between osmotic pressure and biofilm morphology (Cogan and Keener, 2004).
V. GUT FLORA
The number of bacteria in the intestine can be much larger than the total number of human cells (Guarner and Malagelada, 2003). Gut bacteria form a complex ecosystem, known as the gut flora or microbiota, and play important roles in homeostasis and health. The human gut flora has metabolic activity, tropic and immunologic effects, and prevents colonization by pathogens. The microbiota is also linked to disorders such as obesity, colon cancer, inflammatory bowel disease (IBD), and cardiovascular diseases (CVDs). Therefore, predicting and controlling the microbiota are important in medicine and biology.
Biomechanics can be a strong tool for elucidation of gut flora composition and activity, because the distribution and growth of bacteria are dependent on transport phenomena. In this section, we introduce the role of the gut flora in health (Sec. V A). Heterogeneous microbial distributions and the effect of hydrodynamics are discussed in Sec. V B. Zebrafish models of the gut flora have been developed, and so we describe the gut flora of zebrafish (Sec. V C). In Sec. V D, we cover mathematical modeling of the gut flora. We suggest the following as supplementary reading: Guarner and Malagelada (2003) reviewed the gut flora in health and disease, Donaldson et al. (2016) explained the spatial distribution of gut microbiota, Chahill (1990) focused on the bacterial flora of fish, and Song et al. (2014) described modeling of microbial community dynamics.
A. Role of the gut flora
Approximately 1013–1014 bacterial cells and 15 000–36 000 bacterial species reside in the gut lumen (Sartor, 2008). Greater than 99% of the gut flora is composed of four major bacterial divisions—Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria (Frank et al., 2007). Enterobacteriaceae, such as E. coli, are relatively minor components of the division Proteobacteria (8% of all gut bacteria).
The microbiota produces several nutrients, including short-chain fatty acids (SCFAs), B vitamins, and vitamin K. SCFAs are fermented and produced from dietary fiber by the gut microbiota, and they modulate host health by influencing gut barrier function, glucose homeostasis, immunity, appetite, and obesity (Chambers et al., 2018). Lactic acid bacteria (LAB) synthesize vitamin K and water-soluble B vitamins. Many herbivores are dependent on the digestion of plant material (e.g., cellulose, hemicellulose, and pectin) by the gut microbiota (Hanning and Diaz-Sanchez, 2015). Thus, the gut microbiota contributes to the production of dietary components.
Symbiosis with gut flora has both positive and negative effects on host health. The gut flora is linked to pathological disorders, such as allergies, obesity, colon cancer, IBD, and CVD (Zhnag et al., 2015). Allergic disease in infants is caused by delayed establishment of Bifidobacterium and Lactobacillus in the gut, which regulate allergic-type immune responses (Kalliomäki and Isolauri, 2003). Fewer Bacteroidetes and more Firmicutes in the gut microbiota facilitate the extraction of calories from ingested dietary substances and promote their storage in adipose tissue, leading to obesity (DiBaise, et al., 2008). A decrease in gut microbial diversity caused by a shift in the balance between commensal and potentially pathogenic microorganisms is associated with inflammatory bowel diseases, including Crohn's disease and ulcerative colitis (Ni et al., 2017). SCFAs, which are produced by the gut microbiota, are also linked to the risk of CVD. Systolic and diastolic blood pressures are associated with decreased butyrate and plasminogen activator inhibitor-1 levels in early pregnancy (Chambers et al., 2018).
The role of gut microbial biofilms in colorectal cancer is reviewed by Hold and Allen-Vercoe (2019). Initially, small aggregates of bacterial cells attach to mucosal surfaces. Then, mature biofilms develop when these adherent microcolonies become encapsulated in a self-secreted polysaccharide matrix. Dejea et al. (2014) identified invasive polymicrobial bacterial biofilms nearly universally on right-sided tumors but on only 12% of left-sided tumors. The results illustrate that the mucosa-associated microbial community is an important factor in colorectal cancer pathogenesis, particularly in the proximal colon. In addition, a number of healthy individuals also harbored mucosal biofilms. The complex microbial communities of the gut microbiota reside over the intestinal mucus as exopolysaccharide-coated biofilms, which disperse planktonic bacteria (Buret et al., 2019). The mucosal biofilm bacteria differ phylogenetically and metabolically from those living in a planktonic state (Beatty et al., 2017; Macfarlane et al., 2005).
The formation of biofilms also protects the bacterial population from host immune responses and antibiotics by secretion of EPSs and binding bacteria together in layers (Swidsinski et al., 2005). It can prevent the loss of useful secretions and nutrients from the population. IBD is a group of intestinal disorders in which prolonged inflammation occurs in the digestive tract. Biofilm formation in the digestive tract has an adverse effect on the immune response of the host. There is no satisfactory and safe treatment option for IBD. Therefore, the current research aims to disrupt biofilms in IBD and concentrates on improving the drug (Srivastava et al., 2017).
One clear limitation in targeting the distribution of intestinal flora as the cause of chronic disease is the possibility that the inciting microbiota is no longer active at the time the disease is identified, perhaps because of a gradual shift or change in the intestinal environment. Therefore, real-time evaluation of gut motility and microbial diversity would be likely to clarify the causes of intestinal disorders.
B. Dynamics of the gut flora
Studies of microbiota spatial organization in the vertebrate gut have revealed functional relationships between biogeography and health, driven by coincident revolutions in imaging and sequencing technologies (Tropini et al., 2017). It is becoming clear that spatial redistribution of the microbiota can be a common and functionally relevant feature of chronic inflammatory diseases. The density and composition of the microbiota are affected by chemical, nutritional, and immunological fields along the gut (Thursby and Juge, 2017). In the small intestine, there are typically high levels of acids, oxygen, and antimicrobials and a short transit time. These properties limit bacterial growth such that only rapidly growing anaerobes with the ability to adhere to epithelia mucus would survive (Donaldson et al., 2015). In contrast, colonic conditions support a dense and diverse community of bacteria, mainly anaerobes, where Prevotellaceae, Lachnospiraceae, and Rikenellaceae are dominant. Spatial metagenomics can be a strong tool to study microbial biogeography in complex habitats. Sheth et al. (2019) characterized the spatial organization of a microbiome by spatial metagenomics and revealed heterogeneous microbial distributions. They showed that phylogenetically clustered local regions of bacteria were associated with a dietary perturbation.
Heterogeneous microbial distributions have been found in various vertebrate guts. Bacteria within the mouse gut migrate to closer proximity to the epithelium when diet lacks microbiota accessible carbohydrates (Earle et al., 2015). Suzuki and Nachman (2016) investigated microbial communities from ten different segments of the gastrointestinal tract (mouth, esophagus, stomach, duodenum, ileum, proximal cecum, distal cecum, colon, rectum, and feces) in wild house mice. The lower gastrointestinal tract had a greater relative abundance of anaerobic bacteria and greater microbial diversity relative to the upper gastrointestinal tract. A large-scale gene sequencing was performed for newborn piglets. Using intestinal digesta, Liu et al. (2019) showed that the six intestinal segments could be divided into three parts; in the duodenum-jejunum section, the most abundant genera included Lactobacillus and Bacteroides; in the ileum, Fusobacterium and Escherichia; and in the cecum-rectum section, Prevotella. Schlomann et al. (2018) observed bacterial distribution patterns throughout the intestinal volume of live larval zebrafish. Fluorescently tagged strains of seven bacterial symbionts showed large differences in both cohesion and spatial distribution. Significant spatial heterogeneity in composition, diversity, and species of gut microbiota was also found in wild folivorous flying squirrels (Lu et al., 2014).
Hydrodynamic forces in the colon, along with colonic water absorption that manifests as transit time, exert a significant impact on microbiota density and composition. Arnoldini et al. (2018) explained the hydrodynamic effect on colonic pH, which directly affects microbiota competition for food. They also discussed the mixing dynamics of luminal content by wall contractions and its implications for bacterial growth and density. Active mixing of colonic contents was found to be crucial to prevent washout of bacteria. Transport phenomena of bacteria across and along the gastrointestinal tract were experimentally investigated by Takahashi and Sakaguchi (2006). They injected fluorescently labeled viable bacteria into the proximal colon of the guinea pig to observe transport in the large intestinal lumen. Their results showed that bacteria were transported along the radial and longitudinal axes of the intestine and even transported back to the cecum. They speculated that the spreading of bacteria upstream may be caused by the backward flow of dietary residue in the furrow of the proximal colon (Takahashi and Sakaguchi, 2000).
The effect of viscosity on the growth of gut microbiota at physiological conditions was investigated by Tamargo et al. (2018). They performed in vitro experiments using a gastrointestinal simulator, in which the viscosity was controlled by the agar concentrations. The results indicated that changes in intestinal viscosity selectively modify the microbiota composition. Cremer et al. (2016) developed a fluidic channel that allows mimicking active contractions of the colonic wall. They found that repeated contraction was crucial to prevent bacterial washout and maintain a steady-state bacterial population, which illustrates that flow and mixing play a major role in shaping the microbiota of the colon.
Parthasarathy (2018) visualized swimming behaviors of bacteria in the larval zebrafish gut by using a light sheet microscope. Wiles et al. (2020) also investigated the spatial organization and dynamics of bacterial populations within the larval zebrafish gut. They found that a proinflammatory Vibrio symbiont governs its own spatial organization using swimming motility and chemotaxis. The motility did not enhance its growth rate but rather promoted its persistence by resisting the intestinal flow. In contrast, nonmotile Vibrio mutants surrendered to the intestinal flow. Consequently, nonmotile and nonchemotactic mutants were susceptible to intestinal expulsion.
