

Abstract
The current article served to provide the most up-to-date information regarding the causes of keel bone fracture. Although elevated and sustained egg production is likely a major contributing factor toward fractures, new information resulting from the development of novel methodologies suggests complementary causes that should be investigated. We identified 4 broad areas that could explain variation and increased fractures independent of or complementing elevated and sustained egg production: the age at first egg, late ossification of the keel, predisposing bone diseases, and inactivity leading to poor bone health. We also specified several topics that future research should target, which include continued efforts to link egg production and bone health, examination of noncommercial aves and traditional breeds, manipulating of age at first egg, a detailed histological and structural analysis of the keel, assessment of prefracture bone condition, and the relationship between individual activity patterns and bone health.
Key words: pullet, bone, disease, fracture, damage
Introduction
The extremely high frequency and severity of damage to the keel bone in commercial laying hen housing systems represents one of the greatest welfare problems facing the egg production industry as suggested by the United Kingdom's Farm Animal Welfare Committee (FAWC, 2010, FAWC, 2013). Keel bone damage is a more general term that can be subdivided into 1) fractures (keel bone fractures [KBF]), defined as breaks in the bone that typically manifest as a callus around the fracture site after an unknown time, and 2) keel bone deviations, characterized by sharp, unnatural bending from a theoretical, two-dimensional plane (Casey-Trott et al., 2015). The current article focuses primarily on the former. The KBF issue has also been cited as a major problem by the European Food Safety Authority-Panel on Animal Health and Welfare (Welfare, 2015) and a North American–based consortium of welfare researchers (Lay et al., 2011). The pain believed to be associated with KBF (Nasr et al., 2012b) is a primary concern owing to gross anatomical disfigurement, although the likely economic losses (Thiruvenkadan et al., 2010) resulting from reduced egg production (Nasr et al., 2012a; Rufener et al., 2016) are also relevant, with far-ranging effects on the sustainability of production. Unfortunately, despite efforts to increase standards of on-farm animal welfare in general, assessments by a variety of research groups place the percentage of birds with fractures at an alarmingly high rate. The number of affected birds within commercial flocks can range between 20 and 96%, based on reports from various countries including Belgium (Heerkens et al., 2015), Canada (Petrik et al., 2015), Denmark (Riber and Hinrichsen, 2016), The Netherlands (Rodenburg et al., 2008), Switzerland (Käppeli et al., 2011; Stratmann et al., 2015b,a), and the United Kingdom (Wilkins et al., 2011; Tarlton et al., 2013; Toscano et al., 2015). The problem has also been observed in commercial broiler breeder flocks (Gebhardt-Henrich et al., 2017).
The traditional explanation for the cause of fractures is that high calcium demands of contemporary commercial hens required for egg production (approximately 320 eggs over a 365-day period, i.e., nearly an egg per day) induce resorption or the breakdown of the bone matrix and release of contained mineral. The entire process is believed to leave bones weak and brittle (Whitehead and Fleming, 2000; Fleming et al., 2004; Bain et al., 2016). Although not a cause of fractures by itself, the resulting poor bone health leaves the hen relatively susceptible to fractures from more direct causes such as collisions with housing objects (Figure 1). Hazardous movements, such as collisions, being a contributory factor to KBF are supported by correlations between increased KBF frequency and housing features including greater height and the number of perchable items (Wilkins et al., 2011), aviary arrangement within the barn (i.e., portal vs. row units), wire mesh flooring vs. plastic (Heerkens et al., 2015), and the absence of ramps connecting upper and lower aviary tiers (Stratmann et al., 2015a). Although less studied, high-energy collision events have also been reported in caged housing (Baker et al., 2020) and thus could explain the reported occurrence, albeit reduced in frequency and severity relative to noncage systems (Sherwin et al., 2010; Wilkins et al., 2011; Petrik et al., 2015; Casey-Trott and Widowski, 2016).
Figure 1.

Potential explanations for the development of keel bone fractures.
Although increased egg production may well be a major causal factor in predisposing birds to fracture, recent methodological advances have allowed for an improved understanding of KBF and suggest that factors other than high egg production may also have a causative role. For example, methodological advances include improved assessment methods using radiography (L. Candelotto, unpublished data; Eusemann et al., 2018a; Tracy et al., 2019; Baur et al., 2020), sonography (Tracy et al., 2019), computed tomography (L. Candelotto, unpublished work; Regmi et al., 2013, 2015; Chargo et al., 2018a,b); and risk analysis (Toscano et al., 2013, 2018; Candelotto et al., 2017) and use of experimental models that controlled for potentially confounding factors (Regmi et al., 2013; Casey-Trott, 2016; Candelotto et al., 2017; Casey-Trott et al., 2017a,b; Eusemann et al., 2018a).
Further adding to confusion over the role of high egg production as a cause of fracture, deformities have been reported much earlier than the emergence of modern poultry genetics and simultaneous increase in egg production. Observations reported by Darwin (1868) address issues of keel bone quality, wherein 12 breeds selected for observation had so many incidences of deformation such that only 3 breeds could be used for comparison. Although observations were based primarily on large, male specimens, it was reported that up to 88% of chickens examined across the 12 breeds had a deformed sternum. Examination of more recent information, but preceding the application of modern genetic selection programs in poultry, gives further evidence of keel bone deformities. From investigations on what was termed crooked keels of both sexes, Warren (1937) attributed genetics as well as effects of perches and deficient vitamin D concentrations as a causal factor. These and other early studies intriguingly seem to suggest both males and females are equally prone to keel bone damage, apparently in contrast to the modern situation wherein males do not seem to be afflicted. Alternatively, the absence of males without keel bone damage could be due to their presence in breeding populations and not in the production systems, which are predominantly examined for KBF. It is important to note that the types of damage reported in these earlier studies may be a combination of KBF and keel bone deviations. The latter is different in characteristics and likely a cause of KBF with implications discussed in the following section. Although abnormalities predating modern poultry genetics and the current phenomena of keel damage undoubtedly arose from different causes, the existence of keel damage in hens without high and sustained egg production is a reminder that other factors may play a role.
Our understanding of KBF has also improved with the important step of defining the damage in question, which will help to better understand the causes of KBF. Although simplistic in nature, the definition of fracture used in published works has been confusing and inconsistent (reviewed in the study by Harlander-Matauschek et al., 2015). The majority of the published research studies before 2015 used palpation of live birds as a means to detect fracture, wherein the presence of a callus formation was used as an identifying feature. The classification may or may not have included unusual bends in the bone, often referred to as keel bone deviations in an overall category of damage. From a morphological perspective, a fracture would be defined by one or more fracture lines clearly separating a bone into 2 or more separated fragments, a definition that would exclude all examples provided by Wilkins et al. (2011), a commonly cited article used when referencing the palpation technique. Clarity in terms of the definition of KBF and keel bone deviations (Casey-Trott et al., 2015), a visual radiograph evaluation protocol based on those definitions (Rufener et al., 2018a), and improved palpation training (Casey-Trott et al., 2015; Petrik et al., 2015; Buijs et al., 2018; Gebhardt-Henrich et al., 2019; Tracy et al., 2019) are 3 examples of how advances in definitions and assessment have led to an improved understanding of keel bone damage and its causes.
Given the benefits that improved methodologies and the consequent clarification have afforded, combined with the renewed perspective of keel bone damage occurring before modern breeding programs, a revised examination of the causative mechanisms of KBD is warranted. The current article reviews the existing information linking egg production and fractures, including recent evidence suggesting the need to consider other explanations in addition to egg production and then explores several additional explanations that could also lead to poor bone health. The additional explanations included in this article are 1) inappropriate duration of rearing until the hen's first egg, 2) poor ossification at particular keel locations, 3) predisposing bone diseases, and 4) reduced activity leading to a musculoskeletal system poorly suited to current housing (Figure 1). We then conclude with suggestions for future research.
We have purposely not included a specific nutrition-themed explanation as we believe there is insufficient recent information to support a specific alternative explanation. Deficiencies in diet have long been recognized as a problem for bone quality in laying hens, a subject that has been reviewed on numerous occasions (Norman and Hurwitz, 1993; Whitehead, 2004a; Fleming, 2008; Bain et al., 2016; Olgun and Aygun, 2016; Świątkiewicz et al., 2017). Sufficient supply of calcium at lighting up rather than at first egg and feeding the calcium source as a particulate (Fleming et al., 2003, 2006) to ensure delayed absorption during the night period when shell production is occurring is known to be beneficial. If hens are fed to the recommended dietary quantities of calcium and phosphorous with adequate supply of vitamin D, further supplementation has limited effect (Rennie et al., 1997). In practice, there is still the possibility of poor diet formulation, differences in the digestibility of the calcium source (Guinotte et al., 1995; Anwar et al., 2016), or malabsorption of nutrients owing to factors such as poor gut health (Freitas, 2014), although there seems to be little investigation of laying hens in this regard.