C. Zebrafish as a model of the gut flora
The use of mammalian in vivo models of gastrointestinal chyme transport is limited by the physical space needed for housing and by the labor-intensive surgery and invasive imaging techniques used (Field et al., 2009). As a model organism, the zebrafish, Danio rerio, has been widely used to investigate the roles of environmental, genetic, or chemical perturbations in gastrointestinal motility and transport. Zebrafish genetics, organ physiology, and metabolism are similar to those of human (Barbazuk et al., 2000; Quinlivan and Farber, 2017; Zang et al., 2017). The > 500 mutant phenotypes—which are associated with hematopoietic, cardiovascular, kidney, and other-organ disorders—of zebrafish enable development and genetics studies (Dooley and Zon, 2000). The zebrafish also has high fertility and a short generation time (Freifeld et al., 2017).
Real-time in vivo observation enables analysis of the developmental morphology and function of organs. The transparent larval body facilitates noninvasive real-time observation of organ inner structures. In vivo digital motion analysis in Danio rerio zebrafish (3–7 days post fertilization [dpf]) showed that the peristaltic frequency was in the range of 0.015–0.025 Hz, while the velocity of peristaltic wave propagation was around 15 μm/s (Holmberg et al., 2003). The viscosity of fluid in the anterior intestine of zebrafish larvae was measured using oscillating magnetic prolate fluorescent particles as 0.5 mPa s, i.e., approximately fivefold larger than that of water (Taormina et al., 2017). By feeding fluorescent polystyrene microspheres mixed with powdered feed to a zebrafish larva at 7 dpf, intestinal transit was observed under a fluorescent microscope (Field et al., 2009). Even 12 h after feeding, 72% of microspheres remained, while they were absent 24 h after feeding. Cosshiaro et al. (2013) showed that 50% of zebrafish larva at 7 dpf eliminated the injected material in 12 hours.
Kikuchi et al. (2020) performed real-time in vivo imaging of zebrafish larva at 7 dpf for 5 h after feeding and investigated the mechanical roles of anterograde and retrograde intestinal peristalsis in the anterior and posterior intestine. They derived the Péclet number (Fig. 6), which indicates the ratio of advection to diffusion transport. A scaling analysis showed that, after feeding, retrograde peristalsis of the anterior intestine continuously mixes the contents, whereas anterograde peristalsis of the posterior intestine first mixes the contents and then transports them toward the anus. Thus, the role of intestinal peristalsis is drastically changed after feeding.
FIG. 6.
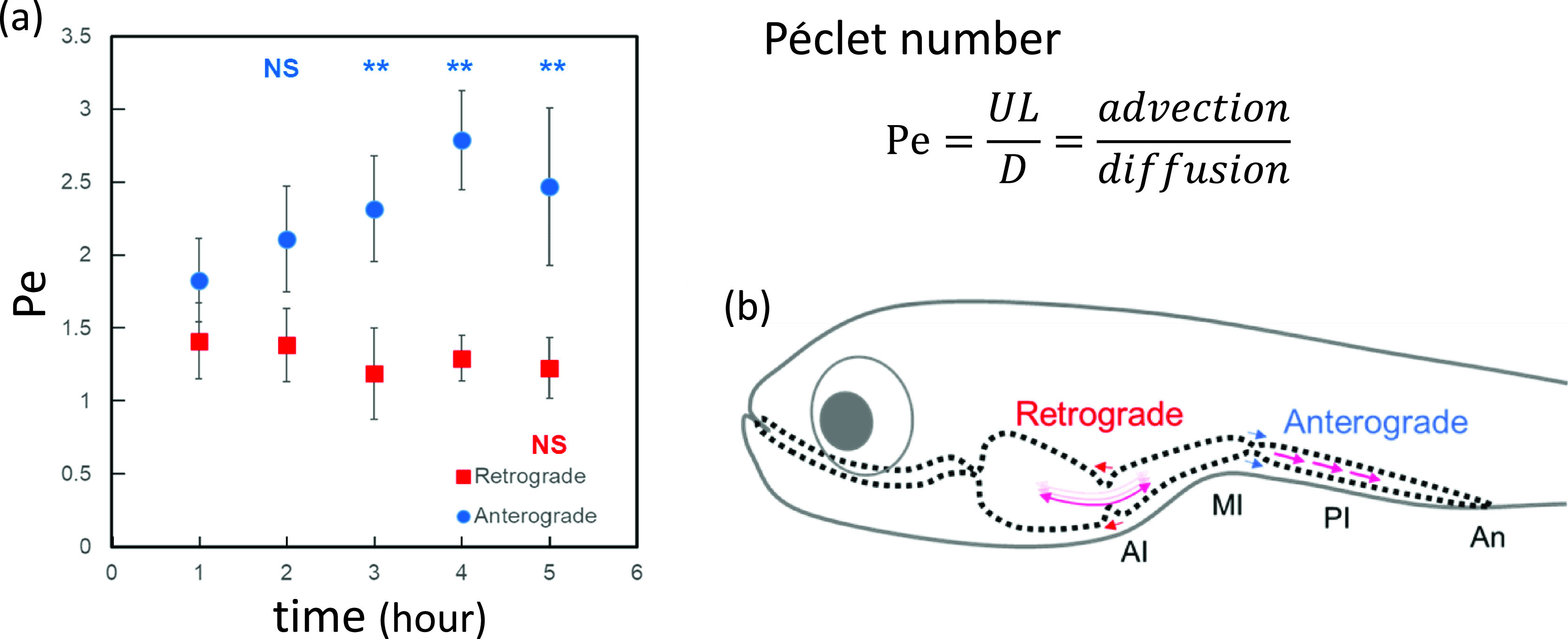
Mechanical roles of anterograde and retrograde intestinal peristalsis in zebrafish larva. (a) Péclet numbers after injection of food into the anterior and posterior intestine. (b) Schematic of food mixing and advection in the zebrafish larval intestine. AI, anterior intestine; MI, middle intestine; PI, posterior intestine; and An, anus. Reproduced with the permission from Kikuchi et al., Am. J. Physiol. 318, G1013–G1021 (2020). Copyright 2020 APS.
The zebrafish larval mouth first opens at 74 h post-fertilization (hpf), followed by the anus at 96 hpf, at which point the gut is a continuous tube accessible to the outside environment (Ng et al., 2005). The gut becomes fully functional by 5 dpf, when uptake of lipid and protein begins (Bates et al., 2006). The zebrafish intestinal microbiota composition changes with development (from 4 to 380 days of age) (Stephens et al., 2016). The spatiotemporal transport of the gut chyme and microbiota in the zebrafish intestine was observed by noninvasive microscopy (Bates et al., 2006; Cocchiaro and Rawls, 2013). Jemielita et al. (2014) reported that Aeromonas in the larval zebrafish intestine exhibits a heterogeneous distribution and aggregates in the middle of the intestinal tract. Wiles et al. (2016) investigated the distribution of Aeromonas by introducing Vibrio into the larval zebrafish intestine. The habitat of Aeromonas was modified by competition with Vibrio, altering its distribution. The intestinal inflammatory damage in a microbiota-containing larval zebrafish intestine was more extensive than in that of germ-free fish (Qi et al., 2014).
D. Modeling of the gut flora
Mathematical modeling enables investigation of the distribution and growth of bacteria by clarifying complex phenomena via well-defined numerical experiments. Mathematical models of the gut bacterial flora have been reported (Ishikawa et al., 2011; Cremer et al., 2016; Yang et al., 2018). In this section, we introduce the computational model of Yang et al. (2018). They attempted to reproduce the findings of Jemielita et al. (2014), in which Aeromonas in the larval zebrafish intestine exhibited a heterogeneous distribution and aggregated in the middle of the intestinal tract.
Yang et al. (2018) expressed the distribution and growth of bacteria in the intestine of a zebrafish larva using four conservation laws: (i) conservation of solvent fluid, (ii) conservation of momentum, (iii) conservation of nutrients, and (iv) conservation of bacteria. The results of these equations, however, did not capture the heterogeneous distribution experimentally observed by Jemielita et al. (2014) [Figs. 7(a)–7(d)]. In the intestine, bacterial taxis can be triggered by nutrients, pH, temperature, oxygen, antimicrobials, and other factors. Yang et al. (2018) mathematically expressed bacterial taxis as a potential field. This enabled reproduction of the heterogeneous distribution [Fig. 7(e)]. The diffusion constants in the governing equations are not equivalent to Brownian diffusivity, because the effects of bacterial swimming need to be considered. To obtain such macroscopic properties, an understanding of the individual and collective motions of cells is needed. This is facilitated by a bottom-up understanding, i.e., from the cellular to the macroscale.
FIG. 7.
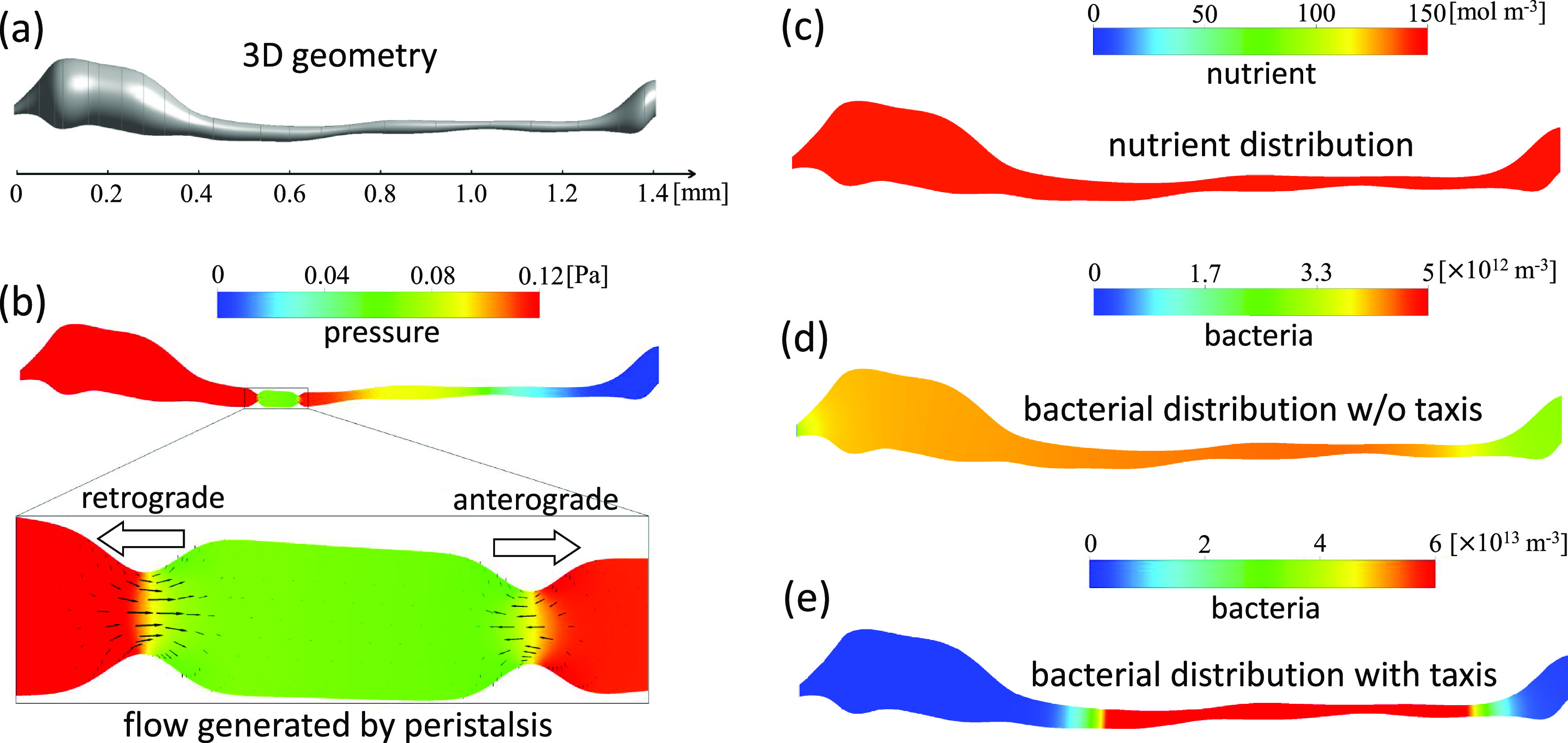
Simulation of the gut flora of a zebrafish larva. (a) Three-dimensional geometry of the intestine. (b) Pressure distribution and velocity vectors. White arrows, retrograde and anterograde peristalsis. (c) Nutrient distribution. (d) Bacterial distribution without taxis. (e) Bacterial distribution with taxis. Reproduced with the permission from Yang et al., J. Theor. Biol. 446, 101–109 (2018). Copyright 2018 Elsevier.
VI. FUTURE PROSPECTS
In this Review, we introduced recent advances in the biomechanics of bacterial swimming, adhesion to surfaces, collective motion, biofilm formation, and the gut flora. Below, we discuss future prospects in bacterial biomechanics.
Swimming behaviors and chemotaxis of bacteria have been clarified in terms of biomechanics, and their bulk motion in a concentration field can be mathematically described. However, bacterial behaviors in complex environments are unclear. Complex environments have geometric constraints (Ishikawa, 2019), surface chemical properties, background flow fields, concentration fields, and non-Newtonian fluids. Because the physical environment of biofilms and the digestive system is complex, knowledge of bacterial behaviors in complex environments has to be strengthened.
Biomechanics of bacterial adhesion on surfaces has been investigated. However, the interplay among adhesion strategies, including appendages like pili and fimbriae and EPS secretion, has not been fully clarified. The physical environment, such as shear stress, can also strengthen (i.e., catch bonds) or weaken (i.e., slip bonds) bonding strength. To fully understand cell adhesion, experimental data at a cellular resolution have to be accumulated. In addition, our understanding of the population dynamics within a biofilm needs to be improved, because cell division and EPS secretion of adhered bacteria are essential in biofilm formation.
Collective swimming of bacteria has been investigated intensively, and the physical mechanisms of coherent structures emerging within a bacterial bath have been understood. However, bacterial swarming has not been modeled precisely, and several ad hoc assumptions have been made. Swarming is a particular biological mode that involves intercellular chemical communication as well as mechanical changes (Beer and Ariel, 2019), which have not been well described mathematically. Transport on an agar gel surface is more complex than that on a solid wall, hampering quantitative investigation of colony-pattern formation. Biomechanics of swarming and colony-pattern formation is an interesting research field to be explored.
Transport phenomena in a biofilm influence cell division and EPS secretion and, consequently, biofilm growth, which in turn influence the surrounding flow field, viscous drag, mass transport, and population dynamics of bacteria. Such crosstalk between bacteria and their environment needs further study. While transport phenomena outside a biofilm can be described mathematically, modeling the population dynamics of bacteria is still challenging. This is in part because (i) we have not obtained sufficient experimental data to establish a mathematical model, (ii) the interactions are complex and contain many variables, and (iii) the parameters in such mathematical models are not well defined or supported experimentally.
Particle-based models can deal with individual bacteria, i.e., cellular resolution. The physical meaning of hydrodynamic, interparticle, and adhesion forces in particle-based models is clear, and the estimation of physical parameters is more straightforward than in continuum models. The drawback of particle-based models is the high computational load required to resolve large tempo-spatial scales. By contrast, macroscopic properties are employed by continuum models, facilitating investigation of large tempo-spatial scales. However, the physical meaning of parameters becomes vague, and ad hoc assumptions are often introduced. Combining the advantages of both models will enable development of a model with cellular resolution that can handle large tempo-spatial scales.
Modeling the gut flora is also a challenging task. Approximately 15 000–36 000 bacterial species live in the gut lumen, and an understanding of the interplay between them and the host is needed. A simplified gut flora model would be an important first step. Jemielita et al. (2014) inoculated germ-free zebrafish larvae with fluorescently labeled Aeromonas species, which is abundant in the zebrafish gut. The presence of a single bacterial species enables interspecies interplay to be ignored. As a second step, Wiles et al. (2016) investigated the distribution of Aeromonas by introducing another bacterium, Vibrio, into the larval zebrafish intestine. This setup enables investigation of the interplay between the two bacterial species. Hence, germ-free zebrafish are expected to provide useful experimental data for modeling of the gut flora.
ACKNOWLEDGMENTS
This study was partly supported by the Japan Society for the Promotion of Science Grant-in-Aid for Scientific Research (JSPS KAKENHI Grant No. 17H00853).