High egg production as a cause for weakened and brittle bone
The bones of laying hens are composed of 3 primary types—cortical, trabecular or cancellous, and medullary (Figure 2). Medullary bone is a special type of woven bone unique to female aves that serves principally as a labile source of calcium, quickly providing the mineral for one eggshell and then absorbing and storing calcium for the next egg (Dacke et al., 1993). Medullary bone is only present when there is high estrogen concentration in the plasma, produced by the developing follicles of the ovary before commencement of egg production, that is, only in lay. In modern commercial hybrids, wherein egg production is sustained over a longer time than in ancestral breeds such as the red jungle fowl, the hen may extract supplemental calcium in a process called resorption from other types of bone including cortical bone. Unlike medullary bone, cortical bone primarily has the function of structural support and is produced throughout the rearing period until the onset of sexual maturity and egg production at approximately 16 wk of age after which it is no longer produced (Hudson et al., 1993) until exiting lay. During the laying period, bone resorption is used to supplement exogenous (dietary) calcium required for eggshell formation. As bone resorption continues throughout the lay cycle (∼50–70 wk in modern flocks) in which eggs are produced at a rate in excess of 9 every 10 D, hens manifest a net loss of mineral in structural (cortical) bone from a peak at sexual maturity (Wilson et al., 1992). It is this gradual but sustained loss of structural cortical bone that is thought to be responsible for the increased frequency of KBF as the hen attempts to support nearly continuous eggshell formation with diminishing endogenous mineral reserves.
Figure 2.

Avian bone can be subdivided into cortical, cancellous, and medullary bone. (A and C) Chicken, tibia, diaphysis: Cortical (synonym: compact) bone is especially prominent in the diaphyses of the long bones of the appendicular skeleton. (B and D) Chicken, femur, distal epiphysis: Cancellous (synonyms: trabecular or spongy) bone is the predominant type of bone in the core of the metaphyses and epiphyses. (A and B) Chicken, 14 wk old: The bone marrow is free of medullary bone before the onset of egg laying. (C and D) Chicken, 32 wk old: Medullary bone builds up within the medullary cavity along the inner border of both cortical (C) and trabecular bone (D) shortly before the onset of egg laying. Hematoxylin and eosin; bars = 200 μm. (Credit: R Ulrich.)
Supporting a direct role of the presence or absence of egg production in development of KBF, Eusemann et al. (2018b) found laying hens whose egg production was abolished by a synthetic agonist of the gonadotropin-releasing hormone (i.e., deslorelin acetate) between 24 to 35 wk of age had no fractures compared with 40% in the control hens. Comparably, using 2 groups of hens selected over 7 generations for a high and low bone index (a composite of strength and mineral density without gains in body mass and described in the study by Bishop et al., 2000), Fleming et al (2004) then assessed bone health in the keel and long bones from males and females at 68 wk of age. Males were found to be free of keel bone damage, whereas the percentage of normal keels in females ranged from 60.3% in the low bone quality line to 100% in the high bone quality line. No males were recorded as having any type of fracture, whereas only the last generation of females with high bone quality lacked evidence of fracture. Unfortunately, although the authors reported evidence of fracture in long bones, they used the term deformities with several categories (i.e., normal, twisted, severe) to describe the keel; hence, it is difficult to ascertain the type of damage that was observed. More recently, Eusemann et al. (2018a) found reduced occurrence of fractures in a low-producing, brown laying line compared with a high-producing, brown line, although the results are confounded by hybrid-specific factors.
Surprisingly, the worst fractures were in a low-producing white line. Similarly, on comparison of a relatively less productive heritage line (Barred Plymouth Rock) with high-producing commercial lines (Hy-Line Brown, Hy-Line Silver), the latter were found to have increased rates of fresh fractures despite comparatively greater bone density (Regmi et al., 2016a). Taken together, these findings highlight the need for direct observation and experimental manipulation of relevant factors to determine the causes of KBF. The authors know of only one article assessing fractures in broiler breeders, in which fractures were present in a maximum of 54.7% of birds when housed with perches (Gebhardt-Henrich et al., 2018), although only limited behavioral assessments were carried out without histological or biomechanical measures.
Evidence of alternative causal factors
To understand of the factors that influence KBF, Toscano et al. (2013, 2018) developed a methodology that uses an impact testing device to create collisions in a controlled, quantifiable manner using recently euthanized hens. The methodology resolved some challenges in linking factors with fracture in that collisions could be viewed essentially in “real time,” i.e., within seconds of collision, and thus, relevant properties could be evaluated at the time of collision and not at a subsequent period after bone began the healing process. By statistically modeling various factors and their relationship to the outcome of whether a fracture occurred and its severity at specific impact energies, the magnitude of change as reflected by the increase or decrease in the factor could be related to fracture susceptibility. The methodology is not examining fractures per se, but modeling the likelihood of their occurrence in relation to key factors. The impact testing protocol was used to relate susceptibility to KBF across 5 time points and then quantify a range of hen-level factors relating to bone health (e.g., bone mineral density) (Toscano et al., 2018). Unexpectedly, the effort identified a post–peak egg production phase, in which, beyond 49 wk of age, susceptibility to new fractures appeared to decrease. The finding was particularly surprising given that hens' daily egg production will remain relatively high with values >85% and that hens are not believed to increase net cortical bone mass while they remain in lay (Hudson et al., 1993). Nonetheless, the findings of Toscano et al. (2018) appear to parallel age-related changes in KBF frequencies reported independently in commercial flocks (Stratmann et al., 2015a; Petrik et al., 2015; Toscano et al., 2015), suggesting the result observed with the impact testing system is not an aberration of the experimental procedure. Previously, the decrease in new fractures could be attributed to altered behavior or improved navigation abilities, although the impact testing data, in combination with mirrored patterns in commercial flocks, suggest some qualitative change within the animal such as bone maturation in combination with or age-related changes in strength (Rath et al., 2000). In other words, if egg production was directly related to bone health and occurrence of fractures, we would expect rates of fracture to continue increasing beyond 49 wk of age, although this is not the case. Rather, the rate of new fractures appears to level off or possibly even decrease in both commercial and experimental settings.
The impact testing system has also provided insight into the relationship between egg production and fracture development of individual hens. In an examination of fracture susceptibility within 5 distinct genetic laying hen lines, Candelotto et al. (2017) reported that increased shell breaking strength and width from eggs of individual hens was associated with reduced fracture susceptibility. Although the authors anticipated the opposite, that is, hens with stronger and thinner eggshells would be more likely to develop fractures as they would be contributing a greater proportion of their available calcium pool toward egg production at a detriment to bone health, the occurrence of the unexpected requires consideration of alternatives. For instance, some hens may have superior calcium absorption, leading to a comprehensive benefit for all tissues—both eggs and bone.
Taken together, these recent findings suggest that although high and sustained egg production does appear to play a major causative role in KBF, multiple other factors have to be considered, especially when some low-producing lines have poorer bone quality than their high-producing counterparts. The problem appears to predate modern production genetics and is not in agreement with expected predictions, according to which KBF frequency is the sole causative factor linked to egg production. Understanding the interaction and relative importance of the factors involved in the pathogenesis of KBF is essential if effective solutions that eliminate or reduce KBF frequency and severity are to be developed and animal welfare is to be optimized.
Additional explanations for KBF susceptibility
Age at First Egg
If the act of egg production and the concomitant adaptations are related to the problem of poor bone quality in laying hens, we should look at other aspects of reproduction. Other than the intensity of egg production, puberty is a parameter with a clear genetic determination and a good candidate for affecting bone quality. Puberty is most usually measured as age at first egg (AFE) in poultry (Wolc et al., 2010) and has been a long-standing factor in breeding programs (Abplanalp, 1957). The AFE trait is influenced by many factors, most notably growth rate and body weight, with which it often shows correlation and overlapping quantitative trait locus (Soller et al., 1984; Kerr et al., 2001; Podisi et al., 2011), although not in every instance (Wolc et al., 2010). Photoperiod (Lewis et al., 2008) and its interaction with diet to influence the growth rate (Dunn and Sharp, 1992) are also contributory. If the shift to an egg laying physiology was to occur early, the transition to development of medullary bone and the calcium requirement for egg formation might prevent or delay full ossification. The effect would be especially relevant in the keel, where ossification occurs relatively late (Buckner et al., 1949). Given the existing knowledge of egg laying physiology and the associated changes in bone development, we need to consider evidence that AFE might affect bone quality.
A comparison by Hocking et al. (2003) of bone quality for 13 different traditional and 12 different commercial breeds at 55 wk of age found the former had considerably better bone quality and a much later onset of sexual maturity by an average of 4 wk. The authors suggested that the modern problems of bone breakage in commercial laying hens were a combination of coming into lay earlier and the continuation of high rates of lay for extended periods of time. Ultimately, however, this study was only correlative. Data for individual breeds' bone quality were not available, and many of the traditional breeds were considerably heavier than the commercial layer breeds, which would positively influence bone strength. A similar study examining bone characteristics before egg laying concluded that intensive selection for elevated egg laying has not changed bone size, shape, or quality before egg laying commenced (Hocking et al., 2009). The result strongly suggested that any effect of selection in egg-laying breeds as seen in an earlier, related study (Hocking et al., 2003) would be manifested after egg laying commenced and could include AFE or persistence.
Silversides et al. (2006) directly tested the hypothesis that delaying sexual maturity using photoperiod would allow greater skeletal maturity before egg formation commenced by using 3 chicken breeds, 2 commercial breeds, and one traditional breed. The paradigm used featured a 2-week difference in age at photostimulation although it delayed AFE by only 4 D. The relatively minor delay illustrates the limitations of photoperiodic manipulation as a solution for improving bone health in modern laying hens that will come into lay even on short-day photoperiods (Morris et al., 1995). Delaying photostimulation resulted in positive results overall, primarily on the density and area of medullary and cortical bone of the radius and humerus, although bone breaking strength was unaffected. The inconsistent results with regard to photostimulation were in contrast to the much clearer effect of the breed, wherein evidence suggested that the humerus may have been weakened in breeds with higher egg production. While the findings were interesting, large differences in body weight make interpretations difficult. In another experiment that also approached the problem directly by using photoperiod to try to delay sexual maturity, the authors found little effect on bone mineralization at the end of lay (Hester et al., 2011). Unfortunately, it was not possible from the publication to determine if a delay in the onset of lay was actually achieved with the photoperiods used, and as the authors conceded, there was no measurement of mineralization earlier in lay or fractures. When the onset of lay was monitored in housing that allowed linking of individual eggs and hens (Gebhardt-Henrich and Fröhlich, 2015), incidents of keel bone damage, as determined by repeated palpation until 43 wk of age, were relatively frequent and associated with AFE, but not with total egg production. Specifically, hens without keel damage came into lay approximately 17 D later than those having some form of keel damage. In contrast, a study looking at bone quality at 70 wk of age, with a wide range of factors, which included photoperiodic manipulation that resulted in a 4-day AFE delay, found bone quality of the humerus and keel radiographic density increased in the early-maturing hens (Fleming et al., 2003), although differences were not observed at any other ages. The changes in bone properties observed may have been linked to increased medullary bone in these structures, perhaps owing to the earlier photostimulation, although the effects were not evident at younger ages. A lack of an observable effect with AFE manipulation by a combination of light and diet was also evident on bone breakage incidence at slaughter at 82 wk of age (Gregory et al., 1990). It may be relevant that hens of these latter 2 studies were kept in battery cages.
To summarize, there are clear genetic effects on bone quality, which can be supported by a number of direct studies in addition to between-breed studies already mentioned (Bishop et al., 2000; Dunn et al., 2007; Podisi et al., 2012; Eusemann et al., 2018a; Raymond et al., 2018). There are some studies that seem to corroborate the role of AFE in bone quality, but it must also be recognized that a number of studies do not show effects. It may well be that effects are dependent on an interaction of genetics and the environment, with more challenging environments being necessary for the full expression of the trait. In other words, genetic parameters and correlations between KBF and other traits such as AFE can only be detected in environments that allow damage to occur at high frequencies and severities, for example, aviary housing. It may also be that some studies lack sufficient power to detect effects that are highly variable by nature. These obstacles, combined with difficulties manipulating the onset of puberty in modern laying hybrids, may have hampered attempts to detect effects of the age at puberty on KBF.
Finally, the keel bone may be more susceptible to the effect of early puberty than other bones. The keel ossifies more slowly in comparison with other parts of the sternum, and much of the keel is still cartilaginous when puberty is reached (Buckner et al., 1948, 1949). The ossification delay may make it more vulnerable to fracture as the physiology of the birds shift to egg laying, resulting in poor calcification of the structure.
Late Ossification of the Full Keel
Although the AFE-related discussion proposes that the keel has not fully developed before the physiological transition to egg laying occurs, resulting in a compromised structure, an extension of this discussion is that the combination of AFE and late ossification of certain areas predisposes those areas to fracture. Late ossification does not provide an explanation for the occurrence of fractures per se but may explain the previously discussed variation in fracture rates and thus aid in identifying true causes of fracture. Bone development is a complex and continuous process through life, including bone formation, bone repair, and bone remodeling. Bone formation occurs through ossification, a process of bone growth in which connective tissue or cartilage is replaced with bone tissue (Bittner et al., 1998; Whitehead, 2004b). Within the keel, ossification begins cranially and progresses to the caudal tip (Buckner et al., 1949). The ossification of the caudal part of the keel is likely not completed until 30 to 40 wk of age based on the historical literature of laying hens from the 1940s (Buckner et al., 1948, 1949), although an examination of tibias in male broiler breeders supports the time frame remains relevant (Rath et al., 2000). To the best of the authors' knowledge, no evidence of longitudinal observations on keel development and bone properties exist. The immature bone in an ossification zone is weaker than mature bone structures and may therefore be more at risk of fractures (Birchard and Sherding, 2005). The elevated rate of fractures seen in the period before 35 wk (Baur et al., 2020) followed by the subsequent decrease could be explained in part by the entire keel being late to complete the ossification process. The cartilaginous composition of the keel and the ongoing ossification during peak production would result in a keel that is weaker than that of older hens and may explain why KBF often are found at the caudal part of the bone (Figure 3), which is the last part to be ossified (Casey-Trott et al., 2017a; Thøfner et al., 2020).
Figure 3.
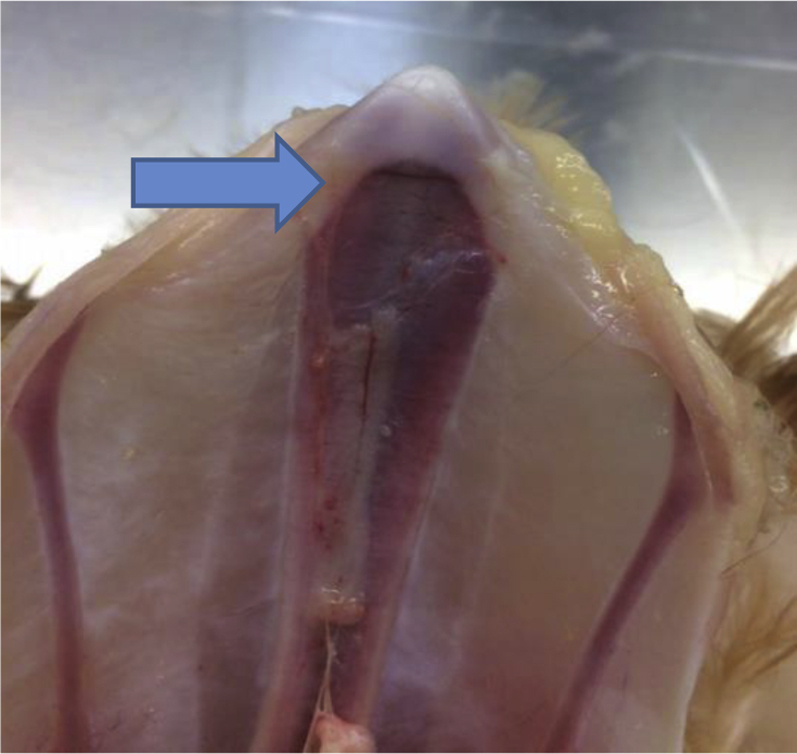
Ventral image of a 20-week-old laying hen, wherein a small fracture can be seen immediately caudal to the line of ossification and indicated by the arrow. (Credit: JP Christensen.)
It could also be speculated that high and sustained egg laying in a period in which the keel bone structure is weak creates excessive strain on the ossification zones, leading to fractures in the immature zones of the keel. Ossification might also be disturbed by the competing demand for calcium to produce eggs. These speculations are supported by greenstick fractures being the type of fracture most commonly reported in the keel (Casey-Trott et al., 2017a; Thøfner et al., 2020). A recent study has demonstrated with the use of histopathological techniques that the fractures indeed look like greenstick fractures owing to the lack of healing (Thøfner et al., 2020). Inflammatory cells were shown to be absent around the fractures, which is a characteristic of stress and greenstick fractures, wherein a constant stress impedes healing. In humans, greenstick fractures are most common in pediatric patients and occur when an immature bone is bent and begins to fracture although the fracture line is not complete (Rodríguez-Merchán, 2005). As laying hens would naturally reach 8 to 10 yr of age, 40 wk of age can be viewed as a pediatric age, with an immature keel capable of flexing to a certain degree, although still prone to fracture. It has not been investigated if the nature of fractures is histologically different when occurring in relatively young animals, animals older than than 40 wk of age, and those older than 1 yr of age, although efforts to understand causes of fracture would benefit from this comparison.