DATA AVAILABILITY
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
- 1. Abriat, C. , Enriquez, K. , Virgilio, N. , Cegelski, L. , Fuller, G. G. , Daigle, F. , and Heuzey, M.-C. , “ Mechanical and microstructural insights of Vibrio cholerae and Escherichia coli dual-species biofilm at the air-liquid interface,” Colloids Surf., B 188, 110786 (2020). 10.1016/j.colsurfb.2020.110786 [DOI] [PubMed] [Google Scholar]
- 2. Abriat, C. , Virgilio, N. , Heuzey, M.-C. , and Daigle, F. , “ Microbiological and real-time mechanical analysis of Bacillus licheniformis and Pseudomonas fluorescens dual-species biofilm,” Microbiology 165, 747–756 (2019). 10.1099/mic.0.000819 [DOI] [PubMed] [Google Scholar]
- 3. Albareda, M. , Dardanelli, M. S. , Sousa, C. , Megias, M. , Temprano, F. et al. , “ Factors affecting the attachment of rhizospheric bacteria to bean and soybean roots,” FEMS Microbiol. Lett. 259, 67–73 (2006). 10.1111/j.1574-6968.2006.00244.x [DOI] [PubMed] [Google Scholar]
- 4. Albero, A. B. , Ehret, A. E. , and Bol, M. , “ A new approach to the simulation of microbial biofilms by a theory of fluid-like pressure-restricted finite growth,” Comput. Methods Appl. Mech. Eng. 272, 271–289 (2014). 10.1016/j.cma.2014.01.001 [DOI] [Google Scholar]
- 5. Ariel, G. et al. , “ Swarming bacteria migrate by Levy walk,” Nat. Commun. 6, 8396 (2015). 10.1038/ncomms9396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Arnoldini, M. , Cremer, J. , and Hwa, T. , “ Bacterial growth, flow, and mixing shape human gut microbiota density and composition,” Gut Microbes 9, 559–566 (2018). 10.1080/19490976.2018.1448741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Batchelor, G. K. , “ The stress system in a suspension of force-free particles,” J. Fluid Mech. 41, 545–570 (1970). 10.1017/S0022112070000745 [DOI] [Google Scholar]
- 8. Batchelor, M. T. , Burne, R. V. , Henry, B. I. , Li, F. , and Paul, J. , “ A biofilm and organomineralisation model for the growth and limiting size of ooids,” Sci. Rep. 8, 559 (2018). 10.1038/s41598-017-18908-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Bates, J. M. , Mittge, E. , Kuhlman, J. , Baden, K. N. , Cheesman, S. E. , and Guillemin, K. , “ Distinct signals from the microbiota promote different aspects of zebrafish gut differentiation,” Dev. Biol. 297, 374–386 (2006). 10.1016/j.ydbio.2006.05.006 [DOI] [PubMed] [Google Scholar]
- 10. Bar-On, Y. M. , Phillips, R. , and Milo, R. , “ The biomass distribution on Earth,” Proc. Natl. Acad. Sci. U. S. A. 115, 6506–6511 (2018). 10.1073/pnas.1711842115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Barbazuk, W. B. , Korf, I. , Kadavi, C. , Heyen, J. , Tate, S. , Wun, E. , Bedell, J. A. , McPherson, J. D. , and Johnson, S. L. , “ The syntenic relationship of the zebrafish and human genomes,” Genome Res. 10, 1351–1358 (2000). 10.1101/gr.144700 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Beatty, J. K. et al. , “ Giardia duodenalis induces pathogenic dysbiosis of human intestinal microbiota biofilms,” Int. J. Parasitol. 47, 311–326 (2017). 10.1016/j.ijpara.2016.11.010 [DOI] [PubMed] [Google Scholar]
- 13. Beer, A. , and Ariel, G. , “ A statistical physics view of swarming bacteria,” Movement Ecol. 7, 9 (2019). 10.1186/s40462-019-0153-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Berg, H. C. , “ The rotary motor of bacterial flagella,” Annu. Rev. Biochem. 72, 19–54 (2003). 10.1146/annurev.biochem.72.121801.161737 [DOI] [PubMed] [Google Scholar]
- 15. Berg, H. C. , E. coli in Motion ( Springer-Verlag, New York, 2004). [Google Scholar]
- 16. Berke, A. P. , Turner, L. , Berg, H. C. , and Lauga, E. , “ Hydrodynamic attraction of swimming microorganisms by surfaces,” Phys. Rev. Lett. 101, 038102 (2008). 10.1103/PhysRevLett.101.038102 [DOI] [PubMed] [Google Scholar]
- 17. Biswas, I. , Ghosh, R. , Sadrzadeh, M. , and Kumar, A. , “ Nonlinear deformation and localized failure of bacterial streamers in creeping flows,” Sci. Rep. 6, 32204 (2016). 10.1038/srep32204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Brooks, J. D. , and Flint, S. H. , “ Biofilms in the food industry: Problems and potential solutions,” Int. J. Food Sci. Technol. 43, 2163–2176 (2008). 10.1111/j.1365-2621.2008.01839.x [DOI] [Google Scholar]
- 19. Brown, A. T. et al. , “ Swimming in a crystal,” Soft Matter 12, 131–140 (2016). 10.1039/C5SM01831E [DOI] [PubMed] [Google Scholar]
- 20. Buret, A. G. , Motta, J.-P. , Allain, T. , Ferraz, J. , and Wallace, J. L. , “ Pathobiont release from dysbiotic gut microbiota biofilms in intestinal inflammatory diseases: A role for iron?,” J. Biomed. Sci. 26, 1 (2019). 10.1186/s12929-018-0495-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Burmolle, M. , Webb, J. , Rao, D. , Hansen, L. , Sorensen, S. , and Kjelleberg, S. , “ Enhanced biofilm formation and increased resistance to antimicrobial agents and bacterial invasion are caused by synergistic interactions in multispecies biofilms,” Appl. Environ. Microbiol. 72, 3916–3923 (2006). 10.1128/AEM.03022-05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Campoccia, D. , Montanaro, L. , and Arciola, C. R. , “ A review of the clinical implications of anti-infective biomaterials and infection-resistant surfaces,” Biomaterials 34, 8018–8029 (2013). 10.1016/j.biomaterials.2013.07.048 [DOI] [PubMed] [Google Scholar]
- 23. Cates, M. E. , “ Diffusive transport without detailed balance in motile bacteria: Does microbiology need statistical physics?,” Rep. Prog. Phys. 75, 042601 (2012). 10.1088/0034-4885/75/4/042601 [DOI] [PubMed] [Google Scholar]
- 24. Chahill, M. M. , “ Bacterial flora of fishes: A review,” Microb. Ecol. 19, 21–41 (1990). 10.1007/BF02015051 [DOI] [PubMed] [Google Scholar]
- 25. Chambers, E. S. , Preston, T. , Frost, G. , and Morrison, D. J. , “ Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health,” Curr. Nutr. Rep. 7, 198–206 (2018). 10.1007/s13668-018-0248-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Chamolly, A. , Ishikawa, T. , and Lauga, E. , “ Active particles in periodic lattices,” New J. Phys. 19, 115001 (2017). 10.1088/1367-2630/aa8d5e [DOI] [Google Scholar]
- 27. Chen, X. , and Berg, H. C. , “ Torque-speed relationship of the flagellar rotary motor of Escherichia coli,” Biophys. J. 78, 1036–1041 (2000). 10.1016/S0006-3495(00)76662-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Cocchiaro, J. L. , and Rawls, J. F. , “ Microgavage of zebrafish larvae,” J. Vis. Express 72, e4434 (2013). 10.3791/4434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Codling, E. A. , Plank, M. J. , and Benhamou, S. , “ Random walk models in biology,” J. R. Soc. Interface 5, 813–834 (2008). 10.1098/rsif.2008.0014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Cogan, N. G. , and Keener, J. P. , “ The role of the biofilm matrix in structural development,” Math. Med. Biol. 21, 147–166 (2004). 10.1093/imammb/21.2.147 [DOI] [PubMed] [Google Scholar]
- 31. Coyte, K. , Tabuteau, H. , Gaffney, E. A. , Foster, K. R. , and Durham, W. M. , “ Microbial competition in porous environments can select against rapid biofilm growth,” Proc. Natl. Acad. Sci. U. S. A. 114, E161–E170 (2017). 10.1073/pnas.1525228113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Cremer, J. , Segota, I. , Yang, C. , Arnoldini, M. , Sauls, J. T. , Zhang, Z. , Gutierrez, E. , Groisman, A. , and Hwa, T. , “ Effect of flow and peristaltic mixing on bacterial growth in a gut-like channel,” Proc. Natl. Acad. Sci. U. S. A. 113, 11414–11419 (2016). 10.1073/pnas.1601306113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Darnton, N. C. , Turner, L. , Rojevsky, S. , and Berg, H. C. , “ On torque and tumbling in swimming Escherichia coli,” J. Bacteriol. 189, 1756–1764 (2007). 10.1128/JB.01501-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Dejea, C. M. et al. , “ Microbiota organization is a distinct feature of proximal colorectal cancers,” Proc. Natl. Acad. Sci. U. S. A. 111, 18321–18326 (2014). 10.1073/pnas.1406199111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. DiBaise, J. K. , Zhang, H. , Crowell, M. D. , Krajmalnik-Brown, R. , Decker, G. A. , and Rittmann, E. B. , “ Gut microbiota and its possible relationship with obesity,” Mayo Clinic Proc. 83, 460–469 (2008). 10.4065/83.4.460 [DOI] [PubMed] [Google Scholar]
- 36. Dillon, R. , Fauci, L. , Fogelson, A. , and Gaver III, D. , “ Modeling biofilm processes using the immersed boundary method,” J. Comput. Phys. 129, 57–73 (1996). 10.1006/jcph.1996.0233 [DOI] [Google Scholar]