Predisposing Bone Diseases
Other possibilities to explain KBF include the existence of underlying disease state that may resemble an injury resulting from collision. Collisions and their resulting injury, referred to as traumatic, would normally take on a morphological characteristic, wherein the bone is split into 2 or more pieces, with one of the several distinctive appearances (Baumgartner and Ulrich, 2015). The healing process of the injury will typically involve callus formation that can be detected by the rather coarse techniques of palpation or direct examination of excised keels (Casey-Trott et al., 2015). However, a callus-like expansion of bone is not a definitive proof of fractured bone as it can also result from other processes such as periosteal irritations, congenital malformations, periostitis, osteomyelitis, and bone tumors (Baumgartner and Ulrich, 2015). The recent use of radiography to accurately detect fractures (Richards et al., 2011; Eusemann et al., 2018a; Rufener et al., 2018b, 2019; Rentsch et al., 2019; Baur et al., 2020) within a relatively narrow time window should be exploited to allow for immediate macroscopic and histopathologic examination. A comprehensive morphological examination of lesions immediately after the onset is needed for proof of principal that the lesion in question is indeed a fracture and identification of the cause. In this scenario, although more severe, or traumatic, fractures may result from collisions, there is also a possibility for a predisposing disease resulting in a weakened skeletal architecture. These predisposed bones would be expected to break easier, even with routine, daily forces that would be considered innocuous. Fractures in such a predisposed bone are referred to as pathological fractures. In contrast, traumatic fractures occur only with increased or massive force acting on normally structured bones such as collisions (Craig et al., 2016). Reports of recurrent fractures at different ages and in multiple locations (Richards et al., 2011; Rufener et al., 2018b, 2019; Baur et al., 2020) are suggestive of a predisposing primary disease.
We propose that traumatic and pathological fractures differ in several meaningful characteristics including the affected anatomic locations and the general pathomorphology of the fractured bone and associated soft tissues. Traumatic fractures of the keel likely manifest as multiple fragments in primarily middle and cranial portions, possibly accompanied by prominent traumatic changes in the surrounding soft tissues. In contrast, pathological fractures will occur at the caudal end of the sternum where only periosteal woven bone formation is seen, suggestive of a previous fracture (Eusemann et al., 2018a). Future work should continue to differentiate these types of fractures and how they can be better classified within experimental scoring systems. In considering these 2 fracture classes, it is important to be aware of the fact that a bird with weakened bone can have fractures owing to both high-impact, traumatic events and low-force stress acting on the least resistant parts of the bone, that is, pathological fractures. In contrast, birds with a normal (nonpredisposed) skeleton would have only traumatic fractures. The existence of pathological fractures is supported by the absence of fractures in males of modern genetic lines (Fleming et al., 2004) and hens in which egg production is suppressed (Eusemann et al., 2018b).
Numerous predisposing conditions of pathological fractures are possible, and only a limited number of them are considered here. Conditions predisposing to pathological fractures theoretically may include those of degenerative (e.g., osteoporosis, osteodystrophia fibrosa, rickets/osteomalacia), dysplastic (e.g., osteogenesis imperfecta or glass bone disease), inflammatory (e.g., osteomyelitis), and neoplastic characteristics. Most of these diseases are exogenous, and discussed endogenous pathologies were obviously not a goal of selection programs. However, it is possible that a form of inadvertent selection occurred. In this scenario, while selecting for desired traits such as efficient egg production, undesirable traits were also selected in a form of genetic hitchhiking, leading to a compromised phenotype. Evidence for genetic hitchhiking is relatively sparse. Specifically for chickens, there was no evidence of coinheritance between pairs of neutral loci in a selection experiment (Derks et al., 2018), whereas the phenomena was shown to exist between pairs of loci within quantitative trait loci (Pinard Van Der Laan et al., 2008).
Focusing on osteoporosis, which is generally cited as relevant to the cause of fractures, it may also impair repair of damaged bone by interfering with the many processes involved in intramembranous ossification or/and endochondral ossification (Cheung et al., 2016). Although healing is generally seen to occur within 6 to 8 wk for the majority of hens (Richards et al., 2011; Baur et al., 2020), longitudinal studies using radiographs in laying hens have also demonstrated relatively delayed (Rufener et al., 2018a) or even a lack of healing, as shown in one study with 16% of surveyed hens (Baur et al., 2020). Delayed healing is also known to occur in response to elevated estrogen concentrations (Engin et al., 1983). It is possible that if hens are unable to adequately repair small, innocuous fractures called greenstick fractures, the bone's ultimate strength could become compromised, leading to fracture. In that sense, the influence of egg laying activity on fracture repair is most likely quantitative but not qualitative.
Unfortunately, the literature on poultry diseases shows an unfavorable trend of confusing the metabolic diseases osteoporosis and osteomalacia because many experiments lack the inclusion of techniques able to truly differentiate these conditions, such as histopathology and bone mineral analysis. The morphologic diagnosis within the piece of bone under investigation is osteopenia and has to be distinguished from the name of the systemic disease which is osteoporosis. Osteopenia can occur owing to different reasons, for example, disuse of a limb leading to an osteopenic limb. In contrast, osteoporosis is a metabolic disease principally weakening all bones although with different speeds and extents.
Genetic predisposition may arise of a congenital nature or later in life. In terms of congenital conditions, although some historical information regarding growth and ossification (Buckner et al., 1948, 1949) and more recent work (Gibson et al., 1995) are available, extensive information about the laying hen's keel ossification is lacking, including enchondral vs. desmal ossification or the number of growth centers that exist. Concerning later development, it is known that hens at the onset of egg laying retain a cartilaginous caudal end of the sternum, suggestive of a zone of ongoing growth (Buckner et al., 1948) that could be a locus of least resistance similar to the characteristic metaphyseal rib swellings in rickets of growing vitamin D–deficient mammals (Baumgartner and Ulrich, 2015; Craig et al., 2016).
Finally, tumors of bone-like osteosarcoma or within the bone marrow–like malignant lymphoma and extramedullary plasmacytoma are common predisposing factors for fractures (Baumgartner and Ulrich, 2015; Craig et al., 2016). Notably avian sarcoma leukosis, which is an alpharetrovirus-induced neoplastic disease, can occur enzootically affecting complete flocks and can induce tumors in multiple soft tissues and bone (Abdul-Aziz and Barnes, 2017). The target cell of the retrovirus is myelomonocytic cells in the bone marrow, and the forming myelocytic tumors referred to as myelocytomas are reported to characteristically occur on the surface of bones and cartilage and distort the contours of the bones, resulting in deformations hard to distinguish without histopathology from callus formation. It is unlikely that tumors are widespread and a dominant cause of fractures although, in pursuit of this review's stated objectives, alternative explanations should be considered even if the implications are limited to a very small subset of occurrences.
Inactivity Leading to Reduced Bone Strength
The mechanostat (Frost, 2003) is an evolving hypothesis that originated with earlier proposals by Wolff, 1892 that theorizes that the biological mechanisms that regulate bone strength and adapt to experienced mechanical loads. At risk of oversimplification, these theories propose that use of a bone or loading will lead to increased strength in that bone through additions of bone mass and changes in the cross-sectional shape via osteocytes within the bone that serve to detect mechanical strain (reviewed by Robling and Turner, 2009).
Interestingly, although noncage systems would be expected to involve more bone loading owing to the additional activity required to move between resources such as litter for dust bathing, nest box for egg laying, feed, and water lines (Fröhlich and Oester, 2001; Blokhuis et al., 2005), noncage systems actually have higher rates of fractures (Rodenburg et al., 2008; Wilkins et al., 2011; Petrik et al., 2015). Wilkins et al. (2011) reasoned that the variation in fractures observed between systems was likely due to perches causing more high-energy collisions, a view supported by the increased fracture occurrence and severity seen in systems with more and higher items available for perching. Paradoxically, birds in noncage systems appear to manifest superior bone strength (Tauson and Abrahamsson, 1994; Jendral et al., 2008; Rodenburg et al., 2008; Regmi et al., 2013, 2015) and cortical density and thickness (Regmi et al., 2016b) compared with birds in the cage system, traits that likely result from greater activity. Rodriguez-Navarro et al. (2018) found that, similar to the aforementioned findings, cortical bone from aviary hens was stronger and had increased cortical thickness but a lower degree of mineralization, and bone mineral was less mature and less organized than in caged birds. Few studies have investigated the effect of exercise on the keel bone, but it is assumed that greater use of the wings should increase its strength, and this is borne out by measurement of keel radiographic density (Fleming et al., 2006). Birds with greater bone strength would be expected to be less susceptible to fractures from collisions, a detail that highlights the difficulty in identifying an appropriate balance between providing an important resource that birds are motivated to use (i.e., perches) while limiting injury (Sandilands et al., 2009). In other words, housing features that lead to improvements in bone health do not necessarily lead to reduced fracture incidence (Regmi et al., 2016a), although the opposite has been reported (Stratmann et al., 2015b,a). The actual benefit of housing adaptations likely lies in whether the adaptations meet the behavior and needs of animals themselves (Stratmann et al., 2015b,a). Within the vein of the current article, given that egg production between noncage and cage systems is similarly high (>85% hens' daily average over the lifetime of the flock) but fracture rates are so different, it suggests the involvement of causal factors other than egg production such as activity levels. The differential rates in fracture are normally attributed to the belief that hens in cage systems would be exposed to collision energies of a lesser magnitude (Harlander-Matauschek et al., 2015), although relatively high-energy collisions do occur in cage systems (Baker et al., 2020). Given the lack of knowledge, the role of activity in general and specific types of activities should be investigated further.
In support of the hypothesis of reduced activity leading to increased fractures, hens within noncage systems were shown to exhibit highly individualized behavior that would likely lead to dramatic but varied effects on bone strength (Rufener et al., 2018c). Rufener et al. (2018c) reported that, despite all hens being in the same aviary, the number of transitions between aviary tiers and the winter garden was highly specific to individual animals, with total daily transitions ranging from 20 to 159. Similar patterns of differential movement patterns were observed in terms of movement between inside and outside barn areas (Y. Gómez, unpublished work; Gebhardt-Henrich et al., 2014; Larsen et al., 2017; Taylor et al., 2017). In this scenario, although all birds are contained within the same housing system, the differential activity levels would be expected to result in substantially different bone loading and consequent bone strength. Future work should investigate if there exists a subpopulation of hens within a flock that adopt relatively reduced activity behavioral patterns and the relationship with keel fracture susceptibility. Y. Gómez (unpublished data) reported consistent movement patterns for groups of individuals that appeared to form an intrapen community, although no data on KBF were taken. Rufener et al. (2019) found that birds with KBF had a similar number of transitions within a commercial aviary, although maintained the majority of their time in upper tiers. Unfortunately, Rufener et al. (2019) were not able to establish causality, so it is not possible to say if the different behavior was a cause or effect of the keel fractures.
Future research areas
The current article is intended to highlight evidence that KBF are likely related to elevated and sustained egg production although multiple published reports suggest the role of complementary or additional contributory factors that were discussed. Unfortunately, much of this information is piecemeal, isolated, and/or conducted with methods or genetic lines and hybrids that limit application to current commercial conditions. With that in mind, we have attempted to identify several key areas where research would be advantageous. Those areas include the following:
-
1.
Continued examination to link egg production and keel bone health and fractures throughout the laying cycle
-
2.
Examination of noncommercial aves and traditional breeds for evidence of keel bone damage
-
3.
Exploration of manipulating AFE with an emphasis on keel-based fracture evaluations and controlling for likely influential factors such as body mass and hybrid. The potential impact on fractures should be analyzed at multiple ages.
-
4.
A detailed histological and structural analysis of the keel from development to full ossification to link age-related changes with the type of fracture occurring and fracture location
-
5.
Assessment of prefracture bone condition to evaluate different occurrence of pathological and traumatic fractures, possibly in combination with techniques to moderate egg production, for example, photostimulation
-
6.
Assess the relationship between individual activity patterns and consequent bone health and KBF susceptibility
Acknowledgments
The authors would like to greatly thank the financial support of the KeelBoneDamageNet (www.keelbonedamage.eu, CA15224), an EU-COST Action, in funding the travel, accommodation, and organizational costs for the meeting, in which the discussions that lead to this manuscript took place. The action also provided for the associated publication costs. The authors also would like to greatly thank all attendees of the associated meeting for the lively discussion of the associated topics. Those attendees, in alphabetical order, were as follows: Ahmet Akkoc, Björn Andersson, Zsolt Becskei, Cristina Benavides, Boris Bilcik, Enver Cavusoglu, Jens Peter Christensen, Dunn Ian, Sokol Duro, Beryl Eusemann, Elisa Folegatti, Marian Johannes Graf, Tone Beate Hansen, Zlatko Janjecic, Lisa Jung, Käthe Elise Kittelsen, Lazarin Lazarov, Maisarah Maidin, Metin Petek, Stefanie Petow, Prafulla Regmi, Alejandro B. Rodríguez Navarro, Victoria Sandilands, Mihaela Saracila, John Tarlton, Freek Thomassen, Michael Toscano, and Reiner Ulrich.
Conflict of Interest Statement: The authors did not provide a conflict of interest statement.
References
- Abdul-Aziz T., Barnes H.J. American Association of Avian Pathologists (AAAP); Madison, WI: 2017. Myelocytic Leukosis (Myelocytomatosis) [Google Scholar]
- Abplanalp H. Genetic and environmental correlations among production traits of poultry. Poult. Sci. 1957;36:226–228. [Google Scholar]
- Anwar M.N., Ravindran V., Morel P.C.H., Ravindran G., Cowieson A.J. Effect of limestone particle size and calcium to non-phytate phosphorus ratio on true ileal calcium digestibility of limestone for broiler chickens. Br. Poult. Sci. 2016;57:707–713. doi: 10.1080/00071668.2016.1196341. [DOI] [PubMed] [Google Scholar]
- Bain M.M., Nys Y., Dunn I.C. Increasing persistency in lay and stabilising egg quality in longer laying cycles. What are the challenges? Br. Poult. Sci. 2016;57:330–338. doi: 10.1080/00071668.2016.1161727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker S.L., Robison C.I., Karcher D.M., Toscano M.J., Makagon M.M. Keel impacts and associated behaviors in laying hens. Appl. Anim. Behav. Sci. 2020;222:104886. [Google Scholar]
- Baumgartner W., Ulrich R. Stütz und Bewegungsapparat. In: Gruber A.D., editor. Spezielle Pathologie für die Tiermedizin. Baumagärtner, W. First. Georg Thieme Verlag; Stuttgart: 2015. pp. 309–341. [Google Scholar]
- Baur S., Rufener C.B., Toscano M.J., Geissbühler U. Radiographic evaluation of keel bone damage in laying hens – morphologic and temporal observations in a longitudinal study. Front. Vet. Sci. 2020;7:129. doi: 10.3389/fvets.2020.00129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birchard S.J., Sherding R.G. Section 8. Osteochondrosis.Page 1203 in Saunders manual of small animal practice-E-Book. In: Duncan L., Fatham L., Gower J., editors. Third. Elsevier Health Sciences; St Louis, MO: 2005. [Google Scholar]
- Bishop S.C., Fleming R.H., McCormack H.A., Flock D.K., Whitehead C.C. Inheritance of bone characteristics affecting osteoporosis in laying hens. Br. Poult. Sci. 2000;41:33–40. doi: 10.1080/00071660086376. [DOI] [PubMed] [Google Scholar]
- Bittner K., Vischer P., Bartholmes P., Bruckner P. Role of the subchondral vascular system in endochondral ossification: endothelial cells specifically derepress late differentiation in resting chondrocytesin vitro. Exp. Cell Res. 1998;238:491–497. doi: 10.1006/excr.1997.3849. [DOI] [PubMed] [Google Scholar]
- Blokhuis H., Cepero R., Colin P., Elson A., Fiks van Niekerk T., Keeling L., Michel V., Nicol C.J., Oester H., Tauson R. Welfare aspects of various systems of keeping laying hens. EFSA J. 2005;197:1–23. [Google Scholar]
- Buckner G.D., Insko W.M., Jr., Henry A.H., Wachs E.F. Rate of growth and calcification of the sternum of male and female New Hampshire chickens. Poult. Sci. 1948;27:430–433. [Google Scholar]
- Buckner G.D., Insko W.M., Jr., Henry A.H., Wachs E.F. Rate of growth and calcification of the sternum of male and female New Hampshire chickens having crooked keels. Poult. Sci. 1949;28:289–292. [Google Scholar]
- Buijs S., Heerkens J.L.T., Ampe B., Delezie E., Rodenburg T.B., Tuyttens F.A.M. Assessing keel bone damage in laying hens by palpation: effects of assessor experience on accuracy, inter-rater agreement and intra-rater consistency. Poult. Sci. 2018;98:514–521. doi: 10.3382/ps/pey326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Candelotto L., Stratmann A., Gebhardt-Henrich S.G., Rufener C., van de Braak T., Toscano M.J. Susceptibility to keel bone fractures in laying hens and the role of genetic variation. Poult. Sci. 2017;96:3539–3549. doi: 10.3382/ps/pex146. [DOI] [PubMed] [Google Scholar]