- 37. Dooley, K. , and Zon, L. I. , “ Zebrafish: A model system for the study of human disease,” Curr. Opin. Genet. Dev. 10, 252–256 (2000). 10.1016/S0959-437X(00)00074-5 [DOI] [PubMed] [Google Scholar]
- 38. Drescher, K. , Shen, Y. , Bassler, B. L. , and Stone, H. A. , “ Biofilm streamers cause catastrophic disruption of flow with consequences for environmental and medical systems,” Proc. Natl. Acad. U. S. A. 110, 4345–4350 (2013). 10.1073/pnas.1300321110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Dombrowski, C. , Cisneros, L. , Chatkaew, S. , Goldstein, R. E. , and Kessler, J. O. , “ Self-concentration and large-scale coherence in bacterial dynamics,” Phys. Rev. Lett. 93, 098103 (2004). 10.1103/PhysRevLett.93.098103 [DOI] [PubMed] [Google Scholar]
- 40. Donaldson, G. P. , Lee, S. M. , and Mazmanian, S. K. , “ Gut biogeography of the bacterial microbiota,” Nat. Rev. Microbiol. 14, 20–32 (2016). 10.1038/nrmicro3552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Dong, J. , and Klumpp, S. , “ Simulation of colony pattern formation under differential adhesion and cell proliferation,” Soft Matter 14, 1908–1916 (2018). 10.1039/C7SM02451G [DOI] [PubMed] [Google Scholar]
- 42. Dufrêne, Y. F. , and Persat, A. , “ Mechanomicrobiology: How bacteria sense and respond to forces,” Nat. Rev. Microbiol. 18, 227–240 (2020). 10.1038/s41579-019-0314-2 [DOI] [PubMed] [Google Scholar]
- 43. Dunkel, J. , Heidenreich, S. , Drescher, K. , Wensink, H. H. , Bar, M. , and Goldstein, R. E. , “ Fluid dynamics of bacterial turbulence,” Phys. Rev. Lett. 110, 228102 (2013). 10.1103/PhysRevLett.110.228102 [DOI] [PubMed] [Google Scholar]
- 44. Earle, K. A. et al. , “ Quantitative imaging of gut microbiota spatial organization,” Cell Host Microbe 18, 478–488 (2015). 10.1016/j.chom.2015.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Elgeti, J. , Winkler, R. G. , and Gompper, G. , “ Physics of microswimmers—single particle motion and collective behavior: A review,” Rep. Prog. Phys. 78, 056601 (2015). 10.1088/0034-4885/78/5/056601 [DOI] [PubMed] [Google Scholar]
- 46. Evans, A. A. , Ishikawa, T. , Yamaguchi, T. , and Lauga, E. , “ Orientational order in concentrated suspensions of spherical microswimmers,” Phys. Fluids 23, 111702 (2011). 10.1063/1.3660268 [DOI] [Google Scholar]
- 47. Even, C. , Marlière, C. , Ghigo, J.-M. , Allain, J.-M. , Marcellan, A. , and Raspaud, E. , “ Recent advances in studying single bacteria and biofilm mechanics,” Adv. Colloid Interface Sci. 247, 573–588 (2017). 10.1016/j.cis.2017.07.026 [DOI] [PubMed] [Google Scholar]
- 48. Farrell, F. D. C. , Hallatschek, O. , Marenduzzo, D. , and Waclaw, B. , “ Mechanically driven growth of quasi-two-dimensional microbial colonies,” Phys. Rev. Lett. 111, 168101 (2013). 10.1103/PhysRevLett.111.168101 [DOI] [PubMed] [Google Scholar]
- 49. Feng, G. , Cheng, Y. , Wang, S.-Y. , Borca-Tasciuc, D. A. , Worobo, R. W. , and Moraru, C. I. , “ Bacterial attachment and biofilm formation on surfaces are reduced by small-diameter nanoscale pores: How small is small enough?,” npj Biofilms Microbiomes 1, 15022 (2015). 10.1038/npjbiofilms.2015.22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Field, H. A. , Kelley, K. A. , Martell, L. , Goldstein, A. M. , and Serluca, F. C. , “ Analysis of gastrointestinal physiology using a novel intestinal transit assay in zebrafish,” Neurogastroenterol. Motil. 21, 304–312 (2009). 10.1111/j.1365-2982.2008.01234.x [DOI] [PubMed] [Google Scholar]
- 51. Flemming, H. C. , and Wingender, J. , “ The biofilm matrix,” Nat. Rev. Microbiol. 8, 623–633 (2010). 10.1038/nrmicro2415 [DOI] [PubMed] [Google Scholar]
- 52. Frank, D. N. , Amand, A. L. St. , Feldman, R. A. , Boedeker, E. C. , Hapaz, N. , and Pace, N. R. , “ Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases,” Proc. Natl. Acad. Sci. U. S. A. 104, 13780–13785 (2007). 10.1073/pnas.0706625104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Freifeld, L. et al. , “ Expansion microscopy of zebrafish for neuroscience and developmental biology studies,” Proc. Natl. Acad. Sci. U. S. A. 114, E10799–E10808 (2017). 10.1073/pnas.1706281114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Friedlander, R. S. , Vlamakis, H. , Kim, P. , Khan, M. , Kolter, R. , and Aizenberg, J. , “ Bacterial flagella explore microscale hummocks and hollows to increase adhesion,” Proc. Natl. Acad. Sci. U. S. A. 110, 5624–5629 (2013). 10.1073/pnas.1219662110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Frymier, P. D. , Ford, R. M. , Berg, H. C. , and Cummings, P. T. , “ Three-dimensional tracking of motile bacteria near a solid planar surface,” Proc. Natl. Acad. Sci. U.S.A. 92, 6195–6199 (1995). 10.1073/pnas.92.13.6195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Fujishige, N. A. , Kapadia, N. N. , De Hoff, P. L. , and Hirsch, A. M. , “ Investigations of Rhizobium biofilm formation,” FEMS Microbiol. Ecol. 56, 195–206 (2006). 10.1111/j.1574-6941.2005.00044.x [DOI] [PubMed] [Google Scholar]
- 57. Galy, O. , Latour-Lamber, P. , Zrelli, K. , Ghigo, J.-M. , Beloin, C. , and Henry, N. , “ Mapping of bacterial biofilm local mechanics by magnetic microparticle actuation,” Biophys. J. 103, 1400–1408 (2012). 10.1016/j.bpj.2012.07.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Gerlee, P. , and Anderson, A. R. A. , “ Stability analysis of a hybrid cellular automaton model of cell colony growth,” Phys. Rev. E 75, 051911 (2007). 10.1103/PhysRevE.75.051911 [DOI] [PubMed] [Google Scholar]
- 59. Ghosh, P. , Mondal, J. , Ben-Jacob, E. , and Levine, H. , “ Mechanically-driven phase separation in a growing bacterial colony,” Proc. Natl. Acad. Sci. U. S. A. 112, E2166–E2173 (2015). 10.1073/pnas.1504948112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Giacché, D. , Ishikawa, T. , and Yamaguchi, T. , “ Hydrodynamic entrapment of bacteria swimming near a solid surface,” Phys. Rev. E 82, 056309 (2010). 10.1103/PhysRevE.82.056309 [DOI] [PubMed] [Google Scholar]
- 61. Giomi, L. , Liverpool, T. B. , and Marchetti, M. C. , “ Sheared active fluids: Thickening, thinning, and vanishing viscosity,” Phys. Rev. E 81, 051908 (2010). 10.1103/PhysRevE.81.051908 [DOI] [PubMed] [Google Scholar]
- 62. Gordon, V. D. , Davis-Fields, M. , Kovach, K. , and Rodesney, C. A. , “ Biofilms and mechanics: A review of experimental techniques and findings,” J. Phys. D: Appl. Phys. 50, 223002 (2017). 10.1088/1361-6463/aa6b83 [DOI] [Google Scholar]
- 63. Goto, T. , Nakata, K. , Baba, K. , Nishimura, M. , and Magariyama, Y. , “ A fluid-dynamic interpretation of the asymmetric motion of singly flagellated bacteria swimming close to a boundary,” Biophys. J. 89, 3771–3779 (2005). 10.1529/biophysj.105.067553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Goto, T. , Nakai, T. , and Aoki, K. , “ Boundary element analysis on transition of distance and attitude of a bacterium near a rigid surface,” J. Biomech. Sci. Eng. 5, 329–339 (2010). 10.1299/jbse.5.329 [DOI] [Google Scholar]
- 65. Goto, T. , and Nakai, T. , “ Steady distribution of cells in a one-dimensional biased random walk model of bacterial chemotaxis,” J. Biomech. Sci. Eng. 11, 15-00587 (2016). 10.1299/jbse.15-00587 [DOI] [Google Scholar]
- 66. Greenberg, E. P. , and Canale-Parola, E. , “ Motility of flagellated bacteria in viscous environments,” J. Bacteriol. 132, 356–358 (1977). 10.1128/JB.132.1.356-358.1977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Guarner, F. , and Malagelada, J.-R. , “ Gut flora in health and disease,” Lancet 361, 512–519 (2003). 10.1016/S0140-6736(03)12489-0 [DOI] [PubMed] [Google Scholar]
- 68. Haines, B. M. , Sokolov, A. , Aranson, I. S. , Berlyand, L. , and Karpeev, D. A. , “ Three-dimensional model for the effective viscosity of bacterial suspensions,” Phys. Rev. E 80, 041922 (2009). 10.1103/PhysRevE.80.041922 [DOI] [PubMed] [Google Scholar]
- 69. Hall-Stoodley, L. , Costerton, J. W. , and Stoodley, P. , “ Bacterial biofilms: From the natural environment to infectious diseases,” Nat. Rev. Microbiol. 2, 95–108 (2004). 10.1038/nrmicro821 [DOI] [PubMed] [Google Scholar]
- 70. Hanning, I. , and Diaz-Sanchez, S. , “ The functionality of the gastrointestinal microbiome in non-human animals,” Microbiome 3, 51 (2015). 10.1186/s40168-015-0113-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Hatwalne, Y. , Ramaswamy, S. , Rao, M. , and Simha, R. A. , “ Rheology of active-particle suspensions,” Phys. Rev. Lett. 92, 118101 (2004). 10.1103/PhysRevLett.92.118101 [DOI] [PubMed] [Google Scholar]