- Casey-Trott T.M. 2016. Opportunities One for Excercise during Pullet Rearing: Effects on Bone Health and Keel Bone Damage in Laying Hens. [Google Scholar]
- Casey-Trott T.M., Guerin M.T., Sandilands V., Torrey S., Widowski T.M. Rearing system affects prevalence of keel-bone damage in laying hens: a longitudinal study of four consecutive flocks. Poult. Sci. 2017;96:2029–2039. doi: 10.3382/ps/pex026. [DOI] [PubMed] [Google Scholar]
- Casey-Trott T., Heerkens J.L.T., Petrik M.T., Regmi P., Schrader L., Toscano M.J., Widowski T.M. Methods for assessment of keel bone damage in poultry. Poult. Sci. 2015;71:461–472. doi: 10.3382/ps/pev223. [DOI] [PubMed] [Google Scholar]
- Casey-Trott T.M., Korver D.R., Guerin M.T., Sandilands V., Torrey S., Widowski T.M. Opportunities for exercise during pullet rearing, Part II: long-term effects on bone characteristics of adult laying hens at the end-of-lay. Poult. Sci. 2017;96:2518–2527. doi: 10.3382/ps/pex060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey-Trott T.M. University of Guelph; Guelph, Ontario, Canada: 2016. Opportunities for Exercise during Pullet Rearing: Effects on Bone Health and Keel Bone Damage in Laying Hens. PhD Thesis. [Google Scholar]
- Chargo N.J., Robison C.I., Akaeze H.O., Baker S.L., Toscano M.J., Makagon M.M., Karcher D.M. Keel bone differences in laying hens housed in enriched colony cages. Poult. Sci. 2018;98:1031–1036. doi: 10.3382/ps/pey421. [DOI] [PubMed] [Google Scholar]
- Chargo N.J., Robison C.I., Baker S.L., Toscano M.J., Makagon M.M., Karcher D.M. Keel bone damage assessment: consistency in enriched colony laying hens. Poult. Sci. 2018;98:1017–1022. doi: 10.3382/ps/pey373. [DOI] [PubMed] [Google Scholar]
- Cheung W.H., Miclau T., Chow S.K.-H., Yang F.F., Alt V. Fracture healing in osteoporotic bone. Injury. 2016;47:S21–S26. doi: 10.1016/S0020-1383(16)47004-X. [DOI] [PubMed] [Google Scholar]
- Craig L.E., Dittmer K.E., Thompson K.G. 6th ed. Elsevier; St Louis, MO: 2016. Bones and joints; pp. 16–163. (Jubb, Kennedy & Palmer's Pathology of Domestic Animals, Volume 1. M. Grant Maxie, eds.). [Google Scholar]
- Dacke C.G., Arkle S., Cook D.J., Wormstone I.M., Jones S., Zaidi M., Bascal Z.A. Medullary bone and avian calcium regulation. J. Exp. Biol. 1993;184:63–88. [Google Scholar]
- Darwin C.R. John Murray; London, UK: 1868. Variation of Plants and Animals under Domestication. [Google Scholar]
- Derks M.F.L., Megens H.-J., Bosse M., Visscher J., Peeters K., Bink M.C.A.M., Vereijken A., Gross C., De Ridder D., Reinders M.J.T. A survey of functional genomic variation in domesticated chickens. Genet. Sel. Evol. 2018;50:17. doi: 10.1186/s12711-018-0390-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunn I.C., Fleming R.H., McCormack H.A., Morrice D., Burt D.W., Preisinger R., Whitehead C.C. A QTL for osteoporosis detected in an F2 population derived from White Leghorn chicken lines divergently selected for bone index. Anim. Genet. 2007;38:45–49. doi: 10.1111/j.1365-2052.2006.01547.x. [DOI] [PubMed] [Google Scholar]
- Dunn I.A.N.C., Sharp P.J. The effect of photoperiodic history on egg laying in dwarf broiler hens. Poult. Sci. 1992;71:2090–2098. doi: 10.3382/ps.0712090. [DOI] [PubMed] [Google Scholar]
- EFSA Panel on Animal Health and Animal Welfare Scientific opinion on welfare aspects of the use of perches for laying hens. EFSA J. 2015;13:4131. [Google Scholar]
- Engin A.E., Toney L.R., Negulesco J.A. Effects of oestrogen upon tensile properties of healing fractured avian bone. J. Biomed. Eng. 1983;5:49–54. doi: 10.1016/0141-5425(83)90078-x. [DOI] [PubMed] [Google Scholar]
- Eusemann B.K., Baulain U., Schrader L., Thöne-Reineke C., Patt A., Petow S. Radiographic examination of keel bone damage in living laying hens of different strains kept in two housing systems. PLoS One. 2018;13:e0194974. doi: 10.1371/journal.pone.0194974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eusemann B.K., Sharifi A.R., Patt A., Reinhard A.-K., Schrader L., Thöne-Reineke C., Petow S. Influence of a sustained release deslorelin acetate implant on Reproductive physiology and associated traits in laying hens. Front. Physiol. 2018;9:1846. doi: 10.3389/fphys.2018.01846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FAWC. Farm Animal Welfare Council; London: 2010. Opinion on Osteoporosis and Bone Fractures in Laying Hens. [Google Scholar]
- FAWC. 2013. An Open Letter to Great Britain Governments: Keel Bone Fracture in Laying Hens. [Google Scholar]
- Fleming R.H. Nutritional factors affecting poultry bone health. Proc. Nutr. Soc. 2008;67:177–183. doi: 10.1017/S0029665108007015. [DOI] [PubMed] [Google Scholar]
- Fleming R.H., McCormack H.A., McTeir L., Whitehead C.C. Effects of dietary particulate limestone, vitamin K3 and fluoride and photostimulation on skeletal morphology and osteoporosis in laying hens. Br. Poult. Sci. 2003;44:683–689. doi: 10.1080/00071660310001643688. [DOI] [PubMed] [Google Scholar]
- Fleming R.H., McCormack H.A., McTeir L., Whitehead C.C. Incidence, pathology and prevention of keel bone deformities in the laying hen. Br. Poult. Sci. 2004;45:320–330. doi: 10.1080/00071660410001730815. [DOI] [PubMed] [Google Scholar]
- Fleming R.H., McCormack H.A., McTeir L., Whitehead C.C. Relationships between genetic, environmental and nutritional factors influencing osteoporosis in laying hens. Br. Poult. Sci. 2006;47:742–755. doi: 10.1080/00071660601077949. [DOI] [PubMed] [Google Scholar]
- Freitas F. Metabolic alterations in broiler chickens experimentally infected with sporulated oocysts of eimeria maxima. Rev. Bras. Parasitol. Vet. 2014;23:309–314. doi: 10.1590/s1984-29612014057. [DOI] [PubMed] [Google Scholar]
- Fröhlich E.K.F., Oester H. From battery cages to aviaries: 20 years of Swiss experiences. Das Wohlergehen von Legehennen Eur. Anal. Schlussfolgerungen. 2001;28:176. [Google Scholar]
- Frost H.M. Bone’s mechanostat: a 2003 update. Anat. Rec. Part A. Discov. Mol. Cell. Evol. Biol. 2003;275:1081–1101. doi: 10.1002/ar.a.10119. [DOI] [PubMed] [Google Scholar]
- Gebhardt-Henrich S.G., Fröhlich E.K.F. Early onset of laying and bumblefoot favor keel bone fractures. Animals. 2015;5:1192–1206. doi: 10.3390/ani5040406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gebhardt-Henrich S.G., Rufener C., Stratmann A. Improving intra-and inter-observer repeatability and accuracy of keel bone assessment by training with radiographs. Poult. Sci. 2019;98:5234–5240. doi: 10.3382/ps/pez410. [DOI] [PubMed] [Google Scholar]
- Gebhardt-Henrich S.G., Toscano M.J., Fröhlich E.K.F. Use of outdoor ranges by laying hens in different sized flocks. Appl. Anim. Behav. Sci. 2014;155:74–81. [Google Scholar]
- Gebhardt-Henrich S.G., Toscano M.J., Würbel H. Perch use by broiler breeders and its implication on health and production. Poult. Sci. 2017;96:3539–3549. doi: 10.3382/ps/pex189. [DOI] [PubMed] [Google Scholar]
- Gebhardt-Henrich S.G., Toscano M.J., Würbel H. Use of aerial perches and perches on aviary tiers by broiler breeders. Appl. Anim. Behav. Sci. 2018;203:24–33. [Google Scholar]
- Gibson G.J., Kohler W.J., Schaffler M.B. Chondrocyte apoptosis in endochondral ossification of chick sterna. Dev. Dyn. 1995;203:468–476. doi: 10.1002/aja.1002030409. [DOI] [PubMed] [Google Scholar]
- Gregory N.G., Wilkins L.J., Eleperuma S.D., Ballantyne A.J., Overfield N.D. Broken bones in domestic fowls: effect of husbandry system and stunning method in end-of-lay hens. Br. Poult. Sci. 1990;31:59–69. [Google Scholar]
- Guinotte F., Gautron J., Nys Y., Soumarmon A. Calcium solubilization and retention in the gastrointestinal tract in chicks (Gallus domesticus) as a function of gastric acid secretion inhibition and of calcium carbonate particle size. Br. J. Nutr. 1995;73:125–139. doi: 10.1079/bjn19950014. [DOI] [PubMed] [Google Scholar]