- 72. Hold, G. L. , and Allen-Vercoe, E. , “ Gut microbial biofilm composition and organization holds the key to CRC,” J. Clin. Invest. 130, 1699–1712 (2019). 10.1038/s41575-019-0148-4 [DOI] [PubMed] [Google Scholar]
- 73. Hill, N. A. , and Pedley, T. J. , “ Bioconvection,” Fluid Dyn. Res. 37, 1–20 (2005). 10.1016/j.fluiddyn.2005.03.002 [DOI] [Google Scholar]
- 74. Hollenbeck, E. C. et al. , “ Mechanical behavior of a Bacillus substillis pellicle,” J. Phys. Chem. B 120, 6080 (2016). 10.1021/acs.jpcb.6b02074 [DOI] [PubMed] [Google Scholar]
- 75. Holmberg, A. , Schwerte, T. , Fritsche, R. , Pelster, B. , and Holmgren, S. , “ Ontogeny of intestinal motility in correlation to neuronal development in zebrafish embryos and larvae,” J. Fish Biol. 63, 318–331 (2003). 10.1046/j.1095-8649.2003.00149.x [DOI] [Google Scholar]
- 76. Horn, H. , and Lackner, S. , “ Modeling of biofilm systems: A review,” Adv. Biochem. Eng. Biotechnol. 146, 53–76 (2014). 10.1007/10_2014_275 [DOI] [PubMed] [Google Scholar]
- 77. Inoue, Y. et al. , “ Torque-speed relationships of Na+ -driven chimeric flagellar motors in Escherichia coli,” J. Mol. Biol. 376, 1251–1259 (2008). 10.1016/j.jmb.2007.12.023 [DOI] [PubMed] [Google Scholar]
- 78. Ishikawa, T. , and Pedley, T. J. , “ The rheology of a semi-dilute suspension of swimming model micro-organisms,” J. Fluid Mech. 588, 399–435 (2007). 10.1017/S0022112007007835 [DOI] [Google Scholar]
- 79. Ishikawa, T. , and Pedley, T. J. , “ Diffusion of swimming model micro-organisms in a semi-dilute suspension,” J. Fluid Mech. 588, 437–462 (2007). 10.1017/S0022112007007847 [DOI] [Google Scholar]
- 80. Ishikawa, T. , Pedley, T. J. , and Yamaguchi, T. , “ Orientational relaxation time of bottom-heavy squirmers in a semi-dilute suspension,” J. Theor. Biol. 249, 296–306 (2007). 10.1016/j.jtbi.2007.07.033 [DOI] [PubMed] [Google Scholar]
- 81. Ishikawa, T. , and Pedley, T. J. , “ Coherent structures in monolayers of swimming particles,” Phys. Rev. Lett. 100, 088103 (2008). 10.1103/PhysRevLett.100.088103 [DOI] [PubMed] [Google Scholar]
- 82. Ishikawa, T. , Locsei, J. T. , and Pedley, T. J. , “ Development of coherent structures in concentrated suspensions of swimming model micro-organisms,” J. Fluid Mech. 615, 401–431 (2008). 10.1017/S0022112008003807 [DOI] [Google Scholar]
- 83. Ishikawa, T. , “ Suspension biomechanics of swimming microbes,” J. R. Soc. Interface 6, 815–834 (2009). 10.1098/rsif.2009.0223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Ishikawa, T. , Yoshida, N. , Ueno, H. , Wiedeman, M. , Imai, Y. , and Yamaguchi, T. , “ Energy transport in a concentrated suspension of bacteria,” Phys. Rev. Lett. 107, 028102 (2011). 10.1103/PhysRevLett.107.028102 [DOI] [PubMed] [Google Scholar]
- 85. Ishikawa, T. , Sato, T. , Mohit, G. , Imai, Y. , and Yamaguchi, T. , “ Transport phenomena of microbial flora in the small intestine with peristalsis,” J. Theor. Biol. 279, 63–73 (2011). 10.1016/j.jtbi.2011.03.026 [DOI] [PubMed] [Google Scholar]
- 86. Ishikawa, T. et al. , “ Separation of motile bacteria using drift velocity in a microchannel,” Lab Chip 14, 1023–1032 (2014). 10.1039/C3LC51302E [DOI] [PubMed] [Google Scholar]
- 87. Ishikawa, T. , “ Vertical dispersion of model microorganisms in horizontal shear flow,” J. Fluid Mech. 705, 98–119 (2012). 10.1017/jfm.2012.116 [DOI] [Google Scholar]
- 88. Ishikawa, T. , “ Swimming of ciliates under geometric constraints,” J. Appl. Phys. 125, 200901 (2019). 10.1063/1.5084764 [DOI] [Google Scholar]
- 89. Jacob, E. B. , Aharonov, Y. , and Shapira, Y. , “ Bacteria harnessing complexity,” Biofilms 1, 239–263 (2004). 10.1017/S1479050505001596 [DOI] [Google Scholar]
- 90. Jang, H. , Rusconi, R. , and Stocker, R. , “ Biofilm disruption by an air bubble reveals heterogeneous age-dependent detachment patterns dictated by initial extracellular matrix distribution,” NPJ Biofilms Microbiomes 3, 6 (2017). 10.1038/s41522-017-0014-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Jemielita, M. et al. , “ Spatial and temporal features of the growth of a bacterial species colonizing the zebrafish gut,” Am. Soc. Microbiol. 5, e01751 (2014). 10.1128/mBio.01751-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Jepson, A. , Martinez, V. A. , Schwarz-Linek, J. , Morozov, A. , and Poon, W. C. K. , “ Enhanced diffusion of nonswimmers in a three-dimensional bath of motile bacteria,” Phys. Rev. E 88, 041002(R) (2013). 10.1103/PhysRevE.88.041002 [DOI] [PubMed] [Google Scholar]
- 93. Kalliomäki, M. , and Isolauri, E. , “ Role of intestinal flora in the development of allergy,” Curr. Opin. Allergy Clini. Immunol. 3, 15–20 (2003). 10.1097/00130832-200302000-00003 [DOI] [PubMed] [Google Scholar]
- 94. Kanehl, P. , and Ishikawa, T. , “ Fluid mechanics of swimming bacteria with multiple flagella,” Phys. Rev. E 89, 042704 (2014). 10.1103/PhysRevE.89.042704 [DOI] [PubMed] [Google Scholar]
- 95. Kikuchi, K. , Noh, H. , Numayama-Tsuruta, K. , and Ishikawa, T. , “ Mechanical roles of anterograde and retrograde intestinal perstalses after feeding in a larval fish (Danio rerio),” Amer. J. Physiol. 318, G1013–G1021 (2020). 10.1152/ajpgi.00165.2019 [DOI] [PubMed] [Google Scholar]
- 96. Kim, M. , Bird, J. C. , Parys, A. J. V. , Breuer, K. S. , and Powers, T. R. , “ A macroscopic scale model of bacterial flagellar bundling,” Proc. Natl. Acad. Sci. U. S. A. 100, 15481–15485 (2003). 10.1073/pnas.2633596100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Kitamura, H. , Omori, T. , and Ishikawa, T. , “ Simulation of a biofilm-formation in a microfluidic channel,” 16th International Conference Fluid Dynamics, Sendai, Japan (2019) [Google Scholar]
- 98. Koch, D. L. , and Subramanian, G. , “ Collective hydrodynamics of swimming microorganisms: Living fluids,” Annu. Rev. Fluid Mech. 43, 637–659 (2011). 10.1146/annurev-fluid-121108-145434 [DOI] [Google Scholar]
- 99. Kostenko, V. , Salek, M. M. , Sattari, P. , and Martinuzzi, R. J. , “ Staphylococcus aureus biofilm formation and tolerance to antibiotics in response to oscillatory shear stresses of physiological levels,” FEMS Immunol. Med. Microbiol. 59, 421–431 (2010). 10.1111/j.1574-695X.2010.00694.x [DOI] [PubMed] [Google Scholar]
- 100. Kyoya, K. , Matsunaga, D. , Imai, Y. , Omori, T. , and Ishikawa, T. , “ Shape matters: Near-field fluid mechanics dominate the collective motions of ellipsoidal squirmers,” Phys. Rev. E 92, 063027 (2015). 10.1103/PhysRevE.92.063027 [DOI] [PubMed] [Google Scholar]
- 101. Lambert, R. A. , Picano, F. , Breugem, W.-P. , and Brandt, L. , “ Active suspensions in thin films: Nutrient uptake and swimmer motion,” J. Fluid Mech. 733, 528–557 (2013). 10.1017/jfm.2013.459 [DOI] [Google Scholar]
- 102. Lauga, E. , “ Bacterial hydrodynamics,” Annu. Rev. Fluid Mech. 48, 105–130 (2016). 10.1146/annurev-fluid-122414-034606 [DOI] [Google Scholar]
- 103. Lauga, E. , DiLuzio, W. R. , Whitesides, G. M. , and Stone, H. , “ Swimming in circles: Motion of bacteria near solid boundaries,” Biophys. J. 90, 400–412 (2006). 10.1529/biophysj.105.069401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Lauga, E. , and Powers, T. R. , “ The hydrodynamics of swimming microorganisms,” Rep. Prog. Phys. 72, 096601 (2009). 10.1088/0034-4885/72/9/096601 [DOI] [Google Scholar]
- 105. Lemelle, L. , Palierne, J.-F. , Chatre, E. , and Place, C. , “ Counterclockwise circular motion of bacteria swimming at the air-liquid interface,” J. Bacteriol. 192, 6307–6308 (2010). 10.1128/JB.00397-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Lieleg, O. , Caldara, M. , Baumgartel, R. , and Ribbeck, K. , “ Mechanical robustness of Pseudomonas aeruginosa biofilms,” Soft Matter 7, 3307–3314 (2011). 10.1039/c0sm01467b [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Liu, J. et al. , “ Coupling between distant biofilms and emergence of nutrient time-sharing,” Science 356, 638–642 (2017). 10.1126/science.aah4204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Liu, Y. , and Tay, J.-H. , “ The essential role of hydrodynamic shear force in the formation of biofilm and granular sludge,” Water Res. 36, 1653–1665 (2002). 10.1016/S0043-1354(01)00379-7 [DOI] [PubMed] [Google Scholar]