- Harlander-Matauschek A., Rodenburg T.B.B., Sandilands V., Tobalske B.W., Toscano M.J. Causes of keel bone damage and their solutions in laying hens. Worlds Poult. Sci. J. 2015;71:461–472. [Google Scholar]
- Heerkens J.L.T., Delezie E., Rodenburg T.B., Kempen I., Zoons J., Ampe B., Tuyttens F.A.M. Risk factors associated with keel bone and foot pad disorders in laying hens housed in aviary systems. Poult. Sci. 2015;95:482–488. doi: 10.3382/ps/pev339. [DOI] [PubMed] [Google Scholar]
- Hester P.Y., Wilson D.A., Settar P., Arango J.A., O’Sullivan N.P. Effect of lighting programs during the pullet phase on skeletal integrity of egg-laying strains of chickens. Poult. Sci. 2011;90:1645–1651. doi: 10.3382/ps.2011-01411. [DOI] [PubMed] [Google Scholar]
- Hocking P.M., Bain M., Channing C.E., Fleming R., Wilson S. Genetic variation for egg production, egg quality and bone strength in selected and traditional breeds of laying fowl. Br. Poult. Sci. 2003;44:365–373. doi: 10.1080/0007166031000085535. [DOI] [PubMed] [Google Scholar]
- Hocking P.M., Sandercock D.A., Wilson S., Fleming R.H. Quantifying genetic (co) variation and effects of genetic selection on tibial bone morphology and quality in 37 lines of broiler, layer and traditional chickens. Br. Poult. Sci. 2009;50:443–450. doi: 10.1080/00071660903110927. [DOI] [PubMed] [Google Scholar]
- Hudson H.A., Britton W.M., Rowland G.N., Buhr R.J. Histomorphometric bone properties of sexually immature and mature White Leghorn hens with evaluation of fluorochrome injection on egg production traits. Poult. Sci. 1993;72:1537–1547. doi: 10.3382/ps.0721537. [DOI] [PubMed] [Google Scholar]
- Jendral M.J., Korver D.R., Church J.S., Feddes J.J. Bone mineral density and breaking strength of White Leghorns housed in conventional, modified, and commercially available colony battery cages. Poult. Sci. 2008;87:828–837. doi: 10.3382/ps.2007-00192. [DOI] [PubMed] [Google Scholar]
- Käppeli S., Gebhardt-Henrich S.G., Fröhlich E., Pfulg A., Stoffel M.H. Prevalence of keel bone deformities in Swiss laying hens (L Lidfors, HJ Blokhuis, and L Keeling, Eds.) Br. Poult. Sci. 2011;52:531–536. doi: 10.1080/00071668.2011.615059. [DOI] [PubMed] [Google Scholar]
- Kerr C.L., Hammerstedt R.H., Barbato G.F. Effects of selection for exponential growth rate at different ages on reproduction in chickens. Avian Poult. Biol. Rev. 2001;12:127–136. [Google Scholar]
- Larsen H., Cronin M.G., Gebhardt-Henrich G.S., Smith L.C., Hemsworth H.P., Rault J.-L. Individual ranging behaviour patterns in commercial free-range layers as observed through RFID Tracking. Animals. 2017;7:21. doi: 10.3390/ani7030021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lay D.C., Fulton R.M., Hester P.Y., Karcher D.M., Kjaer J.B., Mench J.A., Mullens B.A., Newberry R.C., Nicol C.J., O’Sullivan N.P., Porter R.E. Hen welfare in different housing systems. Poult. Sci. 2011;90:278–294. doi: 10.3382/ps.2010-00962. [DOI] [PubMed] [Google Scholar]
- Lewis P.D., Tyler N.C., Gous R.M., Dunn I.C., Sharp P.J. Photoperiodic response curves for plasma LH concentrations and age at first egg in female broiler breeders. Anim. Reprod. Sci. 2008;109:274–286. doi: 10.1016/j.anireprosci.2008.02.012. [DOI] [PubMed] [Google Scholar]
- Morris T.R., Sharp P.J., Butler E.A. A test for photorefractoriness in high-producing stocks of laying pullets. Br. Poult. Sci. 1995;36:763–769. doi: 10.1080/00071669508417820. [DOI] [PubMed] [Google Scholar]
- Nasr M.A.F., Murrell J., Wilkins L.J., Nicol C.J. The effect of keel fractures on egg production parameters, mobility and behaviour in individual laying hens. Anim. Welf. 2012;21:127–135. [Google Scholar]
- Nasr M.A., Nicol C.J., Murrell J.C. Do laying hens with keel bone fractures experience pain? PLoS One. 2012;7:e42420. doi: 10.1371/journal.pone.0042420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norman A.W., Hurwitz S. The role of the vitamin D endocrine system in avian bone biology. J. Nutr. 1993;123:310–316. doi: 10.1093/jn/123.suppl_2.310. [DOI] [PubMed] [Google Scholar]
- Olgun O., Aygun A. Nutritional factors affecting the breaking strength of bone in laying hens. Worlds. Poult. Sci. J. 2016;72:821–832. [Google Scholar]
- Petrik M.T., Guerin M.T., Widowski T.M. On-farm comparison of keel fracture prevalence and other welfare indicators in conventional cage and floor-housed laying hens in Ontario, Canada. Poult. Sci. 2015;94:579–585. doi: 10.3382/ps/pev039. [DOI] [PubMed] [Google Scholar]
- Pinard Van Der Laan M.-H., Pitel F., Verrier E., Bijma P. Evolution of the polymorphism at molecular markers in QTL and non-QTL regions in selected chicken lines. Genet. Sel. Evol. 2008;40:639–661. doi: 10.1186/1297-9686-40-6-639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podisi B.K., Knott S.A., Dunn I.C., Burt D.W., Hocking P.M. Bone mineral density QTL at sexual maturity and end of lay. Br. Poult. Sci. 2012;53:763–769. doi: 10.1080/00071668.2012.747674. [DOI] [PubMed] [Google Scholar]
- Podisi B.K., Knott S.A., Dunn I.C., Law A.S., Burt D.W., Hocking P.M. Overlap of quantitative trait loci for early growth rate, and for body weight and age at onset of sexual maturity in chickens. Reproduction. 2011;141:381–389. doi: 10.1530/REP-10-0276. [DOI] [PubMed] [Google Scholar]
- Rath N.C., Huff G.R., Huff W.E., Balog J.M. Factors regulating bone maturity and strength in poultry. Poult. Sci. 2000;79:1024–1032. doi: 10.1093/ps/79.7.1024. [DOI] [PubMed] [Google Scholar]
- Raymond B., Johansson A.M., McCormack H.A., Fleming R.H., Schmutz M., Dunn I.C., De Koning D.J. Genome-wide association study for bone strength in laying hens. J. Anim. Sci. 2018;96:2525–2535. doi: 10.1093/jas/sky157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Regmi P., Anderson K., DM K. Proceedings of the 2013 Annual Meeting of the Poultry Science Association. Poultry Science Association; San Diego, CA: 2013. Comparison of bone quality between strains and housing systems in end-of-lay hens; p. 83. [Google Scholar]
- Regmi P., Deland T.S., Steibel J.P., Robison C.I., Haut R.C., Orth M.W., Karcher D.M. Effect of rearing environment on bone growth of pullets. Poult. Sci. 2015;94:502–511. doi: 10.3382/ps/peu041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Regmi P., Nelson N., Steibel J.P., Anderson K.E., Karcher D.M. Comparisons of bone properties and keel deformities between strains and housing systems in end-of-lay hens. Poult. Sci. 2016;95:2225–2234. doi: 10.3382/ps/pew199. [DOI] [PubMed] [Google Scholar]
- Regmi P., Smith N., Nelson N., Haut R.C., Orth M.W., Karcher D.M. Housing conditions alter properties of the tibia and humerus during the laying phase in Lohmann white Leghorn hens. Poult. Sci. 2016;95:198–206. doi: 10.3382/ps/pev209. [DOI] [PubMed] [Google Scholar]
- Rennie J.S., Fleming R.H., a McCormack H., McCorquodale C.C., Whitehead C.C. Studies on effects of nutritional factors on bone structure and osteoporosis in laying hens. Br. Poult. Sci. 1997;38:417–424. doi: 10.1080/00071669708418012. [DOI] [PubMed] [Google Scholar]
- Rentsch A.K., Rufener C.B., Spadavecchia C., Stratmann A., Toscano M.J. Laying hens’ mobility is impaired by keel bone fractures though effect is not Reversed by Paracetamol Treatment. Appl. Anim. Behav. Sci. 2019;217:48–56. [Google Scholar]
- Riber A.B., Hinrichsen L.K. Keel-bone damage and foot injuries in commercial laying hens in Denmark. Anim. Welf. 2016;25:179–184. [Google Scholar]
- Richards G.J., Nasr M.A., Brown S.N., Szamocki E.M.G., Murrell J., Barr F., Wilkins L.J. Use of radiography to identify keel bone fractures in laying hens and assess healing in live birds. Vet. Rec. 2011;169:279. doi: 10.1136/vr.d4404. [DOI] [PubMed] [Google Scholar]