- 109. Liu, Y. et al. , “ Examination of the temporal and spatial dynamics of the gut microbiome in newborn piglets reveals distinct microbial communities in six intestinal segments,” Sci. Rep. 9, 3453 (2019). 10.1038/s41598-019-40235-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. López, H. M. , Gachelin, J. , Douarche, C. , Auradou, H. , and Clément, E. , “ Turning bacteria suspensions into superfluids,” Phys. Rev. Lett. 115, 028301 (2015). 10.1103/PhysRevLett.115.028301 [DOI] [PubMed] [Google Scholar]
- 111. Lu, H.-P. , Lai, Y.-C. , Huang, S.-W. , Chen, H.-C. , Hsieh, C.-H. , and Yu, H.-T. , “ Spatial heterogeneity of gut microbiota reveals multiple bacterial communities with distinct characteristics,” Sci. Rep. 4, 6185 (2014). 10.1038/srep06185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Macfarlane, S. , Woodmansey, E. J. , and Macfarlane, G. T. , “ Colonization of mucin by human intestinal bacteria and establishment of biofilm communities in a twostage continuous culture system,” Appl. Environ. Microbiol. 71, 7483–7492 (2005). 10.1128/AEM.71.11.7483-7492.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Marchetti, M. C. , Joanny, J. F. , Ramaswamy, S. , Liverpool, T. B. , Prost, J. , Rao, M. , and Simha, R. A. , “ Hydrodynamics of soft active matter,” Rev. Mod. Phys. 85, 1143–1189 (2013). 10.1103/RevModPhys.85.1143 [DOI] [Google Scholar]
- 114. Marty, A. , Causserand, C. , Roques, C. , and Bacchin, P. , “ Impact of tortuous flow on bacteria streamer development in microfluidic system during filtration,” Biomicrofluidics 8, 014105 (2014). 10.1063/1.4863724 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Matsushita, M. , Hiramatsu, F. , Kobayashi, N. , Ozawa, T. , Yamazaki, Y. , and Matsuyama, T. , “ Colony formation in bacteria: Experiments and modeling,” Biofilms 1, 305–317 (2004). 10.1017/S1479050505001626 [DOI] [Google Scholar]
- 116. Mimura, M. , Sakaguchi, H. , and Matsushita, M. , “ Reaction-diffusion modelling of bacterial colony patterns,” Physica A 282, 283–303 (2000). 10.1016/S0378-4371(00)00085-6 [DOI] [Google Scholar]
- 117. Nilsson, L. M. , Thomas, W. E. , Trintchina, E. , Vogel, V. , and Sokurenko, E. V. , “ Catch bond-mediated adhesion without a shear threshold: Trimannose versus monomannose interactions with the FimH adhesin of Escherichia coli,” J. Biol. Chem. 281, 16656–16663 (2006). 10.1074/jbc.M511496200 [DOI] [PubMed] [Google Scholar]
- 118. Ng, A. N. Y. , De Jong-Curtain, T. A. , Mawdsley, D. J. , White, S. J. , Shin, J. , Appel, B. , Dong, P. D. S. , Stainier, D. Y. R. , and Heath, J. K. , “ Formation of the digestive system in zebrafish: III. Intestinal epithelium morphogenesis,” Dev. Biol. 286, 114–135 (2005). 10.1016/j.ydbio.2005.07.013 [DOI] [PubMed] [Google Scholar]
- 119. Nguyen, D. et al. , “ Active starvation responses mediate antibiotic tolerance in biofilms and nutrient-limited bacteria,” Science 334, 982–986 (2011). 10.1126/science.1211037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Ni, J. , Wum, G. D. , Albenberg, L. , and Tomov, V. T. , “ Gut microbiota and IBD: causation or correlation?,” Nat. Rev. Gastroenterol. Hepatol. 14, 573–584 (2017). 10.1038/nrgastro.2017.88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Or, D. , Smets, B. F. , Wraith, J. M. , Dechesne, A. , and Friedman, S. P. , “ Physical constraints affecting bacterial habitats and activity in unsaturated porous media - a review,” Adv. Water. Resour. 30, 1505–1527 (2007). 10.1016/j.advwatres.2006.05.025 [DOI] [Google Scholar]
- 122. Paramonova, E. , Kalmykowa, O. J. , van der Mei, H. C. , Busscher, H. J. , and Sharma, P. K. , “ Impact of hydrodynamics on oral biofilm strength,” J. Dental Res. 88, 922–926 (2009). 10.1177/0022034509344569 [DOI] [PubMed] [Google Scholar]
- 123. Parthasarathy, R. , “ Monitoring microbial communities using light sheet fluorescence microscopy,” Curr. Opin. Microbiol. 43, 31–37 (2018). 10.1016/j.mib.2017.11.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Pedley, T. J. , and Kessler, J. O. , “ Hydrodynamic phenomena in suspensions of swimming micro-organisms,” Annu. Rev. Fluid Mech. 24, 313–358 (1992). 10.1146/annurev.fl.24.010192.001525 [DOI] [Google Scholar]
- 125. Persat, A. et al. , “ The mechanical world of bacteria,” Cell 161, 988–997 (2015). 10.1016/j.cell.2015.05.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Picioreanu, C. , Kreft, J.-U. , and van Loosdecht, M. C. M. , “ Particle-based multidimensional multispecies biofilm model,” Appl. Environ. Microbiol. 70, 3024–3040 (2004). 10.1128/AEM.70.5.3024-3040.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Potomkin, M. , Gyrya, V. , Aranson, I. , and Berlyand, L. , “ Collision of microswimmers in a viscous fluid,” Phys. Rev. E 87, 053005 (2013). 10.1103/PhysRevE.87.053005 [DOI] [PubMed] [Google Scholar]
- 128. Qi, H. , Lin, W. , Fan, W. , and Qiurong, L. , “ Role of gut microbiota in a zebrafish model with chemically induced enterocolitis involving toll-like receptor signaling pathways,” Zebrafish 11, 255–264 (2014). 10.1089/zeb.2013.0917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Quinlivan, V. H. , and Farber, S. A. , “ Lipid uptake, metabolism, and transport in the larval zebrafish,” Front. Endocrinol. 8, 319 (2017). 10.3389/fendo.2017.00319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Ramaswamy, S. , “ The mechanics and statistics of active matter,” Annu. Rev. Condens. Matter Phys. 1, 323–345 (2010). 10.1146/annurev-conmatphys-070909-104101 [DOI] [Google Scholar]
- 131. Ramia, M. , Tullock, D. L. , and Phan-Thien, N. , “ The role of hydrodynamic interaction in the locomotion of microorganisms,” Biophys. J. 65, 755–778 (1993). 10.1016/S0006-3495(93)81129-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Reigh, S. Y. , Winkler, R. G. , and Gompper, G. , “ Synchronization and bundling of anchored bacterial flagella,” Soft Matter 8, 4363–4372 (2012). 10.1039/c2sm07378a [DOI] [Google Scholar]
- 133. Rittmann, B. E. , “ The effect of shear stress on biofilm loss rate,” Biotechnol. Bioeng. 24, 501–506 (1982). 10.1002/bit.260240219 [DOI] [PubMed] [Google Scholar]
- 134. Rusconi, R. , Lecuyer, S. , Guglielmini, L. , and Stone, H. A. , “ Laminar flow around corners triggers the formation of biofilm streamers,” J. R. Soc. Interface 7, 1293–1299 (2010). 10.1098/rsif.2010.0096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Ryan, S. D. , Haines, B. M. , Berlyand, L. , Ziebert, F. , and Aranson, I. S. , “ Viscosity of bacterial suspensions: Hydrodynamic interactions and self-induced noise,” Phys. Rev. E 83, 050904(R) (2011). 10.1103/PhysRevE.83.050904 [DOI] [PubMed] [Google Scholar]
- 136. Sartor, R. B. , “ Microbial influences in inflammatory bowel diseases,” Gastroenterology 134, 577–594 (2008). 10.1053/j.gastro.2007.11.059 [DOI] [PubMed] [Google Scholar]
- 137. Schlomann, B. H. , Wiles, T. J. , Wall, E. S. , Guillemin, K. , and Parthasarathy, R. , “ Bacterial Cohesion predicts spatial distribution in the larval zebrafish intestine,” Biophys. J. 115, 2271–2277 (2018). 10.1016/j.bpj.2018.10.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Schneider, W. , and Doetsch, R. N. , “ Effect of viscosity on bacterial motility,” J. Bacteriol. 117, 696–701 (1974). 10.1128/JB.117.2.696-701.1974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Sheth, R. U. , Li, M. , Jiang, W. , Sims, P. A. , Leong, K. W. , and Wang, H. H. , “ Spatial metagenomic characterization of microbial biogeography in the gut,” Nat. Biotechnol. 37, 877–883 (2019). 10.1038/s41587-019-0183-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Shimogonya, Y. , “ Torque-induced precession of bacterial flagella,” Sci. Rep. 5, 18488 (2015). 10.1038/srep18488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Shrout, J. D. , Chopp, D. L. , Just, C. L. , Hentzer, M. , Givskov, M. , and Parsek, M. R. , “ The impact of quorum sensing and swarming motility on Pseudomonas aeruginosa biofilm formation is nutritionally conditional,” Mol. Microbiol. 62, 1264–1277 (2006). 10.1111/j.1365-2958.2006.05421.x [DOI] [PubMed] [Google Scholar]
- 142. Simoes, M. , Sillankorva, S. , Pereira, M. O. , Azeredo, J. , and Vieira, M. J. , “ The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms,” Biofouling 23, 249–258 (2007). 10.1080/08927010701368476 [DOI] [PubMed] [Google Scholar]
- 143. Sokolov, A. , Goldstein, R. E. , Feldchtein, F. I. , and Aranson, I. S. , “ Enhanced mixing and spatial instability in concentrated bacterial suspensions,” Phys. Rev. E 80, 031903 (2009). 10.1103/PhysRevE.80.031903 [DOI] [PubMed] [Google Scholar]
- 144. Song, H.-S. , Cannon, W. R. , Beliaev, A. S. , and Konopka, A. , “ Mathematical modeling of microbial community dynamics: A methodological review,” Processes 2, 711–752 (2014). 10.3390/pr2040711 [DOI] [Google Scholar]
- 145. Sowa, Y. , Hotta, H. , Homma, M. , and Ishijima, A. , “ Torque-speed relationship of the Na+ -driven flagellar motor of Vibrio alginolyticus,” J. Mol. Biol. 327, 1043–1051 (2003). 10.1016/S0022-2836(03)00176-1 [DOI] [PubMed] [Google Scholar]
- 146. Srivastava, A. , Gupta, J. , Kumar, S. , and Kumar, A. , “ Gut biofilm forming bacteria in inflammatory bowel disease,” Microb. Pathog. 112, 5–14 (2017). 10.1016/j.micpath.2017.09.041 [DOI] [PubMed] [Google Scholar]
- 147. Stephens, W. , Burns, A. , Stagaman, K. et al. , “ The composition of the zebrafish intestinal microbial community varies across development,” ISME J. 10, 644–654 (2016). 10.1038/ismej.2015.140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Stewart, P. S. , “ Diffusion in biofilms,” J. Bacteriol. 185, 1485–1491 (2003). 10.1128/JB.185.5.1485-1491.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Suzuki, T. A. , and Nachman, M. W. , “ Spatial heterogeneity of gut microbial composition along the gastrointestinal tract in natural populations of house mice,” PLoS One 11, e0163720 (2016). 10.1371/journal.pone.0163720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Swidsinski, A. , Weber, J. , Loening-Baucke, V. , Hale, L. P. , and Lochs, H. , “ Spatial organization and composition of the mucosal flora in patients with inflammatory bowel disease,” J. Clin. Microbiol. 43, 3380–3389 (2005). 10.1128/JCM.43.7.3380-3389.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Taormina, M. J. , Hay, E. A. , and Parthasarathy, R. , “ Passive and active microrheology of the intestinal fluid of the larval zebrafish,” Biophys. J. 113, 957–965 (2017). 10.1016/j.bpj.2017.06.069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Takahashi, T. , and Sakaguchi, E. , “ Role of the furrow of the proximal colon in the production of soft and hard feces in nutrias, Myocastor coypus,” J. Comp. Physiol. B 170, 531–535 (2000). 10.1007/s003600000131 [DOI] [PubMed] [Google Scholar]
- 153. Takahashi, T. , and Sakaguchi, E. , “ Transport of bacteria across and along the large intestinal lumen of guinea pigs,” J. Comp. Physiol. B 176, 173–178 (2006). 10.1007/s00360-005-0039-6 [DOI] [PubMed] [Google Scholar]
- 154. Tamargo, A. , Cueva, C. , Alvarez, M. D. , Herranz, B. , Bartolome, B. , Moreno-Arribas, M. V. , and Laguna, L. , “ Influence of viscosity on the growth of human gut microbiota,” Food Hydrocolloids 77, 163–167 (2018). 10.1016/j.foodhyd.2017.09.031 [DOI] [Google Scholar]
- 155. Taylor, S. W. , and Jaffe, P. R. , “ Biofilm growth and the related changes in the physical properties of a porous medium 3. dispersivity and model verification,” Water Resour. Res. 26, 2171–2180, 10.1029/WR026i009p02171 (1990). [DOI] [Google Scholar]
- 156. Thomas W., “ Catch bonds in adhesion,” Annu. Rev. Biomed. Eng. 10, 39–57 (2008). 10.1146/annurev.bioeng.10.061807.160427 [DOI] [PubMed] [Google Scholar]
- 157. Thursby, E. , and Juge, N. , “ Introduction to the human gut microbiota,” Biochem. J. 474, 1823–1836 (2017). 10.1042/BCJ20160510 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Tropini, C. , Earle, K. A. , Huang, K. C. , and Sonnenburg, J. L. , “ The gut microbiome: Connecting spatial organization to function,” Cell Host Microbe 21, 433–442 (2017). 10.1016/j.chom.2017.03.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Verwey, E. J. W. , and Overbeek, J. T. G. , Theory of the Stability of Lyophobic Colloids ( Elsevier, Amsterdam, 1948). [DOI] [PubMed] [Google Scholar]
- 160. Vieira, M. J. , Melo, L. , and Pinheiro, M. M. , “ Biofilm formation: Hydrodynamic effects on internal diffusion and structure,” Biofouling 7, 67–80 (1993). 10.1080/08927019309386244 [DOI] [Google Scholar]
- 161. Vigeant, M. A. S. , and Ford, R. M. , “ Interactions between motile Escherichia coli and glass in media with various ionic strengths, as observed with a three-dimensional-tracking microscope,” Appl. Environ. Microbiol. 63, 3474–3479 (1997). 10.1128/AEM.63.9.3474-3479.1997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Vigeant, M. A. S. , Ford, R. M. , Wagner, M. , and Tamm, L. K. , “ Reversible and irreversible adhesion of motile Escherichia coli cells analyzed by total internal reflection aqueous fluorescence microscopy,” Appl. Environ. Microbiol. 68, 2794–2801 (2002). 10.1128/AEM.68.6.2794-2801.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Volpe, G. , Buttinoni, I. , Vogt, D. , Kummerer, H.-J. , and Bechinger, C. , “ Microswimmers in patterned environments,” Soft Matter 7, 8810–8815 (2011). 10.1039/c1sm05960b [DOI] [Google Scholar]
- 164. Wakita, J. , Itoh, H. , Matsuyama, T. , and Matsushita, M. , “ Self-affinity for the growing interface of bacterial colonies,” J. Phys. Soc. Jpn. 66, 67–72 (1997). 10.1143/JPSJ.66.67 [DOI] [Google Scholar]
- 165. Watari, N. , and Larson, G. , “ The hydrodynamics of a run-and-tumble bacterium propelled by polymorphic helical flagella,” Biophys. J. 98, 12–17 (2010). 10.1016/j.bpj.2009.09.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Wensink, H. H. , Dunkel, J. , Heidenreich, S. , Drescher, K. , Goldstein, R. E. , Lowena, H. , and Yeomans, J. M. , Proc. Natl. Acad. Sci. U. S. A. 109, 14308–14313 (2012). 10.1073/pnas.1202032109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Wiles, T. J. et al. , “ Host gut motility promotes competitive exclusion within a model intestinal microbiota,” PLoS Biol. 14, e1002517 (2016). 10.1371/journal.pbio.1002517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168. Wiles, T. J. , Schlomann, B. H. , Wall, E. S. , Betancourt, R. , Parthasarathy, R. , and Guillemin, K. , “ Swimming motility of a gut bacterial symbiont promotes resistance to intestinal expulsion and enhances inflammation,” PLoS Biol. 18, e3000661 (2020). 10.1371/journal.pbio.3000661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Wilking, J. N. , Angelini, T. E. , Seminara, A. , Brenner, M. P. , and Weitz, D. A. , “ Biofilms as complex fluids,” MRS Bull. 36, 385–391 (2011). 10.1557/mrs.2011.71 [DOI] [Google Scholar]
- 170. Winstanley, H. F. , Chapwanya, M. , McGuinness, M. J. , and Fowler, A. C. , “ A polymer-solvent model of biofilm growth,” Proc. R. Soc. A 467, 1449–1467 (2011). 10.1098/rspa.2010.0327 [DOI] [Google Scholar]
- 171. Wu, X.-L. , and Libchaber, A. , “ Particle diffusion in a quasi-two-dimensional bacterial bath,” Phys. Rev. Lett. 84, 3017 (2000). 10.1103/PhysRevLett.84.3017 [DOI] [PubMed] [Google Scholar]
- 172. Xavier, J. B. , Picioreanu, C. , and van Loosdecht, M. C. M. , “ A framework for multidimensional modeling of activity and structure of multispecies biofilms,” Environ. Microbiol. 7, 1085–1103 (2005). 10.1111/j.1462-2920.2005.00787.x [DOI] [PubMed] [Google Scholar]
- 173. Yang, J. , Shimogonya, Y. , and Ishikawa, T. , “ What causes the spatial heterogeneity of bacterial flora in the intestine of zebrafish larvae?,” J. Theor. Biol. 446, 101–109 (2018). 10.1016/j.jtbi.2018.03.007 [DOI] [PubMed] [Google Scholar]
- 174. Yang, J. , Shimogonya, Y. , and Ishikawa, T. , “ Bacterial detachment from a wall with a bump line,” Phys. Rev. E 99, 023104 (2019). 10.1103/PhysRevE.99.023104 [DOI] [PubMed] [Google Scholar]
- 175. Yao, J. , and Allen, C. , “ Chemotaxis is required for virulence and competitive fitness of the bacterial wilt pathogen Ralstonia solanacearum,” J. Bacteriol. 188, 3697–3708 (2006). 10.1128/JB.188.10.3697-3708.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176. Young, I. M. , and Crawford, J. W. , “ Interactions and self-organization in the soil-microbe complex,” Science 304, 1634–1637 (2004). 10.1126/science.1097394 [DOI] [PubMed] [Google Scholar]
- 177. Zang, L. , Shimada, Y. , and Nishimura, N. , “ Development of a novel zebrafish model for type 2 diabetes mellitus,” Sci. Rep. 7, 1461 (2017). 10.1038/s41598-017-01432-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178. Zhang, T. , Cogan, N. , and Wang, Q. , “ Phase-field models for biofilms II. 2D numerical simulations of biofilm-flow interaction,” Commun. Comput. Phys. 4, 72–101 (2008). [Google Scholar]
- 179. Zottl, A. , and Stark, H. , “ Hydrodynamics determines collective motion and phase behavior of active colloids in quasi-two-dimensional confinement,” Phys. Rev. Lett. 112, 118101 (2014). 10.1103/PhysRevLett.112.018101 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.