- Robling A.G., Turner C.H. Mechanical signaling for bone modeling and remodeling. Crit. Rev. Eukaryot. Gene Expr. 2009;19:319. doi: 10.1615/critreveukargeneexpr.v19.i4.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodenburg T.B., Tuyttens F.A.M., De Reu K., Herman L., Zoons J., Sonck B. Welfare assessment of laying hens in furnished cages and non-cage systems : an on-farm comparison. Anim. Welf. 2008;17:363–373. [Google Scholar]
- Rodríguez-Merchán E.C. Pediatric skeletal trauma: a review and historical perspective. Clin. Orthop. Relat. Res. 2005;432:8–13. [PubMed] [Google Scholar]
- Rodriguez-Navarro A.B., McCormack H.M., Fleming R.H., Alvarez-Lloret P., Romero-Pastor J., Dominguez-Gasca N., Prozorov T., Dunn I.C. Influence of physical activity on tibial bone material properties in laying hens. J. Struct. Biol. 2018;201:36–45. doi: 10.1016/j.jsb.2017.10.011. [DOI] [PubMed] [Google Scholar]
- Rufener C., Abreu Y., Asher L., Berezowski J., Maximiano Sousa F., Stratmann A., Toscano M.J. Keel bone fractures are associated with individual mobiltiy of laying hen in aviary systems. Appl. Anim. Behav. Sci. 2019;217:48–56. [Google Scholar]
- Rufener C., Baur S., Stratmann A., Toscano M.J. Subjective but reliable: assessing radiographs of keel bone fractures in laying hens with a Tagged Visual Analogue Scale. Front. Vet. Sci. 2018;5:124. doi: 10.3389/fvets.2018.00124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rufener C., Baur S., Stratmann A., Toscano M.J. Keel bone fractures affect egg laying performance but not egg quality in laying hens housed in a commercial aviary system. Poult. Sci. 2018;98:1589–1600. doi: 10.3382/ps/pey544. [DOI] [PubMed] [Google Scholar]
- Rufener C., Baur S., Stratmann A., Würbel H., Geissbühler U., Toscano M. Proceedings of the 50th International Congress of the ISAE. Wageningen; Edinburgh, Scotland: 2016. Effects of keel bone fractures on individual laying hen productivity; p. 284. [Google Scholar]
- Rufener C., Berezowski J., Maximiano Sousa F., Abreu Y., Asher L., Toscano M.J. Finding hens in a haystack: consistency of movement patterns within and across individual laying hens maintained in large groups. Sci. Rep. 2018;8:12303. doi: 10.1038/s41598-018-29962-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandilands V., Moinard C., Sparks N.H.C. Providing laying hens with perches: fulfilling behavioural needs but causing injury? Br. Poult. Sci. 2009;50:395–406. doi: 10.1080/00071660903110844. [DOI] [PubMed] [Google Scholar]
- Sherwin C.M., Richards G.J., Nicol C.J. Comparison of the welfare of layer hens in 4 housing systems in the UK. Br. Poult. Sci. 2010;51:488–499. doi: 10.1080/00071668.2010.502518. [DOI] [PubMed] [Google Scholar]
- Silversides F.G., Korver D.R., Budgell K.L. Effect of strain of layer and age at photostimulation on egg production, egg quality, and bone strength. Poult. Sci. 2006;85:1136–1144. doi: 10.1093/ps/85.7.1136. [DOI] [PubMed] [Google Scholar]
- Soller M., Brody T., Eitan Y., Agursky T., Wexler C. Minimum weight for onset of sexual maturity in female chickens: heritability and phenotypic and genetic correlations with early growth rate. Poult. Sci. 1984;63:2103–2113. doi: 10.3382/ps.0632103. [DOI] [PubMed] [Google Scholar]
- Stratmann A., Fröhlich E.K., Gebhardt-Henrich S.G.G., Harlander-Matauschek A., Würbel H., Toscano M.J. Modification of aviary design reduces incidence of falls, collisions and keel bone damage in laying hens. Appl. Anim. Behav. Sci. 2015;165:112–123. [Google Scholar]
- Stratmann A., F. E. E.K., Fröhlich K.F., Harlander-Matauschek A., Schrader L., Toscano M.J.M.J., Würbel H., Gebhardt-Henrich S.G. Soft perches in an aviary system reduce incidence of keel bone damage in laying hens. PLoS One. 2015;10:e0122568. doi: 10.1371/journal.pone.0122568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Świątkiewicz S., Arczewska-Włosek A., Bederska-Lojewska D., Józefiak D. Efficacy of dietary vitamin D and its metabolites in poultry-review and implications of the recent studies. Worlds. Poult. Sci. J. 2017;73:57–68. [Google Scholar]
- Tarlton J.F.F., Wilkins L.J.J., Toscano M.J.J., Avery N.C.C., Knott L. Reduced bone breakage and increased bone strength in free range laying hens fed omega-3 polyunsaturated fatty acid supplemented diets. Bone. 2013;52:578–586. doi: 10.1016/j.bone.2012.11.003. [DOI] [PubMed] [Google Scholar]
- Tauson R., Abrahamsson P. Foot and skeletal disorders in laying hens: effects of Perch design, hybrid, housing system and Stocking density. Acta Agric. Scand. Sect. A. – Anim. Sci. 1994;44:110–119. [Google Scholar]
- Taylor P.S., Hemsworth P.H., Groves P.J., Gebhardt-Henrich S.G., Rault J.-L. Ranging behaviour of commercial free-range broiler chickens 2: individual variation. Animals. 2017;7:55. doi: 10.3390/ani7070055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiruvenkadan A.K., Panneerselvam S., Prabakaran R. Layer breeding strategies: an overview. Worlds. Poult. Sci. J. 2010;66:477–502. [Google Scholar]
- Thøfner I., Hougen H.P., Villa C., Lynnerup N., Christensen J.P. Pathological characterization of keel bone fractures in laying hens does not support external trauma as the underlying cause. PLoS One. 2020;15:e0229735. doi: 10.1371/journal.pone.0229735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toscano M.J., Booth F., Richards G., Brown S.N., Karcher D.M., Tarlton J.F. Modeling collisions in laying hens as a tool to identify causative factors for keel bone fractures and means to reduce their occurrence and severity (A Yildirim, Ed.) PLoS One. 2018;13:e0200025. doi: 10.1371/journal.pone.0200025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toscano M.J., Booth F., Wilkins L.J.J., Avery N.C.C., Brown S.B.B., Richards G., Tarlton J.F. The effects of long (C20/22) and short (C18) chain omega-3 fatty acids on keel bone fractures, bone biomechanics, behaviour and egg production in free range laying hens. Poult. Sci. 2015;94:823–835. doi: 10.3382/ps/pev048. [DOI] [PubMed] [Google Scholar]
- Toscano M.J., Wilkins L.J., Millburn G., Thorpe K., Tarlton J.F. Development of an ex vivo protocol to model bone fracture in laying hens resulting from collisions. (PE Witten, Ed.) PLoS One. 2013;8:e66215. doi: 10.1371/journal.pone.0066215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tracy L.M., Temple S.M., Bennett D.C., Sprayberry K.A., Makagon M.M., Blatchford R.A. The Reliability and accuracy of palpation, radiography, and sonography for the detection of keel bone damage. Animals. 2019;9:894. doi: 10.3390/ani9110894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warren D.C.D.E. Physiological and genetic studies of crooked keels in chickens. Kans. Agric. Exp. Stn. Tech. Bull. 1937;44:1–32. [Google Scholar]
- Whitehead C.C. Skeletal disorders in laying hens: the problem of osteoporosis and bone fractures. In: Perry G.C., editor. Welfare of the Laying Hen. CABI Publishing; Wallingford: 2004. pp. 259–270. [Google Scholar]
- Whitehead C.C. Overview of bone biology in the egg-laying hen. Poult. Sci. 2004;83:193–199. doi: 10.1093/ps/83.2.193. [DOI] [PubMed] [Google Scholar]
- Whitehead C.C., Fleming R.H. Osteoporosis in cage layers. Poult. Sci. 2000;79:1033–1041. doi: 10.1093/ps/79.7.1033. [DOI] [PubMed] [Google Scholar]
- Wilkins L.J., McKinstry J.L., Avery N.C., Knowles T.G., Brown S.N., Tarlton J., Nicol C.J. Influence of housing system and design on bone strength and keel bone fractures in laying hens. Vet. Rec. 2011;169:414. doi: 10.1136/vr.d4831. [DOI] [PubMed] [Google Scholar]
- Wilson S., Duff S.R.I., Whitehead C.C. Effects of age, sex and housing on the trabecular bone of kaying strain domestic fowl. Res. Vet. Sci. 1992;53:52–58. doi: 10.1016/0034-5288(92)90084-f. [DOI] [PubMed] [Google Scholar]
- Wolc A., Bednarczyk M., Lisowski M., Szwaczkowski T. Genetic relationships among time of egg formation, clutch traits and traditional selection traits in laying hens. J. Anim. Feed Sci. 2010;663:127. [Google Scholar]
- Wolff J. Hirschwald; Berlin: 1892. The Law of Bone Transformation. [Google Scholar]